
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Microbiol. , 18 May 2017
Sec. Food Microbiology
Volume 8 - 2017 | https://doi.org/10.3389/fmicb.2017.00846
 Daniel M. Linares1,2
Daniel M. Linares1,2 Carolina Gómez1Erica Renes3José M. Fresno3María E. Tornadijo3
Carolina Gómez1Erica Renes3José M. Fresno3María E. Tornadijo3 R. P. Ross2
R. P. Ross2 Catherine Stanton1,2*
Catherine Stanton1,2*Consumer interest in healthy lifestyle and health-promoting natural products is a major driving force for the increasing global demand of biofunctional dairy foods. A number of commercial sources sell synthetic formulations of bioactive substances for use as dietary supplements. However, the bioactive-enrichment of health-oriented foods by naturally occurring microorganisms during dairy fermentation is in increased demand. While participating in milk fermentation, lactic acid bacteria can be exploited in situ as microbial sources for naturally enriching dairy products with a broad range of bioactive components that may cover different health aspects. Several of these bioactive metabolites are industrially and economically important, as they are claimed to exert diverse health-promoting activities on the consumer, such as anti-hypertensive, anti-inflammatory, and anti-diabetic, anti-oxidative, immune-modulatory, anti-cholesterolemic, or microbiome modulation. This review aims at discussing the potential of these health-supporting bacteria as starter or adjunct cultures for the elaboration of dairy foods with a broad spectrum of new functional properties and added value.
Today foods are not intended to only satisfy hunger and to provide necessary nutrients for humans, but also to prevent nutrition-related diseases and improve consumers’ health (Siro et al., 2008; Gortzi et al., 2015). Increasing consumer demand and interest in obtaining additional benefits from food has stimulated functional foods to emerge on the market, with USA, Europe, and Japan being the dominant markets.
Although there is no unitary accepted definition, functional foods can be described as an ordinary food that has components or ingredients added to provide a specific health benefit, other than a purely nutritional effect. Ideally, functional food refers to an existing traditional food product that is intended to be consumed as part of a normal diet and has a demonstrated added physiological benefit (Siro et al., 2008). Therefore, it could not be in the form of pill or capsule. The concept of biofunctional foods is generally used when this desirable biological, medical, or physiological effect is exerted by microorganisms (Gobbetti et al., 2010). The health benefits of these microorganisms can be exerted either directly through the interactions of ingested live microorganisms with the host (probiotic effect), or indirectly by ingestion of the microbial metabolites synthesized during fermentation (bioactive effect) (Stanton et al., 2005; Gobbetti et al., 2010; Joshi, 2015).
Lactic acid bacteria (LAB) have been used to ferment foods for at least 4000 years (Rotar et al., 2007). Although the probiotic concept has expanded more recently, we have been unconsciously ingesting beneficial microbes with traditional fermented foods since ancient times. Fermented foods are the main carriers to deliver probiotics (Figure 1). Among them, dairy products (in particular fermented milks and yogurt) are by far the most efficient and widely used (Giraffa, 2012). Cheese is a dairy product which has a good potential for the incorporation of probiotic cultures due to its specific chemical and physical characteristics compared to fermented milks (higher pH value and lower titrable acidity, higher buffering capacity, greater fat content, higher nutrient availability, lower oxygen content, and denser texture). These conditions facilitate survivability of probiotic strains and tolerance to the low pH conditions encountered during gastric transit (Karimi et al., 2011). Utilization of probiotics has been optimized in several cheese varieties such as Cheddar, Gouda, Camembert, Cottage type, white-brined, and traditional cheeses, among others (Araujo et al., 2012; Giraffa, 2012; Martinovic et al., 2016; Oh et al., 2016). Kefir is another milk-fermented product that has health promoting bacteria (Prado et al., 2015). Other non-fermented dairy foods such as low-fat ice cream, chocolate mousse, coconut flan, or infant milk formula have also been supplemented with probiotic strains (Davidson et al., 2000; Aragon-Alegro et al., 2007; Correa et al., 2008; Baglatzi et al., 2016).
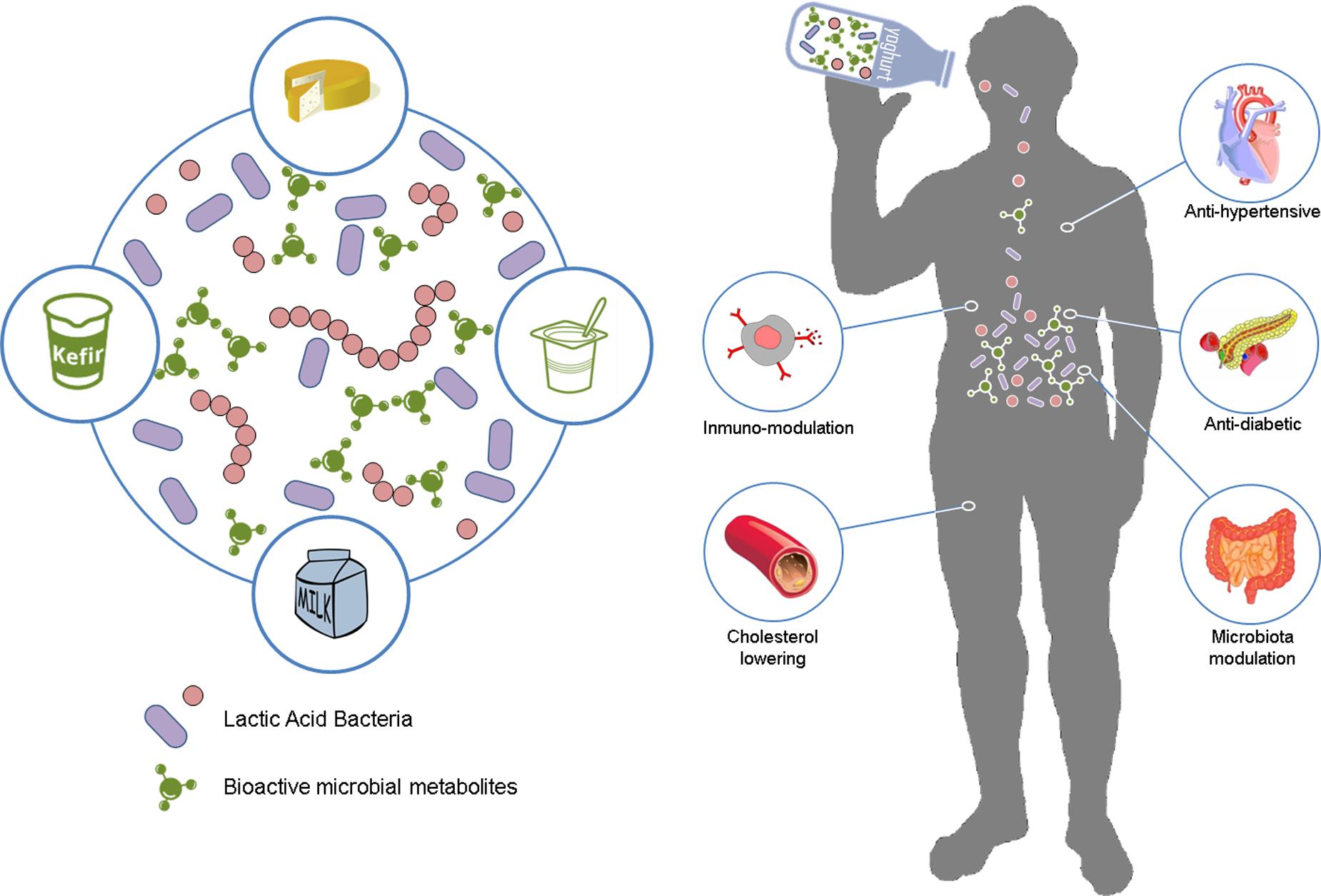
FIGURE 1. Beneficial effects resulting from the consumption of biofunctional fermented dairy foods. Lactic acid bacteria participating in milk fermentation in situ release and naturally enrich the fermented dairy product with a broad range of bioactive metabolites. Subsequent ingestion of this product can exert important health-promoting activities on the consumer, such as anti-hypertensive, and anti-diabetic, immune-modulatory, anti-cholesterolemic or microbiome modulation.
Probiotic microorganisms are generally LAB belonging to the species Lactobacillus acidophilus, L. gasseri, L. helveticus, L. johnsonii, L. (para)casei, L. reuteri, L. plantarum, L. rhamnosus, and L. fermentum, while members of the genus Bifidobacterium are also used, e.g., Bifidobacterium bifidum, B. longum, B. animalis, and B. breve (Tamime et al., 2005; Castro et al., 2015; Linares et al., 2016b). On the basis of the currently available literature, probiotics can balance intestinal microbiota, and thereby regulate proper intestinal function and be effective in the prevention or treatment of several gastrointestinal disorders such as infectious diarrhea, antibiotic-related diarrhea, irritable bowel syndrome or Crohn’s disease (Vanderhoof and Young, 1998) Other examples of health benefits promoted by probiotics supplied via dairy products are inmmunomodulatory effects (L. casei CRL431), reduction of serum cholesterol level (L. reuteri NCIMB 30242) and antihypertensive effects (L. plantarum TENSIATM) (EFSA, 2011; Jones et al., 2012; Aragon et al., 2014).
Probiotics are defined as ‘live micro-organisms, which when consumed in adequate amounts confer a health benefit on the host’ (FAO/WHO, 2001). However, regarding probiotic foods, some considerations are of paramount importance. Firstly, effective levels of the live probiotic in the corresponding food matrix at the time of ingestion are required. In this regard, the minimum effective dose which affects the intestinal environment and provides beneficial effects on human health is considered to be 106–109 live microbial cells per day, although this depends on the particular strain and foodstuff (Williams, 2010; Karimi et al., 2012; Watson and Preedy, 2015). Since probiotics show beneficial health effects on the host once consumed, another precondition for a bacterial strain to be called probiotic is the ability to survive and colonize (at least transiently) the gastrointestinal tract (GIT), which is in part helped by the buffering capacity of the food matrix. In some particular cases, bacterial viability may not be strictly required. As an example, inactivated and dead L. rhamnosus GG cells can maintain immunological and health-promoting effects (Ghadimi et al., 2008; Lopez et al., 2008).
Microorganisms involved in dairy fermentations can produce biologically active molecules and enzymes, giving the final food product an additional health value. Unlike the probiotic concept (the bacteria must be ingested alive and produce the beneficial metabolite in the body), the biofunctional concept is generally used when the healthy metabolite emerge in the food product itself during the fermentation process as a consequence of the bacterial metabolic activity. Consequently, the bacteria can act as a microbial factory to enrich foodstuff, for which bacterial viability through the GIT or during the product storage is not absolutely required (Farnworth and Champagne, 2015). The main bioactive compounds produced by LAB during dairy fermentation are vitamins, gamma-aminobutyric acid, bioactive peptides, bacteriocins, enzymes, conjugated linoleic acid, and exopolysaccharides (Table 1).
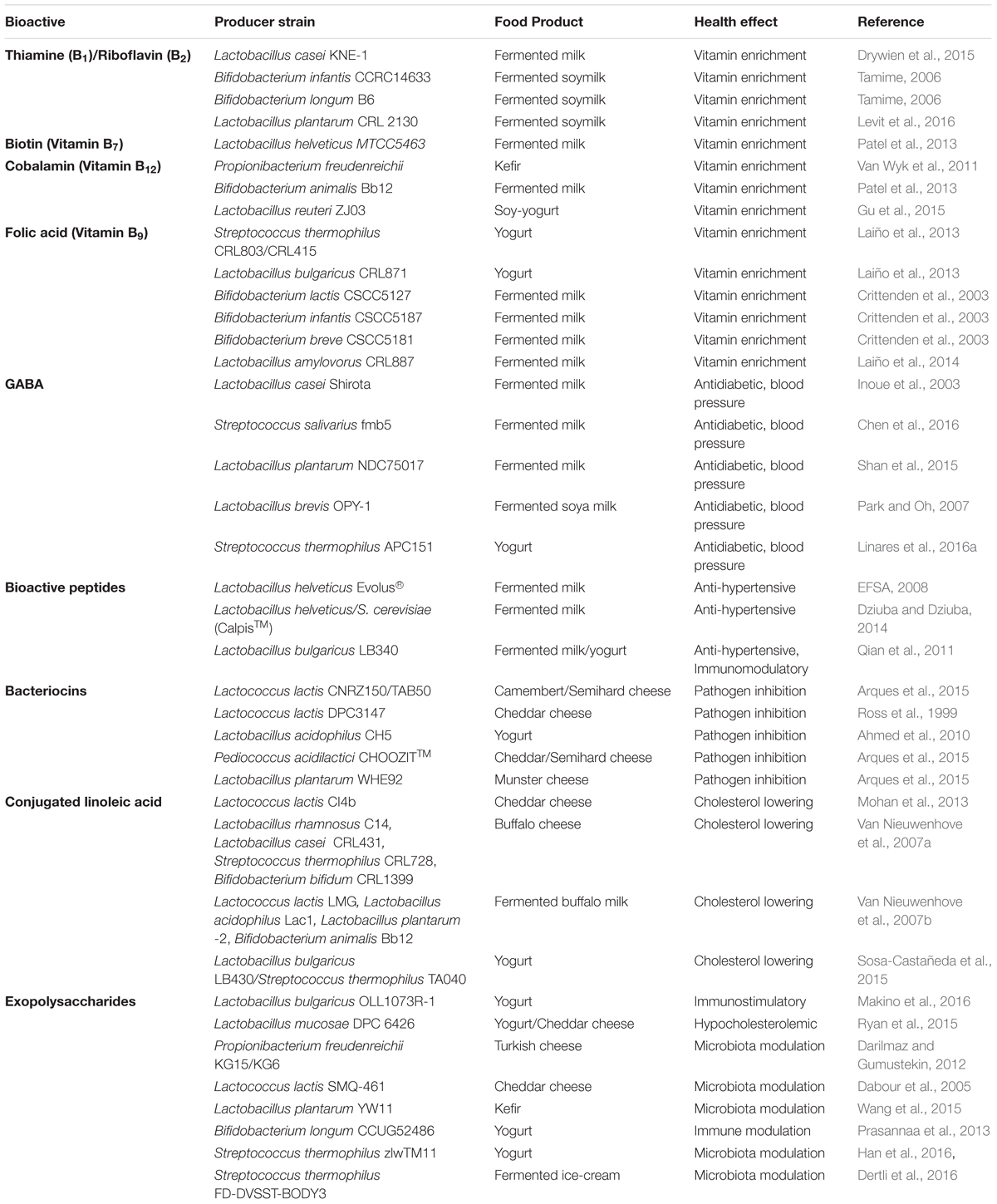
TABLE 1. Some strains of lactic acid bacteria, bifidobacteria and propionibacteria with potential to biosynthesize health-promoting compounds in fermented dairy products.
There are 13 vitamins that must be obtained exogenously due to the inability of humans to synthesize them; thereby they are essential nutrients in the human diet, and although in small amounts, a daily requirement is necessary to prevent deficiencies. Although most vitamins are present in a variety of foods, human vitamin deficiencies still occur in many countries, mainly because of malnutrition, not only as a result of insufficient food intake but also because of unbalanced diets (LeBlanc et al., 2011).
Although milk contains many vitamins fermentation by LAB and bifidobacteria constitute an effective way to increase vitamin levels in milk (Laiño et al., 2013). Some bacterial strains included in the genera Lactobacillus and Bifidobacterium can provide an additional source of B vitamins (thiamine, riboflavin, cobalamin, folate, and biotin) during dairy fermentation. Deficiencies in riboflavin (vitamin B2) or thiamine (vitamin B1) can lead to both liver and skin disorders and alterations in brain glucose metabolism, respectively (Russo et al., 2014). In this regard, L. casei KNE-1 has been shown to produce thiamine and riboflavin in fermented milk drinks (Drywien et al., 2015). B. infantis CCRC14633 and B. longum B6 strains have been reported to produce riboflavin and thiamine during soymilk fermentation (Tamime, 2006). It was recently indicated that soymilk fermented by the riboflavin-producing strain L. plantarum CRL2130 was able to prevent ariboflavinosis and experimental colitis in a murine model (Juarez del Valle et al., 2016; Levit et al., 2016). Some propionibacteria can produce cobalamin, folic acid, and biotin (Hugenholtz et al., 2002).
Folate (vitamin B9) is involved in several vital processes and its deficiency is generally linked to neural tube defects, certain forms of cancer, poor cognitive performance and coronary heart diseases. Even though vitamins are widely present in foods, the prevalence of folate deficiency -especially among women of child bearing age- is a growing concern and thereby folate fortification programs have been implemented in various countries (Divya and Nampoothiri, 2015). Rather than incorporating synthetic folate, foods can be naturally fortified with folate synthesized by LAB and bifidobacteria during manufacture of fermented foods (Lin and Young, 2000; Saubade et al., 2016). The strains Streptococcus thermophilus CRL803/CRL415 and L. bulgaricus CRL871 were reported to be suitable for the elaboration of yogurt naturally bio-enriched in this vitamin (Laiño et al., 2013). High folate concentration (up to 150 μg/l) can be reached in yogurt as a result of the ability of S. thermophilus to produce this vitamin (Hugenholtz et al., 2002). Among bifidobacteria, B. catenulatum ATCC 27539 was shown to produce high levels of folate in vitro (D’Aimmo et al., 2012), and B. lactis CSCC5127, B. infantis CSCC5187, and B. breve CSCC5181 strains increased folate concentration during fermentation of reconstituted skim milk (Crittenden et al., 2003). Similarly, L. amylovorus CRL887 can be used for natural folate bio-enrichment of fermented milk (Laiño et al., 2014).
The deficiency of cobalamin (vitamin B12) can be common, particularly in vegetarians who avoid ingestion of animal protein and use soymilk as an alternative to dairy milk (Gu et al., 2015). Animals, plants and fungi are incapable of producing this vitamin, and hence, it is exclusively produced by microorganisms (LeBlanc et al., 2011). It has been demonstrated that cobalamin can be synthesized by some bacteria such as L. reuteri ZJ03, Propionibacterium freudenreichii, B. animalis Bb12 in soy-yogurt, kefir and fermented milk, respectively (Van Wyk et al., 2011; Patel et al., 2013, Gu et al., 2015; Moslemi et al., 2016). Microorganisms can biosynthesize two different isoforms, the vitamin and the pseudovitamin. For example, in a recent work, the production of vitamin and pseudovitamin B12 by P. freudenreichii was quantified specifically and shows that at the initial phase of the fermentation both isoforms are biosynthesized at similar levels; however, by the end of the fermentation the pseudovitamin is not detected, most likely because it is converted to the vitamin form (Deptula et al., 2017). It seems crucial to differentiate between the two isoforms of this vitamin, as the transporter protein in the human GIT has very low affinity for the pseudovitamin, making it un-available to humans (Varmanen et al., 2016).
Biotin (vitamin B7) deficiency can be caused by inadequate dietary intake or some inborn genetic disorders that affect its metabolism. Subclinical deficiency can cause mild symptoms, such as hair thinning or skin rash typically on the face. Biotin can be synthesized by some LAB in dairy products, e.g., L. helveticus MTCC 5463 increased biotin content in fermented milks (Patel et al., 2013). Some propionibacteria can also produce biotin (Hugenholtz et al., 2002).
Vitamin K is an important promoter of bone and cardiovascular health. It has been associated with the inhibition of arterial calcification and stiffening, and the reduction of vascular risk. This vitamin is nearly non-existent in junk food, with little being consumed even in a healthy Western diet (Maresz, 2015). Its deficiency has been implicated in several clinical ailments such as intracranial hemorrhage in newborn infants and possible bone fracture resulting from osteoporosis (LeBlanc et al., 2011). Vitamin K occurs in two forms: firstly, phylloquinone (vitamin K1), which is present in green plants, and secondly, menaquinone (vitamin K2), which is produced by some intestinal bacteria (LeBlanc et al., 2011). Menaquinone can be biosynthesized by some LAB species (mainly Lactococcus lactis) commonly used in industrial fermentation of cheese, buttermilk, sour cream, cottage cheese, and kefir (Walther et al., 2013). Other LAB have been screened for the ability to produce menaquinone; these included strains from the genera Lactococcus, Bifidobacterium, Leuconostoc, and Streptococcus (Morishita et al., 1999). Although the MK forms are ubiquitous in bacteria, it should be noted that some genera such as Lactobacillus have lost the functional ability to produce them (Lechardeur et al., 2011; Walther et al., 2013).
Gamma-aminobutyric acid (GABA) is the main inhibitory neurotransmitter of the central nervous system (CNS). Several important physiological functions of GABA have been characterized, such as neurotransmission, induction of hypotension, diuretic effects, antidiabetic, relaxing and tranquilizer effects (Inoue et al., 2003; Marques et al., 2016). In fact, some GABAA-receptor agonist drugs (e.g., benzodiazepines) are important pharmacological agents used for clinical treatment of anxiety (Foster and Kemp, 2006).
Gamma-aminobutyric acid is biosynthesized through α-decarboxylation of glutamate, an enzymatic conversion which is catalyzed by glutamate decarboxylase (GAD) (Tajabadi et al., 2015). Several food-grade LAB have been reported to exhibit GABA-producing ability. Among them, most of the GABA-producing strains are lactobacilli (L. brevis, L. paracasei, L. delbrueckii, L. buchneri, L. plantarum, L. helveticus), Streptococcus thermophilus, and Lactococcus lactis (Li and Cao, 2010; Dhakal et al., 2012). Some, Bifidobacterium spp. were also reported to produce GABA, although with lower capacity than LAB (Park et al., 2005; Barrett et al., 2012).
Some fermented dairy products enriched in GABA using GABA-producing LAB as starters have been developed. The strains L. casei Shirota, S. salivarius fmb5 and L. plantarum NDC75017 were utilized to ferment and enrich milk in GABA (Inoue et al., 2003; Shan et al., 2015; Chen et al., 2016). More recently, yogurt enriched with 2 mg GABA/ml was produced using the strain S. thermophilus APC151 (Linares et al., 2016a, 2017). Also, fermented soya milk (using L. brevis OPY-1 as source of GABA) (Park and Oh, 2007), or cheese (Lactococcus lactis as source of GABA) (Nomura et al., 1998; Pouliot-Mathieu et al., 2013) have been produced. Thus, GABA has potential as a health-promoting bioactive component in foods (Li and Cao, 2010).
During milk fermentation, LAB, making use of their proteolytic system can transform milk proteins into biologically active peptides. These peptides can exert a wide range of effects, such as antimicrobial, antihypertensive, antithrombotic, immunomodulatory, and antioxidative (LeBlanc et al., 2002; Nongonierma and FitzGerald, 2015). The most studied mechanism of bioactive peptides is the antihypertensive action displayed by the inhibition of the angiotensin-I-converting enzyme (ACE; peptidyldipeptide hydrolase, EC 3.4.15.1) which regulates blood pressure (Fernandez et al., 2015). ACE inhibitory peptides have been isolated from a variety of fermented dairy products including cheese, fermented milks and yogurt (Fitzgerald and Murray, 2006; Pritchard et al., 2010). The best known ACE-inhibitory biopeptides, Val-Pro-Pro (VPP) and Ile-Pro-Pro (IPP), have been identified in milk fermented by L. helveticus (Slattery et al., 2010). In addition, other dairy starter cultures industrially used in the manufacture of fermented dairy products (e.g., L. helveticus, L. delbrueckii ssp. bulgaricus, L. plantarum, L. rhamnosus, L. acidophilus, Lactococcus Lactis, or S. thermophilus) can generate bioactive peptides (Hajirostamloo, 2010; Hafeez et al., 2014). Other ACE-inhibitory peptides such as β-casein f(72-81), Ser-Lys-Val-Tyr-Pro-Phe-Pro-Gly-Pro-Ile (SLVYPFPGPI) have been produced by L. delbrueckii ssp. bulgaricus LB340 in fermented milk (Qian et al., 2011).
On an industrial scale, two fermented milk products with antihypertensive claims, CalpisTM and Evolus®, have been tested extensively in rats and in clinical trials, and are commercialized as functional foods (Dziuba and Dziuba, 2014). Evolus® is available in the market as a L. helveticus fermented milk -produced in Finland- proven to decrease the systolic blood pressure in hypertensive subjects due to the actions of L. helveticus bioactive peptides (EFSA, 2008). CalpisTM is defined as a milk product marketed in Japan (Calpis Co. Ltd.) with antihypertensive properties. It is prepared by fermenting skimmed milk with L. helveticus and Saccharomyces cerevisiae, which produce VPP and IPP peptides from β-casein and κ-casein (Dziuba and Dziuba, 2014).
Bacteriocins are ribosomally synthesized antimicrobial peptides produced by a particular bacterium that are active against other competitor bacteria; thereby they constitute an important part of the microbial defense system (Nes et al., 2007). Such bacteriocin-producing strains may offer potential as an alternative to antibiotics, and may be useful as a means of controlling pathogen carriage, therefore being highly suitable as microbial food additives (Cotter et al., 2013) (Table 2). Bacteriocins from LAB have attracted much interest because they are frequently produced by commercially useful strains that are generally regarded as safe (GRAS) for human consumption (Nes et al., 2007). These antimicrobial molecules are among the beneficial peptides intrinsically synthesized by some LAB during milk fermentation and they have been traditionally used as naturally produced food biopreservatives. In addition, they may function in the GIT as potential natural biotherapeutic agents facilitating the competition of probiotic strains and/or inhibition of pathogens; thereby they are potential contributors to the microbiota balance and human health (Dobson et al., 2012).
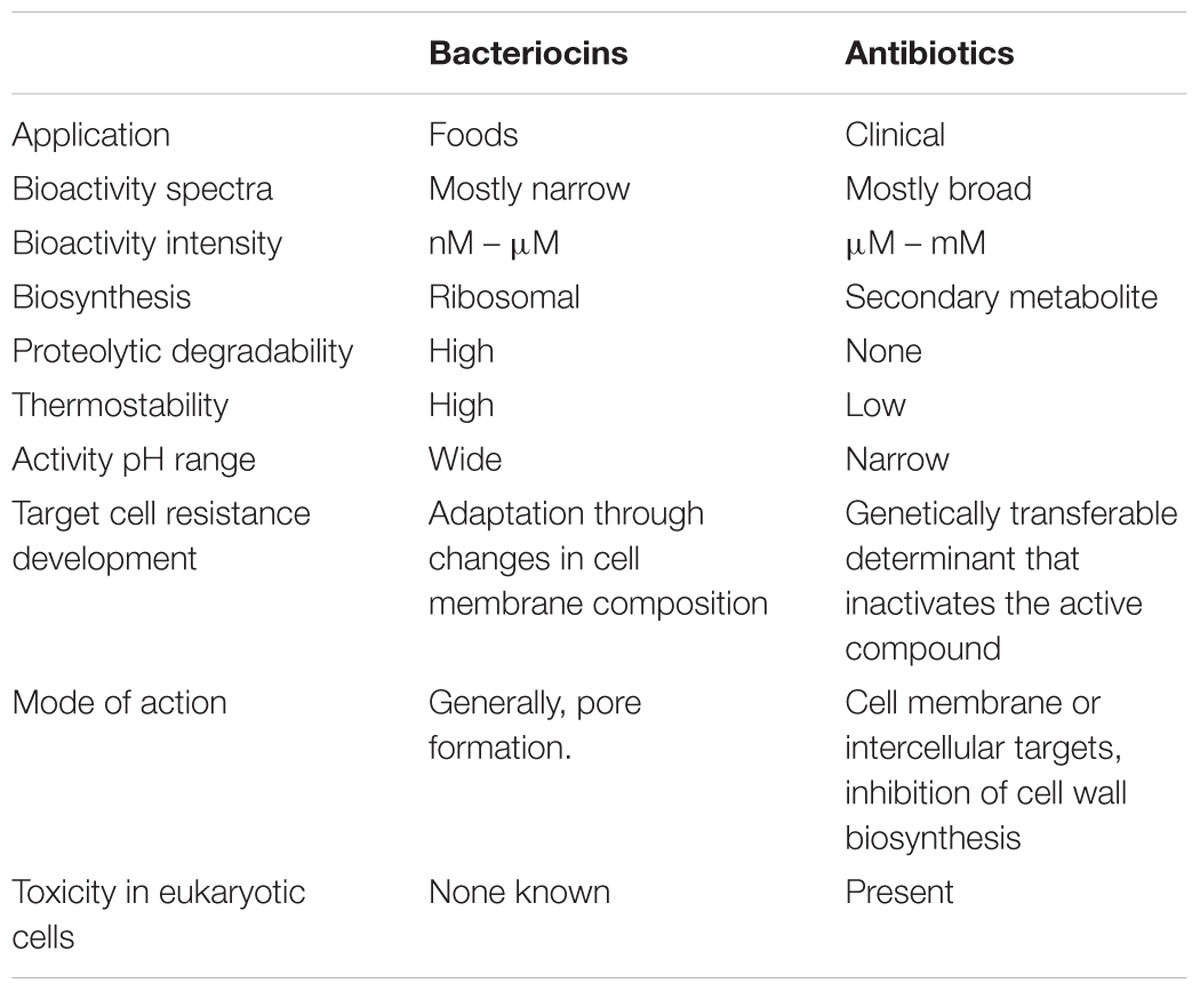
TABLE 2. Characteristic aspects of bacteriocins compared to conventional antibiotics (Adapted from Cleveland et al., 2001).
Nisin is the most well-known bacteriocin used as food preservative due to its antibacterial effect against Listeria, clostridia spores and LAB associated to spoilage. Nisin has been approved as a food additive (E234) in the European Union according to Directive 95/2/EC (EC, 1995) in the following products: semolina and tapioca puddings (3 mg/kg); ripened and processed cheese (12.5 mg/kg), clotted cream (10 mg/kg), and Mascarpone cheese (10 mg/kg). It is also permitted in over 40 countries world-wide including USA, Australia, South Africa, Russia, and India for use as an antimicrobial agent in a variety of food products (EFSA, 2006). Nisin-containing Camembert and semihard cheeses with prolonged shelf-life were made using Lactococcus lactis (strains CNRZ150 or TAB50, respectively) as nisin producers (Arques et al., 2015). Apart from nisin, plantaricins are very well-known bacteriocins. For example, plantaricin C is a broad spectrum bacteriocin produced by L. plantarum, isolated from ripening cheese (Gonzalez et al., 1994). Plantaricins have been reported to produce an immunomodulatory effect on dendritic cells (Meijerink et al., 2010). However, bacteriocins other than nisin have so far only few and limited authorized uses in foods (Yang et al., 2015). Consequently, the use of bacteriocin-producing bacteria as starter culture for in situ biosynthesis during milk fermentation becomes an effective alternative strategy to incorporate bacteriocins in dairy foods. Similarly, the lacticin 3147-producing strain Lactococcus lactis DPC3147 used as a protective culture in Cheddar cheese reduced numbers of Listeria monocytogenes to <10 cfu/g within 5 days at 4°C (Ross et al., 1999; Chen and Hoover, 2006). Other bacterial species such as L. acidophilus can be added as an adjunct in many food fermentation processes to contribute to bacteriocin production and food preservation (Anjum et al., 2014). Other LAB strains such as L. plantarum WHE92 used as adjunct to the starter culture reduced Listeria monocytogenes, Listeria innocua, and Escherichia coli O157:H7 counts in cheese as a consequence of the production of plantaricin (Arques et al., 2015). Using a similar concept, Danisco developed a freeze-dried culture of Pediococcus acidilactici (marketed as CHOOZIT Flav 43) for use as a bacteriocin-producer adjunct in Cheddar and semihard cheeses (Mills et al., 2011).
Studies of the direct impact of dairy foods containing bacteriocins on human health and microbiome are still limited. In vivo antimicrobial activity of nisin and lacticin 3147 has been recently demonstrated in a murine infection model. A nisin-producing Lactococcus lactis CHCC5826 modified the microbiota composition of human microbiota-associated rats increasing bifidobacteria levels and decreasing Enterococcus/Streptococcus populations. Lacticin 3147 has the potential to be employed in the treatment of Clostridium difficile diarrhea and to eliminate the pathogen when added to an anaerobic fecal fermentation (Arques et al., 2015).
Lactic acid bacteria associated to dairy fermentations possess enzymes which can be produced in situ during fermentation of dairy foods and have bioactive potential on the consumer. Examples are hydrolytic enzymes that may exert potential synergistic effects on digestion and alleviate symptoms of intestinal malabsorption (Patel et al., 2013). A well-known example is the β-galactosidase activity, which can achieve lactose degradation and thereby improve health and reduce symptoms of lactose intolerant consumers. Yogurt and other conventional starter cultures and probiotic bacteria in fermented and unfermented milk products improve lactose digestion and alleviate symptoms of intolerance in lactose malabsorbers. These beneficial effects are due to microbial β-galactosidase (de Vrese et al., 2001).
Conjugated linoleic acid (CLA) is a polyunsaturated fatty acid (PUFA) that can be biosynthesized by LAB and bifidobacteria through bioconversion of linoleic acid (LA; cis-9,cis-12 C18:2). The two isomers that have been shown to have bioactive potential are cis-9,trans-11 (c9,t11) and trans-10,cis-12 (t10,c12). The health-promoting properties of CLA include anticarcinogenic, antiatherogenic, anti-inflammatory, and antidiabetic activity, as well as the ability to reduce body fat (Sosa-Castañeda et al., 2015). Although it is a native component of milk, the amount consumed in foods is far from that required in order to obtain desired beneficial effects. Thus, increasing the CLA content in dairy foods through milk fermentation with specific LAB offers a promising alternative. An effective way to increase CLA uptake in humans is to increase CLA levels in dairy products by using strains with high production potential (Lee et al., 2007). A number of food-grade LAB and bifidobacteria were reported to produce CLA in milk products (Sosa-Castañeda et al., 2015; Yang et al., 2015), as is the case of Lactococcus lactis LMG, L. rhamnosus C14, L. casei CRL431, L. acidophilus Lac1, L. plantarum-2, B. bifidum CRL1399 and B. animalis Bb12 (Van Nieuwenhove et al., 2007a; Florence et al., 2009). Some of these strains were also used as adjunct cultures for the manufacture of high CLA-content buffalo cheese (Van Nieuwenhove et al., 2007b). The CLA-producing starter culture of Lactococcus lactis CI4b enhanced levels of total CLA in Cheddar cheese (Mohan et al., 2013). Similarly, L. bulgaricus LB430 and S. thermophilus TA040 are suitable for production of CLA-enriched yogurt (Florence et al., 2009).
In addition, it has been shown that specific microorganisms such as L. plantarum PL60 or B. breve NCIMB702258, are able to produce CLA following dietary administration in animal models (Wall et al., 2009, 2012) and following the administration as a freeze-dried product in humans (Lee and Lee, 2009). Thus, intestinal CLA production by bacteria may contribute to enhance CLA supply in addition to the CLA provided by the producing strain in fermented milks during the manufacture (Teran et al., 2015).
Exopolysaccharides (EPS) are complex extracellular carbohydrate polymers that can be produced by some LAB in situ during dairy fermentations. Some of them promote selective growth of bifidobacteria, thus playing a role on the host microbiota and immune system (Fernandez et al., 2015; Salazar et al., 2016). In this regard, EPS derived from yogurt fermented with L. delbrueckii ssp. bulgaricus OLL1073R-1 exerted immune-stimulatory effects in mice (Makino et al., 2016). Yogurt, Swiss-type, and Cheddar cheeses represent suitable food matrices for the delivery of the hypocholesterolemic EPS-producer strain L. mucosae DPC 6426 (Ryan et al., 2015). Other microorganisms with potential to produce EPS in cheese are P. freudenreichii KG15/KG6, Lactococcus lactis SMQ-461 or S. thermophilus MR-1C (Dabour et al., 2005; Darilmaz and Gumustekin, 2012). Significant levels of EPS can also be produced in kefir by L. plantarum YW11 (Wang et al., 2015). Recently, EPS produced by bifidobacteria have attracted the attention due to their immune modulation capability (Hidalgo-Cantabrana et al., 2012).
Exopolysaccharides can also improve the quality, sensory and rheological properties of the final food product, which in many cases results in a reduction of the amount of chemical stabilizers required, thus favoring a more natural product. For example, strains of B. longum subsp. infantis CCUG 52486 and S. thermophilus were suitable to produce yogurt and fermented ice-cream with improved viscosity and texture and reduced syneresis as a consequence of their high EPS production (Prasannaa et al., 2013; Han et al., 2016; Dertli et al., 2016).
At present, the status of probiotic-based products is full of ambiguities because various regulatory agencies in different countries are defining and categorizing probiotics differently. Despite the emerging interest of consumers toward probiotics and functional foods, in Europe the regulatory framework is still not harmonized and no health claim for probiotics alone (except yogurt starters) has been approved. Paradoxically, probiotics or bioactive bacteria have been introduced into the market as dietary supplements or natural health products (capsules, tablets, and powders) (Arora and Baldi, 2015). Japan was the very first global jurisdiction for implementing a regulatory system for functional foods and nutraceuticals in 1991, and is currently acting as global market leader where probiotics are available as both foods and drugs. The government has designated Foods for Specific Health Uses (FOSHU), which classifies health claims into different subcategories (gastrointestinal health, cholesterol moderation, hypertension moderation, lipid metabolism moderation, sugar absorption moderation, mineral absorption, and bone and tooth health). In China, State Food and Drug Administration (SFDA) has regulated all health foods including functional foods and nutraceuticals, and a well-developed market for functional foods is established (Arora and Baldi, 2015). Currently USA is regulating probiotics as a variety of products as per their intended usage and regulatory bodies are Dietary Supplement Health and Education Act (DSHEA) and Food and Drug administration (FDA). Dietary supplements are considered as ‘foods’ and are regulated by DSHEA and do not need FDA approval before being marketed. However, probiotics and dietary supplements containing a new dietary ingredient without a marketing history are regulated by FDA. In conclusion, a harmonized categorization of probiotics and functional foods may help to regulate these products whenever solid clinical documentation is available to support any health effects and health messages in human subjects. The appropriate level of evidence for determining a health benefit for probiotics should always be put ahead of commercial and labeling industrial interests.
A goal of the dairy industry is to develop novel dairy products with increased nutritional and/or health promoting properties. Food-grade bacteria have the potential to fortify fermented dairy food products with bioactive metabolites by a natural process, thereby reducing the need for fortification with costly chemically synthesized supplements. Nowadays, a number of commercial sources have available synthetic formulations of bioactive substances for their use as a dietary supplement. The use of health-supporting bacteria for naturally enriching dairy foods with bioactives could be a suitable alternative to food fortification with chemical formulations.
The starter cultures must be carefully selected, since the ability of microbial cultures to produce bioactive metabolites is generally a strain-dependent trait and varies considerably among strains within the same species. The yield of bioactive synthesis and the concentration of such compound in dairy products is another critical strain-dependent factor. In this regard, the dose of bioactives ingested with the corresponding food product should remain over the minimum required to meet the human requirements and/or have the claimed therapeutic level on the consumer, according to existing clinical recommendations and studies. An open question when using co-cultures or strain combinations is their interaction in terms of nutrient availability, bacterial growth, as well as the bioactive production yield. In some cases, metabolites (i.e., vitamins etc.) produced by one of the strains could be consumed by the other strains, thus decreasing the final content in food.
Generally, the biosynthetic pathways are genetically encoded. In this regard, the increasing availability of bacterial genome sequences over the last decade has provided a major contribution to the knowledge about microbial production of bioactive molecules. However, the presence of the genes required for the biosynthesis of a particular biomolecule should not be assumed as synonym of metabolite production. Typical exceptions to the correlation genotype-phenotype occur when the genes are not active or when the metabolite is intracellularly biosynthesized and a release system is lacking. This is indeed one of the major bottlenecks during biosynthesis of some vitamins that needs to be overcome through the use of alternative strategies such as autolytic mutants and metabolic engineering (Basavanna and Prapulla, 2013).
Consideration should also be given by manufacturers to the optimum conditions for bioactive compound biosynthesis by LAB during technological processes. The content and activity of a bioactive compound in the dairy fermented foodstuffs is the result of the type of food matrix, the individual bacterial strain properties as well as the processing conditions and storage time. In this regard, it should be noted that the high bioactive biosynthetic rates observed in culture media might not always be extrapolated to dairy products. Therefore, factors such as optimal temperature for microbial growth and viability, food composition or bioactive stability and shelf-life in the final foodstuff are of paramount importance to reach the maximum concentration and activity in the final product.
Overall, the current review updates knowledge about LAB, bifidobacteria and propionibacteria with potential to enrich dairy food products with health-promoting bio-metabolites. Promising applications at commercial level emerge; however, adequate selection of strains is vital to increase the concentration and bioavailability of such biomolecules in fermented foods. The use of LAB and bifidobacteria able to synthesize bioactive components in fermented foods could help to provide these compounds in foods, this being in compliance with current regulatory rules.
DL provided the general concept, and drafted part of the manuscript. CG and ER wrote part of the manuscript. All authors revised and approved the manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
This work was funded by the JPI Food Processing for Health funded by the LONGLIFE Project and the APC Microbiome Institute, a Centre for Science and Technology (CSET) funded by the Science Foundation Ireland (SFI), through the Irish Government’s National Development Plan. The authors are grateful to the University of León (León, Spain) for granting a Ph.D. fellowship to ER.
Ahmed, Z., Wang, Y., Cheng, Q., and Imran, M. (2010). Lactobacillus acidophilus bacteriocin, from production to their application: an overview. Afr. J. Biotechnol. 9, 2843–2850.
Anjum, N., Maqsood, S., Masud, T., Ahmad, A., Sohail, A., and Momin, A. (2014). Lactobacillus acidophilus: characterization of the species and application in food production. Crit. Rev. Food Sci. Nutr. 54, 1241–1251. doi: 10.1080/10408398.2011.621169
Aragon, F., Carino, S., Perdigón, G., and Moreno-LeBlanc, A. (2014). The administration of milk fermented by the probiotic Lactobacillus casei CRL 431 exerts an immunomodulatory effect against a breast tumour in a mouse model. Immunobiology 219, 457–464. doi: 10.1016/j.imbio.2014.02.005
Aragon-Alegro, L. C., Alarcon-Alegro, J. H., Cardarelli, H. R., Chiu, M. C., and Saad, S. M. (2007). Potentially probiotic and synbiotic chocolate mousse. LWT Food Sci. Technol. 40, 669–675. doi: 10.1016/j.lwt.2006.02.020
Araujo, E. A., dos Santos Pires, A. C., Pinto, M. S., Jan, G., and Carvalho, A. F. (2012). Probiotics in Dairy Fermented Products. Rijeja: Intech, 129–148.
Arora, M., and Baldi, A. (2015). Regulatory categories of probiotics across the globe: a review representing existing and recommended categorization. Ind. J. Med. Microbiol. 33, 2–10. doi: 10.4103/0255-0857.150868
Arques, J. L., Rodríguez, E., Langa, S., Landete, J. M., and Medina, M. (2015). Antimicrobial activity of lactic acid bacteria in dairy products and gut: effect on pathogens. Biomed. Res. Int. 2015:584183. doi: 10.1155/2015/584183
Baglatzi, L., Gavrili, S., Stamouli, K., Zachaki, S., Favre, L., Pecquet, S., et al. (2016). Effect of infant formula containing a low dose of the probiotic Bifidobacterium lactis CNCM I-3446 on immune and gut functions in C-section delivered babies: A pilot study. Clin. Med. Insights Pediatr 10, 11–19. doi: 10.4137/CMPed.S33096
Barrett, E., Ross, R. P., O’Toole, P. W., Fitzgerald, G. F., and Stanton, C. (2012). γ-Aminobutyric acid production by culturable bacteria from the human intestine. J. Appl. Microbiol. 113, 411–417. doi: 10.1111/j.1365-2672.2012.05344.x
Basavanna, G., and Prapulla, S. G. (2013). Evaluation of functional aspects of Lactobacillus fermentum CFR 2195 isolated from breast fed healthy infants’ fecal matter. J. Food Sci. Technol. 50, 360–366. doi: 10.1007/s13197-011-0345-9
Castro, J. M., Tornadijo, M. E., Fresno, J. M., and Sandoval, H. (2015). Biocheese: a food probiotic carrier. Biomed. Res. Int 2015, 723056. doi: 10.1155/2015/723056
Chen, H., and Hoover, D. G. (2006). Bacteriocins and their food applications. Compr. Rev. Food Sci. Food Saf. 2, 82–100.
Chen, L., Zhao, H., Zhang, C., Lu, Y., Zhu, X., and Lu, Z. (2016). γ-Aminobutyric acid-rich yogurt fermented by Streptococcus salivarius subsp. thermophiles fmb5 appears to have anti-diabetic effect on streptozotocin-induced diabetic mice. J. Func. Foods 20, 267–275. doi: 10.1016/j.jff.2015.10.030
Cleveland, J., Montville, T. J., Nes, I. F., and Chikindas, M. L. (2001). Bacteriocins: safe, natural antimicrobials for food preservation. Int. J. Food Microbiol. 71, 1–20. doi: 10.1016/S0168-1605(01)00560-8
Correa, S. B., Castro, I., and Saad, S. M. (2008). Probiotic potential and sensory properties of coconut flan supplemented with Lactobacillus paracasei and Bifidobacterium lactis. Int. J. Food Sci. Technol. 43, 1560–1568. doi: 10.1111/j.1365-2621.2007.01585.x
Cotter, P. D., Ross, R. P., and Hill, C. (2013). Bacteriocins: a viable alternative to antibiotics? Nat. Rev. Microbiol. 11, 95–105. doi: 10.1038/nrmicro2937
Crittenden, R. G., Martinez, N. R., and Playne, M. J. (2003). Synthesis and utilisation of folate by yoghurt starter cultures and probiotic bacteria. Int. J. Food Microbiol. 80, 217–222. doi: 10.1016/S0168-1605(02)00170-8
Dabour, N., Kheadr, E. E., Fliss, I., and LaPointe, G. (2005). Impact of ropy and capsular exopolysaccharide-producing strains of Lactococcus lactis subsp. cremoris on reduced-fat Cheddar cheese production and whey composition. Int. Dairy J. 15, 459–471. doi: 10.1016/j.idairyj.2004.08.011
D’Aimmo, M. R., Mattarelli, P., Biavati, B., Carlsson, N. G., and Andlid, T. (2012). The potential of bifidobacteria as a source of natural folate. J. Appl. Microbiol. 112, 975–984. doi: 10.1111/j.1365-2672.2012.05261.x
Darilmaz, D. O., and Gumustekin, Y. (2012). Research on some factors influencing acid and exopolysaccharide produced by dairy Propionibacterium strains isolated from traditional homemade Turkish cheeses. J. Food Prot. 75, 918–926. doi: 10.4315/0362-028X.JFP-11-510
Davidson, R. H., Duncan, S. E., Hackney, C. R., Eigel, W. N., and Boling, J. W. (2000). Probiotic culture survival and implications in fermented frozen yogurt characteristics. J. Dairy Sci. 83, 666–673. doi: 10.3168/jds.S0022-0302(00)74927-7
de Vrese, M., Stegelmann, A., Richter, B., Fenselau, S., Laue, C., and Schrezenmeir, J. (2001). Probiotics: compensation for lactase insufficiency. Am. J. Clin. Nutr. 73, 421S–429S.
Deptula, P., Chamlagain, B., Edelmann, M., Sangsuwan, P., Nyman, T. A., Savijoki, K., et al. (2017). Food-like growth conditions support production of active vitamin B12 by Propionibacterium freudenreichii 2067 without DMBI, the lower ligand base, or cobalt supplementation. Front. Microbiol. 8:368. doi: 10.3389/fmicb.2017.00368
Dertli, E., Toker, O. S., Durak, M. Z., Yilmaz, M. T., Tatlısu, N. B., Sagdic, O., et al. (2016). Development of a fermented ice-cream as influenced by in situ exopolysaccharide production: rheological, molecular, microstructural and sensory characterization. Carbohydr. Polym. 136, 427–440. doi: 10.1016/j.carbpol.2015.08.047
Dhakal, R., Bajpai, V. K., and Baek, K. H. (2012). Production of gaba (γ-Aminobutyric acid) by microorganisms: a review. Braz. J. Microbiol. 43, 1230–1241. doi: 10.1590/S1517-83822012000400001
Divya, J. B., and Nampoothiri, K. M. (2015). Encapsulated Lactococcus lactis with enhanced gastrointestinal survival for the development of folate enriched functional foods. Bioresour. Technol. 188, 226–230. doi: 10.1016/j.biortech.2015.01.073
Dobson, A., Cotter, P. D., Ross, R. P., and Hill, C. (2012). Bacteriocin production: a probiotic trait? Appl. Environ. Microbiol. 78, 1–6. doi: 10.1128/AEM.05576-11
Drywien, M., Frąckiewicz, J., Górnicka, M., Gadek, J., and Jałosiñska, M. (2015). Effect of probiotic and storage time of thiamine and riboflavin content in the milk drinks fermented by Lactobacillus casei KNE-1. Rocz. Panstw. Zakl. Hig. 66, 373–377.
Dziuba, B., and Dziuba, M. (2014). Milk proteins-derived bioactive peptides in dairy products: molecular, biological and methodological aspects. Acta Sci. Pol. Technol. Aliment. 13, 5–25. doi: 10.17306/J.AFS.2014.1.1
EFSA (2006). Opinion of the scientific panel on food additives, flavourings, processing aids and materials in contact with food on a request from the commission related to the use of nisin (E 234) as a food additive: question number EFSA-Q-2005–2031. EFSA J. 314, 1–16.
EFSA (2008). Evolus and reduce arterial stiffness - Scientific substantiation of a health claim related to Lactobacillus helveticus fermented Evolus® low-fat milk products and reduction of arterial stiffness pursuant to article 14 of the regulation (EC) No 1924/2006 - Scientific opinion of the panel on dietetic products, nutrition and allergies; question number EFSA-Q-2008-218. EFSA J. 824, 1–12.
EFSA (2011). Scientific Opinion on the substantiation of a health claim related to “Lactobacillus plantarum TENSIATM in the semihard Edam-type “heart cheese” of HarmonyTM” and maintenance of normal blood pressure pursuant to Article 13(5) of Regulation (EC) No 1924/20061. EFSA panel on dietetic products, nutrition and allergies (NDA). EFSA J. 9, 1981. doi: 10.2903/j.efsa.2011.1981
FAO/WHO (2001). Report of the Joint FAO/WHO Expert Consultation on Evaluation of Health and Nutritional Properties of Probiotics in Food Including Powder Milk with Live Lactic Acid Bacteria. Rome: Food and Agriculture Organization of the United Nations.
Farnworth, E. R., and Champagne, C. P. (2015). “Production of probiotic cultures and their incorporation into foods,” in Probiotics, Prebiotics, and Synbiotics: Bioactive Foods in Health Promotion, Vol. 20, eds R. R. Watson and V. R. Preedy (Amsterdam: Elsevier), 303–318.
Fernandez, M., Hudson, J. A., Korpela, R., and de los Reyes-Gavilán, C. G. (2015). Impact on human health of microorganisms present in fermented dairy products: an overview. Biomed. Res. Int. 2015:412714. doi: 10.1155/2015/412714
Fitzgerald, R. J., and Murray, B. A. (2006). Bioactive peptides and lactic fermentations. Int. J. Dairy Technol. 59, 118–125. doi: 10.1111/j.1471-0307.2006.00250.x
Florence, A., Silva, R., Santo, A., Gioielli, L., Tamime, A., and Oliveira, M. (2009). Increased CLA content in organic milk fermented by Bifidobacteria or yoghurt cultures. Dairy Sci. Technol. 89, 541–553. doi: 10.1051/dst/2009030
Foster, A. C., and Kemp, J. A. (2006). Glutamate- and GABA-based CNS therapeutics. Curr. Opin. Pharmacol. 6, 7–17. doi: 10.1016/j.coph.2005.11.005
Ghadimi, D., Folster-Holst, R., De Vrese, M., Winkler, P., Heller, K. J., and Schrezenmeir, J. (2008). Effects of probiotic bacteria and their genomic DNA on TH1/TH2-cytokine production by peripheral blood mononuclear cells (PBMCs) of healthy and allergic subjects. Immunobiology 213, 677–692. doi: 10.1016/j.imbio.2008.02.001
Giraffa, G. (2012). Selection and design of lactic acid bacteria probiotic cultures. Eng. Life Sci. 12, 391–398. doi: 10.1002/elsc.201100118
Gobbetti, M., Cagno, R. D., and De Angelis, M. (2010). Functional microorganisms for functional food quality. Crit. Rev. Food Sci. Nutr. 50, 716–727. doi: 10.1080/10408398.2010.499770
Gonzalez, B., Arca, P., Mayo, B., and Suarez, J. E. (1994). Detection, purification, and partial characterization of plantaricin C, a bacteriocin produced by a Lactobacillus plantarum strain of dairy origin. Appl. Environm. Microbiol. 60, 2158–2163.
Gortzi, O., Tsakali, E., Chatzilazarou, A., and Galidi, A. (2015). “E-food science project: bio-functional foods,” in Proceedings of the 2nd International Conference on Food and Biosystems Engineering, Vol. 2, Myconos.
Gu, Q., Zhang, C., Song, D., Li, P., and Zhu, X. (2015). Enhancing vitamin B12 content in soy-yogurt by Lactobacillus reuteri. Int. J. Food Microbiol. 206, 56–59. doi: 10.1016/j.ijfoodmicro.2015.04.033
Hafeez, Z., Cakir-Kiefer, C., Roux, E., Perrin, C., Miclo, L., and Dary-Mourot, A. (2014). Strategies of producing bioactive peptides from milk proteins to functionalize fermented milk products. Rev. Article Food Res. Int. 63(Part A), 71–80. doi: 10.1016/j.foodres.2014.06.002
Hajirostamloo, B. (2010). Bioactive component in milk and dairy product. Int. Sci. Index Agric. Biosyst. Eng. 4, 870–874.
Han, X., Yang, Z., Jing, X., Yu, P., Zhang, Y., Yi, H., et al. (2016). Improvement of the texture of yogurt by use of exopolysaccharide producing lactic acid bacteria. Biomed. Res. Int. 2016:7945675. doi: 10.1155/2016/7945675
Hidalgo-Cantabrana, C., López, P., Gueimonde, M., de Los Reyes-Gavilán, C. G., Suárez, A., Margolles, A., et al. (2012). Immune modulation capability of exopolysaccharides synthesised by lactic acid bacteria and bifidobacteria. Probiotics Antimicrob. Proteins 4, 227–237. doi: 10.1007/s12602-012-9110-2
Hugenholtz, J., Hunik, J., Santos, H., and Smid, E. (2002). Nutraceutical production by propionibacteria. Lait 82, 103–112. doi: 10.1051/lait:2001009
Inoue, K., Shirai, T., Ochiai, H., Kasao, M., Hayakawa, K., Kimura, M., et al. (2003). Blood-pressure-lowering effect of a novel fermented milk containing Ɣ-aminobutyric acid (GABA) in mild hypertensives. Eur. J. Clin. Nutr. 57, 490–495. doi: 10.1038/sj.ejcn.1601555
Jones, M. L., Martoni, C. J., Parent, M., and Prakash, S. (2012). Cholesterol-lowering efficacy of a microencapsulated bile salt hydrolase-active Lactobacillus reuteri NCIMB 30242 yoghurt formulation in hypercholesterolaemic adults. Br. J. Nutr. 107, 1505–1513. doi: 10.1017/S0007114511004703
Joshi, V. K. (2015). Indigenous Fermented Foods of South Asia. Fermented Foods and Beverages Series. Boca Raton, FL: CRC Press.
Juarez del Valle, M., Laiño, J. E., de Moreno de LeBlanc, A., Savoy de Giori, G., and LeBlanc, J. G. (2016). Soyamilk fermented with riboflavin-producing Lactobacillus plantarum CRL 2130 reverts and prevents ariboflavinosis in murine models. Br. J. Nutr. 116, 1229–1235. doi: 10.1017/S0007114516003378
Karimi, R., Mortazavian, A. M., and Amiri-Rigi, A. (2012). Selective enumeration of probiotic microorganisms in cheese. Food Microbiol. 29, 1–9. doi: 10.1016/j.fm.2011.08.008
Karimi, R., Mortazavian, A. M., and Gomes Da Cruz, A. (2011). Viability of probiotic microorganisms in cheese during production and storage: a review. Dairy Sci. Technol. 91, 283–308. doi: 10.1007/s13594-011-0005-x
Laiño, J. E., Juarez del Valle, M., Savoy de Giori, G., and LeBlanc, J. G. (2013). Development of a high folate concentration yogurt naturally bio-enriched using selected lactic acid bacteria. LWT Food Sci. Technol. 54, 1–5. doi: 10.1016/j.lwt.2013.05.035
Laiño, J. E., Juarez del Valle, M., Savoy de Giori, G., and LeBlanc, J. G. (2014). Applicability of a Lactobacillus amylovorus strain as co-culture for natural folate bio-enrichment of fermented milk. Int. J. Food Microbiol. 19, 10–16. doi: 10.1016/j.ijfoodmicro.2014.08.031
LeBlanc, J. G., Laiño, J. E., del Valle, M. J., Vannini, V., van Sinderen, D., Taranto, M. P., et al. (2011). B-group vitamin production by lactic acid bacteria–current knowledge and potential applications. J. Appl. Microbiol. 111, 1297–1309. doi: 10.1111/j.1365-2672.2011.05157.x
LeBlanc, J. G., Matar, C., Valdéz, J. C., LeBlanc, J., and Perdigon, G. (2002). Immunomodulating effects of peptidic fractions issued from milk fermented with Lactobacillus helveticus. J. Dairy Sci. 85, 2733–2742. doi: 10.3168/jds.S0022-0302(02)74360-9
Lechardeur, D., Cesselin, B., Fernandez, A., Lamberet, G., Garrigues, C., Pedersen, M., et al. (2011). Using heme as an energy boost for lactic acid bacteria. Curr. Opin. Biotechnol. 22, 143–149. doi: 10.1016/j.copbio.2010.12.001
Lee, K., and Lee, Y. (2009). Production of c9,t11- and t10,c12-conjugated linoleic acids in humans by Lactobacillus rhamnosus PL60. J. Microbiol. Biotechnol. 19, 1617–1619. doi: 10.4014/jmb.0907.07010
Lee, K., Paek, K., Lee, H. Y., Park, J. H., and Lee, Y. (2007). Antiobesity effect of trans-10,cis-12-conjugated linoleic acid-producing Lactobacillus plantarum PL62 on diet-induced obese mice. J. Appl. Microbiol. 103, 1140–1146. doi: 10.1111/j.1365-2672.2007.03336.x
Levit, R., Savoy de Giori, G., de Moreno de LeBlanc, A., and LeBlanc, J. G. (2016). Evaluation of the effect of soymilk fermented by a riboflavin-producing Lactobacillus plantarum strain in a murine model of colitis. Benef. Microbes 8, 65–72. doi: 10.3920/BM2016.0063
Li, H., and Cao, Y. (2010). Lactic acid bacterial cell factories for gamma-aminobutyric acid. Amino Acids 39, 1107–1116. doi: 10.1007/s00726-010-0582-7
Lin, M. Y., and Young, C. M. (2000). Folate levels in cultures of lactic acid bacteria. Int. Dairy J. 10, 409–413. doi: 10.1016/S0958-6946(00)00056-X
Linares, D. M., Arboleya, S., Ross, R. P., and Stanton, C. (2017). Complete genome sequence of the gamma-aminobutyric acid-producing strain Streptococcus thermophilus APC151. Genome Announc. 5, e00205-17. doi: 10.1128/genomeA.00205-17
Linares, D. M., O’Callaghan, T. F., O’Connor, P. M., Ross, R. P., and Stanton, C. (2016a). Streptococcus thermophilus APC151 strain is suitable for the manufacture of naturally GABA-enriched bioactive yoghurt. Front. Microbiol. 7:1876.
Linares, D. M., Ross, P., and Stanton, C. (2016b). Beneficial microbes: the pharmacy in the gut. Bioengineered 7, 11–20. doi: 10.1080/21655979.2015.1126015
Lopez, M., Li, N., Kataria, J., Russell, M., and Neu, J. (2008). Live and ultraviolet-inactivated Lactobacillus rhamnosus GG decrease flagellin-induced interleukin-8 production in Caco-2 cells. J.Nutr. 138, 2264–2268. doi: 10.3945/jn.108.093658
Makino, S., Sato, A., Goto, A., Nakamura, M., Ogawa, M., Chiba, Y., et al. (2016). Enhanced natural killer cell activation by exopolysaccharides derived from yogurt fermented with Lactobacillus delbrueckii ssp. bulgaricus OLL1073R-1. J. Dairy Sci. 99, 915–923. doi: 10.3168/jds.2015-10376
Maresz, K. (2015). Proper calcium use: vitamin K2 as a promoter of bone and cardiovascular health. Integr. Med. 14, 34–39.
Marques, T. M., Patterson, E., Wall, R., O’Sullivan, O., Fitzgerald, G. F., Cotter, P. D., et al. (2016). Influence of GABA and GABA-producing Lactobacillus brevis DPC 6108 on the development of diabetes in a streptozotocin rat model. Benef. Microbes 7, 409–420. doi: 10.3920/BM2015.0154
Martinovic, A., Brede, M. E., Vegarud, G. E., Østlie, H. M., Narvhus, J., and Skeie, S. B. (2016). Survival of lactic acid and propionibacteria in low- and full-fat Dutch-type cheese during human digestion ex vivo. Lett. Appl. Microbiol. 62, 404–410. doi: 10.1111/lam.12561
Meijerink, M., van Hemert, S., Taverne, N., Wels, M., de Vos, P., Bron, P. A., et al. (2010). Identification of genetic loci in Lactobacillus plantarum that modulate the immune response of dendritic cells using comparative genome hybridization. PLoS ONE 5:e10632. doi: 10.1371/journal.pone.0010632
Mills, S., Stanton, C., Hill, C., and Ross, R. P. (2011). New developments and applications of bacteriocins and peptides in foods. Ann. Rev. Food Sci. Technol. 2, 299–329. doi: 10.1146/annurev-food-022510-133721
Mohan, M. S., Anand, S., Kalscheur, K. F., Hassan, A. N., and Hippen, A. R. (2013). Starter cultures and cattle feed manipulation enhances conjugated linoleic acid concentrations in Cheddar cheese. J. Dairy Sci. 96, 2081–2094. doi: 10.3168/jds.2012-6101
Morishita, T., Tamura, N., Makino, T., and Kudo, S. (1999). Production of menaquinones by lactic acid bacteria. J. Dairy Sci. 82, 1897–1903. doi: 10.3168/jds.S0022-0302(99)75424-X
Moslemi, M., Mazaheri-Nezhad-Fard, R., Hosseini, S. M., Homayouni-Rad, A., and Mortazavian, A. M. (2016). Incorporation of propionibacteria in fermented milks as a probiotic. Crit. Rev. Food Sci. Nutr. 56, 1290–1312. doi: 10.1080/10408398.2013.766584
Nes, I. F., Yoon, S. S., and Diep, D. B. (2007). Ribosomally synthesized antimicrobial peptides (bacteriocins) in lactic acid bacteria: a review. Food Sci. Biotechnol. 16, 675–690.
Nomura, M., Kimoto, H., Someya, Y., Furukawa, S., and Suzuki, I. (1998). Production of gamma-aminobutyric acid by cheese starters during cheese ripening. J. Dairy Sci. 81, 1486–1491. doi: 10.3168/jds.S0022-0302(98)75714-5
Nongonierma, A. B., and FitzGerald, R. J. (2015). The scientific evidence for the role of milk protein-derived bioactive peptides in humans: a review. J. Funct. Foods 17, 640–656. doi: 10.1016/j.jff.2015.06.021
Oh, N. S., Joung, J. Y., Lee, J. Y., Kim, S. H., and Kim, Y. (2016). Characterization of the microbial diversity and chemical composition of Gouda cheese made by potential probiotic strains as an adjunct starter culture. J. Agric. Food Chem. 64, 7357–7366. doi: 10.1021/acs.jafc.6b02689
Park, K. B., Ji, G. E., Park, M. S., and Oh, S. H. (2005). Expression of rice glutamate decarboxylase in Bifidobacterium longum enhances gamma-aminobutyric acid production. Biotechnol. Lett. 27, 1681–1684. doi: 10.1007/s10529-005-2730-9
Park, K. B., and Oh, S. H. (2007). Production of yoghurt with enhanced levels of gamma-aminobutyric acid and valuable nutrients using lactic acid bacteria and germinated soybean extract. Bioresour. Technol. 98, 1675–1679. doi: 10.1016/j.biortech.2006.06.006
Patel, A., Shah, N., and Prajapati, J. B. (2013). Biosynthesis of vitamins and enzymes in fermented foods by lactic acid bacteria and related genera - A promising approach. Croat. J. Food Sci. Technol. 5, 85–91.
Pouliot-Mathieu, K., Gardner-Fortier, C., Lemieux, S., St-Gelais, D., Champagne, C. P., and Vuillemard, J. C. (2013). Effect of cheese containing gamma-aminobutyric acid-producing lactic acid bacteria on blood pressure in men. PharmaNutrition 1, 141–148. doi: 10.1016/j.phanu.2013.06.003
Prado, M. R., Blandón, L. M., Vandenberghe, L. P., Rodrigues, C., Castro, G. R., Thomaz-Soccol, V., et al. (2015). Milk kefir: composition, microbial cultures, biological activities, and related products. Front. Microbiol. 6:1177. doi: 10.3389/fmicb.2015.01177
Prasannaa, P. H., Grandisona, A. S., and Charalampopoulosa, D. (2013). Microbiological, chemical and rheological properties of low fat set yoghurt produced with exopolysaccharide (EPS) producing Bifidobacterium strains. Food Res. Int. 5, 15–22. doi: 10.1016/j.foodres.2012.11.016
Pritchard, S. R., Phillips, M., and Kailasapathy, K. (2010). Identification of bioactive peptides in commercial Cheddar cheese. Food Res. Int. 43, 1545–1548. doi: 10.1016/j.foodres.2010.03.007
Qian, B., Xing, M., Cui, L., Deng, Y., Xu, Y., Huang, M., et al. (2011). Antioxidant, antihypertensive, and immunomodulatory activities of peptide fractions from fermented skim milk with Lactobacillus delbrueckii ssp. bulgaricus LB340. J. Dairy Res. 78, 72–79. doi: 10.1017/S0022029910000889
Ross, R. P., Galvin, M., McAuliffe, O., Morgan, S. M., Ryan, M. P., Twomey, D. P., et al. (1999). Developing applications for lactococcal bacteriocins. Antonie Van Leeuwenhoek 76, 337–346. doi: 10.1023/A:1002069416067
Rotar, M. A., Semeniuc, C., Apostu, S., Suharoschi, R., Mureşan, C., Modoran, C., et al. (2007). Researches concerning microbiological evolution of lactic acid bacteria to yoghurt storage during shelf-life. J. Agroalimentary Proc. Technol. 13, 135–138.
Russo, P., Capozzi, V., Arena, M. P., Spadaccino, G., Dueñas, M. T., López, P., et al. (2014). Riboflavin-overproducing strains of Lactobacillus fermentum for riboflavin-enriched bread. Appl. Microbiol. Biotechnol. 98, 3691–3700. doi: 10.1007/s00253-013-5484-7
Ryan, P. M., Guinane, C. M., London, L. E., Kelleher, P. R., Fitzgerald, G. F., Caplice, N. M., et al. (2015). Genome sequence of the heteropolysaccharide-producing strain Lactobacillus mucosae DPC 6426. Genome Announc. 3:e01350-14. doi: 10.1128/genomeA.01350-14
Salazar, N., Gueimonde, M., de Los Reyes-Gavilán, C. G., and Ruas-Madiedo, P. (2016). Exopolysaccharides produced by lactic acid bacteria and bifidobacteria as fermentable substrates by the intestinal microbiota. Crit. Rev. Food Sci. Nutr. 56, 1440–1453. doi: 10.1080/10408398.2013.770728
Saubade, F., Hemery, Y. M., Guyot, J. P., and Humblot, C. (2016). Lactic acid fermentation as a tool for increasing the folate content of foods. Crit. Rev. Food Sci. Nutr. doi: 10.1080/10408398.2016.1192986 [Epub ahead of print].
Shan, Y., Man, C. X., Han, X., Li, L., Guo, Y., Deng, Y., et al. (2015). Evaluation of improved GABA production in yoghurt using Lactobacillus plantarum NDC75017. J. Dairy Sci. 98, 2138–2149. doi: 10.3168/jds.2014-8698
Siro, I., Kápolna, E., Kápolna, B., and Lugasi, A. (2008). Functional food. Product development, marketing and consumer acceptance: a review. Appetite 51, 456–467. doi: 10.1016/j.appet.2008.05.060
Slattery, L., O’Callaghan, J., Fitzgerald, G. F., Beresford, T., and Ross, R. P. (2010). Invited review: Lactobacillus helveticus: a thermophilic dairy starter related to gut bacteria. J. Dairy Sci. 93, 4435–4454. doi: 10.3168/jds.2010-3327
Sosa-Castañeda, J., Hernández-Mendoza, A., Astiazarán-García, H., Garcia, H. S., Estrada-Montoya, M. C., González-Córdova, A. F., et al. (2015). Screening of Lactobacillus strains for their ability to produce conjugated linoleic acid in milk and to adhere to the intestinal tract. J. Dairy Sci. 98, 6651–6659. doi: 10.3168/jds.2014-8515
Stanton, C., Ross, R. P., Fitzgerald, G. F., and Van Sinderen, D. (2005). Fermented functional foods based on probiotics and their biogenic metabolites. Curr. Opin. Biotechnol. 16, 198–203. doi: 10.1016/j.copbio.2005.02.008
Tajabadi, N., Baradaran, A., Ebrahimpour, A., Rahim, R. A., Bakar, F. A., Manap, M. Y., et al. (2015). Overexpression and optimization of glutamate decarboxylase in Lactobacillus plantarum Taj-Apis362 for high gamma-aminobutyric acid production. Microb. Biotechnol. 8, 623–632. doi: 10.1111/1751-7915.12254
Tamime, A. (ed.). (2006). Fermented Milks. Ayr: Blackwell Publishing Ltd. doi: 10.1002/9780470995501
Tamime, A. Y., Saarela, M., Korslund Sødergaard, A., Mistry, V. V., and Shah, N. P. (2005). “Production and maintenance of viability of probiotic micro-organisms in dairy products,” in Probiotic Dairy Products, ed. A. Y. Tamime (Oxford: Blackwell Publishing Ltd), 39–72.
Teran, V., Luna-Pizarro, P., Zacarías, M. F., Vinderola, G., Medina, R., and Van Nieuwenhove, C. (2015). Production of conjugated dienoic and trienoic fatty acids by lactic acid bacteria and bifidobacteria. J. Funct. Foods 19(Part A), 417–425. doi: 10.1016/j.jff.2015.09.046
Van Nieuwenhove, C. P., Oliszewski, R., Gonzalez, S. N., and Perez-Chaia, A. B. (2007a). Conjugated linoleic acid conversion by dairy bacteria cultured in MRS broth and buffalo milk. Lett. Appl. Microbiol. 44, 467–474.
Van Nieuwenhove, C. P., Oliszewski, R., Gonzalez, S. N., and Perez-Chaia, A. B. (2007b). Influence of bacteria used as adjunct culture and sunflower oil addition on conjugated linoleic acid content in buffalo cheese. Food Res. Int. 40, 559–564. doi: 10.1016/j.foodres.2006.08.003
Van Wyk, J., Witthuhn, R. C., and Britz, T. J. (2011). Optimisation of vitamin B12 and folate production by Propionibacterium freudenreichii strains in kefir. Int. Dairy J. 21, 69–74. doi: 10.1016/j.idairyj.2010.09.004
Vanderhoof, J. A., and Young, R. J. (1998). Use of probiotics in childhood gastrointestinal disorders. J. Pediatr. Gastroenterol. Nutr. 27, 323–332. doi: 10.1097/00005176-199809000-00011
Varmanen, P., Deptula, P., Chamlagain, B., and Piironen, V. (2016). Letter to the editor on ‘Enhancing vitamin B12 content in soy-yogurt by Lactobacillus reuteri, IJFM. 206:56-59’. Int. J. Food Microbiol. 228, 33. doi: 10.1016/j.ijfoodmicro.2016.03.029
Wall, R., Marques, T. M., O’Sullivan, O., Ross, R. P., Shanahan, F., Quigley, E. M., et al. (2012). Contrasting effects of Bifidobacterium breve NCIMB 702258 and Bifidobacterium breve DPC 6330 on the composition of murine brain fatty acids and gut microbiota. Am. J. Clin. Nutr. 95, 1278–1287. doi: 10.3945/ajcn.111.026435
Wall, R., Ross, R. P., Shanahan, F., O’Mahony, L., O’Mahony, C., Coakley, M., et al. (2009). Metabolic activity of the enteric microbiota influences the fatty acid composition of murine and porcine liver and adipose tissues. Am. J. Clin. Nutr. 89, 1393–1401. doi: 10.3945/ajcn.2008.27023
Walther, B., Karl, J. P., Booth, S. L., and Boyaval, P. (2013). Menaquinones, bacteria, and the food supply: the relevance of dairy and fermented food products to vitamin K requirements. Adv. Nutr. 4, 463–473. doi: 10.3945/an.113.003855
Wang, J., Zhao, X., Tian, Z., Yang, Y., and Yang, Z. (2015). Characterization of an exopolysaccharide produced by Lactobacillus plantarum YW11 isolated from Tibet Kefir. Carbohydr. Polym. 125, 16–25. doi: 10.1016/j.carbpol.2015.03.003
Watson, R. R., and Preedy, V. R. (2015). Probiotics, Prebiotics and Symbiotics: Bioactive Foods in Health Promotion, 1st Edn. Amsterdam: Elsevier.
Keywords: lactic acid bacteria, bifidobacteria, health, bioactive, probiotic, biofunctional food
Citation: Linares DM, Gómez C, Renes E, Fresno JM, Tornadijo ME, Ross RP and Stanton C (2017) Lactic Acid Bacteria and Bifidobacteria with Potential to Design Natural Biofunctional Health-Promoting Dairy Foods. Front. Microbiol. 8:846. doi: 10.3389/fmicb.2017.00846
Received: 14 December 2016; Accepted: 25 April 2017;
Published: 18 May 2017.
Edited by:
Joaquin Bautista-Gallego, Instituto de la Grasa (CSIC), SpainReviewed by:
Clara G. De Los Reyes-Gavilan, Consejo Superior de Investigaciones Científicas, SpainCopyright © 2017 Linares, Gómez, Renes, Fresno, Tornadijo, Ross and Stanton. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Catherine Stanton, Y2F0aGVyaW5lLnN0YW50b25AdGVhZ2FzYy5pZQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.