
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Vet. Sci. , 12 March 2025
Sec. Parasitology
Volume 12 - 2025 | https://doi.org/10.3389/fvets.2025.1566720
 Michaela Komáromyová1
Michaela Komáromyová1 Daniel Petrič2Klára Demčáková2,3Matej Leško2,3
Daniel Petrič2Klára Demčáková2,3Matej Leško2,3 Klaudia Čobanová2Michal Babják1Alžbeta Königová1
Klaudia Čobanová2Michal Babják1Alžbeta Königová1 Tetiana Kuzmina1,4
Tetiana Kuzmina1,4 Sylwester Ślusarczyk5
Sylwester Ślusarczyk5 Paulina Izabela Fortuna6
Paulina Izabela Fortuna6 Anna Łukomska7Pola Sidoruk8
Anna Łukomska7Pola Sidoruk8 Adam Cieslak8
Adam Cieslak8 Zora Váradyová2*
Zora Váradyová2* Marián Várady1*
Marián Várady1*Bioactive plants provide therapeutic and prophylactic effects to ruminants. We determined the effect of grazing on natural meadow grassland enriched with experimentally sown chicory (Cichorium intybus) on parasitological status, pasture larval infectivity, antioxidant parameters, and the histology of abomasal tissue in lambs experimentally infected with the parasitic gastrointestinal nematode (GIN) Haemonchus contortus. We also qualitatively identified the main polyphenols in the meadow grassland and phenolic metabolites in the feces of the lambs. Sixteen lambs were orally infected with approximately 5,000 infective larvae (L3) of H. contortus. The lambs were divided into two groups: lambs grazing on a plot consisting exclusively of meadow pasture which serves as control group and lambs grazing on a plot where approximately 25% of a meadow grassland was reclaimed with chicory. The experimental period was 144 days. The number of eggs per gram (EPG) of feces was quantified on D21, D34, D48, D62, D76, D89, D103, D118, D131, and D144 post-infection. Pasture contamination with H. contortus L3 was examined. EPG in both groups of lambs was highest at D34. Egg shedding was significantly lower in both groups from D48 onwards, with a reduction of >95% from D103 onwards. Pasture contamination with L3 was highest at D41 but was then significantly lower in both groups. The total antioxidant capacity, the activity of glutathione peroxidase and the concentration of malondialdehyde in the serum changed significantly during the experiment (p < 0.003, < 0.001, and < 0.016, respectively). At least 54 species of meadow plants were identified on both pasture plots; plant bioactive compounds identified were mainly phenolic acids, flavonoids, and glucosides. Phenolic metabolites (e.g., coumaric acid, chicory acid, salvigenin, and esters of gallic acid) were identified in the feces of the lambs. In some lambs, the morphological observation identified small histopathological changes in the abomasal tissues typical of hemonchosis. Both the natural meadow pasture and the pasture enriched with experimentally sown chicory slowed the dynamics of GIN infection and pasture contamination with L3 by mobilizing the antioxidant defensive system and gradually increasing the resistance of the infected lambs, probably due to the beneficial effects of plant bioactive substances.
Gastrointestinal parasites of ruminants represent a wide spectrum of genera and species; gastrointestinal nematodes (GINs) are of the greatest economic importance in Europe (1). Most herds/flocks in grass-based production systems are infected by GINs, and their main economic impact is due to sub-clinical infections that reduce animal growth and milk/wool production. Haemonchus contortus, a highly pathogenic nematode is considered the most important of GIN infections and is responsible for economic losses due to losses in productivity, high mortality, anthelmintic resistance, and cost of treatment (2–5). Antiparasitic drugs play a key role in combatting helminth and protozoan parasitic diseases in both production and pet animals worldwide (6). Effective anthelmintic therapy, however, has several risks and is considered unsustainable and non-ecological mainly due to the accumulation of drug residues in animal tissues and milk and to the development of resistance in parasitic nematodes to all commonly available anthelmintics (7–9). The goal of most methods for controlling parasites is not to eliminate the parasites, but to maintain the population at a level that does not affect the overall health of the host population (10).
The development of new preventive and therapeutic strategies against resistance to synthetic anthelmintics has increased the interest in plant medicines and other alternative methods of parasite control (11–13). Much work has focused on using plants with bioactive compounds in ruminants infected with GINs for their nutritional effects and anthelmintic activities (14–16). The diversity and synergy of bioactive compounds of several medicinal plants and their combinations, described in our previous studies (17, 18), contributed to some pharmacological efficacy against H. contortus. We therefore concluded that using treatments with bioactive plants against H. contortus could directly affect the dynamics of parasitic infection and indirectly mobilize the antioxidant defensive system and antibody or immune response in sheep, thus improving animal resistance (19, 20). Supplementation with only one medicinal bioactive plant (e.g., Artemisia absinthium or Malva sylvestris) also increased the resistance of lambs to H. contortus infection (21, 22). The antiparasitic properties of plant tannins and flavonoids were previously confirmed by several studies, indicating that including plants from the family Fabaceae (≥70% dry matter) in the feed could reduce the parasitic load in the abomasa of sheep infected with H. contortus, Teladorsagia circumcincta, and/or Ostertagia ostertagi and could decrease the number of eggs excreted in feces (23–25). GIN infections may also induce the production of reactive oxygen species, which may damage the parasites but generate oxidative stress (26).
Chicory (Cichorium intybus) is a perennial plant from the family Asteraceae widespread on the European continent that has been used as a feed additive for farm animals due to its high feed value and various therapeutic effects (27, 28). The anthelmintic effect of chicory and its secondary metabolites against GINs have been described in several studies (29–31). Chicory produces natural antioxidants that can be used in animal nutrition to protect from potentially harmful oxidative damage. We hypothesized that the grazing of sheep/lambs on a meadow with different plant species and on a meadow enriched with experimentally sown chicory may influence the level of lamb infection with GINs by reducing the number of parasite eggs in feces and adult worms in the abomasum and improving antioxidant status, which provides better defense against oxidative stress in infected lambs. Haemonchus contortus larvae developing within abomasal glands can cause major damage to the abomasal tissue, accompanied by the dilatation of, and damage to, the glands, hyperplasia in mucosal cells, and the loss of chief and parietal cells, so we were also concerned about the histopathological examination of the abomasa. Our goals were to investigate the effect of grazing on a natural meadow grassland enriched with experimentally sown chicory on (1) parasitological status, (2) pasture larval contamination, (3) antioxidative parameters in sera, and (4) the histopathology of the abomasa of lambs infected with H. contortus. We also qualitatively identified the main polyphenols in the meadow grassland and the phenolic metabolites in the feces.
This study was conducted following the guidelines of the Declaration of Helsinki and National legislation in the Slovak Republic (G.R. 377/2012; Law 39/2007) for the care and use of research animals. The Ethical Committee of the Institute of Parasitology of the Slovak Academy of Sciences approved the experimental protocol on 20 January 2023. The experiment was carried out on pastures of a private farm (PETLAMB) in Petrovce, district of Prešov, Slovakia.
Two meadow grassland plots (0.43 ha each) with mixed plant species were fenced with electric fences; each plot had an automated water trough and a sheepfold. An area approximately 25% of the size of the experimental plot (CHIC) was recultivated with a mixture of three plants: 81.5% chicory (Cichorium intybus), 14.8% sainfoin (Onobrychis viciifolia), and 3.7% alfalfa (Medicago sativa). The control plot (CON) consisted exclusively of meadow grassland with mixed plant species. The plants of both plots (CON and CHIC) were identified based on plant atlas keys (32, 33). The land where these plots were established had never been grazed and was thus considered virtually worm-free.
The experiment was conducted between May and October. Sixteen male and female (1:1) Tsigai breed lambs aged 3–4 months with an average weight of 13.6 ± 0.52 kg from a sheep farm (PD Ružín–farm Ružín, Kysak, Slovakia) were transferred to the PETLAMB private farm. All lambs were dewormed with the recommended dose of albendazole (5 mg/kg body weight; Albendavet 1.9% suspension a.u.v, DIVASA-FARMAVIC S.A., Barcelona, Spain) during a 7-day period of adaptation before the start of the experiment. The animals were evenly divided into two groups of eight lambs based on their live weights and genders: the control group (CON) grazed the control plot, and the chicory group (CHIC) grazed the plot recultivated with chicory. Each animal was fed 300 g dry matter (DM) Mikrop ČOJ, a commercial concentrate (MIKROP, Čebín, Czech Republic) during the entire period of the study. Each lamb was experimentally infected with 5,000 third-stage larvae (L3) of the MHCo1 strain of H. contortus susceptible to anthelmintics (34); the day of infection was considered day 0 (D0) of the experiment. Both groups of lambs grazed the meadow grassland plots with mixed plant species for the first 33 days; the CHIC group started to graze the enriched pasture on D34. All animals were humanely slaughtered at the end of the trial (D144) following the rules of the European Commission (35) for slaughtering procedures.
Fecal samples were collected rectally from each lamb on D21, D34, D48, D62, D76, D89, D103, D118, D131, and D144 post-infection. Part of these feces was stored at −80°C for subsequent analysis of polyphenols. The number of H. contortus eggs per gram (EPG) of feces was quantified using the modified McMaster method described by Coles et al. (36).
The percent fecal egg count reduction (%FECR) in the CON and CHIC groups was calculated using the formula (37):
where Ti1 is EPG on D34, and Ti2 is EPG on the following sampling days (D48, D62, D76, D89, D103, D118, D131, and D144) in a host from a total of n hosts.
The humanely slaughtered animals were necropsied on D144. The gastrointestinal tract of each lamb was examined to count the total number of adult H. contortus in the abomasum. The abomasum of each animal was removed and dissected, and the contents were washed with warm physiological saline and emptied into a jar. The contents were mixed continuously to prevent the clustering of nematodes. Washings were brought up to a volume of two liters with physiological saline. Three 40-mL aliquots (5%) were collected from each animal and fixed with helminthological iodine. The number of adult H. contortus was counted for each animal.
To estimate pasture larval contamination and the survival of L3 over time, we collected grass samples from the experimental (CHIC) and control (CON) plots over 5 months on D0, D41, D78, D103, and D139. Up to 30 grass samples (5–10 g each) were cut from each plot 0–10 cm from the soil surface. We randomly collected grass samples on a “Z-shape trace” twice on each plot, cutting samples every 10–12 m. We also collected grass samples 0–20 cm from the edge of sheep feces in the area around the sheepfold. Samples from the CON and CHIC plots were collected and examined separately.
Infective larvae of H. contortus were collected from the grass samples using the Baermann procedure. Twenty grams of grass subsamples were cut manually by scissors and placed in a Baermann funnel with tap water for 10–12 h (overnight); we used 6 to 12 Baermann funnels for each CON and CHIC plot. L3 were collected from the bottom of the funnel, stained with iodine to separate the L3 from free-living soil nematodes, and counted under a microscope with 40× magnification.
The pasture larval contamination was expressed as the number of L3 per kilogram of dry grass matter (L3/kg DM) using the formula:
where nav is the average number of L3 in 20 g of grass subsample.
Blood samples for sera were collected from each animal on D0, D21, D34, D62, D89, and D131 and placed directly into 10-mL serum separator tubes (Sarstedt AG & Co, Nümbrecht, Germany) on the farm and delivered to the laboratory. The samples were centrifuged at 1200 g for 10 min at room temperature to separate the serum; the sera samples were stored at −80°C until analysis. The total antioxidant capacity (TAC), the activity of glutathione peroxidase (GPx), and the concentration of malondialdehyde (MDA) in the sera were determined as previously described (38).
Samples of plant species from the meadow grassland plots were collected from 10 random quadrats (0.2 × 0.2 m each) of land grazed by the CON and CHIC groups from May to September. The polyphenols were analyzed as previously described (28).
The frozen feces were freeze-dried using a Gamma 2–16 LSC freeze dryer (Christ, Osterode, Germany), ground with a ZM 200 mill (Retsch, Düsseldorf, Germany) equipped with a 1 mm sieve, and stored in the dark for subsequent use and analysis. Three random samples, each 100 mg, were extracted three times with 80% MeOH for 30 min at 40°C (ultrasonic bath). The extract was evaporated to dryness and then dissolved in 2 mL of Milli-Q water (acidified with 0.2% formic acid) and purified by Solid Phase Extraction using an Oasis HLB 3 cc Vac Cartridge, 60 mg (Waters Corp., Milford, USA). The cartridge was washed with 0.5% methanol to remove carbohydrates and then washed with 80% methanol to elute phenolics, then re-evaporated to dryness and dissolved in 1 mL of 80% methanol (acidified with 0.2% formic acid). The sample was then centrifuged (23,000 g, 5 min) before spectrometric analysis. All analyses were performed in triplicate for three independent samples and stored at −20°C before analysis. Polyphenol composition was estimated using liquid chromatography electrospray ionization (ESI) quadrupole time-of-flight (QTOF) mass spectrometry on a Thermo Dionex Ultimate 3,000 RS chromatographic system (Thermo Fisher Scientific, Waltham, USA) coupled to a Bruker Compact QTOF mass spectrometer (Bruker, Billerica, USA), consisting of a binary pump system, sample manager, column manager, and photodiode array detector. Separations were performed on a 2.1 × 100 mm, 2.6 μm Kinetex C18 column (Phenomenex, Torrance, USA), with mobile phase A consisting of 0.1% (v/v) formic acid in water and mobile phase B consisting of 0.1% (v/v) formic acid in acetonitrile. A linear gradient from 5 to 60% phase B in phase A over 20 min was used to separate phenolic compounds. The flow rate was 0.3 mL/min, and the column was held at 30°C. Spectra were acquired in negative-ion mode over a mass range from m/z 100 to 1,500 with a frequency of 5 Hz. The operating parameters of the ESI ion source were: capillary voltage, 3 kV; dry-gas flow, 6 L/min; dry-gas temperature, 200°C; nebulizer pressure, 0.7 bar; collision radio frequency, 700.0 V; transfer time, 100.0 μs; and pre-pulse storage, 7.0 μs. Ultrapure nitrogen was used as the drying and nebulizer gasses, and argon was used as the collision gas. The collision energy was set automatically from 15 to 75 eV depending on the m/z of the fragmented ion. The data acquired were calibrated internally, with sodium formate introduced to the ion source at the beginning of each separation via a 20-μL loop.
The spectra were processed using Bruker DataAnalysis 4.3 software (Bruker Daltonics GmbH, Bremen, Germany), which provides a ranking based on the best fit of measured and theoretical isotopic patterns within a specific window of mass accuracy. The quality of the isotopic fit was expressed using the mSigma-value. SmartFormula3D matched peaks were sent to the MetFrag website in silico fragmentation for the computer-assisted identification of metabolite mass spectra. We also used a few databases to search for the structural identity of the metabolites: the human metabolome database,1 the BiGG database,2 the PubChem database,3 the MassBank database,4 KEGG,5 and the Metlin database,6 supported with appropriate literature information (39–42).
Abomasal tissues were histologically examined similarly to the procedure previously described (43). Samples of fresh tissues from the abomasum of each lamb were washed in a phosphate buffer (0.1 M, pH 7.4), put in plastic containers, and fixed in a 10% buffered formalin solution as pieces of tissue spread on flat polystyrene. The fixed material was processed using a series of reagents and embedded in Paraplast PLUS paraffin blocks (Leica, Buffalo Grove, USA), which were then cut with a rotary microtome into sections 3.5 μm thick. Slides with a paraffin section were automatically stained with hematoxylin and eosin (Varistain Gemini Thermo Scientific, Runcorn, UK). An Axio Lab. A1 microscope (Carl Zeiss, Jena, Germany) equipped with a Zeiss Axiocam ERc5s digital camera was used for the histological examination. Photographs were analyzed and recorded using ZEN 2.3 (blue edition) software (Carl Zeiss Microscopy GmbH, 2011).
The data were statistically analyzed using GraphPad Prism (9.2.0 (332) 2021; GraphPad Software, Inc., San Diego, USA). Parasitological data were analyzed using an unpaired t-test. Data for the antioxidant parameters of the blood sera were analyzed using two-way analyses of variance. The model included treatment, time, and the interaction between treatment and time. Results were considered significant at p < 0.05.
We first noticed a reduction in egg shedding in the feces of both groups of lambs on D48 (Figure 1). %FECR confirmed reductions of 63.94% (p < 0.004) and 40.58% (p < 0.037) of egg number in the CON and CHIC groups, respectively (Table 1). The egg shedding reductions (94.48–99.77%) were highest from D103 onwards in both groups. The mean number of abomasal worms recorded per lamb after the necropsy was lower in the CON (59.33) than the CHIC (88.63) group, but these data were not significant (p > 0.05).
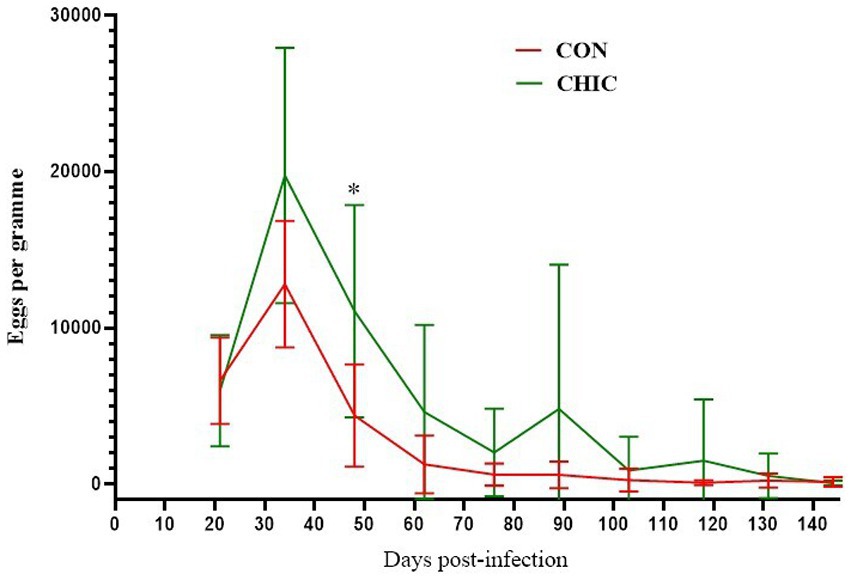
Figure 1. Mean fecal egg counts ± SD for the control (CON) and chicory (CHIC) groups of lambs infected with Haemonchus contortus.
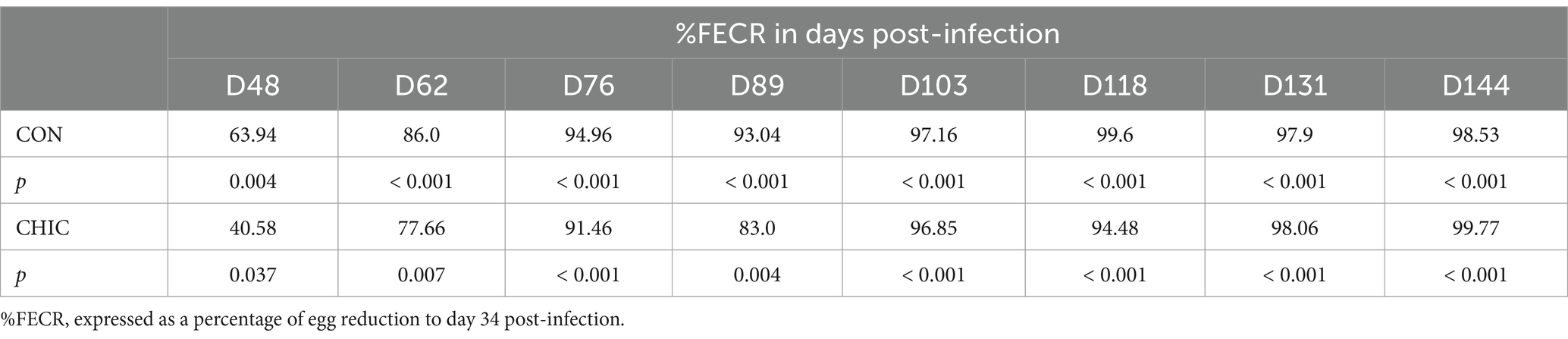
Table 1. Percent fecal egg count reduction (%FECR) of the control (CON) and chicory (CHIC) groups of lambs (n = 8).
The pasture larval count was highest at D41 in both groups of lambs (Table 2); the number of L3 on “random” grass was 20.9-fold higher on the CON (2,820 L3/kg DM) than the CHIC (135 L3/kg) plot. The number of H. contortus larvae on the grass samples collected near feces at D41 was similar on both plots. Subsequent sampling identified significantly reduced pasture larval contamination on both plots and on D139 larvae were not found on the grass of either plot. Haemonchus contortus was the only nematode species found in the pasture grass samples.
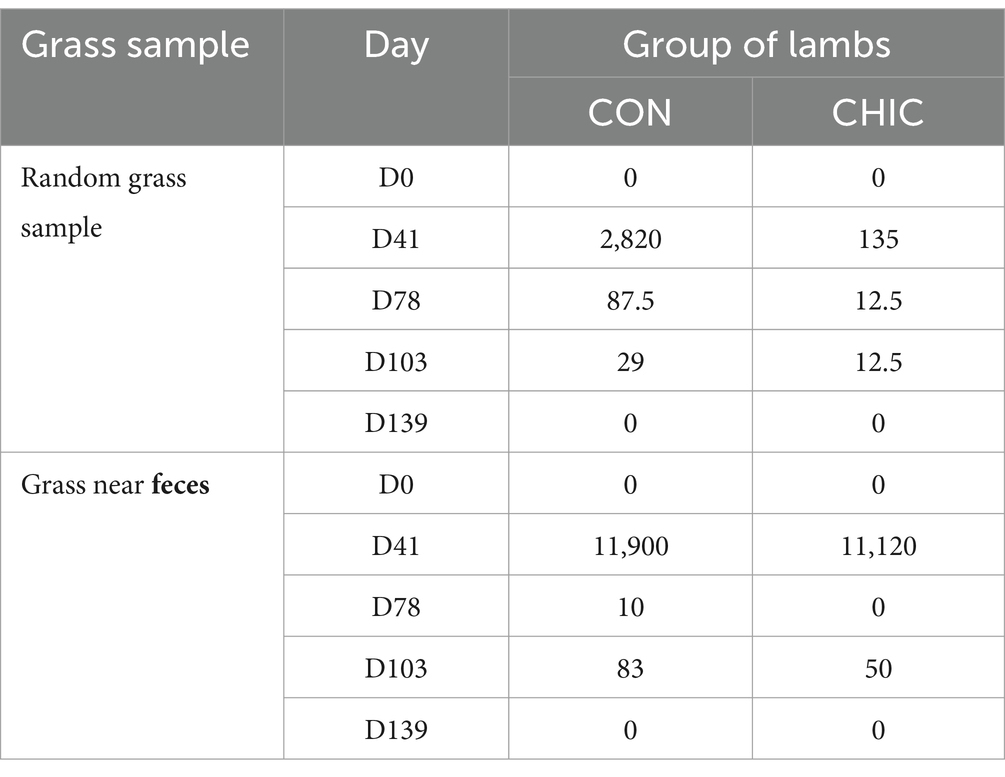
Table 2. Pasture larval contamination expressed as the number of infective third-stage larvae (L3) per kilogram of dry grass matter (L3/kg DM) in the control (CON) and chicory (CHIC) groups of lambs.
TAC, lipid peroxidation (i.e., MDA concentration), and GPx activity in the blood serum changed significantly during the experiment (p < 0.003, < 0.001, and < 0.016, respectively) (Table 3). GPx activity continuously decreased throughout the experiment in both the CON and CHIC groups. TAC was lower on D21 and D34, and lipid peroxidation was higher on D21, D34, and D62, in both the CON and CHIC groups. TAC and MDA concentrations on D131 were similar to their initial values at the beginning of the experiment (D0) for both groups.
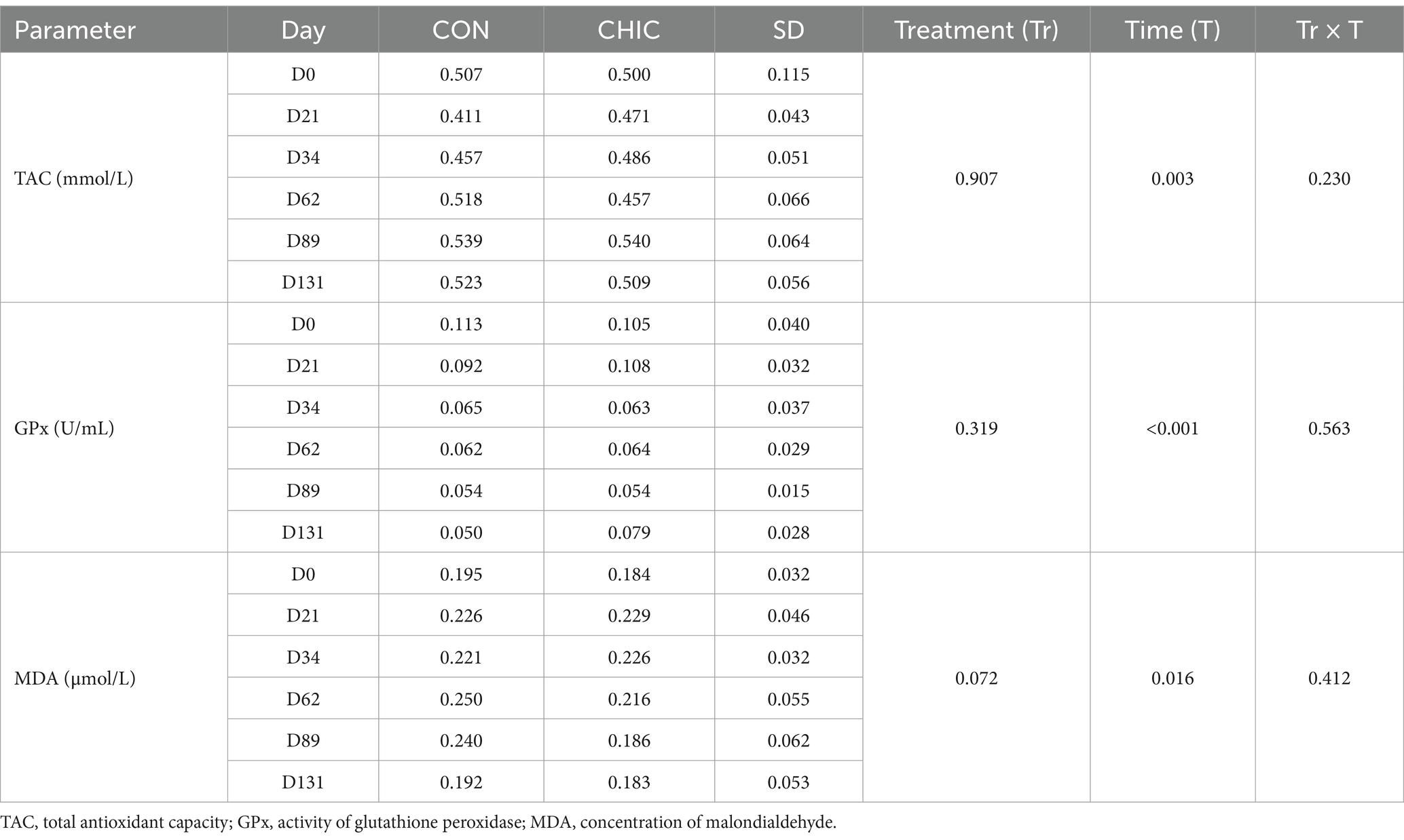
Table 3. Antioxidant parameters in the blood sera of the control (CON) and chicory (CHIC) groups of lambs (n = 8).
The diversity of the meadow plant species in the samples collected from the plots grazed by the CON and CHIC groups from May to September was high; at least 54 species from 20 families were collected and identified (Table 4). The analyses of plant bioactive compounds identified mainly phenolic acids, flavonoids, and glucosides in the samples from the CON and CHIC plots.
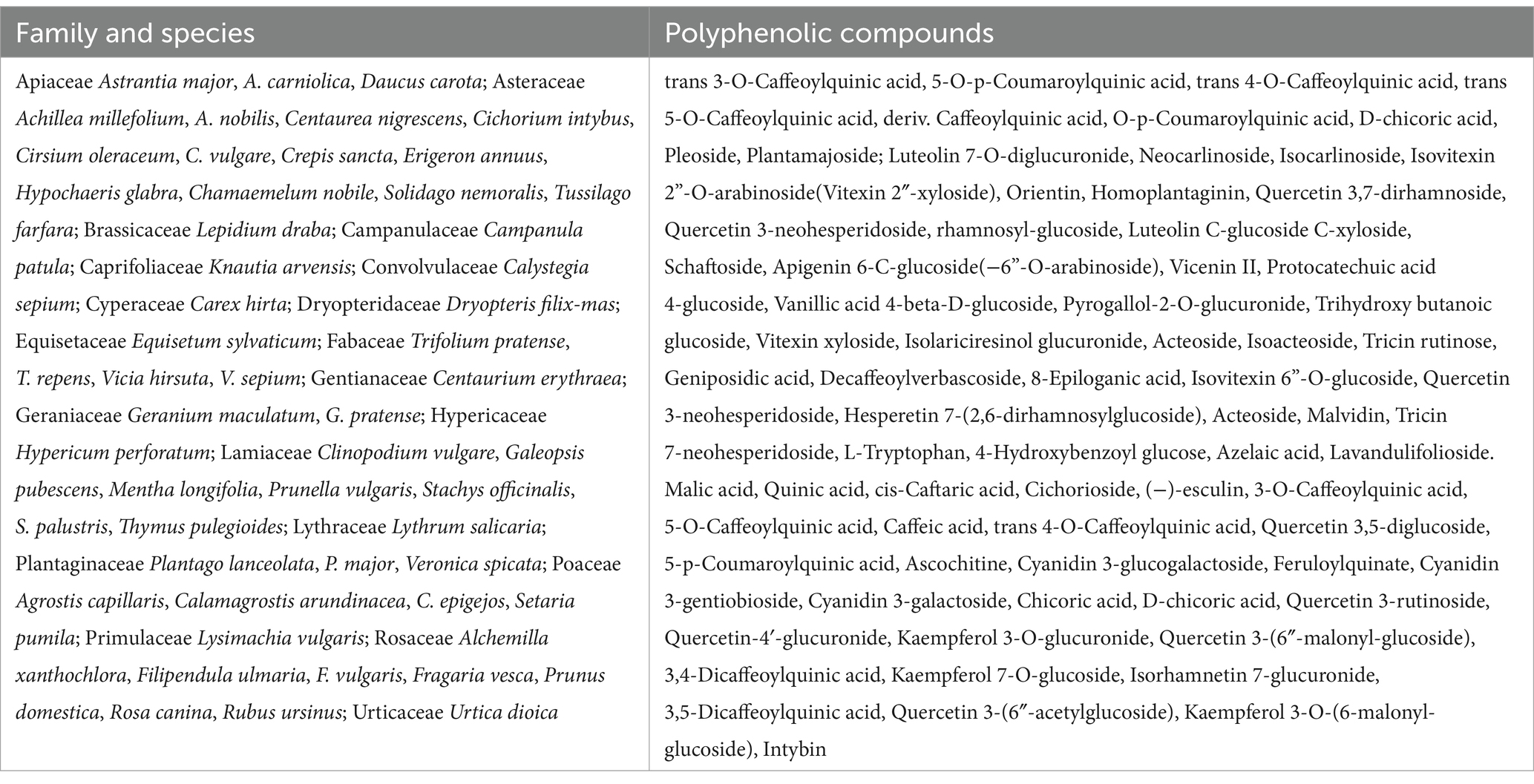
Table 4. Plant species and main polyphenolic compounds in the pasture plots from May to September.
Qualitative analyses of phenolic metabolites in the feces of the lambs from the CON and CHIC groups identified 21 and 28 phenolic metabolites, respectively (Figures 2, 3). The peak numbers in Figures 2, 3 represent the list of the main phenolic metabolites in the feces of the lambs from both groups.
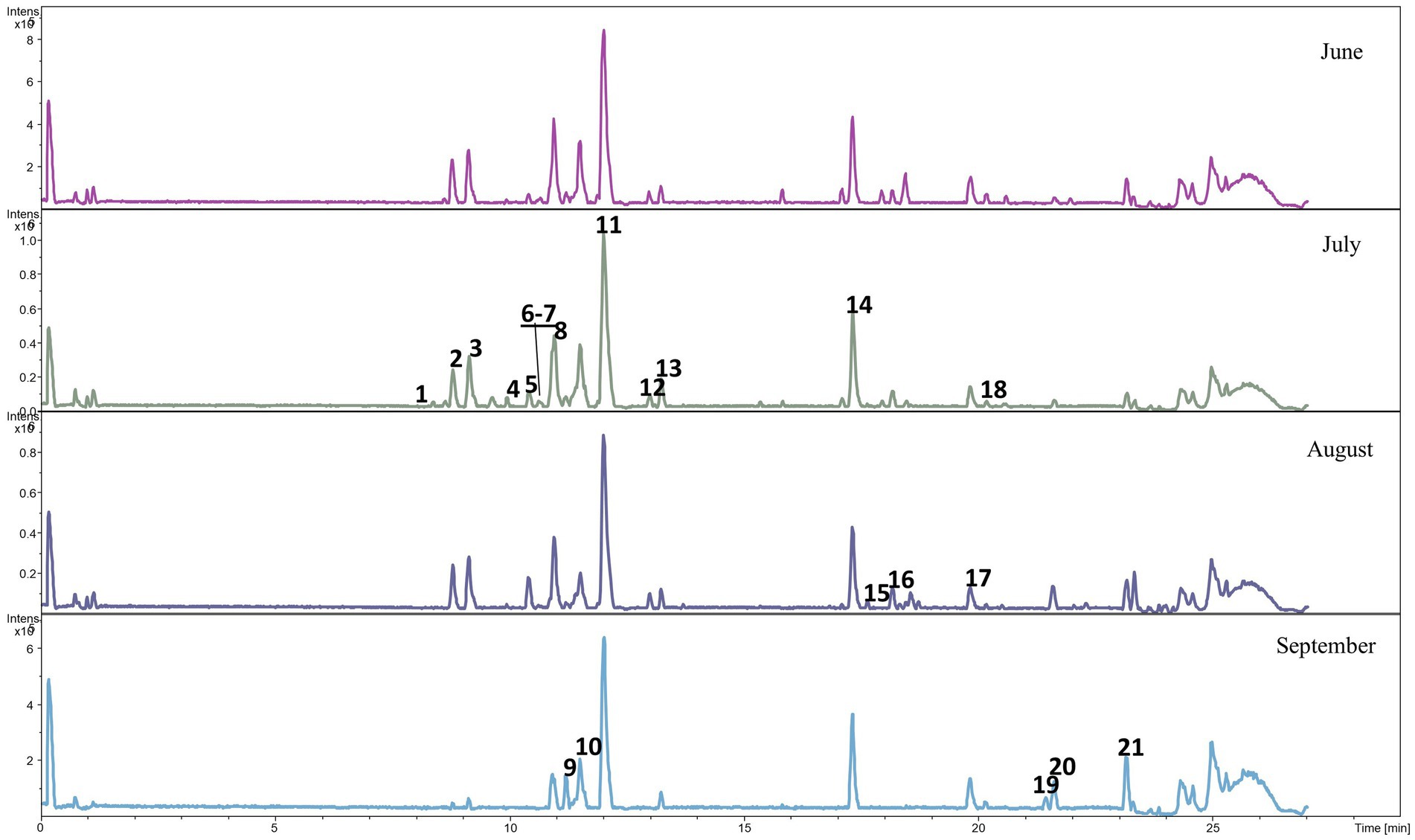
Figure 2. Base peak chromatogram of phenolic metabolites in the feces of lambs from the control group. The peak numbers represent the main phenolic metabolites as follows: 1 cumarinic acid, 2 tridec-3-enedioic acid, 3 D-chicoric acid, 4 dihydrophaseic acid, 5 3′,4′,5′-trimethoxy flavonol, 6 azaleic acid, 7 salvigenin, 8 2′,3-dihydroxy-4,4′,6′-trimethoxychalcone, 9 7-hydroxy-2′,3′,4′-trimethoxyisoflavan, 10 2′,3-dihydroxy-4,4′,6′-trimethoxychalcone (C), 11 2′,3-dihydroxy-4,4′,6′-trimethoxychalcone (diapocynin), 12 4-hydroxyequol, 13 uralenneoside, 14 9-hydroxy-hexadecane-1,16-dioate, 15 dodecanedioic acid, 16 epi-lipoxin, 17 2,3-dihydroxy-5-undecylcyclohexa-2,5-diene-1,4-dione, 18 tetradecanedioic acid, 19 3,7-dihydroxycholan-24-oic acid, 20 7alpha-hydroxy-3-oxo-5beta-cholan-24-oic acid, and 21 isodeoxycholic acid.
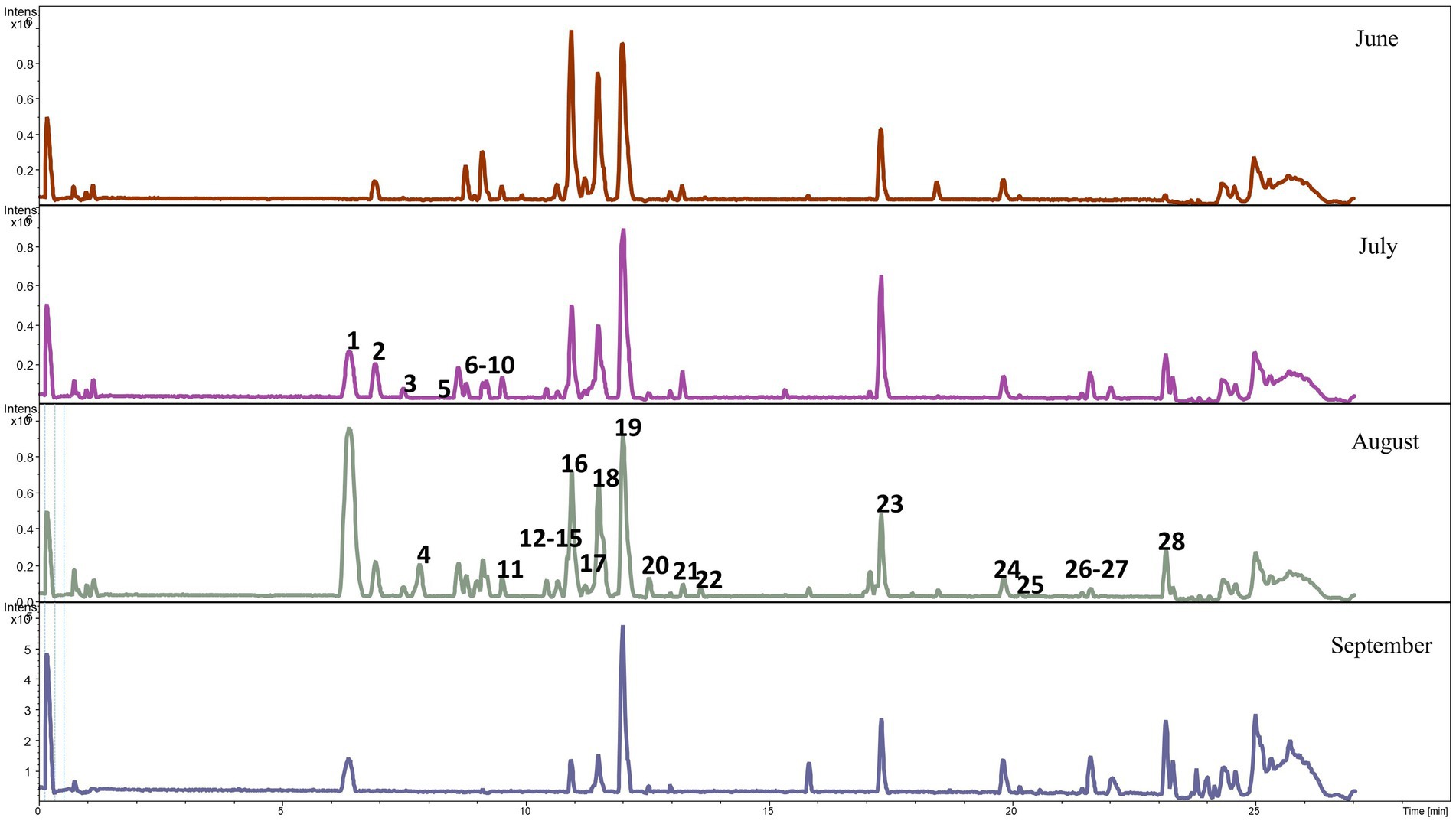
Figure 3. Base peak chromatogram of phenolic metabolites in the feces of lambs from the chicory group. The peak numbers represent the main phenolic metabolites as follows: 1 Not determined, 2 gallic acid octyl ester, 3 gallic acid octyl ester, 4 ND, 5 cumarinic acid, 6 (S)-2,3-dihydro-7-hydroxy-2-methyl-4-oxo-4H-1-benzopyran-5-acetic acid, 7 ND, 8 3-methoxyphenylacetic acid, 9 tridec-3-enedioic acid, 10 pentadeca-4,7,10-trienedioic acid, 11 D-chicoric acid, 12 ND, 13 2′,7-dihydroxy-4′,6-dimethoxyisoflavan, 14 3′,4′,5′-trimethoxyflavonol, 15 salvigenin, 16 2′,3-dihydroxy-4,4′,6′-trimethoxychalcone, 17 7-hydroxy-2′,3′,4′-trimethoxyisoflavan, 18 2′,3-dihydroxy-4,4′,6′-trimethoxychalcone (melilotocarpan C), 19 2′,3-dihydroxy-4,4′,6′-trimethoxychalcone (diapocynin), 20 dihydrophaseic acid, 21 4-hydroxyequol, 22 uralenneoside, 23 dodecanedioic acid, 24 2,3-dihydroxy-5-undecylcyclohexa-2,5-diene-1,4-dione, 25 tetradecanedioic acid, 26 3,7-dihydroxycholan-24-oic acid, 27 7alpha-hydroxy-3-oxo-5beta-cholan-24-oic acid, and 28 isodeoxycholic acid.
Morphological observation identified histological changes in the abomasal tissues of the lambs typical for haemonchosis. The changes in the CON group were aggregates near and in the tunica muscularis and gastric mucosa (Figure 4A) and submucosal oedema with scattered lymphocytes and glandular dilation (Figure 4B). We also observed submucosal oedema and the presence of lymphocytes near the t. muscularis in some cases (Figure 4C). Some animals in the CON group had mucosal hypertrophy and prominent lymphocyte aggregates with infiltration (Figure 4D).
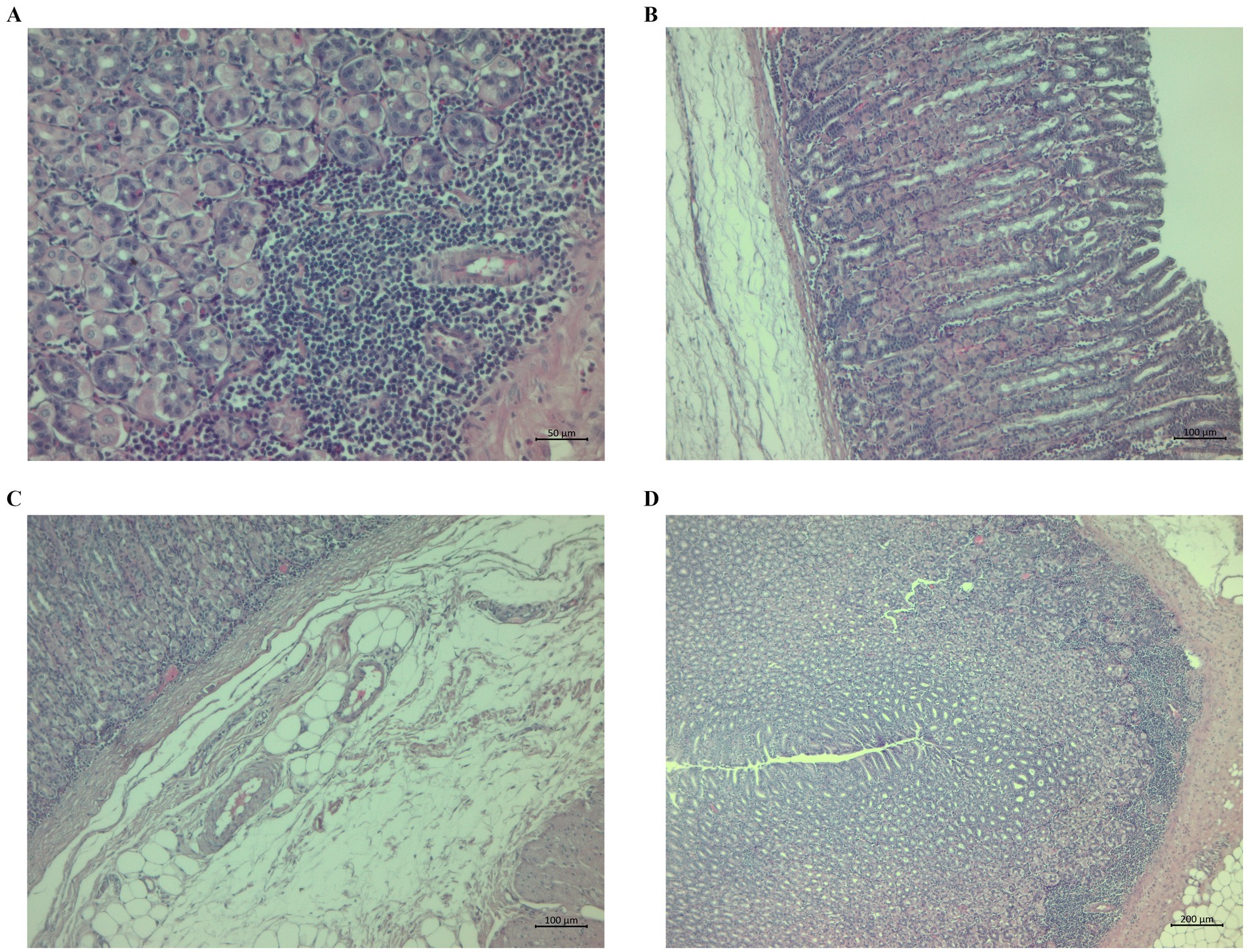
Figure 4. Histopathological sections of abomasa from the control group infected with Haemonchus contortus. (A) Sections stained with hematoxylin and eosin (H&E) (200×) showing aggregates near and in the tunica muscularis and gastric mucosa. (B) Sections stained with H&E (100×) showing submucosal oedema with scattered lymphocytes and dilatation of glands. (C) Sections stained with H&E (100×) showing severe submucosal oedema and the presence of lymphocytes near the t. muscularis. (D) Sections stained with H&E (40×) showing mucosal hypertrophy, marked lymphocyte aggregates, and infiltration.
The changes in the CHIC group were hypertrophy of the mucosa (Figure 5A) and mild hyperaemia and dilatation of glands (Figure 5B). Other pathological changes in the abomasal tissue included aggregates near the t. muscularis or scattered throughout the gastric mucosa (Figure 5C) and lymphocytic aggregates and infiltration (Figure 5D).
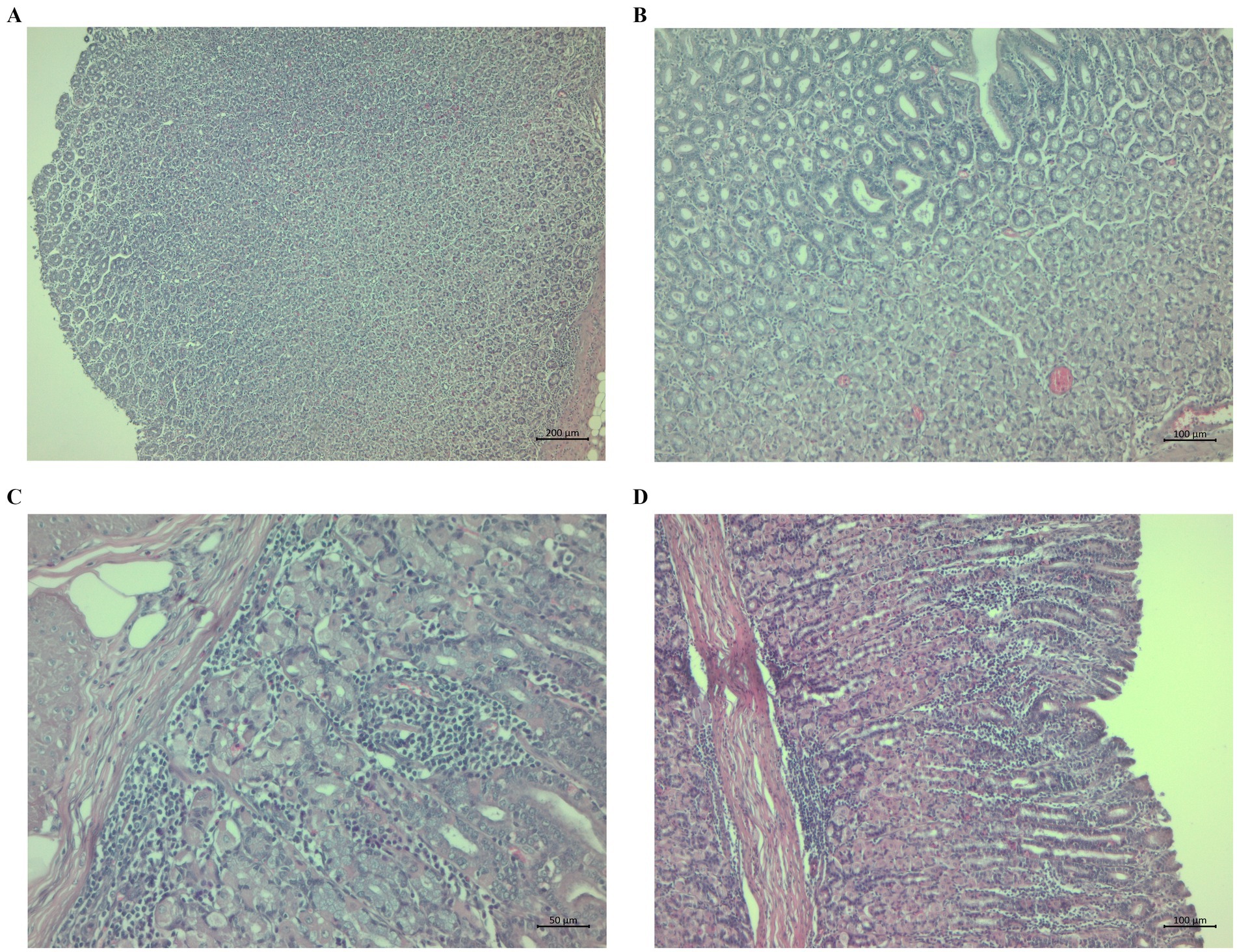
Figure 5. Histopathological sections of abomasa from the chicory group infected with Haemonchus contortus. (A) Sections stained with hematoxylin and eosin (H&E) (40×) showing hypertrophy of the mucosa. (B) Sections stained with H&E (100×) showing hypertrophy of the mucosa, mild hyperaemia, and dilatation of glands. (C) Sections stained with H&E (200×) showing aggregates near the tunica muscularis and gastric mucosa and scattered throughout the gastric mucosa. (D) Sections stained with H&E (100×) showing lymphocyte aggregates and infiltration.
We hypothesized that the natural meadow grassland enriched with experimentally sown chicory would influence the infection of lambs with H. contortus. We initially expected that egg excretion would gradually decrease in the experimental CHIC group throughout the experiment. The results of this study, however, clearly indicated that the fecal shedding of eggs also decreased significantly (p < 0.05) in the CON group from D48 onwards. This decrease was even larger up to D89 than in the CHIC group. The reduced egg production was probably correlated with the reduction in the number of adult worms in the abomasa as a direct correlation between EPG value and the number of GINs recorded for H. contortus in sheep and other ruminants (44).
Different nutritional strategies for managing GINs and H. contortus in ruminants (45–47) have already been examined, but comprehensive recommendations and mechanisms of action are still not unified. Chicory meets the criteria of a suitable antiparasitic plant due to in vitro and in vivo evidence of its anthelmintic effects, is voluntarily grazed by ruminants, can be cultivated as fodder, and is widespread in middle climatic zones (48). In vitro anthelmintic activity of chicory leaves and roots was confirmed by the inhibition of the hatching of Trichostrongylidae eggs with significant efficacies of 62.6% and higher (49). In a study on the effect of chicory compared to meadow hay in sheep infected with mixed Trichostrongylidae infections (50), a progressive increase in egg shedding by 80.83% on day 28 was observed in the group of sheep fed with meadow hay. Continuous reduction (by 24.75% after 28 days) in parasitic load in the chicory hay group was recorded (50). They observed a higher reduction (by 77.6%) after grazing on chicory for 28 days (at D62). The reduction in EPG value in the CHIC group, however, cannot be attributed to the action of chicory alone, because the reduction from D48 to D89 was 3.5–23.4% higher in the CON group. Heckendorn et al. (24) reported a similar course of infection with H. contortus in lambs administered with chicory, where the fecal egg counts peaked at the beginning of the study and then decreased significantly by 69%, confirming that tanniferous forages such as chicory were associated with significant reductions in the total daily fecal egg output of H. contortus. The peak of infection in the control group in the present study was not as high as in the lambs from the chicory group, similar to our experiment; thus, Heckendorn et al. (24) assumed that chicory had a direct antiparasitic effect. Our results indicated that infection in both groups was highest on D34, but the mean EPG was lower in the CON than the CHIC group, which was able to respond to the antiparasitic properties of the meadow grassland in the CON pasture. Our previous studies have described the antiparasitic properties of some plants in meadow grasslands grazed by CON lambs (17, 18, 34, 51).
The pathogenic effect of the GINs of ruminants also increases with the rate of stocking (52). The supplementation of lambs with food concentrates and the availability of proteins to replace lost proteins caused by parasitic infection also play a very important role (53). The combination of commercial food concentrate with the anthelmintic efficacy of some plants in our study (Table 4) could probably account for the significant decrease in excretion of H. contortus eggs in the CON group. EPG on D144 was lower in the CHIC than the CON group (Figure 1), which did not fully correlate with the necropsy results, where the total number of adult worms in the abomasum was higher in the CHIC group. The number of adult worms and the difference between the CHIC and CON groups, however, were not statistically significant. These findings suggest that worms from the CHIC group could be less fertile after exposure to chicory than worms from the CON group. The fecundity of female T. circumcincta exposed to chicory in another study did not differ from the control group (54), but similar data for H. contortus and this phenomenon are not available.
The positive antiparasitic effect of feeding or grazing experimentally infected calves on forage chicory, represented by a significant reduction in parasitic load and EPG value for O. ostertagi, has also been demonstrated (55). The welfare of grazing ruminants could be improved by eliminating negative human-animal interactions, such as the use of pharmacological tools (56). Herbivores can cope with parasitic infections by foraging, which includes mechanisms for preventing infection, resisting parasitism, and self-medicating by eating plants with anti-parasitic properties (57). Infected lambs grazing on rich grasslands are probably able to seek out plants with specific medicinal, antioxidant, immunological (58, 59), and anthelminthic effects and probably increase their consumption when self-medicating against GINs (60). Some polyphenols have synergistic effects. Significant synergies against parasitic infections in sheep have been reported for a combination of medicinal plants containing alkaloids, condensed tannins, flavonoids, and proteases, suggesting complex biochemical interactions that have strong anthelmintic effects (61). These factors may have played a critical role in reducing parasitic infection in both groups of lambs in our experimental study.
Our study of changes in the level of pasture contamination with H. contortus L3 provided interesting results. The level of contamination with L3 on both pasture plots increased in the first stage of the experiment (until D41), which was associated with the peculiarities of the development of H. contortus in the environment (62, 63). L3 contamination of the “random” pasture grass, however, was 20.9-fold higher on the CON than the CHIC plot, despite the similarity of the EPG values in the CON and CHIC lambs during this period and the similar high contamination of grass “near the feces” around the sheepfolds (Table 2). This difference in the contamination of the pasture plots may have been due to the lower survival of L3 on the plot recultivated with chicory compared to the natural meadow grass. Haemonchus contortus L3 survives and migrates differently in pastures with different types of vegetation, probably due to the peculiarities of the composition of the vegetation species and the structure of the meadow plant species (64–67). The physiological and morphological peculiarities of chicory may hinder larval survival on pasture. We also believe that several bioactive compounds and phenolic metabolites in lamb feces also influence the successful development of eggs and free-living larvae in the feces. The number of phenolic metabolites in our study was higher in the feces of the CHIC than in the CON lambs (Figures 2, 3). Chicory and its extracts negatively affect the development of eggs and larvae of various ruminants (54, 68, 69), so we believe that the decrease in the level of pasture contamination with L3 in our study was associated with the influence of bioactive plants, mainly chicory.
The subsequent dramatic reduction of pasture contamination with H. contortus larvae (at D78 and later) was consistent with the results of the FECR test. The lambs were not treated with any anthelmintics during this period, so we believe that the reduction in EPG values in the lambs and the level of L3 contamination in the pastures were associated with the influence of diverse phenolic metabolites on eggs and free-living larvae in the lamb’s feces. Approximately 95% of the parasite population in pastures is presented as eggs and free-living larvae, with only 5% of the population present as adults in animals (70), so the direct effect of bioactive plants and phenolic metabolites on the survival of eggs and larvae on pasture is a very important factor for GIN control. We therefore suggest that using bioactive plants to improve pasture management and reduce L3 intake in grazing ruminants may be a promising approach to control parasites in sheep and other ruminants.
The results of our experiment indicated that both the CON and CHIC pastures containing bioactive compounds had strong antioxidant properties, with a beneficial effect on the antioxidant status of infected lambs. The polyphenols in pasture grass likely provided natural antioxidants to minimize oxidative stress in the lambs, although antioxidant potential may have been influenced by several factors, including intestinal absorption, metabolism, bioavailability, and the presence or absence of co-antioxidants and transition metal ions (71). These findings are consistent with our previous results of using medicinal plants with a wide range of bioactive compounds that had strong antioxidant potential in the diet of lambs infected with H. contortus (18, 19, 34, 38, 51). Similarly, the plants in the grasslands of both the CON and CHIC pastures were good sources of flavonoids and phenols, which have antioxidant activity. Increasing the intake of medicinal plants on pasture likely helped to maintain the proper level of antioxidant potential and reduced the risk of infection associated with oxidative stress (72, 73). The antioxidant status of the infected lambs, however, probably also depended strongly on the efficiencies of the absorption, concentration, and metabolic transformation of phytochemicals and the environment (74).
To the best of our knowledge, this study is the first to perform a large-scale analysis of bioactive compounds on both pasture and the feces of lambs burdened with endoparasites. The natural meadow grassland with a diversity of plant species and enriched with experimentally sown chicory in our experiment was a source of important bioactive compounds that acted synergistically, with important biological effects (e.g., antioxidant, anti-inflammatory, or anthelmintic) in the treatment of GIN infection (75–77). Flavonoids (e.g., quercetin, luteolin, and kaempferol), flavonoid glycosides, and many flavonoid derivatives were the most abundant bioactive compounds identified in the pastures grazed by the CON and CHIC lambs, so they likely influenced GIN infection both directly and indirectly. Flavonoid glycosides and tannins have a similar mechanism of direct action against GINs due to their similar chemical structures (78) and have synergistic anthelmintic effects when tannins are combined with quercetin or luteolin (79). Flavonoids, however, are also known for their indirect activity, by which they increase host resistance to GINs (80, 81) by mobilizing the antioxidant defensive system, allowing animals to protect themselves from new oxidative conditions, as in our experiment. Finally, the phenolic acids, quinic acid, caffeic acid, and coumaric acid derivatives, which also have antioxidant, anti-inflammatory, antibacterial, and anthelmintic properties (82–84), were identified in the plots grazed by the CON and CHIC lambs.
The bioavailability, absorption, and distribution of polyphenols from the plots grazed by the CON and CHIC lambs were likely influenced by direct interactions between polyphenols and feed components in different parts of the digestive tract (85, 86). Some ingested flavonoids may be broken down into phenolic acids, but 90–95% of all ingested polyphenols may accumulate in the colonic lumen, and phenolic metabolites produced by microbes may be absorbed or excreted in the feces (87). The gut microbiota in both the CON and CHIC lambs was likely responsible for the extensive degradation of the original polyphenolic structures to a series of low-molecular-weight molecules that were absorbed and thus responsible for the biological activity. In the feces of both groups, we identified coumaric acid, a phenolic compound with a broad spectrum of biological activities (i.e., antimicrobial, antioxidant, and anti-inflammatory) (88), or chicoric acid, which has anti-inflammatory and antioxidant effects in digestive diseases (89). The flavonoid salvigenin, typical for Salvia species, has a wide range of biological and pharmacological properties and was abundantly represented in the feces of the lambs from both the CON and CHIC groups (90). These identifications of bioactive compounds in the feces were of great importance for the identification of the active phenolic metabolites in particular, because they likely had a specific effect against parasitic GIN larvae on pasture. The composition of polyphenols clearly varies considerably depending on the type of polyphenols and the feed source, but monomeric forms of flavonoids, for example, are best protected from degradation in ruminants, and their bioavailability is high (91). Our results confirmed that lambs can learn to prefer some bioactive plants during therapeutic self-medication or may adjust their intake when sick during nutritional self-medication, but the sustained consumption of bioactive plants can clearly lead to animals being tolerant to disease during prophylactic self-medication (92).
Morphological observation identified histopathological changes in the abomasal tissues typical of haemonchosis, but the changes were small and were only in some lambs compared to our previous experiments (20, 22), probably due to the eradication of parasites in most of the grazing lambs on D144 post-infection (Figure 1). Adult parasites pass into the abomasal lumen, where they feed on blood, move, and cause general histological changes, such as superficial damage to epithelial cells, hypertrophy of the mucosa, and oedema, followed by the formation of lymphoid aggregates and the infiltration of inflammatory cells in response to tissue damage (93, 94). Most of the histopathological changes in the abomasa, e.g., dilatation of glands, lymphoid aggregates near and in the t. muscularis, and oedema, were more notable but similar in both the CON and CHIC groups of lambs. Extensive changes in abomasal tissue previously reported, for example in infected goats, were accompanied by higher larval densities in the abomasal wall and depended on parasite number, immunology, and probably the genetic resistance of the worms (95). Our previous studies with supplementation of dry medicinal plants (Artemisia absinthium and Malva sylvestris (22) or Althaea officinalis, Petasites hybridus, Inula helenium, Plantago lanceolata, Rosmarinus officinalis, Solidago virgaurea, Fumaria officinalis, Hyssopus officinalis, and Foeniculum vulgare (20)) found strong local inflammation in the abomasal tissue, with the formation of lymphoid aggregates and the infiltration of immune cells in the lambs infected with H. contortus compared to unsupplemented infected lambs, even though the number of adult parasites in the abomasum at the end of experiments was generally highest in unsupplemented infected lambs (21, 51). The varying degrees of inflammation or changes in abomasal tissue in the CON and CHIC lambs in the present study may have been due to the delayed and incomplete defense against parasites, where a large number of parasites can cause lesions of the abomasal mucosa, which can lead to stronger local inflammation (96, 97).
Our results indicated that the availability of plants containing bioactive compounds in meadow grasslands may have a high potential to reduce parasite burdens in lambs and pasture contamination with infective nematode larvae which gives a future perspective in the control of parasitism in livestock. This study also performed a large-scale analysis of bioactive compounds on both pasture and the feces of lambs burdened with endoparasites, indicating their rich composition. Haemonchus contortus infections generally damage the abomasal mucosa, but our results suggest that bioactive plants can also likely minimise the adverse effects of GIN infection on the health of abomasal tissue.
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding authors.
The animal study was approved by the Ethical Committee of the Institute of Parasitology of the Slovak Academy of Sciences. The study was conducted in accordance with the local legislation and institutional requirements.
MK: Formal analysis, Investigation, Methodology, Writing – review & editing. DP: Conceptualization, Formal analysis, Methodology, Writing – review & editing. KD: Formal analysis, Writing – review & editing. ML: Formal analysis, Writing – review & editing. KČ: Data curation, Formal analysis, Writing – review & editing. MB: Formal analysis, Writing – review & editing. AK: Formal analysis, Writing – review & editing. TK: Investigation, Writing – review & editing. SS: Formal analysis, Writing – review & editing. PF: Formal analysis, Writing – review & editing. AŁ: Formal analysis, Writing – review & editing. PS: Formal analysis, Writing – review & editing. AC: Data curation, Writing – review & editing. ZV: Supervision, Writing – original draft, Writing – review & editing. MV: Funding acquisition, Project administration, Supervision, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This study was supported by funds from the EU NextGenerationEU of the Recovery and Resilience Plan for Slovakia under project No. 09I03-03-V04-00200/2024/VA. This research was also funded by the Scientific Grant Agency of the Ministry of Education of the Slovak Republic the Slovak Academy of Sciences (VEGA 2/0007/25) and partially funded by the Faculty of Veterinary Medicine and Animal Science, Poznan University of Life Sciences, Poland, by the Department of Animal Nutrition (no. 506.533.04.00). The involvement of T. Kuzmina in the study was partially supported by the NextGenerationEU through the Recovery and Resilience Plan for Slovakia under project No. 09I03-03-V01-00015. The funders had no role in designing the study; collecting, analysing, or interpreting the data; or writing the manuscript.
The authors are grateful to V. Venglovská, P. Jerga, and S. Spišáková for their laboratory and technical assistance. The English has been revised by a native English language editor, Dr. William Blackhall.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors declare that no Gen AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fvets.2025.1566720/full#supplementary-material
1. Charlier, J, Rinaldi, L, Musella, V, Loeger, HW, Chartier, C, Vineer, HR, et al. Initial assessment of the economic burden of major parasitic helminth infections to the ruminant livestock industry in Europe. Prev Vet Med. (2020) 182:105103. doi: 10.1016/j.prevetmed.2020.105103
2. Mavrot, F, Hertzberg, H, and Torgerson, P. Effect of gastro-intestinal nematode infection on sheep performance: a systematic review and meta-analysis. Parasit Vectors. (2015) 8:557. doi: 10.1186/s13071-015-1164-z
3. Mahmoud, MAM, Elfadil, AAM, Yagoup, EA, Adam, IA, Mohamed, EGS, Bushara, SB, et al. Epidemiological study of Haemonchus contortus among sheep in north Kordufan state. Sudan Int J Vet Sci. (2017) 6:209–15.
4. Hassan, M, Saeed, Z, Shaukat, A, Qureshi, MA, Aleem, MT, and Ahmad, Z. Comparing FAMACHA® scores and hematological parameters in Nachi, Beetal, and Pak angora goats naturally infected with Haemonchus contortus. Continental Vet J. (2024) 4:14–27. doi: 10.71081/cvj/2024.003
5. Gareh, A, Elhawary, NM, Tahoun, A, Ramez, AM, El-Shewehy, DMM, Elbaz, E, et al. Epidemiological, morphological, and morphometric study on Haemonchus spp. recovered from goats in Egypt. Front Vet Sci. (2021) 8:705619. doi: 10.3389/fvets.2021.705619
6. Selzer, PM, and Epe, C. Antiparasitics in animal health: quo vadis? Trends Parasitol. (2021) 37:77–89. doi: 10.1016/j.pt.2020.09.004
7. Jedziniak, P, Szprengier-Juszkiewicz, T, and Olejnik, M. Determination of benzimidazoles and levamisole residues in milk by liquid chromatography-mass spectrometry: screening method development and validation. J Chromatogr A. (2009) 1216:8165–72. doi: 10.1016/j.chroma.2009.07.036
8. Jackson, F, Várady, M, and Bartley, DJ. Managing anthelmintic resistance in goats – can we learn lessons from sheep? Small Rumin Res. (2012) 103:3–9. doi: 10.1016/j.smallrumres.2011.10.012
9. Bihaqi, SJ, Allaie, IM, Banday, MAA, Sankar, M, Wani, ZA, and Prasad, A. Multiple anthelmintic resistance in gastrointestinal nematodes of Caprines on mountain research Centre for Sheep and Goat at Kashmir Valley, India. Parasite Epidemiol Control. (2020) 11:e00163. doi: 10.1016/j.parepi.2020.e00163
10. Maqbool, I, Wani, ZA, Shahardar, RA, Allaie, IM, and Shah, MM. Integrated parasite management with special reference to gastro-intestinal nematodes. J Parasit Dis. (2017) 41:1–8. doi: 10.1007/s12639-016-0765-6
11. Pavičić, A, Zajíčková, M, Šadibolová, M, Svobodová, G, Matoušková, P, Szotáková, B, et al. Anthelmintic activity of European fern extracts against Haemonchus contortus. Vet Res. (2023) 54:59. doi: 10.1186/s13567-023-01192-8
12. Qamar, W, and Alkheraije, KA. Anthelmintic resistance in Haemonchus contortus of sheep and goats from Asia–a review of in vitro and in vivo studies. Pak Vet J. (2023) 43:376–87. doi: 10.29261/pakvetj/2023.088
13. Abubakar, M, Oneeb, M, Rashid, M, Ashraf, K, Chisti, GA, Awan, F, et al. In vitro anthelmintic efficacy of three plants extracts against various developmental stages of Haemonchus contortus. Pak Vet J. (2024) 44:238–43. doi: 10.29261/pakvetj/2024.174
14. Githiori, JB, Athanasiadou, S, and Thamsborg, SM. Use of plants in novel approaches for control of gastrointestinal helminths in livestock with emphasis on small ruminants. Vet Parasitol. (2006) 139:308–20. doi: 10.1016/j.vetpar.2006.04.021
15. Manolaraki, F, Sotiraki, S, Stefanakis, A, Skampardonis, V, Volanis, M, and Hoste, H. Anthelmintic activity of some Mediterranean browse plants against parasitic nematodes. Parasitology. (2010) 137:685–96. doi: 10.1017/S0031182009991399
16. Hoste, H, Torres-Acosta, JF, Quijada, J, Chan-Perez, I, Dakheel, MM, Kommuru, DS, et al. Interactions between nutrition and infections with Haemonchus contortus and related gastrointestinal nematodes in small ruminants. Adv Parasitol. (2016) 93:239–351. doi: 10.1016/bs.apar.2016.02.025
17. Váradyová, Z, Kišidayová, S, Čobanová, K, Grešáková, Ľ, Babják, M, Königová, A, et al. The impact of a mixture of medicinal herbs on ruminal fermentation, parasitological status and haematological parameters of the lambs experimentally infected with Haemonchus contortus. Small Rumin Res. (2017) 151:124–32. doi: 10.1016/j.smallrumres.2017.04.023
18. Mravčáková, D, Váradyová, Z, Kopčáková, A, Čobanová, K, Grešáková, Ľ, Kišidayová, S, et al. Natural chemotherapeutic alternatives for controlling of haemonchosis in sheep. BMC Vet Res. (2019) 15:302. doi: 10.1186/s12917-019-2050-2
19. Komáromyová, M, Petrič, D, Kucková, K, Batťányi, D, Babják, M, Dolinská, MU, et al. Impact of sainfoin (Onobrychis viciifolia) pellets on parasitological status, antibody responses, and antioxidant parameters in lambs infected with Haemonchus contortus. Pathogens. (2022) 11:301. doi: 10.3390/pathogens11030301
20. Batťányi, D, Petrič, D, Babják, M, Dvorožňáková, E, Łukomska, A, Cieslak, A, et al. Antibody response and abomasal histopathology of lambs with haemonchosis during supplementation with medicinal plants and organic selenium. Vet Anim Sci. (2023) 19:100290. doi: 10.1016/j.vas.2023.100290
21. Mravčáková, D, Komáromyová, M, Babják, M, Urda Dolinská, M, Königová, A, Petrič, D, et al. Anthelmintic activity of wormwood (Artemisia absinthium L.) and mallow (Malva sylvestris L.) against Haemonchus contortus in sheep. Animals. (2020) 10:219. doi: 10.3390/ani10020219
22. Mravčáková, D, Sobczak-Filipiak, M, Váradyová, Z, Kucková, K, Čobanová, K, Maršík, P, et al. Effect of Artemisia absinthium and Malva sylvestris on antioxidant parameters and abomasal histopathology in lambs experimentally infected with Haemonchus contortus. Animals. (2021) 11:462. doi: 10.3390/ani11020462
23. Marley, CL, Cook, R, Keatinge, R, Barrett, J, and Lampkin, NH. The effect of birdsfoot trefoil (Lotus corniculatus) and chicory (Cichorium intybus) on parasite intensities and performance of lambs naturally infected with helminth parasites. Vet Parasitol. (2003) 112:147–55. doi: 10.1016/s0304-4017(02)00412-0
24. Heckendorn, F, Häring, DA, Maurer, V, Senn, M, and Hertzberg, H. Individual administration of three tanniferous forage plants to lambs artificially infected with Haemonchus contortus and Cooperia curticei. Vet Parasitol. (2007) 146:123–34. doi: 10.1016/j.vetpar.2007.01.009
25. Kidane, A, Houdijk, J, Athanasiadou, S, Tolkamp, B, and Kyriazakis, I. Nutritional sensitivity of periparturient resistance to nematode parasites in two breeds of sheep with different nutrient demands. Br J Nutr. (2010) 104:1477–86. doi: 10.1017/S0007114510002503
26. Machado, V, Da Silva, AS, Schafer, AS, Aires, AR, Tonin, AA, Oliveira, CB, et al. Relationship between oxidative stress and pathological findings in abomasum of infected lambs by Haemonchus contortus. Pathol Res Pract. (2014) 210:812–7. doi: 10.1016/j.prp.2014.09.006
27. Li, G, and Kemp, PD. Forage chicory (Cichorium intybus L.): a review of its agronomy and animal production. Adv Agron. (2005) 88:187–222. doi: 10.1016/S0065-2113(05)88005-8
28. Marley, CL, Fychan, R, Davies, JW, Scollan, ND, Richardson, RI, Theobald, VJ, et al. Effects of chicory/perennial ryegrass swards compared with perennial ryegrass swards on the performance and carcass quality of grazing beef steers. PLoS One. (2014) 9:e86259. doi: 10.1371/journal.pone.0086259
29. Tzamaloukas, O, Athanasiadou, S, Kyriazakis, I, Huntley, JF, and Jackson, F. The effect of chicory (Cichorium intybus) and sulla (Hedysarum coronarium) on larval development and mucosal cell responses of growing lambs challenged with Teladorsagia circumcincta. Parasitology. (2006) 132:419–26. doi: 10.1017/S0031182005009194
30. Athanasiadou, S, Gray, D, Younie, D, Tzamaloukas, O, Jackson, F, and Kyriazakis, I. The use of chicory for parasite control in organic ewes and their lambs. Parasitology. (2007) 134:299–307. doi: 10.1017/S0031182006001363
31. Hoste, H, Torres-Acosta, JF, Alonso-diaz, MÁ, Brunet, S, Sandoval-Castro, C, and Adote, SH. Identification and validation of bioactive plants for the control of gastrointestinal nematodes in small ruminants. Trop Biomed. (2008) 25:56–72.
32. Kresánek, J Jr, and Kresánek, J. Atlas of medicinal plants and berries. Slovakia: Osveta (2008).
34. Váradyová, Z, Mravčáková, D, Babják, M, Bryszak, M, Grešáková, Ľ, Čobanová, K, et al. Effects of herbal nutraceuticals and/or zinc against Haemonchus contortus in lambs experimentally infected. BMC Vet Res. (2018) 14:78. doi: 10.1186/s12917-018-1405-4
35. European Commission. (2009). Council regulation (EC) 1099/2009 of 24 September 2009 on the protection of animals at the time of killing. Available online at:. (https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=celex:32009R1099)
36. Coles, GC, Bauer, C, Borgsteede, FH, Geerts, S, Klei, TR, Taylor, MA, et al. World Association for the Advancement of veterinary parasitology (W.A.A.V.P.) methods for the detection of anthelmintic resistance in nematodes of veterinary importance. Vet Parasitol. (1992) 44:35–44. doi: 10.1016/0304-4017(92)90141-u
37. Cabaret, J, and Berrag, B. Faecal egg count reduction test for assessing anthelmintic efficacy: average versus individually based estimations. Vet Parasitol. (2004) 121:105–13. doi: 10.1016/j.vetpar.2004.01.020
38. Čobanová, K, Váradyová, Z, Grešáková, Ľ, Kucková, K, Mravčáková, D, and Várady, M. Does herbal and/or zinc dietary supplementation improve the antioxidant and mineral status of lambs with parasite infection? Antioxidants. (2020) 9:1172. doi: 10.3390/antiox9121172
39. Theodorou, MK, Kingston-Smith, AH, Winters, AL, Lee, MRF, Minchin, FR, Morris, P, et al. Polyphenols and their influence on gut function and health in ruminants: a review. Environ Chem Lett. (2006) 4:121–6. doi: 10.1007/s10311-006-0061-2
40. Lewandowska, U, Szewczyk, K, Hrabec, E, Janecka, A, and Gorlach, S. Overview of metabolism and bioavailability enhancement of polyphenols. J Agric Food Chem. (2013) 61:12183–99. doi: 10.1021/jf404439b
41. Lee, MR. Forage polyphenol oxidase and ruminant livestock nutrition. Front Plant Sci. (2014) 5:694. doi: 10.3389/fpls.2014.00694
42. Stevens, JF, and Maier, CS. The chemistry of gut microbial metabolism of polyphenols. Phytochem Rev. (2016) 15:425–44. doi: 10.1007/s11101-016-9459-z
43. Petrič, D, Mravčáková, D, Kucková, K, Kišidayová, S, Cieslak, A, Szumacher-Strabel, M, et al. Impact of zinc and/or herbal mixture on ruminal fermentation, microbiota, and histopathology in lambs. Front Vet Sci. (2021) 8:630971. doi: 10.3389/fvets.2021.630971
44. Cabaret, J, Gasnier, N, and Jacquiet, P. Faecal egg counts are representative of digestive-tract strongyle worm burdens in sheep and goats. Parasite. (1998) 5:137–42. doi: 10.1051/parasite/1998052137
45. Haider, A, Hussain, K, Mares, MM, Abbas, A, Mohsin, M, Rehman, A, et al. In vitro and in vivo anthelmintic activity of Nicotiana tabacum against Haemonchus placei in cattle. Pak Vet J. (2024) 44:745–50. doi: 10.29261/pakvetj/2024.245
46. Cissokho, B, Sambe Babacar, S, Diome, T, Ndiaye, MR, and Sembene, M. In vitro nematocidal effects of aqueous extracts of leaves and bark of Khaya senegalensis and Azadirachta indica on the mortality of adults of Haemonchus contortus, cobb, 1899 (Rhabditida, Trichostrongylidae). Agrobiol Rec. (2024) 18:105–12. doi: 10.47278/journal.abr/2024.042
47. Batool, S, Munir, F, Sindhu, ZD, Abbas, RZ, Aslam, B, Khan, MK, et al. In vitro anthelmintic activity of Azadirachta indica (neem) and Melia azedarach (bakain) essential oils and their silver nanoparticles against Haemonchus contortus. Agrobiol Rec. (2023) 11:6–12. doi: 10.47278/journal.abr/2023.002
48. Rodríguez-Hernández, P, Reyes-Palomo, C, Sanz-Fernández, S, Rufino-Moya, PJ, Zafra, R, Martínez-Moreno, FJ, et al. Antiparasitic tannin-rich plants from the south of Europe for grazing livestock: a review. Animals. (2023) 13:201. doi: 10.3390/ani13020201
49. Bosco, A, Prigioniero, A, Falzarano, A, Maurelli, MP, Rinaldi, L, Cringoli, G, et al. Use of perennial plants in the fight against gastrointestinal nematodes of sheep. Front Parasitol. (2023) 2:1186149. doi: 10.3389/fpara.2023.1186149
50. Cireșan, CA, Cocan, I, Alexa, E, Cărpinișan, L, Sîrbu, CB, Obiștioiu, D, et al. Research on the control of gastrointestinal strongyles in sheep by using Lotus corniculatus or Cichorium intybus in feed. Pathogens. (2023) 12:986. doi: 10.3390/pathogens12080986
51. Komáromyová, M, Mravčáková, D, Petrič, D, Kucková, K, Babják, M, Dolinská, MU, et al. Effects of medicinal plants and organic selenium against ovine haemonchosis. Animals. (2021) 11:1319. doi: 10.3390/ani11051319
52. Thamsborg, SM, Jørgensen, RJ, Ranvig, H, Bartlett, P, Waller, PJ, and Nansen, P. The performance of grazing sheep in relation to stocking rate and exposure to nematode infections. Livest Prod Sci. (1998) 53:265–77. doi: 10.1016/S0301-6226(97)00105-X
53. Poli, CHEC, Monteiro, ALG, Devincenzi, T, de Albuquerque, FHMAR, da Motta, JH, Borges, LI, et al. Management strategies for lamb production on pasture-based systems in subtropical regions: a review. Front Vet Sci. (2020) 7:543. doi: 10.3389/fvets.2020.00543
54. Tzamaloukas, O, Athanasiadou, S, Kyriazakis, I, Jackson, F, and Coop, RL. The consequences of short-term grazing of bioactive forages on established adult and incoming larvae populations of Teladorsagia circumcincta in lambs. Int J Parasitol. (2005) 35:329–35. doi: 10.1016/j.ijpara.2004.11.013
55. Peña-Espinoza, M, Thamsborg, SM, Desrues, O, Hansen, TV, and Enemark, HL. Anthelmintic effects of forage chicory (Cichorium intybus) against gastrointestinal nematode parasites in experimentally infected cattle. Parasitology. (2016) 143:1279–93. doi: 10.1017/S0031182016000706
56. Bricarello, PA, Longo, C, da Rocha, RA, and Hötzel, MJ. Understanding animal-plant-parasite interactions to improve the management of gastrointestinal nematodes in grazing ruminants. Pathogens. (2023) 12:531. doi: 10.3390/pathogens12040531
57. Hutchings, MR, Athanasiadou, S, Kyriazakis, I, and Gordon, IJ. Can animals use foraging behaviour to combat parasites? Proc Nutr Soc. (2003) 62:361–70. doi: 10.1079/pns2003243
58. Hosoda, K, Kuramoto, K, Eruden, B, Nishida, T, and Shioya, S. The effect of three herbs as feed supplements on blood metabolites, hormones, antioxidant activity, IgG concentration, and ruminal fermentation in Holstein steers. Asian Australas J Anim Sci. (2006) 19:35–41. doi: 10.5713/ajas.2006.35
59. Fraisse, D, Carnat, A, Viala, D, Pradel, P, Besle, JM, Coulon, JB, et al. Polyphenolic composition of a permanent pasture: variations related to the period of harvesting. J Sci Food Agric. (2007) 87:2427–35. doi: 10.1002/jsfa.2918
60. Villalba, JJ, Miller, J, Ungar, ED, Landau, SY, and Glendinning, J. Ruminant self-medication against gastrointestinal nematodes: evidence, mechanism, and origins. Parasite. (2014) 21:31. doi: 10.1051/parasite/2014032
61. Fomum, SW, and Nsahlai, IV. In vitro evaluation of anthelmintic interaction of plant species combinations putatively containing different bioactive macromolecules in sheep. J Altern Compl Integr Med. (2019) 6:1–9. doi: 10.24966/ACIM-7562/100087
62. Coadwell, WJ, and Ward, PF. Observations on the development of Haemonchus contortus in young sheep given a single infection. Parasitology. (1975) 71:505–15. doi: 10.1017/s0031182000047260
63. Rahman, WA, and Collins, GH. The establishment and development of Haemonchus contortus in goats. Vet Parasitol. (1990) 35:189–93. doi: 10.1016/0304-4017(90)90053-e
64. O'Connor, LJ, Walkden-Brown, SW, and Kahn, LP. Ecology of the free-living stages of major trichostrongylid parasites of sheep. Vet Parasitol. (2006) 30:1–15. doi: 10.1016/j.vetpar.2006.08.035
65. Oliveira, AL, Costa, C, Rodella, RA, Silva, BF, and Amarante, AF. Effect of plant trichomes on the vertical migration of Haemonchus contortus infective larvae on five tropical forages. Trop Anim Health Prod. (2009) 41:775–82. doi: 10.1007/s11250-008-9251-1
66. de Pires, CRS, Cavalcante, GG, Prado, RS, Costa Junior, LM, and GRC, R. Survival of Haemonchus contortus larvae in forage species in the eastern Amazon. Pesq Agrop Brasileira. (2021) 56:e02664. doi: 10.1590/S1678-3921.pab2021.v56.02664
67. Gasparina, JM, Baby, RG, Fonseca, L, Bricarello, PA, and Rocha, RA. Infective larvae of Haemonchus contortus found from the base to the top of the grass sward. Braz J Vet Parasitol. (2021) 30:e028120. doi: 10.1590/S1984-29612021032
68. Deane, JC, Warren, J, Findlay, E, Dagleish, MP, Cork, SC, Jackson, F, et al. The effect of Cichorium intybus and Lotus corniculatus on nematode burdens and production in grazed lambs. In: Proceedings of the UK organic research conference, 26–28. (2002). Organic Centre Wales, Institute of Rural Studies, University of Wales, Aberystwyth.
69. Peña-Espinoza, M, Valente, AH, Thamsborg, SM, Simonsen, HT, Boas, U, Enemark, HL, et al. Antiparasitic activity of chicory (Cichorium intybus) and its natural bioactive compounds in livestock: a review. Parasit Vectors. (2018) 11:475. doi: 10.1186/s13071-018-3012-4
70. Bowman, DD, Lynn, RC, Eberhard, ML, and Alcaraz, A. Georgi’s parasitology for veterinarians. Philadelphia: Saunders (2003).
71. Kasote, DM, Katyare, SS, Hegde, MV, and Bae, H. Significance of antioxidant potential of plants and its relevance to therapeutic applications. Int J Biol Sci. (2015) 11:982–91. doi: 10.7150/ijbs.12096
72. Singh, A, Singh, S, and Prasad, SM. Role of medicinal plants for health perspective: special references to antioxidant potential. J Chem Biol Ther. (2016) 1:106. doi: 10.4172/2572-0406.1000106
73. Zafar, F, Asif, HM, Shaheen, G, Ghauri, AO, Rajpoot, SR, Tasleem, MW, et al. A comprehensive review on medicinal plants possessing antioxidant potential. Clin Exp Pharmacol Physiol. (2023) 50:205–17. doi: 10.1111/1440-1681.13743
74. Surai, PF. Polyphenol compounds in the chicken/animal diet: from the past to the future. J Anim Physiol Anim Nutr. (2014) 98:19–31. doi: 10.1111/jpn.12070
75. Lavecchia, T, Rea, G, Antonacci, A, and Giardi, MT. Healthy and adverse effects of plant-derived functional metabolites: the need of revealing their content and bioactivity in a complex food matrix. Crit Rev Food Sci Nutr. (2013) 53:198–213. doi: 10.1080/10408398.2010.520829
76. Brusotti, G, Cesari, I, Dentamaro, A, Caccialanza, G, and Massolini, G. Isolation and characterization of bioactive compounds from plant resources: the role of analysis in the ethnopharmacological approach. J Pharm Biomed Anal. (2014) 87:218–28. doi: 10.1016/j.jpba.2013.03.007
77. Spiegler, V, Liebau, E, and Hensel, A. Medicinal plant extracts and plant-derived polyphenols with anthelmintic activity against intestinal nematodes. Nat Prod Rep. (2017) 34:627–43. doi: 10.1039/c6np00126b
78. Barrau, E, Fabre, N, Fouraste, I, and Hoste, H. Effect of bioactive compounds from sainfoin (Onobrychis viciifolia Scop.) on the in vitro larval migration of Haemonchus contortus: role of tannins and flavonol glycosides. Parasitology. (2005) 131:531–8. doi: 10.1017/S0031182005008024
79. Klongsiriwet, C, Quijada, J, Williams, AR, Mueller-Harvey, I, Williamson, EM, and Hoste, H. Synergistic inhibition of Haemonchus contortus exsheathment by flavonoid monomers and condensed tannins. Int J Parasitol Drugs Drug Resist. (2015) 5:127–34. doi: 10.1016/j.ijpddr.2015.06.001
80. Iqbal, Z, Sarwar, M, Jabbar, A, Ahmed, S, Nisa, M, Sajid, MS, et al. Direct and indirect anthelmintic effects of condensed tannins in sheep. Vet Parasitol. (2007) 144:125–31. doi: 10.1016/j.vetpar.2006.09.035
81. Hoste, H, Torres-Acosta, JF, Sandoval-Castro, CA, Mueller-Harvey, I, Sotiraki, S, Louvandini, H, et al. Tannin containing legumes as a model for nutraceuticals against digestive parasites in livestock. Vet Parasitol. (2015) 212:5–17. doi: 10.1016/j.vetpar.2015.06.026
82. Duarte-Almeida, JM, Salatino, A, Genovese, MI, and Lajolo, FM. Phenolic composition and antioxidant activity of culms and sugarcane (Saccharum officinarum L.) products. Food Chem. (2011) 125:660–4. doi: 10.1016/j.foodchem.2010.09.059
83. Sato, Y, Itagaki, S, Kurokawa, T, Ogura, J, Kobayashi, M, Hirano, T, et al. In vitro and in vivo antioxidant properties of chlorogenic acid and caffeic acid. Int J Pharm. (2011) 403:136–8. doi: 10.1016/j.ijpharm.2010.09.035
84. Xiong, J, Li, S, Wang, W, Hong, Y, Tang, K, and Luo, Q. Screening and identification of the antibacterial bioactive compounds from Lonicera japonica Thunb. Leaves. Food Chem. (2013) 138:327–33. doi: 10.1016/j.foodchem.2012.10.127
85. Rice-Evans, C, Spencer, JP, Schroeter, H, and Rechner, AR. Bioavailability of flavonoids and potential bioactive forms in vivo. Drug Metabol Drug Interact. (2000) 17:291–310. doi: 10.1515/DMDI.2000.17.1-4.291
86. D'Archivio, M, Filesi, C, Varì, R, Scazzocchio, B, and Masella, R. Bioavailability of the polyphenols: status and controversies. Int J Mol Sci. (2010) 11:1321–42. doi: 10.3390/ijms11041321
87. Bešlo, D, Golubić, N, Rastija, V, Agić, D, Karnaš, M, Šubarić, D, et al. Antioxidant activity, metabolism, and bioavailability of polyphenols in the diet of animals. Antioxidants. (2023) 12:1141. doi: 10.3390/antiox12061141
88. Garg, SS, Gupta, J, Sharma, S, and Sahu, D. An insight into the therapeutic applications of coumarin compounds and their mechanisms of action. Eur J Pharm Sci. (2020) 152:105424. doi: 10.1016/j.ejps.2020.105424
89. Peng, Y, Sun, Q, Gao, R, and Park, Y. AAK-2 and SKN-1 are involved in chicoric-acid-induced lifespan extension in Caenorhabditis elegans. J Agric Food Chem. (2019) 67:9178–86. doi: 10.1021/acs.jafc.9b00705
90. Mansourabadi, AH, Sadeghi, HM, Razavi, N, and Rezvani, E. Anti-inflammatory and analgesic properties of salvigenin, Salvia officinalis flavonoid extracted. Adv Herb Med. (2015) 1:31–41.
91. Bešlo, D, Došlić, G, Agić, D, Rastija, V, Šperanda, M, Gantner, V, et al. Polyphenols in ruminant nutrition and their effects on reproduction. Antioxidants. (2022) 11:970. doi: 10.3390/antiox11050970
92. Villalba, JJ, Ramsey, RD, and Athanasiadou, S. Review: herbivory and the power of phytochemical diversity on animal health. Animal. (2024) 6:101287. doi: 10.1016/j.animal.2024.101287
93. Simpson, HV. Pathophysiology of abomasal parasitism: is the host or parasite responsible? Vet J. (2000) 160:177–91. doi: 10.1053/tvjl.2000.0491
94. Stear, MJ, Bishop, SC, Henderson, NG, and Scott, I. A key mechanism of pathogenesis in sheep infected with the nematode Teladorsagia circumcincta. Anim Health Res Rev. (2003) 4:45–52. doi: 10.1079/ahrr200351
95. Al-Zubaidy, AJ, Altaif, KI, Al-Qaisy, HH, and Makkawi, TA. Gross pathology and histopathology of haemonchosis in sheep and goats in Iraq. Vet Parasitol. (1987) 23:249–56. doi: 10.1016/0304-4017(87)90010-0
96. Toscano, JHB, Okino, CH, Dos Santos, IB, Giraldelo, LA, von Haehling, MB, Esteves, SN, et al. Innate immune responses associated with resistance against Haemonchus contortus in Morada Nova sheep. J Immunol Res. (2019) 2019:3562672. doi: 10.1155/2019/3562672
Keywords: antioxidant status, chicory, histology, lambs, parasitological status, phytochemicals
Citation: Komáromyová M, Petrič D, Demčáková K, Leško M, Čobanová K, Babják M, Königová A, Kuzmina T, Ślusarczyk S, Fortuna PI, Łukomska A, Sidoruk P, Cieslak A, Váradyová Z and Várady M (2025) Insights into the role of bioactive plants for lambs infected with Haemonchus contortus parasite. Front. Vet. Sci. 12:1566720. doi: 10.3389/fvets.2025.1566720
Edited by:
Nicola Pugliese, University of Bari Aldo Moro, ItalyReviewed by:
Rao Zahid Abbas, University of Agriculture, PakistanCopyright © 2025 Komáromyová, Petrič, Demčáková, Leško, Čobanová, Babják, Königová, Kuzmina, Ślusarczyk, Fortuna, Łukomska, Sidoruk, Cieslak, Váradyová and Várady. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Zora Váradyová, dmFyYWR5ekBzYXNrZS5zaw==; Marián Várady, dmFyYWR5QHNhc2tlLnNr
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.