 Miguel Criado1,2*
Miguel Criado1,2* Marta Silva1,2
Marta Silva1,2 Noive Arteche-Villasol1,2
Noive Arteche-Villasol1,2 David Zapico1,2
David Zapico1,2 Natalia Elguezabal3
Natalia Elguezabal3 Elena Molina3
Elena Molina3 José Espinosa1,2
José Espinosa1,2 María del Carmen Ferreras1,2
María del Carmen Ferreras1,2 Julio Benavides1
Julio Benavides1 Valentín Pérez1,2
Valentín Pérez1,2 Daniel Gutiérrez-Expósito1,2
Daniel Gutiérrez-Expósito1,2- 1Departamento de Sanidad Animal, Instituto de Ganadería de Montaña (IGM) CSIC-ULE, León, Spain
- 2Departamento de Sanidad Animal, Facultad de Veterinaria, Universidad de León, Campus de Vegazana, León, Spain
- 3Departamento de Sanidad Animal, NEIKER-BRTA, Instituto Vasco de Investigación y Desarrollo Agrario, Derio, Spain
Paratuberculosis is a chronic granulomatous enteritis, caused by Mycobacterium avium subspecies paratuberculosis (Map), that affects ruminants worldwide. Vaccination has been considered the most cost-effective method for the control of this disease in infected dairy herds. However, currently available vaccines do not provide complete protection and interfere with the diagnosis of both paratuberculosis and bovine tuberculosis, limiting its use. Because of that, efforts are being made for the development of new vaccines. The primary objective of this study was to evaluate the efficacy of two whole-cell inactivated experimental vaccines against paratuberculosis in goats, administered through the oral (OV) and intradermal (IDV) routes, and compare them with that of the commercial subcutaneous vaccine Gudair® (SCV). Over an 11-month period, the effect of vaccination and a subsequent Map challenge on the specific peripheral immune responses and Map-DNA fecal shedding were recorded. At the end of the experiment, tissue bacterial load and lesion severity were assessed. The experimental vaccines did not induce specific humoral immune responses and only elicited mild and delayed cellular immune responses. Although the OV reduced lesion severity, neither this vaccine nor the IDV prototype was able to reduce fecal shedding or tissue bacterial load. Moreover, although the SCV did not confer sterile immunity, it outperformed both experimental vaccines in all these parameters.
1 Introduction
Paratuberculosis (PTB) is a chronic granulomatous enteritis produced by Mycobacterium avium subspecies paratuberculosis (Map). Domestic and wild ruminants are the most commonly affected species by this worldwide distributed disease, which is officially recognized by the World Organisation for Animal Health (WOAH) (1), and regulated under the EU Animal Health Law (2). It causes important economic losses in cattle, sheep, and goat livestock industries, where PTB is highly prevalent (3), as seen by the estimated herd-level prevalences in southern Spain, which are 66.3% for sheep and 90% for goats (4). Control programs based on hygiene and management measures are hampered by the ability of Map to persist in the environment. In addition, although diagnostic tests are improving, sensible detection of infected animals is still challenging, especially in the early stages of infection. Therefore, vaccination has been considered the most cost-effective control tool in affected herds (5–9), as it reduces the colonization of intestinal tissues, Map shedding, and the number of clinically affected animals (10, 11). However, currently available commercial vaccines present several drawbacks, as they do not fully protect against infection or prevent transmission (12), they induce the formation of a large granuloma in the injection site (13), and they could interfere with PTB diagnosis (14–16), and with the official bovine tuberculosis (bTB) diagnostic tests (17). As a result of this interference, vaccination has been prohibited in several countries with eradication programs against PTB (18), and many countries with bTB control programs do not allow vaccination against PTB in cattle with current vaccines (5).
Due to this diagnostic interference, there has been widespread reluctance to adopt vaccination strategies for goats in countries where vaccines are available for this species. Furthermore, the relatively smaller economic importance of goats in developed countries likely contributes to the lower number of vaccination trials conducted for goats than those for cattle and sheep. Nevertheless, goats are highly susceptible to PTB and frequently develop diffuse forms of the disease (5, 19), and in this species, hygiene and management measures are particularly challenging; for example, strategies such as individual testing and culling are prohibitively expensive. Previous efforts relying on these measures have not successfully reduced the incidence of the disease. In contrast, vaccination has been shown to be both highly effective and economically viable (5, 8). However, differences in immune responses between cattle and goats to other vaccines (20) should be considered in developing new vaccines against PTB.
All currently approved vaccines and most which were approved until recently are inactivated and administered subcutaneously. One exception to this was Neoparasec®, which used a live-attenuated 316F Map strain and was discontinued in 2001. To avoid the drawbacks previously mentioned, efforts have been made in the development of alternative vaccination strategies against PTB over the last decades. Many attenuated vaccines have been studied, although, in addition to the risks associated with this type of vaccine, they share most of the disadvantages of the available inactivated vaccines. One of the most comprehensive recent trials conducted in five different laboratories, evaluated 22 attenuated Map strains, finding that none of them constituted an optimal vaccine candidate (21). Two of these attenuated strains have been tested in other in vivo studies. A K10 ΔpknG mutant did not protect goats against wild-type K10 and was able to persist viable in the studied tissues, highlighting one of the issues regarding live-attenuated vaccines. On the other hand, a K10 ΔrelA mutant has demonstrated the capacity of eliciting a cytotoxic T-cell response when orally inoculated, and reduced tissue colonization in goats and cattle (22–24). Similarly, an oral live-attenuated MAP A1-157 ΔBacA has been very recently tested in a short-term study in calves, inducing a proinflammatory profile in peripheral PBMCs and reducing Map burden in some tissues (25). However, these studies lasted under 3 months after the Map challenge, and though a reduction in bacterial load was observed, no vaccinated animal reached sterile immunity. In addition, different recombinant proteins and protein cocktails have failed to offer complete protection in both murine and goat models (26, 27). Well-designed DNA vaccines have similar advantages to subunit vaccines but share limited immunogenicity (28), a few of them, encoding different Map, antigens have also been studied in murine and sheep models, offering variable levels of protection (29, 30). To summarize, all the alternatives tested do not outperform inactivated vaccines in terms of protection.
Among other different strategies, alternative to the currently available vaccines, recent studies have explored the combination of different adjuvants (31–33) and non-subcutaneous administration routes (33–35). Regarding the latter, the oral (36–42) and intradermal (43–45) vaccination routes have been studied in mycobacteria species such as M. bovis and M. tuberculosis mainly in ruminants and several wildlife species, as well as in mice and humans, respectively. The oral route has been proposed to achieve an increased activation of mucosal immunity, as this is the entry site for Map. Studies have shown that it may stimulate the peripheral immune response to a lesser extent which could be advantageous as it would not interfere with the diagnosis of PTB by ELISA, and it could also decrease the interference with bTB diagnostic tests (35). Similarly, it has been proven that oral vaccination with inactivated M. bovis does not interfere with bTB diagnosis in cattle (46) and goats (39). In contrast, regarding intradermal vaccination, the dermis and epidermis are rich in antigen-presenting cells, and therefore, this delivery route should be more efficient at eliciting immune responses with smaller amounts of antigen (47). This route is widely employed in human tuberculosis vaccination (43–45) and has been shown to be more effective than intramuscular or subcutaneous vaccination against other diseases caused by intracellular pathogens, such as influenza virus and Hepatitis B virus (48, 49). Additionally, intradermal delivery is gaining attention due to the development of microneedle and needle-free injection devices, which decrease the risk of disease transmission, and the injection site lesions eliminate the risk of residual needles and improve animal welfare (43–45, 50).
Regarding experimental trials with non-subcutaneous vaccines against PTB in ruminants, several oral live-attenuated vaccines have failed to protect sheep (51) or goats (21, 34) from infection. Nevertheless, orally administered, whole-cell, inactivated, vaccines have been recently tested in the rabbit model: a non-adjuvanted vaccine (35) and one adjuvanted with Quil A® (33), have been shown to exert protection against Map infection. However, to our knowledge, no intradermal PTB vaccine has been tested in ruminants up to date. In rabbits, an inactivated, non-adjuvanted whole-cell intradermal vaccine against Map induced a higher bacterial clearance and less cross-reactivity with the tuberculin skin test than Silirum® and was more protective against infection than a similar oral vaccine in rabbits (35).
Based on recent promising results, alternative immunization routes could potentially overcome some of the disadvantages of the currently available subcutaneous PTB vaccines. However, Map infection is hard to model (52) due to several factors like Map strain (53), passage level (54) host species and breed (55), exposure age (56), and the long incubation period of the disease, all of which have a great influence on disease outcome. Because of this, there is still a lack of research in this field, particularly on ruminants whose susceptibility and immune response against Map may differ from that of laboratory animals such as rabbits or mice (21). On this basis, the main objective of the present study was to test the efficacy of two whole-cell, inactivated vaccines against Map, administered through the intradermal and oral routes, and compare it with a commercial subcutaneous vaccine in goats (Gudair®, CZ Vaccines, Porriño, Spain).
2 Materials and methods
2.1 Animals and experimental design
A total of 28, 1-month-old goat kids of the Murciano-Granadina breed, randomly selected from a commercial flock without a previous history of PTB or TB were used in this study. Animals were kept in the experimental facilities of the Instituto de Ganadería de Montaña (ULE-CSIC). Before the study, all animals were tested negative for both PTB and tuberculosis, using the PTB antibody ELISA (ID Screen® Paratuberculosis Indirect, IDvet), and the WOAH-validated interferon-γ release assay (BOVIGAM™ TB Kit) against both avian (PPDa) and bovine (PPDb) protein derivatives (57).
After a 2-week adaptation period, animals were first divided into four groups: oral vaccine (OV) (n = 6), intradermal vaccine (IDV) (n = 6), subcutaneous vaccine (SCV) (n = 6), and non-vaccinated (NV) (n = 10) (see Figure 1). Animals of the three first groups were vaccinated using different inoculation routes, and the last group was not vaccinated. Subsequently, 1-month post-vaccination (1 mpv), half of the goats immunized with each different vaccine (n = 3), and seven goats of the non-vaccinated group were challenged with Map (Figure 1). Before the experimental challenge, the orally vaccinated animals were kept isolated from the others. Afterward, challenged and non-challenged groups were housed in separate areas. Details on the vaccines and challenge inoculum composition and administration are indicated in the next subsection. This study aimed to include non-challenged, vaccinated groups, to enable the comparison of the immune responses elicited by the different immunization routes in non-challenged animals, something that was not possible in previous studies studying alternative vaccination routes (21, 33–35). This, and the logistical limitations, conditioned a small sample size of the groups.
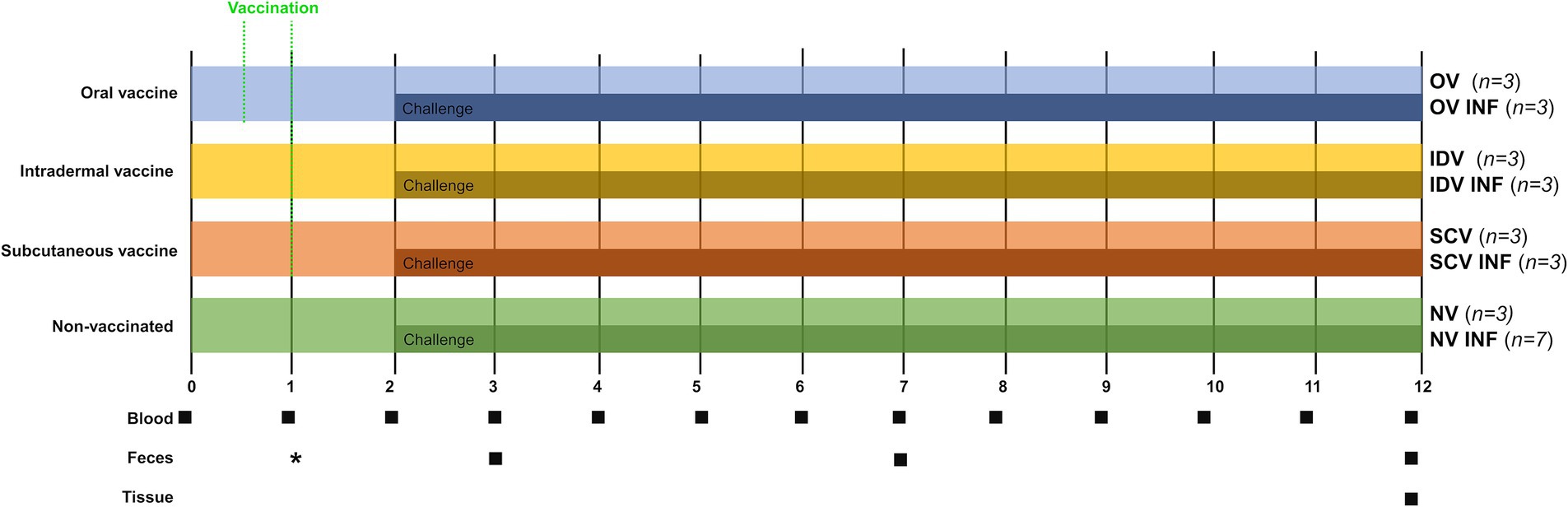
Figure 1. Experimental design scheme. After a 2-week adaptation period, goat kids were divided into four groups and vaccinated through different routes [oral (OV), intradermal (IDV), and subcutaneous (SCV) or left unvaccinated (NV)]. Experimental (OV and IDV) and commercial vaccines (SCV—Gudair®) were used. One month after vaccination, three or seven animals from each group were orally challenged with Map (INF). A total of eight groups were formed: OV (oral vaccine), OV-INF (oral vaccine, challenged), IDV (intradermal vaccine), IDV-INF (intradermal vaccine, challenged), SCV (subcutaneous vaccine), SCV-INF (subcutaneous vaccine, challenged), NV (non-vaccinated), and NV-INF (non-vaccinated, challenged). Squares indicate sampling time points (blood, feces, and tissues) from the beginning of the study to 12 months. *Fecal samples to assure the vaccine reached the intestine were taken from the orally vaccinated animals at 3, 7, and 14 dpv.
The total duration of the experiment was 12 months. Blood sampling was performed monthly throughout the entire experiment, for the determination of the specific peripheral immune response. The injection sites of the IDV and SCV animals were also checked throughout the study. Fecal samples were taken 1, 5, and 10 months post-infection for Map fecal shedding quantification through qPCR, and at this time point, fecal consistency was checked. Additionally, fecal samples from the OV group were also collected at 3, 7, and 14 days post-vaccination (dpv). At the end of the study, all animals were euthanized, and tissue samples were taken for Map detection and lesion assessment (Figure 1).
Goat handling and blood sample collection were carried out in accordance with European Union legislation (Law 6/2013), concerning animals, their exploitation, transportation, experimentation, and sacrifice; R. D. 118/2021 for the protection of animals employed in research and teaching; Directive 2010/63/UE, related to the protection of animals used for scientific goals. All the procedures were approved by the corresponding animal welfare body (OEBA) and the Consejería de Agricultura y Ganadería de la Junta de Castilla y León (authorization code ULE-02-2021). All animals used in this study were handled in strict accordance with good clinical practices, and all efforts were made to minimize suffering.
2.2 Vaccines and challenge inoculum
Three different administration routes using the same Map strain (316F) with equivalent antigen quantity, but different adjuvants, were employed in this study. Both experimental vaccines contained 12.5 mg of antigen per dose to match Gudair® antigen quantity as calculated by wet weight following guidelines for PTB vaccination trials (52). The experimental oral vaccine (OV) was composed of chemically inactivated Map and 5 mg of the adjuvant Quil A® (InvivoGen, San Diego, CA, United States), diluted in 10 mL of physiological saline solution (B. Braun, Melsungen, Germany) and was administered orally using an automated syringe in two doses separated by 14 days. The efficacy of this vaccine and its effects on mucosal, trained, and innate immunity have been previously studied in the rabbit model (33). The experimental intradermal vaccine (IDV) was composed of chemically inactivated Map diluted in 0.4 mL of physiological saline solution and a single dose was administered, using a 25-G needle. The dose was inoculated in four previously shaved injection sites (0.1 mL per injection), two at each side of the dorsal thoracic area. The efficacy of this vaccine has been studied in the rabbit model, and its interference in the diagnosis of bTB through the skin test has been studied in guinea pigs (35). Finally, Gudair®, a commercial subcutaneous vaccine (SCV), composed of heat-inactivated Map, and a mineral oil-based adjuvant (containing Marcol 52, Montanide 80, and Montanide 103) was injected subcutaneously in a single 1 mL dose in the right ventral abdominal area, using an 18-G needle, as per manufacturer’s instructions.
The challenge inoculum, a low-passage type C field isolate of Map (strain 832) (58, 59), was grown on Middlebrook 7H9 broth enriched with 10% oleic acid–albumin–dextrose–catalase (OADC) and 2 mgL−1 Mycobactin J (7H9 OADC MJ) for 3 weeks at 37 ± 1°C. Then, cultures were harvested by centrifugation at 3,000 g for 10 min, and bacterial pellets were washed two times and resuspended in PBS. To disrupt bacterial clumps, the resultant suspension was passed up and down through a 27-G needle several times and vortexed. Bacterial concentration was estimated by optical density (OD) and colony-forming units (CFU) of 10-fold serial dilutions plated onto agar-solidified 7H9 OADC MJ. Finally, suspensions were adjusted to 2 × 1010 CFUs/ml, maintained at 4°C throughout the challenge period, and disrupted before oral inoculation as mentioned above. Specifically, each animal from the experimentally infected groups (INF) was orally inoculated with 8 × 109 Map CFUs diluted in 10 mL of PBS, using an automatic syringe. The inoculation was performed four times separated by 3 days, as previously described by Fernández et al. (53), for a total dose of 3.2 × 1010 CFUs. Ten milliliters of PBS was administered orally to non-infected animals at the same time.
2.3 Sampling
Blood samples were monthly collected from the jugular vein into Vacutainer® tubes (Becton Dickinson and Company, Erembodegem, Belgium) with lithium heparin and without anticoagulant. Then, heparinized samples were processed immediately for IFN-γ release test in response to PPDa and PPDb. Non-heparinized samples were processed for the study of the Map-specific antibody dynamics in the sera samples. Fecal samples were obtained per rectum using individual single-use polyethylene gloves and frozen at −80°C for quantification of Map-DNA through real-time quantitative polymerase chain reaction (qPCR).
At 11 mpv, all animals were humanely euthanized by deep sedation with xylazine (Xilagesic®, Calier, Barcelona, Spain) and subsequent intravenous injection of embutramide, mebezonium iodide, and tetracaine hydrochloride, (T61®, MSD Animal Health) followed by exsanguination. Then, a regulated, orderly, and complete necropsy was performed, and gross lesions were annotated. Immediately after gross examination of the viscera, a distal jejunal Peyer’s patch (DJPP), a draining jejunal lymph node (JLN), and the distal ileum (DI) were collected and frozen at either −20°C for Map culture or −80°C for quantification of Map-DNA through qPCR. In addition, tissue samples from the ileocecal valve, ileum [distal (DI), medial (MI), and proximal zones (PI)], jejunum [distal (DJ), medial (MJ), and proximal zones (PJ)], and Peyer’s patches [from the distal (DJPP), medial (MJPP), and proximal jejunum (PJPP)], together with ileocecal (ICLN), caudal mesenteric (CMLN), and jejunal (JLN)—lymph nodes, were taken, thoroughly washed in PBS, and taken into buffered formol saline fixative for histological examination.
2.4 Determination of the peripheral immune responses
To measure the Map-specific IgG in serum samples, the ID Screen® Paratuberculosis Indirect (IDvet, Grabels, France) ELISA test was employed. Interpretation was performed following manufacturer’s instructions. Briefly, the optical density was measured spectrophotometrically at a wavelength of 450 nm (OD450) and the results were expressed as a ratio of the mean OD450 of each sample sera duplicates and the mean OD450 of the positive control sera duplicates from each plate (OD450 ratio).
For the determination of the specific cellular immune responses, within 3 h of blood collection, 1.5 mL aliquots of the heparinized blood samples were incubated in duplicate, in 24-well sterile plates, with either 100 μL of sterile phosphate-buffered saline (PBS), PPDa or PPDb (CZ Veterinaria, Porriño, Spain), at a final concentration of 20 μg/mL. After incubation (20 h at 37°C, 5% CO2), plates were centrifuged at 750 g for 15 min, and plasma was collected and stored at −20°C (60). Then, the assay for IFN-γ determination BOVIGAM® TB Kit (Thermo Fisher Scientific, Waltham, MA, United States) was carried out in the plasma samples following the manufacturer’s instructions, interpreted as previously described (60), and results were expressed as a quotient (OD index), between the mean OD of the PPD-stimulated blood plasma and the mean OD of the plasma from the blood incubated with PBS.
Additionally, the standard interpretation of the BOVIGAM™ TB Kit was performed, to evaluate the possible interference of the different immunization strategies or the Map challenge with bTB diagnosis. According to the manufacturer’s instructions, PPDb-stimulated blood plasma with an OD value greater than 0.100 above that of both the PPDa and nil (PBS) antigen samples is indicative of the presence of M. bovis infection.
2.5 Histopathological examination and lesion classification
The formaldehyde-fixed tissue samples were conventionally processed for paraffin embedding and stained with hematoxylin–eosin (HE) for histopathological examination. Following the previously established classification for small animals, lesions consistent with PTB infection were classified as focal, when granulomas were restricted to the lymphoid tissue of PP; multifocal, when small granulomas were also located in the lamina propria; and diffuse, when the granulomas were widespread throughout the intestinal mucosa in some of the sections (19, 61). Additionally, three cross sections from the ileum (DI, MI, and PI), jejunum (DJ, MJ, and PJ), JPP (DJPP, MJPP, and PJPP), one longitudinal section from the ICV, and one section from each lymph node (ICLN, CMLN, and JLN) were selected. Then, the whole sections from ileal, jejunal, JPP, and ICV were analyzed, and the total number of granulomas per section was recorded. In the lymph nodes, a total of 5.46 mm2 of cortical area was analyzed per section (10 random 100× fields, Nikon® Eclipse E600 microscope, coupled with a Nikon® DS-Fi1 digital camera—Nikon, Tokyo, Japan).
2.6 Tissue and fecal detection and quantification of Map-DNA through qPCR
DNA was extracted from 50 mg of the homogenized tissue samples (DJPP, JLN, and DI) using the Maxwell® RSC 16 Tissue DNA Purification Kit with the Maxwell® RSC 16 Instrument (Promega, WI, United States), following the manufacturer’s instructions. Fecal DNA was extracted from 200 to 250 mg of homogenized fecal samples (three to four fecal pellets) using the Maxwell® RSC Fecal Microbiome DNA Kit, with the Maxwell 16 Instrument (Promega, WI, United States), following the manufacturer’s instructions, including the optional bead-beating steps, which were performed using the Fisherbrand™ Bead Mill 24 (Thermo Fisher Scientific, Waltham, MA, United States). Thereupon, DNA was quantified using QuantiFluor™ ONEdsDNA System Kit (Promega, WI, United States) and Quantus™ Fluorometer (Promega, WI, United States). Extracted DNA was diluted to 50 ng · μL−1 for tissue samples and 20 ng · μL−1 for fecal samples and stored at −80°C until qPCR was performed, and a total of 100 ng of DNA were added to each qPCR reaction.
The primers employed for Map IS900 qPCR were: forward (MP10-1, [5′-ATGCGCCACGACTTGCAGCCT-3′]) and reverse (MP11-1, [5′-GGCACGGCTCTTGTTGTAGTCG-3′]) (62). The quantification was performed as described by Arteche-Villasol et al. (63), a 10-fold diluted standard curve was constructed using Map genomic DNA, obtained from 108 CFUs of Map K10 strain, ranging from 10−1 to 10−8 ng of total DNA mixed with 20 ng of tissue DNA from a non-infected animal per reaction. Samples were analyzed in triplicate and considered as positive when the dissociation peak (Tm) was 89.1 ± 1°C and threshold cycles (Ct) were ≤ 37. The qPCR results were analyzed using 7,500 Software v2.0.6 (Applied Biosystems™). Map-DNA concentration (fg Map-DNA/g of tissue or feces) was calculated by interpolation of their Ct values with the standard curve, and results were expressed as the mean quantity of the triplicates.
2.7 Map detection through Ziehl–Neelsen and immunohistochemistry
The Ziehl–Neelsen (ZN) technique for acid-fast bacilli (AFB) detection and an immunohistochemical (IHC) staining against Map were used in formaldehyde-fixed paraffin-embedded tissue samples, for detecting Map bacilli and assessing its distribution. One DJPP, one JLN, and one DI section were employed for each technique. These tissues were chosen as they showed the most severe lesions (a mean of 53.6, 67.3, and 57.4 granulomas per section/area, respectively, in the challenged animals). The ZN staining was performed conventionally (19, 64), and sections were classified as positive or negative, based on the presence of AFB in the cytoplasm of macrophages.
The IHC staining was performed on 3-μm-thick tissue sections, placed onto poly-L-lysine-coated slides (SuperFrost Plus Adhesion slides, Thermo Fisher Scientific, Waltham, United States), as previously described (64). After deparaffinization and hydration, sections were washed two times using wash buffer (Agilent Technologies, Santa Clara, United States), for 5 min. Then, endogenous peroxidase was blocked by immersion of the sections into a 3% H2O2 in methanol for 30 min in darkness at room temperature. After washing two times, sections were incubated with an in-house rabbit anti-Map antibody (64), diluted 1:2000 in antibody diluent (Agilent Technologies, Santa Clara, United States), overnight, at 4°C, in a humidified chamber. After washing, sections were incubated for 40 min at room temperature with a secondary polyclonal antibody, horseradish peroxidase labeled polymer (Agilent Technologies, Santa Clara, USA), and after washing, antibody localization was determined using 3,3-diaminobenzidine (Agilent Technologies, Santa Clara, United States) as a chromogenic substrate for peroxidase. Sections were counterstained with Mayer’s hematoxylin for 10 s. Appropriate species- and isotype-matched immunoglobulins were used as negative controls. An ileal sample from a naturally Map-infected sheep with diffuse lesions was used as a positive control. IHC sections were classified as positive or negative, based on the presence of positively stained macrophages. Both ZN and IHC sections were observed under the Nikon® Eclipse E600 light microscope, coupled with a Nikon® DS-Fi1 digital camera (Nikon, Tokyo, Japan).
2.8 Map culture
DJPP, JLN, and DI samples from each animal were cultured for viable Map detection. Briefly, 1 g of each tissue was decontaminated with 19 mL of 0.8% hexadecylpyridinium chloride (Sigma-Aldrich, St. Louis, MO, United States) and homogenized in a stomacher blender (Masticator, IUL) for approximately 30 s. After 18 h of decontamination, 200 μL of the suspension was dispensed per tube containing Herrold’s egg yolk medium supplemented with sodium pyruvate and mycobactin J (MJ). Cultures were incubated at 37 ± 1°C, and growth was checked by examination under a stereoscopic microscope after 8, 12, 16, and 20 weeks post-inoculation. Cultures were considered positive if one or more characteristic Map colonies were observed in any tube. Colonies isolated were confirmed by a real-time multiplex PCR detecting IS900 and ISMap02 Map sequences (65).
2.9 Statistical analysis
The normality of the data was assessed using the Shapiro–Wilk test; all variables were non-normally distributed, so non-parametric tests were employed for the analysis. The antibody OD450 ratio and the avian and bovine indices were logarithmically transformed [log2(x + 1)]. Data collected at a single time point (PCR quantification of the tissue and fecal bacterial load, Map detection through culture, IHC results, and granuloma counts) were analyzed using Pearson’s chi-squared test of independence or Kruskal–Wallis test. Data collected at multiple time points (antibody OD450 ratio, the avian and bovine indices, and the Map fecal shedding quantification) were analyzed using the Friedman test. If statistically significant differences were detected, post-hoc tests were used to perform pairwise multiple comparisons: the Wilcoxon test (with Bonferroni–Holm correction for differences in total granuloma counts between groups) or the Dunn test. The Spearman rank test was performed to assess the correlation between the results of the different assays used for Map detection, and the results are included in Supplementary file S1. All statistical analyses were performed with the R software version 4.2.3 (66). The statistical packages psych (2.4.3), corrplot (0.92), rstatix (0.7.2), ggpubr (0.6.0.999), lubridate (1.9.2), forcats (1.0.0), stringr (1.5.0), dplyr (1.1.1), purrr (1.0.1), readr (2.1.4), tidyr (1.3.0), tibble (3.2.1), ggplot2 (3.4.2), and tidyverse (2.0.0) were used in the different statistical tests.
3 Results
3.1 Peripheral humoral and cell-mediated immune responses
Results from the indirect antibody ELISA are represented in Figure 2A, and its statistical analysis is provided in Supplementary Table S1. A rapid increase in the mean Map-specific antibody levels occurred after subcutaneous vaccination in the animals from the SCV and SCV-INF groups, with no apparent influence of the Map challenge in the latter group. A delayed, slower, and sustained increase took place in the remaining challenged groups (OV-INF, IDV-INF, and NV-INF), but no significant differences were detected between them. Regarding their non-challenged counterparts, animals in the OV and IDV groups did not show any increase in Map-specific antibody levels despite vaccination. In addition, animals in the NV group did not produce specific antibodies against Map.
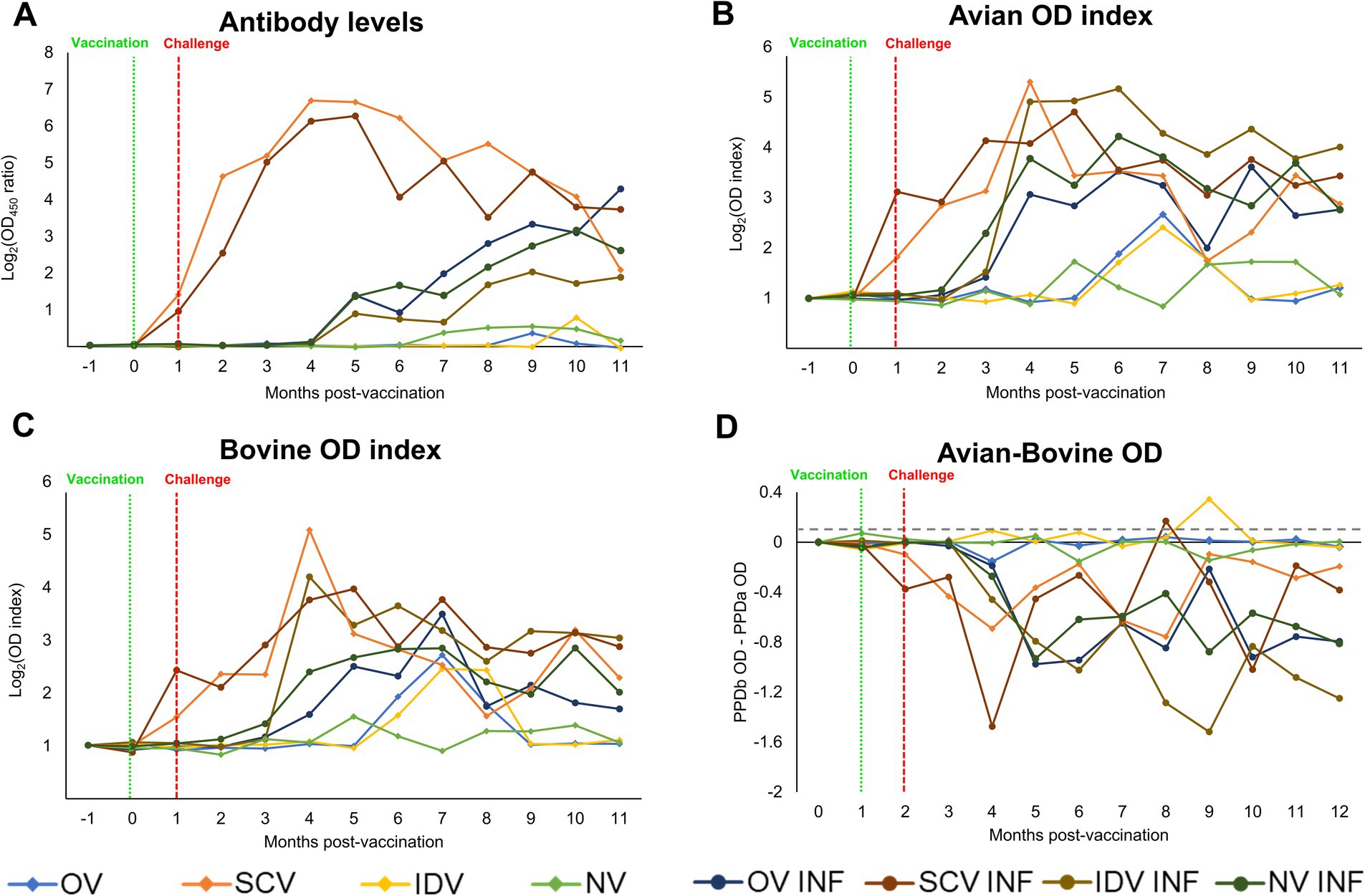
Figure 2. Kinetics of the peripheral immune response. (A) Dynamics of the specific anti-Map antibody response through the experiment, expressed as the mean Log2 (OD450 ratio) of each group. (B) Kinetics of the IFN-γ production by whole blood stimulated with PPDa. (C) Kinetics of the IFN-γ production by whole blood stimulated with PPDb, expressed as the mean avian and bovine Log2 (OD index) of each group. (D) bTB interference, calculated by subtracting the OD of the PPDa from the PPDb stimulated samples, results are expressed as a mean, and the gray horizontal dotted line represents the cutoff ratio of the BOVIGAM™ test (0.100). The experimental groups were as follows: OV (oral vaccine), OV-INF (oral vaccine, challenged), IDV (intradermal vaccine), IDV-INF (intradermal vaccine, challenged), SCV (subcutaneous vaccine), SCV-INF (subcutaneous vaccine, challenged), NV (non-vaccinated), and NV-INF (non-vaccinated, challenged). The vertically dotted red line represents the vaccine administration time-point, and the vertically dotted green line represents the challenge time-point. The results from the statistical analysis are shown in Supplementary Table S1.
The cellular immune responses (Figures 2B,C; Supplementary Table S1) were in line with the humoral response for most groups; in the SCV and SCV-INF groups, the avian and bovine index showed an early increase after vaccination. In contrast, in the other challenged groups (OV-INF, IDV-INF, and NV-INF), this increase in the cellular responses was slightly delayed, starting 2 to 3 months after the infectious challenge, and was sustained up to the end of the experiment. However, the OV and IDV groups behaved differently, with only subtle and time-limited cellular responses initiating at a late time point of approximately 5–6 months after vaccination. The mentioned increases in the avian and bovine OD index values were significant (p < 0.05), with respect to the basal levels, at some point in all groups except for the NV group. This increase took place sooner in the challenged groups, and overall, in all groups, higher mean values were reached for the avian index. Throughout the experiment, at several time points, all the challenged groups, except for the SCV-INF, showed significantly higher avian and bovine indexes (p < 0.05) than their respective only challenged counterparts.
Following the standard interpretation of the BOVIGAM™ TB Kit, several animals would have been considered positive for bTB (PPDb OD–PBS OD ≥ 0.1 and PPDb OD–PPDa OD ≥ 0.1) at some point during the experiment. Specifically, one from the SCV group (animal 52 at 2 MPI and 4 MPI), two from the IDV group (animals 31 and 32 at 7 MPI), all three animals from the SCV-INF group (animal 61 at 4 MPI, 62 at 6 and 7 MPI, and 63 at 6 MPI), and one animal from the NV-INF group (animal 81 at 8 MPI). The quantitative results of the PPDb OD-PPDa OD subtraction are represented in Figure 2D.
3.2 Clinical and pathological findings
In the SCV group, vaccine injection site nodules, which were already present since the first few weeks after vaccination, persisted at the time of necropsies. However, animals from the IDV group did not develop any noticeable lesions at the injection site during the experiment. Fecal consistency remained unaffected in all animals, exhibiting firm, well-formed pellets at all the selected time points (1, 5, and 10 mpi).
Regarding gross lesions, none of the non-challenged animals showed any lesion during necropsies, whereas gross lesions compatible with PTB were observed in the ileum, jejunum, and mesenteric lymph nodes of some of the challenged animals except for the SCV-INF group. The most affected group was the non-vaccinated (three of seven animals), whereas only one of three animals showed gross lesions in the remaining groups (OV-INF and IDV-INF) (Table 1). The main gross lesions observed were a diffuse thickening of the ileal and/or jejunal mucosa (Supplementary file S2A) and cortical enlargement of the mesenteric lymph nodes (Supplementary file S2B). In some of the challenged goats, in addition to the diffuse thickening, congestion was observed in some of the Peyer’s patches (Supplementary file S2D) and one of the animals from the OV-INF group presented multiple foci of caseous necrosis, located in the Peyer’s patches (Supplementary file S2C).
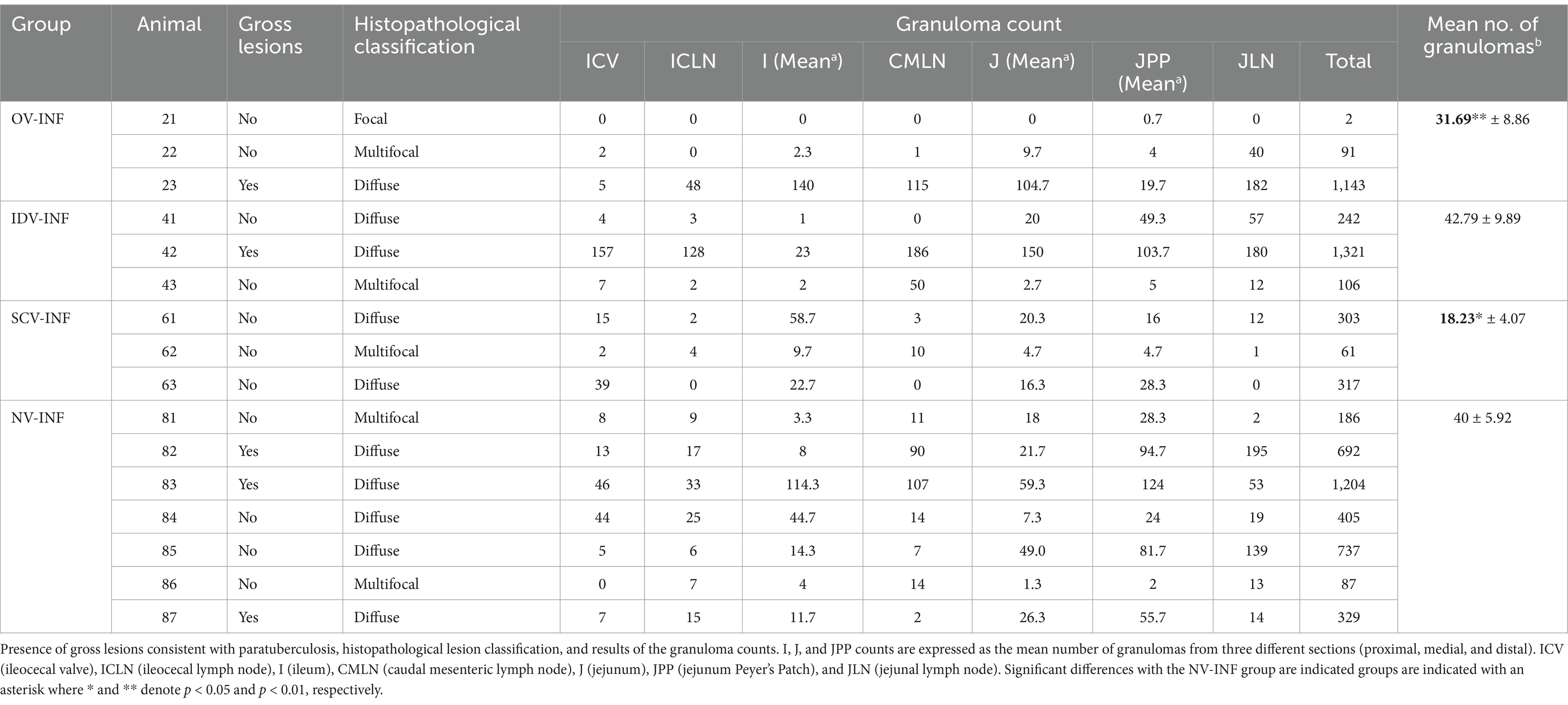
Table 1. Lesion assessment in infected groups.
Microscopic lesion classification and granuloma counts for each challenged animal and tissue are also indicated in Table 1. All challenged animals developed microscopic granulomatous lesions despite vaccination; however, a wide range of lesion severity was observed (Figures 3A–D), both between individuals and between different tissues in the same individuals (Table 1). The mean number of granulomas was significantly lower in the OV-INF (p < 0.01) and SCV-INF (p < 0.05) groups with respect to the NV-INF. Additionally, the number of granulomas present in the ICLN and JLN lymph nodes from the animals in the SCV-INF group was significantly lower (p < 0.05) than that found in the animals from the NV-INF group. Overall, microscopic lesion severity was high; of the 16 challenged animals, 10 developed diffuse lesions and 5 developed multifocal lesions, and the tissues with the highest number of granulomas were DJPP, JLN, and DI, with a mean of, 67.3, 57.4, and 53.6 granulomas per section/area, respectively, in the challenged animals.
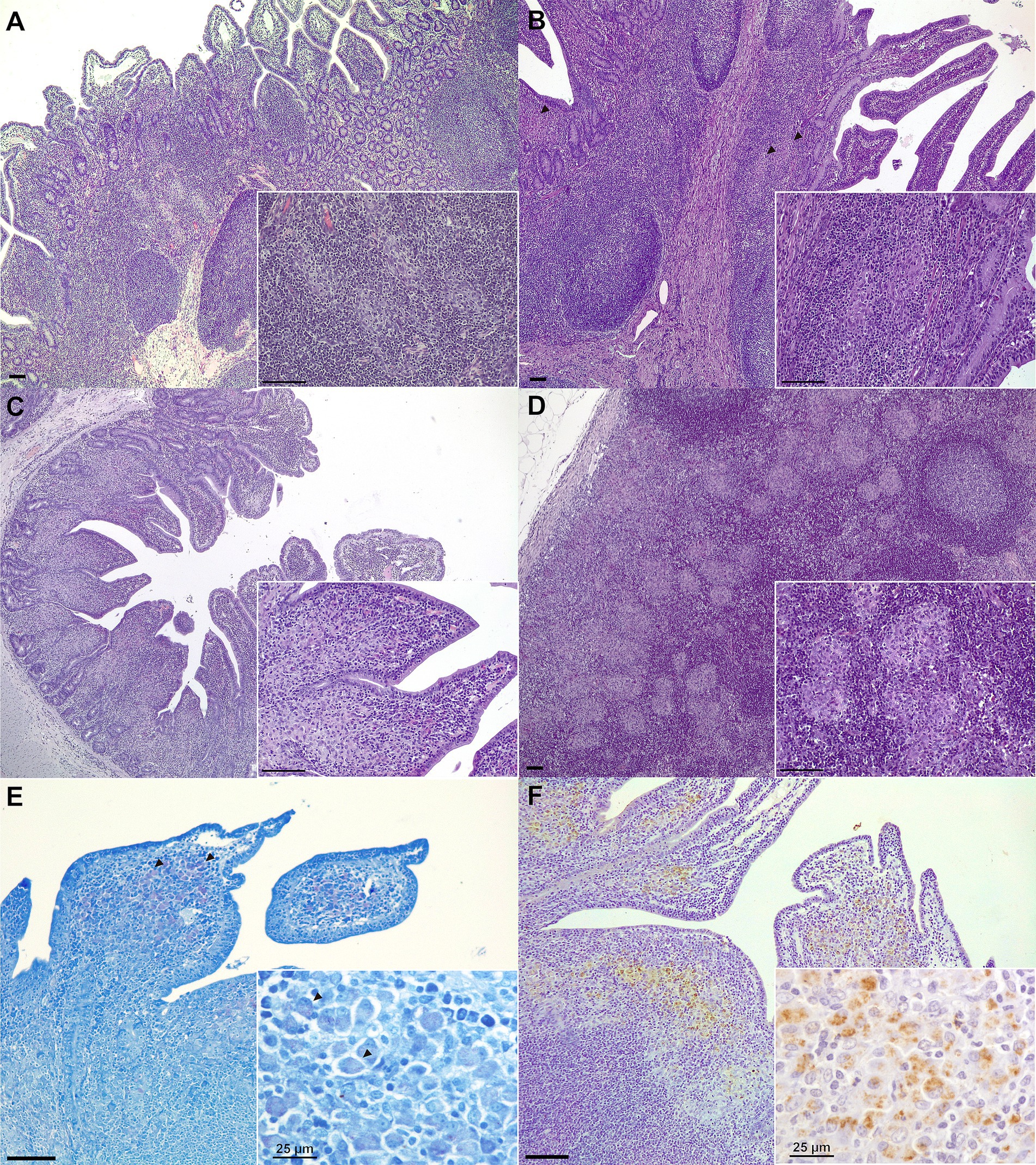
Figure 3. Representative microscopic lesions and Map detection in tissue samples. (A) Focal lesion: multiple well-demarcated granulomas located in the interfollicular area of the Peyer’s patches (medial ileum, Goat 64, SCV-INF group); (B) Multifocal lesion: granulomas (arrowheads) can be seen in both the lymphoid tissue and lamina propria (medial jejunum Peyer’s patch, Goat 22, OV-INF group); (C) Diffuse lesion: the lamina propria is a diffuse granulomatous infiltrate affects the lamina propria (medial jejunum, Goat 42, IDV-INF group). (D) Multifocal to coalescing granulomatous foci present in the cortex of a jejunal lymph node (goat 83, NV-INF group). Representative images of Map-positive samples as detected through: (E) Ziehl–Neelsen staining. Some acid-fast staining can be observed in some of the macrophages present in the apex of the villi (arrowheads) (distal jejunum Peyer’s patch, Goat 82, NV-INF group). (F) Immunohistochemistry. Macrophages positive to Map immunostaining are observed in the diffuse granulomatous infiltrate affecting the lamina propria over the lymphoid tissue of a distal jejunum Peyer’s patch (distal jejunum Peyer’s patch, Goat 82, NV-INF group). Unless indicated otherwise, scale bars equal to 100 μm.
3.3 Map fecal shedding
After oral vaccination, Map-DNA pass-through was detected in at least one sampling during the first 14 dpv in five out of six vaccinated animals. By 3, 7, and 14 dpv, Map-DNA was detected in four of six, one of six, and two of six of the orally vaccinated animals, respectively, in each time point. The mean DNA quantified was low in all samplings: 0.97 ± 0.34, 0.22 ± 0.22, and 0.57 ± 0.37 fg of Map-DNA/g feces, respectively.
After the challenge, the evolution of Map fecal shedding can be seen in Table 2. Before infection, all fecal samples from non-challenged animals were negative throughout the entire experiment, except for the OV group as mentioned above. After infection, in challenged animals, inter- and intraindividual variability in Map fecal shedding was very high, and no significant differences between challenged groups were detected. The number of animals shedding and the mean Map-DNA quantity shed progressively increased throughout the experiment; this increase was only significant in the NV-INF group (p < 0.05). Specifically, at 1 month post-infection (mpi), two of three animals from the IDV-INF group, and four of seven from the NV-INF group were positive and the remaining were negative. By 5 mpi, two of three animals from the OV-INF group, one of three animals from the IDV-INF group, two of three animals from the SCV-INF group, and four of seven animals from the NV-INF group were positive by qPCR. By the end of the experiment (10 mpi), all fecal samples from challenged animals, except for one animal from the NV-INF group and another from the SCV-INF group, were positive to Map.
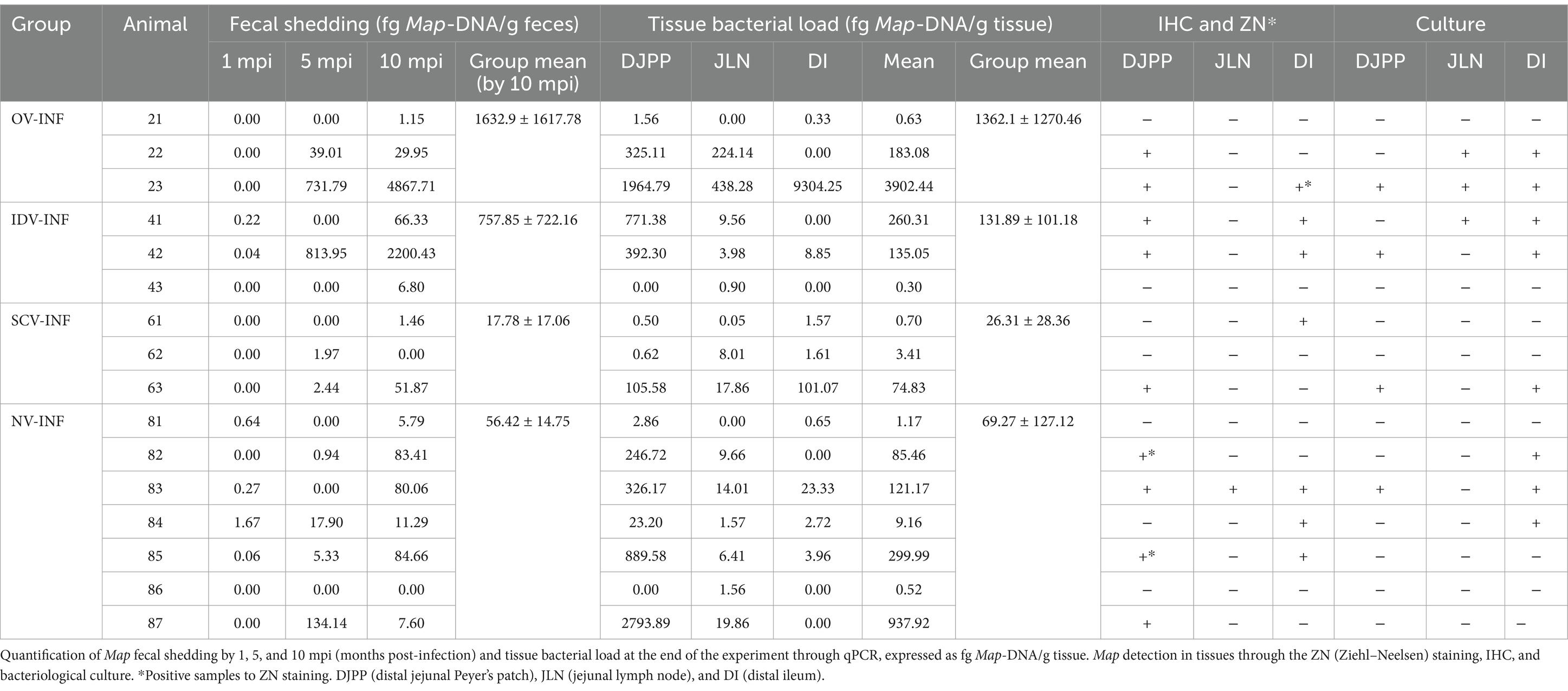
Table 2. Map detection and quantification in feces and tissues in challenged animals.
3.4 Map detection in tissues
The bacterial load per tissue, as assessed through qPCR as well as ZN, IHC, and Map-culture results can be seen in Table 2. The tissue bacterial load results align with the fecal shedding data, as Map was detected via qPCR in at least one tissue sample from all challenged animals (Table 2). In addition, in accordance with the microscopic lesions observed, the samples with a higher Map-DNA load in most animals (11/16) were the DJPP, and the SCV group showed the lowest mean Map load. However, the Map load varied greatly both between samples and animals, and no significant differences were observed in tissue distribution or between groups.
In this line, Map was detected through bacteriological culture, in at least one tissue, in one of three animals from the SCV-INF group in two of three animals from the OV-INF and IDV-INF groups and in three of seven animals from the NV-INF group (results are indicated in Table 2), but no significant relation between culture detection of Map and the vaccine employed in challenged animals was detected by the chi-square test (p > 0.05).
Representative examples of ZN and Map IHC positive staining can be seen in Figures 3E,F, respectively, and the results for the individual animals and tissues are collected in Table 2. AFB was only observed in a small number of macrophages, in extensive areas of granulomatous infiltrate in samples from one of three and two of seven animals from the OV-INF and NV-INF groups, respectively. Map was detected through IHC mostly in DI and DJPP sections; positively stained macrophages were mainly present in the diffuse granulomatous infiltrates and large granulomas. Two out of three animals from the OV-INF and IDV-INF groups, 1/3 from the SCV-INF group, and 6/7 from the NV-INF group were positive to Map IHC. No significant relation between the immunohistochemical detection of Map and the vaccine employed in challenged animals was detected by the chi-square test (p > 0.05).
The detection of Map in feces and tissues, through the different techniques employed, showed strong correlations in most cases (Supplementary Figure S1). An interesting exception was the results from the JLN samples, as most of them were negative to IHC and culture.
4 Discussion
Alternative routes of vaccination like the oral and intradermal are new strategies that could aid to circumvent some of the drawbacks of the currently available subcutaneous vaccines against paratuberculosis. Few studies have been made on ruminants on vaccines employing the oral route and most of them used live-attenuated, non-adjuvanted vaccines (21, 34, 51). Furthermore, the intradermal route of vaccination against paratuberculosis remains unexplored in ruminants. Thus, the present study has compared the effect on the specific peripheral immune responses and the protective efficacy of two experimental PTB vaccines, administered through the oral and intradermal routes, with that of a commercial SCV. As expected, the SCV vaccine induced strong peripheral immune responses which remained unaffected by the subsequent Map challenge. Conversely, mild, delayed, and short-spanned peripheral cellular responses were induced by the OV and IDV, suggesting its potential of not interfering with bTB diagnosis. However, although none of the three vaccines conferred complete protection against the experimental Map challenge, the SCV (Gudair®) was the most effective at reducing bacterial load, fecal shedding, and lesion severity, whereas the OV only slightly reduced lesion severity when compared to the NV-INF animals and the IDV failed to reduce any of these protection parameters.
The efficacy of these immunization strategies has been previously tested in rabbits (33, 35), and its interference with bTB diagnosis has been assessed in guinea pigs (35). In those experiments, the IDV induced a higher bacterial clearance and lower bTB diagnosis interference than the SCV control. Additionally, the OV-induced mucosal immunity (significantly increasing TNF, IL-1β, IL-10, IL-12B, and IL-23A expression in the GALT) (33) and a comparable OV, albeit without adjuvant, induced similar clearance to that of the control vaccine while not interfering with bTB diagnosis (35). However, it is important to emphasize that experimental results obtained in laboratory animal models of paratuberculosis vaccination do not always correlate with those observed in ruminants (21). Because of this, studies in the relevant host are necessary to assess vaccine efficacy, and because of its pathogenesis, long-term studies are also necessary to study Map infection progression. Cattle require significantly higher upkeep and larger facilities than small ruminants, whereas goats are more susceptible to developing clinical PTB than sheep, particularly to type C Map strains (54, 55, 67, 68). Additionally, goats are also more susceptible to bTB infection (69, 70). Therefore, the caprine model was considered the most suitable for studying the efficacy of PTB vaccination and its interference with bTB diagnosis. Nevertheless, PTB is difficult to model, starting from the challenge inoculum, as culture passages significantly decrease Map virulence (71, 72), and therefore, a common problem faced by experimental infections is the lack of infectivity and virulence of cultured strains (73). In addition, the short time periods used in most experimental settings, of a few months versus the several years that natural infections would require to produce severe lesions and reach the clinical disease stage (68, 74), further complicate the interpretation of the challenge outcome. To solve these challenges, in the present study, we have used a low-passage, type C field Map isolate, at a relatively high dose for the experimental infection and a study duration of 10 months post-challenge. This has resulted in a high infection rate of the challenged animals; however, tissue bacterial load, Map shedding, and lesion severity were highly variable between individuals, which would be in concordance with that observed in natural and experimental infections (12, 75, 76).
The peripheral immune response elicited by PTB infection is characterized by an initial strong cell-mediated immune response, followed by a late, steady increase in the humoral immune response (77). This switch is generally thought to signify a breakdown in disease control (78) and can be seen in the evolution of the response developed by the animals from the infected groups. This pattern of response was followed by the OV-INF and IDV-INF animals but not by their non-challenged counterparts, which suggests that these peripheral responses could be elicited by infection and were not affected by vaccination. The lack of induction of humoral responses could be expected, particularly in oral vaccines, as they primarily stimulate local (intestinal) immunity (79). In this sense, the non-challenged OV- and IDV-vaccinated goats did not produce significant levels of specific IgG at any point in the 10 mpv period. Similarly, in a previous experiment carried out by Hines et al. (34), using live-attenuated oral vaccines in goats, antibody production started 5 months after the Map challenge, which suggests that it was not elicited by vaccination but by the subsequent challenge. In line with this, Arrazuria et al. (35) also observed an absence of humoral responses during 160 dpv, in orally or intradermally vaccinated rabbits.
This humoral immunity has not been traditionally considered necessary for protection against Map infection, although some recent studies point out in the opposite direction (78). On the other hand, the cell-mediated response, and particularly Th1, is widely considered critical in the effective response against mycobacterial infections (80). In our experiment, in the OV and IDV groups, a peak in the peripheral cell-mediated immune responses against avian and bovine PPDs was observed between 6 and 8 mpv. This peak was short-lived, of low intensity, and occurred late with respect to the response induced by subcutaneous vaccination or infection. In this sense when applying the standard interpretation of the BOVIGAM™ test, by 7 MPI, two of the IDV-vaccinated animals would be considered positive to bTB, but no OV, OV-INF, or IDV-INF animal was positive throughout the experiment. Few studies can serve as a comparison; in the mentioned study carried out by Arrazuria et al. (35), for example, the cell-mediated response was only measured through skin testing in the guinea pig model, and in this case, the animals vaccinated through the oral route did not show skin reaction against both PPDa and PPDb, but the intradermal vaccine induced a significant skin reaction against both PPD, although it was lower than that of Silirum®. Furthermore, a study carried out in goats testing the same oral vaccine showed the absence of interference with PPDa and PPDb in the skin test 48 days post-vaccination (81). In accordance with the results presented, oral, whole-cell heat-inactivated bTB vaccines did not induce specific antibody production or cell-mediated immune responses before the M. bovis challenge, in goats (39), cattle (46), red deer (82), or boar (83). As previously stated, no intradermal PTB vaccines have been tested in ruminants; however, the subtle peripheral responses observed contrast with the strong cell-mediated responses induced by several intradermal vaccines against human tuberculosis. Nevertheless, without considering species differences, the formulation and/or posology of these vaccines differ substantially from the one tested in this study, with most studies employing attenuated or vectored vaccines (43, 45) or multiple doses of an inactivated vaccine (84).
The peripheral responses induced by the control SCV were substantially different from those induced by the experimental vaccines; strong humoral and cell-mediated immune responses were detectable early after vaccination in the subcutaneously vaccinated animals, reaching their peak by 3–5 mpv. This is the typical pattern of response induced by Gudair® (85) and other inactivated whole-cell, oil-adjuvanted vaccines like Silirum® (86) and Mycopar® (87). In addition, in contrast with the experimental vaccines, the infectious challenge did not significantly affect the kinetics of the peripheral immune response in the animals of the SCV-INF group. Regarding the interference of this vaccine with bTB diagnosis, as per BOVIGAM standard interpretation, only one SCV animal was positive by 4 MPI, but the three SCV-INF animals were positive at least once during the experiment. This vaccine (88) and PTB exposure or infection (89) are known to interfere with bTB diagnosis. In field conditions, both factors are present, as PTB-vaccinated herds are those where Map is circulating, so vaccinated animals are likely exposed to Map and therefore prone to yielding false positives to bTB diagnostic tests. Overall, regarding the peripheral immune responses, it should be stated that they do not always perfectly align with the mucosal responses (90, 91). In this sense, our general knowledge of the local immunity elicited by PTB vaccination, particularly through the oral route, is still very limited.
Herein, the intestinal gross lesions observed in some of the animals are compatible with typical PTB lesions, including the caseous necrotic areas and lymphangiectasia, which constitute a specific finding of small ruminants, usually found in naturally infected animals (19, 92, 93). These findings suggest the effective establishment of Map infection in some of the challenged animals and the usefulness of the caprine model for the study of PTB. Microscopic lesion classification revealed a wide range of severity, indicating varied individual and tissue-specific responses to infection. Despite this high variability, the overall progressive increase in shedding quantity, and the detection of Map through different techniques and in multiple tissues, further suggest the successful establishment and persistence of the infection in animals from all groups. However, the significantly lower granuloma counts in the OV-INF and SCV-INF groups, when compared to the NV-INF group, indicate potential differences in disease progression or immune control among these groups, in spite of the differences observed in the peripheral immune responses between both. This immune control is further supported in the SCV-INF group, where the lower number of granulomas present in the mesenteric lymph nodes (JLN, CMLN, and ICLN), with respect to the NV-INF group suggests a more effective containment of Map spread in the intestine. In this sense, interestingly, the low number of JLN samples found positive through Map IHC and culture in all groups, suggests that Map integrity was low in this tissue and that a large proportion of the Map-DNA detected in this tissue may pertain to degraded bacteria.
In general terms, the diffuse lesions observed, which are considered the pathological form of patent PTB disease (94), together with the continuous increase in fecal shedding and/or high tissue bacterial load, suggest that the disease is progressing in animals 22, 23, 41, 42, 63, 82, 83, 84, 85, and 87. This would mean that none of the vaccines was able to completely protect animals from PTB infection and lesion development despite differences in bacterial load, shedding, and lesion severity. Similarly, previous studies have demonstrated partial protection against Map or M. bovis infection conferred by OV and IDV, but when compared with parenterally administered vaccines, they were often outperformed. For example, in rabbits challenged with a different, less virulent strain, Silirum® reduced PTB lesion severity more than IDV and OV (35). In goats, two of five live-attenuated OV tested slightly decreased lesion severity, but none of them had a significant effect on Map fecal shedding when compared to non-vaccinated controls, and all were outperformed by Silirum® in both parameters (34). In Eurasian boar, both an inactivated M. bovis vaccine and live BCG, administered orally, reduced lesion severity but both were outperformed by a vaccine administered parenterally (83). In red deer, these vaccines were not compared with a parenterally administered one, but they only conferred partial lesion reduction (95). As mentioned earlier, most of these IDV and OV vaccines induced poor peripheral immune responses. While reducing the diagnostic interferences caused by the peripheral responses induced by subcutaneous vaccines would be beneficial, those findings suggest that eliminating the peripheral responses elicited by vaccination might compromise protection. Further studies are needed to better understand these mechanisms.
Histopathological and tissue bacterial load assessments are however constrained, providing insights on Map infection at a single time point. This, coupled with the substantial variability in Map distribution across the intestinal tract, and the intricate dynamics of PTB pathogenesis, presents a challenge to understanding the ongoing processes in the infected individuals. In addition, the differences in qPCR, IHC, ZN, and culture results emphasize the complexity of assessing Map presence and distribution, and the need for multiple diagnostic approaches, being the qPCR the most sensitive technique for direct Map detection. However, it should be noted that this method is not able to assess the viability of Map in feces, which would indicate whether the vaccine is reducing the risk of transmission by reducing viable Map. In addition, in the light of the results, to better analyze the effect of the alternative strategies tested in this study, a higher sample size would have been needed due to the high inter- and intraindividual variability observed, not only in lesion severity but also in the fecal shedding patterns and tissue bacterial load. This finding was expected, the variability observed in lesion severity and tissue bacterial load is characteristic of Map infection, even in non-vaccinated, naturally infected animals (19, 96). Moreover, differences between low and high shedders are often several orders of magnitude apart (68), because of this, the results of all the individual animals have been presented, instead of plotting the means. In this sense, for example, animal 23 (OV-INF) exhibited exceptionally high levels of Map-DNA in the studied tissues, surpassing all other animals. In this line, it tested positive for Map culture in all tissues, demonstrated the highest levels of Map-DNA fecal shedding, and showed elevated granuloma counts in all tissues. Though this could be an incidental result, a recent study in European badgers (Meles meles), found that two out of eight of the animals, vaccinated with an inactivated OV against M. bovis, responded in a clearly divergent manner to this vaccination strategy, showing increased lesion severity and bacterial load. The authors proposed several hypotheses like tolerization induced by the oral vaccine or T-cell exhaustion (97).
In this study, to compare the efficacy of each immunization route, the same amount of antigen was administered in the three tested vaccines. However, it needs to be pointed out that a well-designed intradermal vaccine should be effective using lower amounts of antigen than a subcutaneously administered one (47), whereas oral vaccines require higher antigen doses or multiple administrations, as used for human oral vaccines, to achieve the same potency as injected vaccines, due to degradation in the digestive system (79). Thus, the results here presented should be different by adjusting the amount of antigen according to the route of immunization. In this sense, as mentioned before, inactivated OV vaccines have been tested with some success in a rabbit model (33, 35), leaving aside the differences between rabbits and ruminant’s immune systems, rabbits are monogastric and show considerably shorter retention times of ingesta and cecotrophy (98, 99). In contrast, the extended retention times in the upper digestive tract in ruminants mean that a significantly greater quantity of antigen can be potentially degraded in the forestomachs before reaching the intestinal gut-associated lymphoid tissue (GALT), the primary site for antigen uptake, processing, and initiation of immune responses in the intestine (79). To increase this OV effectiveness, an adequate delivery system, targeted for release in the small intestine, could be used to protect both the antigen and the adjuvant from degradation in the gastrointestinal tract (100). Additionally, a higher dose should compensate for the possible degradation and the differences in live weight between rabbits and ruminants. Regarding the OV vaccine, fecal shedding of Map-DNA, by most of the orally vaccinated animals, was detected throughout the first 2 weeks after vaccine administration. Moreover, even though the shed quantity was low, it would be of concern in a live-attenuated vaccine and highlights one of the main potential limitations of the oral administration of this type of vaccine. Regarding IDV, multiple doses could be administered to improve its immunogenicity, given that the amount of antigen inoculated at each injection site cannot be increased. However, it would also incur increased costs. Alternatively, the addition of an adjuvant could increase the IDV potency (101–103). Up to the present, most currently approved adjuvants have been considered as not suitable for intradermal delivery due to the high risk of local reactions (101). However, recently, several adjuvants, some of them previously approved for subcutaneous or intramuscular delivery, have been proven to be safe for use in intradermal vaccines (102, 103), including Quil A®, and its purified fraction QS-21 (104). Quil A® has also been tested in a live-attenuated subcutaneous vaccine against PTB, and it induced a strong IFN-γ response in the goat model without antibody production, and showed significant protective efficacy, reducing histopathological lesions, challenging strain tissue colonization and eliminating fecal shedding (105). The strong IFN-γ response induction, along with the reduction in fecal shedding, was confirmed in a bovine model. However, in this model, the challenge strain employed did not establish itself in the host tissues in any of the study groups (106). Further studies, testing an inactivated subcutaneous Quil A®-adjuvanted vaccine could provide valuable information.
To conclude, new immunization strategies against PTB have been tested in a caprine model confirming that the oral and intradermal immunization routes barely influence the Map-specific peripheral immune responses and therefore induce a low and predictable interference with the bTB IGRA testing. Even though the OV slightly reduced lesion severity, the IDV did not show any evidence of protective efficacy and both were outperformed by the SCV; these results and those of previous studies suggest that achieving vaccine protection against PTB in ruminants without inducing peripheral responses is likely complicated. Alternative PPDs, molecularly defined antigens, as well as DIVA vaccines and diagnostic techniques, may provide a solution to reduce diagnostic interferences (5, 107); however, implementing these options would require changes in the current legislation. Nevertheless, the peripheral immune response parameters induced by vaccination and challenge, in the present study, were not predictive of the infection outcome or lesion severity in the challenged animals. Thus, further investigation into the significance of the peripheral responses induced by these alternative vaccines in protection against PTB, and its effect in the local immunity, would be of interest, especially given the mentioned lack of correlation between the peripheral and mucosal responses induced by parenteral vaccines (90, 91). Our general knowledge on the local immunity elicited by PTB vaccination, particularly through the oral route, is still very limited. It would be of interest to analyze both the local cellular (e.g., cytokine expression and immune cell subpopulations) and humoral responses (e.g., IgA production and B-cell subpopulations), established after vaccination and infection, using larger sample sizes and longer terms, in further studies. This could aid in the development of alternative vaccination strategies that offer protection without the compromises of currently available vaccines.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
Ethics statement
The animal study was approved by Subcomité de Experimentación y Bienestar Animal (OEBA) del Comité de Ética de la Universidad de León, Campus de Vegazana, Calle Profesor s/n 24071, León, Spain. The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
MC: Data curation, Formal analysis, Investigation, Methodology, Visualization, Writing – original draft. MS: Investigation, Methodology, Writing – review & editing. NA-V: Investigation, Methodology, Writing – review & editing. DZ: Investigation, Methodology, Writing – review & editing. NE: Investigation, Supervision, Writing – review & editing, Methodology. EM: Investigation, Methodology, Writing – review & editing. JE: Investigation, Methodology, Writing – review & editing. MF: Investigation, Methodology, Writing – review & editing, Visualization. JB: Conceptualization, Investigation, Methodology, Supervision, Visualization, Writing – review & editing. VP: Conceptualization, Funding acquisition, Investigation, Methodology, Project administration, Supervision, Writing – review & editing. DG-E: Conceptualization, Investigation, Methodology, Supervision, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This study was funded by the projects RTI2018-099496-B-I00 and PID2021-125807OB-C21, from the “Agencia Estatal de Investigación,” of the Spanish Ministry of Science and Innovation. MC acknowledges the receipt of a pre-doctoral grant (PRE2019-087309) from MCIN/AEI/10.13039/501100011033 and FSE “El FSE invierte en tu futuro.”
Acknowledgments
We would like to thank the staff of the Instituto de Ganadería de Montaña (CSIC-ULE) for the handing of the experimental animals and the technical help from María Murillas, Lucía Sanz, Maria Teresa Carro, Maria Jesús Gonzalez, Carmen Espiniella, and Ainara Badiola.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fvets.2024.1457849/full#supplementary-material
References
1. WOAH . Chapter 3.1.16. Paratuberculosis (Johne’s disease) In: Manual of diagnostic tests and vaccines for terrestrial animals. Paris, France: World Organisation for Animal Health (WOAH). (2024).
2. More, S, Bøtner, A, Butterworth, A, Calistri, P, Depner, K, Edwards, S, et al. Assessment of listing and categorisation of animal diseases within the framework of the animal health law (regulation (EU) no 2016/429): paratuberculosis. EFSA J. (2017) 15:e04960. doi: 10.2903/j.efsa.2017.4960
3. Fecteau, M-E. Paratuberculosis in cattle. Vet Clin North Am Food Anim Pract. (2018) 34:209–22. doi: 10.1016/j.cvfa.2017.10.011
4. Diéguez, FJ, Arnaiz, I, Sanjuán, ML, Vilar, MJ, López, M, and Yus, E. Prevalence of serum antibodies to Mycobacterium avium subsp. paratuberculosis in cattle in Galicia (Northwest Spain). Prev Vet Med. (2007) 82:321–6. doi: 10.1016/j.prevetmed.2007.08.006
5. Bastida, F, and Juste, RA. Paratuberculosis control: a review with a focus on vaccination. J Immune Based Ther Vaccines. (2011) 9:8. doi: 10.1186/1476-8518-9-8
6. Groenendaal, H, Zagmutt, FJ, Patton, EA, and Wells, SJ. Cost-benefit analysis of vaccination against Mycobacterium avium ssp. paratuberculosis in dairy cattle, given its cross-reactivity with tuberculosis tests. J Dairy Sci. (2015) 98:6070–84. doi: 10.3168/jds.2014-8914
7. Garcia, AB, and Shalloo, L. Invited review: the economic impact and control of paratuberculosis in cattle. J Dairy Sci. (2015) 98:5019–39. doi: 10.3168/jds.2014-9241
8. Juste, RA, and Perez, V. Control of paratuberculosis in sheep and goats. Vet Clin N Am Food Anim Pract. (2011) 27:127–38. doi: 10.1016/j.cvfa.2010.10.020
9. Rasmussen, P, Barkema, HW, and Hall, DC. Effectiveness and economic viability of Johne’s disease (paratuberculosis) control practices in dairy herds. Front Vet Sci. (2021) 7:614727. doi: 10.3389/fvets.2020.614727
10. Reddacliff, L, Eppleston, J, Windsor, P, Whittington, R, and Jones, S. Efficacy of a killed vaccine for the control of paratuberculosis in Australian sheep flocks. Vet Microbiol. (2006) 115:77–90. doi: 10.1016/j.vetmic.2005.12.021
11. Corbiere, F, Guellouz, D, Tasca, C, Foures, L, Dubaux, E, and Foucras, G. Effects of Silirum®-based vaccination programs on map fecal shedding and serological response in seven French dairy herds. Animals. (2023) 13:1569. doi: 10.3390/ani13091569
12. Dhand, NK, Eppleston, J, Whittington, RJ, and Windsor, PA. Changes in prevalence of ovine paratuberculosis following vaccination with Gudair®: results of a longitudinal study conducted over a decade. Vaccine. (2016) 34:5107–13. doi: 10.1016/j.vaccine.2016.08.064
13. Criado, M, Reyes, LE, Marín, JFG, Gutiérrez-Expósito, D, Zapico, D, Espinosa, J, et al. Adjuvants influence the immune cell populations present at the injection site granuloma induced by whole-cell inactivated paratuberculosis vaccines in sheep. Front Vet Sci. (2024) 11:1284902. doi: 10.3389/fvets.2024.1284902
14. Jayaraman, S, Jain, M, Dhama, K, Singh, SV, Datta, M, Jain, N, et al. DIVA technology: indispensable tool for the control of Johne’s disease. J Exp Biol Agricult Sci. (2016) 4:16–25. doi: 10.18006/2015.4(1).16.25
15. Luo, L, and De Buck, J. Inducing cellular immune responses with a marked Mycobacterium avium subsp. paratuberculosis strain in dairy calves. Vet Microbiol. (2020) 244:108665. doi: 10.1016/j.vetmic.2020.108665
16. Stabel, JR, Waters, WR, Bannantine, JP, and Lyashchenko, K. Mediation of host immune responses after immunization of neonatal calves with a heat-killed Mycobacterium avium subsp. paratuberculosis vaccine. Clin Vaccine Immunol. (2011) 18:2079–89. doi: 10.1128/CVI.05421-11
17. Pérez de Val, B, Nofrarías, M, López-Soria, S, Garrido, JM, Vordermeier, H, Villarreal-Ramos, B, et al. Effects of vaccination against paratuberculosis on tuberculosis in goats: diagnostic interferences and cross-protection. BMC Vet Res. (2012) 8:191. doi: 10.1186/1746-6148-8-191
18. Gupta, S, Singh, SV, Singh, M, Chaubey, KK, Karthik, K, Bhatia, AK, et al. Vaccine approaches for the “therapeutic management” of Mycobacterium avium subspecies paratuberculosis infection in domestic livestock. Vet Q. (2019) 39:143–52. doi: 10.1080/01652176.2019.1667042
19. Corpa, J, Garrido, J, Garcia Marin, J, and Pérez, V. Classification of lesions observed in natural cases of paratuberculosis in goats. J Comp Pathol. (2000) 122:255–65. doi: 10.1053/jcpa.1999.0368
20. Oliveira, R d C, de Oliveira Júnior, CA, Alves, GG, Assis, RA, Silva, ROS, de Sousa Xavier, MA, et al. Cattle and goats’ humoral response to vaccination with Clostridium perfringens type D purified epsilon toxoids. Anaerobe. (2021) 72:102465. doi: 10.1016/j.anaerobe.2021.102465
21. Bannantine, JP, Hines, ME, Bermudez, LE, Talaat, AM, Sreevatsan, S, Stabel, JR, et al. A rational framework for evaluating the next generation of vaccines against Mycobacterium avium subspecies paratuberculosis. Front Cell Infect Microbiol. (2014) 4:126. doi: 10.3389/fcimb.2014.00126
22. Park, KT, Allen, AJ, Bannantine, JP, Seo, KS, Hamilton, MJ, Abdellrazeq, GS, et al. Evaluation of two mutants of Mycobacterium avium subsp. paratuberculosis as candidates for a live attenuated vaccine for Johne’s disease. Vaccine. (2011) 29:4709–19. doi: 10.1016/j.vaccine.2011.04.090
23. Park, KT, Allen, AJ, Barrington, GM, and Davis, WC. Deletion of relA abrogates the capacity of Mycobacterium avium paratuberculosis to establish an infection in calves. Front Cell Infect Microbiol. (2014) 4:89674. doi: 10.3389/FCIMB.2014.00064/BIBTEX
24. Abdellrazeq, GS, Elnaggar, MM, Bannantine, JP, Park, KT, Souza, CD, Backer, B, et al. A Mycobacterium avium subsp. paratuberculosis relA deletion mutant and a 35 kDa major membrane protein elicit development of cytotoxic T lymphocytes with ability to kill intracellular bacteria. Vet Res. (2018) 49:53. doi: 10.1186/s13567-018-0549-3
25. Eshraghisamani, R, Facciuolo, A, and De Buck, J. Oral paratuberculosis vaccine efficacy and mucosal immunity in cattle. Vaccine. (2024) 42:126447. doi: 10.1016/j.vaccine.2024.126447
26. Koets, A, Hoek, A, Langelaar, M, Overdijk, M, Santema, W, Franken, P, et al. Mycobacterial 70 kD heat-shock protein is an effective subunit vaccine against bovine paratuberculosis. Vaccine. (2006) 24:2550–9. doi: 10.1016/J.VACCINE.2005.12.019
27. Kathaperumal, K, Kumanan, V, McDonough, S, Chen, LH, Park, SU, Moreira, MAS, et al. Evaluation of immune responses and protective efficacy in a goat model following immunization with a coctail of recombinant antigens and a polyprotein of Mycobacterium avium subsp. paratuberculosis. Vaccine. (2009) 27:123–35. doi: 10.1016/J.VACCINE.2008.10.019
28. Hobernik, D, and Bros, M. DNA vaccines—how far from clinical use? Int J Mol Sci. (2018) 19:3605. doi: 10.3390/IJMS19113605
29. Roupie, V, Viart, S, Leroy, B, Romano, M, Trinchero, N, Govaerts, M, et al. Immunogenicity of eight Mycobacterium avium subsp. paratuberculosis specific antigens in DNA vaccinated and map infected mice. Vet Immunol Immunopathol. (2012) 145:74–85. doi: 10.1016/J.VETIMM.2011.10.012
30. Sechi, LA, Mara, L, Cappai, P, Frothingam, R, Ortu, S, Leoni, A, et al. Immunization with DNA vaccines encoding different mycobacterial antigens elicits a Th1 type immune response in lambs and protects against Mycobacterium avium subspecies paratuberculosis infection. Vaccine. (2006) 24:229–35. doi: 10.1016/J.VACCINE.2005.08.086
31. Thakur, A, Andrea, A, Mikkelsen, H, Woodworth, JS, Andersen, P, Jungersen, G, et al. Targeting the Mincle and TLR3 receptor using the dual agonist cationic adjuvant formulation 9 (CAF09) induces humoral and polyfunctional memory T cell responses in calves. PLoS One. (2018) 13:e0201253. doi: 10.1371/journal.pone.0201253
32. Begg, DJ, Dhungyel, O, Naddi, A, Dhand, NK, Plain, KM, de Silva, K, et al. The immunogenicity and tissue reactivity of Mycobacterium avium subsp paratuberculosis inactivated whole cell vaccine is dependent on the adjuvant used. Heliyon. (2019) 5:e01911. doi: 10.1016/j.heliyon.2019.e01911
33. Ladero-Auñon, I, Molina, E, Oyanguren, M, Barriales, D, Fuertes, M, Sevilla, IA, et al. Oral vaccination stimulates neutrophil functionality and exerts protection in a Mycobacterium avium subsp. paratuberculosis infection model. NPJ Vaccines. (2021) 6:102–15. doi: 10.1038/S41541-021-00367-8
34. Hines, ME, Turnquist, SE, Ilha, MRS, Rajeev, S, Jones, AL, Whittington, L, et al. Evaluation of novel oral vaccine candidates and validation of a caprine model of Johne’s disease. Front Cell Infect Microbiol. (2014) 5:26. doi: 10.3389/FCIMB.2014.00026/BIBTEX
35. Arrazuria, R, Ladero, I, Molina, E, Fuertes, M, Juste, R, Fernández, M, et al. Alternative vaccination routes against paratuberculosis modulate local immune response and interference with tuberculosis diagnosis in laboratory animal models. Vet Sci. (2020) 7:7. doi: 10.3390/vetsci7010007
36. Cross, ML, and Aldwell, F. Oral vaccination against bovine tuberculosis with Mycobacterium bovis BCG. Expert Rev Vaccines. (2007) 6:323–31. doi: 10.1586/14760584.6.3.323
37. Doherty, TM, Olsen, AW, Van Pinxteren, L, and Andersen, P. Oral vaccination with subunit vaccines protects animals against aerosol infection with Mycobacterium tuberculosis. Infect Immun. (2002) 70:3111–21. doi: 10.1128/IAI.70.6.3111-3121.2002
38. Buddle, BM, Aldwell, FE, Skinner, MA, De Lisle, GW, Denis, M, Vordermeier, HM, et al. Effect of oral vaccination of cattle with lipid-formulated BCG on immune responses and protection against bovine tuberculosis. Vaccine. (2005) 23:3581–9. doi: 10.1016/J.VACCINE.2005.01.150
39. Roy, A, Risalde, MA, Casal, C, Romero, B, de Juan, L, Menshawy, AM, et al. Oral vaccination with heat-inactivated Mycobacterium bovis does not interfere with the antemortem diagnostic techniques for tuberculosis in goats. Front Vet Sci. (2017) 4:124. doi: 10.3389/fvets.2017.00124
40. Gormley, E, Bhuachalla, DN, O’Keeffe, J, Murphy, D, Aldwell, FE, Fitzsimons, T, et al. Oral vaccination of free-living badgers (Meles meles) with Bacille Calmette Guérin (BCG) vaccine confers protection against tuberculosis. PLoS One. (2017) 12:e0168851. doi: 10.1371/JOURNAL.PONE.0168851
41. Palmer, MV, Thacker, TC, Waters, WR, and Robbe-Austerman, S. Oral vaccination of white-tailed deer (Odocoileus virginianus) with Mycobacterium bovis Bacillus Calmette-Guerin (BCG). PLoS One. (2014) 9:e97031. doi: 10.1371/JOURNAL.PONE.0097031
42. Aldwell, FE, Tucker, IG, de Lisle, GW, and Buddle, BM. Oral delivery of Mycobacterium bovis BCG in a lipid formulation induces resistance to pulmonary tuberculosis in mice. Infect Immun. (2003) 71:101–8. doi: 10.1128/IAI.71.1.101-108.2003
43. Vierboom, MPM, Dijkman, K, Sombroek, CC, Hofman, SO, Boot, C, Vervenne, RAW, et al. Stronger induction of trained immunity by mucosal BCG or MTBVAC vaccination compared to standard intradermal vaccination. Cell Rep Med. (2021) 2:100185. doi: 10.1016/J.XCRM.2020.100185
44. Lee, H, Cho, SN, Kim, HJ, Anh, YM, Choi, JE, Kim, CH, et al. Evaluation of cell-mediated immune responses to two BCG vaccination regimes in young children in South Korea. Vaccine. (2011) 29:6564–71. doi: 10.1016/J.VACCINE.2011.07.003
45. Meyer, J, Harris, SA, Satti, I, Poulton, ID, Poyntz, HC, Tanner, R, et al. Comparing the safety and immunogenicity of a candidate TB vaccine MVA85A administered by intramuscular and intradermal delivery. Vaccine. (2013) 31:1026–33. doi: 10.1016/J.VACCINE.2012.12.042
46. Jones, GJ, Steinbach, S, Sevilla, IA, Garrido, JM, Juste, R, and Vordermeier, HM. Oral vaccination of cattle with heat inactivated Mycobacterium bovis does not compromise bovine TB diagnostic tests. Vet Immunol Immunopathol. (2016) 182:85–8. doi: 10.1016/J.VETIMM.2016.10.010
47. Hickling, JK, Jones, KR, Friede, M, Zehrung, D, Chen, D, and Kristensenc, D. Intradermal delivery of vaccines: potential benefits and current challenges. Bull World Health Organ. (2011) 89:221–6. doi: 10.2471/BLT.10.079426
48. Holland, D, Booy, R, De Looze, F, Eizenberg, P, McDonald, J, Karrasch, J, et al. Intradermal influenza vaccine administered using a new microinjection system produces superior immunogenicity in elderly adults: a randomized controlled trial. J Infect Dis. (2008) 198:650–8. doi: 10.1086/590434
49. Propst, T, Propst, A, Lhotta, K, Vogel, W, and Konig, P. Reinforced intradermal hepatitis B vaccination in hemodialysis patients is superior in antibody response to intramuscular or subcutaneous vaccination. Am J Kidney Dis. (1998) 32:1041–5. doi: 10.1016/S0272-6386(98)70081-2
50. Temple, D, Jiménez, M, Escribano, D, Martín-Valls, G, Díaz, I, and Manteca, X. Welfare benefits of intradermal vaccination of piglets. Animals (Basel). (2020) 10:1898. doi: 10.3390/ANI10101898
51. Gilmour, NJL, and Angus, KW. Absence of immunogenicity of an Oral vaccine against Mycobacterium johnei in sheep. Res Vet Sci. (1974) 16:269–70. doi: 10.1016/S0034-5288(18)33764-0
52. Hines, ME, Stabel, JR, Sweeney, RW, Griffin, F, Talaat, AM, Bakker, D, et al. Experimental challenge models for Johne’s disease: a review and proposed international guidelines. Vet Microbiol. (2007) 122:197–222. doi: 10.1016/j.vetmic.2007.03.009
53. Fernández, M, Benavides, J, Sevilla, IA, Fuertes, M, Castaño, P, Delgado, L, et al. Experimental infection of lambs with C and S-type strains of Mycobacterium avium subspecies paratuberculosis: immunological and pathological findings. Vet Res. (2014) 45:1–15. doi: 10.1186/1297-9716-45-5/FIGURES/7
54. Begg, DJ, and Whittington, RJ. Experimental animal infection models for Johne’s disease, an infectious enteropathy caused by Mycobacterium avium subsp. paratuberculosis. Vet J. (2008) 176:129–45. doi: 10.1016/j.tvjl.2007.02.022
55. Idris, SM, Eltom, KH, Okuni, JB, Ojok, L, Elmagzoub, WA, El, WAA, et al. Paratuberculosis: the hidden killer of small ruminants. Animals (Basel). (2022) 12:12. doi: 10.3390/ANI12010012
56. Windsor, PA, and Whittington, RJ. Evidence for age susceptibility of cattle to Johne’s disease. Vet J. (2010) 184:37–44. doi: 10.1016/j.tvjl.2009.01.007
57. Pérez, V, Tellechea, J, Corpa, JM, Gutiérrez, M, and Marín, JFG. Relation between pathologic findings and cellular immune responses in sheep with naturally acquired paratuberculosis. Am J Vet Res. (1999) 60:123–7. doi: 10.2460/ajvr.1999.60.01.123
58. Sevilla, I, Li, L, Amonsin, A, Garrido, JM, Geijo, MV, Kapur, V, et al. Comparative analysis of Mycobacterium avium subsp. paratuberculosis isolates from cattle, sheep and goats by short sequence repeat and pulsed-field gel electrophoresis typing. BMC Microbiol. (2008) 8:1–8. doi: 10.1186/1471-2180-8-204
59. Oyanguren, M, Molina, E, Mugica, M, Ladero-Auñon, I, Fuertes, M, Fernández, M, et al. Probiotic bacteria can modulate immune responses to paratuberculosis vaccination. Front Cell Infect Microbiol. (2024) 14:1394070. doi: 10.3389/FCIMB.2024.1394070
60. Delgado, L, Juste, RA, Muñoz, M, Morales, S, Benavides, J, Ferreras, MC, et al. Differences in the peripheral immune response between lambs and adult ewes experimentally infected with Mycobacterium avium subspecies paratuberculosis. Vet Immunol Immunopathol. (2012) 145:23–31. doi: 10.1016/j.vetimm.2011.10.005
61. Pérez, V, Marín, JFG, and Badiola, JJ. Description and classification of different types of lesion associated with natural paratuberculosis infection in sheep. J Comp Pathol. (1996) 114:107–22. doi: 10.1016/S0021-9975(96)80001-6
62. Kawaji, S, Taylor, DL, Mori, Y, and Whittington, RJ. Detection of Mycobacterium avium subsp. paratuberculosis in ovine faeces by direct quantitative PCR has similar or greater sensitivity compared to radiometric culture. Vet Microbiol. (2007) 125:36–48. doi: 10.1016/j.vetmic.2007.05.002
63. Arteche-Villasol, N, Gutiérrez-Expósito, D, Vallejo, R, Espinosa, J, Elguezabal, N, Ladero-Auñon, I, et al. Early response of monocyte-derived macrophages from vaccinated and non-vaccinated goats against in vitro infection with Mycobacterium avium subsp. paratuberculosis. Vet Res. (2021) 52:69–12. doi: 10.1186/s13567-021-00940-y
64. Delgado, L, Marín, JFG, Muñoz, M, Benavides, J, Juste, RA, García-Pariente, C, et al. Pathological findings in young and adult sheep following experimental infection with 2 different doses of Mycobacterium avium subspecies paratuberculosis. Vet Pathol. (2013) 50:857–66. doi: 10.1177/0300985813476066
65. Sevilla, IA, Garrido, JM, Molina, E, Geijo, MV, Elguezabal, N, Vázquez, P, et al. Development and evaluation of a novel multicopy-element-targeting triplex PCR for detection of Mycobacterium avium subsp. paratuberculosis in feces. Appl Environ Microbiol. (2014) 80:3757–68. doi: 10.1128/AEM.01026-14
66. R Core Team . R: A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing. (2022).
67. de Juan, L, Álvarez, J, Aranaz, A, Rodríguez, A, Romero, B, Bezos, J, et al. Molecular epidemiology of types I/III strains of Mycobacterium avium subspecies paratuberculosis isolated from goats and cattle. Vet Microbiol. (2006) 115:102–10. doi: 10.1016/J.VETMIC.2006.01.008
68. Stewart, DJ, Vaughan, JA, Stiles, PL, Noske, PJ, Tizard, MLVV, Prowse, SJ, et al. A long-term study in merino sheep experimentally infected with Mycobacterium avium subsp. paratuberculosis: clinical disease, faecal culture and immunological studies. Vet Microbiol. (2004) 104:165–78. doi: 10.1016/j.vetmic.2004.09.007
69. O’Brien, A, Whelan, C, Clarke, JB, Hayton, A, Watt, NJ, and Harkiss, GD. Serological analysis of tuberculosis in goats by use of the enferplex caprine TB multiplex test. Clin Vaccine Immunol. (2017) 24:518–34. doi: 10.1128/CVI.00518-16
70. Gelalcha, BD, Zewude, A, and Ameni, G. Tuberculosis caused by Mycobacterium bovis in a sheep flock Colocated with a tuberculous dairy cattle herd in Central Ethiopia. J Vet Med. (2019) 2019:1–6. doi: 10.1155/2019/8315137
71. Fernández, M, Delgado, L, Sevilla, IA, Fuertes, M, Castaño, P, Royo, M, et al. Virulence attenuation of a Mycobacterium avium subspecies paratuberculosis S-type strain prepared from intestinal mucosa after bacterial culture. Evaluation in an experimental ovine model. Res Vet Sci. (2015) 99:180–7. doi: 10.1016/J.RVSC.2015.02.001
72. Watkins, C, Schock, A, May, L, Denham, S, Sales, J, Welch, L, et al. Assessing virulence of vaccine strains of Mycobacterium avium subspecies paratuberculosis in a calf model. Vet Microbiol. (2010) 146:63–9. doi: 10.1016/J.VETMIC.2010.04.017
73. Arteche-Villasol, N, Gutiérrez-Expósito, D, Criado, M, Benavides, J, and Pérez, V. Assessment of paratuberculosis vaccination effect on in vitro formation of neutrophil extracellular traps in a sheep model. Vaccine. (2022) 10:1403. doi: 10.3390/VACCINES10091403
74. Begg, DJ, Plain, KM, de Silva, K, Gurung, R, Gunn, A, Purdie, AC, et al. Immunopathological changes and apparent recovery from infection revealed in cattle in an experimental model of Johne’s disease using a lyophilised culture of Mycobacterium avium subspecies paratuberculosis. Vet Microbiol. (2018) 219:53–62. doi: 10.1016/j.vetmic.2018.03.029
75. Navarro-Gonzalez, N, Fourichon, C, Blanquefort, P, Delafosse, A, Joly, A, Ngwa-Mbot, D, et al. Longitudinal study of Mycobacterium avium ssp. paratuberculosis fecal shedding patterns and concurrent serological patterns in naturally infected dairy cattle. J Dairy Sci. (2019) 102:9117–37. doi: 10.3168/jds.2018-15897
76. Juste, RAA, García Marín, JFF, Peris, B, Sáez de Ocáriz, C, and Badiola, JJJ. Experimental infection of vaccinated and non-vaccinated lambs with Mycobacterium paratuberculosis. J Comp Pathol. (1994) 110:185–94. doi: 10.1016/S0021-9975(08)80189-2
77. Coussens, PM. Mycobacterium paratuberculosis and the bovine immune system. Anim Health Res Rev. (2001) 2:141–62. doi: 10.1079/AHRR200134
78. Pooley, HB, Begg, DJ, Plain, KM, Whittington, RJ, Purdie, AC, and De Silva, K. The humoral immune response is essential for successful vaccine protection against paratuberculosis in sheep. BMC Vet Res. (2019) 15:223. doi: 10.1186/s12917-019-1972-z
79. Kwong, KW-YY, Xin, Y, Lai, NC-YY, Sung, JC-CC, Wu, K-CC, Hamied, YK, et al. Oral vaccines: a better future of immunization. Vaccines (Basel). (2023) 11:1232. doi: 10.3390/vaccines11071232
80. Koets, AP, Eda, S, and Sreevatsan, S. The within host dynamics of Mycobacterium avium ssp. paratuberculosis infection in cattle: where time and place matter. Vet Res. (2015) 46:61. doi: 10.1186/s13567-015-0185-0
81. Molina, E, Geijo, M, Múgica, M, Oyanguren, M, Garrido, J, and Elguezabal, N. Un prototipo de vacuna oral frente a la paratuberculosis no interfiere con el diagnóstico de la tuberculosis bovina y aumenta la actividad de los fagocitos en un modelo caprino. XXVI AVEDILA. (2023):84.
82. López, V, González-Barrio, D, Lima-Barbero, JF, Ortiz, JA, Domínguez, L, Juste, R, et al. Oral administration of heat-inactivated Mycobacterium bovis reduces the response of farmed red deer to avian and bovine tuberculin. Vet Immunol Immunopathol. (2016) 172:21–5. doi: 10.1016/j.vetimm.2016.03.003
83. Garrido, JM, Sevilla, IA, Beltrán-Beck, B, Minguijón, E, Ballesteros, C, Galindo, RC, et al. Protection against tuberculosis in Eurasian wild boar vaccinated with heat-inactivated Mycobacterium bovis. PLoS One. (2011) 6:e24905. doi: 10.1371/journal.pone.0024905
84. Lahey, T, Arbeit, RD, Bakari, M, Horsburgh, CR, Matee, M, Waddell, R, et al. Immunogenicity of a protective whole cell mycobacterial vaccine in HIV-infected adults: a phase III study in Tanzania. Vaccine. (2010) 28:7652–8. doi: 10.1016/j.vaccine.2010.09.041
85. Fernández, M, Royo, M, Fuertes, M, Arteche-Villasol, N, Ferreras, MC, Benavides, J, et al. Effects of Paratuberculosis vaccination at different ages in a dairy goat herd: a 2-year follow-up. Animals. (2022) 12:3135. doi: 10.3390/ANI12223135
86. Arteche-Villasol, N, Gutiérrez-Expósito, D, Elguezabal, N, Sevilla, IA, Vallejo, R, Espinosa, J, et al. Influence of heterologous and homologous vaccines, and their components, on the host immune response and protection against experimental caprine Paratuberculosis. Front Vet Sci. (2022) 8:1568. doi: 10.3389/FVETS.2021.744568/BIBTEX
87. Faisal, SM, Chen, JW, Yan, F, Chen, TT, Useh, NM, Yan, W, et al. Evaluation of a mycobacterium avium subsp. paratuberculosis leuD mutant as a vaccine candidate against challenge in a caprine model. Clin Vaccine Immunol. (2013) 20:572–81. doi: 10.1128/CVI.00653-12
88. Santema, W, Hensen, S, Rutten, V, and Koets, A. Heat shock protein 70 subunit vaccination against bovine paratuberculosis does not interfere with current immunodiagnostic assays for bovine tuberculosis. Vaccine. (2009) 27:2312–9. doi: 10.1016/J.VACCINE.2009.02.032
89. Álvarez, J, de Juan, L, Bezos, J, Romero, B, Sáez, JL, Gordejo, FJR, et al. Interference of paratuberculosis with the diagnosis of tuberculosis in a goat flock with a natural mixed infection. Vet Microbiol. (2008) 128:72–80. doi: 10.1016/j.vetmic.2007.08.034
90. Facciuolo, A, Lee, AH, Trimble, MJ, Rawlyk, N, Townsend, HGG, Bains, M, et al. A bovine enteric Mycobacterium infection model to analyze parenteral vaccine-induced mucosal immunity and accelerate vaccine discovery. Front Immunol. (2020) 11:1. doi: 10.3389/FIMMU.2020.586659/FULL
91. Purdie, AC, Plain, KM, Pooley, H, Begg, DJ, de Silva, K, and Whittington, RJ. Correlates of vaccine protection against Mycobacterium avium sub-species paratuberculosis infection revealed in a transcriptomic study of responses in Gudair® vaccinated sheep. Front Vet Sci. (2022) 9:1004237. doi: 10.3389/fvets.2022.1004237
92. Stamp, JT, and Watt, JA. Johne’s disease in sheep. J Comp Pathol. (1954) 64:26–40. doi: 10.1016/S0368-1742(54)80005-1
93. Derakhshandeh, A, Namazi, F, Khatamsaz, E, Eraghi, V, and Hemati, Z. Goat paratuberculosis in shiraz: histopathological and molecular approaches. Vet Res Forum. (2018) 9:253–7. doi: 10.30466/VRF.2018.32080
94. Blanco Vázquez, C, Alonso-Hearn, M, Juste, RA, Canive, M, Iglesias, T, Iglesias, N, et al. Detection of latent forms of Mycobacterium avium subsp. paratuberculosis infection using host biomarker-based ELISAs greatly improves paratuberculosis diagnostic sensitivity. PLoS One. (2020) 15:e0236336. doi: 10.1371/journal.pone.0236336
95. Thomas, J, Risalde, MÁ, Serrano, M, Sevilla, I, Geijo, M, Ortíz, JA, et al. The response of red deer to oral administration of heat-inactivated Mycobacterium bovis and challenge with a field strain. Vet Microbiol. (2017) 208:195–202. doi: 10.1016/j.vetmic.2017.08.007
96. Englund, S, Bölske, G, Ballagi-Pordány, A, and Johansson, K-E. Detection of Mycobacterium avium subsp. paratuberculosis in tissue samples by single, fluorescent and nested PCR based on the IS900 gene. Vet Microbiol. (2001) 81:257–71. doi: 10.1016/S0378-1135(01)00348-0
97. Juste, RA, Blanco-Vázquez, C, Barral, M, Prieto, JM, Varela-Castro, L, Lesellier, S, et al. Efficacy of heat-inactivated Mycobacterium bovis vaccine delivered to European badgers (Meles meles) through edible bait. Heliyon. (2023) 9:e19349. doi: 10.1016/J.HELIYON.2023.E19349
98. Gidenne, T, Perez, J-M, Gidenne, T, and Perez, J-M. Replacement of digestible fibre by starch in the diet of the growing rabbit. I. Effects on digestion, rate of passage and retention of nutrients. Ann Zootech. (2000) 49:357–68. doi: 10.1051/ANIMRES:2000127
99. Mambrini, M, and Peyraud, J. Retention time of feed particles and liquids in the stomachs and intestines of dairy cows. Direct measurement and calculations based on faecal collection. Reprod Nutr Dev. (1997) 37:427–42. doi: 10.1051/rnd:19970404
100. Kang, SH, Hong, SJ, Lee, YK, and Cho, S. Oral vaccine delivery for intestinal immunity—biological basis, barriers, delivery system, and M cell targeting. Polymers. (2018) 10:948. doi: 10.3390/POLYM10090948
101. Chen, X. Emerging adjuvants for intradermal vaccination. Int J Pharm. (2023) 632:122559. doi: 10.1016/j.ijpharm.2022.122559
102. Wang, J, Li, P, and Wu, MX. Natural STING agonist as an “ideal” adjuvant for cutaneous vaccination. J Invest Dermatol. (2016) 136:2183–91. doi: 10.1016/j.jid.2016.05.105
103. Bardel, E, Doucet-Ladeveze, R, Mathieu, C, Harandi, AM, Dubois, B, and Kaiserlian, D. Intradermal immunisation using the TLR3-ligand poly (I:C) as adjuvant induces mucosal antibody responses and protects against genital HSV-2 infection. NPJ Vaccines. (2016) 1:16010. doi: 10.1038/npjvaccines.2016.10
104. Vassilieva, EV, Li, S, Korniychuk, H, Taylor, DM, Wang, S, Prausnitz, MR, et al. cGAMP/Saponin adjuvant combination improves protective response to influenza vaccination by microneedle patch in an aged mouse model. Front Immunol. (2021) 11:583251. doi: 10.3389/fimmu.2020.583251
105. Shippy, DC, Lemke, JJ, Berry, A, Nelson, K, Hines, ME, and Talaat, AM. Superior protection from live- attenuated vaccines directed against Johne’s disease. Clin Vaccine Immunol. (2017) 24:e00478-16. doi: 10.1128/CVI.00478-16
106. Phanse, Y, Wu, C-W, Venturino, AJ, Hansen, C, Nelson, K, Broderick, SR, et al. A protective vaccine against Johne’s disease in cattle. Microorganisms. (2020) 8:1427. doi: 10.3390/microorganisms8091427
Keywords: paratuberculosis, oral vaccine, intradermal vaccine, immunization strategies, goat
Citation: Criado M, Silva M, Arteche-Villasol N, Zapico D, Elguezabal N, Molina E, Espinosa J, Ferreras MC, Benavides J, Pérez V and Gutiérrez-Expósito D (2024) Evaluation of alternative vaccination routes against paratuberculosis in goats. Front. Vet. Sci. 11:1457849. doi: 10.3389/fvets.2024.1457849
Edited by:
Jesús Hernández, National Council of Science and Technology (CONACYT), MexicoReviewed by:
Douwe Bakker, Independent Researcher, Lelystad, NetherlandsMihaela Niculae, University of Agricultural Sciences and Veterinary Medicine of Cluj-Napoca, Romania
Copyright © 2024 Criado, Silva, Arteche-Villasol, Zapico, Elguezabal, Molina, Espinosa, Ferreras, Benavides, Pérez and Gutiérrez-Expósito. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Miguel Criado, bWNyaWJAdW5pbGVvbi5lcw==