 Ondřej Máca
Ondřej Máca Naglis Gudiškis
Naglis Gudiškis Dalius Butkauskas
Dalius Butkauskas David González-Solís
David González-Solís Petras Prakas
Petras Prakas- 1Department of Pathology and Parasitology, State Veterinary Institute Prague, Prague, Czechia
- 2Department of Zoology and Fisheries, Faculty of Agrobiology, Food and Natural Resources, Czech University of Life Sciences Prague, Prague, Czechia
- 3Nature Research Centre, Vilnius, Lithuania
- 4Department of Systematics and Aquatic Ecology, El Colegio de la Frontera Sur, Chetumal, Mexico
Background: Sarcocystis includes a global group of apicomplexan parasites with two-host life cycle frequently circulating in wildlife and domestic hosts, including humans. Two of the most important wild terrestrial carnivores acting as definitive hosts are the red fox and raccoon dog, due to their wide distribution in Europe and usage of wild and farmed animals as prey. This study was conducted to determine the prevalence of Sarcocystis in hunted red foxes and raccoon dogs from nine regions of the Czech Republic and to identify isolated sporocysts by molecular techniques.
Methods: Approximately 5 g of the contents of large intestine from 200 animals (197 red foxes and three raccoon dogs) were examined by flotation centrifugation coprological method. Only samples of 50 red foxes and one raccoon dog positive to Sarcocystis spp. were used for the nested PCR (nPCR) method to amplify a fragment or partial sequence on the cox1 gene. Ten species-specific primer pairs for detection of Sarcocystis spp. using farm animals as intermediate hosts were utilized.
Results: In total, 38.1% of the red foxes and 66.7% of the raccoon dogs were positive to Sarcocystis by light microscopy. The molecular characterization resulted in the identification of five species in the red fox: S. arieticanis, S. capracanis, S. cruzi, S. miescheriana, and S. tenella, while the PCR was negative for the sole raccoon dog. The highest intraspecific variation was found for S. miescheriana, while S. tenella was the most prevalent. Co-infections occurred in the large intestine of the red fox. No zoonotic species were found in our samples.
Conclusion: This is the first study where the potential role of the red fox and raccoon dogs as spreaders of Sarcocystis to farm animals in the Czech Republic is shown. The use of species-specific primers provides a fast and easy method for screening multiple samples for a particular Sarcocystis species.
Introduction
Members of the genus Sarcocystis are apicomplexan parasites of reptiles, birds and mammals including humans (1). They have compulsory two-host prey–predator life cycle. Asexual sarcocysts are found mainly in muscle tissues of intermediate hosts (herbivores, omnivores, and carnivores), while sexual sporocysts develops in the lamina propria of the small intestine of definitive hosts (carnivores, scavengers) (1–3). Definitive hosts get infected through consumption of animal tissues containing mature sarcocyst, while intermediate hosts acquire Sarcocystis infection via food or water contaminated with sporocysts. Some of Sarcocystis spp. (e.g., S. canis, S. calchasi, S. falcatula, S. neurona) are highly pathogenic for domestic and wildlife animals (1, 4–6). Furthermore, the livestock industry suffers losses due to macroscopic sarcocysts, reduced quality of meat due to intensive Sarcocystis infections or due to the rarely encountered clinical symptom induced by acute infections (1, 7, 8).
Morphological distinguishment of Sarcocystis spp. according to the sexual stages of the parasites is virtually not possible in the final hosts (1, 2, 9, 10). Therefore, definitive hosts of Sarcocystis spp. have historically been identified through laboratory transmission experiments (11–14). However, co-infection with several Sarcocystis spp. is very common in wild and domestic ungulates, which complicates the implementation and reliability of transmission experiments (15, 16). Furthermore, the ethical considerations related to the use of wild predatory mammals or birds make it crucial to find other approaches in revealing the life cycles of these parasites. Therefore, DNA analysis methods are now increasingly used to identify Sarcocystis spp. in intestinal or fecal samples of definitive hosts (9, 10, 17–23).
Representatives of the family Canidae (e.g., red fox [Vulpes vulpes], Arctic fox [Vulpes lagopus], coyote [Canis latrans], gray wolf [Canis lupus], raccoon dogs [Nyctereutes procyonoides], jackal [Canis aureus], dingo [Canis lupus dingo] and dog [Canis lupus familiaris]) are involved in the prey–predator life cycle of numerous Sarcocystis spp. Most of these parasite species employ domesticated and wild ungulates as their intermediate hosts (1). The red fox is widely distributed throughout the Northern Hemisphere (24) and suggested as the definitive host of about 20 Sarcocystis spp. forming sarcocysts in muscles of domestic and wild ungulates, small mammals, and birds (1, 19, 25–27). The raccoon dog primarily originated from the Far East, although nowadays it is one of the most prevalent invasive mammal species in Europe (28). It has been shown that raccoon dogs serve as definitive hosts of several Sarcocystis spp. employing roe deer (Capreolus capreolus), reindeer (Rangifer tarandus), pigs and wild boar (Sus scrofa), and ducks (Anas platyrhynchos) as their intermediate hosts (18, 26, 29). However, there is a lack of molecular or epidemiological investigations addressing the role of red foxes and raccoon dogs in the transmission of Sarcocystis spp. to farm animals. Since the red fox and raccoon dog serve as definitive and reservoir hosts for a wide variety of Sarcocystis spp., the main goal of the present study was to determine the Sarcocystis spp. using these canid hosts as definitive hosts and farmed animals as intermediate hosts.
Methods
The whole intestinal tracts of 200 animals (197 red foxes and three raccoon dogs) were obtained during the monitoring on rabies and Echinococcus in 2019 from nine regions of the Czech Republic (Figure 1). These samples were sent to the State Veterinary Institute Prague and approximately 5 g of the contents of large intestine were taken and examined by flotation centrifugation coprological method according to Breza (30) using a Leica DMLB optical microscope with a Leica DFC420 digital camera (Leica Microsystems, Wetzlar, Germany) and concentrated sporocysts were transferred directly from glass slide by pipette to 2 mL Eppendorf safe-lock tubes with InhibitEX buffer. Only samples of 50 red foxes and one raccoon dog positive to Sarcocystis spp. were used for the molecular identification. Total genomic DNA (gDNA) was extracted from purified sporocysts using a QIAamp® Fast DNA Stool Mini Kit (Qiagen, Hilden, Germany) according to the manufacturer’s instructions, except disruption of the sporocysts mixed with InhibitEX buffer with glass beads. The eluted DNA was kept at −20°C until further use.
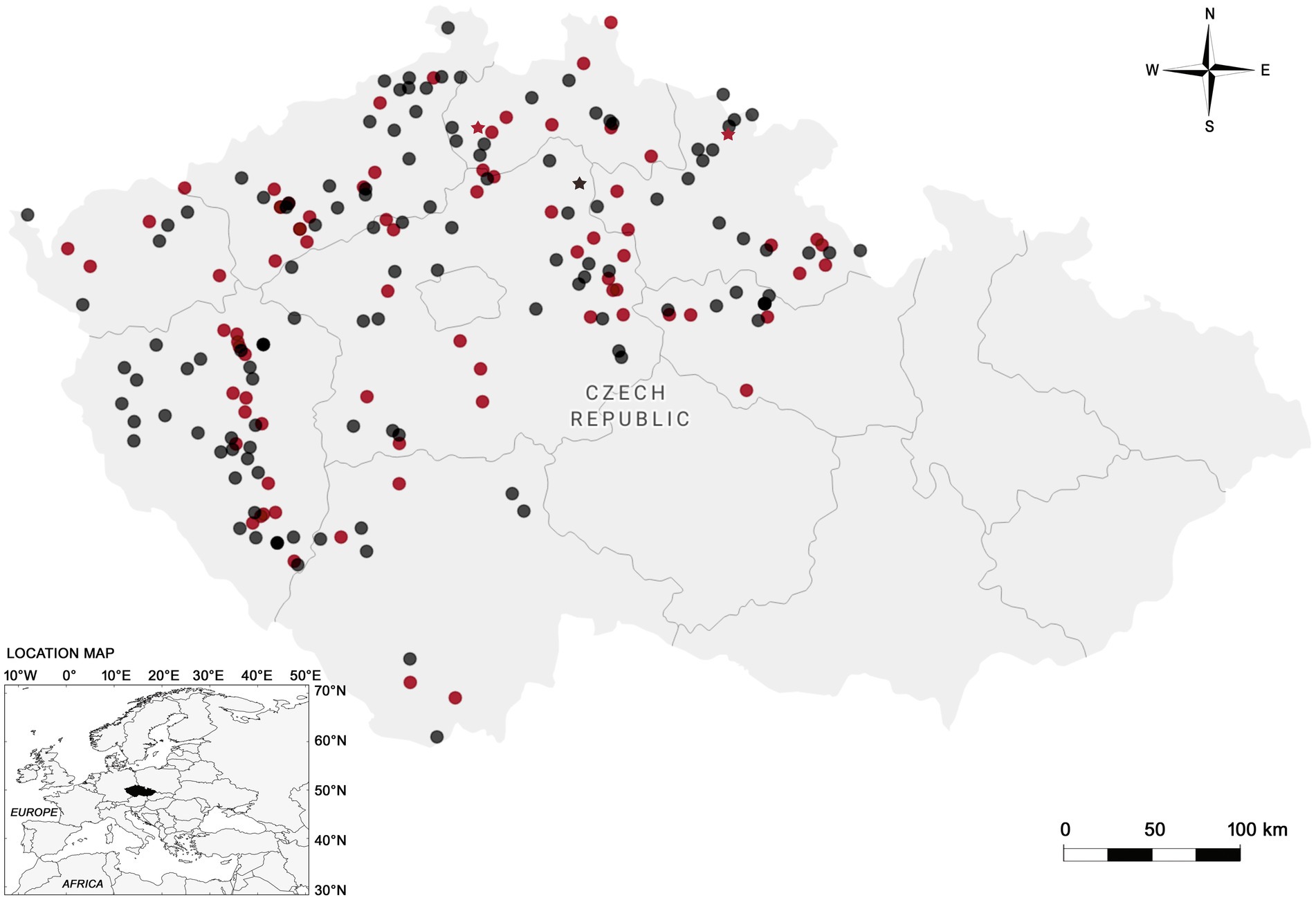
Figure 1. Map of the Czech Republic showing the sampling sites and regions (outlined areas) where red foxes (circles) and raccoon dogs (stars) were collected. Negative samples are represented in black color, while positive in red.
The nested PCR (nPCR) method was used to amplify a fragment or partial sequence on the cox1 gene from the collected DNA samples. Ten species-specific primer pairs for detection of Sarcocystis spp. using cattle (Bos taurus), goat (Capra hircus), horse (Equus caballus), pig, and sheep (Ovis aries) as intermediate hosts were utilized (see Table 1). Two of the tested Sarcocystis spp. (i.e., S. hominis and S. suihominis) employ humans as definitive hosts (3). The first round of amplification was carried out with a reaction mixture of 25 μL comprising 12.5 μL of DreamTaq PCR Master Mix (Thermo Fisher Scientific Baltics, Vilnius, Lithuania), 4 μL of DNA template, 0.5 μM of both forward and reverse primers, and nuclease-free water added up to 25 μL. nPCR was carried out using a Veriti 96-Well Thermal Cycler (Applied Biosystems, Foster City, CA, United States). The thermal cycling conditions began with 5 min at 95°C, followed by 35 cycles of 35 s at 94°C, 45 s at the species-specific annealing temperature (depending on the primer pair), and 60 s at 72°C, finishing with 5 min at 72°C. In the second round of amplification, 2 μL of the first round PCR product, 12.5 μL of DreamTaq PCR Master Mix, 0.5 μM of each internal primer specific to the species, and nuclease-free water were added up to 25 μL. Positive and negative controls, including nuclease-free water as a negative control and positive controls with previously acquired DNA samples from the sarcocysts of the corresponding Sarcocystis spp., were used for both rounds of nPCR. To visualize amplified products, 1% agarose gel (Thermo Fisher Scientific Baltics, Vilnius, Lithuania) electrophoresis was used. Obtained gel was documented using a BioDocAnalyze (Biometra, Gottingen, Germany) system.

Table 1. Data of the PCR primers of Sarcocystis species using farm animals as intermediate hosts used for the isolates from red fox and raccoon dogs in the Czech Republic.
Obtained PCR samples were purified using phosphatase FastAP and exonuclease ExoI (Thermo Fisher Scientific Baltics, Vilnius, Lithuania). Positive PCR samples were subjected to sequencing performed using a Big-Dye® Terminatorv3.1 Cycle Sequencing Kit (Thermo Fisher Scientific Baltics, Vilnius, Lithuania) and a 3500 Genetic Analyzer (Applied Biosystems, Foster City, CA, United States), following the manufacturer’s recommendations. After obtaining the cox1 sequences of Sarcocystis spp., nucleotide BLAST function was employed to compare them with similar ones available in NCBI GenBank.1 The phylogenetic analysis was carried out by the help of MEGA v11.0.13 (36). Multiple alignments were obtained using ClustalW algorithm. The Kimura 2-parameter evolution model with a gamma distribution (K2 + G) was chosen as the best fit to the data for all analyses. Phylogenetic trees were rooted on S. hirsuta. The robustness of phylogenetic trees was tested using bootstrap test with 1,000 replicates. The map was drawn using Datawrapper server.2
Results
In total, 75 out of 197 (38.1%; 95% CI = 31.45-45.15) intestinal mucosa samples of the red fox and two out of three (66.7%; 95% CI = 13.54-98.30) of the raccoon dog were found to be Sarcocystis-positive by light microscopy. Of these, 50 samples of red fox and one of raccoon dog were used for further molecular characterization, which resulted in the identification of five species in the red fox: S. arieticanis, S. capracanis, S. cruzi, S. miescheriana, and S. tenella. At least one Sarcocystis species isolate was present in 21 out of 51 (41.2%) red foxes, while the PCR was negative for the sole raccoon dog.
Sarcocystis arieticanis, S. capracanis, S. cruzi, and S. miescheriana were amplified by using nPCR, and amplified fragments were visible only after the second round of nPCR, whereas S. tenella was observed using the direct PCR. Sequences of these five species obtained in our study (GenBank accession numbers: PP358805–PP358830) were compared to those of the same and closely related Sarcocystis spp. available in GenBank (see Table 2). When comparing the sequences found in this study, the highest intraspecific variation was found for S. miescheriana (94.3–99.7%). Notably, the obtained intraspecific and interspecific genetic variability values for all detected species did not overlap, thus showing that species have been correctly identified.
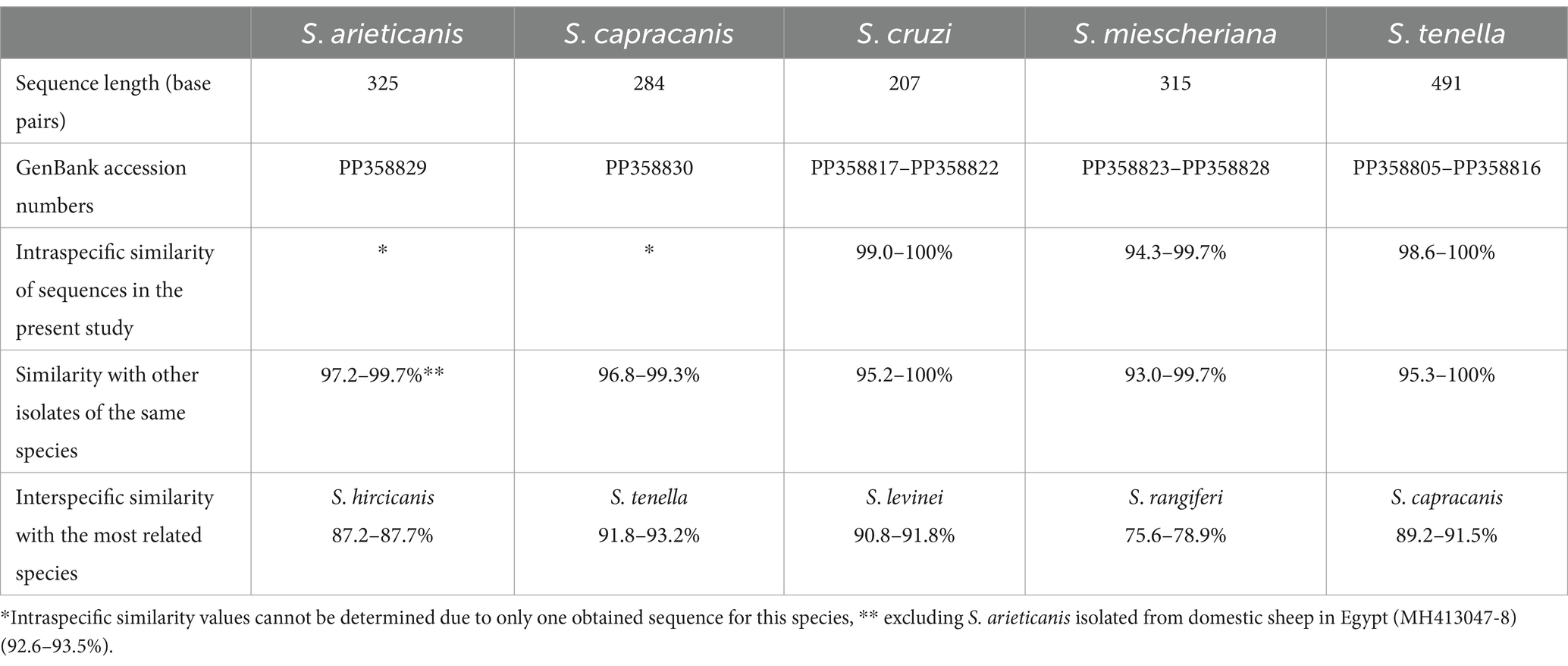
Table 2. Molecular information of the cox1 sequences of five Sarcocystis species found in the large intestine of red fox (Vulpes vulpes) from the Czech Republic.
Sarcocystis spp. coinfections occurred in the large intestine of the red fox. Four different parasite species (S. arieticanis, S. capracanis, S. cruzi, and S. tenella) were found in a single specimen of red fox, S. cruzi/S. tenella and S. cruzi/S. miescheriana were identified in two separate foxes, while the remaining 18 samples were confirmed with a single Sarcocystis species. Among the molecularly confirmed species, S. tenella was the most prevalent, whereas the detection rates of other four Sarcocystis spp. were lower, and two parasite species were only detected in a single fox (Table 3).
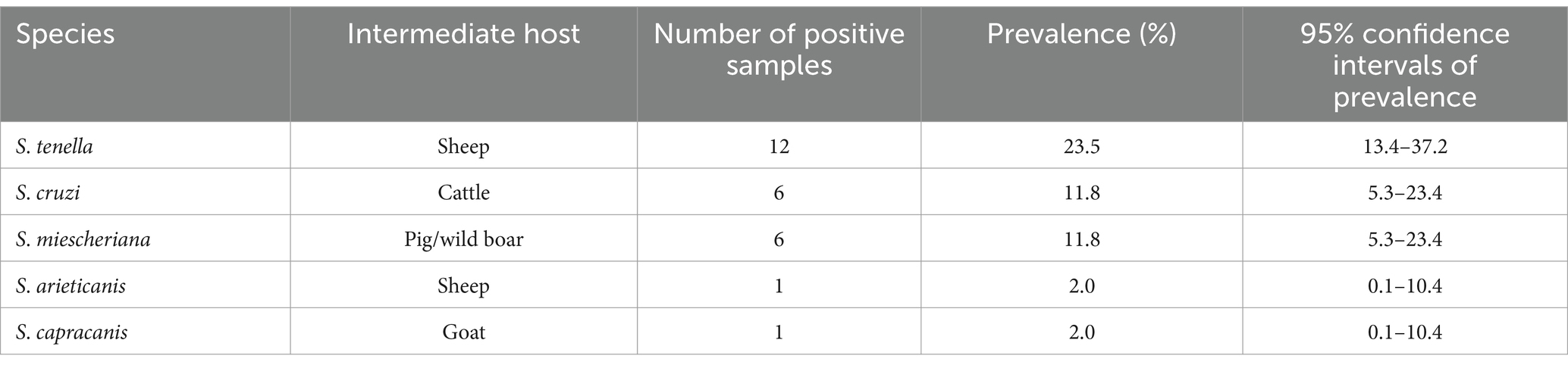
Table 3. Infection parameter (prevalence) of the five Sarcocystis species molecularly confirmed in the red fox (Vulpes vulpes) from the Czech Republic.
The phylogenetic analysis also confirmed the identification of five Sarcocystis species in the large intestines of red foxes. Based on phylogenetic results close relationship was established between S. capracanis and S. tenella (Figure 2A), S. arieticanis was a sister species to S. hircicanis (Figure 2D). Additionally, S. cruzi was sister taxa to S. levinei (Figure 2C), while a relatively high genetic distance was determined comparing S. miescheriana with other Sarcocystis spp. (Figure 2B).
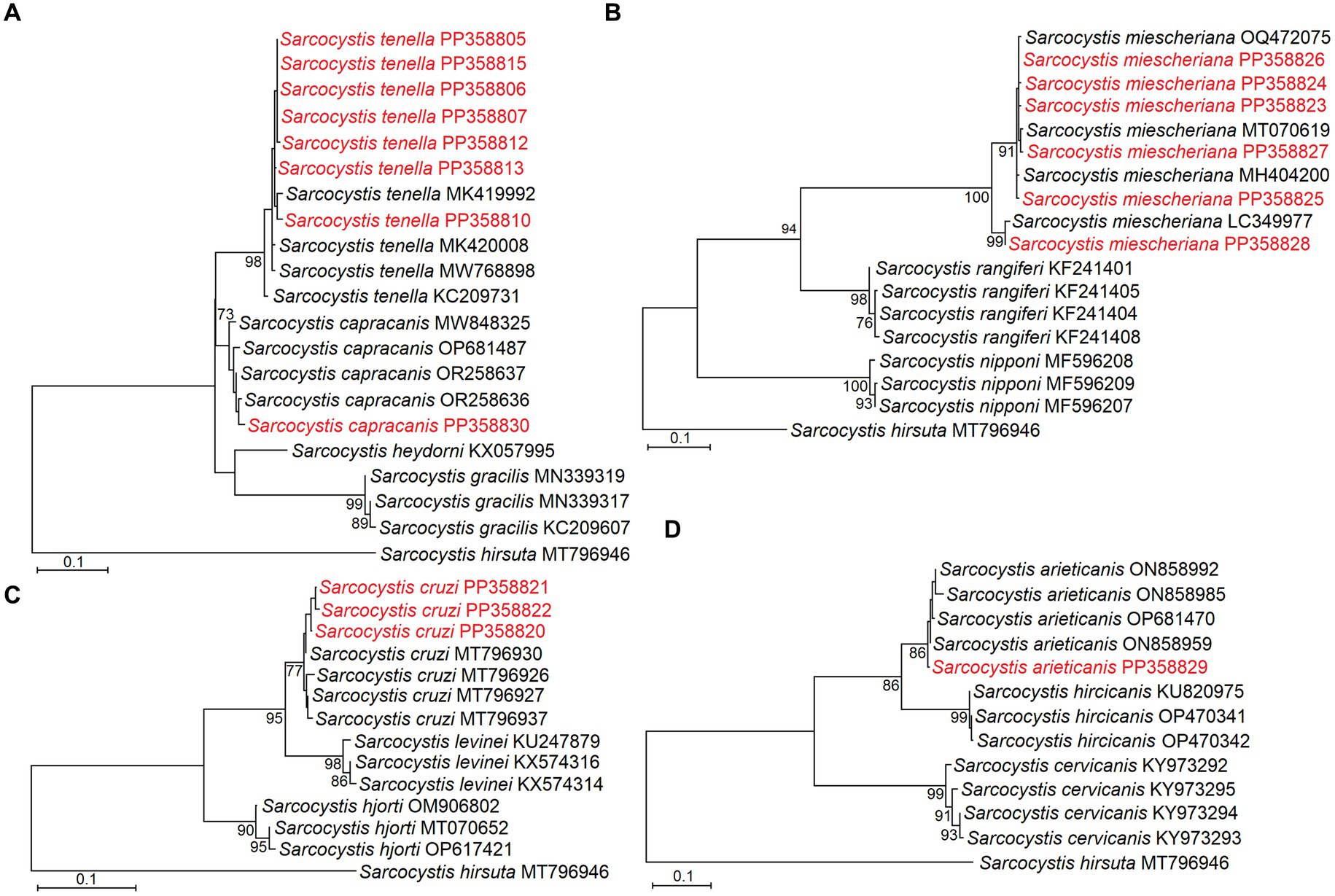
Figure 2. Phylogenetic trees based on cox1 sequences showing placement of Sarcocystis tenella and S. capracanis (A), S. miescheriana (B), S. cruzi (C), and S. arieticanis (D). Sequences representing different haplotypes identified in the present study are marked in red.
The sequences generated in the present study were submitted to the GenBank database under the accession numbers: PP358805–PP358816; PP358817–PP358822; PP358823–PP358828; PP358829; PP358830.
Discussion
The red fox and raccoon dog are one of the most widespread and invasive wild terrestrial carnivores that have been involved in the life cycle of Sarcocystis as either intermediate or definitive hosts (37–40). In the present case, the red fox might serve as definitive host of five Sarcocystis spp., which use farmed animals (e.g., sheep, cattle, pig/wild boar, and goat) as intermediate hosts (41, 42). On the other hand, the raccoon dog was parasitized by sporocysts of a Sarcocystis species, although their molecular characterization resulted in negative PCR and require the use of other sets of primers. Therefore, this is the first report of the red fox as definitive host of Sarcocystis spp. molecularly characterized in the Czech Republic.
The examination of the small intestinal mucosa is a common technique for detecting apicomplexans in individual animals (1, 10, 43–45), since it minimizes the risk of reporting sporocysts coming from the prey (“passage sporocysts”) and allows the finding of higher number of oocysts/sporocysts, which are released in small amounts in feces (18). In this survey, the presence of developmental stages of Sarcocystis spp. in the anterior large intestine demonstrated that this part of the digestive tract is also useful for obtaining epizootiological data on these parasites.
In this study, the values of prevalence in the red fox were higher after the examination of the large intestinal mucosa in comparison to those of molecular analysis. During the first method, the whole Sarcocystis richness is pooled together and might generate overestimated prevalence, while in the second method each species is individually identified, thus resulting in more particular values. The microscopical and molecular approaches are mandatory for the study of these protozoans, although the latter is the best to categorize the Sarcocystis spp. (46). Moré et al. (18) found similar prevalence in the red fox (38.0%) and lower in the raccoon dog (52.6%), whereas Prakas et al. (26) reported lower prevalence (20.0%) of apparently various Sarcocystis spp. and especially of S. rileyi in the red fox and raccoon dog from Lithuania. The contrasting results between surveys should be taken cautiously since data come from different number of samples, climatic seasons, age of hosts, locality, availability of intermediate hosts, parasitological skills of the observer, and proper molecular analysis. Unfortunately, the sporocysts in the raccoon dog were not molecularly characterized and their identity remains uncertain. The present values of prevalence are determined for the first time for five Sarcocystis spp. in the red fox.
Out of the five species herein molecularly identified, S. arieticanis predominantly occurs in sheep [(e.g., 33, 47)], S. capracanis in goat [(e.g., 33, 48)], S. cruzi in cattle [(e.g., 49, 50)], S. miescheriana in wild boar [(e.g., 51–53)], and S. tenella also in sheep [(e.g., 47, 54)]. The role of canids (e.g., dog, jackal, raccoon dog, red fox, and gray wolf) as definitive hosts of these Sarcocystis spp. has been previously confirmed (18). Particularly, the red fox is known as the main scavenger of wildlife (55) and commonly feeds on pigs or wild boars, so its role as definitive host for these five Sarcocystis spp. is possible. The occurrence of these parasite species is likely linked to the presence of canid hosts and the close trophic interaction between predator and prey, as already stated (52).
Previously, DNA of zoonotic S. hominis was detected in a single small intestine mucosal sample of European pine marten (Martes martes) from Lithuania (56). Fortunately, none of the five Sarcocystis spp. found in the present investigation is known to be zoonotic. However, the diagnostic and monitoring of Sarcocystis and other parasites in farm animals should be imperative, since, for example, wild boar might be infected by S. suihominis (52) and cattle by S. hominis and S. heydorni (16, 36, 57), which actually are zoonotic and might be potentially transmitted to humans through the consumption of raw or undercooked meat. On the other hand, domestic pigs experimentally infected with S. miescheriana showed symptoms as reduced weight gain, cutaneous purpura, dyspnea, muscle tremors, abortion, and death (52). The transmission of Sarcocystis spp. through canids to farm animals is more dangerous and cause similar symptoms than those mentioned, as well as fever, anemia, and reduction in milk yield (1).
Since the present survey was based on the use of species-specific primers, some Sarcocystis spp. were absent from the analysis, like S. capreolicanis, S. gracilis, and S. rileyi, that use wild animals as intermediate hosts (1, 26, 40). The cox1 gene clearly differentiated the closely related Sarcocystis spp. in the present study and being very useful for those species having ungulates as intermediate hosts (16, 50). If the complete role of canids in the life cycle of Sarcocystis pretends to be elucidated, samples from more farm animals and wildlife should be examined and characterized.
The presence of Sarcocystis in the intermediate hosts might lead to economic losses or represent a zoonotic risk for humans (8). However, most of studies lacks proper molecular characterization of the species and thus their proper diagnosis and assessment of their importance is overlooked. Therefore, the present findings suggest a potential role of red fox populations in the transmission of Sarcocystis to wild and farmed animals in the study area.
The occurrence of sporocysts in the red fox and raccoon dogs indicates that both wild animal species might be spreading and transmitting these developmental stages (via feces, water, or food) to farm or zoo animals, but probably also to breeders or the staff from zoological gardens. The human activities and destruction of habitats produce a more frequent interaction between canids with farm animals that might produce higher prevalence, as occurred with the European gray wolf and its prey in Central Europe (58), although mesopredators may maintain Sarcocystis life cycles in the absence of the suitable definitive host (18). Particular attention should be paid to the handling process of hunters and/or shepherds in leaving carcasses or viscera infected with Sarcocystis on the ground and that might promote the dissemination of the parasite (59), because after feeding on infected meat, canids begin shedding sporocysts in the environment that might be infective for farmed and wild animals. If possible, reduce the free access of canids to pasture, feeders and water sources in the farms and the exposure of farm animals to feces of wild canids. Frequently, the species of Sarcocystis are non-pathogenic for farm animals, but when sarcocysts are large and evident, significant losses occur in the animal husbandry industry (60).
This is the first study where the potential role of the red fox and raccoon dogs as spreaders of Sarcocystis to farm animals in the Czech Republic is shown. However, more data from other definitive hosts and countries are needed to fulfil the missing data on the main definitive hosts or environmental samples around farms or zoological gardens. Moreover, the huge populations of red fox and raccoon dogs need to be controlled by hunting to avoid the transmission of these and other parasites.
Conclusion
The proper morphological molecular characterization of Sarcocystis spp. is extremely important to identify and thus take the actions to control their spreading through the environment and hosts and ensure food safety and avoid economic losses. This could lead to proper prevention for breeders and avoid potential risks for their animals, as well as to detect pathogenic or zoonotic species that might be transferred to humans. The use of species-specific primers provides a fast and easy method for screening multiple samples for a particular Sarcocystis species. However, it is necessary to use more general primers or cloning of PCR products and sequence a few samples in order to detect a mixed infection with unexpected species, not targeted by species-specific primers.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/supplementary material.
Ethics statement
Ethical approval was not required for the study involving animals in accordance with the local legislation and institutional requirements because animals were killed by the hunters.
Author contributions
OM: Conceptualization, Formal analysis, Funding acquisition, Investigation, Methodology, Writing – original draft, Writing – review & editing. NG: Formal analysis, Methodology, Writing – review & editing. DB: Formal analysis, Methodology, Writing – review & editing. DG-S: Conceptualization, Formal analysis, Supervision, Writing – original draft, Writing – review & editing. PP: Conceptualization, Formal analysis, Methodology, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. The Faculty of Agrobiology, Food and Natural Resources, Czech University of Life Sciences Prague provided open access funding.
Acknowledgments
The authors thank all colleagues from the Pathology and Parasitology Department (SVI Prague). To Janneth Padilla Saldívar from El Colegio de la Frontera Sur for help with the map editing.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Footnotes
1. ^http://blast.ncbi.nlm.nih.gov/, accessed on 14 February 2023.
References
1. Dubey, JP, Calero-Bernal, R, Rosenthal, BM, Speer, CA, and Fayer, R. Sarcocystosis of animals and humans. 2nd ed. Boca Raton, FL: CRC Press (2016).
2. Mehlhorn, H, and Heydorn, AO. The sarcosporidia (Protozoa, Sporozoa): life cycle and fine structure. Adv Parasitol. (1978) 16:43–91. doi: 10.1016/s0065-308x(08)60572-2
3. Rosenthal, BM . Zoonotic Sarcocystis. Res Vet Sci. (2021) 136:151–7. doi: 10.1016/j.rvsc.2021.02.008
4. Dubey, JP, Chapman, JL, Rosenthal, BM, Mense, M, and Schueler, RL. Clinical Sarcocystis neurona, Sarcocystis canis, toxoplasma gondii, and Neospora caninum infections in dogs. Vet Parasitol. (2006) 137:36–49. doi: 10.1016/j.vetpar.2005.12.017
5. Verma, SK, Trupkiewicz, JG, Georoff, T, and Dubey, JP. Molecularly confirmed acute, fatal Sarcocystis falcatula infection in the rainbow lorikeets (Trichoglossus moluccanus) at the Philadelphia zoo. J Parasitol. (2018) 104:710–2. doi: 10.1645/18-78
6. Gonzales-Viera, O, Arranz-Solís, D, Smith, J, Saeij, JPJ, and Mete, A. Fatal Sarcocystis calchasi hepatitis in a captive Indian ringneck parakeet (Psittacula krameri manillensis). Vet Parasitol Reg Stud Rep. (2023) 39:100841. doi: 10.1016/j.vprsr.2023.100841
7. Dessì, G, Tamponi, C, Pasini, C, Porcu, F, Meloni, L, Cavallo, L, et al. Survey on Apicomplexa protozoa in sheep slaughtered for human consumption. Parasitol Res. (2022) 121:1437–45. doi: 10.1007/s00436-022-07469-9
8. Dubey, JP, and Rosenthal, M. Bovine sarcocystosis: Sarcocystis species, diagnosis, prevalence, economic and public health considerations, and association of Sarcocystis species with eosinophilic myositis in cattle. Int J Parasitol. (2023) 53:463–75. doi: 10.1016/j.ijpara.2022.09.009
9. Gjerde, B, Vikøren, T, and Hamnes, IS. Molecular identification of Sarcocystis halieti n. sp., Sarcocystis lari and Sarcocystis truncata in the intestine of a white-tailed sea eagle (Haliaeetus albicilla) in Norway. Int J Parasitol Parasites Wildl. (2018) 7:1–11. doi: 10.1016/j.ijppaw.2017.12.001
10. Máca, O, and González-Solís, D. White-tailed eagle (Haliaeetus albicilla) as the definitive host of Sarcocystis lutrae in the Czech Republic. Front Vet Sci. (2022) 9:981829. doi: 10.3389/fvets.2022.981829
11. Juozaitytė-Ngugu, E, Švažas, S, Šneideris, D, Rudaitytė-Lukošienė, E, Butkauskas, D, and Prakas, P. The role of birds of the family Corvidae in transmitting Sarcocystis protozoan parasites. Animals. (2021) 11:3258. doi: 10.3390/ani11113258
12. Gjerde, B, and Hilali, M. Domestic cats (Felis catus) are definitive hosts for Sarcocystis sinensis from water buffaloes (Bubalus bubalis). J Vet Med Sci. (2016) 78:1217–21. doi: 10.1292/jvms.16-0127
13. Hu, J, Sun, J, Guo, Y, Zeng, H, Zhang, Y, and Tao, J. Infection of the Asian gray shrew Crocidura attenuata (Insectivora: Soricidae) with Sarcocystis attenuati n. sp. (Apicomplexa: Sarcocystidae) in China. Parasit Vectors. (2022) 15:13. doi: 10.1186/s13071-021-05136-z
14. Jäkel, T, Raisch, L, Richter, S, Wirth, M, Birenbaum, D, Ginting, S, et al. Morphological and molecular phylogenetic characterization of Sarcocystis Kani sp. nov. and other novel, closely related Sarcocystis spp. infecting small mammals and colubrid snakes in Asia. Int J Parasitol Parasites Wildl. (2023) 22:184–98. doi: 10.1016/j.ijppaw.2023.10.005
15. Gjerde, B, Giacomelli, S, Bianchi, A, Bertoletti, I, Mondani, H, and Gibelli, LR. Morphological and molecular characterization of four Sarcocystis spp., including Sarcocystis linearis n. sp., from roe deer (Capreolus capreolus) in Italy. Parasitol Res. (2017) 116:1317–38. doi: 10.1007/s00436-017-5410-5
16. Rubiola, S, Civera, T, Panebianco, F, Vercellino, D, and Chiesa, F. Molecular detection of cattle Sarcocystis spp. in north-West Italy highlights their association with bovine eosinophilic myositis. Parasit Vectors. (2021) 14:223. doi: 10.1186/s13071-021-04722-5
17. Gjerde, B, and Dahlgren, SS. Corvid birds (Corvidae) act as definitive hosts for Sarcocystis ovalis in moose (Alces alces). Parasitol Res. (2010) 107:1445–53. doi: 10.1007/s00436-010-2017-5
18. Moré, G, Maksimov, A, Conraths, FJ, and Schares, G. Molecular identification of Sarcocystis spp. in foxes (Vulpes vulpes) and raccoon dogs (Nyctereutes procyonoides) from Germany. Vet Parasitol. (2016) 220:9–14. doi: 10.1016/j.vetpar.2016.02.011
19. Basso, W, Alvarez Rojas, CA, Buob, D, Ruetten, M, and Deplazes, P. Sarcocystis infection in red deer (Cervus elaphus) with eosinophilic myositis/fasciitis in Switzerland and involvement of red foxes (Vulpes vulpes) and hunting dogs in the transmission. Int J Parasitol Parasites Wildl. (2020) 13:130–41. doi: 10.1016/j.ijppaw.2020.09.005
20. Prakas, P, Rudaitytė-Lukošienė, E, Šneideris, D, and Butkauskas, D. Invasive American mink (Neovison vison) as potential definitive host of Sarcocystis elongata, S. entzerothi, S. japonica, S. truncata and S. silva using different cervid species as intermediate hosts. Parasitol Res. (2021) 120:2243–50. doi: 10.1007/s00436-021-07180-1
21. Rogers, KH, Arranz-Solís, D, Saeij, JPJ, Lewis, S, and Mete, A. Sarcocystis calchasi and other Sarcocystidae detected in predatory birds in California, USA. Int J Parasitol Parasites Wildl. (2021) 17:91–9. doi: 10.1016/j.ijppaw.2021.12.008
22. Máca, O, and González-Solís, D. Role of three bird species in the life cycle of two Sarcocystis spp. (Apicomplexa, Sarcocystidae) in the Czech Republic. Int J Parasitol Parasites Wildl. (2022) 17:133–7. doi: 10.1016/j.ijppaw.2022.01.002
23. Šukytė, T, Butkauskas, D, Juozaitytė-Ngugu, E, Švažas, S, and Prakas, P. Molecular confirmation of Accipiter birds of prey as definitive hosts of numerous Sarcocystis species, including Sarcocystis sp., closely related to pathogenic S. calchasi. Pathogens. (2023) 12:752. doi: 10.3390/pathogens12060752
24. Walton, Z, Samelius, G, Odden, M, and Willebrand, T. Variation in home range size of red foxes vulpes along a gradient of productivity and human landscape alteration. PLoS One. (2017) 12:e0175291. doi: 10.1371/journal.pone.0175291
25. Jo, Y, Lee, SJ, Bia, MM, Choe, S, and Jeong, DH. First report of Sarcocystis pilosa from a red fox (Vulpes vulpes) released for the re-introduction project in South Korea. Animals. (2023) 14:89. doi: 10.3390/ani14010089
26. Prakas, P, Liaugaudaitė, S, Kutkienė, L, Sruoga, A, and Švažas, S. Molecular identification of Sarcocystis rileyi sporocysts in red foxes (Vulpes vulpes) and raccoon dogs (Nyctereutes procyonoides) in Lithuania. Parasitol Res. (2015) 114:1671–6. doi: 10.1007/s00436-015-4348-8
27. Irie, T, Uraguchi, K, Ito, T, Yamazaki, A, Takai, S, and Yagi, K. First report of Sarcocystis pilosa sporocysts in feces from red fox, Vulpes schrencki, in Hokkaido, Japan. Int J Parasitol Parasites Wildl. (2019) 11:29–31. doi: 10.1016/j.ijppaw.2019.12.001
28. Kauhala, K, and Kowalczyk, R. Invasion of the raccoon dog Nyctereutes procyonoides in Europe: history of colonization, features behind its success, and threats to native fauna. Curr Zool. (2011) 57:584–98. doi: 10.1093/czoolo/57.5.584
29. Gjerde, B . The raccoon dog (Nyctereutes procyonoides) as definitive host for Sarcocystis spp. of reindeer (Rangifer tarandus). Acta Vet Scand. (1984) 25:419–24. doi: 10.1186/BF03547256
30. Breza, M . Some practical knowledge and suggestions for helminth coprological diagnostics. Helminthology. (1957) 1:57–63.
31. Strazdaitė-Žielienė, Ž, Baranauskaitė, A, Butkauskas, D, Servienė, E, and Prakas, P. Molecular identification of parasitic Protozoa Sarcocystis in water samples. Vet Sci. (2022) 9:412. doi: 10.3390/vetsci9080412
32. Baranauskaitė, A, Strazdaitė-Žielienė, Ž, Servienė, E, Butkauskas, D, and Prakas, P. Molecular identification of protozoan Sarcocystis in different types of water bodies in Lithuania. Life (Basel). (2022) 13:51. doi: 10.3390/life13010051
33. Marandykina-Prakienė, A, Butkauskas, D, Gudiškis, N, Juozaitytė-Ngugu, E, Bagdonaitė, DL, Kirjušina, M, et al. Sarcocystis species richness in sheep and goats from Lithuania. Vet Sci. (2023) 10:520. doi: 10.3390/vetsci10080520
34. Prakas, P, Rehbein, S, Rudaitytė-Lukošienė, E, and Butkauskas, D. Molecular identification of Sarcocystis species in diaphragm muscle tissue of European mouflon (Ovis gmelini musimon) from Austria. Parasitol Res. (2021) 120:2695–702. doi: 10.1007/s00436-021-07212-w
35. Prakas, P, Strazdaitė-Žielienė, Ž, Januškevičius, V, Chiesa, F, Baranauskaitė, A, Rudaitytė-Lukošienė, E, et al. Molecular identification of four Sarcocystis species in cattle from Lithuania, including S. hominis, and development of a rapid molecular detection method. Parasit Vectors. (2020) 13:610. doi: 10.1186/s13071-020-04473-9
36. Tamura, K, Stecher, G, and Kumar, S. MEGA11: molecular evolutionary genetics analysis version 11. Mol Biol Evol. (2021) 38:3022–7. doi: 10.1093/molbev/msab120
37. Šneideris, D, Moskaliova, D, Butkauskas, D, and Prakas, P. The distribution of Sarcocystis species described by ungulates-canids life cycle in intestines of small predators of the family Mustelidae. Acta Parasitol. (2024) 69:747–58. doi: 10.1007/s11686-024-00814-1
38. Pavlásek, I, and Máca, O. Morphological and molecular identification of Sarcocystis arctica sarcocysts in three red foxes (Vulpes vulpes) from the Czech Republic. Parasitol Int. (2017) 66:603–5. doi: 10.1016/j.parint.2017.05.003
39. Kirillova, V, Prakas, P, Calero-Bernal, R, Gavarāne, I, Fernández-García, JL, Martínez-González, M, et al. Identification and genetic characterization of Sarcocystis arctica and Sarcocystis lutrae in red foxes (Vulpes vulpes) from Baltic States and Spain. Parasit Vectors. (2018) 11:173–9. doi: 10.1186/s13071-018-2694-y
40. Dahlgren, SS, and Gjerde, B. The red fox (Vulpes vulpes) and the arctic fox (Vulpes lagopus) are definitive hosts of Sarcocystis alces and Sarcocystis hjorti from moose (Alces alces). Parasitology. (2010) 137:1547–57. doi: 10.1017/S0031182010000399
41. Rubiola, S, Moré, G, Civera, T, Hemphill, A, Frey, CF, Basso, W, et al. Detection of Sarcocystis hominis, Sarcocystis bovifelis, Sarcocystis cruzi, Sarcocystis hirsuta and Sarcocystis sigmoideus sp. nov. in carcasses affected by bovine eosinophilic myositis. Food Waterborne Parasitol. (2024) 34:e00220. doi: 10.1016/j.fawpar.2024.e00220
42. Helman, E, Dellarupe, A, Steffen, KD, Bernstein, M, and Moré, G. Morphological and molecular characterization of Sarcocystis spp. in pigs (Sus scrofa domestica) from Argentina. Parasitol Int. (2024) 100:102859. doi: 10.1016/j.parint.2024.102859
43. Scioscia, NP, Olmos, L, Gorosábel, A, Bernad, L, Pedrana, J, Hecker, YP, et al. Pampas fox (Lycalopex gymnocercus) new intermediate host of Sarcocystis svanai (Apicomplexa: Sarcocystidae). Parasitol Int. (2017) 66:214–8. doi: 10.1016/j.parint.2017.01.021
44. Máca, O, Kouba, M, Korpimäki, E, and González-Solís, D. Molecular identification of Sarcocystis sp. (Apicomplexa, Sarcocystidae) in offspring of Tengmalm's owls, Aegolius funereus (Aves, Strigidae). Front Vet Sci. (2021) 8:804096. doi: 10.3389/fvets.2021.804096
45. Máca, O, Kouba, M, Langrova, I, Panská, L, Korpimäki, E, and González-Solís, D. The Tengmalm's owl Aegolius funereus (Aves, Strigidae) as the definitive host of Sarcocystis funereus sp. nov. (Apicomplexa). Front Vet Sci. (2024) 11:1356549. doi: 10.3389/fvets.2024.1356549
46. Elshahawy, IS, Fawaz, M, Gomaa, A, and Mohammed, E. Prevalence and first molecular identification of Sarcocystis species in feces of domestic dogs (Canis familiaris) in Egypt. BMC Vet Res. (2023) 19:278. doi: 10.1186/s12917-023-03841-8
47. El-Morsey, A, Abdo, W, Sultan, K, Elhawary, NM, and AbouZaid, AA. Ultrastructural and molecular identification of the sarcocysts of Sarcocystis tenella and Sarcocystis arieticanis infecting domestic sheep (Ovis aries) from Egypt. Acta Parasitol. (2019) 64:501–13. doi: 10.2478/s11686-019-00070-8
48. Hu, JJ, Liu, TT, Liu, Q, Esch, GW, Chen, JQ, Huang, S, et al. Prevalence, morphology, and molecular characteristics of Sarcocystis spp. in domestic goats (Capra hircus) from Kunming, China. Parasitol Res. (2016) 115:3973–81. doi: 10.1007/s00436-016-5163-6
49. Moré, G, Basso, W, Bacigalupe, D, Venturini, MC, and Venturini, L. Diagnosis of Sarcocystis cruzi, Neospora caninum, and Toxoplasma gondii infections in cattle. Parasitol Res. (2008) 102:671–5. doi: 10.1007/s00436-007-0810-6
50. Gjerde, B . Molecular characterisation of Sarcocystis bovifelis, Sarcocystis bovini n. sp., Sarcocystis hirsuta and Sarcocystis cruzi from cattle (Bos taurus) and Sarcocystis sinensis from water buffaloes (Bubalus bubalis). Parasitol Res. (2016) 115:1473–92. doi: 10.1007/s00436-015-4881-5
51. Coelho, C, Gomes, J, Inácio, J, Amaro, A, Mesquita, JR, Pires, I, et al. Unraveling Sarcocystis miescheriana and Sarcocystis suihominis infections in wild boar. Vet Parasitol. (2015) 212:100–4. doi: 10.1016/j.vetpar.2015.08.015
52. Gazzonis, AL, Gjerde, B, Villa, L, Minazzi, S, Zanzani, SA, Riccaboni, P, et al. Prevalence and molecular characterisation of Sarcocystis miescheriana and Sarcocystis suihominis in wild boars (Sus scrofa) in Italy. Parasitol Res. (2019) 118:1271–87. doi: 10.1007/s00436-019-06249-2
53. Pacifico, L, Rubiola, S, Buono, F, Sgadari, M, D'Alessio, N, Scarcelli, S, et al. Molecular differentiation of Sarcocystis miescheriana and Sarcocystis suihominis using a new multiplex PCR targeting the mtDNA cox1 gene in wild boars in southern Italy. Res Vet Sci. (2023) 164:105039. doi: 10.1016/j.rvsc.2023.105039
54. Hussein, NM, Hassan, AA, and Abd Ella, OH. Morphological, ultrastructural, and molecular characterization of Sarcocystis tenella from sheep in Qena governorate, upper Egypt. Egypt Acad J Biol Sci E Med Entomol Parasitol. (2018) 10:11–9. doi: 10.21608/EAJBSE.2018.14456
55. Bassi, E, Battocchio, D, Marcon, A, Stahlberg, S, and Apollonio, M. Scavenging on ungulate carcasses in a mountain forest area in northern Italy. Mammal Stud. (2018) 43:1–11. doi: 10.3106/ms2016-0058
56. Prakas, P, Balčiauskas, L, Juozaitytė-Ngugu, E, and Butkauskas, D. The role of mustelids in the transmission of Sarcocystis spp. using cattle as intermediate hosts. Animals. (2021) 11:822. doi: 10.3390/ani11030822
57. Zeng, H, van Damme, I, Kabi, TW, Šoba, B, and Gabriël, S. Sarcocystis species in bovine carcasses from a Belgian abattoir: a cross-sectional study. Parasit Vectors. (2021) 14:271–10. doi: 10.1186/s13071-021-04788-1
58. Lesniak, I, Heckmann, I, Franz, M, Greenwood, AD, Heitlinger, E, Hofer, H, et al. Recolonizing gray wolves increase parasite infection risk in their prey. Ecol Evol. (2018) 8:2160–70. doi: 10.1002/ece3.3839
59. Guardone, L, Armani, A, Mancianti, F, and Ferroglio, E. A review on Alaria alata, Toxoplasma gondii and Sarcocystis spp. in mammalian game meat consumed in Europe: epidemiology, risk management and future directions. Animals. (2022) 12:263. doi: 10.3390/ani12030263
Keywords: red fox, raccoon dog, Czech Republic, farm animals, molecular characterization, Protozoa
Citation: Máca O, Gudiškis N, Butkauskas D, González-Solís D and Prakas P (2024) Red foxes (Vulpes vulpes) and raccoon dogs (Nyctereutes procyonoides) as potential spreaders of Sarcocystis species. Front. Vet. Sci. 11:1392618. doi: 10.3389/fvets.2024.1392618
Edited by:
Vikrant Sudan, Guru Angad Dev Veterinary and Animal Sciences University, IndiaReviewed by:
Giovanni Sgroi, Experimental Zooprophylactic Institute of Southern Italy (IZSM), ItalyIraj Mohammadpour, Shiraz University of Medical Sciences, Iran
Copyright © 2024 Máca, Gudiškis, Butkauskas, González-Solís and Prakas. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: David González-Solís, ZGdvbnphbGVAZWNvc3VyLm14
†ORCID: Ondřej Máca, http://orcid.org/0000-0002-6311-0851
Naglis Gudiškis, http://orcid.org/0009-0002-7184-8167
Dalius Butkauskas, http://orcid.org/0000-0003-2892-0493
David González-Solís, http://orcid.org/0000-0001-7765-2865
Petras Prakas, http://orcid.org/0000-0002-5115-2659