 Ezequiel Hidalgo-Hermoso1*
Ezequiel Hidalgo-Hermoso1* Javier Cabello2Rodrigo Lopez3Vicente Vergara-Wilson4Frank Vera5Carola Valencia5Carlos Sanchez6Sebastian Celis4Alejandra Silva7Aintzane Cariñanos7Ismael Barria7Rocio Vicencio2
Javier Cabello2Rodrigo Lopez3Vicente Vergara-Wilson4Frank Vera5Carola Valencia5Carlos Sanchez6Sebastian Celis4Alejandra Silva7Aintzane Cariñanos7Ismael Barria7Rocio Vicencio2 Sebastián Muñoz-Leal8Paula Aravena8Rocio Lagos9Juan Toro-Letelier10Sebastián Verasay-Caviedes1,9
Sebastián Muñoz-Leal8Paula Aravena8Rocio Lagos9Juan Toro-Letelier10Sebastián Verasay-Caviedes1,9 Antonio Garnham11Irene Peña12,13
Antonio Garnham11Irene Peña12,13 Fernando Sánchez13
Fernando Sánchez13 Dario Moreira-Arce14,15
Dario Moreira-Arce14,15 Pablo M. Vergara14
Pablo M. Vergara14 Raul Alegria-Moran16
Raul Alegria-Moran16 Galaxia Cortés-Hinojosa13*
Galaxia Cortés-Hinojosa13*- 1Fundacion Buin Zoo, Buin, Chile
- 2Centro de Conservación de la Biodiversidad Chiloé-Silvestre, Ancud, Chile
- 3Aumen ONG, Coyhaique, Chile
- 4Departamento de Veterinaria, Parque Zoológico Buin Zoo, Buin, Chile
- 5School of Veterinary Medicine, Facultad de Ciencias de la Naturaleza, Universidad San Sebastian, Puerto Montt, Chile
- 6Veterinary Medical Center, Oregon Zoo, Portland, OR, United States
- 7Departamento de Areas Silvestres Protegidas, Corporacion Nacional Forestal, Punta Arenas, Chile
- 8Departamento de Ciencia Animal, Facultad de Ciencias Veterinarias, Universidad de Concepción, Chillán, Chile
- 9Laboratorio Clínico, Hospital Veterinario SOS Buin Zoo, Buin, Chile
- 10Facultad de Cs Veterinarias y Pecuarias, Univeridad de Chile, Santiago, Chile
- 11Escuela de Medicina Veterinaria, Universidad Mayor, Santiago, Chile
- 12Facultad de Ciencias de la Vida, Universidad Andrés Bello, Santiago, Chile
- 13Escuela de Medicina Veterinaria, Facultad de Agronomía e Ingeniería Forestal, Facultad de Ciencias Biológicas, Facultad de Medicina, Pontificia Universidad Católica de Chile, Santiago, Chile
- 14Departamento de Gestión Agraria, Universidad de Santiago de Chile (USACH), Santiago, Chile
- 15Institute of Ecology and Biodiversity (IEB), Santiago, Chile
- 16Escuela de Medicina Veterinaria, Sede Santiago, Facultad de Recursos Naturales y Medicina Veterinaria, Universidad Santo Tomás, Santiago, Chile
Introduction: Herpesvirus infections have been highlighted as emerging diseases affecting wildlife health and the conservation of several taxa. Malignant catarrhal fever (MCF) and infectious keratoconjunctivitis (IKC) are two viruses that infect wild ruminants. Nevertheless, epidemiological data on herpesviruses in South American wild ruminants are limited. An outbreak of caprine gammaherpesvirus-2 (CpHV-2) that recently was suspected as the cause of MCF in southern pudus (Pudu puda) prompted the need to conduct molecular screenings in Chilean cervids to understand the epidemiology of herpesviruses. The aim of this study was to determine the occurrence and genetic diversity of herpesviruses in free-ranging cervids from Chile.
Methods: Herpesvirus infection was assessed in antemortem blood samples (n = 86) from pudus (n = 81) and huemuls (Hippocamelus bisulcus) (n = 5), as well as postmortem samples of spleen (n = 24) and lung (n = 3) from pudus, using a nested pan-herpesvirus PCR assay.
Results: Combining all suitable sample types, DNA of pudu gammaherpesvirus-1 was detected in five pudues and five huemuls, with an overall prevalence of 9.90% (n = 10/101; 95% CI = 5.11–17.87%). One pudu tested positive for ovine gammaherpesvirus-2 (n = 1/96; 1.04%; 95% CI = 0.05–6.49%), and one pudu tested positive for a Macavirus sequence with 98.63 similarity to ovine gammaherpesvirus-2 (n = 96; 1.04%; 95% CI = 0.05–6.49%).
Discussion: To the best of our knowledge, this is the first report of a herpesvirus in huemul and of ovine gammaherpesvirus-2 in Chile. Our results also confirm the active circulation of herpesvirus in free-ranging cervids in Chilean Patagonia, and as such, MCF should be considered as a possible cause of disease in free-ranging Chilean pudus and livestock species. Further research is necessary to develop a plan of systematic monitoring (serological and pathological screening) of herpesviruses in Chilean wild and domestic ruminants to understand their diversity and impact on animal health and conservation.
1 Introduction
Herpesviruses are a large group of linear, non-segmented, double-stranded, enveloped DNA viruses that infect many vertebrate and invertebrate species (1). The Herpesviridae family includes three subfamilies, Alphaherpesvirinae, Betaherpesvirinae, and Gammaherpesvirinae, all of which can establish latency in different organs (1, 2). Herpesvirus infections have been cataloged as emerging diseases in wild mammals. For example, hemorrhagic diseases in elephants are caused by elephant endotheliotropic herpesvirus (3, 4), and malignant catarrhal fever (MCF) is caused by several macaviruses in ruminant species (5–8). All these agents can cause devastating illness at the individual or population level and represent a threat to animal health and conservation. In cervids, different species of the Alpha-, Beta-, and Gammaherpesvirinae subfamilies have been described as potential agents of various diseases, including cervid herpesvirus-2 (CvHV-2), which causes infectious keratoconjunctivitis (9), alcelaphine gammaherpesvirus-1 (AlHV-1), alcelaphine gammaherpesvirus-2 (AlHV-2), ovine gammaherpesvirus-2 (OvHV-2), caprine gammaherpesvirus-2 (CpHV-2), hippotragine HV-1 (HiHV-1), ibex-MCFV, and caprine gammaherpesvirus-3, which cause MCF (10–16), and bovine herpesvirus-1, which causes infectious bovine rhinotracheitis (17). Although herpesviruses have been widely investigated, little is known about herpesvirus prevalence and host specificity in cervid species in wild environments. Indeed, to date, most of the reports come from populations under human care (18–21). In Chile, caprine gammaherpesvirus-2 was recently reported in southern pudus (Pudu puda) from a zoological collection (or zoo), possibly associated with an MCF outbreak (22). Additionally, a new pudu gamma-herpesvirus-1 was detected in a rescued pudu at Chiloé Island (22). However, there have been no reports of gammaherpesvirus infection or cases of MCF in livestock in Chile to date.
There are three species of cervids native to Chile: the taruka (Hippocamelus antisensis) restricted to the north of the country; the huemul (Hippocamelus bisulcus) distributed in Patagonia and central Chile; and the southern pudu [with the largest populations throughout temperate coastal and Andean forests (36–49°S)] (23, 24). In Chile, the pudu is classified as vulnerable, and the huemul is classified as endangered (25). The main threats to the conservation of both species include fragmentation of their habitats, traumatic events such as road kills by cars, attacks by dogs, and emerging diseases potentially transmitted via livestock (22, 24, 26–32). The present study aimed to investigate the presence of herpesvirus DNA and genetically characterize it in the blood, spleen, and lung samples from free-ranging southern pudus and huemuls in Chile.
2 Materials and methods
2.1 Animal sampling
2.1.1 Pudu
Between July 2017 and June 2022, 96 healthy or ill pudus from three wildlife rescue centers, two in the Los Lagos region (USS: Universidad San Sebastian, Ch S: Chiloe Silvestre) and one in the Ñuble region (UC: Universidad de Concepcion), were prospected. Blood or samples of organs were collected on the day of admission or 24 h later during necropsy in the case of animals that perished after being admitted. A physical examination of the animals was performed before taking blood samples. Using a standard vacutainer system (Vacutainer, Beckon, Dickson, and Company, Franklin Lakes, New Jersey, USA), 2–4 mL of blood was collected from the cephalic or saphenous vein and stored at −20°C until processing. This procedure was performed after sedating the animals with 0.04 mg/kg of dexmedetomidine (Dexdomitor, Zoetis, Santiago, Chile), and subsequently, the sedation was reversed with 0.4 mg/kg of atipamezole (Antised, Zoetis), both of which were administered intramuscularly (26). The spleen and lung samples were collected using fine-needle aspirates. None of the animals were euthanized; instead all of them died because of different causes, including trauma, such as being hit by a car, attack by dogs, or presumptive infectious diseases. All samples were stored at −20°C until DNA extraction. According to the guidelines of the authors' institutions, a formal approval by the ethical committee was not required.
2.1.2 Huemul
Blood samples were taken from healthy adult free-ranging huemuls captured in April–May 2019 in Torres del Paine National Park (468010 580 0S, 718580 370 0W), Chilean Patagonia. Their captures were carried out under the supervision of wildlife veterinarians, biologists, and park rangers and authorized by permits 2107/2019 and 256/2019 of the Servicio Agricola y Ganadero and National Forest Corporation, respectively. The animals were anesthetized with a combination of medetomidine (0.09 mg/kg) and ketamine (2 mg/kg) administered intramuscularly (33). Blood was collected from the cephalic vein, placed in EDTA-coated tubes, and kept at −20°C until analysis. All animals were released in the same place immediately after recovery from anesthesia.
2.2 Molecular characterization
DNA extractions were carried out using a Quick-DNA/RNA Viral Kit (Zymo Research, Irvine, CA, USA) and quality-checked with PCR targeting the mammalian mitochondrial cytochrome oxidase 1 (COX1; internal control) (34). Nucleic acid concentrations were measured with a Qubit 4 Fluorometer (Invitrogen) or spectrophotometer (MaestroGen). Up to 100 ng of DNA/RNA were used for each PCR reaction. The first screening for herpesviruses targeting a ~200 bp fragment of the herpesviral DNA polymerase gene (Dpol) was implemented from a previously published protocol (35). Positive and potential samples were then submitted to a second PCR protocol to amplify the ~500 bp fragment of the glycoprotein B gene (GlyB) (36). For all PCR assays, we used the Sapphire Amp Fast PCR Master Mix (Cat. No. RR350B, Takara Bio), with all protocols adapted. The PCR products were electrophoresed in 1% agarose gels. Fragments of the expected size were excised, and the gel was extracted using an E.Z.N.A.® MicroElute® Gel Extraction Kit (Omega Bio-tek) following the manufacturer's instructions. Products were then submitted for sequencing to the Plataformas Omicas UC, where they used an ABI PRISM 3500 xL DNA sequencer (Applied Biosystems®, USA). Sequences obtained were assembled with Geneious Prime v. 2022.2.2, and the primers were removed. The edited sequences were then submitted to GenBank, and BLASTN (megablast)/BLASTP (protein-protein BLAST) analyses were performed under a default setting to infer similarities with sequences available in GenBank.
2.3 Phylogenetic analysis
Phylogenetic analyses were performed with the amino acid sequences. Alignments were constructed for 59 herpesviruses and 44 sequences of Dpol and GlyB using MAFFT (37). An alphaherpesvirus was designated as the outgroup for the analysis. The phylogeny was inferred by Bayesian statistics with MrBayes 3.2.7a in the CIPRES science gateway (38). The trees were run for a maximum of 1,000,000 generations, with four chains, three hot chains, and one cold chain with the default heating parameter (temperature = 0.2), sampling every 100 generations, and the first 20% of the MCMC samples were discarded as burn-in. Figures were edited in FigTree v1.4.4 (39).
2.4 Data analysis
Positivity rate (prevalence) and 95% confidence intervals were estimated (when possible) using approximation and null hypothesis testing (positivity rate equals to 0) (40). Statistically significant differences were set at a p-value of < 0.05.
3 Results
Overall, 81 samples of blood were collected antemortem and 24 samples of spleen and three samples of lung were collected during the necropsy of pudus. Furthermore, five huemuls (two male and three female huemuls) were captured, and blood samples were collected as stated above. Most samples were obtained from individuals with no clinical signs of MCF at the time of sampling, except for 10 animals suspected to be clinically ill, including one animal with clinical signs consistent with MCF, such as fever (40.2°C temp), weakness, lethargy, and nasal mucous discharge.
All DNA extractions were positive for the COX1 gene, with concentrations ranging from 31.5 to 56.2 ng/μL. Regarding herpesvirus detection, partial sequences were acquired from seven pudus and five huemuls, with a partial Dpol sequence identified in four pudus and five huemuls. Additionally, five pudus and four huemuls tested positive for the GlyB gene (Table 1).
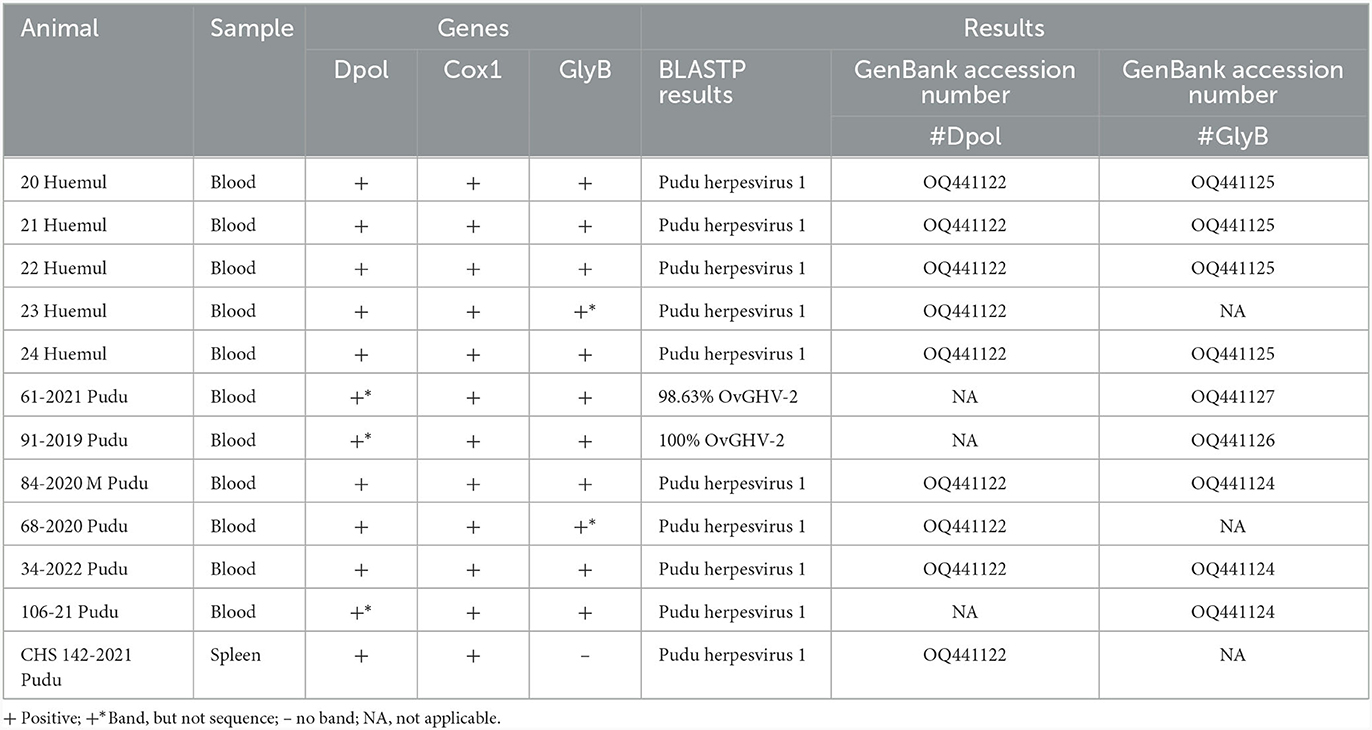
Table 1. Samples of Chilean Cervids positive for herpesvirus.
In the case of Dpol, a host species-specific consensus sequence of 175 bp was generated after primer removal for all positive pudus (GenBank accession OQ441122) and all positive huemuls (GenBank accession OQ441123). Pudu and huemul consensus and individual sequences were submitted for BlastN and BlastP analyses independently. In the case of pudu, BLASTN and BLASTP indicated that our sequences were 100% identical to pudu gammaherpesvirus-1 (LC667446.1/BDD79276.1). In the case of huemul, the consensus BLASTN indicated a 95.62% sequence identity to pudu gammhaerpesvirus-1 (LC667446.1). However, BLASTP analysis yielded a 100% sequence identity to pudu gammhaerpesvirus-1 (BDD79276.1), which indicated that both species were positive for the same pudu gammaherpesvirus-1. For the second gene, GlyB, we obtained a 467 bp length sequence from two pudus (GenBank accession OQ441124). Furthermore, we acquired sequences for three animals, all of which were exclusively identified as positive with GlyB. This contributes to the total of positive pudues by sequencing to seven (Table 2). One of them had a sequence that matches with pudu gammaherpesvirus-1. A second pudu with clinical signs consistent with MCF (GenBank accession OQ441127) had a 440 bp sequence with a 98.63% sequence identity with BLASTP to ovine gammaherpesvirus-2 (AAK28846.1), and the third animal (GenBank accession OQ441126) had a 484 bp sequence with a 100% sequence identity with BLASTP to ovine gammaherpesvirus-2. Finally, four of the five huemuls had a sequence for GlyB (GenBank accession OQ441125). In the GlyB of pudu gammhaerpesvirus-1, similar to Dpol, the sequences obtained had high intraspecies similarity with a 100% match at the amino acid level, among sequences of the same host. Consequently, the consensus sequences of the pudu samples and huemul samples were further evaluated. The BLASTP analysis of the pudu consensus sequence confirmed a 100% sequence identity with pudu gammaherpesvirus-1 (BDD79278.1). In the case of BLASTP, the analysis revealed a sequence identity of 99.24% to pudu gammaherpesvirus-1 (BDD79278.1). In this case, we identified an aminoacidic change from S to N at the 25th position of GlyB in our alignment. This result is consistent with the data obtained from the comparison of the pudu amino acid sequence obtained in the laboratory with 99.24% sequence identity and the same point of aminoacidic change from S to N. Pudu gammaherpesvirus-1 was detected in 9.90% (10/101; 95% CI = 5.11–17,87%) of the samples: all huemul specimens (100%; 5/5; 95% CI = 46.29–100%) and five pudus (5.20%; 5/96; 95% CI = 1.93–12.30%), with one pudu testing positive for ovine gammaherpesvirus-2 (1.04%; 1/96; 95% CI = 0.05–6.49%) and one pudu testing positive for a 98% compatible ovine gamma-herpesvirus-2 (1.04%; 1/96; 95% CI = 0.05–6.49%). Details related to the detection on the different sampled tissues are presented in Table 2.

Table 2. Herpesvirus-positive samples by species and tissue type.
Regarding phylogenetic reconstruction, pudu gammhaerpesvirus-1 clustered into a clade with Rhadinovirus (Figures 1, 2). The phylogenetic analysis for the GlyB gene indicates that the virus forms detected in pudu 91 2019 and pudu 61 2021 are clade within the Macavirus genus (Figure 2), and both are sister taxa of ovine gammaherpesvirus-2.
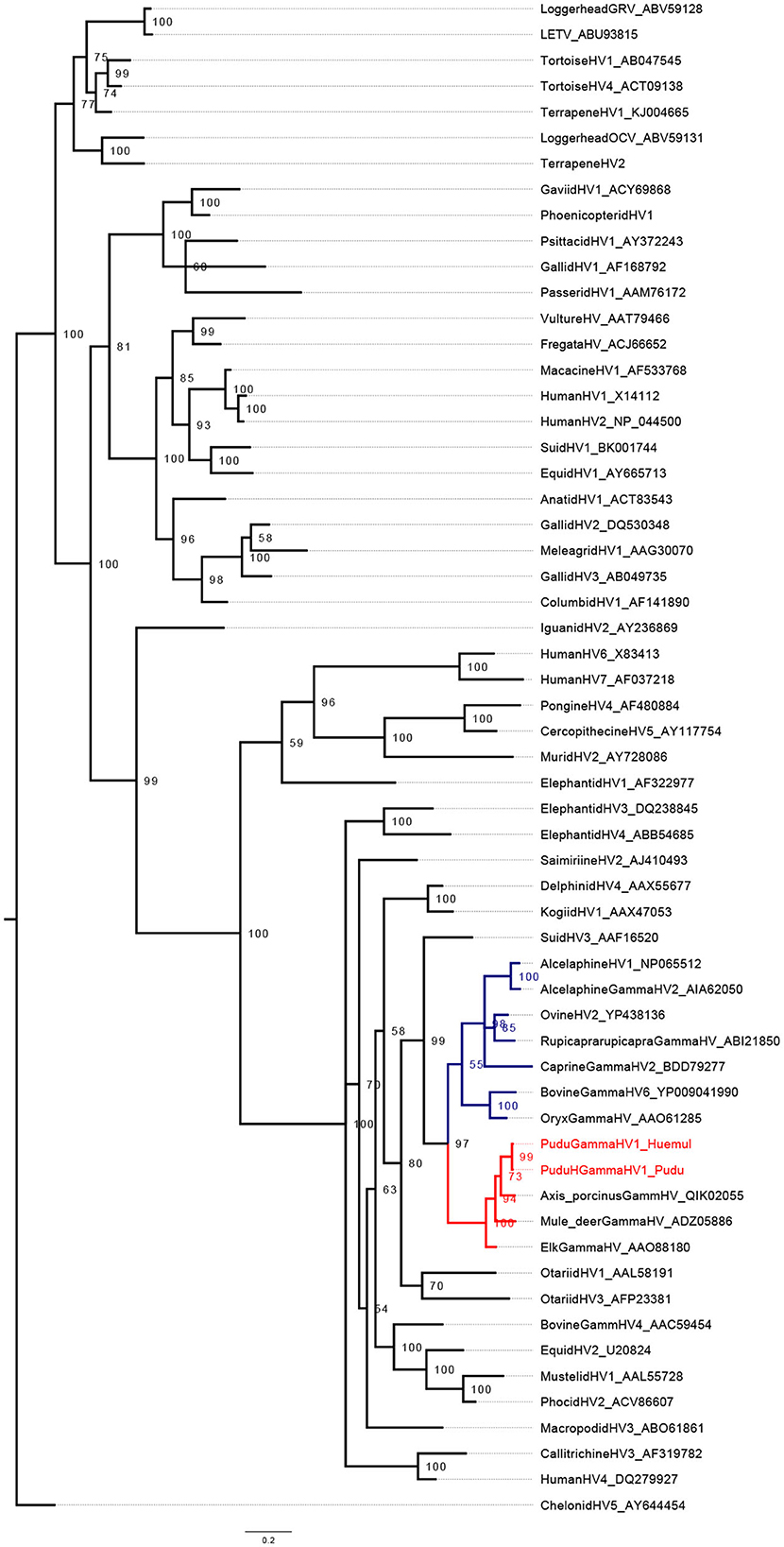
Figure 1. Midpoint-rooted Bayesian phylogenetic tree showing the relationship of pudu gammaherpesvirus-1 clustering in the genus Rhadinovirus (in red). The genus Macavirus is highlighted in blue. The analysis was carried out on amino acid (AA) sequences of partial herpes Dpol (176 AA characters including gaps).
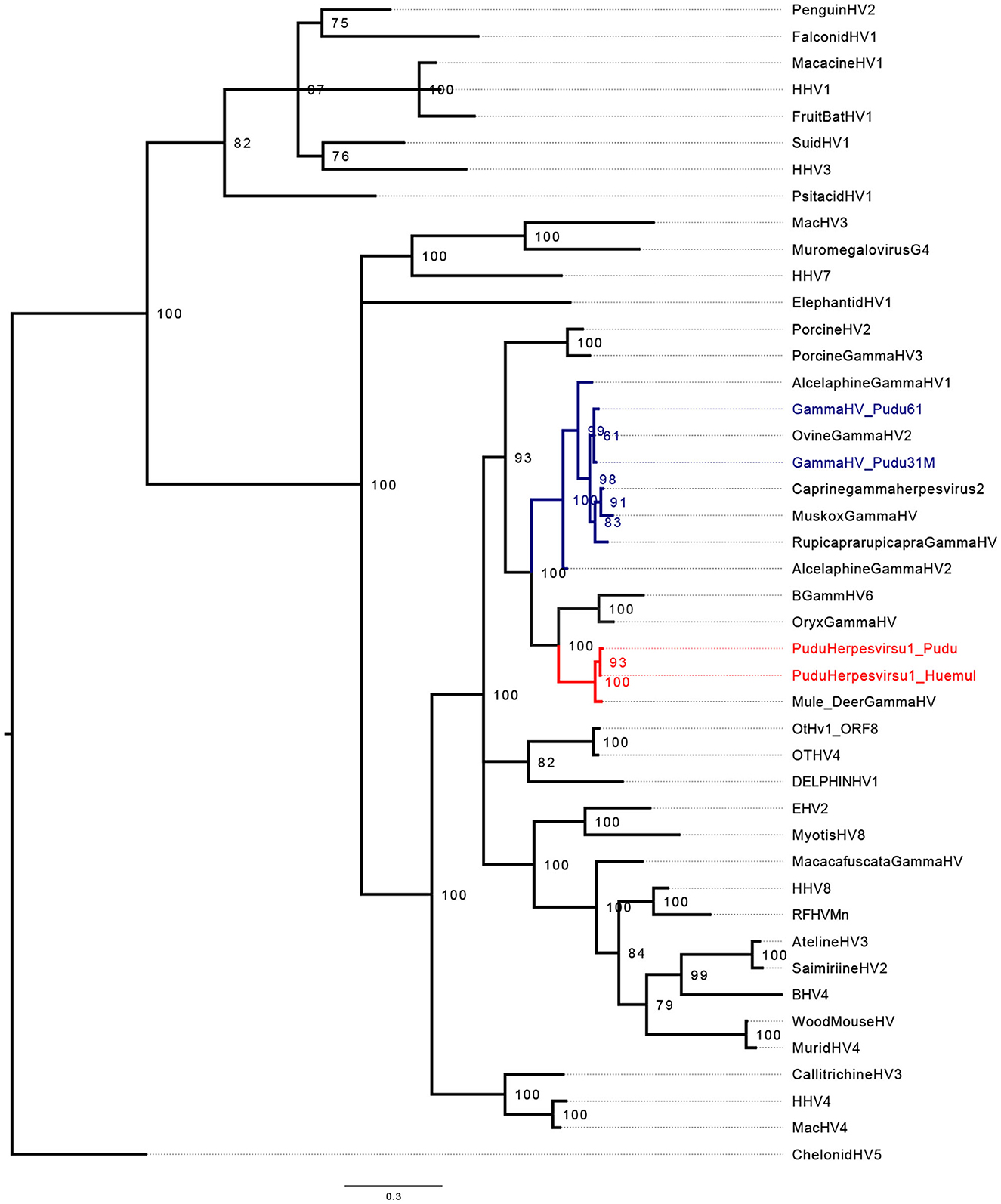
Figure 2. Midpoint-rooted Bayesian phylogenetic tree showing the relationship of pudu gammaherpesvirus-1 clustering in the genera Rhadinovirus (in red) and Macavirus (in blue). The analysis was carried out using the amino acid (AA) sequences for partial herpes GlyB (203 AA characters, including gaps and poorly aligned sequences).
4 Discussion
This is the first systematic large-scale screening study conducted over a period of 5 years seeking herpesvirus DNA in any threatened free-ranging cervid species. This study describes the first report of ovine gammaherpesvirus-2 in Chilean ruminants. Our results confirmed that pudu gammaherpesvirus-1 is circulating in free-ranging Chilean pudus and huemuls (22). The occurrence of pudu gammaherpesvirus-1 in free-ranging huemuls in Torres del Paine National Park and in pudus of Chiloe, an island located ~1,100 km to the north, suggests that the detected virus circulates in Chilean wild deer despite the distance and geographic barriers between populations.
Previous molecular surveys of herpesvirus in cervid species showed variable levels of occurrence, ranging from 0.01% of ovine gammaherpesvirus-2 in sika deer (Cervus nippon) (19) in China to 4.7% of Elk gammaherpesvirus in Javan rusa (Rusa timorensis) in the USA (20). Both studies were conducted on animals under human care. In free-ranging cervids, the only comparative study was carried out among Eurasian tundra reindeer (Rangifer tarandus tarandus), moose (Alces alces), and red deer (Cervus elaphus), where herpesvirus was detected in 48.6% (17/35) of reindeers, while all moose and red deer were negative (2). The obtained sequences from this study were clearly different from other ruminant gammaherpesvirus sequences available from GenBank.
This is the first report of a pudu with clinical signs of MCF confirmed to be infected by a macavirus that is 98.63% similar to ovine gammaherpesvirus-2 for the GlyB gene. The pudu positive to with a to ovine gammaherpesvirus-2 was remitted dead to the rescue center as a consequence of an attack by dogs. However, no other clinical signs were observed on admission. Moreover, coinfections of different herpesvirus strains in the same individual were not observed. None of the screened samples from either pudu or huemul were positive for caprine gammaherpesvirus-2, suggesting that only deer in contact with goats, which are the natural reservoir of this virus, would be exposed to the contagion. Additional molecular surveys in goats of Patagonia will be necessary to rule out the caprine gammaherpesvirus-2 presence in the region.
Mortalities in captive or free-ranging artiodactyls associated with gammaherpesvirus are caused mainly by Macavirus and MCF (15, 16, 18–20, 41–51), suggesting that the current and future causes of mortality in both species of Chilean deer could be linked to infection by ovine gammaherpesvirus-2 or caprine gammaherpesvirus-2. The fact that fatal cases of infection by MCF have not been reported in Chilean cervids to date could be because of the inability to detect sick and dead animals in the field, calling for further efforts. A fluid communication between investigators and rescue centers that receive wild deer could be a solution to fill this gap. Notably, ovine gammaherpesvirus-2 usually causes diseases in domestic ruminants (8, 52, 53) and represents an emerging health threat to the herds of Chiloe Island and Chilean Patagonia.
The detection of pudu gammaherpesvirus-1 (100%) in five apparently healthy huemuls in Torres del Paine calls into question their origin and potential pathogenic role in Chilean cervids. Previously, the genus Rhadinovirus included all herpesviruses that cause MCF in cervids and other ungulates; in 2005, this clade was moved to the Macavirus genus by the International Committee on Taxonomy of Viruses. Currently, only one report of encephalitis induced by a newly discovered ruminant Rhadinovirus (RuRv) has been reported, in a free-ranging Formosan sambar deer (Rusa unicolor swinhoei) (54). It is notable that seven uteruses from hinds that were aborted were positive for cervid Rhadinovirus type-2 (CRhV-2) through PCR in a deer farm in New Zealand; however, the role of the virus in abortion needs clarification (55). Recent reports show novel Rhadinovirus sequences in cervid species with no evidence of disease (20, 56, 57); therefore, whether all Rhadinoviruses are pathogenic or not needs support. Finally, the pathogenicity and epidemiology of pudu gammaherpesvirus-1 in livestock is unknown. It is currently necessary to perform genetic screenings in sheep, goats, and cattle sharing the ecosystem with pudus from Patagonia to tackle this question.
5 Conclusion
A novel gammaherpesvirus, herein named pudu gammaherpesvirus-1, was detected in two species of cervids in Chilean Patagonia with no apparent health problems. Some gammaherpesviruses should be considered as a possible cause of illness in free-ranging pudus and domestic ruminants in Chilean Patagonia. The ovine gammaherpesvirus-2 detected in one rescued pudu with clinical signs of MCF could represent a risk for the health and conservation of the species due to its negative effects on wild populations as a direct (fatal infection) or indirect cause (weakness in the pudu that facilitates predation by feral dogs). Further studies, including serological and pathological screening, are needed to understand the diversity of herpesviruses in Chilean wild and domestic ruminants and their impact on animal health and conservation.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/supplementary material.
Ethics statement
Ethical approval was not required for the study involving animals in accordance with the local legislation and institutional requirements because this is not required for wild animal captures/management (Huemul) under the Chilean Legislation. The pudu samples proven from rehabilitation centers Frozen Bank.
Author contributions
EH-H: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Writing – original draft, Writing – review & editing. JC: Data curation, Funding acquisition, Resources, Writing – review & editing. RL: Data curation, Funding acquisition, Investigation, Writing – review & editing. VV-W: Investigation, Resources, Writing – review & editing. FV: Data curation, Resources, Writing – review & editing. CV: Data curation, Resources, Writing – review & editing. CS: Funding acquisition, Investigation, Resources, Writing – original draft, Writing – review & editing. SC: Data curation, Funding acquisition, Resources, Writing – review & editing. AS: Resources, Writing – review & editing. AC: Investigation, Resources, Writing – review & editing. IB: Investigation, Resources, Writing – review & editing. RV: Data curation, Formal analysis, Investigation, Resources, Writing – review & editing. SM-L: Data curation, Resources, Writing – review & editing. PA: Data curation, Resources, Writing – review & editing. RL: Investigation, Resources, Writing – review & editing. JT-L: Investigation, Writing – review & editing. SV-C: Data curation, Investigation, Writing – original draft, Writing – review & editing. AG: Data curation, Investigation, Writing – original draft, Writing – review & editing. IP: Investigation, Methodology, Resources, Writing – review & editing. FS: Investigation, Resources, Writing – review & editing. DM-A: Investigation, Resources, Writing – review & editing. PV: Investigation, Resources, Writing – review & editing. RA-A: Formal analysis, Investigation, Methodology, Resources, Writing – original draft, Writing – review & editing. GC-H: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. The research for this study was funded by Fundacion Buin Zoo and Chiloe Silvestre. The funds for publication of this article were supported by ISA INTERCHILE and ISA INTERVIAL and Oregon Zoo. DM-A thanks the grant received from ANID/BASAL with the grant number FB210006. GC-H thanks the grant received from Fondecyt with the grant number 11200302. SM-L was funded by Fondecyt Iniciación 11220177.
Acknowledgments
The authors acknowledge all staff from the Universidad San Sebastian and Universidad de Concepción wildlife rescue centers and Chiloe Silvestre for their support in providing the samples. Additionally, the authors thank CONAF personnel for their valuable support with huemul captures.
Conflict of interest
EH-H and SV-C were employed by Fundacion Buin Zoo. RL was employed by Aumen ONG. VV-W and SC were employed by Parque Zoológico Buin Zoo. CS was employed by Oregon Zoo. AS, AC, and IB were employed by Corporacion Nacional Forestal.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Wozniakowski G, Samorek-Salamonowicz E. Animal herpesviruses and their zoonotic potential for cross-species infection. Ann Agric Environ Med. (2015) 22:191–4. doi: 10.5604/12321966.1152063
2. das Neves CG, Sacristán C, Madslien K, Tryland M. Gammaherpesvirus in cervid species from Norway: characterization of a new virus in wild and semi-domesticated Eurasian tundra reindeer (Rangifer tarandus tarandus). Viruses. (2020) 12:876. doi: 10.3390/v12080876
3. Guntawang T, Sittisak T, Kochagul V, Srivorakul S, Photichai K, Boonsri K, et al. Pathogenesis of hemorrhagic disease caused by elephant endotheliotropic herpesvirus (EEHV) in Asian elephants (Elephas maximus). Sci Rep. (2021) 11:12998. doi: 10.1038/s41598-021-92393-8
4. Richman LK, Montali RJ, Garber RL, Kennedy MA, Lehnhardt J, Hildebrandt T, et al. Novel endotheliotropic herpesviruses fatal for Asian and African elephants. Science. (1999) 283:1171–6. doi: 10.1126/science.283.5405.1171
5. Plowright W, Ferris RD, Scott GR. Blue Wildebeest and the ætiological agent of bovine malignant catarrhal fever. Nature. (1960) 188:1167–9. doi: 10.1038/1881167a0
6. Gong M, Myster F, van Campe W, Roels S, Mostin L, van den Berg T, et al. Wildebeest-derived malignant catarrhal fever: a bovine peripheral T cell lymphoma caused by cross-species transmission of alcelaphine gammaherpesvirus 1. Viruses. (2023) 15:526. doi: 10.3390/v15020526
7. Pfitzer S, Last R, Espie I, van Vuuren M. Malignant catarrhal fever: an emerging disease in the African buffalo (Syncerus caffer). Transbound Emerg Dis. (2015) 62:288–94. doi: 10.1111/tbed.12131
8. Li H, Cunha CW, Taus NS, Knowles DP. Malignant catarrhal fever: inching toward understanding. Annu Rev Anim Biosci. (2014) 2:209–33. doi: 10.1146/annurev-animal-022513-114156
9. das Neves CG, Roth S, Rimstad E, Thiry E, Tryland M. Cervid herpesvirus 2 infection in reindeer: a review. Vet Microbiol. (2010) 143:70–80. doi: 10.1016/j.vetmic.2010.02.015
10. Cunha CW, Slater OM, Macbeth B, Duignan PJ, Warren A, Highland MA Li H. Domestic sheep and bighorn sheep carry distinct gammaherpesviruses belonging to the genus macavirus. Virus Res. (2019) 272:197729. doi: 10.1016/j.virusres.2019.197729
11. Metzler AE. The malignant catarrhal fever complex. Comp Immunol Microbiol Infect Dis. (1991) 14:107–24. doi: 10.1016/0147-9571(91)90125-W
12. Li H, Gailbreath K, Flach EJ, Taus NS, Cooley J, Keller J, et al. novel subgroup of rhadinoviruses in ruminants. J Gen Virol. (2005) 86:3021–26. doi: 10.1099/vir.0.81238-0
13. Russell GC, Stewart JP, Haig DM. Malignant catarrhal fever: a review. Vet J. (2009) 179:324–35. doi: 10.1016/j.tvjl.2007.11.007
14. Hussain I, Kashoo ZA, Wani AH, Hasin D, Pampori ZA, Wani SA. Malignant catarrhal fever: recent update. Indian J Anim Sci. (2017) 87:260–9. doi: 10.56093/ijans.v87i3.68792
15. Seeley KE, Junge RE, Jennings RN, Cunha CW Li H. Moose (Alces alces) mortality associated with caprine herpesvirus 2 (CpHV-2). J Zoo Wildl Med. (2018) 49:774–8. doi: 10.1638/2016-0207.1
16. Carvallo FR, Uzal FA, Moore JD, Jackson K, Nyaoke AC, Naples L, et al. Ibex-associated malignant catarrhal fever in duikers (Cephalophus Spp). Vet Pathol. (2020) 57:577–81. doi: 10.1177/0300985820918313
17. Fabisiak M, Sałamaszyńska A, Stadejek T. Detection of seroconversion to bovine herpesvirus 1 related alphaherpesvirus and bovine viral diarrhea virusin polish free-living deer. Pol J Vet Sci. (2018) 21:437–40.
18. Zhu H, Huang Q, Hu X, Chu W, Zhang J, Jiang L, et al. Caprine herpesvirus 2-associated malignant catarrhal fever of captive sika deer (Cervus nippon) in an intensive management system. BMC Vet Res. (2018) 14:38. doi: 10.1186/s12917-018-1365-8
19. Zhu H, Sun N, Li Y, Feng T, Jiang L, Yu X, et al. Malignant catarrhal fever: an emerging yet neglected disease in captive sika deer (Cervus nippon) herds in China. Transbound Emerg Dis. (2020) 67:149–58. doi: 10.1111/tbed.13334
20. Partin TG, Schrenzel MD, Braun J, Witte CL, Kubiski SV, Lee J, et al. Herpesvirus surveillance and discovery in zoo-housed ruminants. PLoS ONE. (2021) 16:e0246162. doi: 10.1371/journal.pone.0246162
21. Auer A, Schweitzer L, Kübber-Heiss A, Posautz A, Dimmel K, Seitz K, et al. Porcine circoviruses and herpesviruses are prevalent in an Austrian game Population. Pathogens. (2022) 11:305. doi: 10.3390/pathogens11030305
22. Hidalgo-Hermoso E, Celis S, Cabello J, Kemec I, Ortiz C, Lagos R, et al. Molecular survey of selected viruses in Pudus (Pudu puda) in Chile revealing first identification of caprine herpesvirus – 2 (CpHV-2) in South American ungulates. Vet Q. (2023) 46:1–7. doi: 10.1080/01652176.2022.2149879
23. Pavez-Fox M, Estay SA. Correspondence between the habitat of the threatened pudú (Cervidae) and the national protected-area system of Chile. BMC Ecol. (2016) 16:1. doi: 10.1186/s12898-015-0055-7
24. Silva-Rodríguez E, Pastore H, Jiménez J. Pudu puda. The IUCN Red List of Threatened Species. (2016). Available online at: https://www.iucnredlist.org/es/species/18848/22164089 (accessed September 22, 2021).
25. Biblioteca del Congreso Nacional de Chile. Supreme Decree no. 151 of the Ministerio Secretaria General de la Presidencia de Chile. [First Species Classification Process, MMA]. (2007). Available online at: https://www.bcn.cl/leychile/navegar?idNorma=259402 (accessed July 18, 2022).
26. Hidalgo-Hermoso E, Cabello J, Novoa I, Celis S, Ortiz C, Kemec I, et al. Molecular detection and characterization of hemoplasmas in the Pudu (Pudu puda), a native cervid from Chile. J Wildl Dis. (2022) 58:8–14. doi: 10.7589/JWD-D-21-00057
27. Morales N, Aldridge D, Bahamonde A, Cerda J, Araya C, Muñoz R, et al. Corynebacterium pseudotuberculosis infection in Patagonian huemul (Hippocamelus bisulcus). J Wildl Dis. (2017) 53:621–4. doi: 10.7589/2016-09-213
28. Moreno-Beas E, Abalos P, Hidalgo-Hermoso E. Seroprevalence of nine Leptospira interrogans serovars in wild carnivores, ungulates, and primates from a zoo population in a Metropolitan region of Chile. J Zoo Wildl Med. (2015) 46:774–8. doi: 10.1638/2014-0139.1
29. Salgado R, Hidalgo-Hermoso E, Pizarro-Lucero J. Detection of persistent pestivirus infection in pudú (Pudu puda) in a captive population of artiodactyls in Chile. BMC Vet Res. (2018) 14:37. doi: 10.1186/s12917-018-1363-x
30. Santodomingo A, Robbiano S, Thomas R, Parragué-Migone C, Cabello-Stom J, Vera-Otarola F, et al. search for piroplasmids and spirochetes in threatened pudu (Pudu puda) and associated ticks from Southern Chile unveils a novel Babesia sp. and a variant of Borrelia chilensis. Transbound Emerg Dis. (2022) 69:3737–48. doi: 10.1111/tbed.14743
31. Silva-Rodríguez EA, Sieving KE. Domestic dogs shape the landscape-scale distribution of a threatened forest ungulate. Biol Conserv. (2012) 150:103–10. doi: 10.1016/j.biocon.2012.03.008
32. Vila AR, Briceño C, McAloose D, Seimon TA, Armién AG, Mauldin EA, et al. Putative parapoxvirus-associated foot disease in the endangered huemul deer (Hippocamelus bisulcus) in Bernardo O'Higgins National Park, Chile. PLoS ONE. (2019) 14:e0213667. doi: 10.1371/journal.pone.0213667
33. Corti P, Arnemo JM. Partially reversible immobilization of free-ranging huemul deer (Hippocamelus bisulcus) with medetomidine-ketamine and atipamezole. J Wildl Dis. (2021) 57:927–31. doi: 10.7589/JWD-D-20-00199
34. Ward RD, Zemlak TS, Innes BH, Last PR, Hebert PD. DNA barcoding Australia's fish species. Philos Trans R Soc Lond B Biol Sci. (2005) 360:1847–57. doi: 10.1098/rstb.2005.1716
35. VanDevanter DR, Warrener P, Bennett L, Schultz ER, Coulter S, Garber RL, et al. Detection and analysis of diverse herpesviral species by consensus primer PCR. J Clin Microbiol. (1996) 34:1666–71. doi: 10.1128/jcm.34.7.1666-1671.1996
36. Ehlers B, Dural G, Yasmum N, Lembo T, de Thoisy B, Ryser-Degiorgis MP, et al. Novel mammalian herpesviruses and lineages within the gammaherpesvirinae: cospeciation and interspecies transfer. J Virol. (2008) 82:3509–16. doi: 10.1128/JVI.02646-07
37. Katoh K, Toh H. Recent developments in the MAFFT multiple sequence alignment program. Brief Bioinform. (2008) 9:286–98. doi: 10.1093/bib/bbn013
38. Miller MA, Pfeiffer W, Schwartz T. The CIPRES science gateway: a community resource for phylogenetic analyses. In: Proceedings of the 2011 TeraGrid Conference: Extreme Digital Discovery. New York, NY: ACM (2011), p. 41. doi: 10.1145/2016741.2016785
39. Rambaut A. FigTree v1. 3.1 Institute of Evolutionary Biology. Edinburgh: University of Edinburgh (2010).
40. Rothman KJ. Epidemiology: An Introduction, 2nd ed. Oxford: Oxford University Press (2012), p. 280.
41. Grinde B. Herpesviruses: latency and reactivation - viral strategies and host response. J Oral Microbiol. (2013) 5. doi: 10.3402/jom.v5i0.22766
42. Blake JE, Nielsen NO, Heuschele WP. Lymphoproliferation in captive wild ruminants affected with malignant catarrhal fever: 25 cases (1977-1985). J Am Vet Med Assoc. (1990) 196:1141–3. doi: 10.2460/javma.1990.196.07.1141
43. Crawford TB Li H, Rosenburg SR, Norhausen RW, Garner MM. Mural folliculitis and alopecia caused by infection with goat-associated malignant catarrhal fever virus in two sika deer. J Am Vet Med Assoc. (2002) 221:843–7. doi: 10.2460/javma.2002.221.843
44. Li H, Cunha CW, Abbitt B, deMaar TW, Lenz SD, Hayes JR, et al. Goats are a potential reservoir for the herpesvirus (MCFV-WTD), causing malignant catarrhal fever in deer. J Zoo Wildl Med. (2013) 44:484–6. doi: 10.1638/2012-0087R.1
45. Li H, Wunschmann A, Keller J, Hall DG, Crawford TB. Caprine herpesvirus-2- associated malignant catarrhal fever in white tailed deer (Odocoileus virginianus). J Vet Diagn. (2003) 15:46–9. doi: 10.1177/104063870301500110
46. Foyle KL, Fuller HE, Higgins RJ, Russell GC, Willoughby K, Rosie WG, et al. Malignant catarrhal fever in sika deer (Cervus nippon) in the UK. Vet Rec. (2009) 165:445–7. doi: 10.1136/vr.165.15.445
47. Keel MK, Patterson JG, Noon TH, Bradley GA, Collins JK. Caprine herpesvirus-2 in association with naturally occurring malignant catarrhal fever in captive sika deer (Cervus nippon). J Vet Diagn. (2003) 15:179–83. doi: 10.1177/104063870301500215
48. Frontoso R, Autorino GL, Friedrich KG Li H, Eleni C, Cocumelli C, Di Cerbo P, et al. An acute multispecies episode of sheep-associated malignant catarrhal fever in captive wild animals in an Italian zoo. Transbound Emerg Dis. (2016) 63:621–7. doi: 10.1111/tbed.12321
49. Vikøren T, Li H, Lillehaug A, Jonassen CM, Böckerman I. Malignant catarrhal fever in free-ranging cervids associated with OvHV-2 and CpHV-2 DNA. J Wildl Dis. (2006) 42:797–807. doi: 10.7589/0090-3558-42.4.797
50. Schultheiss PC, Van Campen H, Spraker TR, Bishop C, Wolfe L. Malignant catarrhal fever associated with ovine herpesvirus-2 in free-ranging mule deer in Colorado. J Wildl Dis. (2007) 43:533–7. doi: 10.7589/0090-3558-43.3.533
51. Neimanis AS, Hill JE, Jardine CM. Sheep-associated malignant catarrhal fever in free-ranging moose (Alces alces) in Saskatchewan, Canada. J Wildl Dis. (2009) 45:213–7. doi: 10.7589/0090-3558-45.1.213
52. Headley SA, Dall Agnol AM, Bessegato JA, Frucchi APS, Maturana ÉFL, Rodrigues RV, et al. Association of ovine gammaherpesvirus 2 with an outbreak of acute respiratory disease in dairy cattle. Sci Rep. (2023) 13:5623. doi: 10.1038/s41598-023-30133-w
53. Oliveira TES, Scuisato GS, Pelaquim IF, Cunha CW, Cunha LS, Flores EF, et al. The participation of a malignant catarrhal fever virus and Mycoplasma bovis in the development of single and mixed infections in beef and dairy cattle with bovine respiratory disease. Front Vet Sci. (2021) 8:691448. doi: 10.3389/fvets.2021.691448
54. Chang AM, Chen CC, Chang CD, Huang YL, Ke GM, Walther BA. Encephalitis induced by a newly discovered ruminant rhadinovirus in a free-living Formosan sambar deer (Rusa unicolor swinhoei). J Vet Med Sci. (2018) 80:810–3. doi: 10.1292/jvms.17-0477
55. Patel KK, Stanislawek WL, Burrows E, Heuer C, Asher GW, Wilson PR, et al. Investigation of association between bovine viral diarrhoea virus and cervid herpesvirus type-1, and abortion in New Zealand farmed deer. Vet Microbiol. (2019) 228:1–6. doi: 10.1016/j.vetmic.2018.11.001
56. McKillen J, Hogg K, Lagan P, Ball C, Doherty S, Reid N, et al. Detection of a novel gammaherpesvirus (genus Rhadinovirus) in wild muntjac deer in Northern Ireland. Arch Virol. (2017) 162:1737–40. doi: 10.1007/s00705-017-3254-z
Keywords: gammaherpesvirus, malignant catarrhal fever, endangered species, pudu, huemul
Citation: Hidalgo-Hermoso E, Cabello J, Lopez R, Vergara-Wilson V, Vera F, Valencia C, Sanchez C, Celis S, Silva A, Cariñanos A, Barria I, Vicencio R, Muñoz-Leal S, Aravena P, Lagos R, Toro-Letelier J, Verasay-Caviedes S, Garnham A, Peña I, Sánchez F, Moreira-Arce D, Vergara PM, Alegria-Moran R and Cortés-Hinojosa G (2024) Molecular and phylogenetic analysis of herpesviruses in endangered free-ranging cervids of Chile: ovine gammaherpesvirus-2—A novel threat to wild and domestic animal health in Chilean Patagonia. Front. Vet. Sci. 10:1321172. doi: 10.3389/fvets.2023.1321172
Received: 13 October 2023; Accepted: 29 December 2023;
Published: 01 February 2024.
Edited by:
Magdalena Larska, National Veterinary Research Institute (NVRI), PolandReviewed by:
Wojciech Socha, National Veterinary Research Institute (NVRI), PolandCristina Cunha, United States Department of Agriculture (USDA), United States
Copyright © 2024 Hidalgo-Hermoso, Cabello, Lopez, Vergara-Wilson, Vera, Valencia, Sanchez, Celis, Silva, Cariñanos, Barria, Vicencio, Muñoz-Leal, Aravena, Lagos, Toro-Letelier, Verasay-Caviedes, Garnham, Peña, Sánchez, Moreira-Arce, Vergara, Alegria-Moran and Cortés-Hinojosa. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ezequiel Hidalgo-Hermoso, ZXplcXVpZWxoaWRhbGdvdmV0QHlhaG9vLmNvbQ==; Galaxia Cortés-Hinojosa, Z2FsYXhpYS5jb3J0ZXNAdWMuY2w=