 Carlos Iglesias Pastrana1
Carlos Iglesias Pastrana1 Francisco Javier Navas González1*
Francisco Javier Navas González1* Elena Ciani2
Elena Ciani2 Carmen Marín Navas1
Carmen Marín Navas1 Juan Vicente Delgado Bermejo1
Juan Vicente Delgado Bermejo1- 1Department of Genetics, Faculty of Veterinary Sciences, University of Cordoba, Cordoba, Spain
- 2Department of Biosciences, Biotechnologies and Environment, University of Bari ‘Aldo Moro’, Bari, Italy
Despite the relatively wide knowledge of camel biomechanics, research into the immediate functional response that accompanies the execution of physical exercise remains unapproached. Therefore, selective breeding programs lack an empirical basis to achieve genetic improvement of physical stress tolerance traits and monitor camel welfare in this regard. Given the fact that physical exercise increases net heat production, infrared thermography (IRT) was selected to study the temperature changes at the skin surface of the different body areas in clinically normal dromedary camels, mostly relegated to leisure activities. Specifically, a lower dispersion at the individual level of the surface temperature at the scapular cartilage region, shoulder joint, and pelvis region, as well as lower values for Tmax and Tmin at the region of the ocular region, pectoral muscles, semimembranosus-semitendinosus muscles, and hind fetlock after exercise, have to be considered as breeding criteria for candidate selection. Such thermophysiological responses can be used as indirect measures of tissue activity in response to exercise and hence are reliable indicators of animal tolerance to physical exercise-induced stress. Additionally, sex, castration, age, and iris pigmentation significantly impacted the thermo-physiological response to exercise in the study sample, which can be attributed to hormones, general vigor, and visual acuity-mediated effects. These specific factors’ influence has to be considered for the evaluation of physical performance and the design of selection schemes for physical-related traits in dromedaries.
1 Introduction
The socioeconomic interest in camel breeding fundamentally resides in the production of food (milk and meat) and other products and by-products such as wool, dung, skin, and fat. Abundant literature does exist on the characterization of camel-based food products and subproducts, with special reference to the different animal-and management-related variables that potentially influence yield and thus economic benefits in these camel farming systems (1–5). However, several camel breeds are typically selected for their power and speed within a functional scenario of leisure, load-carrying, and traction work (6). Comparatively, little information is available for these specific tasks.
Elementary analyses of the biomechanics of gait (7–9), the elastic extension of tendons (10), the morphology of some parts of the distal skeleton (7, 11–13), and the pedal anatomy (14) in one-humped camels shed light on the normal movements, osteomuscular composition, and locomotor adaptations of these animals to sandy terrains in desert ecosystems. More specifically, the repercussions that different exercise conditions have at a biochemical and hormonal level are relatively widely studied (15–17), highlighting the near-Newtonian behavior of camel blood as a response to endurance activities (18). In regards to animal-dependent factors, Al-Shorepy (19) concludes that age and sex have a significant effect on racing performance.
Notwithstanding this conglomerate of empirical knowledge on camel biomechanics, research into the basic physiology and function of the musculature in the working camel still remains unapproached. The knowledge of the immediate functional responses and organic adaptations that accompany the execution of a session of physical activity or exercise allows the accurate, early diagnosis of decreased levels of sports performance and the design of training/rehabilitation programs (20, 21). Since physical exercise has a remarkable direct effect on net heat production (22), infrared thermography (IRT) is the technique of choice to study the underlying physiological response of muscles and other organic tissues during and after exercise (23). This non-invasive imaging method is capable of detecting temperature changes (thermogenesis) at the skin surface that are an indirect measure of tissue activity (local blood flow and metabolism rate) in response to exercise (24, 25).
In this study, using thermography, we examine 22 body regions to determine their significance in monitoring the effects of exercise in clinically normal dromedary camels. Additionally, we will analyze the varying impact of different factors specific to the animals on the thermo-physiological responses observed in these body regions. The results obtained will serve to evaluate the potential of infrared thermography to be used as a tool for reliable assessment of camel physical fitness and, thus, welfare impacts when these animals participate in physically demanding activities. The animal model used is the Canarian camel (Camelus dromedarius), an autochthonous endangered breed mainly relegated to leisure activities in which the physical health and performance of the animals have large significance (26).
2 Materials and methods
2.1 Animal sample and data selection
A total of 130 Canarian dromedaries (72 males and 58 females, aged between 18 months and 35 years old) were included in this study. All dromedary camels had resided at the respective participating farm for at least 1 year prior to the study. Animals were checked before the evaluations to ensure they were in good health.
2.2 Study duration
The study was carried out over 5 consecutive days in January 2021. For the study of the thermo-physiological response to exercise in leisure dromedaries, each dromedary camel participating in this study was exercised for 15 min by the same familiar handler. The exercise involved walking the animal for 15 min at a comfortable, active walking speed for each individual.
2.3 Thermal data collection
Thermal images for this study were captured using an Hti-Xintai HT-18 thermal imaging camera (220 × 160 IR resolution, 35,200 pixels). The camera has a thermal sensitivity of <0.05°C at temperatures of ≥30°C, a temperature detection range between −20°C and + 120°C, and an emissivity of 0.95.
To minimize the impact of environmental factors, thermographic evaluations were consistently conducted within an enclosed pen by the same technicians. Static thermal images were taken at a 90° angle from a distance of 1 m ± 50 cm from the animals, following recommendations from Yarnell et al. (23).
Thermal images were captured in 22 body regions. Thermal images were taken at three separate sampling time points: pre-exercise, post-exercise 0 min, and post-exercise 5 min.
2.4 Evaluated areas and thermal quantitative variables
The software used for thermographic assessment was Batch Flir Image Converter.1 Quantitative variables extracted from each thermal image included average temperature, maximum temperature, minimum temperature, and standard deviation, all of which were measured in degrees Celsius.
The evaluated areas from which the aforementioned quantitative variables were sampled comprised the ocular region (Regio orbitalis), scapular cartilage region (Regio cartilaginis scapulae), shoulder joint (Articulatio humeri), thorax (Regio pectoris), antebrachial region (Regio antebrachii), carpus region (Regio carpi), metacarpal region (Regio metacarpi), region of the proximal phalanx (Regio phalangis proximalis) of the thoracic limb, digital flexor tendons of the thoracic limb, coronary region of the thoracic limb (Regio coronalis), lateral and medial hoof cartilage of the thoracic limb (Cartilago ungularis medialis et lateralis), palmary bulb of the heel (Torus ungulae), lumbar region (Regio lumbalis), pelvis region (Regio pelvis), semimembranosus and semitendinosus muscles, tarsus region (Regio tarsi), metatarsus region (Regio metatarsi), region of the proximal phalanx (Regio phalangis proximalis) of the hind limb, digital flexor tendons of the hind limb, coronary region of the hind limb (Regio coronalis), the lateral and medial hoof cartilage of the hind limb (Cartilago ungularis medialis et lateralis), and plantary bulb of the heel (Torus ungulae). The “Nomina anatomica veterinaria” (Sixth Edition) (27) was used as a guideline to select the pertinent anatomical nomenclature used for the reference of the body regions considered in the present study.
The hump region was not evaluated due to its primarily fatty tissue composition, where fat thermogenesis primarily depends on the types of fatty acids present, which, in turn, are influenced by the animal’s diet and other factors related to the animal’s overall health status, factors that were not controlled in this study. Additionally, the femoro-patellar joint or stifle joint (Articulatio genus) was not assessed due to the presence of a notable hard callus (active osteoblasts that mineralize the callus matrix) with significant dimensions (area and thickness) in this region, which functions to protect this area when the camel sits on hot surfaces such as sand.
For the forelimb or thoracic limb (Membrum thoracicum) and hindlimb or pelvic limb (Membrum pelvinum), the same image was used to extract the average and standard deviation temperatures for each individual region evaluated within each limb. This approach provided a more precise evaluation of the workload and physiological stress relative to each individual region compared to the overall limb (forelimb or hindlimb).
2.5 Statistical analysis
In line with the methodologies outlined in González Ariza et al. (28), we initiated our evaluation by employing discriminant canonical analysis. This allowed us to create a tool that assesses optimal linear combinations of thermo-physiological response-related variables and animal-dependent factors able to determine within and between population clustering patterns across different moments of exercise [pre and post (0 and 5 min)].
Our independent variables comprised average temperature, maximum temperature, minimum temperature, and standard deviation per evaluated region. Additional variables such as sex, age (months), coat color (white, blonde, cinnamon, chestnut, bay, black, ashed, and roan), coat particularities (solid color, white-haired areas in extremities, white-haired areas in head and neck, white-haired areas in the thorax, white-haired areas in extremities, head and neck, white-haired areas in extremities and thorax, white-haired areas in head, neck and thorax, and white-haired areas across almost the body), eye color (brownish, brownish with blue spots, and bluish), neutering status (neutered and non-neutered), and active involvement (involved and not involved) in desensitization protocols were included.
Calculation of centroids for different moments of exercise [pre and post (0 and 5 min)] was conducted. The relative positions of each centroid were determined by substituting the mean values for the observations represented in each of the two detected discriminant functions (F1 and F2).
Subsequently, we computed squared Mahalanobis distances to measure dissimilarities between pre-exercise moments and post-exercise (0 and 5 min) moments. These squared Mahalanobis distances were determined using the following formula:
where is the distance between population i and j; and are the means of the variable x in the ith and jth populations, respectively; and COV−1 is the inverse of the covariance matrix of measured variable x. Finally, we used the squared Mahalanobis distance to visually illustrate the clustering patterns arising from variations in thermal information among the different exercise times [pre and post (0 and 5 min)] considered in this study. A dendrogram was then created using the underweighted pair-group method arithmetic averages (UPGMA) from Universität Rovira i Virgili (URV), Tarragona, Spain, and the phylogeny procedure of MEGA X 10.0.5 (Institute of Molecular Evolutionary Genetics, Pennsylvania State University, State College, PA, United States).
3 Results
A summary of the mean values for thermal information reported by all the variables considered across the body region sampled is shown in Figure 1.
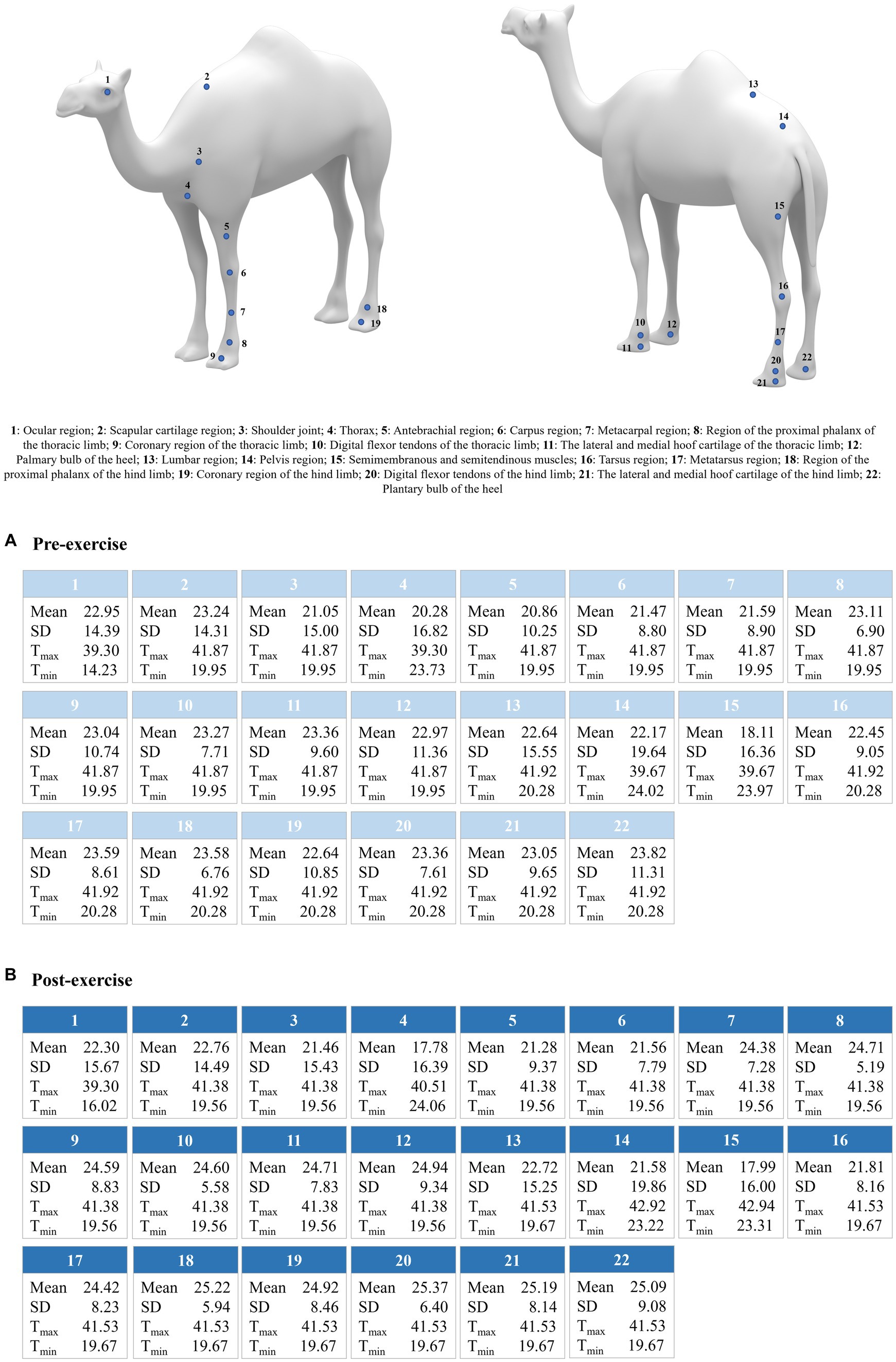
Figure 1. Average values for mean, standard deviation (SD), maximum (Tmax), and minimum (Tmin) temperature per body region and evaluation state (pre- and post-exercise 0 min). Data for post-exercise 5 min were not presented due to the lack of differences with pre-exercising, which suggests total thermal recovery of the animals after this time has passed.
Concerning the reliability of the discriminant canonical analysis model, after conducting 49 rounds of multicollinearity analyses, the variables included in the discriminant canonical analysis are shown in Table 1.
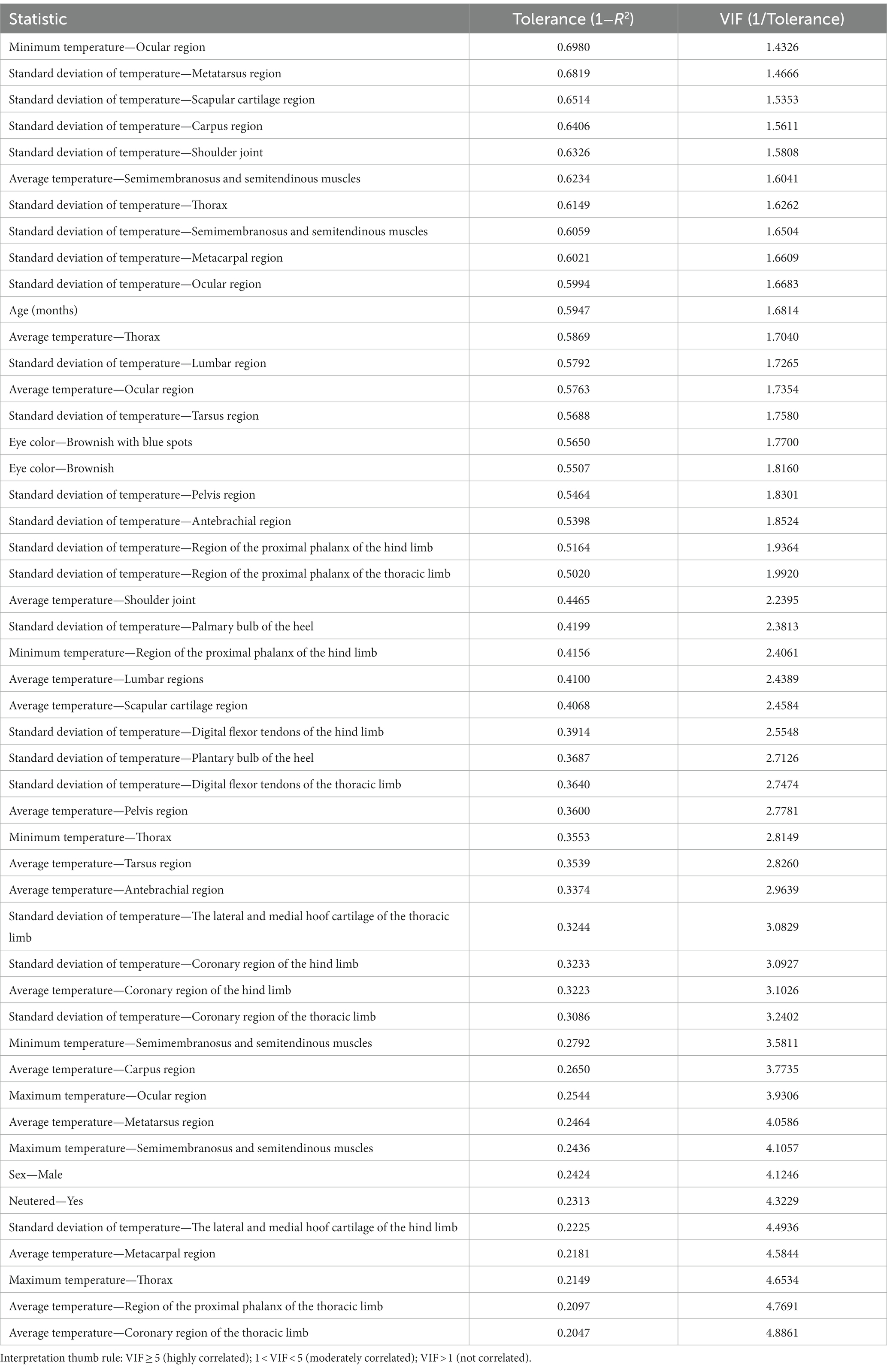
Table 1. Summary of the value of tolerance and VIF after multicollinearity analysis of thermal information-related variables in the Canarian camel breed.
Pillai’s trace criterion reported a significant difference between pre- and post-exercise moments (Pillai’s trace criterion: 0.4434, F (observed value): 1.9764, F (critical value): 1.2695, df1: 98, df2: 680, p-value<0.0001), confirming the validity of the discriminant canonical analysis.
Out of the two functions identified through discriminant analysis, one was found to be significant for their discriminant ability (Table 2). Among these, the F1 function exhibited the highest discriminatory power, with an eigenvalue of 0.5374, explaining 83.84% of the variance.
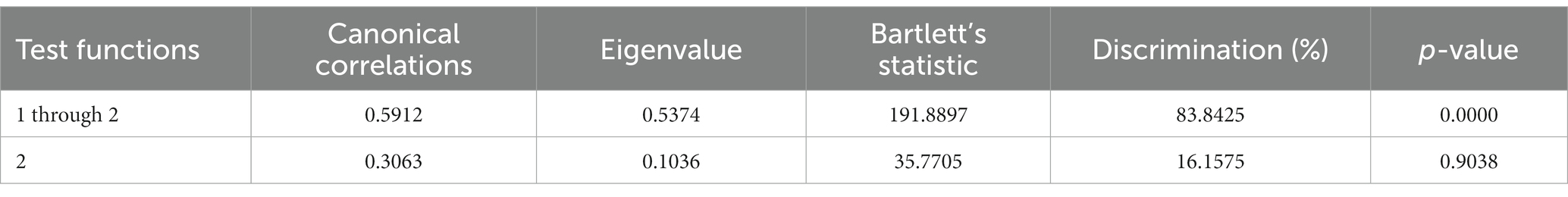
Table 2. Canonical discriminant analysis efficiency parameters to determine the significance of each canonical discriminant function.
The various variables examined in this study were ranked based on their discriminative capacity. An evaluation of the equality of group means of the dependent variables involved in the discriminant canonical analysis is presented in Table 3.
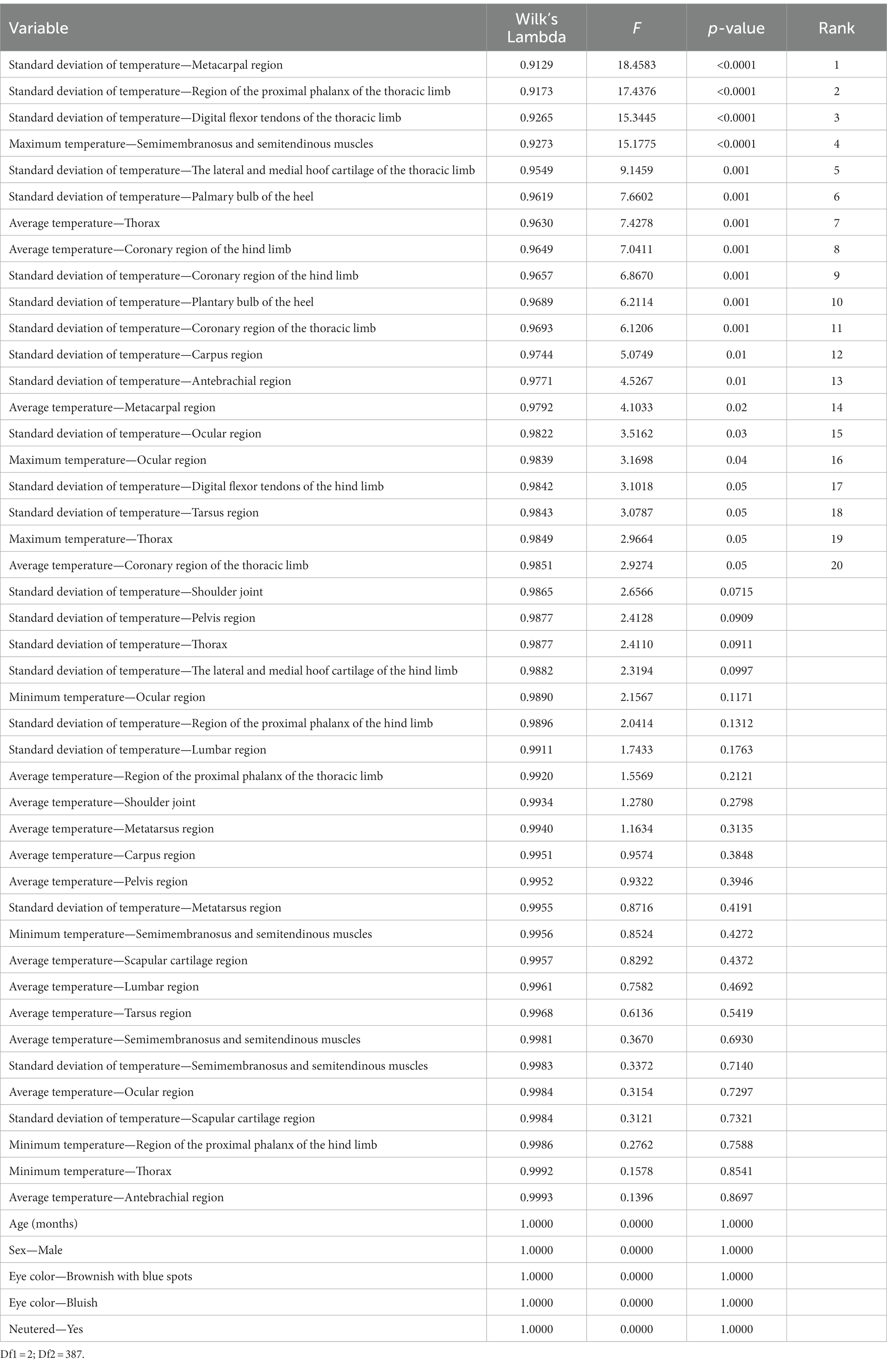
Table 3. Results for the tests of equality of group means to test for differences in the means across sample groups once redundant variables were removed.
Higher values of F and lower values of Wilks’ lambda indicate greater discriminating power. The analysis revealed that standard deviation of temperature in the metacarpal region, standard deviation of temperature in the region of the proximal phalanx of the thoracic limb, standard deviation of temperature in the digital flexor tendons of the thoracic limb, maximum temperature in the semimembranosus and semitendinous muscles, standard deviation of temperature in the lateral and medial hoof cartilage of the thoracic limb, standard deviation of temperature in the palmary bulb of the heel, average temperature in the thorax, average temperature in the coronary region of the hind limb, standard deviation of temperature in the coronary region of the hind limb, standard deviation of temperature in the plantary bulb of the heel, standard deviation of temperature in the coronary region of the thoracic limb, standard deviation of temperature in the carpus region, standard deviation of temperature in the antebrachial region, average temperature in the metacarpal region, standard deviation of temperature in the ocular region, maximum temperature in the ocular region, standard deviation of temperature in the digital flexor tendons of the hind limb, standard deviation of temperature in the tarsus region, maximum temperature in the thorax, and average temperature in the coronary region of the thoracic limb made highly significant contributions (p < 0.0001) to the discriminant functions when qualitative gait evaluation levels were the clustering criteria.
Standardized discriminant coefficients were used to assess the relative weight of each dependent variable across the two established discriminant functions (Supplementary Table S1).
A Press’ Q-value of 51.80 (n = 390; n’ = 197; K = 3) was computed for thermal information across exercise moment levels [pre and post (0 and 5 min)], indicating that predictions can be considered better than chance at a 95% confidence level (29).
The results for the functions at the centroids are presented in Table 4.
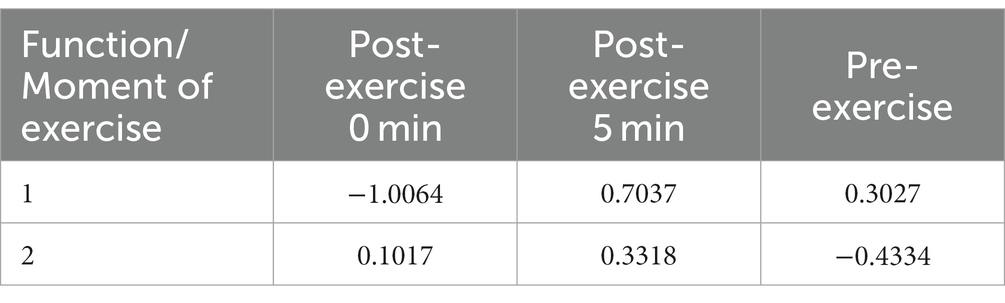
Table 4. Functions at the centroids for the two discriminant functions detected in this study.
Concerning the dendrogram constructed using the squared Mahalanobis distance (Figure 2), the absence of differences between pre- and post-exercise thermal information 5 min after exercise may be explained by the fact that camels are energy-efficient animals, hence it takes them less than 5 min to completely recover at a thermo-physiological level (14, 17, 30).
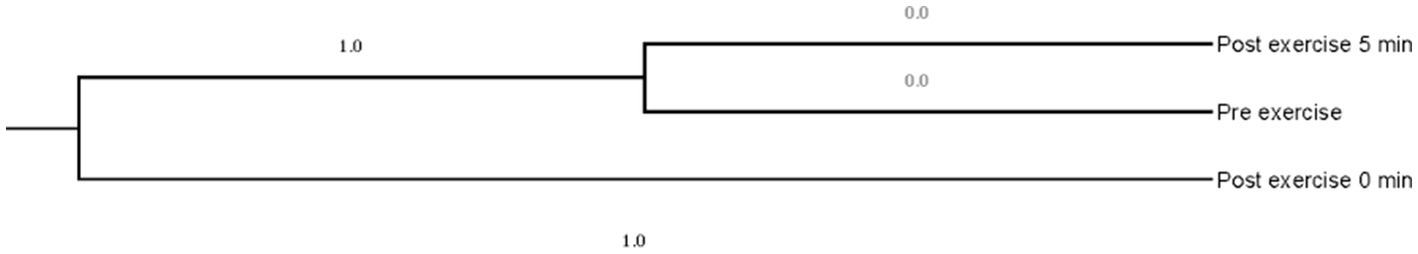
Figure 2. Dendrogram constructed from Mahalanobis distances across exercise moments [pre and post (0 and 5 min)].
4 Discussion
The thermo-physiological response to physical exercise was evaluated through infrared thermography at 22 body regions of clinically normal dromedary camels. The mean temperature was generally higher after exercise in almost all the evaluated body regions, while the opposite trend was observed for the values of standard deviation, maximum temperature (Tmax), and minimum temperature (Tmin). As Tmax and Tmin do not decrease by approximately the same amount between evaluated time periods, the standard deviation is expected to decrease and the mean to move up, and vice versa (31). From a pure physiological viewpoint, the mean temperature of the body regions can be anticipated to be increased after physical exercise due to the near-immediate local rise in blood flow, nutrient supply, and metabolic rate, but also the temperature to be more or less homogeneous along each local body area depending on their specific functional repercussion in locomotion. In addition, considering the natural selection for energy efficiency of movement and optimization in extreme environments in camelids (32), a low-temperature dispersion should appear.
At the individual level, a more irregular regional temperature dispersion could be indicative of local functional alteration (e.g., microcirculation failures or inflammation) that might compromise animal physical health and thus performance (33). On the other hand, a higher regional temperature dispersion at the group level might be indicative of substantial interindividual variability for the thermo-physiological local response, which could be attributed to the effects of different animal-and environment-related factors and therefore plausible to be used as a reliable measure of individual tolerance to physical exercise-induced stress. This could also be reflecting the higher latency of thermo-physiological response signals to exercise at these body areas, which then indicates that thermographic evaluation of these areas should be prioritized to accurately examine tolerance to physical exercise.
This last particular thermo-physiological behavior is indeed patent for some of the examined body regions. For the ocular region, scapular cartilage region, pectoral muscles, pelvis region, and semimembranosus-semitendinosus muscles, the mean temperature is lower and the mean standard deviation and/or Tmax and Tmin are higher after exercise. Only at the shoulder joint area are the mean temperature and the mean standard deviation higher, and/or Tmax and Tmin are lower, after exercise. First, during exercise, as a potential stress-inducing activity, eyelid contractions and sensory inputs from the visual, vestibular, and somatosensory systems for the visual-motor coordination to maintain postural balance are enhanced (34, 35), which might be manifested in an increase in local temperature. Such visual-motor coordination, and thus the level of stress with which the animal has to deal, is in turn governed by different influencing factors related to the anatomy and general biomechanics of each individual. Moreover, such idiosyncrasy will be reflected in the variability of the thermo-physiological response at the scapular cartilage region, shoulder joint, and pelvis region, which are largely implicated in the attenuation of impact forces from the feet to the head and back during quadrupedal locomotion (36).
In regards to the generally higher mean maximum and minimum temperatures at the ocular region, pectoral muscles, pelvis region, and semimembranosus-semitendinosus muscles after exercise, the mean maximum temperature of the ocular region does not significantly vary between the resting state and after exercise, which could be attributed to the existence of a local critical temperature due to the greater risk of heat damage at the tissues of the eye and adjacent brain structures for their elevated sensitivity to the effects of heat (37). Furthermore, a higher mean Tmax and/or Tmin at the pectoral muscles, pelvis region, and semimembranosus-semitendinosus muscles after exercise could be explained by the potential role of such areas as external projection windows for cardiac work and blood flow at the level of the brachial plexus (38) and for the work of the caudal region of the musculature at hindlimb in the flexion-extension of both the hip or coxofemoral and stifle joints and their respective propulsive forces (23), respectively.
However, for the specific case of the tarsal region, the mean temperature, standard deviation, Tmax, and Tmin are lower in the post-exercise state. Such a finding reveals the importance of the early physiological recovery of this area to maintain good mechanical performance (39), hence it can be used as a reliable indicator of physical health in these animal species. In the animal sample studied, as they are functionally relegated to riding activities, the mechanics of the hindlimbs are crucial for good performance and maintenance of good health status. This peculiar thermal behavior could also suggest the potential existence of an extraordinary blood vessel circulation involved in heat dissipation at this local body region, and that should be specifically approached in future applied studies.
In summary, the quantitative trends discussed, together with the discriminant potential between the resting and post-exercise states of the standard deviation values for surface temperature in practically all the regions evaluated, and to a lesser extent the mean temperature values and the mean edge values of the range, serve to construct a non-invasive protocol for the monitoring of the impact of physical exercise and welfare status in dromedary camels subjected to physical exercise. Such a statement is additionally reinforced with the percentages of correct classification of individuals based on discriminant analysis, from which a significant number of camels in resting state are classified by the discriminant analysis as if they would have performed physical exercise. Following the same logic, those camels that are classified as in a resting state after having performed exercise would be the individuals to be selected for breeding purposes since they are able to physiologically recover quickly after physical effort.
Specifically, the greater the variability in surface temperature values at the discriminating regions after exercise, the induced stress will presumably be greater, or, if it is the same, the individual tolerance to exercise and physical fitness will be lower. At a pragmatic level, such variability would be observed in the thermographic camera as a relatively heterogeneous map of colors and tones in the pertinent region(s). The Tmax and the Tmin at the region of the ocular region, pectoral muscles, semimembranosus-semitendinosus muscles, and the region of the proximal phalanx of the hind limb also have discriminant potential between the resting and post-exercise states’ surface temperatures. For the specific case of the ocular region, additional evidence is thus provided to the existing literature for other animal species, which reports that eye temperature is one of the most accurate, non-invasive tools to study animal welfare (40). For the thorax region, its discriminating potential reinforces the abovementioned significance of its role as a window of external projection for cardiac work and brachial plexus blood flow. Concerning the semimembranosus-semitendinosus muscles and the region of the proximal phalanx of the hind limb, these structures have a prominent implication in the coordination of movement and effectiveness of locomotion, mainly due to the control that they exert at the propulsive forces and their support of body weight in the animal model used (riding camels) (41). Although the Tmin at the region of the proximal phalanx of the hind limb is the same as the other structures evaluated at the rear limbs for methodological reasons, its discriminating potential derives from the statistical relationships that exist between the thermo-physiological behavior at this body area and the pertinent for the remaining regions evaluated at the population level.
Then, the detection at these discriminating areas of thermo-physiological responses contrary to the indicated, specific trend or exceptional extreme values could be interpreted as the incidence of a functional alteration, probably subclinical, with local (e.g., osteomuscular alteration in hindlimbs) or systemic (e.g., cardiac pathology) repercussions. Notwithstanding, the determination of the possible deviation from the thermo-physiological behavior of a single individual and their preference to be included in the breeding schemes should be carefully implemented with consideration of the effects of other qualitative and quantitative factors such as sex, age, neutering status, and eye color. Males, neutered and young (sexually immature) animals, and camels with brownish and relatively spotted blue irises display greater variability in the thermo-physiological response to physical exercise. In contrast, ambient temperature, breed, and training level, but neither gender nor age, have significant effects on skin temperature in other sport animal species such as racehorses (42).
The impact of sex and neutering status on individual physiological recovery capacity after exercise in camels would be basically explained by hormonal mechanisms (e.g., androgen-mediated effects) (43) and the higher body corpulence after castration (44), as well as the differences between sexes for cell composition and skin structure in dromedaries (45). In relation to age, the lower general vigor and functional maturity of the thermoregulatory system in the early stages of development would determine a relative greater variability in the individual thermoregulatory abilities (46). Finally, the higher the heterogeneity in iris pigmentation, the lower the visual acuity may be (47), which may cause further stress when the visual effort to maintain proprioception during a physical exercise needs to be consistent.
Contrastingly, coat color particularities and training regimes did not significantly affect the body surface temperature in the study sample. The camel hair medulla, independent of their color and the possible influence of color on light reflection capacity, provides animals with a notable thermoregulatory capacity (48). These results are in accordance with those reported by Abdoun et al. (49), who found a non-significant effect of coat color on thermo-physiological responses and heat tolerance in Saudi dromedaries. Such an outcome would also explain why there are camels of very different colors, even dark ones, in desert-like environments. Finally, the non-significant influence of training regimens on thermo-physiological responses in camels could be derived from the above-cited physiological adaptations of these animals to energy optimization in arid and semi-arid environments.
Seeking routine, on-field application of the results derived from this research study, the thermographic ranges presented and discussed constitute a valuable tool for enhancing monitoring protocols to assess dromedary camel welfare before and after physically demanding activities. Camel breeders are provided with guidelines to refine their management practices and increase the list of selection criteria for genetic improvement and welfare-oriented purposes. Additionally, veterinarians will be able to detect, at early stages, subtle health alterations that are potentially limiting the physical performance of the leisure dromedary camels. Ultimately, incorporating thermographic technology into leisure-oriented activities not only promotes the ethical treatment of dromedary camels but also lays the foundation for a robust protocol that benefits both the animals and those responsible for their care. Furthermore, future applied thermography studies should focus on overcoming some current limitations, such as the measurement of rectal temperature and the analysis of biochemistry data, so that correlations between thermographic and physiological parameters can be established. By including age as a covariate in this information set, we could also estimate the life span of sports dromedaries. With this aim, when animals of different ages show appreciably higher surface temperatures and levels of stress-related biomarkers during the same physical activity, it can be deduced that their ability to tolerate physical exercise is lower. As a result, there is a greater likelihood of adverse effects on their overall wellbeing.
5 Conclusion
The lower the regional dispersion at the thermo-physiological response after physical exercise, the more likely it is to discard potential local functional alterations with a possible systemic impact. Moreover, greater individual tolerance to physical exercise-induced stress and, thus, a lesser negative impact on animal welfare can be assumed. Specifically, a lower dispersion of the surface temperature at the scapular cartilage region, shoulder joint, and pelvis region should be preferably used to select dromedary camels for traits related to tolerance to physical exercise-related stress, given that larger overall data variability was detected at these body regions within the study population. In addition, the edge values of the temperature range at the local level for the ocular region, thorax, semimembranosus-semitendinosus muscles, and region of the proximal phalanx of the hind limb have a discriminating effect in this regard, and camels with lower values for Tmax and Tmin at these areas after exercise have to be prioritized for selection with genetic improvement purposes. Sex, castration, age, and iris pigmentation also significantly affect the thermo-physiological response to exercise in dromedaries, which can be related to variations in stress levels because of hormones, general vigor, and visual acuity-mediated effects. The discussed thermographic ranges in this research study serve as a reliable, non-invasive tool for improving monitoring protocols for dromedary camel welfare, especially before and after physically demanding activities. The results offer guidelines for camel breeders to enhance their management practices and enable veterinarians to detect early signs of health alterations. Future applied studies are also encouraged to correlate thermographic measurements with other physiological data as well as estimate the life span of sports dromedaries by including age as a covariate in the combined monitoring (non-invasive and invasive techniques) of tolerance to physical exercise.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
Ethics statement
All farms included in our study adhered to specific codes of good practices, ensuring humane care for the animals, in accordance with the national guidelines for the care and use of laboratory and farm animals in research. Written consent was obtained from the owners for their participation in the study. Our research was conducted in compliance with the Declaration of Helsinki. The Spanish Ministry of Economy and Competitiveness, as well as the Ethics Committee of Animal Experimentation at the University of Córdoba, authorized the application of the protocols used in this study. This authorization was granted in accordance with the 5th section of the 2nd article of the Royal Decree Law 53/2013, as the animals assessed were intended for accredited zootechnical purposes. This national decree adheres to the European Union Directive 2010/63/EU, issued on September 22, 2010.
Author contributions
CP: Data curation, Formal analysis, Investigation, Software, Visualization, Writing – original draft, Writing – review & editing. FN: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. EC: Conceptualization, Funding acquisition, Project administration, Resources, Supervision, Validation, Visualization, Writing – review & editing. CM: Data curation, Formal analysis, Investigation, Methodology, Software, Writing – review & editing. JD: Funding acquisition, Project administration, Resources, Supervision, Validation, Visualization, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. The present research was carried out in the financing framework of the international project CA.RA.VA.N – “Toward a Camel Transnational Value Chain” (Reference APCIN-2016-00011-00-00) and during the covering period of a predoctoral contract (FPU Fellowship) funded by the Spanish Ministry of Science and Innovation and a Ramón y Cajal Post-Doctoral Contract with the reference MCIN/AEI/10.13039/501100011033 and the European Union “NextGenerationEU”/PRTR.
Acknowledgments
The authors would also like to thank “Aires Africanos” Eco-tourism Company, Oasis Park Fuerteventura, “Camelus” Camellos de Almería, and Clara Isabel Sotomayor Dorado for their direct technical help and assistance.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer CB declared a shared affiliation with the author(s) CP, FN, CM, and JD to the handling editor at the time of review.
The reviewer AG declared a shared affiliation with the author(s) CP, FN, CM, and JD to the handling editor at the time of review.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fvets.2023.1297412/full#supplementary-material
Footnotes
References
1. Abdel-Rahman, EH, Alzahrani, FS, and Sulieman, AMEH. Potential of camel dung as promising organic manure in Saudi Arabia. Adv Life Sci. (2020) 7:202–7.
2. Al-Hassan, A. Gelatin from camel skins: extraction and characterizations. Food Hydrocoll. (2020) 101:105457. doi: 10.1016/j.foodhyd.2019.105457
3. Al Kanhal, HA. Compositional, technological and nutritional aspects of dromedary camel milk. Int Dairy J. (2010) 20:811–21. doi: 10.1016/j.idairyj.2010.04.003
4. Kadim, IT, Mahgoub, O, Faye, B, and Farouk, MM. Camel meat and meat products. CABI: CABI Digital Library, (2013).
5. Li, HJ, and Guo, KK. Research on structure and properties of camel wool. Adv Mat Res. (2013) 750:2313–6. doi: 10.4028/www.scientific.net/AMR.750-752.2313
6. Burger, PA, Ciani, E, and Faye, B. Old World camels in a modern world–a balancing act between conservation and genetic improvement. Anim Genet. (2019) 50:598–612. doi: 10.1111/age.12858
7. Abdo, M, Haddad, SS, Aziz, EK, Abdeen, A, and Sabek, A. Kinematics biomechanical analysis and three dimensional reconstruction diagnostic technique of carpal joint during gait in one-humped camel (Camelus dromedarius). Alex J Vet Sci. (2019) 60:23. doi: 10.5455/ajvs.30333
8. Dagg, AI. The locomotion of the camel (Camelus dromedarius). J Zool. (1974) 174:67–78. doi: 10.1111/j.1469-7998.1974.tb03144.x
9. Pfau, T, Parsons, K, and Wilson, A. Mechanics of over-ground locomotion in the dromedary camel (Camelus dromedarius). J Biomech. (2006):39:S359. doi: 10.1016/S0021-9290(06)84436-2
10. Alexander, RM, Maloiy, G, Ker, R, Jayes, A, and Warui, C. The role of tendon elasticity in the locomotion of the camel (Camelus dromedarius). J Zool. (1982) 198:293–313. doi: 10.1111/j.1469-7998.1982.tb02077.x
11. Alsafy, MA, El-Gendy, SA, and Kamal, B. Computed tomographic and radiographic morphology of the pastern and coffin joints of one-humped camel (Camelus dromedarius). Anat Histol Embryol. (2021) 50:108–13. doi: 10.1111/ahe.12607
12. Badawy, AM. Computed tomographic anatomy of the fore foot in one-humped camel (Camelus dromedrus). Glob Vet. (2011) 6:417–23.
13. El-Shafey, A, and Kassab, A. Computed tomography and cross-sectional anatomy of the metatarsus and digits of the one-humped camel (C amelus dromedarius) and Buffalo (B os bubalis). Anat Histol Embryol. (2013) 42:130–7. doi: 10.1111/j.1439-0264.2012.01174.x
14. Janis, CM, Theodor, JM, and Boisvert, B. Locomotor evolution in camels revisited: a quantitative analysis of pedal anatomy and the acquisition of the pacing gait. J Vertebr Paleontol. (2002) 22:110–21. doi: 10.1671/0272-4634(2002)022[0110:LEICRA]2.0.CO;2
15. El-Deeb, W., and Abdelghani, M. A. (2022). Investigation of lameness in racing dromedary camels (Camelus Dromedarius) and associated oxidative stress biomarkers. J Camel Prac Res. 29:110–21.
16. Kumar, S, Lamo, D, Gahlawat, G, Bharti, VK, and Kumar, K. Effect of endurance load exercise on physio-biochemical and hormonal parameters of single-humped camels (Camelus dromedarius) at high altitude. Indian J Anim Sci. (2022) 92:837–42. doi: 10.56093/ijans.v92i7.115253
17. Maloiy, G, Rugangazi, B, and Rowe, M. Energy expenditure during level locomotion in large desert ungulates: the one-humped camel and the domestic donkey. J Zool. (2009) 277:248–55. doi: 10.1111/j.1469-7998.2008.00535.x
18. Windberger, U, Auer, R, Seltenhammer, M, Mach, G, and Skidmore, JA. Near-Newtonian blood behavior–is it good to be a camel? Front Physiol. (2019) 10:906. doi: 10.3389/fphys.2019.00906
19. Al-Shorepy, SS. Identification of environmental factors affecting the racing performance of race camels in the United Arab Emirates. Emir J Food Agric. (2011):424–30.
20. Soroko, M, Jodkowska, E, and Dudek, K. Thermography diagnosis in monitoring the annual training cycle of racehorses. Med Weter. (2015) 71:52–8.
21. Turner, T. A., Pansch, J., and Wilson, J. H. (2001). Thermographic assessment of racing thoroughbreds. Proceeding of the American Association of Equine Practitioners, 344–346.
22. Simon, EL, Gaughan, EM, Epp, T, and Spire, M. Influence of exercise on thermographically determined surface temperatures of thoracic and pelvic limbs in horses. J Am Vet Med Assoc. (2006) 229:1940–4. doi: 10.2460/javma.229.12.1940
23. Yarnell, K, Fleming, J, Stratton, TD, and Brassington, R. Monitoring changes in skin temperature associated with exercise in horses on a water treadmill by use of infrared thermography. J Therm Biol. (2014) 45:110–6. doi: 10.1016/j.jtherbio.2014.08.003
24. Eddy, A, Van Hoogmoed, L, and Snyder, J. The role of thermography in the management of equine lameness. Vet J. (2001) 162:172–81. doi: 10.1053/tvjl.2001.0618
25. Tunley, B, and Henson, F. Reliability and repeatability of thermographic examination and the normal thermographic image of the thoracolumbar region in the horse. Equine Vet J. (2004) 36:306–12. doi: 10.2746/0425164044890652
26. Iglesias, C, Navas, F, Ciani, E, Arbulu, AA, González, A, Marín, C, et al. Biokinematics and applied thermography in the Canarian camel breed. Arch Zootec. (2020) 69:102–7. doi: 10.21071/az.v69i265.5045
27. Nav (Nomina anatomica veterinaria), Sixth edition. (2017). Prepared by the international committee on veterinary gross anatomical nomenclature Icvgan and published by the world Association of Veterinary Anatomists. Available at: https://www.wava-amav.org/.
28. González Ariza, A, Arando Arbulu, A, Navas González, FJ, León Jurado, JM, Delgado Bermejo, JV, and Camacho Vallejo, ME. Data mining-based discriminant analysis as a tool for the study of egg quality in native hen breeds. Sci Rep. (2022) 12:15873. doi: 10.1038/s41598-022-20111-z
30. Alexander, RM. Energy-saving mechanisms in walking and running. J Exp Biol. (1991) 160:55–69. doi: 10.1242/jeb.160.1.55
31. Delmas, R, and Liu, Y. Exploring Students’conceptions of the standard deviation. Stat Educ Res J. (2005) 4:55–82. doi: 10.52041/serj.v4i1.525
33. Kirimtat, A, Krejcar, O, Selamat, A, and Herrera-Viedma, E. Flir vs seek thermal cameras in biomedicine: comparative diagnosis through infrared thermography. BMC Bioinformatics. (2020) 21:1–10. doi: 10.1186/s12859-020-3355-7
34. Merkies, K, Ready, C, Farkas, L, and Hodder, A. Eye blink rates and eyelid twitches as a non-invasive measure of stress in the domestic horse. Animals. (2019) 9:562. doi: 10.3390/ani9080562
35. Ogard, WK. Proprioception in sports medicine and athletic conditioning. Strength Cond J. (2011) 33:111–8. doi: 10.1519/SSC.0b013e31821bf3ae
36. Dunbar, DC, Macpherson, JM, Simmons, RW, and Zarcades, A. Stabilization and mobility of the head, neck and trunk in horses during overground locomotion: comparisons with humans and other primates. J Exp Biol. (2008) 211:3889–907. doi: 10.1242/jeb.020578
37. Yarmolenko, PS, Moon, EJ, Landon, C, Manzoor, A, Hochman, DW, Viglianti, BL, et al. Thresholds for thermal damage to normal tissues: an update. Int J Hyperth. (2011) 27:320–43. doi: 10.3109/02656736.2010.534527
38. Kozlov, A, and Son’kin, V. Infrared thermography diagnostics of subcutaneous thermogenerators of non-shivering thermogenesis. Hum Physiol. (2019) 45:658–72. doi: 10.1134/S0362119719060070
39. Khumsap, S, Lanovaz, J, and Clayton, H. Three-dimensional kinematic analysis of horses with induced tarsal synovitis. Equine Vet J. (2004) 36:659–63. doi: 10.2746/0425164044848073
40. Ijichi, C, Evans, L, Woods, H, and Yarnell, K. The right angle: validating a standardised protocol for the use of infra-red thermography of eye temperature as a welfare indicator. Anim Welf. (2020) 29:123–31. doi: 10.7120/09627286.29.2.123
42. Soroko, M, Howell, K, Dudek, K, Henklewski, R, and Zielińska, P. The influence of breed, age, gender, training level and ambient temperature on forelimb and back temperature in racehorses. Anim Sci J. (2017) 88:347–55. doi: 10.1111/asj.12631
43. Wood, RI. Reinforcing aspects of androgens. Physiol Behav. (2004) 83:279–89. doi: 10.1016/j.physbeh.2004.08.012
44. Pigière, F, and Henrotay, D. Camels in the northern provinces of the Roman empire. J Archaeol Sci. (2012) 39:1531–9. doi: 10.1016/j.jas.2011.11.014
45. Hamdi, I, Benaissa, A, Babelhadj, B, Bedda, H, Aboub, S, and Loubaki, R. Composition and structure of the skin of dromedary (Camelus dromedarius, L. 1758) from two Algerian populations. J Anim Behav Biometeorol. (2022) 10:2217–2022. doi: 10.31893/jabb.22017
46. Dreiss, AN, Séchaud, R, Béziers, P, Villain, N, Genoud, M, Almasi, B, et al. Social huddling and physiological thermoregulation are related to melanism in the nocturnal barn owl. Oecologia. (2016) 180:371–81. doi: 10.1007/s00442-015-3491-3
47. Frank, RN, Puklin, JE, Stock, C, and Canter, LA. Race, iris color, and age-related macular degeneration. Trans Am Ophthalmol Soc. (2000) 98:109.
48. Hasi, S, Amu, G, and Zhang, W. Camel hair structure, properties, and commercial products. Handbook of research on health and environmental benefits of camel products Igi Global (2020).
Keywords: thermo-physiological response, exercise tolerance, camel welfare, camel phenotyping, genetic selection
Citation: Iglesias Pastrana C, Navas González FJ, Ciani E, Marín Navas C and Delgado Bermejo JV (2023) Thermographic ranges of dromedary camels during physical exercise: applications for physical health/welfare monitoring and phenotypic selection. Front. Vet. Sci. 10:1297412. doi: 10.3389/fvets.2023.1297412
Edited by:
Vladimír Jekl, University of Veterinary and Pharmaceutical Sciences Brno, CzechiaReviewed by:
Marcelo Ghezzi, Universidad Nacional del Centro de Buenos Aires, ArgentinaAntonio González Ariza, University of Córdoba, Spain
Emiliano Lasagna, University of Perugia, Italy
Cecilio Barba, Universidad de Córdoba, Spain
Copyright © 2023 Iglesias Pastrana, Navas González, Ciani, Marín Navas and Delgado Bermejo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Francisco Javier Navas González, ZmpuYXZhc0B1Y28uZXM=