 Laice A. Silva1
Laice A. Silva1 Monique F. S. Souza1Camila G. Torquetti1Daniele C. O. Freitas1Larissa G. A. Moreira1Thaynara P. Carvalho1
Monique F. S. Souza1Camila G. Torquetti1Daniele C. O. Freitas1Larissa G. A. Moreira1Thaynara P. Carvalho1 Clarissa H. Santana1Julia R. Thompson2Daniela C. O. Rosa2Tatiana R. Jesus2Tatiane A. Paixão3
Clarissa H. Santana1Julia R. Thompson2Daniela C. O. Rosa2Tatiana R. Jesus2Tatiane A. Paixão3 Renato L. Santos1*
Renato L. Santos1*- 1Departamento de Clínica e Cirurgia Veterinárias, Escola de Veterinária, Universidade Federal de Minas Gerais, Belo Horizonte, Minas Gerais, Brazil
- 2Sete Soluções e Tecnologia Ambiental, Belo Horizonte, Minas Gerais, Brazil
- 3Departamento de Patologia Geral, Instituto de Ciências Biológicas, Universidade Federal de Minas Gerais, Belo Horizonte, Minas Gerais, Brazil
This study included 47 free-ranging bats from the State of Minas Gerais, Brazil. Six bats (12.8%) had genital inflammatory lesions, and two of them (one Artibeus lituratus and one Glossophaga soricina, a frugivorous and a nectarivorous, respectively) were diagnosed with Brucella sp. infection through PCR, and antigens in intralesional macrophages were detected using immunohistochemistry.
Introduction
Brucella are gram-negative facultative intracellular bacteria belonging to the class of Alphaproteobacteria. Some species of the genus are causative agents of brucellosis, a disease associated with economic losses in livestock and public health issues (1). Classical Brucella species, originally isolated from terrestrial mammals—mostly domestic animals, were taxonomically classified according to their pathogenicity and host specificity, including B. abortus (cattle), B. melitensis (small ruminants), B. suis (pigs), B. ovis (sheep), B. canis (dogs), and B. neotomae (wood desert rats) (1). More recently, B. microti was isolated from the common vole (2), B. pinnipedialis and B. ceti were isolated from pinnipeds and cetaceans, respectively (3), B. papionis was isolated from baboons (4), and B. inopinata was isolated from a human breast implant (5), and both were recognized as new species. Atypical Brucella spp. have also been isolated from cold-blooded animals, such as amphibians (6), reptiles (7), and fish (8). Therefore, the recent expansion of the Brucella genus supports the notion that possibly many other Brucella species may infect wildlife (1).
Although Brucella spp. may have a broad cell and tissue tropism depending on the pathogen and host species (9), they are often associated with genital lesions in domestic animals (9, 10). For instance, B. abortus is an important cause of abortion in pregnant heifers and cows (11), and it is also an important cause of orchitis and epididymitis in males (10). In contrast, B. ovis infection in sheep is primarily associated with epididymitis in rams (12), although it may also be a cause of abortion in pregnant ewes (10). Bats have been investigated as potential hosts for Brucella spp. (13–16). Importantly, anti-Brucella antibodies have been detected in vampire bats (Desmodus rotundus) in Brazil (17), and recently, a novel Brucella species has been identified infecting vampire bats in Costa Rica (18). Thus, the finding of genital lesions, which are known to be associated with Brucella infection in many animal species (10–12), in free-ranging bats prompted us to investigate the possibility of Brucella infection. Therefore, this study provides evidence of Brucella sp. infection associated with genital lesions in frugivorous and nectarivorous bats, Artibeus lituratus and Glossophaga soricina, respectively.
Methods
Samples of testes and epididymides were obtained from 47 free-ranging male bats belonging to eight species: Carollia perspicillata (n = 17), Artibeus obscurus (n = 11), Glossophaga soricina (n = 7), Platyrrhinus lineatus (n = 6), Artibeus lituratus (n = 3), Dermanura cinerea (n = 1), Desmodus rotundus (n = 1), and Phyllostomus discolor (n = 1). These bats were captured in the northwestern region of the State of Minas Gerais, Brazil (Figure 1) using mist nets. The bats were euthanized by anesthetic overdose. Capturing and sampling were performed as part of an environmental monitoring program that followed all applicable laws and regulations and was authorized by the Instituto Estadual de Florestas (IEF) under protocol number 024-002/2021. Tissue samples were fixed in 10% buffered formalin, embedded in paraffin, and processed for histological analysis.
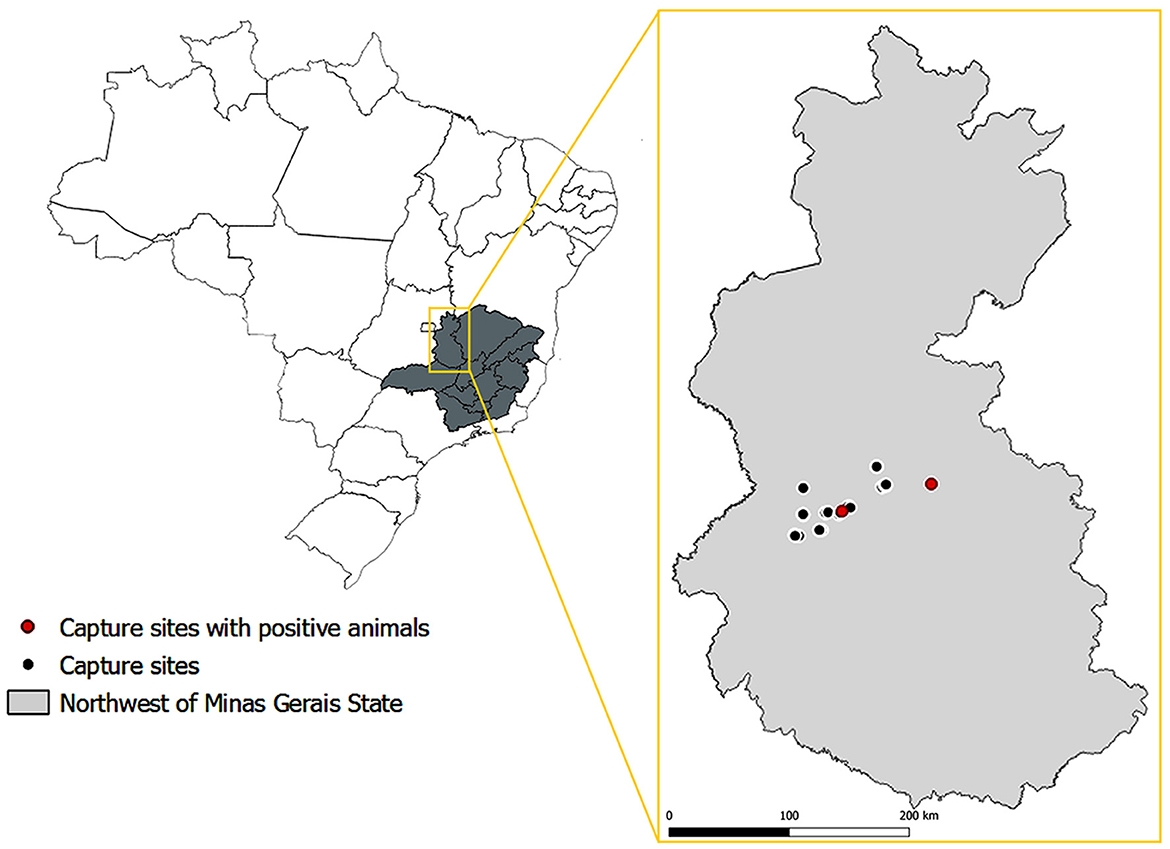
Figure 1. Location of sites where bats included in this study were captured, with the indication of locations where the capture of Brucella sp. infected bats took place. Source of maps: IBGE 2017. SRC: EPSG: 4674—SIRGAS 2000 (QGIS Version 3.28.2).
DNA was extracted from paraffin-embedded samples of testes and epididymides of all bats using a commercially available kit (ReliaPrep FFPE gDNA Miniprep System, Promega, Madison, USA), following the manufacturer's instructions, and used as a template DNA for polymerase chain reaction (PCR) targeting Brucella spp. genomic sequences. PCR targeted the Brucella spp. bcsp31 gene with primers 5′-TGGCTCGGTTGCCAATATCAA-3′ and 5′-CGCGCTTGCCTTTCAGGTCTG-3′, as previously described (19). PCR was performed with 2.5 μL of 10X Taq Buffer with KCl (Invitrogen, USA), 2.5 μL of 2 mM dNTP set solution (Invitrogen, USA), 1.5 μL of 25 mM MgCl2, 2.5 μL of each primer, 0.2 μL of Taq DNA polymerase (Invitrogen, USA), and 12 μL of template DNA samples with concentrations ranging from 64 to 200 ng/μL. The final reaction volume was 25 μL, with an expected product of 223 base pairs (bp). Amplification parameters were as follows: 94°C for 3 min; 40 cycles of 94°C for 30 s, 60°C for 30 s, and 72°C for 30 s, followed by a final extension at 72°C for 10 min. Amplified products were subjected to 1.5% agarose gel electrophoresis stained with SYBRSafe DNA Gel Stain (Invitrogen, USA) and examined in an ultraviolet light transilluminator.
Immunohistochemistry for in situ detection of Brucella sp. antigens was performed in samples from bats with genital lesions, as previously described (11), with modifications. Paraffin-embedded tissues were sectioned (4-μm-thick sections), dewaxed, hydrated, and washed three times in PBS. The sections were incubated with a 10% solution of hydrogen peroxide for 1 h for blocking endogenous peroxidase, rinsed in PBS, and then incubated with skimmed milk (25 μg/ml) for 45 min for blocking non-specific labeling. Sections were then incubated with a polyclonal anti-Brucella spp. antibody and diluted in the ratio of 1:1000 in 1% bovine serum albumin (BSA) for 1 h. The primary antibody employed in this study has been previously developed and characterized in our laboratory, as described in Xavier et al. (11), and it has been demonstrated to cross-react with various Brucella species (11, 20, 21). Sections were rinsed three times in PBS and incubated with a detection system (EnVision FLEX+; Dako, Carpinteria, CA, USA) for 30 min, followed by rinsing in PBS and development with chromogen 3′3-diaminobenzidine (DAB+ Substrate Chromogen; Dako) for 80 s, and sections were counterstained with Mayer's hematoxylin for 35 s. Positive controls included tissue sections from mice experimentally infected with Brucella ovis. Negative controls had the primary antibody replaced with sterile PBS.
Results
Six bats had inflammatory lesions in the testes and epididymis, including two Artibeus obscurus and one bat of each of the following species: Carollia perspicillata, Glossophaga soricina, Artibeus lituratus, and Phyllostomus discolor (Table 1). Histological changes were characterized by mild-to-severe multifocal to coalescing interstitial inflammatory infiltrate composed of lymphocytes and macrophages (Figure 2A). A necrotizing and histiocytic epididymitis was observed in one of the affected bats (bat #10; Figure 2B).
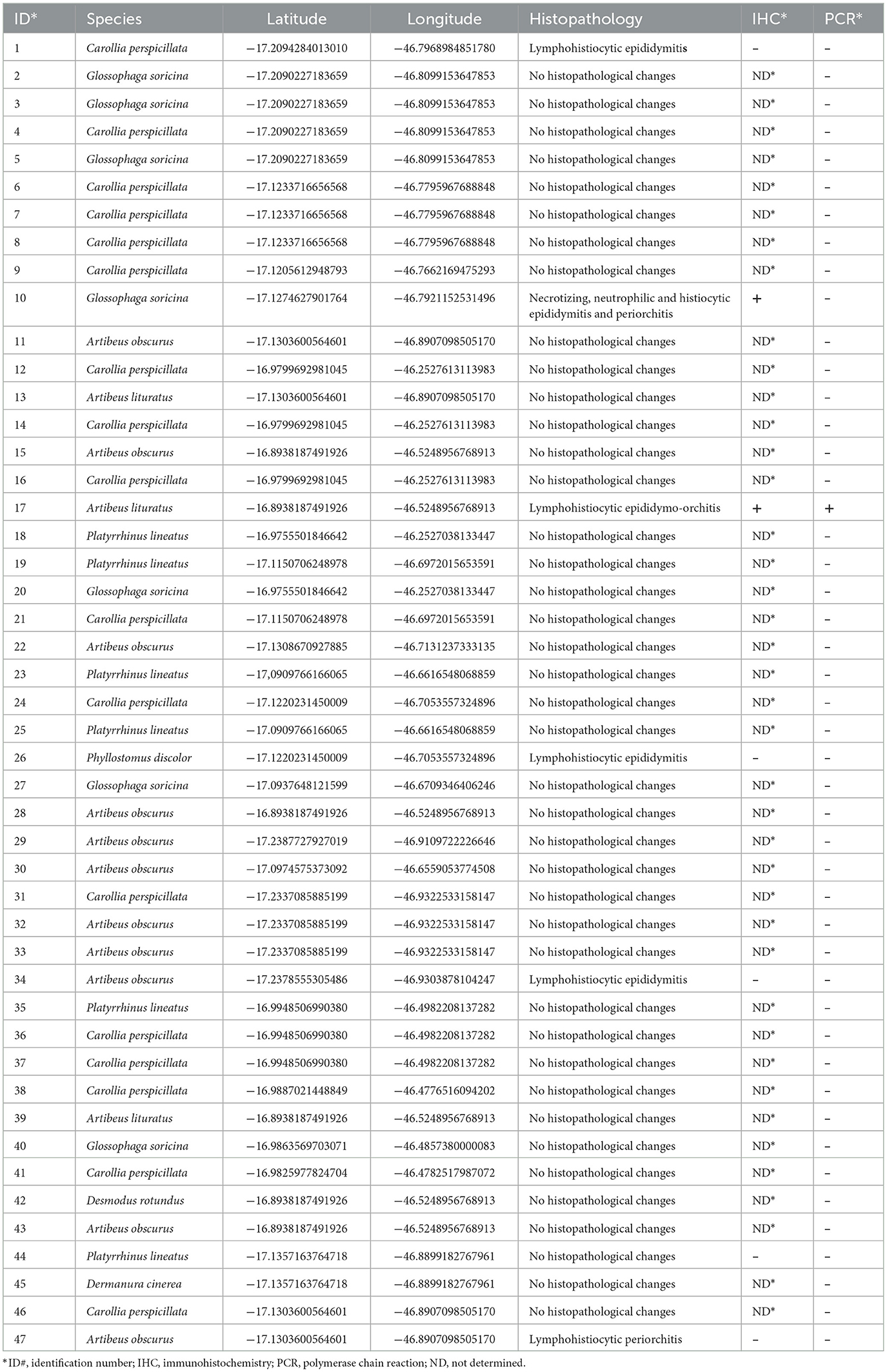
Table 1. Species, identification, geographic location, histopathology, immnunohistochemistry, and PCR of bats captured in Minas Gerais, Brazil, and analyzed in the study.
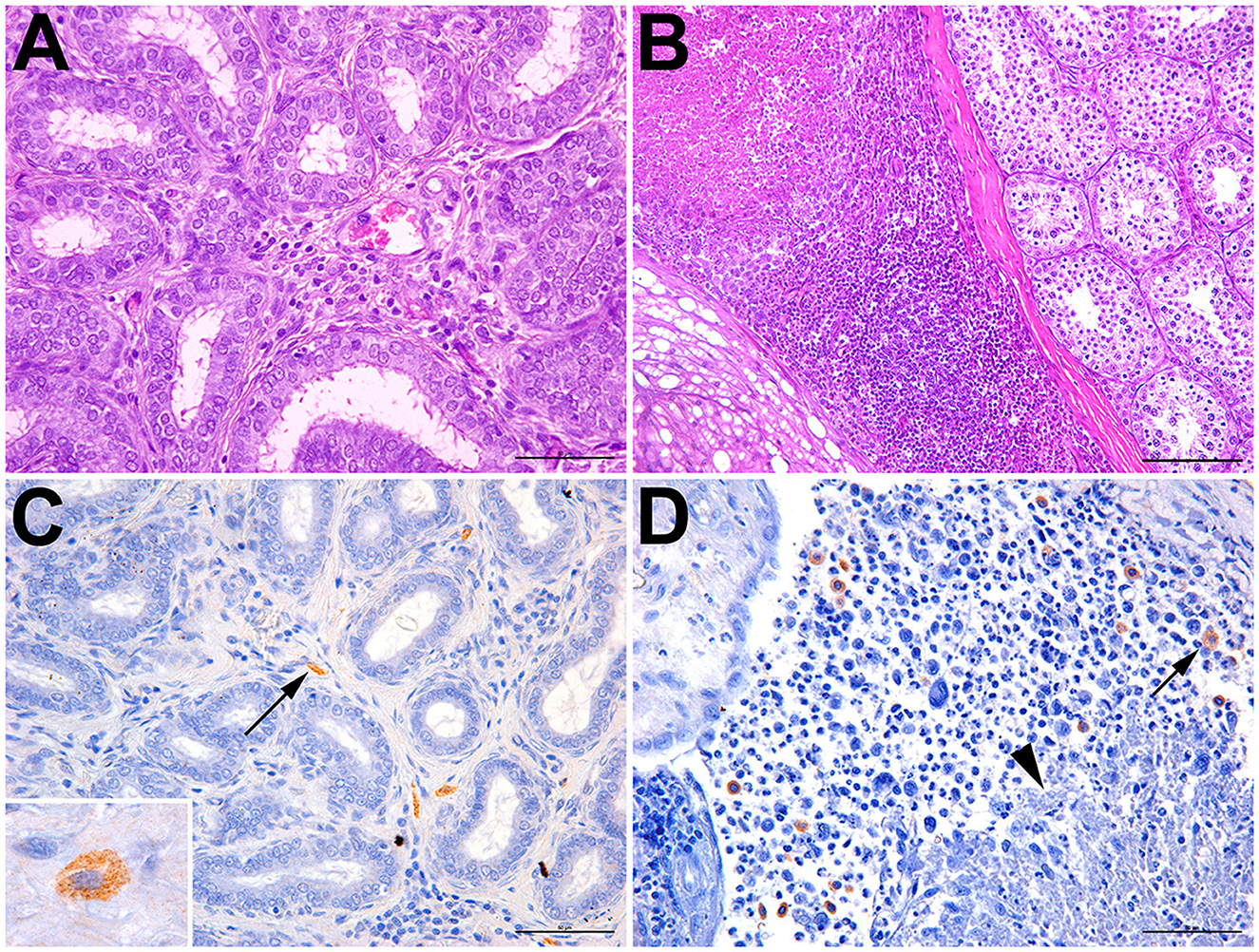
Figure 2. Epididymitis and orchitis in Artibeus lituratus (bat #17) and Glossophaga soricina (bat #10). (A) Bat #17; mild interstitial lymphohistiocytic epididymitis; hematoxylin and eosin, bar = 50μm. (B) Bat #10; necrotizing and lymphohistiocytic epididymitis and periorchitis; hematoxylin and eosin, bar = 100μm. (C) Bat #17; interstitial macrophages with cytoplasmic immunolabeled Brucella sp. (arrow); bar = 50μm. Inset: immunolabeled cytoplasmic granular contents morphologically compatible with coccobacilli. (D) Bat #10; multiple macrophages with cytoplasmic immunolabeled Brucella sp. (arrow) adjacent to an area of necrosis (arrowhead); bar = 50μm.
DNA samples extracted from paraffin-embedded testes and epididymides from all 47 bats were subjected to PCR targeting a Brucella spp. genomic sequence. Of the six bats, one bat (bat #17; Artibeus lituratus) had genital lesions tested positive by PCR, whereas none of the bats without genital lesions tested positive (Table 1).
Brucella spp. antigens were detected by immunohistochemistry in two bats, mostly associated with intralesional macrophages (Figures 2C, D; Table 1), including bat #17 (Artibeus lituratus), which also tested positive by PCR; and bat #10 (Glossophaga soricina). In some cells, the immunolabeled cytoplasmic contents of macrophages were morphologically compatible with coccobacilli (Figure 2C).
Considering the combined results of PCR and immunohistochemistry, in only two affected genera, Glossophaga and Artibeus (in total 21 bats), an estimated prevalence for that particular area [latitude extending from −16.8938187491926 to −17.2387727927019; and longitude from −46.2527038133447 to −46.9322533158147 (Figure 1)] was 9.52%, with a confidence interval between 1.18 and 30.38% (95% confidence level). Only the two genera that had positive animals were included in this estimate of prevalence since our data did not demonstrate the susceptibility of the other genera included in this study.
Discussion
These are the first reported cases of Brucella sp. infection in Artibeus lituratus and Glossophaga soricina, which are frugivorous and nectarivorous bats, respectively. Importantly, molecular evidence of Brucella sp. infection, based on the same PCR protocol employed in this study along with real-time PCR, was reported in bats of the species Myotis schreibersii and Myotis blythii from Georgia, where four cases were identified among 236 bats included in that study (13). Importantly, in addition to PCR, our study provided a second line of evidence of Brucella sp. infection, which was the intralesional in situ detection of Brucella sp. antigens by immunohistochemistry. In these cases, immunolabeling of Brucella antigens was morphologically compatible with coccobacilli associated with intralesional macrophages in sites of epididymal and testicular inflammation (epididymitis and orchitis, respectively), which are common Brucella-induced lesions in other host species (10). This is a relevant finding since it strongly supports the hypothesis that Brucella sp. has pathogenic potential for these animals. Although previous surveys (14–16), including one previous survey in the State of Minas Gerais, Brazil (16), failed to demonstrate evidence of Brucella sp. infection in various species of bats from different parts of the world, a pioneering study has demonstrated serologic evidence of Brucella infection in vampire bats (Desmodus rotundus) in Brazil (17). Importantly, during the course of this study, infection of Desmodus rotundus vampire bats with a novel Brucella species was described, and the newly identified species was named Brucella nosferati (18). Unfortunately, only formalin-fixed samples were available in this study, which prevented us from attempting isolation and characterization of the Brucella sp. that infected these bats.
The criterion for investigating Brucella spp. infection in these cases was to find inflammation in the testis and epididymis. In many domestic animal species, Brucella has a well-established tropism for the genital system (9), and it is an important cause of epididymitis and orchitis in various species (10, 22). The recent expansion of the Brucella genus with the recognition of Brucella sp. infecting many wildlife species (1) and the finding of epididymitis and orchitis in these South American bats lead us to these diagnoses, which are relevant in the context of the expanding knowledge on brucellosis in wildlife and its zoonotic potential, with obvious relevance in the context of One Health. Importantly, the tropism of newly recognized Brucella spp. from wildlife is still poorly characterized (9). However, there are a few cases of genital infections with B. ceti affecting various cetacean species (23), including a well-characterized case of necrotizing epididymo-orchitis with intralesional labeling of Brucella sp. and isolation of B. ceti from the testis and epididymis of a harbor porpoise (Phocoena phocoena) (24).
Surveillance studies, such as the present study, are highly relevant since bats play an important ecological role by providing pest control, plant pollination, and seed dispersal (25). Many bat species have territorial behavior, so they tend to return to the same refuge. However, due to increasing anthropogenic activities and habitat losses, there is a tendency for agglomeration of bat populations in restricted areas (26). Furthermore, bats differ from other mammals of the same size in several ways, including their ability to disperse rapidly and widely, the highly gregarious nature of their social structures, and their long lifespans, which are features that may favor disease transmission and dissemination (27). Bats have been identified as reservoirs or disseminators of many zoonoses or diseases that may be transmitted to other animal species (27–29). Therefore, future studies should assess the zoonotic potential or risk to farm animals posed by bat-derived Brucella strains. Importantly, although brucellosis usually has high morbidity and low mortality and is often associated with reproductive failure (10), the emergence of certain infectious diseases may result in marked population decline as observed after the introduction of Pseudogymnoascus destructans in North America, which causes white-nose syndrome in bats, and resulted in the collapse of bat populations as one of the most devastating outbreaks affecting wildlife ever recorded (30).
Conclusion
In conclusion, this study demonstrated Brucella sp. infection through two complementary diagnostic methods (immunohistochemistry and PCR) in Artibeus lituratus and Glossophaga soricina. Detection of Brucella sp. in these cases was associated with genital lesions. Importantly, demonstration of intralesional Brucella sp. in organs for which Brucella spp. have tropism in other host species (9) associated with lesions, namely epididymitis and orchitis, with a pattern of inflammation that have been classically recognized as Brucella spp.-induced lesions (10), supports the hypothesis that Brucella sp. is pathogenic for these species of bats. Unfortunately, the biological samples available in this study prevented us from isolating the organism, which could allow a proper identification at the species level.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
Ethics statement
The animal study was reviewed and approved by Instituto Estadual de Florestas (IEF) under protocol number: 024-002/2021.
Author contributions
TP and RS: conceptualization, data curation, supervision, validation, and project administration. LS, MS, CT, DF, LM, TC, CS, JT, DR, and TJ: methodology. LS, MS, TP, and RS: formal analysis. LS, MS, DF, LM, TP, and RS: investigation. LS, MS, and RS: writing—original draft preparation and writing—reviewing and editing. All authors have read and agreed to the published version of the manuscript.
Funding
The study in RS laboratory was supported by CNPq (Conselho Nacional de Desenvolvimento Científico e Tecnológico, Brazil), FAPEMIG (Fundação de Amparo a Pesquisa do Estado de Minas Gerais, Brazil), and CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior, Brazil).
Conflict of interest
JT, DR, and TJ were employed by Sete Soluções e Tecnologia Ambiental.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Moreno E. The one hundred year journey of the genus Brucella (Meyer and Shaw 1920). FEMS Microbiol Rev. (2021) 45:fuaa045. doi: 10.1093/femsre/fuaa045
2. Scholz HC, Hofer E, Vergnaud G, Fleche PL, Whatmore AM, Dahouk S, et al. Isolation of Brucella microti from mandibular lymph nodes of red foxes, Vulpes vulpes, in lower Austria. Vector Borne Zoonotic Dis. (2009) 9:153–6. doi: 10.1089/vbz.2008.0036
3. Foster G, Osterman BS, Godfroid J, Jacques I, Cloeckaert A. Brucella ceti sp. nov. and Brucella pinnipedialis sp. nov. for Brucella strains with cetaceans and seals as their preferred hosts. Int J Syst Evol Microbiol. (2007) 57:2688–93. doi: 10.1099/ijs.0.65269-0
4. Whatmore AM, Davison N, Cloeckaert A, Al Dahouk S, Zygmunt MS, Brew S, et al. Brucella papionis sp. nov., isolated from baboons (Papio spp.). Int J Syst Evol Microbiol. (2014) 64:4120. doi: 10.1099/ijs.0.065482-0
5. Scholz HC, Nöckler K, Göllner C, Bahn P, Vergnaud G, Tomaso H, et al. Brucella inopinata sp. nov., isolated from a breast implant infection. Int J Syst Evol Microbiol. (2010) 60:801–8. doi: 10.1099/ijs.0.011148-0
6. Eisenberg T, Hamann HP, Kaim U, Schlez K, Seeger H, Schauerte N, et al. Isolation of potentially novel Brucella spp. from frogs. Appl Environ Microbiol. (2012) 78:3753–5. doi: 10.1128/AEM.07509-11
7. Eisenberg T, Schlez K, Fawzy A, Völker I, Hechinger S, Curić M, et al. Expanding the host range: infection of a reptilian host (Furcifer pardalis) by an atypical Brucella strain. Antonie Van Leeuwenhoek. (2020) 113:1531–7. doi: 10.1007/s10482-020-01448-9
8. Eisenberg T, Riße K, Schauerte N, Geiger C, Blom J, Scholz HC. Isolation of a novel ‘atypical' Brucella strain from a blue spotted ribbontail ray (Taeniura lymma). Antonie Van Leeuwenhoek. (2017) 110:221–34. doi: 10.1007/s10482-016-0792-4
9. Carvalho TP, Silva LA, Castanheira TLL, Souza TD, Paixão TA, Lazaro-Anton L, et al. Cell and tissue tropism of Brucella spp. Infect Immun. (2023) 91:e0006223. doi: 10.1128/iai.00062-23
10. Poester FP, Samartino LE, Santos RL. Pathogenesis and pathobiology of brucellosis in livestock. Rev Sci Tech. (2013) 32:105–15. doi: 10.20506/rst.32.1.2193
11. Xavier MN, Paixão TA, Poester FP, Lage AP, Santos RL. Pathological, immunohistochemical and bacteriological study of tissues and milk of cows and fetuses experimentally infected with Brucella abortus. J Comp Pathol. (2009) 140:149–57. doi: 10.1016/j.jcpa.2008.10.004
12. Carvalho Júnior CA, Moustacas VS, Xavier MN, Costa EA, Costa LF, Silva TMA, et al. Andrological, pathologic, morphometric, and ultrasonographic findings in rams experimentally infected with Brucella ovis. Small Rumin Res. (2012) 102:213–22. doi: 10.1016/j.smallrumres.2011.08.004
13. Bai Y, Urushadze L, Osikowicz L, McKee C, Kuzmin I, Kandaurov A, et al. Molecular survey of bacterial zoonotic agents in bats from the country of Georgia (Caucasus). PLoS ONE. (2017) 12:e0171175. doi: 10.1371/journal.pone.0171175
14. McKee CD, Osikowicz LM, Schwedhelm TR, Bai Y, Castle KT, Kosoy MY. Survey of parasitic bacteria in bat bugs, Colorado. J Med Entomol. (2018) 55:237–41. doi: 10.1093/jme/tjx155
15. Imnadze T, Natradze I, Zhgenti E, Malania L, Abazashvili N, Sidamonidze K, et al. Identification of a novel Yersinia enterocolitica strain from bats in association with a bat die-off that occurred in Georgia (Caucasus). Microorganisms. (2020) 8:1000. doi: 10.3390/microorganisms8071000
16. Ferreira ACR, Vieira TM, Custódio DAC, Melo MN, Gontijo CMF, Lage AP, et al. Cross-sectional study on Brucella spp, Leptospira spp and Salmonella spp in bats from Montes Claros, Minas Gerais, Brazil. Comp Immunol Microbiol Infect Dis. (2021) 78:101692. doi: 10.1016/j.cimid.2021.101692
17. Ricciardi ID, Nunes MP, Andrade CM, Silva AG. Anti-Brucella agglutinins in bats and “Callithrix” monkeys. J Wildl Dis. (1976) 12:52–4. doi: 10.7589/0090-3558-12.1.52
18. Hernández-Mora G, Chacón-Díaz C, Moreira-Soto A, Barrantes-Granados O, Suárez-Esquivel M, Viquez-Ruiz E, et al. Virulent Brucella nosferati infecting Desmodus rotundus has emerging potential due to the broad foraging range of its bat host for humans, wild and domestic animals. mSphere. (2023) 61:23. doi: 10.1128/msphere.00061-23
19. Mayfield JE, Bricker BJ, Godfrey H, Crosby RM, Knight DJ, Hailing SM, et al. The cloning, expression, and nucleotide sequence of a gene coding for an immunogenic Brucella abortus protein. Gene. (1988) 63:1–9. doi: 10.1016/0378-1119(88)90540-9
20. Eckstein C, Mol JPS, Costa FB, Nunes PP, Lima PA, Melo MM, et al. Brucella ovis mutant in ABC transporter protects against Brucella canis infection in mice and it is safe for dogs. PLoS ONE. (2020) 15:e0231893. doi: 10.1371/journal.pone.0231893
21. Braz HMB, Silva MF, Carvalho TP, Silva LAD, Soares JB, Costa FB, et al. Pathogenesis of Brucella ovis in pregnant mice and protection induced by the candidate vaccine strain B. ovis ΔabcBA. Vaccine. (2022) 40:4617–24. doi: 10.1016/j.vaccine.2022.06.044
22. Santos RL, Souza TD, Mol JPS, Eckstein C, Paixão TA. Canine brucellosis: an update. Front Vet Sci. (2021) 8:594291. doi: 10.3389/fvets.2021.594291
23. Guzmán-Verri C, González-Barrientos R, Hernández-Mora G, Morales JA, Baquero-Calvo E, Chaves-Olarte E, et al. Brucella ceti and brucellosis in cetaceans. Front Cell Infect Microbiol. (2012) 2:3. doi: 10.3389/fcimb.2012.00003
24. Dagleish MP, Barley J, Finlayson J, Reid RJ, Foster G. Brucella ceti associated pathology in the testicle of a harbour porpoise (Phocoena phocoena). J Comp Pathol. (2008) 139:54–9. doi: 10.1016/j.jcpa.2008.03.004
25. Baldwin JW, Dechmann DKN, Thies W, Whitehead SR. Defensive fruit metabolites obstruct seed dispersal by altering bat behavior and physiology at multiple temporal scales. Ecology. (2020) 101:e02937. doi: 10.1002/ecy.2937
26. Frick WF, Kingston T, Flanders J. A review of the major threats and challenges to global bat conservation. Ann N Y Acad Sci. (2020) 1469:5–25. doi: 10.1111/nyas.14045
27. Tian J, Sun J, Li D, Wang N, Wang L, Zhang C, et al. Emerging viruses: cross-species transmission of coronaviruses, filoviruses, henipaviruses, and rotaviruses from bats. Cell Rep. (2022) 39:110969. doi: 10.1016/j.celrep.2022.110969
28. Streicker DG, Turmelle AS, Vonhof MJ, Kuzmin IV, McCracken GF, Rupprecht CE. Host phylogeny constrains cross-species emergence and establishment of rabies virus in bats. Science. (2010) 329:676–9. doi: 10.1126/science.1188836
29. Letko M, Seifert SN, Olival KJ, Plowright RK, Munster VJ. Bat-borne virus diversity, spillover and emergence. Nat Rev Microbiol. (2020) 18:461–71. doi: 10.1038/s41579-020-0394-z
Keywords: Brucella, brucellosis, epididymitis, orchitis, bats, Artibeus lituratus, Glossophaga soricina
Citation: Silva LA, Souza MFS, Torquetti CG, Freitas DCO, Moreira LGA, Carvalho TP, Santana CH, Thompson JR, Rosa DCO, Jesus TR, Paixão TA and Santos RL (2023) Antigenic and molecular evidence of Brucella sp.-associated epididymo-orchitis in frugivorous (Artibeus lituratus) and nectarivorous (Glossophaga soricina) bats in Brazil. Front. Vet. Sci. 10:1235299. doi: 10.3389/fvets.2023.1235299
Received: 06 June 2023; Accepted: 18 July 2023;
Published: 28 August 2023.
Edited by:
Maryam Dadar, Razi Vaccine and Serum Research Institute, IranReviewed by:
Elías Barquero-Calvo, National University of Costa Rica, Costa RicaClifton McKee, Johns Hopkins University, United States
Copyright © 2023 Silva, Souza, Torquetti, Freitas, Moreira, Carvalho, Santana, Thompson, Rosa, Jesus, Paixão and Santos. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Renato L. Santos, cmxzQHVmbWcuYnI=