 Ondřej Máca
Ondřej Máca- 1Department of Pathology and Parasitology, State Veterinary Institute Prague, Prague, Czechia
- 2Department of Zoology and Fisheries, Faculty of Agrobiology, Food and Natural Resources, Czech University of Life Sciences Prague, Prague, Czechia
- 3El Colegio de la Frontera Sur, Chetumal, Mexico
Species of Sarcocystis use various vertebrates as intermediate or definitive hosts in their life cycles. One of these is snakes, whose role as intermediate hosts for these protozoans is scarce; in fact, there are six records, but only three with molecular characterization. An imported green tree python was involved in the morphological and molecular characterization (four loci) of a new species of Sarcocystis localized in skeletal muscles. Sarcocystis moreliae sp. nov. has a type 1 sarcocyst with a smooth wall and is genetically similar (at the 18S rRNA gene) to two unnamed species of Sarcocystis found in Lytorhynchus diadema from Oman and Varanus salvator macromaculatus from Malaysia, but their detailed comparison is impossible. The new species showed lower similarity to its congeners in other loci (28S rRNA, ITS1, and cox1). This is the first morphological and genetic characterization of a Sarcocystis species in snakes of the genus Morelia, particularly M. viridis, using four loci, but more data are needed to fill the knowledge gap about snakes as intermediate hosts of Sarcocystis.
Introduction
Among the protozoans of the genus Sarcocystis Lankester, 1882 are parasites of reptiles (avian and non-avian), mammals, and fish that act as intermediate (IHs) and/or definitive hosts (DHs) in an obligatory heteroxenous life cycle. To date, mammals harbor the vast majority of known species of Sarcocystis by acting as IHs (148) and DHs (56), followed by reptiles [IH: 23, DH: 26; (see 1–3)]. However, most of those Sarcocystis species in reptiles have only been found in lizards and have been solely evaluated morphologically and morphometrically; they lack a molecular analysis and cannot be compared with other congeners.
In particular, two named and four unnamed species of Sarcocystis that use snakes as IHs have so far been reported worldwide [i.e., S. atractaspidis Parenzan, 1947 in Atractaspis leucomelas Boulenger, S. pythonis Tiegs, 1931 in Morelia spilota (Lacépède) (syns. M. argus, Python spilotes), and Sarcocystis spp. in Crotalus durissus terrificus Laurenti, Lytorhynchus diadema (Duméril, Bibron & Duméril), Malpolon monspessulanus (Hermann), and Psammophis schokari Forskall]. Of them, S. atractaspidis and S. pythonis are listed as species inquirendae by Dubey et al. (2) due to the absence of differential morphological descriptions and molecular characterization, while the sequence of Sarcocystis sp. in C. d. terrificus is absent from GenBank, and the three others were genetically analyzed and are compared herein with the new species.
The occurrence of Sarcocystis in snakes of the genus Morelia has been scarcely studied. In addition to the report of S. pythonis in M. spilota (Pythonidae) as an IH, there are also some records of DHs, such as Sarcocystis sp. in M. spilota variegata Gray from Australia (4), and Sarcocystis spp. (KC201639, KC201640) in M. viridis (Schlegel) from Germany (5).
The green tree python, M. viridis, is an arboreal snake inhabiting lowland and lower montane environments (0–2,000 m above sea level) from New Guinea Island up to north-eastern Australia, except the Bismarck Archipelago (6, 7). This python species feeds on lizards, mammals, and birds (8) and has become one of the most traded reptiles in the world (9). Since the molecular characterization of Sarcocystis species infecting and using snakes as IHs is scarce, and the role of M. cf. viridis in the life cycle of those protozoans is barely known, we report here the findings of sarcocysts in the skeletal muscles of a green tree python imported from Indonesia to the Czech Republic.
Methods
In 2021, a green tree python (Morelia cf. viridis) was imported to the Czech Republic from an unknown locality in Southwestern Papua, Indonesia. Due to the unknown collection location and the various species of Morelia inhabiting this geographic area (see (10)), we decided to name it M. cf. viridis. In October 2022, this 2-year-old snake died following long-term health problems and poor body condition (0.3 kg in weight). The carcass was sent to the State Veterinary Institute (SVI) Prague for necropsy, where bacteriological and parasitological analyses were applied through blood smears and the examination of the intestine, stomach, subcutaneous tissue, body cavity, liver, and muscles from the heart, head, trunk, and tail. These examinations revealed mixed bacterial and parasitic infections (unpublished data), but only data on Sarcocystis are shown herein. Muscles were examined by wet mounts for the presence of sarcocysts under 10 − 100x objective magnifications of a Leica DM2500 LED optical microscope with a Leica DMC5400 digital camera and Leica Application Suite X microscope software (both Leica Microsystems, Wetzlar, Germany). For histology, muscles with sarcocysts were fixed in 10% formalin solution and tissue sections were stained with hematoxylin and eosin. All measurements are given in micrometers unless otherwise mentioned.
Individual sarcocysts (four isolates) or muscles with numerous sarcocysts (two isolates) were transferred to Eppendorf tubes for DNA extraction. Genomic DNA was extracted by glass bead disruption from six isolates collected from skeletal muscles (head, trunk, and tail) using the NucleoSpin tissue XS kit (Macherey-Nagel, Düren, Germany) according to the manufacturer’s recommendations. DNA was stored at –20°C until use in PCR assays of 18S rRNA, 28S rRNA, ITS1, and cox1 loci. PCR and nested-PCR were carried out using primers for 18S rRNA (11), 28S rRNA (KL_P1F/KL_P2R) (12), ITS1 region (12), and cox1 (SF1/SR5) (13). The amplification cycles for all genes were as follows: 95°C for 3 min, 5 cycles of 94°C for 45 s, 64°C for 60 s, 72°C for 90 s; followed by 30 cycles of 95°C for 30 s, 52–60°C for 30 s, 72°C for 1 min; and 72°C for 10 min. RNase/Dnase-free water was used as a negative control in each PCR test. PCR products were later analyzed by electrophoresis on a 1% agarose gel, visualized by ethidium bromide staining, and purified using the ExoSAP-IT™ Express PCR Product Cleanup Reagent Kit (Thermo Fisher Scientific) according to the manufacturer’s protocol. Cleaned amplicons were sequenced through the commercial company Eurofins Genomics (Ebersberg, Germany) using both forward and reverse primers. Nucleotide sequences of the four loci derived in this study have been deposited in GenBank1 under accession numbers (OQ296416–OQ296418, OQ295898). Sequences were analyzed and edited using FinchTV software (Geospiza Inc., Seattle, WA, United States) and compared with those of the valid species of Sarcocystis in the GenBank NCBI database using Basic Local Alignment Search Tool (BLAST; http://blast.ncbi.nlm.nih.gov/Blast.cgi). Alignment of sequences was performed using MAFFT software version 7.2
Phylogenetic relationships of S. moreliae sp. nov. at 18S rRNA, 28S rRNA, and ITS1 loci were analyzed using the MEGA11 software (14). Phylogenetic trees were inferred using the maximum-likelihood (ML) method, with the Hasegawa-Kishino-Yano parameter model with a gamma distribution rate and a proportion of invariant sites (HKY + G + I) according to the best-fit substitution model using Bayesian Information Criterion scores (BIC) in MEGA 11. Phylogenetic analysis involved 52 nucleotide sequences, with a total of 1,776 aligned nucleotide positions for 18S rRNA, 28 sequences with 1,611 positions for 28S rRNA, and 22 sequences with 1,714 positions for ITS1 loci. In the case of the cox1 locus, phylogenetic methods used the ML method and a General Time Reversible model with a gamma distribution rate (GTR + G) because this best fitted the data involving 25 sequences in 1,038 positions. Bootstrap analyses were carried out using 1,000 replicates. Phylogenetic trees were constructed with sequences obtained in the present study along with reference sequences deposited in GenBank and rooted using the Toxoplasma gondii Nicolle et Manceaux, 1908 sequence and S. neurona Dubey, Davis, Speer, Bowman, de Lahunta, Granstrom, Topper, Hamir, Cummings et Suter, 1991 (ITS1).
Results
Examination of the sole green tree python revealed the presence of sarcocysts in the skeletal muscles (head, trunk, tail; Figures 1A, 2A); muscles from the heart were negative for the presence of parasites. All sarcocysts were microscopic and divided into compartments by septa (Figure 1B). Based only on LM, the cyst wall appeared to be smooth without visible protrusions on its surface (Figures 2A−C), thus belonging to type 1, although determining the subtype is impossible without transmission electron microscopy studies. The largest sarcocyst measured 1,118 × 123 μm, with a wall thickness of 0.55 μm. The banana-shaped bradyzoites (n = 20) were 6.63–7.99 × 1.43–1.87 μm in size (Figures 2B,C). Histological examination showed that no visible inflammatory reaction occurred (Figures 2A−C).
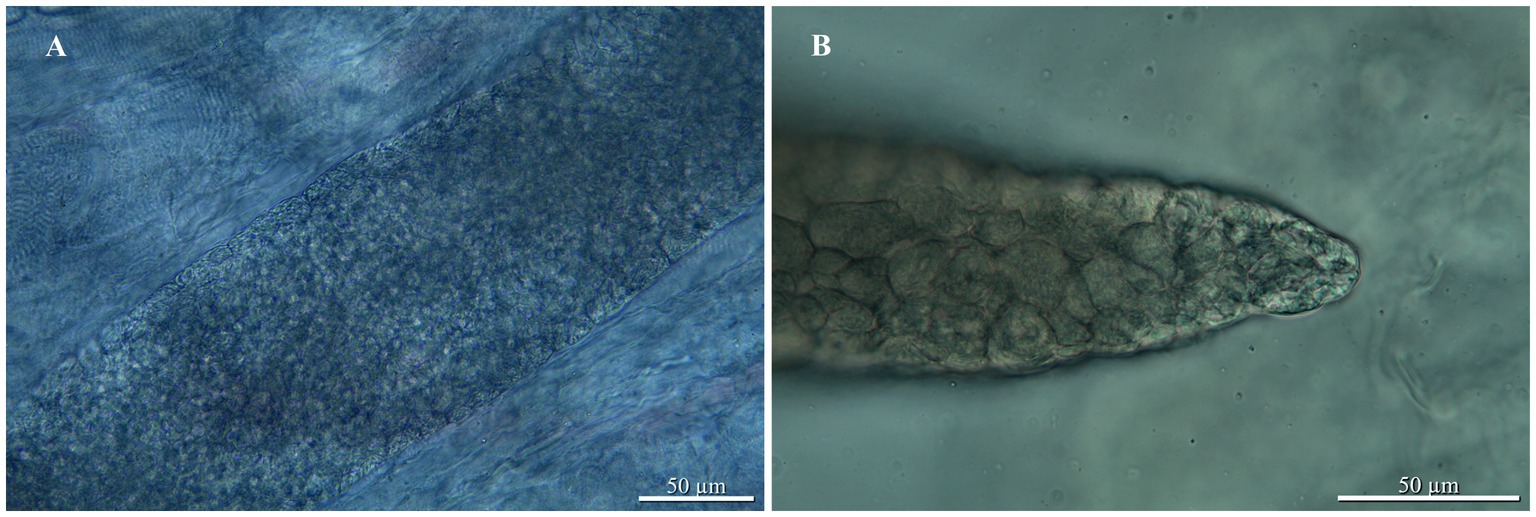
Figure 1. Sarcocystis moreliae sp. nov. from Morelia cf. viridis, light micrographs: (A) Sarcocyst in skeletal muscle of host; (B) Sarcocyst with septa.
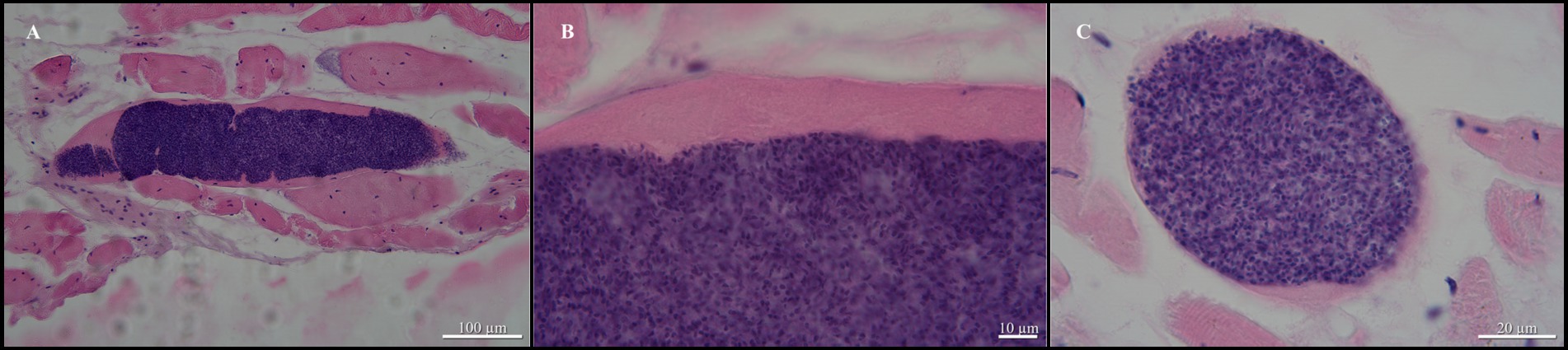
Figure 2. Sarcocystis moreliae sp. nov. from Morelia cf. viridis, hematoxylin, and eosin-stained histological sections of skeletal muscle. (A) Sarcocyst in skeletal muscle of host; (B) Same higher magnification showing sarcocyst wall without villar protrusions; (C) Transversal cut of sarcocyst showing bradyzoites.
Taxonomic summary
Family Sarcocystidae Poche, 1913
Sarcocystis moreliae sp. nov. (Figures 1, 2).
Intermediate host: Green tree python Morelia cf. viridis (Serpentes, Pythonidae).
Definitive host: Unknown.
Original distribution: Southwestern Papua, Indonesia.
Deposited material: Symbiotype (frozen muscle with sarcocysts), and genomic DNA in an Eppendorf tube were stored at SVI Prague. GenBank sequences OQ296418 (18S rRNA gene), OQ296417 (28S rRNA gene), OQ296416 (ITS1 region), and OQ295898 (cox1 gene).
ZooBank registration: To comply with the regulations set out in article 8.5 of the amended 2012 version of the International Code of Zoological Nomenclature (15), details of the new species have been submitted to ZooBank. The Life Science Identifier (LSID) for Sarcocystis moreliae sp. nov. is lsid:zoobank.org:pub:6A62F67A-CA7F-4938-AFFA-8EF0E62708F8.
Etymology: The specific epithet is derived from the generic name of its intermediate host, i.e., Morelia.
Molecular sequences of the 18S rRNA, 28S rRNA, ITS1, and cox1 loci from six sarcocyst isolates were successfully obtained with no intraspecific variability. The 18S rRNA representative sequence of S. moreliae sp. nov. showed query cover 31–100% with high similarity to two unnamed Sarcocystis species in the crowned leaf nose snake and in the common water monitor, respectively (see Table 1 for details). Sarcocystis moreliae sp. nov. formed a single clade with these two unnamed species (Figure 3A). The new species was also shown to be similar to Sarcocystis sp. in the California kingsnake, S. speeri Dubey and Lindsey, 1999 in the white-eared opossum, S. lari Prakas, Kutkienė, Butkauskas, Sruoga et Žalakevičius, 2014 in the white-tailed sea eagle, S. halieti Gjerde, Vikøren et Hamnes, 2018 in the red kite, S. neurona in the horse, and S. lutrae Gjerde et Josefsen, 2014 in the European otter (see Table 1 for details).
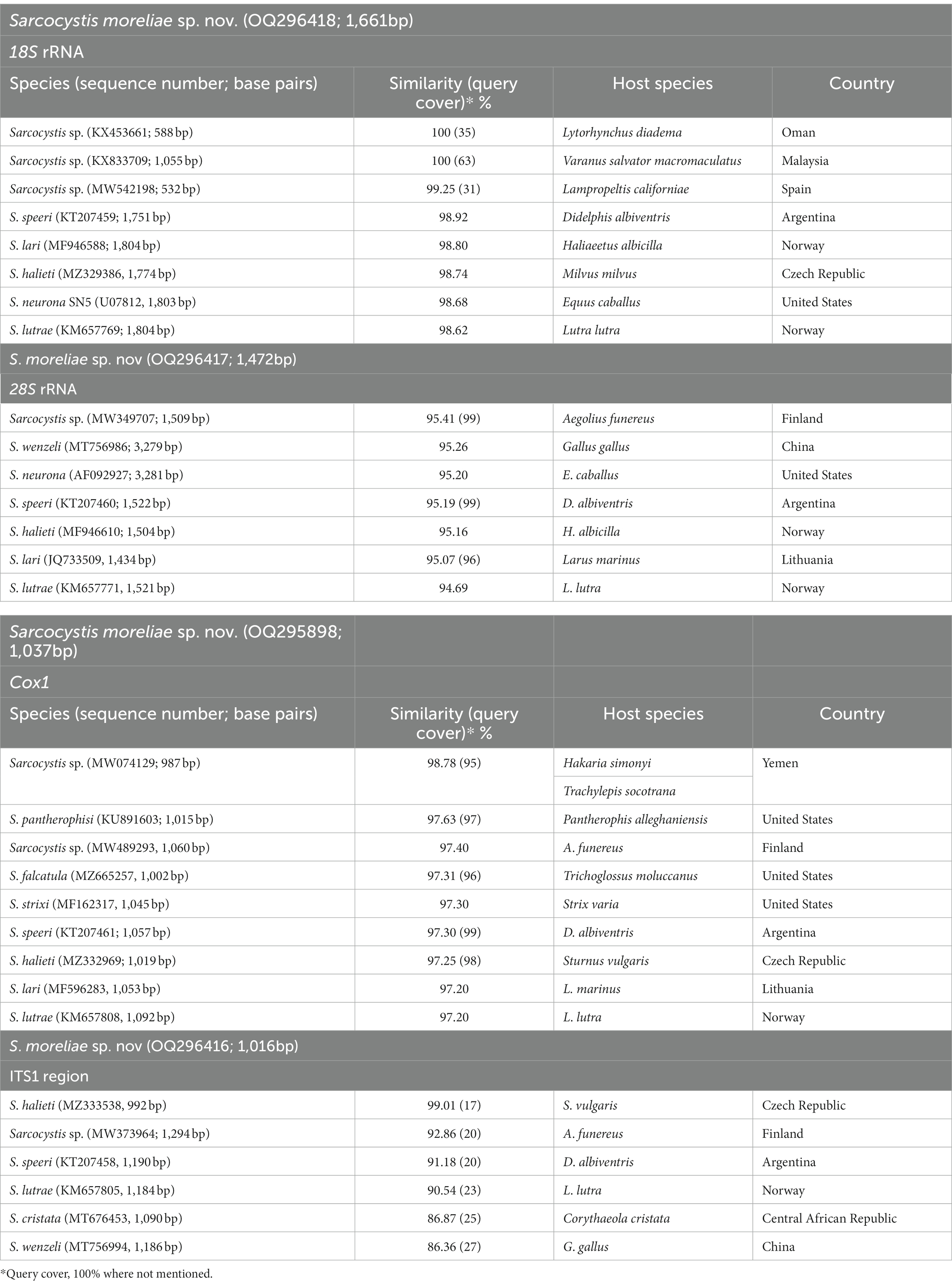
Table 1. Degree of similarity among Sarcocystis moreliae sp. nov with other Sarcocystis species retrieved from GenBank, by using for loci (18S rRNA, 28S rRNA, ITS1, and cox1).
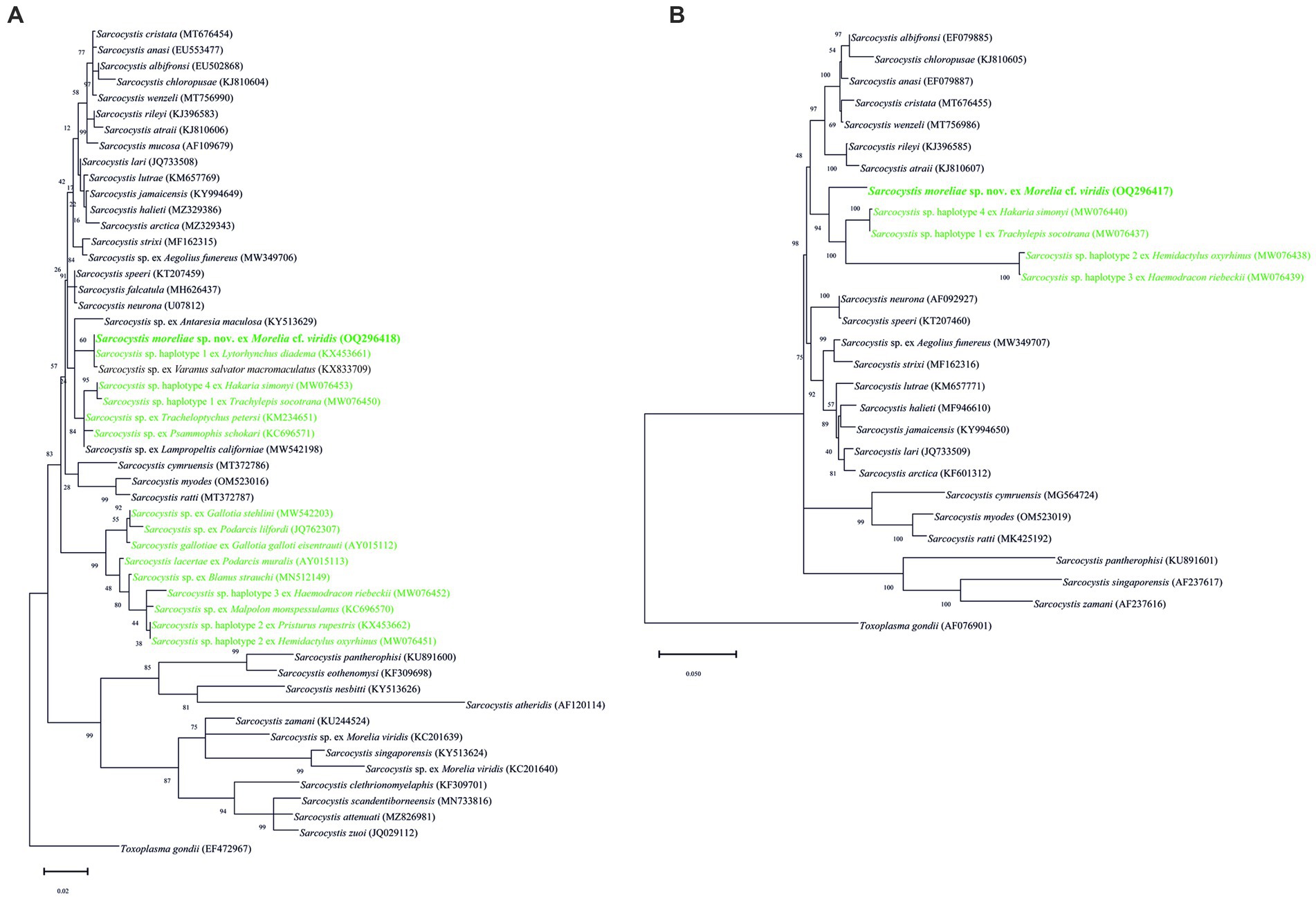
Figure 3. Phylogenetic trees of the species of Sarcocystis; those in green are for reptiles being used as intermediate hosts, based on (A) 18S rRNA and (B) 28S rRNA loci sequences. The numbers on phylogenetic trees represent bootstrap values based on 1,000 replications, while those after Sarcocystis species are GenBank accession numbers.
The sequence of the new species at 28S rRNA contained single nucleotide polymorphisms (SNPs) at positions 52 (C/T) and 669 (A/G). This sequence was most similar to Sarcocystis sp. in the Tengmalm’s owl and S. lari in the great black-backed gull, S. wenzeli Wenzel, Erber, Boch et Schellner, 1982 in the chicken, S. neurona in the horse, S. speeri in D. albiventris, S. halieti in H. albicilla, and S. lutrae in L. lutra (see Table 1 for details).
The cox1 sequence was most similar to Sarcocystis sp. in Hakaria simonyi and Trachylepis socotrana, S. pantherophisi (3) in the Eastern rat snake, Sarcocystis sp. in A. funereus, S. falcatula Stiles, 1893 in the rainbow lorikeet, S. strixi Verma, Rosypal von Dohlen, Mowery, Scott, Cerqueira-Cézar, Rosenthal, Dubey et Lindsay, 2017 in the barred owl, S. speeri in D. albiventris, S. halieti in the common starling, S. lari in the great black-backed gull, and S. lutrae (KM657808, 1,092 bp) in L. lutra (see Table 1 for details).
Amplification and sequencing of the ITS1 region were successful, and a representative sequence showed only one SNP at position 329, one double peak with A dominant over G, with no intraspecific variability for this marker. This sequence was similar to S. wenzeli in G. gallus, S. cristata Máca et González-Solís, 2021 in the great blue turaco, S. halieti in S. vulgaris, S. lutrae in L. lutra, Sarcocystis sp. in A. funereus, and S. speeri in D. albiventris (see Table 1 for details).
The topologies and phylogenetical relationships between S. moreliae sp. nov. with its congeners depended on the availability of sequences. The phylogenetic trees based on 18S rRNA, 28S rRNA, and cox1 genes grouped the new species along with Sarcocystis spp. in Ha. simonyi and T. socotrana, as well as with other species using reptiles as IHs, such as Sarcocystis spp. in Hemidactylus oxyrhinus Boulenger and Haemodracon riebeckii (Peters) at 28S and cox1 genes (Figures 3A,B, 4B). In particular, and due to the higher availability of sequences in the 18S rRNA gene, the tree displayed a larger branch where several unnamed Sarcocystis using reptiles as IHs and DHs were included (Figure 3A). On the other hand, the tree of the ITS1 region clearly isolated S. moreliae sp. nov. in a single clade within a group of species parasitizing snakes as DH (Figure 4A). None of the unnamed Sarcocystis used in the 18S rRNA tree were so far sequenced at the ITS1 region, so its comparison was not possible.

Figure 4. Phylogenetic tree of the species of Sarcocystis; those in green are for reptiles being used as intermediate hosts, based on (A) ITS1 and (B) cox1 loci sequences. The numbers on phylogenetic trees represent bootstrap values based on 1,000 replications, while those after Sarcocystis species are GenBank accession numbers.
Discussion
Some Sarcocystis have so far used six snake species as IHs, excluding S. moreliae sp. nov. Of them, S. atractaspidis in A. leucomelas seems to belong to a different genus (e.g., Besnoitia Henry; (see 16)), since its description was scarce and apparently incomplete and illustrations are missing (see (17)). On the other hand, the sarcocyst of S. pythonis in M. spilota and that of the new species in M. cf. viridis were similar in size (1,100 vs. 1,118 μm) and came from a congeneric snake species (see (18)), although S. pythonis is considered species inquirenda due to the lack of its molecular characterization. The sarcocysts of Sarcocystis sp. in C. d. terrificus also have septa, wall type 1, although slightly dissimilar in size, and with a thicker cyst wall than S. moreliae sp. nov. (2,000 μm × 50–150 μm; 1.00 vs. 0.55 μm, respectively; (see 19)). Moreover, it was molecularly analyzed, but the sequence was not uploaded in GenBank. The sarcocysts found in L. diadema, M. monspessulanus (20), and P. schokari (21) were molecularly analyzed (18S rRNA), but their morphological descriptions are missing. Thus, the morphological comparison between the new species and those using snakes as IHs is practically impossible, coupled with the ineffectiveness of using the morphology of developmental stages to separate species. Therefore, counting on a detailed morphological description, complemented with a good molecular characterization (high-quality sequences with a large number of base pairs) and their availability will help with the phylogenetic analysis and clear identification of these protozoans.
The 18S rRNA sequence of the new species matched completely with those of two unnamed Sarcocystis species in L. diadema and V. s. macromaculatus from Oman and Malaysia, respectively. However, it is difficult to say whether they belong to the same species since the latter two have not been phylogenetically analyzed (18S rRNA partially sequenced, no other loci), and morphological descriptions are lacking. In the case of V. s. macromaculatus, the site of infection, developmental stages (sarcocysts, sporocysts), and the role of the hosts as IHs or DHs are unknown. Moreover, the 18S rRNA sequence length of Sarcocystis sp. in L. diadema was too short (588 bp; (see 20)), and that in V. s. macromaculatus was only deposited in GenBank without formal publication. Interestingly, these two unnamed Sarcocystis have been recently grouped with Sarcocystis sp. using the California kingsnake (L. californiae) from Gran Canaria as a DH, although at a slightly lower percentage (99.7 and 99.3%, respectively) and with a short sequence (532 bp; (see 22)). Thus, we consider our finding to be a new species and encourage the examination of the three unnamed Sarcocystis to elucidate their taxonomic status.
At 28S rRNA and cox1 loci, the new species was not 100% the same as any other Sarcocystis species, although in both trees, it seems to be a sister species of four unnamed Sarcocystis found in other reptiles of the families Blanidae (Blanus strauchi Bedriaga), Gekkonidae (H. oxyrhinus), Phyllodactylidae (Hae. riebeckii), and Scincidae (Ha. simonyi, T. socotrana). These unnamed species were closely related to other unnamed Sarcocystis that used lizards, lacertids, geckos, and reptiles as IHs (e.g., M. monspessulanus, P. schokari; (see 23)), but they are all different from S. moreliae sp. nov. The role of various reptile species as IHs of Sarcocystis is still unknown but represents a possibility after considering their phylogenetic relationships. The use of the cox1 gene to delineate species of Sarcocystis in snake hosts has already been proven, although the number of sequences at this locus is still small (24).
On the other hand, the similarity of the new species with its congeners at the ITS1 region was very low, but it formed a group with species using snakes as DHs, namely S. attenuati Hu, Sun, Guo, Zeng, Zhang et Tao, 2022, S. pantherophisi, S. singaporensis (Zaman et Colley, 1975), S. zamani Beaver et Maleckar, 1981, and S. zuoi Hu, Ma, et Li, 2005. Unfortunately, no Sarcocystis species using reptiles as IHs have been sequenced at this genetic region, so a comparison of the new species with others is impossible, and the use of this locus to separate species occurring in reptiles is still uncertain. The 18S rRNA gene has been useful for the discrimination of sarcocystid protozoans (24), although the multigene phylogenetic analysis might be more robust in resolving relationships among species.
Since data on Sarcocystis using snake species of the genus Morelia as IHs are not available, of those in M. viridis as DHs that have been molecularly analyzed (i.e., KC201639 and KC201640), the new species clearly differs phylogenetically from them and forms a different clade. In fact, the above-mentioned sequences showed a close phylogenetic relationship with S. singaporensis and S. zamani, as well as being grouped along with other Sarcocystis species using snakes as DHs, as already stated by Moré et al. (5). These authors also mentioned that both sequences might either represent a known species of Sarcocystis, whose sequences are unavailable, or a new taxon. Whereas Wassermann et al. (25) pointed out that S. singaporensis is widely distributed in the Indo-Australian Archipelago and is probably frequently present in the green tree python in New Guinea, our results showed that this is not completely true, since other Sarcocystis spp. might also be present. Thus, M. viridis acts as IH and DH for at least three apparently different species of Sarcocystis, and all future parasitological examinations of this snake should include a review of feces and the intestinal tract, as well as the skeletal muscles in order to determine their possible presence.
None of the Sarcocystis species found in reptiles to date produce clinical illness. However, the present report of a new species of this genus in M. cf. viridis is important from diagnostic and epidemiological points of view because the parasite needs to find a definitive host to accomplish its life cycle. Unfortunately, only one green tree python was examined, and it is impossible to evaluate the impact of infection on other snakes acting as IHs, even for the one described here. This is an important finding, especially in connection with captive breeding, as this species is a great favorite of breeders worldwide, and with the importation of green tree pythons from areas where the complete life cycle can be realized, the risk of deterioration of the overall host condition is possible. This may be subject to further monitoring.
Snakes are considered potential carriers of human pathogens (see (25)). For example, S. nesbitti Mandour, 1969 uses snakes as DHs and could accidentally infect humans (see (26)) when the latter feed on food or water contaminated with sporocysts. However, when snakes act as IH, sporocysts might also be transmitted to humans when they eat improperly cooked sarcocyst-infected meat of the IH. Therefore, the presence of the new species should be monitored, especially in those places where the meat of snakes is used for human consumption.
The life cycle of S. moreliae sp. nov. is unknown, but its DHs might be monitors, birds, birds of prey (27, 28), quolls, or even other snakes. The black butcherbird (Cracticus quoyi) is distributed in Australia and New Guinea and is considered to be one of the most important predators of yellow individuals of M. viridis and might act as a DH of a Sarcocystis species. However, there are no records of protozoans in the feces of this bird species, except that of the informal report of Isospora sp. in C. nigrogularis from Queensland by O’Donoghue (29) at a conference.
As described above, there is only one case worldwide where a Morelia snake (M. spilota) was reported as an IH of a Sarcocystis species. Unfortunately, this was not molecularly analyzed, and its comparison with the new species is impossible. Therefore, this is the first morphological and genetic characterization of a Sarcocystis species in the Morelia genus and M. viridis, in particular, using four loci (18S rRNA, 28S rRNA, ITS1, and cox1). This kind of genetic analysis is mandatory for distinguishing between members of this protozoan species since the morphologies of their developmental stages are rather subjective and have low taxonomic relevance.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: https://www.ncbi.nlm.nih.gov/genbank/, OQ296418, OQ296417, OQ296416, and OQ295898.
Author contributions
OM conceived and designed the study and performed analyses. OM and DG-S wrote the manuscript. All authors contributed to the article and approved the submitted version.
Funding
Open access funding was provided by the Faculty of Agrobiology, Food and Natural Resources, Czech University of Life Sciences Prague.
Acknowledgments
The authors would like to thank all colleagues from the Pathology and Parasitology Department (SVI Prague), mainly I. Křičková for their technical support and a private breeder for providing valuable information. This research was performed within the Center for Infectious Animal Diseases.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abbreviations
bp, Base pairs; cox1, Cytochrome c oxidase subunit 1; DNA, Deoxyribonucleic acid; ITS1, Internal transcribed spacer 1; MAFFT, Multiple alignment using Fast Fourier Transform; rRNA, Ribosomal ribonucleic acid.
Footnotes
References
1. Odening, K. The present state of species-systematics in Sarcocystis Lankester, 1882 (Protista, Sporozoa, Coccidia). Syst Parasitol. (1998) 41:209–33. doi: 10.1023/A:1006090232343
2. Dubey, JP, Calero-Bernal, R, Rosenthal, BM, Speer, CA, and Fayer, R. Sarcocystosis of Animals and Humans 2nd Edn 53. Boca Raton, Florida: CRC Press (2016). 948–949
3. Verma, SK, Lindsay, DS, Mowery, JD, Rosenthal, BM, and Dubey, JP. Sarcocystis pantherophisi n. sp., from eastern rat snakes (Pantherophis alleghaniensis) as definitive hosts and interferon gamma gene knockout mice as experimental intermediate hosts. J Parasitol. (2017) 103:547–54. doi: 10.1645/17-2
4. Rzepczyk, CM. Evidence of a rat-snake life cycle for Sarcocystis. Int J Parasitol. (1974) 4:447–9. doi: 10.1016/0020-7519(74)90058-7
5. Moré, G, Pantchev, N, Herrmann, DC, Vrhovec, MG, Öfner, S, Conraths, FJ, et al. Molecular identification of Sarcocystis spp. helped to define the origin of green pythons (Morelia viridis) confiscated in Germany. Parasitology. (2014) 141:646–51. doi: 10.1017/S0031182013001960
6. O'Shea, M. A Guide to the Snakes of Papua New Guinea: The First Comprehensive Guide to the Snake Fauna of Papua New Guinea. Papua New Guinea, Singapore: Independent Publishing (1996).
7. Natusch, DJD, and Natusch, DFS. Distribution, abundance and demography of green pythons (Morelia viridis) in Cape York peninsula, Australia. Aust J Zool. (2011) 59:145–55. doi: 10.1071/ZO11031
8. Natusch, DJ, and Lyons, JA. Ecological attributes and trade of white-lipped pythons (genus Leiopython) in Indonesian New Guinea. Aust J Zool. (2012) 59:339–43. doi: 10.1071/ZO12017
9. Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES) (2022) CITES trade database. (Accessed 12 December, 2022)
10. Natusch, DJ, Esquerré, D, Lyons, JA, Hamidy, A, Lemmon, AR, Lemmon, EM, et al. Species delimitation and systematics of the green pythons (Morelia viridis complex) of Melanesia and Australia. Mol Phylogenet Evol. (2020) 142:106640. doi: 10.1016/j.ympev.2019.106640
11. Dubey, JP, Sykes, JE, Shelton, GD, Sharp, N, Verma, SK, Calero-Bernal, R, et al. Sarcocystis caninum and Sarcocystis svanai n. sp. (Apicomplexa: Sarcocystidae) associated with severe myositis and hepatitis in the domestic dog (Canis familiaris). J Eukaryot Microbiol. (2015) 62:307–17. doi: 10.1111/jeu.12182
12. Kutkienė, L, Prakas, P, Sruoga, A, and Butkauskas, D. The mallard duck (Anas platyrhynchos) as intermediate host for Sarcocystis wobeseri sp. nov. from the barnacle goose (Branta leucopsis). Parasitol Res. (2010) 107:879–88. doi: 10.1007/s00436-010-1945-4
13. Gjerde, B. Phylogenetic relationships among Sarcocystis species in cervids, cattle and sheep inferred from the mitochondrial cytochrome c oxidase subunit I gene. Int J Parasitol. (2013) 43:579–91. doi: 10.1016/j.ijpara.2013.02.004
14. Tamura, K, Stecher, G, and Kumar, S. MEGA11: molecular evolutionary genetics analysis version 11. Mol Biol Evol. (2021) 38:3022–7. doi: 10.1093/molbev/msab120
15. International Commission on Zoological Nomenclature (ICZN). Amendment of articles 8, 9, 10, 21 and 78 of the international code of zoological nomenclature to expand and refine methods of publication. Bull Zool Nomen. (2012) 69:161–9. doi: 10.21805/bzn.v69i3.a8.161
16. Matuschka, FR. Sarcocystis chalcidicolubris n. sp.: recognition of the life cycle in skinks of the genus Chalcides and snakes of the genus Coluber. J Parasitol. (1987) 73:1014–8. doi: 10.2307/3282526
17. Parenzan, P. Sarcosporidiosi (psorospermosi) da nuova specie (Prot.: Sarcocystis atractaspidis n. sp.) in reptile (Atractaspis). B Soc Nat Napoli. (1947) 55:117–9.
18. Tiegs, OW. Note on the occurrence of Sarcocystis in muscle of python. Parasitology. (1931) 23:412–4. doi: 10.1017/S0031182000013767
19. Roberts, JF, Wellehan, JFX Jr, Weisman, JL, Rush, M, Childress, AL, and Lindsay, DS. Massive muscular infection by a Sarcocystis species in a south American rattlesnake (Crotalus durissus terrificus). J Parasitol. (2015) 101:386–9. doi: 10.1645/14-642.1
20. Maia, JP, Harris, DJ, Carranza, S, and Gomez-Diaz, E. Assessing the diversity, host-specificity and infection patterns of Apicomplexan parasites in reptiles from Oman. Arabia Parasitol. (2016) 143:1730–47. doi: 10.1017/S0031182016001372
21. Tomé, B, Maia, JP, and Harris, DJ. Molecular assessment of apicomplexan parasites in the snake Psammophis from North Africa: do multiple parasite lineages reflect the final vertebrate host diet? J Parasitol. (2013) 99:883–7. doi: 10.1645/12-95.1
22. Santana-Hernández, KM, Javorská, K, Rodríguez-Ponce, E, Fecková, B, Šlapeta, J, and Modrý, D. Sarcocystis sp. infection (Apicomplexa: Sarcocystidae) in invasive California kingsnake Lampropeltis californiae (Serpentes: Colubridae) in gran Canaria. Parasitology. (2022) 149:1419–24. doi: 10.1017/S0031182022000841
23. Tomé, B, Maia, J, Perera, A, Carranza, S, and Vasconcelos, R. Parasites in a hotspot: diversity and specificity patterns of apicomplexans infecting reptiles from the Socotra archipelago. Parasitology. (2021) 148:42–52. doi: 10.1017/S0031182020002000
24. Ortega Pérez, P, Wibbelt, G, Brinkmann, A, Galindo Puentes, JA, Tuh, FY, Lakim, MB, et al. Description of Sarcocystis scandentiborneensis sp. nov. from treeshrews (Tupaia minor, T. tana) in northern Borneo with annotations on the utility of COI and 18S rDNA sequences for species delineation. Int J Parasitol Parasites Wildl. (2020) 12:220–31. doi: 10.1016/j.ijppaw.2020.07.003
25. Wassermann, W, Raisch, L, Lyons, JA, Natusch, DJD, Richter, S, Wirth, M, et al. Examination of Sarcocystis spp. of giant snakes from Australia and Southeast Asia confirms presence of a known pathogen—Sarcocystis nesbitti. PLoS One. (2017) 12:e0187984. doi: 10.1371/journal.pone.0187984
26. Lau, YL, Chang, PY, Tan, CT, Fong, MY, Mahmud, R, and Wong, KT. Sarcocystis nesbitti infection in human skeletal muscle: possible transmission from snakes. Am J Trop Med Hyg. (2014) 90:361–4. doi: 10.4269/ajtmh.12-0678
27. Wilson, D. Foraging ecology and diet of an ambush predator: the green python (Morelia viridis) In: RW Henderson and R Powell, editors. Biology of the Boas and Pythons. Eagle Mountain: Utah Eagle Mountain Publishing. 141–50.
28. San Diego Zoo Willdlife Alliance Animals & Plants: Python (2023). Available at: https://animals.sandiegozoo.org/ (Accessed on March 9, 2023).
Keywords: Indonesia, Sarcocystis, reptile, intermediate host, molecular characterization, new species
Citation: Máca O and González-Solís D (2023) Sarcocystis moreliae sp. nov. in the imported green tree python Morelia cf. viridis (Reptilia, Pythonidae). Front. Vet. Sci. 10:1211522. doi: 10.3389/fvets.2023.1211522
Edited by:
Vikrant Sudan, Guru Angad Dev Veterinary and Animal Sciences University, IndiaReviewed by:
Dean Konjević, University of Zagreb, CroatiaRanju Manoj, Cornell University, United States
Copyright © 2023 Máca and González-Solís. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: David González-Solís, dgonzale@ecosur.mx