 Imen Belhadj Slimen
Imen Belhadj Slimen Houari Yerou3,4
Houari Yerou3,4 Manel Ben Larbi
Manel Ben Larbi Naceur M’Hamdi
Naceur M’Hamdi
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Vet. Sci. , 17 August 2023
Sec. Animal Nutrition and Metabolism
Volume 10 - 2023 | https://doi.org/10.3389/fvets.2023.1200031
This article is part of the Research Topic Efforts To Reduce Feed-Food Competition View all 6 articles
This review summarizes the most relevant scientific literature related to the use of insects as alternative protein sources in poultry diets. The black soldier fly, the housefly, the beetle, mealworms, silkworms, earthworms, crickets, and grasshoppers are in the spotlight because they have been identified as an important future source of sustainable animal proteins for poultry feeding. Insect meals meet poultry requirements in terms of nutritional value, essential amino acid composition, nutrient digestibility, and feed acceptance. Furthermore, they are enriched with antimicrobial peptides and bioactive molecules that can improve global health. Results from poultry studies suggest equivalent or enhanced growth performances and quality of end-products as compared to fish meal and soybean meal. To outline this body of knowledge, this article states established threads of research about the nutrient profiles and the digestibility of insect meals, their subsequent effects on the growth and laying performances of poultry as well as the quality of meat, carcass, and eggs. To fully exploit insect-derived products, the effects of insect bioactive molecules (antimicrobial peptides, fatty acids, and polysaccharides) were addressed. Furthermore, as edible insects are likely to take a meaningful position in the feed and food chain, the safety of their derived products needs to be ensured. Some insights into the current knowledge on the prevalence of pathogens and contaminants in edible insects were highlighted. Finally, the effect of insect farming and processing treatment on the nutritive value of insect larvae was discussed. Our overview reveals that using insects can potentially solve problems related to reliance on other food sources, without altering the growth performances and the quality of meat and eggs.
“Feed-food competition” was defined as “the tensions and trade-offs between two alternative uses for edible crops: direct consumption by humans versus feeding livestock” (1). However, feed-food competition includes the use of production resources, such as land, wild fish, and water, and labor, capital, and ecosystem services. The allocation of these resources between all their possible uses is often determined by which end use is most profitable. Mottet et al. (2) estimated that 0.4 billion ha of cropland is oriented to feed production, in a competing way with human food. In the same context, Alexander et al. (3) estimated that feed crops and processed feed (i.e., oilseed cakes) account for 27% of livestock feed and 30% of crop consumption. Mottet et al. (2) reported similar estimations. If human food accounts for 45% of crops, the quantity of crops available for direct human consumption could rise by two-thirds if feed crops and processed feed were no longer given to livestock (1).
Digging in and scratching the ground is a natural behavior of poultry. This innate instinct is triggered by the numerous ‘delicacies’ that are found underground, including worms and insect larvae. Birds prefer insect-based diets, probably because of their taste and nutritive value. In this trend, insects can provide an alternative protein source for poultry production. They can generate income for both industrial and smallholder farmers and are potentially able to reduce feed-food competition. Recently, there has been an escalating interest in insects as an alternative protein source in poultry diets. There are 13 types of insects that have been identified and confirmed to be incorporated in poultry feeding. Among them, eight insects were approved by the EU regulations to be used as food (4) and feed (text of the commission regulation (EU) 2017/893 of 24 Mai 2017 amending annexes I and IV to Regulation (EC) No 999/2001 of the European Parliament and of the council, as well as the annexes X, XIV and XV to the commission regulation (EU) No 142/2011 as regards the provisions on processed animal protein). The most studied insects (Figure 1) include mainly the black soldier fly (BSF), and other insect types such as the housefly (HF), the mealworm (MW), the beetle, the locusts, the silkworm (SW), and the cockroach (5–7).

Figure 1. Some insects used in poultry feeding.
Insect meal is a low-cost and environmental-friendly ingredient. It contributes to decreasing environmental pollution as organic wastes are used in insect breeding (8–10). Insect meals are considered as potential substitution candidates for soybean meal (SBM) and fish meal (FM) since they have been used as partial or total substituents (9–13) without adverse effects on production performances or product quality.
This review article provides an overview of the nutritional value and digestibility of some insect meals, as well as their effects on animal growth performance, meat and egg quality, and overall health. Concerns about the safety of rearing insects or feeding insect-based diets are also addressed.
Increasing meat production requires the expansion of agricultural areas and the increase of feed and water consumption. The reduction of arable lands, global climate change, and the shortage of freshwater resources make it hard to meet the escalating demand for proteins. In this context, it makes sense to look for alternative and sustainable protein sources, which may contribute to global food security. Edible insects indeed require much less land than conventional livestock (14), but they also produce much fewer greenhouse gases (14) allowing them to mitigate the negative effects of climate change (15). Indeed, insects have a high food conversion ratio (FCR). To produce the same amount of proteins, crickets, for example, require half as much feed as broiler chickens, four times less feed than sheep, and twelve times less feed than cattle (16). Moreover, insect meals are a good source of protein, energy, digestible essential and nonessential amino acids, and saturated (SFA), monounsaturated (MUFA), and polyunsaturated fatty acids (PUFA), in addition to vitamins and minerals.
The black soldier fly (BSF) is a common and widespread fly of the family Stratiomyidae. BSF meal is a naturally nutritious, protein-packed, and calcium-rich source. The concentration of crude protein (CP) in BSF meal varies from 35 to 61%, and the content of crude fat (CF) ranges from 7 to 42% (Table 1). High concentrations of lauric and palmitic acids are also reported (48). Methionine, lysine, threonine, and valine contents were found to be between 0.08 and 0.90% (21, 22, 49), 0.34 and 3.30% (19–21), 0.22 and 2.26% (21, 22), and 0.33 and 3.38% (21, 22) respectively. The concentration of calcium ranges from 1.21% (22) to 4.39% (17), and that of phosphorous varies from 0.74% (23) to 0.95% (22). The nutritive value of BSF meal depends on the rearing substrate, the age of harvesting, and the technology of processing. The pressing and defatting treatments could decrease fat content, from 37% (original) to 27.36% (steamed), 17.05% (methanol extracted), 17.95% (normal pressing), or 13.05% with heated pressing (50).
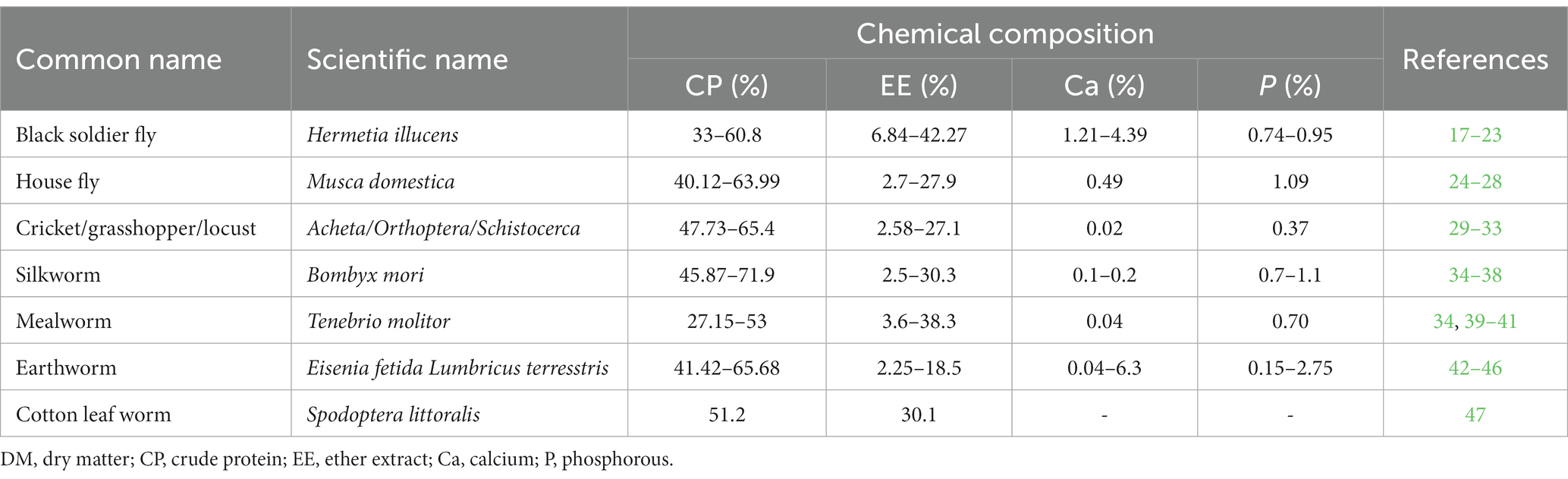
Table 1. Chemical composition of some common insect meals used in poultry nutrition (DM basis).
The housefly (HF) can be found in all countries and can be reared on animal manure and food waste. HF meal contains considerable amounts of CP and CF ranging from 40 to 64%, and from 2.5 to 28%, respectively (Table 1). In HF larvae, the content of CP decreases with age, whereas the content of CF increases (51, 52). Lysine and methionine are the most limiting amino acids in poultry nutrition. Both of these amino acids are available in high amounts in HF meal (53). Interestingly, the amino acid profile of HF meal resembles that of FM. Similarly to BSF meal, the processing method of the insect impacts the nutritive value of the meal (52).
Mealworms (MW) can be found worldwide where warm, dark, and damp places are available. MW meal is considered a good source of protein and fat, with a CP amount ranging from 27 to 54%, and CF content from 4 to 34% (Table 1). Values of ash, crude fibers, and energy in MW larvae range from 3.0 to 4.5% (53, 54), 5.0 to 8.8% (53, 54), and 1,378 to 4029.63 Kcal/Kg DM (45, 49), respectively. As compared to SBM, MW larvae had higher CP, CF, and ADF (11). Methionine + cysteine, lysine, leucine, isoleucine, histidine, threonine, valine, and arginine contents are higher in SBM, whereas MWs had greater tryptophan content (11). Moreover, MW larvae contain low amounts of Ca (434.59 mg/Kg) and high levels of K and P [9479.73 and 7060.70 mg/Kg, respectively (41)]. Consequently, the Ca/P ratio is not suitable for poultry production, especially for hens, and Ca supplementation is recommended (5). In addition to these minerals, Zn, Fe, and Cu are found in MW in considerable amounts [104.28, 66.87, and 13.27 mg/kg, respectively (41)].
Crickets and grasshoppers are also valuable protein sources for poultry. The amount of CP ranges from 48 to 65% and CF is between 3 and 21% (Table 1). Of course, these contents varied between African grasshoppers (Acanthacris ruficornis), Chinese grasshoppers (Acrida cinerea), wild edible grasshoppers (Ruspolia nitidual), short-horned grasshoppers (Oxya hyla hyla), and desert locusts (Schistocerca gregaria) (32). The crude fiber content varies between 1.06 and 9.21% (55, 56). Ojewola and his coauthors reported GE values of 1,618 and 1917 Kcal/Kg (55, 57), and Sun and his collaborators reported ME values ranging from 3,923 to 4,018 Kcal/Kg (58). The amino acid (AA) content matched well with those of FM. Regarding methionine, cysteine, and lysine, the relative contents were evaluated at 1.70, 0.69, and 3.79% on DM basis, respectively (30).
.In addition to crickets, earthworms (EW) are a good source of protein, energy, and amino acids (42, 43, 49). The CP content of EW meal ranges from 41 to 66%, and CF is between 3.5 and 18% (Table 1). Important contents of lysine and methionine were recorded in EWs (45, 60). However, it is important to mention that these values depend on the freshness and dryness of the EWs.
The nutritional value of a feed ingredient is determined by its chemical composition and the bioavailability of its nutrients. These two factors impact the use of nutrients in forming cells and tissues and in performing the maintenance and production functions of the bird body. Hence, it is important to examine the content of balanced proteins; the profile of amino acids, especially the essential ones; the content and composition of fats; and the amounts of vitamins and minerals. Nowadays, there is evidence that insects are rich in nutrients (proteins, energy, fat, and ash), including those considered essentials for the growth and production of birds. Scientific data indicate that edible insects are a source of complete animal protein and contain all essential amino acids. They can therefore be utilized in poultry diets. Also, they can be incorporated in cereal-based poultry feed, since cereals are poor in essential amino acids such as lysine, threonine, and tryptophan. In addition, insects are good sources of lysine, which is the reference amino acid in poultry and all remaining essential amino acids are related to it.
Adult insects have lower ME content than larvae and pupae because of the decrease of the crude fat content due to age. The nutritional value of lipids is related to the content and profile of their fatty acids. Among saturated fatty acids, insects have high contents of palmitic acid (C16:0), which is involved in stimulating blood cholesterol, especially the LDL fraction. Insects contain also both MUFA and PUFA, especially oleic (C18:1) and linoleic (C18:2) which were noticed for their positive effects on broiler meat quality (61, 62). Knowledge of nutrients and the amounts in which they occur in various insect species indicates the possibility to switch to insect-based diets in poultry feeding. However, knowledge of the best combination ratios of insect proteins/cereals proteins should be deepened.
Some decades ago, BSF larvae were reared on beef cattle feces and urine. They were then collected, dried, and fed to growing pigs. In an assay, researchers demonstrated that BSF larvae are a suitable ingredient for pig diets, although the fat content may affect both the palatability and digestibility of the diet (63). Many other reports have confirmed that using insect meals as an alternative protein source in broilers resulted in a reduced FCR and improved meat quality indicators. These findings may be attributed to the enhanced digestibility of dry matter, crude protein, crude fiber, ether extract, and ash (34, 64). Cullere et al. (65) reported no significant differences in DM, OM, and starch digestibility between growing broilers that received BSF meal and controls. However, they calculated superior EE digestibility for the group of broilers that received 15% defatted BSF larvae meal. The total tract digestibility (TTD) of DM and ash did not differ significantly between housefly larvae and pupae meals. However, AME and EE were different (66). Bosch et al. (67) evaluated the in-vitro OM digestibility of MW larvae to be 91.5%. De Marco et al. (49) compared MW and BSF meals regarding the TTD of nutrients and energy, and concluded that there were no significant differences between the two insect meals for all nutrients, except for the value of EE which was higher in MW larvae meal than in BSF meal (Table 2). Similarly, no statistical difference was reported between these two insect meals for AME (4026.94 and 4151.14 kcal/kg DM for MW and BSF, respectively) and AMEn (AMEn = 3826.31 and 3964.84 kcal/kg DM for MW and BSF, respectively).
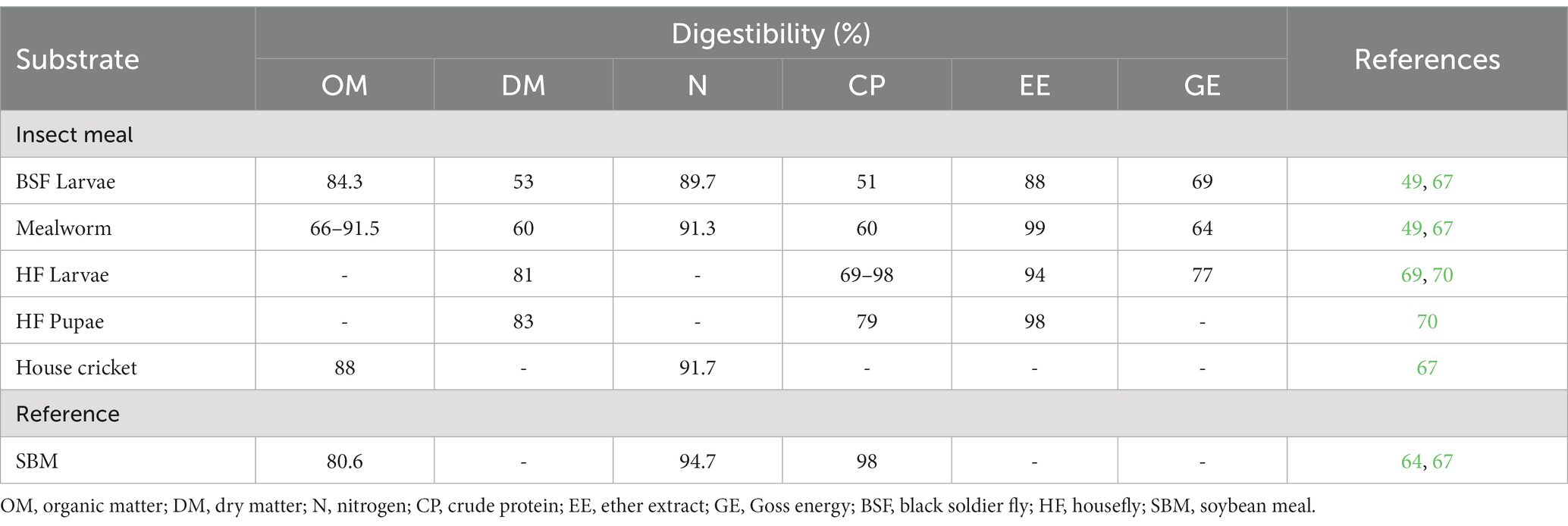
Table 2. Total tract apparent digestibility of some insect meals (%).
The coefficient of total tract digestibility (CTTD) of crude fibers (CF) of the housefly meal (Musca domestica) ranges from 0.58 to 0.62 (66). The CTTD of neutral detergent fibers (NDF) was evaluated at 0.87 and that of acid detergent fibers (ADF) varied from 0.35 to 0.67 for larvae and pupae meals (66). In poultry, dietary fibers are associated with reduced digestion as they lower the transit rate of almost all nutrients in the upper digestive tract, and thus a longer time for digestion was registered in the lower digestive tract (68). Bosch et al. (67) reported an in vitro nitrogen digestibility of MW larvae of 91.3%. The proteins of the housefly larvae meal (Musca domestica) are highly digestible in broilers. Zuidhof et al. (69) and Hwangbo et al. (64) evaluated the TTDs for CP of housefly larvae meal at 98.8 and 98%, respectively. Lower values were obtained by Pieterse and Pretorius (66) who compared between pupae and larvae meals of houseflies, and calculated TTDs of 79 and 69%, respectively. The TTD values reported in the literature of the CP of pupae meal are comparable or higher to those of soybean meal (Table 2). Similarly, Cullere et al. (65) tested the digestibility of CP from BSF in growing broiler quail. They concluded that there are no significant differences between the groups that received 10 and 15% defatted BSF larvae meal compared to the control group.
The AA of insects are very digestible in broilers. The CTTD of both essential and non-essential amino acids ranges from 0.83 to 1.00 (66). The reported TTD values of essential amino acids of houseflies are higher than those of soya oil cake meal. De Marco et al. (49) stated that the average of the apparent ileal digestibility coefficient (AIDC) of the AAs was greater in MW (0.86) as compared to BSF (0.68). Zuidhof et al. (69), Hwangbo et al. (64), and Pieterse and Pretorius (66) concluded that the CTTDs for certain essential amino acids (threonine, lysine, methionine, isoleucine, phenylalanine, and leucine) and all non-essential amino acids, except tyrosine, calculated for pupae meal are significantly (p < 0.05) higher than those calculated for larvae meal. In another study, Hall et al. (53) compared the AIDC and the true ileal digestibility coefficient (TIDC) of AA from housefly larvae and those from fish meal. They concluded that these two protein sources had similar values of AIDC and TIDC, and these values are close to those reported in the literature (71, 72). Nevertheless, in another study, Ravindran et al. (73) reported intermediate digestibility coefficients of amino acids in FM. This notable mismatch could be attributed to the method used to assess digestibility, or to the drying process which may damage the AAs. Grasshoppers were also reported for the elevated digestibility of their AAs. The true amino acid digestibility (TAAD) coefficients of methionine, cysteine, and lysine in grasshoppers were 0.97, 0.84, and 0.95, respectively, determined in cecectomized roosters (30). According to this latter study, the average TAAD coefficients of field crickets (92.9%) were higher than that of fish meal (91.3%), and the TAAD coefficients for EAAs in field crickets ranged from 82% for cysteine to 99% for asparagines.
It is worth stating that the findings regarding TTDs of CP and amino acids of larvae and pupae meals are surprising since pupae hold a chitin layer, an indigestible polysaccharide found in the cuticle of insects, which may decrease the availability of nutrients once fed to broilers (74). Decreased CTTD for CP from the housefly larvae meal may be due to overheating during processing, which is the factor most often responsible for damaging the quality of proteins and reducing the digestibility of amino acids (75). Indeed, the milling process of the pupae may be responsible for reducing the particle size. Thus, the surface area available for digestion is increased and the substances of the chitin layer are more available to the animal (76). It is certain that the drying and processing of insects’ larvae affect the chemical composition and the digestibility of nutrients. Raw MWs had a higher digestibility of CP compared to cooked ones (77). However, it is worth stating that using raw insects in poultry diets maybe not be feasible because of storage, transportation, and product safety issues. Oven-dried larvae at 60°C had a greater AA digestibility than microwave-dried ones (78). Despite the cooking process, the quality of insect proteins depends on the stage of insect production (77). In fact, the amount of chitin is higher in adult crickets than in larvae. Consequently, the digestibility of adult crickets is reduced (77). In the same trend, an assay was carried out to evaluate the effect of including 0.05% cricket chitosan or 0.05% cricket chitin in Cobb 500 male broiler chickens. The results showed that the intestinal morphology was negatively affected and the expression of some nutrient transporters (PepT1, EAAT3, SGLT1, and SGLT5) was downregulated (79).
The high content of digestible CP and amino acids in insect-derived products introduces them as a potent future-proof solution to upgrade protein self-sufficiency in animal feed. Since they are enriched in essential and limiting amino acids (lysine, threonine, methionine, and tryptophan), insects meet the nutritional requirements of poultry and contribute to the development of a highly formulated feed.
Bioactive substances derived from insects can be allocated into three main categories: antimicrobial peptides (AMPs), fatty acids, and polysaccharides [Figure 2 (80)].
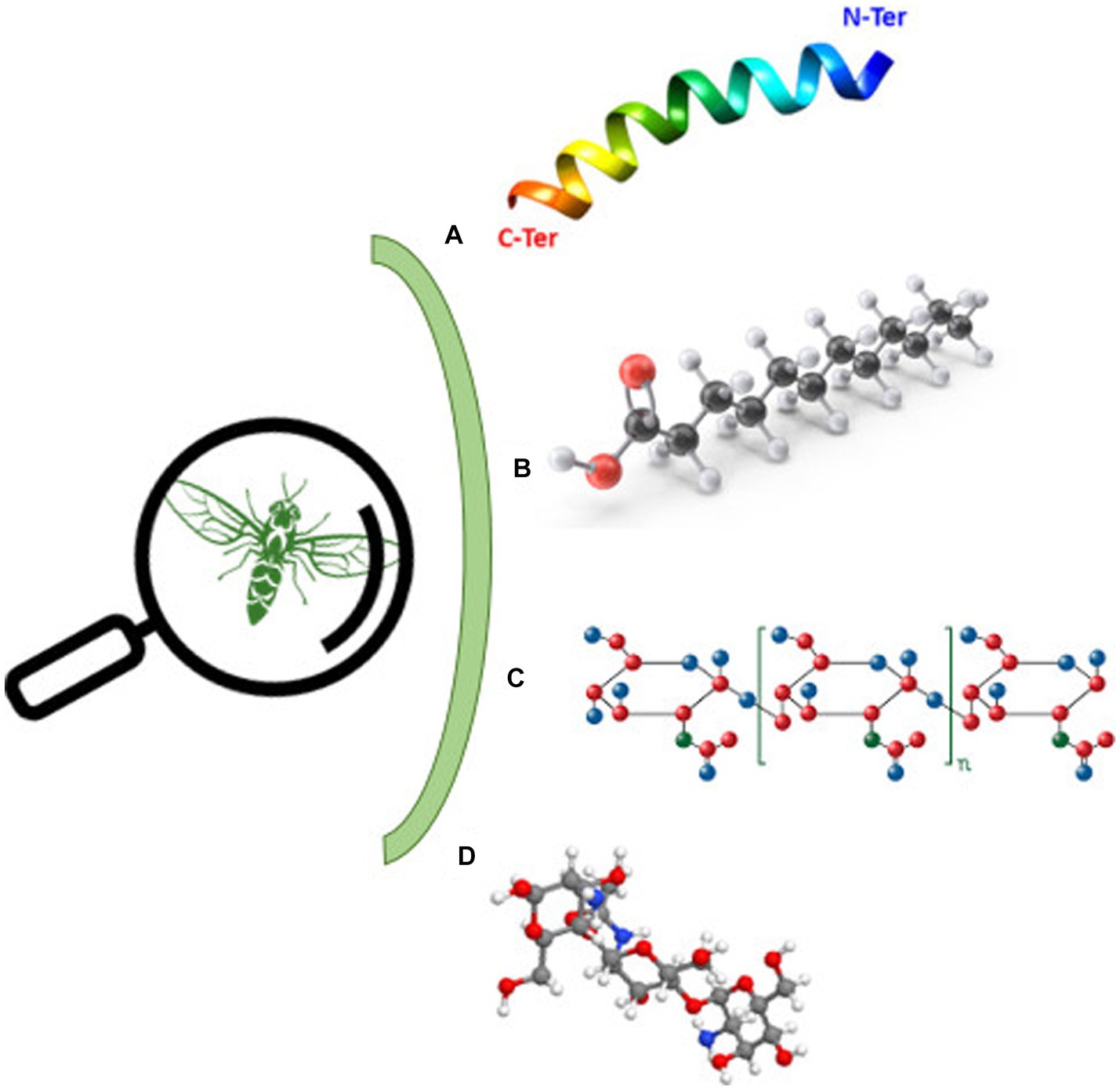
Figure 2. Bioactive molecules derived from insects. (A) Antimicrobial peptides, (B) lauric acid, (C) chitin, and (D) chitosan.
AMPs have received great attention as natural antibiotics that can likely confer protection against the development of bacterial resistance. Nisin is the most explored bacteriocin. In addition to its use as a preservative in several human foodstuffs, animal feed is supplemented with nisin as an anti-bacterial additive. In broilers, nisin was shown to reduce the number of Bacteroides and Enterobacteriaceae in ileal digesta (81, 82). Choi et al. (83, 84) assessed the effects of dietary administration of the antimicrobial peptide P5 (AMP-P5) and antimicrobial peptide-A3 (AMP-A3) in Ross 308 broilers. They concluded that supplementation with 60 mg AMP-P5/Kg or with 90 mg AMP-A3/Kg allowed for improved growth performances, nutrient retention, intestinal morphology, and reduced coliforms in intestines and excreta. They concluded that both of these AMPs can be used as potential antimicrobial growth promoters. In another study (85), Wen and He investigated the effect of the supplementation of synthetic cecropin (an insect-derived AMP) in broilers. The authors reported a decrease in the aerobic bacterial counts in jejunal and caecal digesta in a dose-dependent manner, as well as an improved height of the duodenal villus. Cecropin was then suggested as a possible alternative to antibiotic growth promoters in broiler production. Interestingly, the scientific literature implies that AMPs improve the intestinal health of the host animal by creating a microbial ecology, allowing it to deteriorate harmful microorganisms and favor the proliferation of beneficial ones. In this trend, Wang and his collaborators (86) described that the AMP sublancin significantly decreased the cecal count of Clostridium perfringens in broilers and enhanced the Lactobacilli count under lincomycin treatment (an antibiotic used to treat severe bacterial infections). These findings highlight the advantage of the use of insect-derived AMPs, as compared to traditional antibiotics, in the modulation of the gut microflora.
Insect-derived AMPs include α-helical peptides, cysteine-rich peptides, proline-rich (PR) peptides, and glycine-rich (GR) peptides. Most of them are cationic molecules. AMPs target the anionic phospholipids and phosphate groups of the lipopolysaccharides (LPS) of Gram-negative bacteria, and the peptidoglycan layer of Gram-positive bacteria (80). They bind to these specific sites and disturb the cell membrane of the pathogen by creating ion channels or transmembrane pores, leading to the killing of bacterial cells (87).
The α-helical peptides contain nine amino acid peptides in their α-helix region (LLCIALRKK) that are responsible for the antimicrobial activity (88). In a mouse model, acute intestinal inflammation caused by Clostridium difficile infection was treated by administrating an analog of the α-helical peptide. The treatment was shown to be efficient against inflammatory diarrhea associated with Clostridium difficile infection and Pseudomembranous colitis in mice (88).
Cysteine-rich peptides or insect defensins are cationic polypeptides, composed of 34 to 51 amino acids, containing multiple cysteine residues, and have three or four disulfide bridges (89). Cysteine-rich peptides are produced by fat body cells and hemolymph cells in insects. They essentially target Gram-positive bacteria (90). Some Cysteine-rich peptides were reported to be active against Gram-negative bacteria such as E. coli (91), as well as fungi (92), yeasts, and protozoa (93, 94). Some maggot-based medical preparations are commercially produced and used by professionals in wound disinfection and healing (95). Maggot secretions were reported for their antibacterial activity. They contain a defensin called lucifensin, which was shown to be active against Staphylococcus aureus, Staphylococcus carnosus, Streptococcus pyogenes, and Streptococcus pneumoniae (96).
Proline-rich peptides are cationic peptides composed of high proline residues, often associated with arginine residues in repeating motifs. PR peptides produced by insects are usually composed of 20 to 35 amino acid residues. They are mainly active against Gram-negative bacteria. They infiltrate the outer membrane and enter the periplasmic space where they impede the intracellular processes of the pathogen’s cell (97). They can also interfere with the synthesis of DNA and RNA by binding to nucleic acids (98, 99). PR peptides were described to exhibit antimicrobial effects in E. coli, Klebsiella pneumoniae, P. aeruginosa, and Acinetobacter baumannii (100, 101). Apidaecins are a group of PR peptides that destroys many Gram-negative bacteria, and hence, may be used as new candidates for peptide antibiotics (98).
Glycine-rich peptides include many molecules such as sarcotoxin IIA, hymenoptaecin, attacin, diptericin, and coleoptericin. They are rich in glycine residues (14 to 22%) and are active against fungi, Gram-negative bacteria (90), and cancer cells (102). The high content of glycine plays a key role in the tertiary structure of GR peptides and their mechanism of action against pathogens (103).
Lauric acid (LA, C12:0) is a saturated medium-chain fatty acid (MCFA) with a 12-carbon backbone. MCFAs were reported for their antimicrobial activity and suggested for preventing and treating gastrointestinal disease in piglets after weaning (104), as well as pig enteritis (105). MCFAs are also used directly by enterocytes to produce energy, thus they help to strengthen gut integrity in young piglets (106).
In Cobb 500 broiler chickens, supplementing 22.8 g of free LA per 100 g of total fatty acids allowed enhancing feed: gain ratio and breast meat yield (107). In male Ross 308 broiler chickens, LA significantly ameliorated BW and ADG (108). The intestinal mucosal barrier was enhanced and the immunoglobulins (IgA, IgM, and IgY) were upregulated. Regarding inflammatory cytokines, IL-1β, IL-6, TNF-α, IL-4, and IL-10 were downregulated (108). Concerning lipid metabolism, the levels of phosphatidylcholines were decreased, those of lysophosphatidylcholines were increased and the sphingolipid metabolism pathway was inhibited (108). Moreover, Phascolarctobacterium, the Christensenellaceae_R7 group, and Bacteroides were reduced, and Faecalibacterium and Ruminococcaceae UCG-014 were increased, indicating the ability of LA to significantly (p < 0.05) regulate cecal microbiota composition (108). These findings argue for the potential of LA as an additive in poultry feed and suggest a new alternative for antibiotics in poultry husbandry and a new way to ensure feed safety.
Lauric acid has been described to have antiviral and antibacterial activities (109). It is active mainly against Gram-positive bacteria, even in the presence of solid particles and at a pH greater than 6 (110). In animal models, C12:0 was effective in pigs with D-streptococcal infections (111). It is important to add that monolaurin, a monoester from lauric acid, can be formed, and has a biological bactericide activity greater than lauric acid (109).
Undissociated forms of MCFA can infiltrate the lipid membrane of bacteria, and move to the cytoplasm where they dissociate and decrease the pH. In an attempt to maintain its neutral pH, the bacterial cell exports many protons. Cellular ATP is then consumed, leading to energy depletion and bacterial cell death (112, 113).
Lauric acid was detected in high levels (up to 60%) in BSF prepupae reared in organic waste streams with high amounts of starch (114), but it is important to note that this level depends on the rearing substrate and the insect feed (115). In order to gain maximum benefit from the antimicrobial effects of lauric acid, it is recommended to feed whole larvae or prepupae to poultry. The utilization of less sustainable protein and fat sources could therefore be decreased.
Both chitin and chitosan are polysaccharides available in high amounts in insects. Chitosan, β-(1–4)-linked 2-amino-2-deoxy-β-D-glucopyranose, is an N-deacetylated derivative of chitin, and the main organic skeleton material of the exoskeleton of insects (116). In chitin, the acetamide groups are converted into primary amino groups (117).
Chitosan was reported to act as a chelating agent in biological systems and exhibited antimicrobial activity against bacteria, yeasts, and fungi (118). Indeed, it is considered a potential target for recognition by the immune system in many species, including birds (119). In other words, the innate immune system is stimulated by chitin and chitosan from insects (120, 121). In this context, a decreased albumin/globulin ratio was recorded in broiler chickens fed MW larvae, indicating an improved immune status (11, 122). Furthermore, improved immune activity was seen in laying hens fed BSF larvae meal-based diet with 1 g of chitin offered daily (123). In their trial, Islam and Yang (121) described decreased caecal E. coli and S. enteritidis contents in broilers fed 0.4% dry MW or Z. morio larvae, associated with an increase in IgG and IgA levels, which play a protective role against microbial infections. Interestingly, chitin is known for its hypolipidemic and hypocholesterolemic properties (124). In broiler chickens, chitin supplementation resulted in a decrease in body fat and possibly the production of leaner meat (125). Moreover, chitin and chitosan were recognized for their antiparasitic activity against L. major in mice (126) and intestinal infections with Eimeria papillata (127).
Chitin and chitosan exhibit their antimicrobial and antiparasitic effects directly or indirectly by supporting immune activity. The direct mechanism resembles that of AMPs and induces cell lysis, penetration of cytoplasmic membranes, and cation chelation (128). It is worth stating that chitosan was shown to be more effective in treating microbial infections, while chitin was more effective in treating parasitic infections (129), but they have in common their immunomodulatory effects. Immune cells can recognize both of these polymers thanks to their galactin-3, mannose receptor, RegIIIγ, dectin-1, and various toll-like receptors. These receptors have been shown to interact with chitin and/or chitosan (119).
Health benefits attributed to insects can create an additional value chain for poultry health and be antibiotic alternatives. The antimicrobial properties and the immuno-modulating effects of insect-derived bioactive molecules may contribute to enhancing global health, reducing the use of antibiotics, and avoiding antibiotic resistance when these insect products are included in poultry diets.
Insects are a sustainable, attractive, safe, and promising alternative protein source that could satisfy the world’s growing demand for food. This is especially true for poultry nutrition. Housefly larvae and pupae have been widely investigated in broilers (130–134). The different studies agreed that this protein source, either fresh or dried, can be used as an alternative to fish meal, soybean meal, and other protein ingredients. No adverse effects were registered regarding survival, daily FI, BWG, and FCR. It is worth stating that combining housefly meals and conventional meals in the same diet allowed for enhanced growth performances (130, 134). This may point toward a more balanced feed when comparing conventional and insect meals. Furthermore, housefly meal can partially or totally replace FM in broiler diets. Inaoka et al. (135) and Awoniyi et al. (130) reported that housefly maggots and pupae may be included in the diet up to 7%. If the inclusion rate exceeds 10%, a decrease in FI and growth performance was observed. This seems to be related to the dark color of the produced feed, which is not attractive to chickens (136, 137), and to the imbalanced AAs profile (5). In this latter case, methionine supplementation might improve broiler growth performance. Nevertheless, Hwangbo et al. (64) concluded that growth performance was enhanced when supplementing broiler feed with 10–15% housefly larvae. Similarly, Okah and Onwujiariri (133) described that the experimental group of chickens fed 20 and 30% of housefly maggot meal registered the best BW, as compared with the control group. It seems that this discordance regarding the inclusion rate is related to the processing method of the insects, which influences the availability of nutrients, as well as the inclusion rate of the insect meal.
Mealworm meal is also an alternative protein source that can be included in poultry feed without affecting the growth performance of the birds. In broilers, and without affecting feed intake, increasing levels of MW meal (from 0.1 to 0.3%) allowed significant improvement (p < 0.05) in both the BWG and the FCR from 1322.0 g to 1423.3 g, and from 1.88 to 1.75, respectively. The highest dressing percentage was recorded in the group supplemented with 0.3% MW (39). Interestingly, and according to this latter study, the gross return and the net profit were significantly (p < 0.05) higher for the group fed 0.3% MW (0.41 US$). The control group registered the lowest return (0.34 US$). The findings of this study were corroborated by Ramos-Elorduy et al. (53) who concluded that MW, once incorporated in the broiler diets up to 10%, has no negative effects on BWG, FI, and FCR. These authors noted a consequent decrease in the inclusion rate of SBM from 31 to 20% after incorporating dried yellow MW up to 10%, without any significant differences in the performance results. According to Schiavone et al. (138), the best BWG was obtained using 25% MW. Another assay aiming to fully substitute SBM by MW larvae in the broilers’ diet was conducted by Bovera and his collaborators (6). No significant effects on growth performance, carcass traits, and meat quality were observed. In addition, the FCR was significantly enhanced (p < 0.05) in the MW group. As compared to the control group (SBM group), broilers fed MW as a unique protein source had a more developed digestive system, a greater percentage of the spleen, longer intestines, ileum, and ceca (p < 0.05). Compared with an iso-proteic and isoenergetic SBM diet, MW larvae did not affect the FI of broilers. Indeed, a better FCR was calculated in the experimental group (11). Importantly, the lowest albumin/globulin ratio was recorded in broilers that received MW diets, which indicates a better disease resistance and immune response in birds. This finding may be attributed to the prebiotic effects of chitin and other bioactive molecules produced by the insect. Furthermore, the values of aspartate aminotransferase of the MW group were within the normal range for broilers and were not accompanied by an increase in gamma-glutamyl transferase, lactic dehydrogenase, or lactic dehydrogenase. Such results indicate that no alteration occurred in liver and muscle cells, and suggest that MW larvae can be used as a unique protein source in broiler diets.
BSF is another potent candidate to be included in poultry feed as SBM and FM substitute. The nutritional profile of these worms is comparable to that of FM and, in some aspects, better than that of SBM. Maggot meal could replace FM, without altering BWG, FI, and FCR. As a substitute for SBM, dried BSF larvae allowed for decreasing FI and enhancing FCR, without adverse effects on BWG (5). In growing broiler quails, no significant statistical differences were reported regarding BWG, FI, FCR, and mortality rate between control, 10% defatted BSF larvae meal, and 15% defatted BSF larvae meal groups (65). Regarding palatability, no special preference for broilers was shown toward control or defatted BSF larvae meals. It is interesting to mention that in this latter study, 10% defatted BSF larvae substituted for 28.4% of soybean oil and 16.1% of SBM, and 15% of defatted BSF larvae meal allowed the replacement of 100% of soybean oil and 24.8% of SBM. It can be concluded that BSF meal is a good alternative to FM and SBM in broiler diets, without any negative impacts on the growth performances of the birds.
Grasshoppers were also assessed in broilers’ feed. Grasshopper meal was used to replace 50 and 100% of FM (the inclusion rate was 5 and 10%, respectively) in Arbor Acres broiler chickens, which exhibited improved growth (29). These findings were corroborated by the findings of Sanusi et al. (139) and Sun et al. (140) who reported that grasshopper meal could totally replace FM without any adverse effects in Anak 2000 broiler chickens. However, diets containing more than 25% wild edible grasshopper (Ruspolia nitidual) meal were found to lower FI in indigenous chickens (141).
Earthworms were also recommended as a potential protein source that could replace FM and SBM in poultry feeding. Loh et al. (142) reported that incorporating EWM in Ross male broiler chickens diets at 5, 10, 15 and 20% allowed enhancing FI (p < 0.05), increasing BW (p < 0.05), improving FCR (p < 0.05) and reducing fecal lactic acid bacteria count. Other related studies confirmed that Ross 308 broiler chickens fed on diets containing 2, 3%, or 4% EWM increased their BW and FI (143–145). An inclusion rate of 5% EWM (Eudrilus eugeniae) enhanced FCR (87). Lower inclusion rates (1% EWM and 1% vermi-humus) resulted in a negative impact on BWG, despite the observed improvements in the immune functions in broilers (145). This was probably due to the unbalanced amino acid profile of the diet.
The effect of insect meal feed on the carcass quality of broilers is quite different. Although some studies did not disclose any significant impact, several authors reported enhancing effects. Hwangbo et al. (64) did not find significant differences in carcass traits of Ross broiler chickens when replacing SBM with HF larvae or pupae. The trial used 600 one-day-old male chicks and lasted 35 days. Similarly, Awonyi et al. (130) assessed the effect of substituting FM with maggot meal on 102 ANAK 3000-strain broiler chicks. The authors stated that the live, dressed, and eviscerated weights, as well as the relative length, breadth, and weights of the pectoral and gastrocnemius muscles, were not significantly affected by HF maggot meal diets. The dressing percentage was not affected as well. Nevertheless, Hwangbo et al. (64) described higher dressing percentages in broilers fed 5 to 20% HF meal. Likewise, Pieterse et al. (146) reported that unsexed Ross 308 broiler chickens fed HF meal had heavier carcasses and higher breast and thigh muscle weights than those fed SBM-based diet.
Several studies (147–149) have been conducted in order to investigate the effect of using MW on growth performance and carcass traits of male Shaver brown broiler chickens, female Label Hubbard hybrid chickens, and male Ross 708 broiler chickens. The authors concluded that using MW meal in broiler diets had no significant effects on carcass traits. However, Ballitoc and Sun (150) found that unsexed Ross broiler chickens fed on a diet supplemented with 2% MW meal over 35 days exhibited an increased carcass yield, slaughter weight, dressed weight, eviscerated weight, and a reduced abdominal fat weight. In the same way, Hussain et al. (39) reported that broiler chickens fed a diet containing 3% MW meal exhibited increased dressing percentage, feed cost, gross return, and net profit.
With regard to BSF meal, Cullere et al. (65) have found that both breast meat weight and yield did not differ among control and experimental groups in growing broiler quails when replacing FM with 10 and 15% BSF meal in broiler quails diets (a total of 450 10-day-old Coturnix coturnix japonica chicks were used in this study). Another similar finding was reported by Schiavone et al. (61) who showed that a diet containing 100% BSF fat as a substitute for soyabean oil did not affect the carcass quality of Ross 308 broiler chickens. Nonetheless, Altmann et al. (151) reported that Ross 308 male birds fed up to 15% BSF meal over 35 days had heavier carcasses compared to those fed an SBM-based diet. In another trial conducted on 256 one-day-old male broiler chicks for 35 days, it was reported that the diet containing 5% BSF meal resulted in a reduction of the abdominal fat percentage, and 10% BSF meal resulted in heavier carcass weight and greater breast percentage (152). Carcass composition was also improved in Hubbard S757 broilers fed 7.8% BSF meal in combination with 5.2% alfalfa meal (153).
Regarding silkworms, Khatun et al. (154) reported that increasing the levels of SW meal in replacement of SBM in poultry diet allowed increasing linearly the FCR, dressing percentage, and profitability in broilers. Ullah et al. (155) described that the FCR and the dressing percentage were not affected by increasing levels of SW (from 25 to 100%) in broilers. Hence, it can be concluded that SWM can be used to substitute FM or SBM without adverse effects on broiler performances, carcass traits and profitability. The literature explained that the improved broiler production traits could be related to the higher content of EAA in SWs, and to the growth prompting factors they contain (156), such as the ecdysteroid activity (a hormone involved in protein synthesis and tissue formation). However, this latter argument needs to be elucidated.
Live grasshoppers were also reported to improve live weight and carcass composition, and total lipids phospholipids, and anti-oxidative potential of the meat, of Qinjiaoma broiler chickens (85). In two other studies (56, 157), the effect of replacing FM with grasshopper meal (50 and 100%) on the carcass traits and the profitability of broiler chickens was assessed. The carcass analysis showed significant differences (p < 0.05) between the control and experimental groups, with the exception of breast, pancreas, proventriculus, heart, spleen, liver, lungs, and crop and chest weights. The highest profitability was calculated for the group fed 100% grasshopper meal, followed by the group that received 50% of this insect meal, followed by the control group. These literature reports imply that grasshoppers can substitute for FM without negative effects on carcass traits and economic return in broiler chickens. These findings may be attributed to the enhanced CP and EE values of the insect-based diets and their improved digestibility as well.
A general improvement in meat quality in broiler chicken was observed when feeding insect meal-based diets. Hwangbo et al. (64) described enhanced contents of lysine and tryptophan in the breast muscle, despite the unchanged content of CP. This may be due to the greater AA profile and the high protein digestibility (98.5%) of HF larvae meal. In the same context, Sun et al. (140) reported that a live grasshoppers-based diet improved the total lipids, phospholipids, and antioxidant potential of broiler chicken meat. In addition, a full replacement of FM by EW meal resulted in a reduced fat content in the breast and thigh meat of broiler chicken (43). The antioxidant capacity of the meat was improved using a diet containing 5% EW meal. An inclusion rate of 7% EW meal allowed for obtaining greater aroma, juiciness, residues, and flavor in Cobb 500 broiler meat (144). Similarly, Gholami et al. (143) concluded that feeding EW meal to Ross 308 broiler chickens increased breast meat yield, but had no effect on thigh and abdominal fat percent.
Cullere et al. (65) reported that BSF meal feed did not affect the breast meat weight of broiler quails, and the meat quality remained unchanged (61). In the same way, Leiber et al. (153) described an improved meat redness using BSF meal in Hubbard S757 broilers. Schiavone et al. (152) noted that feeding 5% BSF meal lowered the abdominal fat percentage, 10% BSF meal resulted in greater carcass weight and breast percentage, and 15% BSF meal resulted in an increased abdominal fat percentage, meat redness, meat protein percentage, MUFAs percentage in breast meat, and reduced breast meat PUFAs percentage. The diet containing 100% BSF fat generated an elevated content of saturated fatty acids (SFAs), and a reduced level of PUFAs in the breast meat. In fact, chickens assimilate unsaturated fatty acids (UFAs) better than SFA. UFAs form mixed micelles with monoglycerides and conjugated bile salts. They are transported to the surface of the intestinal mucosa where they are absorbed (158). PUFAs play a myriad of roles in the body and are precursors of cellular functional molecules (159). It was shown that BSF oil supplementation increased n-3 and n-6 fatty acids in the breast meat of chickens (62), without influencing the level of MUFAs (61).
Several studies agreed that insect meal feed did not have any negative effect on poultry products Hussain et al. (39) indicated that MWs, when used in broiler chicken feeding, do not affect the organoleptic properties of the meat (i.e., taste, tenderness, juiciness, flavor, and color). Similarly, SW meal does not impact the taste of poultry products (13) and does not cause a fishy taint in the meat (160). Furthermore, HF meal fed to Cobb 500 broiler chickens enhanced meat flavor, meat juiciness, meat aroma, meat tenderness, and meat desirability (27). The breast muscle yield and the water-holding capacity were improved as well, and the thawing loss and cooking loss were decreased (146). Regarding color, it is known that the pigments contained in the feed originate from the raw materials and ingredients used in feed formulation.
The pH value is an important parameter for the detection of meat defects such as pale, soft, and exudative meat (PSE). This anomaly occurs when the post-mortem pH value measured 15 min after slaughtering is lower than 5.6. Cullere et al. (65) reported a pH value of 5.67 using an HF meal-based diet, and Bovera et al. (147) measured higher pH values in the poultry group fed insect meal-based feed. Pieterse et al. (161) concluded that there are no differences regarding the initial and ultimate pH of the thigh muscles between broiler chickens fed a control diet and those fed a BSF meal-based diet. It can be concluded that feeding insect meal-based diets to broiler chickens does not cause abnormal pH values, and consequently are not responsible for causing meat defects.
It seems that insect meals used in broiler chickens have the potential to produce meat with comparable chemical and quality traits compared to those fed diets containing traditional feed ingredients.
In addition to broilers, BSF was also investigated in laying hens as a substitute for SBM and FM. The inclusion rates varied from 5 to 100% (10, 13, 162, 163). In the assay carried out by Maurer et al. (164), SBM was partially (50%) and completely (100%) replaced by dried and defatted BSF larvae meal in a laying hens diet. No significant differences were noted between the control and the experimental groups with regard to egg production, FI, egg weight, and feed efficiency. Although there was a trend (p = 0.06) for lower albumen weight in the 24% BSF group, there were no differences in yolk and shell weights. Kawasaki and his collaborators (165) investigated the effect of BSF larvae and pupae meals on the egg quality of laying hens. They reported that the highest weights of egg and albumen were recorded in the poultry group fed BSF pupae. The eggshell thickness remained unchanged between these two experimental groups. Compared to the control group, feeding BSF meal improved the eggshell thickness, probably due to its high content of available calcium (163, 166). In another study, laying hens receiving a diet containing 10% BSFM have shown an increased egg weight, albumen weight, eggshell thickness, albumen height, and plasma calcium. The egg yolk color score was enhanced as well (165). An inclusion rate of 3% BSFM in Xuefeng black-bone laying hens improved egg weight; Haugh unit; eggshell weight; yolk C14:00, C17:00, and C20:2 fatty acids; yolk amino acids (glutamic acid, methionine, phenylalanine, and leucine), plasma total superoxide dismutase; and plasma avian influenza virus antibody. However, eggshell thickness and plasma interleukin-2 decreased (23). It is also important to note that feeding Lohman Brown Classic laying hens a diet with 17% BSFM to substitute for SBM resulted in poor growth and production percentage, as well as a reduction in blood lipids, blood chloride, and blood creatinine. However, an increase in the percentage of small, medium, and extra-large size eggs, blood globulin, blood calcium (123), and fecal dry matter (164) was noted.
HF maggots and pupae were assessed in rearing layers (167, 168). In Isa brown and Nera black layer hens, HF larvae meal was used as a 50% (7.08% HF meal and 1.50% FM) and total (9.44% HF meal and 0% FM) replacement of FM. This study showed that the experimental diets did not affect feed intake (FI) and feed conversion ratio (FCR) but significantly (p < 0.05) improved hen-day production, from 3.00% for the FM group to 4.72% for the insect meal group (168). This result seems to be attributed to the enhanced AA profile of the diet when HF larvae meal and FM were supplied together. No differences between the control and the experimental diets were reported by the authors regarding the egg quality traits. Nevertheless, the shell thickness weight was decreased in the experimental groups (168). This reduction may be explained by the lower calcium content of HF larvae meals. Moreover, Dankwa et al. (167) found that supplementation with 30–50 g of maggots results in a significant (p < 0.05) improvement in clutch size, number of hatched eggs, and egg and chick weight. The results of the above studies suggest that HF meal could be used as a partial replacement for FM in laying hens’ feed.
Grasshoppers (Ornithacris cavroisi), silkworms, bee propolis, and pollen were also investigated in laying hens’ diets. An enhanced Haugh unit in Isa Brown laying hens was observed when feeding 25% grasshopper meal as a substitution for FM. The egg yolk color was improved using an incorporation rate of 75% grasshopper meal (29). White Leghorn layer hens fed 5.6% SW meal as a replacement for SBM showed a significant (p < 0.05) improvement in live weight, FCR, and egg production, with a significant (p < 0.05) reduction in FI and feed cost (169). An inclusion rate of 8% SWM was reported to improve survivability (169). Bee propolis was included at 0.025 and 0.05% in Lohmann LSL laying hens’ diets. The results showed an increase in egg mass and production, Haugh unit, albumen height, yolk height, index and weight, blood total protein, globulin, hemoglobin, and lymphocytes. A simultaneous decrease of FCR, yolk diameter, blood cholesterol, heterophil, and lymphocyte ratio was reported (170). Bee pollen was supplemented to Sinai laying hens at 0.05 and 0.15%. A subsequent increase in the egg number and mass, production percentage, feed intake, red blood cells, white blood cells, and lymphocytes was described. But BW, BWG, heterophils, heterophil lymphocyte ratio, blood cholesterol, and blood triglycerides were reduced (171).
Food security arguments for switching to an alternative protein source are powerful motivators for using insects in poultry diets, especially since scientific reports have approved insect meals as a valuable and sustainable protein source that is able to meet the production requirements of these birds. However, the nutritive value of insects varies throughout the production chain, from the insect-rearing step to the feed-manufacturing phase.
Insect farming conditions have critical effects on the nutritional value of the insect larvae. In fact, larval density as well as the quantity and the quality of the insect diet determine the body composition of the larvae. Barragan-Fonseca et al. (172) investigated the effect of dietary nutrient concentration and larval rearing density on the growth performance and the chemical composition of BSF larvae. Four (04) rearing densities (50, 100, 200, or 400 larvae per container) and three nutrient concentrations (low, medium, and high) were tested. The authors reported that individual larval weight and total larval yield increased in all groups that received high nutrient concentration, in spite of the larval density. Larval crude fat content was higher in groups with low rearing density and received high nutrient concentrations. Larval crude protein was higher in groups at low rearing density and low nutrient concentration. This study revealed that larval CP content is affected by nutrient concentration, rearing density, as well as their interaction. Indeed, this study suggested that larval CP content is regulated within narrow limits, and larval crude fat depends on the nutrient concentration and the larval density. In fact, it was stated that high-fat and high-carbohydrate diets increase larval crude fat content (173) compared to low-fat and/or high-fiber meals which decrease the larval crude fat content (174, 175). These findings highlight the importance of rearing practices and insect diets in producing larvae with valuable nutritive profiles.
In animal feed, insects are usually added as a meal. Insects are first dehydrated or roasted and then ground into a fine powder, the meal (176). Some popular processing methods include steaming, boiling, frying, smoking, drying, and toasting (177). Although a significant reduction of microbial hazards can be ensured through thermal treatments, the nutritive value of the insects can be affected. Nyangena et al. (178) demonstrated that all thermal treatments (boiling, toasting, oven-drying, and their combinations) except solar-drying lowered bacterial counts and eliminated yeast and molds in BSF prepupae. Meanwhile, these heat treatments decreased the crude fat content up to 14.3–28.2 g/Kg DM in the order: toasting > boiling > oven-drying > solar-drying. They also increased the CP content up to 37–41.3 g/Kg DM in the same order and improved the available carbohydrates. Another study conducted by Dobermann et al. (179) examined the effect of freeze-drying and heat processing at low and high temperatures (45°C and 120°C) on the nutritional profile of the black cricket Gryllus bimaculatus. The results showed that drying at 45°C obtained high protein and calcium contents. Moreover, freeze-drying conserved long-chain polyunsaturated fatty acids, more than drying at 120°C. Consequently, more attention is needed in order to protect the nutritive value of insects during heat processing.
As in the case of soy and whey protein concentrates and isolates, developing insect protein powders involves defatting, protein solubilization and recovery, purification, and drying (180). The methods used for chitin separation include enzymatic proteolysis, thermal treatments, solvent extractions, or alkali/acidic reactions. Free amino acids and peptides of different sizes are then produced. These processes may modify the insect proteins and affect their functionalities through structural modifications, molecular weight decreases, or polarity increases (181). Longer proteolysis and a high enzyme concentration result in lower molecular weight peptides with affected techno-functional properties. Also, the selection of the enzyme protease will determine the peptide amino acid sequence and/or amino acid residues which are responsible for the nutritional properties of the protein hydrolysates (182).
Fat extraction is a critical step to produce insect proteins and to obtain a high yield of good quality insect fat. Conventional extraction methods include chemical extraction using ethanol and methanol as extraction solvents, mechanical oil press, and three-phase partitioning (TPP). Non-conventional extraction techniques involve supercritical CO2. Laroche et al. (183) compared the effects of conventional solvents, TPP, and supercritical CO2 on the fat extraction yield and the fatty acid profiles of cricket and mealworm meals. They reported that supercritical CO2 was efficient only in cricket meal, whereas ethanol extraction and TPP increased the fat yield from both meals. Regardless, the defatting method should be selected with care.
Producing safe insects as an alternative protein source in poultry feeding is a challenge. The safety of edible insects always needs to be guaranteed. Contamination is very likely to occur and to be transferred to the animals unless safety rules are not ensured during the industrial rearing cycle. Insects for food have been associated with Staphylococcus aureus, pathogenic Clostridium spp., and pathogenic species of the Bacillus cereus group. When regarding insects for feed, microbes from the substrate can enter the gut, proliferate and become part of the gut microbiota (184, 185). The bacterial species vary depending on the region of the gut they colonize since each region has its distinct properties, and represents, therefore, a different ecological niche (184). Regarding the BSF, a set of insect species were screened, including Actinomyces sp., Dysgonomonas sp., Enterococcus sp., and Morganella sp. It is important to note that these species have been tracked in other insects as well (186). Regarding industrially produced HF, there are no studies that have been performed to investigate its microbiota to our current knowledge. Little is known about other types of biological contaminants, fungi, viruses, protozoa, and prions.
Substrates and feed for insects can be adulterated with heavy metals, pesticides, fungi, and bacteria. Some studies reported potential risks in BSF larvae. Wynants et al. (187) observed contamination with Salmonella enterica serovar Agona in the rearing residue. Wu et al. (188) and Jiang et al. (185) reported an infestation by Campylobacter and Clostridium species, respectively. Presumptive B. cereus was revealed in the BSF larvae with counts up to 6,000 cfu/g. With respect to viruses, Chen et al. (189) isolated an Escherichia phage from the BSF larvae gut. Indeed, prions can infest non-processed insects from contaminated substrates, even insects were shown to be not able to produce them (190).
Mycotoxins from feed or rearing substrates are another potential risk that can affect growth and increase the mortality of insects. Consuming a mycotoxin-contaminated insect meal can affect poultry health and performance. Schrogel and Wätjen (191) found that the mycotoxin content of the substrate was 25-fold higher than the maximum limit. Therefore, they recommended starving insects for 24 h before harvesting. In addition, insects may accumulate not only residues of pesticides, veterinary drugs, and hormones but also dioxins and PCBs as well (192).
Another potential risk is the contamination of feed and/or substrate with heavy metals (e.g., cadmium, arsenic, cobalt, copper, nickel, etc.). In fact, Vijver et al. (193) proved the accumulation in BSF larvae of cadmium, copper, lead, and zinc from the soil. Schrogel and Wätjen (191) stated that HF is able to concentrate cadmium, whereas BSF is able to accumulate arsenic. Insect larvae are able to accumulate cadmium or plumbum in their exoskeleton (194).
The presence of pathogenic bacteria in insects can be avoided through farming or processing conditions (195). Rearing insects on pollutant-free substrate is key for preventing insect and animal contamination.
Insects can fulfill a major role in upgrading the value of poultry feed, coping with competing uses of food system resources, and contributing to food safety. Insect meal represents an alternative protein source enriched with highly digestible essential amino acids, fat, and other important nutrients such as MUFA, PUFA, calcium, phosphorous, copper, iron, magnesium, manganese, selenium, and zinc, in addition to riboflavin, pantothenic acid, biotin, and, in some cases, folic acid. However, the quality of insect meals and their nutrient profile depends on the rearing medium, insect feed, and processing methods. Insect farming is able to provide an alternative source of income for small-scale farmers and large-scale industries. Nevertheless, the cost of production needs to be reduced. Also, the risk of contamination and the spreading of pathogens needs to be kept under control. Beyond their nutritional value, insects produce bioactive compounds that could act as health stimulators in livestock. Hence, insects may be used as antibiotic alternatives because of the biological properties of their AMPs and their synergic effects with fatty acids and either chitin or chitosan. They may be also used in the future to combat antibiotic resistance. With the unceasing reliable advancements in scientific research in this field, insect protein is being touted as the answer to a consistent and tenable feed group, and more importantly, it is predicted to be the number-one sustainable protein source for the poultry feed industry in future generations.
IBS and HY: conceptualization. IBS, TN, and NM’H: validation. IBS, HY, and MBL: resources and writing–original draft preparation. IBS and NM’H: writing–review and editing. TN and NM: supervision. All authors contributed to the article and approved the submitted version.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Breewood, H., and Garnett, T. What is feed-food competition? (Foodsource: Building blocks). Food Climate Research Network, University of Oxford. (2020).
2. Mottet, A, de Haan, C, Falcucci, A, Tempio, G, Opio, C, and Gerber, P. Livestock: on our plates or eating at our table? A new analysis of the feed/food debate. Glob Food Secur. (2017) 14:1–8. doi: 10.1016/j.gfs.2017.01.001
3. Alexander, P, Brown, C, Arneth, A, Finnigan, J, Moran, D, and Rounsevell, MDA. Losses, inefficiencies and waste in the global food system. Agric Syst. (2017) 153:190–200. doi: 10.1016/j.agsy.2017.01.014
4. Turck, D, Castenmiller, J, De Henauw, S, Hirsch-Ernst, KI, Kearney, J, Maciuk, A, et al. Scientific opinion on the safety of dried yellow mealworm (Tenebrio molitor larva) as a novel food pursuant to regulation (EU) 2015/2283. EFSA J. (2021) 19:6343. doi: 10.2903/j.efsa.2021.6343
5. Makkar, HP, Tran, G, Heuzé, V, and Ankers, P. State-of-the art on use of insects as animal feed. Anim Feed Sci Technol. (2014) 197:1–33. doi: 10.1016/j.anifeedsci.2014.07.008
6. Bovera, F, Loponte, R, Marono, S, Piccolo, G, Parisi, G, Iaconisi, V, et al. Use of Tenebrio molitor larvae meal as protein source in broiler diet: effect on growth performance, nutrient digestibility, and carcass and meat traits. J Anim Sci. (2015) 94:639–47. doi: 10.2527/jas.2015-9201
7. Abd-El-Hack, A, Mohamed, E, Shafi, ME, Alghamdi, WY, Abdelnour, SA, Shehata, AM, et al. Black soldier fly (Hermetia Illucens) meal asa promising feed ingredient for poultry: A comprehensive review. Agriculture. (2020) 10:339. doi: 10.3390/agriculture10080339
8. Sayed, WA, Ibrahim, NS, Hatab, MH, Zhu, F, and Rumpold, BA. Comparative study of the use of insect meal from Spodoptera Littoralis and Bactrocera Zonata for feeding Japanese quail chicks. Animals. (2019) 9:136. doi: 10.3390/ani9040136
9. Secci, G, Bovera, F, Nizza, S, Baronti, N, Gasco, L, Conte, G, et al. Quality of eggs from Lohmann Brown classic laying hens fed black soldier fly meal as substitute for soya bean. Animal. (2018) 12:2191–7. doi: 10.1017/S1751731117003603
10. Secci, G, Bovera, F, Parisi, G, and Moniello, G. Quality of eggs and albumen technological properties as affected by Hermetia Illucens larvae meal in hens’ diet and hen age. Animals. (2020) 10:81. doi: 10.3390/ani10010081
11. Bovera, F, Piccolo, G, Gasco, L, Marono, S, Loponte, R, Vassalotti, G, et al. Yellow mealworm larvae (Tenebrio molitor, L.) as a possible alternative to soybean meal in broiler diets. Br Poult Sci. (2015) 56:569–75. doi: 10.1080/00071668.2015.1080815
12. Marono, S, Piccolo, G, Loponte, R, Di Meo, C, Attia, YA, Nizza, A, et al. In vitro crude protein digestibility of Tenebrio Molitor and Hermetia Illucens insect meals and its correlation with chemical composition traits. Ital J Anim Sci. (2015) 14:3889. doi: 10.4081/ijas.2015.3889
13. Khan, SH. Recent advances in role of insects as alternative protein source in poultry nutrition. J App Anim Res. (2018) 46:1144–57. doi: 10.1080/09712119.2018.1474743
14. Oonincx, DGAB, and de Boer, IJM. Environmental impact of the production of mealworms as a protein source for humans–A life cycle assessment. PLoS One. (2012) 7:51145. doi: 10.1371/journal.pone.0051145
15. Imathiu, S. Benefits and food safety concerns associated with consumption of edible insects. Benefits and food safety concerns associated with consumption of edible insects. NFS J. (2020) 18:1–11. doi: 10.1016/j.nfs.2019.11.002
16. Van Huis, A., Klunder, JVI, Merten, E., Halloran, A., and Vantomme, P. (2013). Future prospects for food and feed security. Food and agriculture organisation of the United Nations (FAO). Rome, Italy: Edible Insects, FAO Forestry Papers.
17. Chu, X, Li, M, Wang, G, Wang, K, Shang, R, Wang, Z, et al. Evaluation of the low inclusion of full-fatted hermetia illucens larvae meal for layer chickens: growth performance, nutrient digestibility, and gut health. Front Vet Sci. (2020) 7:585843. doi: 10.3389/fvets.2020.585843
18. Neumann, C, Velten, S, and Liebert, F. N balance studies emphasize the superior protein quality of pig diets at high inclusion level of algae meal (Spirulina platensis) or insect meal (Hermetia illucens) when adequate amino acid supplementation is ensured. Animals. (2018) 8:1–14. doi: 10.3390/ani8100172
19. Neumann, C, Velten, S, and Liebert, F. Improving the dietary protein quality by amino acid fortification with a high inclusion level of micro algae (Spirulina platensis) or insect meal (Hermetia illucens) in meat type chicken diets. Open J Anim Sci. (2018) 8:12–26. doi: 10.4236/ojas.2018.81002
20. Neumann, C, Velten, S, and Liebert, F. The graded inclusion of algae (Spirulina platensis) or insect (Hermetia illucens) meal as a soybean meal substitute in meat type chicken diets impacts on growth, nutrient deposition and dietary protein quality depending on the extent of amino acid supplementation. Open J Anim Sci. (2018) 8:163–83. doi: 10.4236/ojas.2018.82012
21. Brede, A, Wecke, C, and Liebert, F. Does the optimal dietary methionine to cysteine ratio in diets for growing chickens respond to high inclusion rates of insect meal from Hermetia illucens? Animals. (2018) 8:187. doi: 10.3390/ani8110187
22. Mwaniki, Z, Neijat, M, and Kiarie, E. Egg production and quality responses of adding up to 7.5% defatted black soldier fly larvae meal in a corn–soybean meal diet fed to shaver white leghorns from wk 19 to 27 of age. Poult Sci. (2018) 97:2829–35. doi: 10.3382/ps/pey118
23. Liu, X, Liu, X, Yao, Y, Qu, X, Chen, J, Xie, K, et al. Effects of different levels of Hermetia illucens larvae meal on performance, egg quality, yolk fatty acid composition and oxidative status of laying hens. Ital J Anim Sci. (2021) 20:256–66. doi: 10.1080/1828051X.2021.1878946
24. Đorđević, M, Radenković-Damnjanović, B, Vučinić, M, Baltić, R, Teodorović, L, Janković, M, et al. Effects of substitution of fish meal with fresh and dehydrated larvae of the house fly (Musca domestica L) on productive performance and health of broilers. Acta Vet Brno. (2008) 58:357–68. doi: 10.2298/AVB0804357D
25. Dillak, SYFG, Suryatni, NPF, Handayani, HT, Temu, ST, Nastiti, HP, Osa, DB, et al. The effect of fed maggot meal as a supplement in the commercial diets on the performance of finisher broiler chickens. IOP Conf Ser Earth Environ Sci. (2019) 260:012056. doi: 10.1088/1755-1315/260/1/012056
26. Elahi, U, Ma, Y, Wu, S, Wang, J, Zhang, H, and Qi, G. Growth performance, carcass characteristics, meat quality and serum profile of broiler chicks fed on housefly maggot meal as a replacement of soybean meal. J Anim Physiol Anim Nutr. (2020) 104:1075–84. doi: 10.1111/jpn.13265
27. Khan, S, Khan, RU, Sultan, A, Khan, M, Hayat, SU, and Shahid, MS. Evaluating the suitability of maggot meal as a partial substitute of soya bean on the productive traits, digestibility indices and organoleptic properties of broiler meat. J Anim Physiol Anim Nutr. (2016) 100:649–56. doi: 10.1111/jpn.12419
28. Hussein, M, Pillai, VV, Goddard, JM, Park, HG, Kothapalli, KS, Ross, DA, et al. Sustainable production of housefly (Musca domestica) larvae as a protein-rich feed ingredient by utilizing cattle manure. PLoS One. (2017) 12:e0171708. doi: 10.1371/journal.pone.0171708
29. Brah, N, Issa, S, and Houndonougbo, F. Effect of grasshopper meal on laying hens’ performance and eggs quality characteristics. Indian J Anim Sci. (2017) 87:1005–10. doi: 10.56093/ijans.v87i8.73520
30. Wang, D, Zhai, S-W, Zhang, CX, Zhang, Q, and Chena, H. Nutrition value of the Chinese grasshopper Acrida cinerea (Thunberg) for broilers. Anim Feed Sci Technol. (2007) 135:66–74. doi: 10.1016/j.anifeedsci.2006.05.013
31. Ghosh, S, Haldar, P, and Mandal, D. Evaluation of nutrient quality of a short horned grasshopper, Oxya hyla hyla Serville (Orthoptera: Acrididae) in search of new protein source. J Entomol Zool Stud. (2016) 4:193–7.
32. Amobi, MI, Saleh, A, Okpoko, VO, and Abdullahi, AM. Growth performance of broiler chickens based on grasshopper meal inclusions in feed formulation. Fortschr Zool. (2020) 18:39–43. doi: 10.4314/tzool.v18i1.7
33. Kinyuru, J, Kenji, GM, Muhoh, SN, and Ayieko, M. Nutritional potential of longhorn grasshopper (Ruspolia Differens) consumed in Siaya District, Kenya. J Agric Sci Technol. (2009) 2009:32–46.
34. Khan, S, Khan, RU, Alam, W, and Sultan, A. Evaluating the nutritive profile of three insect meals and their effects to replace soya bean in broiler diet. J Anim Physiol Anim Nutr. (2018) 102:e662–8. doi: 10.1111/jpn.12809
35. Jintasataporn, O. Production performance of broiler chickens fed with silkworm pupa (Bombyx mori). J Agric Sci Technol Iran. (2012) 2:505–10.
36. Acay, RP. Silkworm pupa meal as feed supplement for growing-finishing broilers. Benguet, Philippines: Benguet State University (2011).
37. Rangacharyulu, PV, Giri, SS, Paul, BN, Yashoda, KP, Rao, RJ, Mahendrakar, NS, et al. Utilization of fermented silkworm pupae silage in feed for carps. Bioresour Technol. (2003) 86:29–32. doi: 10.1016/S0960-8524(02)00113-X
38. Lakshminarayana, T, and Thirumala Rao, SD Silkworm pupae, silkworm meal. World feeds and Protein News, 3, p. 2. (1971).
39. Hussain, I, Sarzamin, K, Asad, S, Naila, C, Rafiullah, K, Waqas, A, et al. Mealworm (Tenebrio molitor) as potential alternative source of protein supplementation in broiler. Int J Biol Sci. (2017) 10:255–62. doi: 10.12692/ijb/10.4.255-262
40. Liu, C, Masri, J, Perez, V, Maya, C, and Zhao, J. Growth performance and nutrient composition of mealworms (Tenebrio Molitor) fed on fresh plant materials-supplemented diets. FoodReview. (2020) 9:151. doi: 10.3390/foods9020151
41. Ravzanaadii, N, Kim, SH, Choi, WH, Hong, SJ, and Kim, NJ. Nutritional value of mealworm, Tenebrio molitor as food source. Int J Indust Entomol. (2012) 25:93–8. doi: 10.7852/IJIE.2012.25.1.093
42. Janković, LJ, Petrujkić, B, Aleksić, N, Vučinić, M, Teodorović, R, Karabasil, N, et al. Carcass characteristics and meat quality of broilers fed on earthworm (Lumbricus rubellus) meal. J Hellenic Vet Med Soc. (2020) 71:2031–40. doi: 10.12681/jhvms.22953
43. Istiqomah, L, Sofyan, A, Damayanti, E, and Julendra, H. Amino acid profile of earthworm and earthworm meal (Lumbricus Rubellus) for animal feedstuff. J Indones Trop Anim Agric. (2009) 34:253–7. doi: 10.14710/jitaa.34.4.253-257
44. Sun, Z, and Jiang, H. Nutritive evaluation of earthworms as human food. Future Foods. (2016) 2016:70271. doi: 10.5772/intechopen.70271
45. Finke, MD. Complete nutrient composition of commercially raised invertebrates used as food for insectivores. Zoo Biol. (2002) 21:269–85. doi: 10.1002/zoo.10031
46. Heuzé, V, Tran, G, Sauvant, D, Bastianelli, D, and Lebas, F. Earthworm meal. Feedipedia, a programme by INRAE, CIRAD, AFZ and FAO. (2020). Available at: https://feedipedia.org/node/665 (Accessed November 3, 2020).
47. Amer, AA, El-Nabawy, M, Els, M, Gouda, AH, and Dawood, MAO. The addition of insect meal from Spodoptera littoralis in the diets of Nile tilapia and its effect on growth rates, digestive enzyme activity and health status. Aquac Res. (2021) 52:5585–94. doi: 10.1111/are.15434
48. Moula, N, Scippo, ML, Douny, C, Degand, G, Dawans, E, Cabaraux, JF, et al. Performances of local poultry breed fed black soldier fly larvae reared on horse manure. Anim Nutr. (2018) 4:73–8. doi: 10.1016/j.aninu.2017.10.002
49. De Marco, M, Martínez, S, Hernandez, F, Madrid, J, Gai, F, Rotolo, L, et al. Nutritional value of two insect larval meals (Tenebrio molitor and Hermetia illucens) for broiler chickens: apparent nutrient digestibility, apparent ileal amino acid digestibility and apparent metabolizable energy. Anim Feed Sci Technol. (2015) 209:211–8. doi: 10.1016/j.anifeedsci.2015.08.006
50. Astuti, DA, Damanik, RH, Anggraeny, A, and Aidismen, YDP. Utilization of insect as a protein alternative for goat rations. Proceeding of the 4th international Asian-Australasian dairy goat conference. Vietnam 17-19 October 2018. (2018).
51. Aniebo, AO, Erondu, ES, and Owen, OJ. Proximate composition of housefly larvae (Musca domestica) meal generated from mixture of cattle blood and wheat bran. Livest Res Rural Dev. (2008) 20:205
52. Aniebo, AO, and Owen, OJ. Effects of age and method of drying on the proximate composition of housefly larvae (Musca domestica Linnaeus) meal (HFLM). Pak J Nutr. (2010) 9:485–7. doi: 10.3923/pjn.2010.485.487
53. Hall, HN, Masey O’Neill, HV, Scholey, D, Burton, E, Dickinson, M, and Fitches, EC. Amino acid digestibility of larval meal (Musca domestica) for broiler chickens. Poult Sci. (2018) 97:1290–7. doi: 10.3382/ps/pex433
54. Józefiak, D, and Engberg, RM. Insects as poultry feed. 20th European symposium on poultry nutrition (august). Prague, Czech Republic. (2015).
55. Ojewola, GS, Okoye, FC, and Ukoha, OA. Comparative utilization of three animal protein sources by broiler chickens. Int J Poult Sci. (2005) 4:462–7. doi: 10.3923/ijps.2005.462.467
56. Hassan, AA, Sani, I, Maiangwa, MW, and Rahman, SA. The effect of replacing graded levels of fishmeal with grasshopper meal in broiler starter diet. PAT. (2009) 5:30–8.
57. Ojewola, GS, and Udom, SF. Chemical evaluation of the nutrient composition of some unconventional animal protein sources. Int J Poult Sci. (2005) 4:745–7. doi: 10.3923/ijps.2005.745.747
58. Sun, T, Long, RJ, Liu, ZY, Ding, WR, and Zhang, Y. Aspects of lipid oxidation of meat from free-range broilers consuming a diet containing grasshoppers on alpine steppe of the Tibetan plateau. Poult Sci. (2012) 91:224–31. doi: 10.3382/ps.2011-01598
59. Istiqomah, L, Sakti, AA, Suryani, AE, Karimy, MF, Anggraeni, AS, and Herdian, H. Effect of feed supplement containing earthworm meal (Lumbricus rubellus) on production performance of quail (Coturnix coturnix japonica). IOP Conf Ser Earth Environ Sci. (2017) 2017:012032. doi: 10.1088/1755-1315/101/1/012032
60. Sun, ZJ, Liu, XC, Sun, LH, and Chunyang, S. Earthworm as a potential protein resource. Ecol Food Nutr. (1997) 36:221–36.
61. Schiavone, A, Cullere, M, De Marco, M, Meneguz, M, Biasato, I, Bergagna, S, et al. Partial or total replacement of soybean oil by black soldier fly larvae (Hermetia illucens L.) fat in broiler diets: effect on growth performances, feed-choice, blood traits, carcass characteristics and meat quality. Ital J Anim Sci. (2017) 16:93–100. doi: 10.1080/1828051X.2016.1249968
62. Benzertiha, A, Kierończyk, B, Rawski, M, Kołodziejski, P, Bryszak, M, and Józefiak, D. Insect oil as an alternative to palm oil and poultry fat in broiler chicken nutrition. Animals. (2019) 9:116. doi: 10.3390/ani9030116
63. Newton, GL, Booram, CV, Barker, RW, and Hale, OM. Dried Hermetia illucens larvae meal as a supplement for swine. J Anim Sci. (1977) 44:395–400. doi: 10.2527/jas1977.443395x
64. Hwangbo, J, Hong, EC, Jang, A, Kang, HK, Oh, JS, Kim, BW, et al. Utilization of house fly-maggots, a feed supplement in the production of broiler chickens. J Envir Biol. (2009) 30:609–14.
65. Cullere, M, Tasoniero, G, Giaccone, V, Miotti-Scapin, R, Claeys, E, DeSmet, S, et al. Black soldier fly as dietary protein source for broiler quails: apparent digestibility, excreta microbial load, feed choice, performance, carcass and meat traits. Animal. (2016) 10:1923–30. doi: 10.1017/S1751731116001270
66. Pieterse, E, and Pretorius, Q. Nutritional evaluation of dried larvae and pupae meal of the housefly (Musca domestica) using chemical and broiler-based biological assays. Anim Prod Sci. (2013) 54:347–55.
67. Bosch, G, Zhang, S, Oonincx, DG, and Hendriks, WH. Protein quality of insects as potential ingredients for dog and cat foods. J Nutr Sci. (2014) 3:1–4. doi: 10.1017/jns.2014.23
68. Wenk, C. The role of dietary fiber in the digestive physiology of the pig. Anim Feed Sci Technol. (2001) 90:21–33. doi: 10.1016/S0377-8401(01)00194-8
69. Zuidhof, MJ, Molnar, CL, Morley, FM, Wray, TL, Robinson, FE, Khan, BA, et al. Nutritive value of house fly (Musca domestica) larvae as a feed supplement for Turkey poults. Anim Feed Sci Technol. (2003) 105:225–30. doi: 10.1016/S0377-8401(03)00004-X
70. Pretorius, Q. The evaluation of larvae of Musca domestica (common fly) as protein source for broiler production. Thesis presented in partial fulfilment of the requirements for the degree of master of science in agriculture (animal sciences) at Stellenbosch University, p. 95. (2011).
71. Lemme, A, Ravindran, V, and Bryden, WL. Ileal digestibility of amino acids in feed ingredients for broilers. World Poult Sci J. (2004) 60:421–35. doi: 10.1079/WPS200426
72. Kim, EJ, Utterback, PL, and Parsons, CM. Comparison of amino acid digestibility coefficients for soyabean meal, canola meal, fish meal, and meat and bone meal among 3 different bioassays. Poult Sci. (2012) 91:1350–5. doi: 10.3382/ps.2011-01861
73. Ravindran, V, Hew, LI, Ravindran, G, and Bryden, WL. Apparent ileal digestibility of amino acids in dietary ingredients for broiler chickens. Anim Sci. (2005) 81:85–97. doi: 10.1079/ASC42240085
74. Ludwig, D, Crowe, PA, and Hassemer, MM. Free fat and glycogen during metamorphosis of Musca domestica L. J NY Entom Soc. (1964) 72:23–8.
75. Parsons, CM. Digestible amino acids for poultry and swine. Anim Feed Sci Technol. (1996) 59:147–53. doi: 10.1016/0377-8401(95)00895-0
77. Poelaert, C, Beckers, Y, Despret, X, Portetelle, D, Francis, F, and Bindelle, J. In vitro evaluation of fermentation characteristics of two types of insects as potential novel protein feeds for pigs. J Anim Sci. (2016) 94:198–201. doi: 10.2527/jas.2015-9533
78. Huang, C, Feng, W, Xiong, J, Wang, T, Wang, W, Wang, C, et al. Impact of drying method on the nutritional value of the edible insect protein from black soldier fly (Hermetia illucens L.) larvae: amino acid composition, nutritional value evaluation, in vitro digestibility, and thermal properties. Eur Food Res Technol. (2019) 245:11–21. doi: 10.1007/s00217-018-3136-y
79. Ibitoye, EB, Lokman, IH, Hezmee, MNM, Goh, YM, Zuki, ABZ, Jimoh, AA, et al. Gut health and serum growth hormone levels of broiler chickens fed dietary chitin and chitosan from cricket and shrimp. Poult Sci. (2019) 98:745–52. doi: 10.3382/ps/pey419
80. Veldkamp, T, Dong, L, Paul, A, and Govers, C. Bioactive properties of insect products for monogastric animals–a review. J Insects Food Feed. (2022) 8:1027–40. doi: 10.3920/JIFF2021.0031
81. Józefiak, D, Kierończyk, B, Juśkiewicz, J, Zduńczyk, Z, Rawski, M, Długosz, J, et al. Dietary nisin modulates the gastrointestinal microbial ecology and enhances growth performance of the broiler chickens. PLoS One. (2013) 8:e85347. doi: 10.1371/journal.pone.0085347
82. Kierończyk, B, Pruszyńska Oszmałek, E, Świątkiewicz, S, Rawski, M, Długosz, J, Engberg, RM, et al. The nisin improves broiler chicken growth performance and interacts with salinomycin in terms of gastrointestinal tract microbiota composition. J Anim Feed Sci. (2016) 25:309–16. doi: 10.22358/jafs/67802/2016
83. Choi, SC, Ingale, SL, Kim, JS, Park, YK, Kwon, IK, and Chae, BJ. Effects of dietary supplementation with an antimicrobial peptide-P5 on growth performance, nutrient retention, excreta and intestinal microflora and intestinal morphology of broilers. Anim Feed Sci Technol. (2013) 185:78–84. doi: 10.1016/j.anifeedsci.2013.07.005
84. Choi, SC, Ingale, SL, Kim, JS, Park, YK, Kwon, IK, and Chae, BJ. An antimicrobial peptide-A3: effects on growth performance, nutrient retention, intestinal and faecal microflora and intestinal morphology of broilers. Br Poult Sci. (2013) 54:738–46. doi: 10.1080/00071668.2013.838746
85. Wen, L-F, and He, J-G. Dose-response effects of an antimicrobial peptide, a cecropin hybrid, on growth performance, nutrient utilisation, bacterial counts in the digesta and intestinal morphology in broilers. Br J Nutr. (2012) 108:1756–63. doi: 10.1017/S0007114511007240
86. Wang, S, Zeng, XF, Wang, QW, Zhu, JL, Peng, Q, Hou, CL, et al. The antimicrobial peptide sublancin ameliorates necrotic enteritis induced by Clostridium perfringens in broilers. J Anim Sci. (2015) 93:4750–60. doi: 10.2527/jas.2015-9284
87. Park, SI, and Hahm, KS. Antimicrobial peptides (AMPs): peptide structure and mode of action. Biochem Mol Biol J. (2005) 38:766. doi: 10.5483/bmbrep.2005.38.5.507
88. Kang, JK, Hwang, JS, Nam, HJ, Ahn, KJ, Seok, H, Kim, S-K, et al. The insect peptide coprisin prevents Clostridium difficile-mediated acute inflammation and mucosal damage through selective antimicrobial activity. Antimicrob Agents Chemother. (2011) 55:4850–7. doi: 10.1128/AAC.00177-11
89. Thevissen, K, Warnecke, D, François, I, Leipelt, M, Heinz, E, Ott, C, et al. Defensins from insects and plants interact with fungal glucosylceramides. J Biol Chem. (2004) 279:3900–5. doi: 10.1074/jbc.M311165200
90. Otvos, JL. Antibacterial peptides isolated from insects. J Peptide Sci. (2000) 6:497–511. doi: 10.1002/1099-1387(200010)6:10<497::AID-PSC277>3.0.CO;2-W
91. Yi, H-Y, Chowdhury, M, Huang, Y-D, and Yu, X-Q. Insect antimicrobial peptides and their applications. App Microbiol Biotechnol. (2014) 98:5807–22. doi: 10.1007/s00253-014-5792-6
92. Jozefiak, D, Jozefiak, A, Kieronczyk, B, Rawski, M, Swiatkiewicz, S, Dlugosz, J, et al. Insects – a natural nutrient source for poultry – a review. Ann Anim Sci. (2016) 16:297–313. doi: 10.1515/aoas-2016-0010
93. Chapuisat, M, Oppliger, A, Magliano, P, and Christe, P. Wood ants use resin to protect themselves against pathogens. Proc Roy Soc B: Biol Sci. (2007) 274:2013–7. doi: 10.1098/rspb.2007.0531
94. Viljakainen, L, and Pamilo, P. Selection on an antimicrobial peptide defensin in ants. J Mol Evol. (2008) 67:643–52. doi: 10.1007/s00239-008-9173-6
95. Andersen, AS, Sandvang, D, Schnorr, KM, Kruse, T, Neve, S, Joergensen, B, et al. A novel approach to the antimicrobial activity of maggot debridement therapy. J Antimicrob Chemother. (2010) 65:1646–54. doi: 10.1093/jac/dkq165
96. Schmitt, P, Rosa, RD, and Destoumieux-Garzón, D. An intimate link between antimicrobial peptide sequence diversity and binding to essential components of bacterial membranes. Biochim Biophys Acta. (2016) 1858:958–70. doi: 10.1016/j.bbamem.2015.10.011
97. Scocchi, M, Tossi, A, and Gennaro, R. Proline-rich antimicrobial peptides: converging to a non-lytic mechanism of action. Cel Mol Life Sci. (2011) 68:2317–30. doi: 10.1007/s00018-011-0721-7
98. Li, W-F, Ma, G-X, and Zhou, X-X. Apidaecin-type peptides: biodiversity, structure-function relationships and mode of action. Peptides. (2006) 27:2350–9. doi: 10.1016/j.peptides.2006.03.016
99. Nicolas, P. Multifunctional host defense peptides: intracellular targeting antimicrobial peptides. FEBS J. (2009) 276:6483–96. doi: 10.1111/j.1742-4658.2009.07359.x
100. Mattiuzzo, M, Bandiera, A, Gennaro, R, Benincasa, M, Pacor, S, Antcheva, N, et al. Role of the Escherichia coli SbmA in the antimicrobial activity of proline-rich peptides. Mol Microbiol. (2007) 66:151–63. doi: 10.1111/j.1365-2958.2007.05903.x
101. Hansen, A, Schäfer, I, Knappe, D, Seibel, P, and Hoffmann, R. Intracellular toxicity of proline-rich antimicrobial peptides shuttled into mammalian cells by the cell-penetrating peptide penetratin. Antimicrob Agents Chemother. (2012) 56:5194–201. doi: 10.1128/AAC.00585-12
102. Lee, VS, Tu, WC, Jinn, TR, Peng, CC, Lin, LJ, and Tzen, JT. Molecular cloning of the precursor polypeptide of mastoparan B and its putative processing enzyme, dipeptidyl peptidase IV, from the black-bellied hornet, Vespa basalis. Insect Mol Biol. (2007) 16:231–7. doi: 10.1111/j.1365-2583.2006.00718.x
103. Carlsson, A, Nyström, T, De Cock, H, and Bennich, H. Attacin–an insect immune protein–binds LPS and triggers the specific inhibition of bacterial outer-membrane protein synthesis. Microbiol. (1998) 144:2179–88. doi: 10.1099/00221287-144-8-2179
104. Rossi, R, Pastorelli, G, Cannata, S, and Corino, C. Recent advances in the use of fatty acids as supplements in pig diets: a review. Anim Feed Sci Technol. (2010) 162:1–11. doi: 10.1016/j.anifeedsci.2010.08.013
105. Liu, Y. Fatty acids, inflammation and intestinal health in pigs. J Anim Sci Biotechnol. (2015) 6:41. doi: 10.1186/s40104-015-0040-1
106. Guillot, E, Vaugelade, P, Lemarchali, P, and Re Rat, A. Intestinal absorption and liver uptake of medium-chain fatty acids in nonanaesthetized pigs. Br J Nutr. (1993) 69:431–42. doi: 10.1079/BJN19930045
107. Zeitz, JO, Fennhoff, J, Kluge, H, Stangl, GI, and Eder, K. Effects of dietary fats rich in lauric and myristic acid on performance, intestinal morphology, gut microbes, and meat quality in broilers. Poult Sci. (2015) 94:2404–13. doi: 10.3382/ps/pev191
108. Wu, Y, Zhang, H, Zhang, R, Cao, G, Li, Q, Zhang, B, et al. Serum metabolome alterations in broiler chickens supplemented with lauric acid. Poult Sci. (2021) 100:101315. doi: 10.1016/j.psj.2021.101315
109. Lieberman, S, Enig, MG, and Preuss, PHG. A review of monolaurin and lauric acid: natural virucidal and bactericidal agents. Altern Complement Ther. (2006) 12:310–4. doi: 10.1089/act.2006.12.310
110. Skřivanová, E, Marounek, M, Dlouhá, G, and Kaňka, J. Susceptibility of Clostridium perfringens to C2-C18 fatty acids. Lett App Microbiol. (2005) 41:77–81. doi: 10.1111/j.1472-765X.2005.01709.x
111. Spranghers, T, Michiels, J, Vrancx, J, Ovyn, A, Eeckhout, M, De Clercq, P, et al. Gut antimicrobial effects and nutritional value of black soldier fly (Hermetia illucens L.) prepupae for weaned piglets. Anim Feed Sci Technol. (2018) 235:33–42. doi: 10.1016/j.anifeedsci.2017.08.012
112. Roth, FX, and Kirchgessner, M. Organic acids as feed additives for young pigs: nutritional and gastrointestinal effects. J Anim Feed Sci. (1998) 7:25–33. doi: 10.22358/jafs/69953/1998
113. Ricke, SC. Perspectives on the use of organic acids and short chain fatty acids as antimicrobials. Poult Sci. (2003) 82:632–9. doi: 10.1093/ps/82.4.632
114. Spranghers, T, Ottoboni, M, Klootwijk, C, Ovyn, A, Deboosere, S, De Meulenaer, B, et al. Nutritional composition of black soldier fly (Hermetia illucens) prepupae reared on different organic waste substrates. J Sci Food Agric. (2017) 97:2594–600. doi: 10.1002/jsfa.8081
115. St-Hilaire, S, Cranfill, K, McGuire, MA, Mosley, EE, Tomberlin, JK, Newton, L, et al. Fish offal recycling by the black soldier fly produces a foodstuff high in omega-3 fatty acids. J World Aquac Soc. (2007) 38:309–13. doi: 10.1111/j.1749-7345.2007.00101.x
116. Suyatma, NE, Copinet, A, Tighzert, L, and Coma, V. Mechanical and barrier properties of biodegradable films made from chitosan and poly (lactic acid) blends. J Polym Envir. (2004) 12:1–6. doi: 10.1023/B:JOOE.0000003121.12800.4e
117. Islam, S, Bhuiyan, MAR, and Islam, MN. Chitin and chitosan: structure, properties and applications in biomedical engineering. J Polym Environ. (2017) 25:854–66. doi: 10.1007/s10924-016-0865-5
118. Vartiainen, J, Motion, R, Kulonen, H, Rättö, M, Skyttä, E, and Ahvenainen, R. Chitosan-coated paper: effects of nisin and different acids on the antimicrobial activity. J App Polym Sci. (2004) 94:986–93. doi: 10.1002/app.20701
119. Komi, DEA, Sharma, L, and Dela Cruz, CS. Chitin and its effects on inflammatory and immune responses. Clinic Rev Allerg Immunol. (2018) 54:213–23. doi: 10.1007/s12016-017-8600-0
120. Lee, CG, Da Silva, CA, Lee, J-Y, Hartl, D, and Elias, JA. Chitin regulation of immune responses: an old molecule with new roles. Curr Opin Immunol. (2008) 20:684–9. doi: 10.1016/j.coi.2008.10.002
121. Islam, MM, and Yang, CJ. Efficacy of mealworm and super mealworm larvae probiotics as an alternative to antibiotics challenged orally with Salmonella and E. coli infection in broiler chicks. Poult Sci. (2017) 96:27–34. doi: 10.3382/ps/pew220
122. Bovera, F, Loponte, R, Pero, ME, Cutrignelli, MI, Calabrò, S, Musco, N, et al. Laying performance, blood profiles, nutrient digestibility and inner organs traits of hens fed an insect meal from Hermetia illucens larvae. Res Vet Sci. (2018) 120:86–93. doi: 10.1016/j.rvsc.2018.09.006
123. Marono, S, Loponte, R, Lombardi, P, Vassalotti, G, Pero, ME, Russo, F, et al. Productive performance and blood profiles of laying hens fed Hermetia illucens larvae meal as total replacement of soybean meal from 24 to 45 weeks of age. Poult Sci. (2017) 96:1783–90. doi: 10.3382/ps/pew461
124. Koide, SS. Chitin-chitosan: properties, benefits and risks. Nutr Res. (1998) 18:1091–101. doi: 10.1016/S0271-5317(98)00091-8
125. Hossain, S, and Blair, R. Chitin utilization by broilers and its effect on body composition and blood metabolites. Br Poult Sci. (2007) 48:33–8. doi: 10.1080/00071660601156529
126. Hoseini, MHM, Moradi, M, Alimohammadian, MH, Shahgoli, VK, Darabi, H, and Rostami, A. Immunotherapeutic effects of chitin in comparison with chitosan against Leishmania major infection. Parasitol Int. (2016) 65:99–104. doi: 10.1016/j.parint.2015.10.007
127. Abdel-Latif, M, Abdel-Haleem, HM, and Abdel-Baki, A-AS. Anticoccidial activities of chitosan on Eimeria papillata-infected mice. Parasitol Res. (2016) 115:2845–52. doi: 10.1007/s00436-016-5035-0
128. Dijksteel, GS, Ulrich, MMW, Middelkoop, E, and Boekema, BKHL. Review: lessons learned from clinical trials using antimicrobial peptides (AMPs). Front Microbiol. (2021) 12:616979. doi: 10.3389/fmicb.2021.616979
129. Dong, L, Wichers, HJ, and Govers, C. Beneficial health effects of chitin and chitosan. In: LAM BroekVan den and CG Boeriu. Chitin and chitosan. Wiley, Hoboken, NJ, USA, pp. 145–167. (2019).
130. Awoniyi, AM, Aletor, VA, and Aina, M. Performance of broiler chickens fed on maggot meal in place of fishmeal. Int J Poult Sci. (2003) 2:271–4.
131. Adeniji, AA. Effect of replacing groundnut cake with maggot meal in the diet of broilers. Int J Poult Sci. (2007) 6:822–5. doi: 10.3923/ijps.2007.822.825
132. Adesina, MA, Adejinmi, OO, Omole, AJ, Fayenuwo, JA, and Osunkeye, O. Performance of broilers’ finishers fed graded levels of cassava peelmaggot meal-based diet mixtures. J Agric Forest Soc Sci. (2011) 9:226–31. doi: 10.4314/joafss.v9i1.25
133. Okah, U, and Onwujiariri, EB. Performance of finisher broiler chickens fed maggot meal as a replacement for fish meal. J Agric Tech. (2012) 8:471–7.
134. Téguia, A, Mpoame, M, and Mba, JAO. The production performance of broiler birds as affected by the replacement of fish meal by maggot meal in the starter and finisher diets. Tropicultura. (2002) 20:187–92.
135. Inaoka, T, Okubo, G, Yokota, M, and Takemasa, M. Nutritive value of house fly larvae and pupae fed on chicken feces as food source for poultry. J Poultry Sci. (1999) 36:174–80. doi: 10.2141/jpsa.36.174
136. Atteh, JO, and Ologbenla, FD. Replacement of fish meal with maggots in broiler diets: effects on performance and nutrient retention. Niger J Anim Prod. (1993) 20:44–9.
137. Bamgbose, AM. Utilization of maggot-meal in cockerel diets. Indian J Anim Nutr. (1999) 69:1056–8.
138. Schiavone, A, DeMarco, M, Rotolo, L, Belforti, M, Martinez Mirò, S, Madrid Sanchez, J, et al. Nutrient digestibility of Hermetia illucens and Tenebrio molitor meal in broiler chickens. Proceedings on conference insects to feed the world, Abstract Book, The Netherlands, May 14–17. p. 84. (2014).
139. Sanusi, M, Garba, A, Saidu, I, and Ali, YZ. Performance of broiler chickens fed graded levels of grasshopper meals. Int J Appl Res Technol. (2013) 2:235–40.
140. Sun, T, Long, RJ, and Liu, ZY. The effect of a diet containing grasshoppers and access to free-range on carcass and meat physicochemical and sensory characteristics in broilers. Br Poult Sci. (2013) 54:130–7. doi: 10.1080/00071668.2012.756575
141. Nginya, ES, Ondiek, JO, King’ori, AM, and Nduko, JM. Evaluation of grasshoppers as a protein source for improved indigenous chicken growers. Livest Res Rural Dev. (2019) 31:2.
142. Loh, TC, Fong, LY, Foo, HL, Thanh, NT, and Sheikh-Omar, AR. Utilisation of earthworm meal in partial replacement of soybean and fish meals in diets of broilers. J Appl Anim Res. (2009) 36:29–32. doi: 10.1080/09712119.2009.9707024
143. Gholami, H, Shargh, MS, Zarabi, M, and Zerehdaran, S. Effect of different levels of earthworm meal (Eisenia Fetida) on performance, carcass characteristics and blood parameters of broiler chickens. Anim Prod Res. (2016) 7:70–6. doi: 10.18869/acadpub.rap.7.13.76
144. Nalunga, A, Komakech, AJ, Jjagwe, J, Magala, H, and Lederer, J. Growth characteristics and meat quality of broiler chickens fed earthworm meal from Eudrilus eugeniae as a protein source. Livest Sci. (2021) 245:104394. doi: 10.1016/j.livsci.2021.104394
145. Zang, YT, Bing, S, Zhang, YZ, Sheng, XW, and Shu, DQ. Effects of dietary supplementation with earthworm powder on production performance, blood characteristics, and heavy metal residues of broiler pullets. J Appl Poult Res. (2018) 27:609–15. doi: 10.3382/japr/pfy024
146. Pieterse, E, Pretorius, Q, Hoffman, LC, and Drew, D. The carcass quality, meat quality and sensory characteristics of broilers raised on diets containing either Musca domestica larvae meal, fish meal or soya bean meal as the main protein source. Anim Prod Sci. (2014) 54:622–8. doi: 10.1071/AN13073
147. Bovera, F, Loponte, R, Marono, S, Piccolo, G, Parisi, G, Iaconisi, V, et al. Use of Tenebrio molitor larvae meal as protein source in broiler diet: effect on growth performance, nutrient digestibility, and carcass and meat traits. J Anim Sci. (2016) 94:639–47. doi: 10.2527/jas.2015-9201
148. Biasato, I, De Marco, M, Rotolo, L, Renna, M, Dabbou, S, Capucchio, MT, et al. Effects of dietary Tenebrio molitor meal inclusion in freerange chickens. J Anim Physiol Anim Nutr. (2016) 100:1104–12. doi: 10.1111/jpn.12487
149. Biasato, I, Gasco, L, De Marco, M, Renna, M, Rotolo, L, Dabbou, S, et al. Yellow mealworm larvae (Tenebrio molitor) inclusion in diets for male broiler chickens: effects on growth performance, gut morphology, and histological findings. Poult Sci. (2018) 97:540–8. doi: 10.3382/ps/pex308
150. Ballitoc, DA, and Sun, S. Ground yellow mealworms (Tenebrio molitor L.) feed supplementation improves growth performance and carcass yield characteristics in broilers. Open Sci Reposit Agric. (2013) 2013:e23050425. doi: 10.7392/OPENACCESS.23050425
151. Altmann, BA, Neumann, C, Velten, S, Liebert, F, and Mörlein, D. Meat quality derived from high inclusion of a micro-alga or insect meal as an alternative protein source in poultry diets: a pilot study. Foods Basel Switz. (2018) 7:34. doi: 10.3390/foods7030034
152. Schiavone, A, Dabbou, S, Petracci, M, Zampiga, M, Sirri, F, Biasato, I, et al. Black soldier fly defatted meal as a dietary protein source for broiler chickens: effects on carcass traits, breast meat quality and safety. Animal. (2019) 13:2397–405. doi: 10.1017/S1751731119000685
153. Leiber, F, Gelencsér, T, Stamer, A, Amsler, Z, Wohlfahrt, J, Früh, B, et al. Insect and legume-based protein sources to replace soybean cake in an organic broiler diet: effects on growth performance and physical meat quality. Renew Agric Food Syst. (2019) 32:21–7.
154. Khatun, R, Howlider, MAR, Rahman, MM, and Hasanuzzama, M. Replacement of fish meal by silkworm pupae in broiler diets. Pak J Biol Sci. (2003) 6:955–8. doi: 10.3923/pjbs.2003.955.958
155. Ullah, R, Khan, S, Hafeez, A, Asad, S, Nazir, AK, Naila, C, et al. Silkworm (Bombyx mori) meal as alternate protein ingredient in broiler finisher ration. Pak J Zool. (2017) 49:1463–70. doi: 10.17582/journal.pjz/2017.49.4.1463.1470
156. Fagoonee, I. Possible growth factors for chickens in silkworm pupae meal. Br Poult Sci. (1983) 24:295–300. doi: 10.1080/00071668308416743
157. Muftau, MA, and Olorede, BR. Carcass characteristics and economics of broiler chickens fed different levels of grasshopper meal in place of fish meal. Anim Prod Res Adv. (2009) 5:248–51.
158. Tancharoenrat, P, Ravindran, V, Zaefarian, F, and Ravindran, G. Digestion of fat and fatty acids along the gastrointestinal tract of broiler chickens. Poult Sci. (2014) 93:371–9. doi: 10.3382/ps.2013-03344
159. Abdulla, NR, Loh, TC, Foo, HL, Alshelmani, MI, and Akit, H. Influence of dietary ratios of n-6: n-3 fatty acid on gene expression, fatty acid profile in liver and breast muscle tissues, serum lipid profile, and immunoglobulin in broiler chickens. J Appl Poult Res. (2019) 28:454–69. doi: 10.3382/japr/pfz008
160. Ssepuuya, G, Namulawa, V, Mbabazi, D, Mugerwa, S, Fuuna, P, Nampijja, Z, et al. Use of insects for fish and poultry compound feed in sub-saharan Africa- A systematic review. JInsects FOOD FEED. (2017) 3:289–302. doi: 10.3920/JIFF2017.0007
161. Pieterse, E, Erasmus, SW, Uushona, T, and Hoffman, LC. Black soldier fly (Hermetia illucens) pre-pupae meal as a dietary protein source for broiler production ensures a tasty chicken with standard meat quality for every pot. J Sci Food Agric. (2019) 99:893–903. doi: 10.1002/jsfa.9261
162. Moula, N, and Detilleux, J. A Meta-analysis of the effects of insects in feed on poultry growth performances. Animals. (2019) 9:201. doi: 10.3390/ani9050201
163. Star, L, Arsiwalla, T, Molist, F, Leushuis, R, Dalim, M, and Paul, A. Gradual provision of black soldier fly (Hermetia Illucens) larvae to older laying hens: effect on production performance, egg quality, feather condition and behavior. Animals. (2020) 10:216. doi: 10.3390/ani10020216
164. Maurer, V, Holinger, M, Amsler, Z, Früh, B, Wohlfahrt, J, Stamer, A, et al. Replacement of soybean cake by Hermetia illucens meal in diets for layers. J Insect Food Feed. (2016) 2:83–90. doi: 10.3920/JIFF2015.0071
165. Kawasaki, K, Hashimoto, Y, Hori, A, Kawasaki, T, Hirayasu, H, Iwase, SI, et al. Evaluation of black soldier fly (Hermetia illucens) larvae and pre-pupae raised on household organic waste, as potential ingredients for poultry feed. Animals. (2019) 9:98. doi: 10.3390/ani9030098
166. Al-Qazzaz, MFA, Ismail, D, Akit, H, and Idris, LH. Effect of using insect larvae meal as a complete protein source on quality and productivity characteristics of laying hens. Revista Brasileira De Zoot. (2016) 45:518–23. doi: 10.1590/s1806-92902016000900003
167. Dankwa, D, Nelson, FS, Oddoye, EOK, and Duncan, JL. Housefly larvae as a feed supplement for rural poultry. Ghana J Agric Sci. (2002) 35:185–7.
168. Agunbiade, JA, Adeyemi, OA, Ashiru, OM, Awojobi, HA, Taiwo, AA, Oke, DB, et al. Replacement of fish meal with maggot meal in cassava-based layers’ diets. J. Poultry Sci. (2007) 44:278–82. doi: 10.2141/jpsa.44.278
169. Ullah, R, Khan, S, Khan, N, Mobashar, M, Sultan, A, Ahmad, N, et al. Replacement of soybean meal with silkworm meal in the diets of white leghorn layers and effects on performance, apparent total tract digestibility, blood profile and egg quality. Int J Vet Health Sci Res. (2017) 5:200–7. doi: 10.19070/2332-2748-1700040
170. Abdel-Kareem, AAA, and El-Sheikh, TM. Impact of supplementing diets with propolis on productive performance, egg quality traits and some haematological variables of laying hens. J Anim Physiol Anim Nutr. (2017) 101:441–8. doi: 10.1111/jpn.12407
171. Rizk, YS, Abuoghaba, AA, Ismail, II, and Awadien, NB. Egg production, egg quality traits and some hematological parameters of Sinai chicken strain treated with different levels of bee pollen. Egypt Poult Sci. (2018) 38:427–38.
172. Barragan-Fonseca, KB, Dicke, M, and van Loon, JJA. Influence of larval density and dietary nutrient concentration on performance, body protein, and fat contents of black soldier fly larvae (Hermetia illucens). Entomol Exp Appl. (2018) 166:761–70. doi: 10.1111/eea.12716
173. Zheng, L, Qing, L, Jibin, Z, and Ziniu, Y. Double the biodiesel yield: rearing black soldier fly larvae, Hermetia illucens, on solid residual fraction of restaurant waste after grease extraction for biodiesel production. Renew Energy. (2012) 41:75–9. doi: 10.1016/j.renene.2011.10.004
174. Nguyen, TT, Tomberlin, JK, and Vanlaerhoven, S. Ability of black soldier fly (Diptera: Stratiomyidae) larvae to recycle food waste. Envir Entomol. (2015) 44:406–10. doi: 10.1093/ee/nvv002
175. Tschirner, M, and Simon, A. Influence of different growing substrates and processing on the nutrient composition of black soldier fly larvae destined for animal feed. J Insects Food Feed. (2015) 1:1–12. doi: 10.3920/JIFF2014.0008
176. Liceaga, AM. Processing insects for use in the food and feed industry. Curr Op Insect Sci. (2021) 48:32–6. doi: 10.1016/j.cois.2021.08.002
177. Mutungi, C, Irungu, FG, Nduko, J, Mutua, F, Affognon, H, Nakimbugwe, D, et al. Postharvest processes of edible insects in Africa: A review of processing methods, and the implications for nutrition, safety and new products development. Crit Rev Food Sci Nutr. (2019) 59:276–98. doi: 10.1080/10408398.2017.1365330
178. Nyangena, DN, Mutungi, C, Imathiu, S, Kinyuru, J, Affognon, H, Ekesi, S, et al. Effects of traditional processing techniques on the nutritional and microbiological quality of four edible insect species used for food and feed in East Africa. Foods. (2020) 9:574. doi: 10.3390/foods9050574
179. Dobermann, D, Field, LM, and Michaelson, LV. Impact of heat processing on the nutritional content of Gryllus bimaculatus (black cricket). Nutr Bull. (2019) 44:116–22. doi: 10.1111/nbu.12374
180. Gravel, A, and Doyen, A. The use of edible insect proteins in food: challenges and issues related to their functional properties. Innov Food Sci Emerg Technol. (2020) 59:102272. doi: 10.1016/j.ifset.2019.102272
181. Hall, FG, Jones, OG, O’Haire, ME, and Liceaga, AM. Functional properties of tropical banded cricket (Gryllodes sigillatus) protein hydrolysates. Food Chem. (2017) 224:414–22. doi: 10.1016/j.foodchem.2016.11.138
182. Liceaga, AM. Approaches for utilizing insect protein for human consumption: effect of enzymatic hydrolysis on protein quality and functionality. Ann Entomol Soc Am. (2019) 112:529–32. doi: 10.1093/aesa/saz010
183. Laroche, M, Perreault, V, Marciniak, A, Gravel, A, Chamberland, J, and Doyen, A. Comparison of conventional and sustainable lipid extraction methods for the production of oil and protein isolate from edible insect meal. Foods. (2019) 8:572. doi: 10.3390/foods8110572
184. Bruno, D, Bonelli, M, De Filippis, F, Di Lelio, I, Tettamanti, G, Casartelli, M, et al. The intestinal microbiota of Hermetia illucens larvae is affected by diet and shows a diverse composition in the different midgut regions. Appl Environ Microbiol. (2019) 85:e01864–18. doi: 10.1128/AEM.01864-18
185. Jiang, C-L, Jin, W-Z, Tao, X-H, Zhang, Q, Zhu, J, Feng, S-Y, et al. Black soldier fly larvae (Hermetia illucens) strengthen the metabolic function of food waste biodegradation by gut microbiome. Microb Biotechnol. (2019) 12:528–43. doi: 10.1111/1751-7915.13393
186. Klammsteiner, T, Walter, A, Bogataj, T, Heussler, CD, Stres, B, Steiner, FM, et al. The core gut microbiome of black soldier fly (Hermetia illucens) larvae raised on low-bioburden diets. Front Microbiol. (2020) 11:993. doi: 10.3389/fmicb.2020.00993
187. Wynants, E, Frooninckx, L, Crauwels, S, Verreth, C, De Smet, J, Sandrock, C, et al. Assessing the microbiota of black soldier fly larvae (Hermetia illucens) reared on organic waste streams on four different locations at laboratory and large scale. Microb Ecol. (2018) 77:913–30. doi: 10.1007/s00248-018-1286-x
188. Wu, N, Wang, X, Xu, X, Cai, R, and Xie, S. Effects of heavy metals on the bioaccumulation, excretion and gut microbiome of black soldier fly larvae (Hermetia illucens). Ecotoxicol Envir Safety. (2020) 192:110323. doi: 10.1016/j.ecoenv.2020.110323
189. Chen, Y, Li, X, Song, J, Yang, D, Liu, W, Chen, H, et al. Isolation and characterization of a novel temperate bacteriophage from gut-associated Escherichia within black soldier fly larvae (Hermetia illucens L. [Diptera: Stratiomyidae]). Arch Virol. (2019) 164:2277–84. doi: 10.1007/s00705-019-04322-w
190. European Food Safety Authority Scientific Committee. Risk profile related to production and consumption of insects as food and feed. EFSA J. (2015) 13:4257. doi: 10.2903/j.efsa.2015.4257
191. Schrogel, P, and Wätjen, W. Insects for food and feed-safety aspects related to mycotoxins and metals. Foods. (2019) 8:288. doi: 10.3390/foods8080288
192. Fels-Klerx, HJ, van der Camenzuli, L, Belluco, S, Meijer, N, and Ricci, A. Food safety issues related to uses of insects for feeds and foods. Comp Rev Food Sci Food Saf. (2018) 17:1172–83. doi: 10.1111/1541-4337.12385
193. Vijver, M, Jager, T, Posthuma, L, and Peijnenburg, W. Metal uptake from soils and soil– sediment mixtures by larvae of Tenebrio molitor (L.) (Coleoptera). Ecotoxicol Environ Saf. (2003) 54:277–89. doi: 10.1016/S0147-6513(02)00027-1
194. Diener, S, Zurbrügg, C, and Tockner, K. Bioaccumulation of heavy metals in the black soldier fly, Hermetia illucens and effects on its life cycle. J Insects Food Feed. (2015) 1:261–70. doi: 10.3920/JIFF2015.0030
Keywords: insect meal, broiler chickens, laying hens, feed, digestibility, safety, immune system, bioactive molecules
Citation: Belhadj Slimen I, Yerou H, Ben Larbi M, M’Hamdi N and Najar T (2023) Insects as an alternative protein source for poultry nutrition: a review. Front. Vet. Sci. 10:1200031. doi: 10.3389/fvets.2023.1200031
Edited by:
Domenico Pietro Lo Fiego, University of Modena and Reggio Emilia, ItalyReviewed by:
Sihem Dabbou, University of Trento, ItalyCopyright © 2023 Belhadj Slimen, Yerou, Ben Larbi, M’Hamdi and Najar. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Imen Belhadj Slimen, aW1lbi5iZWxoYWRqc2xpbWVuQGluYXQudWNhci50bg==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.