 Ghazal Nemati
Ghazal Nemati Alicia Romanó
Alicia Romanó Fabian Wahl
Fabian Wahl Thomas Berger
Thomas Berger Laura Vazquez Rojo2
Laura Vazquez Rojo2- 1Food Microbial Systems, Risk Assessment and Mitigation Group, Agroscope, Bern, Switzerland
- 2Food Microbial Systems, Microbiological Safety of Foods of Animal Origin Group, Agroscope, Bern, Switzerland
- 3Food Microbial Systems, Agroscope, Bern, Switzerland
In dairy herds managements, mastitis is the leading cause of economic losses. One of the most important pathogens responsible for intra-mammary infections is Staphylococcus aureus. The genetic properties of S. aureus have a strong influence on its pathogenicity and contagiousness. In this study, we aimed to obtain a comprehensive overview of the key bovine S. aureus clinical properties, such as contagiousness and antimicrobial resistance, present in European strains. For this, 211 bovine S. aureus strains from ten European countries that were used in a previous study were used in this study. Contagiousness was assessed using qPCR for the detection of the marker gene adlb. Antimicrobial resistance was evaluated using a broth microdilution assay and mPCR for the detection of genes involved in penicillin resistance (blaI, blaR1, and blaZ). It was found that adlb was present in CC8/CLB strains; however, in Germany, it was found in CC97/CLI and in an unknown CC/CLR strains. CC705/CLC strains from all countries were found to be susceptible to all tested antibiotics. Major resistance to penicillin/ampicillin, chloramphenicol, clindamycin and tetracycline was detected. Resistance to oxacillin, trimethoprim/sulfamethoxazole and cephalosporins was rarely observed. In addition, contagiousness and antibiotic resistance seem to correlate with different CCs and genotypic clusters. Hence, it is recommended that multilocus sequence typing or genotyping be utilized as a clinical instrument to identify the most appropriate antibiotic to use in mastitis treatment. Actualization of the breakpoints of veterinary strains is necessary to address the existing antibiotic resistance of the bacteria involved in veterinary mastitis.
1. Introduction
In veterinary medicine, mastitis is the leading cause of economic losses in dairy herds management. It contributes to reductions in milk quality and production, there are costs associated with its treatment, and animal culling can be a consequence of treatment failures (1, 2). In Switzerland, the total cost of mastitis is ~$131 million annually, according to Heiniger et al. (3). One of the most important pathogens responsible for intramammary infections (IMIs) is Staphylococcus aureus (4). S. aureus may infect only some individual animals or may be contagious and infect the entire herd; infections usually resulting in subclinical chronic mastitis (5, 6). As shown previously (5, 7–12), the genetic properties of S. aureus have a strong influence on its pathogenicity and contagiousness, making subtyping necessary to improve treatment success and dairy herd management. Using ribosomal spacer PCR (RS-PCR), it has been shown that the rate of infected cows in a herd is highly dependent on the bacterial genotype (GT) (7–10), and, S. aureus genotype B (GTB) and its variants may infect up to 100% of cows in the same herd (7–9, 11) due to its high contagiousness (13, 14). In contrast, other genotypes and their variants (e.g., GTC, GTF, GTS) are restricted to one or a few cows in a herd (7–10, 15, 16). In the electrophoresis of the RS-PCR product, variants differ in 1 electrophoretic band and as consequence, are named by superscripted roman numerals (e.g., GTRI). For further simplification, genotypes and their variants are combined into genotypic clusters (CL). For example, GTB and its variants form a cluster named CLB. Multilocus sequence typing (MLST) (17) results have shown that CLB is almost exclusively associated with clonal complex 8 (CC8), whereas CLC corresponds largely to CC705, and CLR to CC97 and CC133 (9, 18). In Europe, CLB, CLC, CLF, CLI, and CLR account for 76.6% of all S. aureus isolates obtained from clinical milk samples (19).
RS-PCR is particularly suitable for clinical applications as it is a low-cost, high-throughput method that provides analytical resolution at least as good as spa typing in bovine strains (9, 18). However, it is more appropriate to use MLST for subtyping at the biological level because a S. aureus clone can be used (17) and, consequently, evolutionary identity established (20, 21). To sanitize Swiss dairy herds infected with the contagious S. aureus CLB, Sartori et al., developed a real-time quantitative PCR (qPCR) assay to identify this pathogen in milk samples and achieved diagnostic sensitivity and specificity at the cow level of 99 and 100%, respectively (22). This new assay has been used to detect, with high specificity, the gene adlb which encodes the bovine adhesion-like protein located in the GTB-specific staphylococcal cassette chromosome SCCgtb (16, 22). It is a marker for contagiousness and high prevalence of intra-mammary infection (IMI) in dairy herds (11, 16).
Antibiotic (AB) treatment is still one of the most important measures for controlling bovine mastitis (23). However, the frequently unsatisfactory cure rates remain a serious concern, particularly for IMI caused by S. aureus (6, 24–27). One major reason for this drawback is the improper use of ABs (28, 29). Additionally, AB treatments applied at the herd level are usually not reported, even though various mastitis control plans strongly recommend performing these analyses and collecting the resultant data (30). Since 2019, it has been required for Swiss's farms to declare the AB treatments used at the herd level (31). In terms of the ABs used to treatment bovine IMIs caused by S. aureus, various classes of AB are used: typically, ß-lactams (penicillins and cephalosporins), aminoglycosides, lincosamides, and macrolides (32, 33). Penicillin G is the most commonly used AB for treating IMI in cows caused by S. aureus and other Gram-positive mastitis pathogens. In S. aureus, the bla operon mediates AB resistance against penicillin G and other β-lactamase-sensitive penicillins. The bla operon can be located on plasmids (as transposon) or on the chromosome (34, 35) and contains three genes: (1) blaZ, which encodes the βlactamase that hydrolyzes the β-lactam ring of AB, rending them inactive; (2) blaI, which encodes the repressor; (3) blaR1 which encodes the sensor and antirepressor (35, 36). Ivanovic et al., recently showed that the bla operon plays a key role in phenotypic resistance to penicillin. Furthermore, for S. aureus, they highlighted the importance of using the minimum inhibitory concentration (MIC) value as the gold standard when assessing resistance to penicillin and probably other ABs (33).
As contagiousness and antimicrobial resistance (AMR) are critical pathogenic factors of the S. aureus strains responsible for bovine mastitis, a comprehensive study was performed to assess the distribution of these key clinical properties in strains from across Europe. Contagiousness was assessed using qPCR to detect the adlb gene, which is a staphylococcal marker for contagiousness and for high prevalence of intra-mammary infection in dairy herds. Furthermore, AMR was evaluated using an MIC assay and melting curve PCR (mPCR) to detect genes involved in penicillin resistance (blaI, blaR1, and blaZ).
2. Materials and methods
2.1. Strain collection
A total of 211 bovine strains of S. aureus were used in this study that had been collected from 10 European countries; Austria, Belgium, France, Germany, Ireland, Italy, Macedonia, Norway, Slovenia, and Switzerland. These strains were originally collected during two previous studies by Boss et al. (18) and Cosandey et al. (19). As described by Cosandey et al., the strains were aseptically collected from milk samples from individual quarters (19). The strains had been stored in skim milk at −20°C. They were plated onto Columbia agar plates containing 5% sheep blood (Biomérieux Suisse s.a., Geneva, Switzerland) and incubated at 37°C for 24 h (18, 19). The genotypes (GT) and the clonal complexes (CCs) information was obtained from previous studies. The distribution of the different CCs and the GT across the 10 European countries is shown in Table 1 (19).
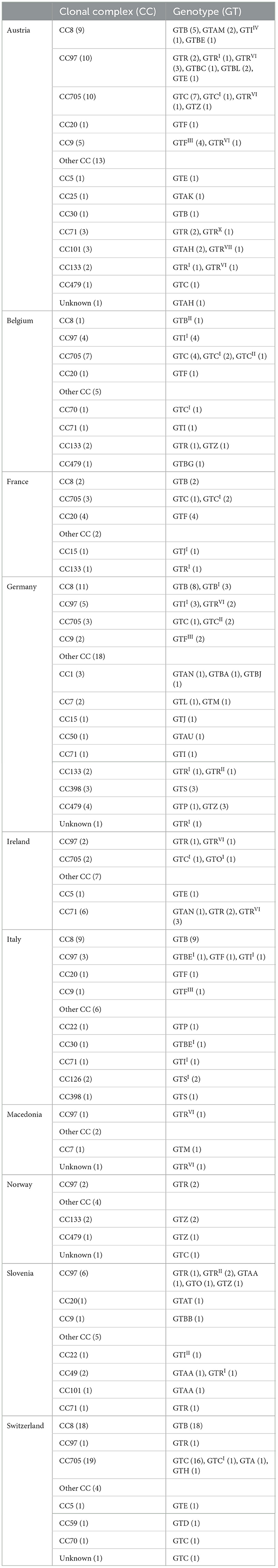
Table 1. Distribution of Staphylococcus aureus genotypes and clonal complexes across 10 European countries.
2.2. DNA extraction
DNA was extracted from single S. aureus colonies. One colony was picked and resuspended in 100 μL of 10 mM Tris-HCl and 10 mM EDTA (pH = 8.5), incubated at 95°C for 10 min, and immediately placed on ice. The lysates were diluted 1:100 in qPCR H2O (SINTETICA S.A, Mendrisio, Switzerland) for use as templates. The samples were stored at −20°C and were analyzed within 2 weeks of extraction (18).
2.3. Quantitative PCR (qPCR) with adlb and internal control gene
Real-time qPCR was performed with adlb and the internal control gene (N gene of canine distemper virus [CDVN]) according to the protocol of Sartori et al. (22). The characteristics of the utilized primers are listed in Supplementary Table S1. DNA amplification was performed using a Magnetic Induction Cycler qPCR real-time thermal cycler (Bio Molecular Systems, Australia) and the following cycling conditions: initial denaturation at 95°C for 3 min followed by 45 running cycles of denaturation at 95°C for 3 s and annealing/elongation at 60°C for 20 s. Two reference strains that were positive for both targets were included as positive controls.
2.4. PCR analysis of the bla operon genes
The mPCR was performed according to the protocol of Ivanovic et al. (33). Each of the 211 strains was analyzed for the presence of blaI, blaR1, and blaZ; each gene was detected separately. As per Ivanovic et al., amplicons with a single melting peak identical to the positive control for blaI, blaR1, or blaZ were considered positive. The characteristics of the utilized primers are listed in Supplementary Table S2.
2.5. Assessment of antimicrobial sensitivity
The sensitivity of each strain to 30 antimicrobial agents was tested by minimum inhibitory concentration (MIC) using a PM32 panel (Beckman Coulter, Inc., Brea, CA, USA) following the manufacturer's instructions. The tested ABs concentrations (μg/mL) were as follows: amoxicillin/K clavulanate (0.5/0.25–8/4), ampicillin (0.5–8), azithromycin (1–2), cefepime (4–8), cefotaxime (1–2), cefuroxime (4–8), chloramphenicol (8), ciprofloxacin (0.5–1), clindamycin (0.25–0.5, 2), daptomycin (0.5–4), ertapenem (0.5–1), erythromycin (1–2), fosfomycin (32), fusic acid (2), gentamycin (1–4), imipenem (2–8), levofloxacin (1–2), linezolid (0.5–4), meropenem (2–8), moxifloxacin (0.5–1), nitrofurantoin (64), oxacillin (0.25–2), penicillin (0.03–0.25, 2), rifampin (0.5–2), synercid (1–4), teicoplanin (1–8), tetracycline (1–2), tobramycin (1–4), trimethoprim/sulfamethoxazole (1/19–4/76), and vancomycin (0.25–8). Additionally, cefoxitin (4 μg/mL) screening was performed to determine the presence of methicillin resistant Staphylococcus aureus (MRSA) strains. When possible, the current clinical breakpoint of the EUCAST was used (37), otherwise the range specified by the CLSI was applied (38). All the ABs tested and their breakpoints are listed in Supplementary Table S3.
2.6. Statistical analysis
Data are expressed as absolute numbers or percentage. To assess the associations among different AB, the corresponding phi coefficients were computed and plotted using R 4.0.5 (39) together with the corrplot package v. 0.84. Phi values range from −1 to 1 (40). Negative phi values indicate a negative, inverse association among both variables, whereas positive phi values indicate a positive association. The Kappa test was performed using R 4.0.5 (39) to evaluate the agreement between the MIC and the bla mPCR results. Kappa values range from 0 to 1, with values of 0 and 1 indicating no and perfect agreement, respectively (41). To assess penicillin resistance, a loglinear model was computed to analyze the relationships among the factors penicillin, CC, country, and their interactions. The analysis was performed using Systat 13 (Systat Software Inc., Richmond, CA).
3. Results
3.1. Presence of adlb in European S. aureus strains
The 211 S. aureus strains collected from 10 European countries were assessed using qPCR for the presence of adlb and its association with GTs and CCs. Among the 211 strains, 46 were positive for adlb. The distribution of adlb among the different GTs and CCs and among the 10 European countries is shown in Table 2.
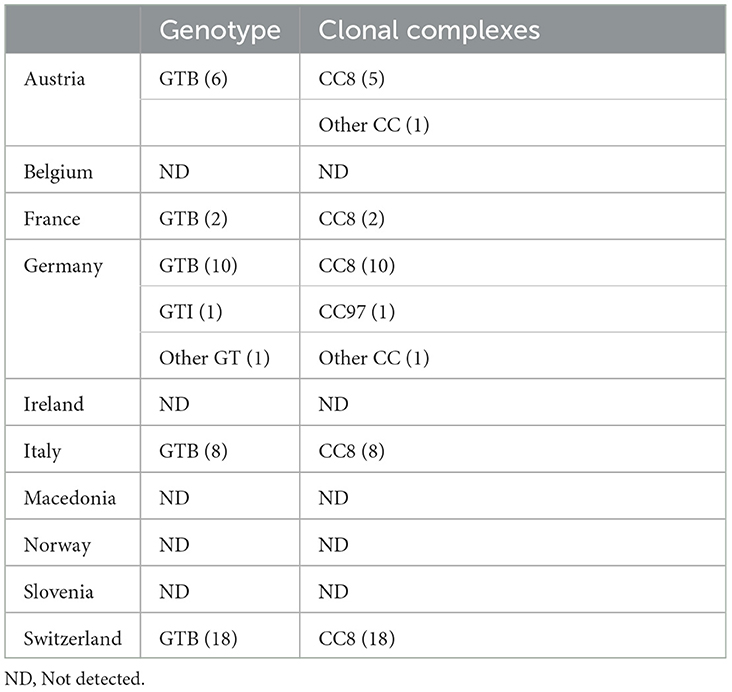
Table 2. Detailed distribution of adlb across different genotypes and clonal complexes, listed by country.
An analysis of the GTs found to contain adlb, showed that 44 of 47 (94%) CLB strains were positive for adlb and that only two strains were positive for adlb in the remaining 164 strains (1.2%). Furthermore, the gene was also observed in a German GTII and a GTRI strain. GTB was not detected in Ireland, Macedonia, Slovenia, or Norway. In Italy, Germany, and Belgium, three GTB strains were found that did not contain adlb.
3.2. AMR overview in European S. aureus strains
An analysis of the MIC data showed that 65% of the strains (n = 137) were inhibited by all the tested ABs. Table 3 shows the strains that demonstrated AMR, sorted by CC. Only the ABs to which resistance was exhibited are included.
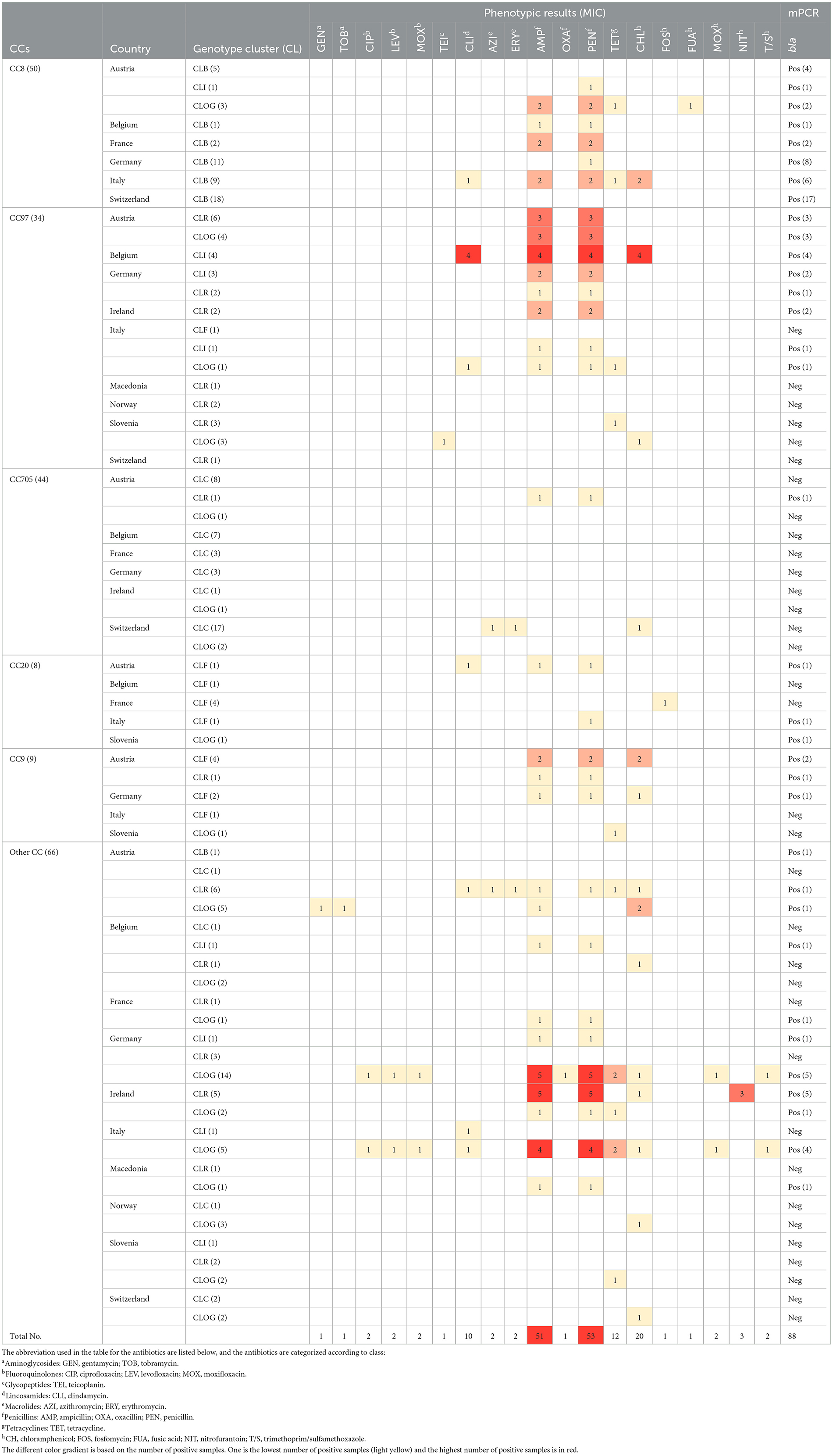
Table 3. Detailed description of the isolates (n = 211), their genotypes, and their phenotypic (and mPCR) resistance to the tested antibiotics.
Among all the ABs, the greatest number of AMR strains were found to be resistant to penicillin/ampicillin, chloramphenicol, clindamycin and tetracycline. There was no AMR observed against most of the tested antibiotics, including vancomycin, trimethoprim/sulfamethoxazole, rifampin, synercid, meropenem, linezolid, imipenem, daptomycin, and ertapenem. Interestingly, no MRSA strains were found.
A total of nine strains (4.3%) were multidrug resistant (MDR). The MDR strains were detected in only four countries: Belgium (n = 4, 1.8%), Austria (n = 1, 0.5%), Italy (n = 3, 1.4%) and Germany (n = 1, 0.5%). It is worth noting that the four Belgian strains showed the same pattern of resistance to β-lactams (ampicillin and penicillin), chloramphenicol, and clindamycin. The most resistant strain originated in Italy and showed resistance to β-lactams (ampicillin and penicillin), chloramphenicol, quinolones (ciprofloxacin, levofloxacin, and moxifloxacin), tetracycline, and trimethoprim/sulfamethoxazole.
Supplementary Figure S1 shows the AMR associations found among different ABs (ampicillin, chloramphenicol, clindamycin, penicillin and tetracycline). A strong association was found between the β-lactam ABs (ampicillin and penicillin, phi = 1.0; P < 0.001). Additionally, a strong association (phi = 0.79; P < 0.001) was found between clindamycin and chloramphenicol.
To analyze the observed penicillin resistance in more detail, a statistical model was computed to analyze the relationships among the following factors: resistance to penicillin, the most abundant CCs (CC8, CC97, and CC705), countries, and their interactions. For penicillin (n = 54), significant interactions (P < 0.001 in each case) were observed between penicillin resistance and CCs and between penicillin resistance and countries. Significant values (P < 0.001 in each case) were also obtained for the interaction between the CCs and countries, and for individual factors except the CCs (P = 0.055). Regarding the CCs, 50% and 14% of the CC97 and CC8 strains, respectively, showed resistance to penicillin. In contrast, CC705 was always sensitive to penicillin. Resistance to penicillin was particularly prominent in Austria, Belgium, Germany, and Ireland, and was absent in Slovenia and Switzerland. An identical loglinear model was also calculated for the genotypic clusters; the most observed CCs were replaced by the three most common CLs (CLB, CLC, and CLR). Significant interactions were found between penicillin resistance and CLs (P = 0.014) and between the penicillin resistance and countries (P < 0.001). CLC strains were always sensitive to penicillin, whereas 13% of CLB strains and 37% of CLR strains were resistant to penicillin. The distribution of penicillin resistance among the countries was identical to the found in the CCs model. Similar analyses for ABs other than penicillin were not performed due to a lack of sufficient data. In fact, for chloramphenicol and tetracycline, the next most common resistance targets after penicillin, only 20 (9.5%) and 12 (5.7%) of strains demonstrated resistance to these ABs, respectively.
CC705 was not only susceptible to penicillin but also to all other ABs except for one strain that was resistant to azithromycin and erythromycin (both macrolides) and another one that was resistant to chloramphenicol (Table 3). CC97 showed resistance to penicillin, chloramphenicol, and clindamycin. Increased AMR rates, in particular to penicillin/ampicillin and chloramphenicol, were also detected in CC9, CC20, and CC133 (Supplementary Table S4).
3.3. Association between MIC and bla operon genes
All 54 strains that exhibited phenotypic resistance to penicillin (26% of all strains) showed the simultaneous presence of all bla operon genes. In contrast, in 34 strains that were positive for all bla genes, the corresponding MIC value was always <0.12 μg/mL. Interestingly, this discrepancy was observed exclusively in CC8/CLB strains with the exception of one strain CC20/GTAT. For 123 trains, the MIC assay and mPCR for bla operon genes gave negative results.
4. Discussion
4.1. Prevalence of adlb in European S. aureus strains
Previous studies demonstrated that S. aureus CC8/CLB is highly contagious (13, 14) and can be detected very specifically by the qPCR assay for adlb (22) as also used in the present study. Indeed, with an inclusivity of 97% and exclusivity of 98%, the specificity of this test is very high (22), a fact that was recently confirmed by Gazzola et al. (42). Because of the tight association between CC8/CLB (contagious) and adlb, the gene turned out to be a marker for contagiousness and for high prevalence of IMI in dairy herds as shown by Sartori et al. in Swiss and by Maisano et al. in Italian dairy herds (11, 16). Based on the present results we further suggest that high staphylococcal IMI prevalence is also present in Austrian, French, and German dairy herds as adlb was regularly observed in the corresponding strains. Indeed, a recent examination of an Austrian and German dairy herd with high IMI prevalence caused by S. aureus revealed again the presence of the adlb gene. Whether adlb is the only staphylococcal marker for contagiousness and high IMI prevalence remains to be elucidated. In fact, the study by Maisano et al. demonstrated that in a small percentage of herds adlb was not linked to high staphylococcal IMI prevalence (16).
Interestingly, we detected the adlb gene in a German GTII and a GTRI strain, genotypes that are not part of CLB/CC8. From ongoing studies, we know that the adlb gene is located on the staphylococcal cassette chromosome (SCC). As reviewed by Malachowa et al., SCCs may be transmitted among S. aureus strains by horizontal gene transfer; hence, the presence of adlb gene in GTII and GTRI strains may be the result of this mechanism, with an S. aureus CC8/CLB most likely being the SCC donor (43).
4.2. Prevalence of AMR in 10 European countries
In recent years, a general increase in AMR has been reported, and this increase is thought to mainly be due to AB misuse and abuse in agriculture (44, 45). In the worst-case scenario, this AMR could be transmitted to humans, which would aggravate the existing AMR situation faced in human medicine (29). Nevertheless, ABs continue to be a key factor in the treatment of bovine mastitis caused by S. aureus (11, 23, 46). Hence, it is vital to use the AB to which an isolate is fully susceptible to guarantee the successful of the therapy. According to our research, despite the large amounts of ABs that have been used to treat bovine IMIs in the past, the AMR status of S. aureus isolates from European mastitis cases is promising (47). In fact, all strains were susceptible to most of the 31 ABs tested. AMR was only observed for penicillin (25.6%) ampicillin (24.2%), chloramphenicol (9.5%), clindamycin (4.7%), and tetracyclines (5.7%). Penicillin, chloramphenicol, and tetracycline are ABs that have been widely used in cattle medicine over the past 50 years (48–50). These findings demonstrate and confirm previous observations that the regular use of ABs against S. aureus increases the possibility of the emergence of AMR (51, 52). This is in line with our observations that AMR was absent for all ABs whose application, at least in Switzerland, has not been approved for treatment of cattle (50); this is true for all the ABs on the World Health Organization (WHO) reserve list (53, 54), such as daptomycin, linezolid, and fifth-generation cephalosporins. This also holds true for most of the ABs on the WHO watch list (54) including quinolones, carbapenems, fusidic acid (one strain resistant), rifampin, teicoplanin, tobramycin (one strain resistant), and vancomycin; the exceptions were the very limited macrolide (0.9%) and tetracycline (5.7%) resistance. Interestingly, all strains were susceptible to oxacillin and all (except two strains) were susceptible to gentamicin and to trimethoprim/sulfamethoxazole. Obviously, these ABs are still efficient despite their extensive use in cattle medicine. In Switzerland, trimethoprim/sulfamethoxazole is exclusively used as a systemic treatment and is not applied intramammarily (55), so IMI-associated S. aureus strains are not in direct contact with this AB, which explain their susceptibility. This contrasts with oxacillin and gentamicin, which have been widely used for the treatment of IMIs in the past 40 years. The minimal AMR prevalence for these AB in bovine S. aureus demonstrates that the occurrence of AMRs is not only a matter of frequent use (penicillin and tetracycline). But that it considerably depends on the AB class (aminoglycosides) and even on the properties of the individual compound (oxacillin and penicillin). Considering MRSA, the present study and the one by El Garch co-authors (47) show that MRSA are of no to little concern in the field of bovine mastitis. These observations are in clear contrast to the situation in Swiss human isolates, where the prevalence of MRSA is 6.6% (56). These findings largely suggest that bovine mastitis isolates are not the source of MRSA at the human level.
With a prevalence of 25.6%, penicillin resistance was the most frequently observed type of AMR in our study. This finding aligns with the results of another European study (25.5%) (47) and of an international study (19.4%) that included strains from South America (Argentina, Brazil, and Colombia), South Africa, and the USA (57). Penicillin was introduced for the treatment of bovine mastitis as early as 1945 (58) and is still considered the AB of choice to treat Gram-positive mastitis pathogens (29), which demonstrates its importance in modern medicine.
It is worth noting that resistance to penicillin in bovine S. aureus strains can be misreported, as recently shown by Ivanovic et al. (33). Using whole genome sequencing and bioinformatics, the authors showed that the MIC assay, which was also used in the present study, provided the correct results, while analyses conducted using disk diffusion and PCR methods were remarkably flawed (33). Depending on the protocol applied, either too many false negative or false positive results were generated, and false positive results were also generated when the mPCR method was used to assed the bla operon genes (blaI, blaR1, blaZ). In the case of mPCR, it turned out that the discrepant results were always associated with S. aureus CC8/CLB strains. Further genomic analyses of these strains showed that the promoter of the bla operon present in the plasmid of the S. aureus CC8/CLB strains was inactivated by a 31-bp deletion (33); consequently, the bla operon genes that mediate penicillin resistance, were no longer expressed but could be detected by mPCR. The same association, which was explicit for the CC8/CLB strain, between negative MIC values and positive mPCR results was confirmed in the present study. Compared to the previous study (33), however, considerably more strains were evaluated here.
The present study further revealed two more very relevant findings. First, for the three major CCs (CC8, CC97, and CC705) and CLs (CLB, CLC, and CLR), penicillin resistance was highly dependent on the CC and CL. In fact, the CC705 and CLC strains were always susceptible to penicillin whereas penicillin resistance in the CC97 and CLR strains was high, at 50 and 37%, respectively. Penicillin resistance in the CC8 and CLB strains was intermediate, at 14 and 13%, respectively. Importantly, the CC705 and CLC strains were not only susceptible to penicillin but, with two exceptions, also to all other ABs, a property that was not observed for strains in the other CCs and CLs. Second, the prevalence of penicillin resistance is country dependent. Indeed, resistance to penicillin was particularly observed in strains from Austria, Belgium, Germany, and Ireland; however, it was completely absent in strains from Slovenia and Switzerland. It is likely that resistance to other ABs (i.e., chloramphenicol and tetracycline) is also country dependent, although this could not be assessed in the present study because the rate of resistance of other ABs were low and the data set was too small for statistical analyses. Unfortunately, the reason for the difference in penicillin resistance among countries remains unknown and requires further clinical and epidemiological investigations. Nevertheless, our findings demonstrate at least for penicillin, that the prevalence of AMR is country dependent and that caution is required when interpreting results. However, from a statistical and interpretative perspective there are no concerns about analyzing data from multiple-countries as a single entity. In our case, this means that, except for penicillin resistance, the observed prevalence of AMR reflects that at the European level.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
Author contributions
HG conceived and planned the experiments. GN and LR performed the experimental analyses. AR and HG performed the statistical analyses. All authors discussed the results, and critically revised and approved the final submitted manuscript.
Funding
This work was financed by the Agroscope (Bern, Switzerland) research program. Agroscope is the center of excellence for agricultural research of the Swiss administration. Open access funding by Agroscope.
Acknowledgments
The authors thank Aurélie Cosandey and Renate Boss for their valuable support and for sharing samples collected during previous studies.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fvets.2023.1154550/full#supplementary-material
References
1. Halasa T, Nielen M, De Roos AP, Van Hoorne R, de Jong G, Lam TJ, et al. Production loss due to new subclinical mastitis in dutch dairy cows estimated with a test-day model. J Dairy Sci. (2009) 92:599–606. doi: 10.3168/jds.2008-1564
2. Ruegg PL. A 100-year review: mastitis detection, management, and prevention. J Dairy Sci. (2017) 100:10381–97. doi: 10.3168/jds.2017-13023
3. Heiniger D, van den Borne BH, Lechner I, Tschopp A, Strabel D, Steiner A, et al. [Cost-benefit analysis of an intervention to improve udder health in swiss dairy farms]. Schweiz Arch Tierheilkd. (2014) 156:473–81. doi: 10.1024/0036-7281/a000634
4. Peton V, Le Loir Y. Staphylococcus aureus in veterinary medicine. Infect Genet Evol. (2014) 21:602–15. doi: 10.1016/j.meegid.2013.08.011
5. Leuenberger A, Sartori C, Boss R, Resch G, Oechslin F, Steiner A, et al. Genotypes of Staphylococcus aureus: on-farm epidemiology and the consequences for prevention of intramammary infections. J Dairy Sci. (2019) 102:3295–309. doi: 10.3168/jds.2018-15181
6. Sears PM, McCarthy KK. Management and treatment of Staphylococcal mastitis. Vet Clin North Am Food Anim Pract. (2003) 19:171–85. doi: 10.1016/S0749-0720(02)00079-8
7. Fournier C, Kuhnert P, Frey J, Miserez R, Kirchhofer M, Kaufmann T, et al. Bovine Staphylococcus aureus: association of virulence genes, genotypes and clinical outcome. Res Vet Sci. (2008) 85:439–48. doi: 10.1016/j.rvsc.2008.01.010
8. Graber HU, Naskova J, Studer E, Kaufmann T, Kirchhofer M, Brechbuhl M, et al. mastitis-related subtypes of bovine Staphylococcus aureus are characterized by different clinical properties. J Dairy Sci. (2009) 92:1442–51. doi: 10.3168/jds.2008-1430
9. Cremonesi P, Pozzi F, Raschetti M, Bignoli G, Capra E, Graber HU, et al. Genomic characteristics of Staphylococcus aureus strains associated with high within-herd prevalence of intramammary infections in dairy cows. J Dairy Sci. (2015) 98:6828–38. doi: 10.3168/jds.2014-9074
10. Capra E, Cremonesi P, Pietrelli A, Puccio S, Luini M, Stella A, et al. Genomic and transcriptomic comparison between Staphylococcus aureus strains associated with high and low within herd prevalence of intra-mammary infection. BMC Microbiol. (2017) 17:21. doi: 10.1186/s12866-017-0931-8
11. Sartori C, Boss R, Bodmer M, Leuenberger A, Ivanovic I, Graber HU. Sanitation of Staphylococcus aureus genotype B-positive dairy herds: a field study. J Dairy Sci. (2018) 101:6897–914. doi: 10.3168/jds.2017-13937
12. Sartori C, Perreten V, Ivanovic I, Hardi-Landerer MC, Graber HU. Short communication: lack of intramammary niche recolonization during a sanitation program for the contagious mastitis pathogen Staphylococcus aureus genotype B. J Dairy Sci. (2018) 101:8296–300. doi: 10.3168/jds.2017-14313
13. Voelk V, Graber HU, van den Borne BH, Sartori C, Steiner A, Bodmer M, et al. A longitudinal study investigating the prevalence of Staphylococcus aureus genotype B in seasonally communal dairy herds. J Dairy Sci. (2014) 97:4184–92. doi: 10.3168/jds.2013-7291
14. van den Borne BHP, Graber HU, Voelk V, Sartori C, Steiner A, Haerdi-Landerer MC, et al. A longitudinal study on transmission of Staphylococcus aureus genotype B in swiss communal dairy herds. Prev Vet Med. (2017) 136:65–8. doi: 10.1016/j.prevetmed.2016.11.008
15. Luini M, Cremonesi P, Magro G, Bianchini V, Minozzi G, Castiglioni B, et al. Methicillin-resistant Staphylococcus aureus (Mrsa) is associated with low within-herd prevalence of intra-mammary infections in dairy cows: genotyping of isolates. Vet Microbiol. (2015) 178:270–4. doi: 10.1016/j.vetmic.2015.05.010
16. Maisano AM, Luini M, Gazzola A, Sala L, Vezzoli F, Bertocchi L, et al. Staphylococcus aureus adlb gene is associated with high prevalence of intramammary infection in dairy herds of northern Italy: A cross-sectional study. J Dairy Sci. (2023). doi: 10.3168/jds.2022-22496. [Epub ahead of print].
17. Feil EJ, Cooper JE, Grundmann H, Robinson DA, Enright MC, Berendt T, et al. How clonal is Staphylococcus aureus? J Bacteriol. (2003) 185:3307–16. doi: 10.1128/JB.185.11.3307-3316.2003
18. Boss R, Cosandey A, Luini M, Artursson K, Bardiau M, Breitenwieser F, et al. Bovine Staphylococcus aureus: subtyping, evolution, and zoonotic transfer. J Dairy Sci. (2016) 99:515–28. doi: 10.3168/jds.2015-9589
19. Cosandey A, Boss R, Luini M, Artursson K, Bardiau M, Breitenwieser F, et al. Staphylococcus aureus genotype B and other genotypes isolated from cow milk in European countries. J Dairy Sci. (2016) 99:529–40. doi: 10.3168/jds.2015-9587
20. Schwendimann L, Merda D, Berger T, Denayer S, Feraudet-Tarisse C, Kläui AJ, et al. Staphylococcal enterotoxin gene cluster: prediction of enterotoxin (Seg and Sei) production and of the source of food poisoning on the basis of Vsaβ typing. Appl Environ Microbiol. (2021) 87:e0266220. doi: 10.1128/AEM.02662-20
21. Kläui AJ, Boss R, Graber HU. Characterization and comparative analysis of the Staphylococcus aureus genomic island Vsa: an in silico approach. J Bacteriol. (2019) 201:e00777. doi: 10.1128/JB.00777-18
22. Sartori C, Boss R, Ivanovic I, Graber HU. Development of a new real-time quantitative Pcr assay for the detection of Staphylococcus aureus genotype B in cow milk, targeting the new gene Adlb. J Dairy Sci. (2017) 100:7834–45. doi: 10.3168/jds.2017-12820
23. Algharib SA, Dawood A, Xie S. Nanoparticles for treatment of bovine Staphylococcus aureus. mastitis. Drug Deliv. (2020) 27:292–308. doi: 10.1080/10717544.2020.1724209
24. Sol J, Sampimon OC, Snoep JJ, Schukken YH. Factors associated with bacteriological cure during lactation after therapy for subclinical mastitis caused by Staphylococcus aureus. J Dairy Sci. (1997) 80:5. doi: 10.3168/jds.S0022-0302(97)76243-X
25. Sol J, Sampimon OC, Barkema HW, Schukken YH. Physiology and management factors associated with cure after therapy of clinical mastitis caused by Staphylococcus aureus. J Dairy Sci. (2000) 83:6. doi: 10.3168/jds.S0022-0302(00)74875-2
26. Gruet P, Maincent P, Berthelot X, Kaltsatos V. Bovine mastitis and intramammary drug delivery: review and perspectives. Adv Drug Deliv Rev. (2001) 50:14. doi: 10.1016/s0169-409x(01)00160-0
27. Barkema HW, Schukken YH, Zadoks RN. Invited review: the role of cow, pathogen, and treatment regimen in the therapeutic success of bovine Staphylococcus aureus mastitis. J Dairy Sci. (2006) 89:18. doi: 10.3168/jds.S0022-0302(06)72256-1
28. Pyorala S. In vitro studies as supporting evidence on the efficacy of antimicrobials for mastitis treatment. J Appl Microbiol. (2009) 107:2116–7. doi: 10.1111/j.1365-2672.2009.04538.x
29. Preine F, Herrera D, Scherpenzeel C, Kalmus P, McCoy F, Smulski S, et al. Different European perspectives on the treatment of clinical mastitis in lactation. Antibiotics. (2022) 11:1107. doi: 10.3390/antibiotics11081107
30. NMC. Bovine Mastitis Pathogens and Trends in Resistance to Antibacterial Drugs. (2004). Available online at: https://www.nmconline.org/wp-content/uploads/2020/09/Bovine-Mastitis-Pathogens-and-Trends-in-Resistance-to-Antibacterial-Drugs-FINAL-May-2020.pdf (accessed 2022).
31. Swiss Antibiotic Resistance Report 2020. Usage of Antibiotics and Occurrence of Antibiotic Resistance in Switzerland. (2020). 2020-OEG-64.
32. Molineri AI, Camussone C, Zbrun MV, Suarez Archilla G, Cristiani M, Neder V, et al. Antimicrobial resistance of Staphylococcus aureus isolated from bovine mastitis: systematic review and meta-analysis. Prev Vet Med. (2021) 188:105261. doi: 10.1016/j.prevetmed.2021.105261
33. Ivanovic I, Boss R, Romano A, Guedon E, Le-Loir Y, Luini M, et al. Penicillin resistance in bovine Staphylococcus aureus: genomic evaluation of the discrepancy between phenotypic and molecular test methods. J Dairy Sci. (2023) 106:462–75. doi: 10.3168/jds.2022-22158
34. Lowy FD. Antimicrobial resistance: the example of Staphylococcus aureus. J Clin Invest. (2003) 111:1265–73. doi: 10.1172/JCI18535
35. Llarrull LI, Toth M, Champion MM, Mobashery S. Activation of Blar1 protein of methicillin-resistant Staphylococcus aureus, its proteolytic processing, and recovery from induction of resistance. J Biol Chem. (2011) 286:38148–58. doi: 10.1074/jbc.M111.288985
36. Clarke SR, Dyke KGH. The signal transducer (Blari) and the repressor (Blai) of the Staphylococcus aureus?-lactamase operon are inducible. Microbiology. (2001) 147:7. doi: 10.1099/00221287-147-4-803
37. Breakpoint Tables for Interpretation of Mics and Zone Diameters. EUCAST (2022) (accessed January 01, 2022).
38. CLSI. M100-Performance Standards for Antimicrobial Susceptibility Testing. Wayne, PA: Informational Supplements ECaLSI (2012).
39. R Core Team. R: A Language and Environment for Statistical Computing. R 766 Foundation for Statistical Computing V. A Language and Environment for Statistical Computing. In: 766 R, editor. Vienna: Austria: Foundation for Statistical Computing (2021).
40. Yule GU. On the methods of measuring association between two attributes. J R Stat Soc. (1912) 75:73. doi: 10.2307/2340126
41. Cohen J. A coefficient of agreement for nominal scales. Educ Psychol Measur. (1960) 20:9. doi: 10.1177/001316446002000104
42. Gazzola A, Maisano AM, Bianchini V, Vezzoli F, Romanò A, Graber HU, et al. Short communication: characterization of staphylococcus aureus from bulk tank milk of dairy cattle in Lombardy (Northern Italy). J Dairy Sci. (2020) 103:2685. doi: 10.3168/jds.2019-17181
43. Malachowa N, DeLeo FR. Mobile genetic elements of Staphylococcus aureus. Cell Mol Life Sci. (2010) 67:3057–71. doi: 10.1007/s00018-010-0389-4
44. Manyi-Loh C, Mamphweli S, Meyer E, Okoh A. Antibiotic use in agriculture and its consequential resistance in environmental sources: potential public health implications. Molecules. (2018) 23:795. doi: 10.3390/molecules23040795
45. Mann A, Nehra K, Rana JS, Dahiya T. Antibiotic resistance in agriculture: perspectives on upcoming strategies to overcome upsurge in resistance. Curr Res Microb Sci. (2021) 2:100030. doi: 10.1016/j.crmicr.2021.100030
46. Ruegg PL. Making antibiotic treatment decisions for clinical mastitis. Vet Clin North Am Food Anim Pract. (2018) 34:413–25. doi: 10.1016/j.cvfa.2018.06.002
47. El Garch F, Youala M, Simjee S, Moyaert H, Klee R, Truszkowska B, et al. Antimicrobial susceptibility of nine udder pathogens recovered from bovine clinical mastitis milk in Europe 2015-2016: vetpath results. Vet Microbiol. (2020) 245:108644. doi: 10.1016/j.vetmic.2020.108644
48. Cargill CF, Bootes BW. Field observations on the control of Staphylococcal Bovine Mastitis. Aust Vet J. (1970) 46:249–52. doi: 10.1111/j.1751-0813.1970.tb15769.x
49. Third Joint Inter-Agency Report on Integrated Analysis of Consumption of Antimicrobial Agents Occurrence of Antimicrobial Resistance in Bacteria from Humans Food-Producing Animals in the Eu/Eea: Jiacra Iii 2016-2018. (2021). Available online at: https://www.ncbi.nlm.nih.gov/pubmed/34221148 (accessed October 01, 2022).
50. Umsichtiger Einsatz Von Antibiotika Bei Rindern Schweinen Kleinen Wiederkauern Und Neuweltkameliden. BLV (2022). Available online at: https://www.blv.admin.ch/blv/de/home/tiere/tierarzneimittel/antibiotika/nationale-strategie-antibiotikaresistenzen–star–/sachgemaesser-antibiotikaeinsatz.html (accessed March, 2022).
51. Prestinaci F, Pezzotti P, Pantosti A. Antimicrobial resistance: a global multifaceted phenomenon. Pathog Glob Health. (2015) 109:309–18. doi: 10.1179/2047773215Y.0000000030
52. Guo Y, Song G, Sun M, Wang J, Wang Y. Prevalence and therapies of antibiotic-resistance in Staphylococcus aureus. Front Cell Infect Microbiol. (2020) 10:107. doi: 10.3389/fcimb.2020.00107
53. Critically Important Antimicrobials for Human Medicine (6th Revision) 2018—Ranking of Medically Important Antimicrobials for Risk Management of Antimicrobial Resistance Due to Non-Human Use (2018).
54. World Health Organization. WHO Access, Watch, Reserve (AWaRe) Classification of Antibiotics for Evaluation and Monitoring of Use, 2021. World Health Organization (2021). Available online at: https://apps.who.int/iris/handle/10665/345555
55. Toxikologie IfVu. Wirkstoff: Trimethoprim - Präparate. (2022). Available online at: https://www.vetpharm.uzh.ch/Zubereitungen/000000000073/8705_99.html (accessed October 01, 2022).
56. Resistance Data Human Medicine ANRESIS. (2022). Available online at: https://www.anresis.ch/antibiotic-resistance/resistance-data-human-medicine (accessed October 01, 2022).
57. Monistero V, Barberio A, Biscarini F, Cremonesi P, Castiglioni B, Graber HU, et al. Different distribution of antimicrobial resistance genes and virulence profiles of Staphylococcus aureus. strains isolated from clinical mastitis in six countries. J Dairy Sci. (2020) 103:3431–46. doi: 10.3168/jds.2019-17141
Keywords: Staphylococcus aureus, adlb, antimicrobial resistance, minimum inhibitory concentration, multidrug resistant
Citation: Nemati G, Romanó A, Wahl F, Berger T, Rojo LV and Graber HU (2023) Bovine Staphylococcus aureus: a European study of contagiousness and antimicrobial resistance. Front. Vet. Sci. 10:1154550. doi: 10.3389/fvets.2023.1154550
Received: 30 January 2023; Accepted: 04 April 2023;
Published: 03 May 2023.
Edited by:
Kohei Makita, Rakuno Gakuen University, JapanReviewed by:
Amjad Islam Aqib, Cholistan University of Veterinary and Animal Sciences, PakistanAneliya Milanova, Trakia University, Bulgaria
Copyright © 2023 Nemati, Romanó, Wahl, Berger, Rojo and Graber. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ghazal Nemati, Z2hhemFsLm5lbWF0aUBhZ3Jvc2NvcGUuYWRtaW4uY2g=
†These authors share first authorship