
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Vet. Sci. , 23 June 2023
Sec. Veterinary Infectious Diseases
Volume 10 - 2023 | https://doi.org/10.3389/fvets.2023.1153069
This article is part of the Research Topic Host response to Veterinary Infectious Diseases: Role of Coding and Non-coding RNAs as Biomarkers and Disease Modulators View all 7 articles
 María Augusta Chávez-Larrea1,2Cristina Cholota-Iza2Jorge Cueva-Villavicencio2Michelle Yugcha-Díaz2Jorge Washington Ron-Román3
María Augusta Chávez-Larrea1,2Cristina Cholota-Iza2Jorge Cueva-Villavicencio2Michelle Yugcha-Díaz2Jorge Washington Ron-Román3 Andrea Rodríguez-Cabezas2
Andrea Rodríguez-Cabezas2 Claude Saegerman1
Claude Saegerman1 Armando Reyna-Bello2*
Armando Reyna-Bello2*Trypanosoma theileri is a worldwide distributed haemoparasite that has been reported throughout the American continent in various species, including bovines, buffaloes and bats. In bovines, high incidence of T. theileri can be harmful when associated with other infections or under stress situations. There is little information on this hemoflagellate in Ecuador, which prompted the study and molecular identification of the trypanosomes collected in two slaughtering centers. Between February and April 2021, a total of 218 samples of bovine blood were collected in abattoirs located in the Andean region of Quito (n = 83) and in the coastal region, in Santo Domingo (n = 135). Quito public Slaughterhouse is the biggest in Ecuador, and for that, they receive animals from all country; on the other hand, Santo Domingo's Slaughterhouse is a small one where mainly females from the region are sacrificed and some males. The samples were evaluated using two molecular tests, the PCR cathepsin L-like (CatL) specific for T. theileri and for the positive samples, a Nested PCR that targets the ITS of the 18S gene. The corresponding PCR products were sequenced, analyzed by BLAST/NCBI and the sequences were used to build a concatenated phylogenetic tree, using the MEGA XI software. Overall, 34 out of the 218 samples, (15.6%) were positive to T. theileri by PCR CatL, resulting from 20/83 (24.1%) positives from the Quito abattoir and 14/135 (10.4%) from the Santo Domingo slaughterhouse. These prevalence rates were found to be significantly different (p = 0.006). According to the phylogenetic tree based on the CatL and ITS concatenated sequences (n = 13), the two novel Equatorial T. theileri isolates, ThI (n = 7) and ThII (n = 6) are closely related and associated to the IC, IB and IIB genotypes, present in Brazil, Venezuela and Colombia. Thirty-one out of the thirty-four T. theileri-positive bovines were co-infected with other haemotropic pathogens, Anaplasma marginale Babesia spp and T. vivax. This coinfection could be responsible for additional pathologies and harmful effects on the affected cattle. This study presents the molecular identification and genotypification of T. theileri isolated from cattle in Ecuador through the analysis of CAtL and ITS sequences, and the high frequency of coinfection of this hemoflagellate with other blood haemotropic organisms.
In Ecuador, livestock production represents an important component of its economy with a 7.7% contribution to the Gross Domestic Product (GDP), and an estimated bovine population of 4.34 million that is distributed in four regions: 41.24% in the Coast region, 46.11% in the Mountain range (Andes), 9.65% in the Amazon and 0.43% in the Insular or Galapagos region (1). The division of Ecuador in four natural regions, coastal, Andes, Amazon (2) and Insular (Galapagos Islands) influences the distribution and management of various bovine breeds, as well as disease prevalence and disease risk factors (3).
Bovine trypanosomosis is a hemoparasitic disease distributed throughout parts of the African and American continents. In Latin-America, Trypanosoma vivax, Trypanosoma evansi and Trypanosoma theileri are the main trypanosome species that affect bovines (4). T. theileri is the least studied since it has been considered a nonpathogenic parasite (5). However, some recent studies show that T. theileri is an opportunistic parasite that can cause anemia, fever, swollen lymph nodes and lower hemoglobin concentration (6, 7).
Trypanosoma theileri has been classified within the Megatrypanum subgenera. It is larger than T. evansi and T. vivax, reaching a length between 69 to 109 μm it. This parasite has a free flagellum, with a well-developed undulating membrane and its posterior part is conical (4). The sequencing and subsequent transcriptome of T. theileri has revealed greater proximity to Trypanosoma cruzi and Trypanosoma rangeli, in the Stercoraria clade, as compared to T. brucei and T. vivax, which belong to the Salivarian clade. Like other trypanosomes, T. theileri persists in the host for a long time due to an efficient evasion mechanism that involves the synthesis of five diverse families of GPI-anchored surface proteins with conserved N- and C-terminal and over 1,000 genes that encode surface proteins distinct from the Variable Surface Glycoprotein (VSG) characteristic of the Salivarian group (8).
T. theileri has a cosmopolitan distribution throughout the world from Asia to America (5), infecting different species of the Order Artiodactyl, especially cattle and buffaloes (9). In Colombia, a northern neighbor country to Ecuador, the reported prevalence values for T. theileri appear to be higher in cattle (38.6 and 50.9%) than in buffaloes (28.2%) (9, 10). Lower prevalence values have been reported by Pacheco et al. (11), using a PCR based on Cathepsin L-like gene (PCR-TthCATL) in Brazil, with prevalence values of 42.19% per farm and 12.19% per animal.
The transmission of T. theileri can occur cyclically, mainly by horseflies, where the infective form is found in the intestine. This hemoflagellate like others within the Stercoraria group, is transmitted to the vertebrate host through contamination of wounds caused by horsefly bites (4, 10). Studies in Poland and Germany showed that 33.68 and 39% of T. theileri infected horseflies, respectively (12, 13). In Brazil, the reported prevalence of T. theileri in horseflies was 40 and 70% in two different geographical areas. In Ecuador there are few studies concerning tabanid fauna, however, in 2009 Cárdenas et al. (14) revealed a high density and species diversity for the country. In addition, iatrogenic and mechanical transmission by Phlebotomus (10, 15) and Aedes mosquitoes has also been reported (16).
The presence of trypanosomosis represents a limiting factor for livestock productivity due to economic losses attributed mainly to low milk production (17).
Despite this, there is little information on bovine trypanosomosis in Ecuador; however, Coello Peralta et al. found 20% positive samples of Trypanosoma spp. in sheep blood smears (18). Other studies by Medina-Naranjo et al. found a seroprevalence of 31.3% for Trypanosoma spp. using ELISA in the province of Pastaza (Amazon region) (19). In 2020 the first report of T. vivax was made in an outbreak in cattle in the Canton El Carmen, in the Manabí province, in the coastal region (20), and, regarding T. theileri, this hemoflagellate was recently described in the Ecuadorian Amazon region, with a prevalence of 11.4% (21).
Other homeotropic agents have also been reported in Ecuador; A. marginale was first described in the Santo Domingo Province after analysis by PCR of 151 blood samples, which resulted in a high, 86.1% rate of rickettsia infection (22). In Zamora-Chinchipe (South-Eastern Ecuador, close to Peru), a 68.8% prevalence was reported, indicating the widespread distribution and genetic variability of this bacterial pathogen in the country (21%) (23).
Even in the Galapagos Islands, this Rickettsia has been described, with a prevalence higher than 90%, which indicates the endemic nature of anaplasmosis in the islands (24). Regarding bovine babesiosis, Chávez-Larrea et al. (25) determined the presence of Babesia spp. by PCR in 18.94% (14.77% Babesia bovis and 4.17% Babesia bigemina) of twenty farms around El Carmen at 300 m.a.s.l.. Curiously, in this study, they found a prevalence of 20.28% (14.69% B. bovis and 5.59% B. bigemina) in Quito at 2469 m. a. s. l., demonstrating the adaptation of the vectors at higher altitudes (25).
The identification of the Trypanosoma species and its relationship with other haemotropic that are present in Ecuador, their distribution and prevalence are important aspects to clarify their epidemiology and to set the basis to implement timely and adequate diagnostic protocols and treatments. Similar studies led to the description of a novel trypanosome species, Trypanosoma (Megatrypanum) trinaperronei n. sp. in the white-tailed deer in Venezuela (26). For this reason, the present study focused, on the molecular identification of T. theileri in cattle originating in two slaughterhouses from the Pichincha and Santo Domingo de los Tsachilas provinces and its relationship with other haemotropic.
To study the presence of Trypanosoma theileri in Ecuador, blood specimens were collected in the Quito and Santo Domingo slaughterhouses (Figure 1). Three interventions were done in the Quito slaughterhouses between February and March 2020, while four interventions were performed in the Santo Domingo abattoir between March and April 2021.
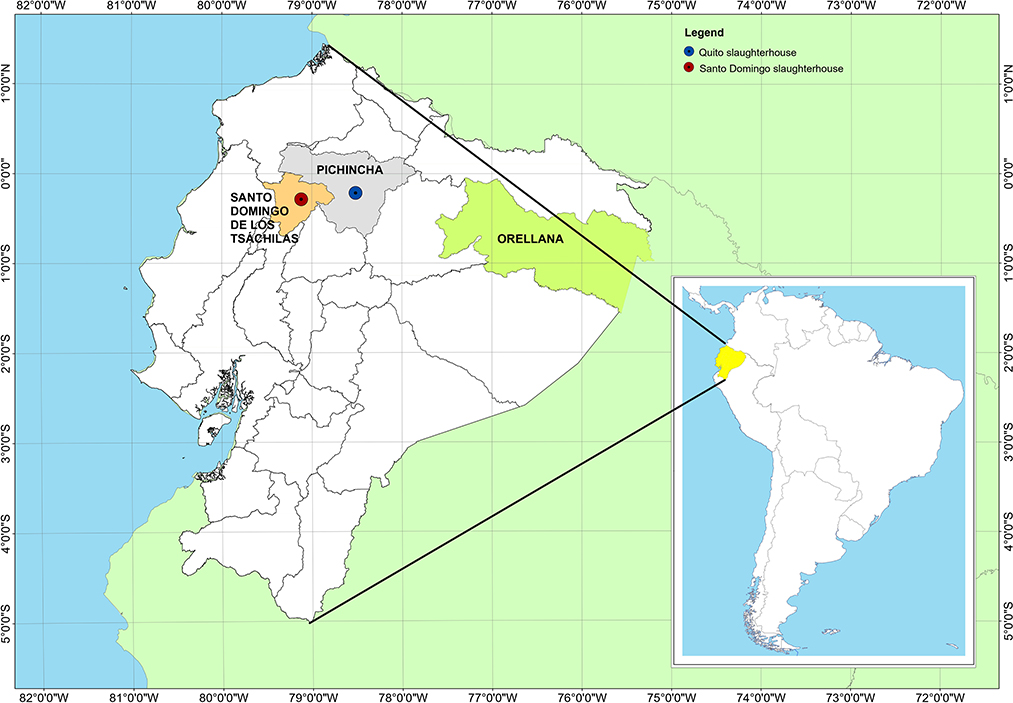
Figure 1. Location of the slaughterhouses of Quito and Santo Domingo and Provincia de Orellana in Ecuador.
The Quito slaughterhouse is located in the capital city of Quito, in the Pichincha province, belonging to the Sierra (Andes) region, at 2.850 m above sea level. The Santo Domingo slaughterhouse is in the Santo Domingo de los Tsachilas province, 133 km from the city of Quito, at 655 m above sea level, in the Coastal zone (Figure 1).
A total of 218 cattle were sampled, including two species: Bos indicus and Bos taurus. In the slaughterhouses of Quito, n = 83, and Santo Domingo, n = 135. Blood samples were collected in 10 mL tubes with EDTA from the jugular vein at the time of the sacrifice of the animals. Depending on the number of animals slaughtered in the Quito slaughterhouse (27), one out of 10 animals was selected for each intervention. Given the low number of animals slaughtered daily, one out of every two animals was sampled for each intervention in the Santo Domingo slaughterhouse.
The Quito slaughterhouse is one of the largest in Ecuador, where many male and female animals are slaughtered. On the other hand, the Santo Domingo slaughterhouse is small, where mainly females from surrounding areas are killed. The males in the area are transferred to the slaughterhouse in Quito or another private.
Samples were temporarily stored at room temperature while transferred to the laboratory and stored at−20°C at the Animal Biotechnology Laboratory of the Universidad de las Fuerzas Armadas ESPE and assigned identification numbers with information on sex and breed.
The PCV value of the blood samples was determined using capillary tubes with heparin (TECNAN, Navarra, Spain). The tubes were centrifuged (TG12M Madell Technology Corporation, Riverside, California, USA) at 12.000 rpm for 5 min. To evidence the parasite in the blood, within the first 4 h of sample collection, the heparinized capillary tubes were observed under a microscope (KRUSS, Hamburg, Germany) according to the HCT technique (28).
Deoxyribonucleic Acid (DNA) extraction from blood samples was performed using the Genejet Whole Blood Genomic DNA Purification Mini Kit (Thermo Scientific, Waltham, Massachusetts, U.S.). The DNA integrity was verified on a 0.8% agarose gel and DNA concentration was quantified by UV spectrophotometry, using the NanoDrop 2000 (Thermo Fisher Scientific, Waltham, Massachusetts, USA).
Several primer sets were used to diagnose haemotropic pathogens (Table 1) and all PCR reactions were carried out in 25 μl reaction volumes, using the Proflex Thermal Cycler (Life Technologies, Carlsbad, California, U.S.A). The positive controls for T. theileri were the samples that tested positive for HCT. On the other hand, for Babesia, T. vivax, and A. margianle, previously prepared plasmids were used (29–31).

Table 1. List of primers used for molecular analysis by PCR for the detection of haemoparasites in cattle.
A specific catalytic domain of cathepsin L-like (CATL-like) PCR, that amplified a partial sequence of the CATL gene (29), was performed to detect the presence of T. theileri, using between 100 and 150 ng of DNA, as modified by Yokoyama et al. (36). Samples that were positive for the CATL-like PCR were analyzed with a Nested PCR based on the internal transcribed spacers (ITS) region, as described by Cox et al. (31), except for the temperature of annealing which was increased 59°C to improve specificity. This genetic marker was used to distinguish the T. theileri lineages and phenotypes. The first-round reaction mixture for the nested PCR contained 100–150 ng of DNA and the external primers ITS1 and ITS2, while the second round used 1 μL of the PCR product from the first reaction and the ITS3 and ITS4 internal primers (Table 1).
All positive samples for the CATL-like PCR were further analyzed with different PCR assays to determine coinfection. For T. vivax, a specific CATL-like PCR was performed (20, 30), for T. evansi, the ESAG primer set was used (32), for A. marginale the msp5 PCR was utilized (22, 33), while B. bovis and B. bigemina were diagnosed by RAP-1 PCR and HYP PCR, respectively (34, 35).
Positive PCR products for CATL-like PCR specific for T. theileri and positive PCR products for ITSs Nested PCR, were sequenced in (Macrogen, Seoul, Korea), using the Sanger technique. The consensus sequences were deposited in GenBank (Accession numbers for CATL-like: OQ304106, OQ304107, OQ304108, OQ304109, OQ304110, OQ304111, OQ304112, OQ304113, OQ304114, OQ304115, OQ304116, OQ304117, OQ304118 and Accession numbers for ITS: OQ341204, OQ341205, OQ341206, OQ341207, OQ341208, OQ341209, OQ341210, OQ341211, OQ341212, OQ341213, OQ341214, OQ341215, OQ3412166). The similarity of the consensus sequences obtained for both the CATL-like and the ITS were analyzed with the BLAST tool (Basic Local Alignment Search Tool the National Center for Biotechnology Information). Phylogenetic relationships were based on the sequences obtained in this study and on previously described sequences available in the GenBank database (9, 30, 37). The construction of a Maximum Likelihood tree, Tamura-Nei model (38) was carried out using the MEGAXI program.
A Microsoft Excel sheet was used to organize, clean, and validate the laboratory results. Significant differences in the distribution of T. theileri lineages and biotypes between the two abattoirs, between individuals of different sex and animal breeds, were analyzed by the Fisher's exact test, a statistical test used to determine nonrandom associations between a small group of observations. Bovine origin was excluded in the analysis of the Santo Domingo abattoir, since that information was not considered reliable.
Out of the 218 analyzed samples, 38.1% were collected in the Quito slaughterhouse, while the remaining 62% were from the Santo Domingo slaughterhouse. Regarding the origin of the animals, we relied only on the information collected in the Quito slaughterhouse, where 11/83 (13.3%) of the animals originated in the Santo Domingo Province, 13/83 (15.7%) in the Orellana province, and 59/83 (71.1%) with unknown provenance. The results of the diagnostic tests regarding the sex variable, were only analyzed for the Quito slaughterhouse, where the sampling included similar percentages of males (44.6%) and females (55.4%). By contrast, an unintentional bias was introduced in the sampling of slaughter groups at the Santo Domingo abattoir, with a very low percentage of males (3%), as compared to females (97%).
From the 218 total bovine samples, 13 (6.0%) were positive with Trypanosoma spp. by the HCT technique, corresponding to a significant difference (p < 0.05) in the prevalence (14.5%) in the Quito abattoir, as compared to the prevalence (0.74%) in the Santo Domingo slaughterhouse. The average values of the packed cell volume (PCV) for the tested animals and those positive to T. theileri were 37.7 and 38.42%, with standard deviations of 10.0 and 7.6%, respectively.
The CATL-like PCR test identified 34 samples positive to T. theileri (15.6%), 20 (24.1%) originating in the Quito abattoir and 14 (10.4%) in the Santo Domingo slaughterhouse, with a significant difference between these locations (p = 0.006). Among the 20 samples that were positive by CATL-like PCR in the Quito abattoir, there were 10 males (27.0%) and 10 females (21.7%), which resulted in no significant difference in the sex variable (p > 0.05). In the Quito abattoir, no statistically significant differences were found among the bovines positive to T. theileri and the two species that were examined, Bos indicus and Bos taurus.
The use of the CATL-like PCR showed that 7 of the 13 HCT-positive bovines, corresponded to T. theileri infections and 5 to T. vivax infections. Through the use of molecular tests, the prevalence of five haemotropic agents was obtained: T. theileri (15.6%), T. vivax (7.3%), A. marginale (60.1%), B. bovis (3.2%) and B. bigemina (2.3%). No animals were positive to T. evansi, using the ESAG PCR. Table 2 presents distribution details of the positive cases of the various haemopathogens that were analyzed, regarding the two abattoirs, sex and species of the sampled individuals.
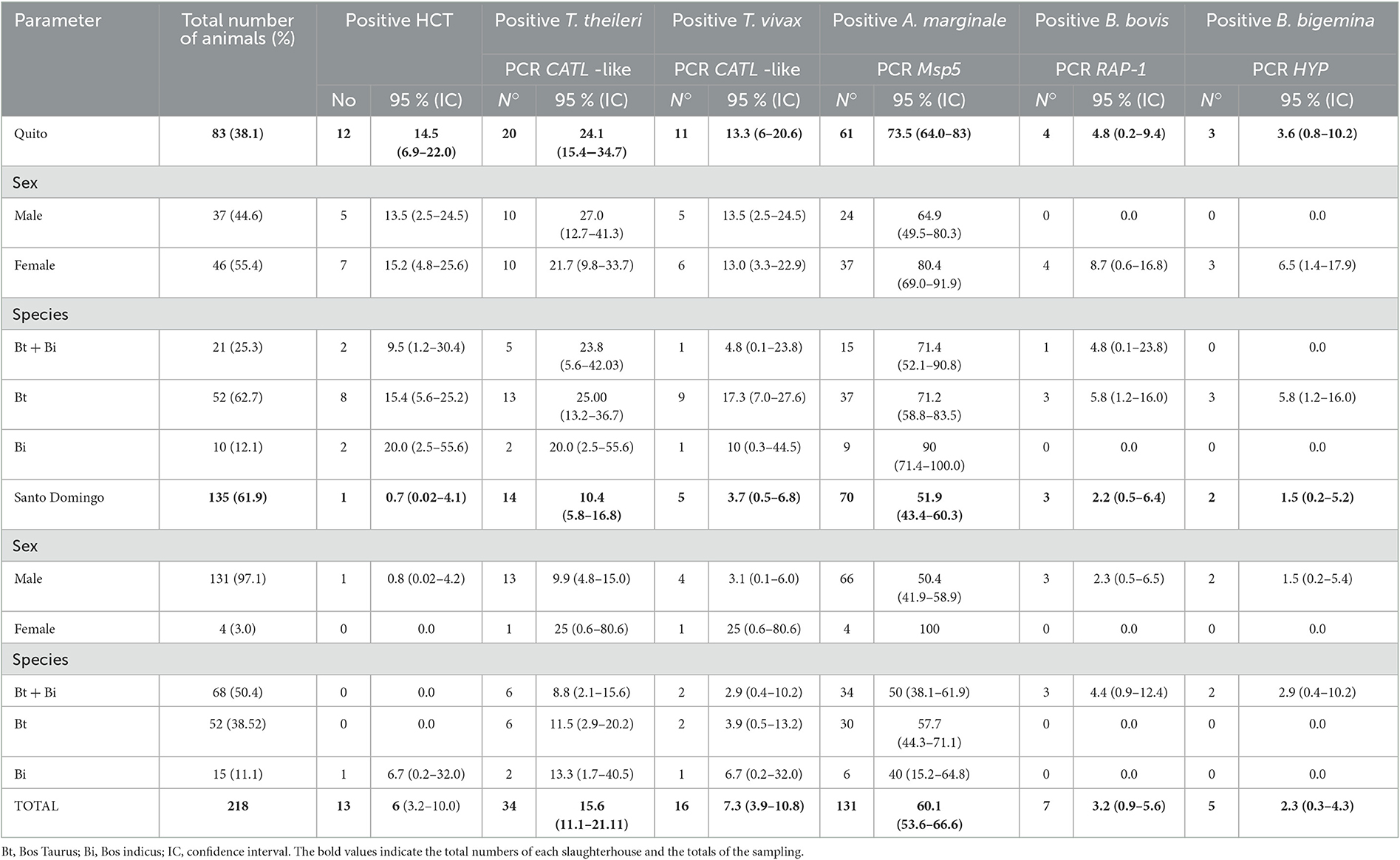
Table 2. Distribution of positive cases for T. theileri and other haemopathogens in the slaughterhouses of Quito and Santo Domingo.
All the T. theileri positive bovines (n = 20) in the Quito slaughterhouse were coinfected, 15 (18.1%) with A. marginale and 5 (6.0%) with T. vivax and A. marginale. With regard to the Santo Domingo slaughterhouse 11 of the 14 bovine samples that were positive to T. theileri showed coinfection; 6 (4.4%) with A. marginale, 2 (1.5%) with T. vivax and 3 (2.2%) with T. vivax and A. marginale (Table 3).
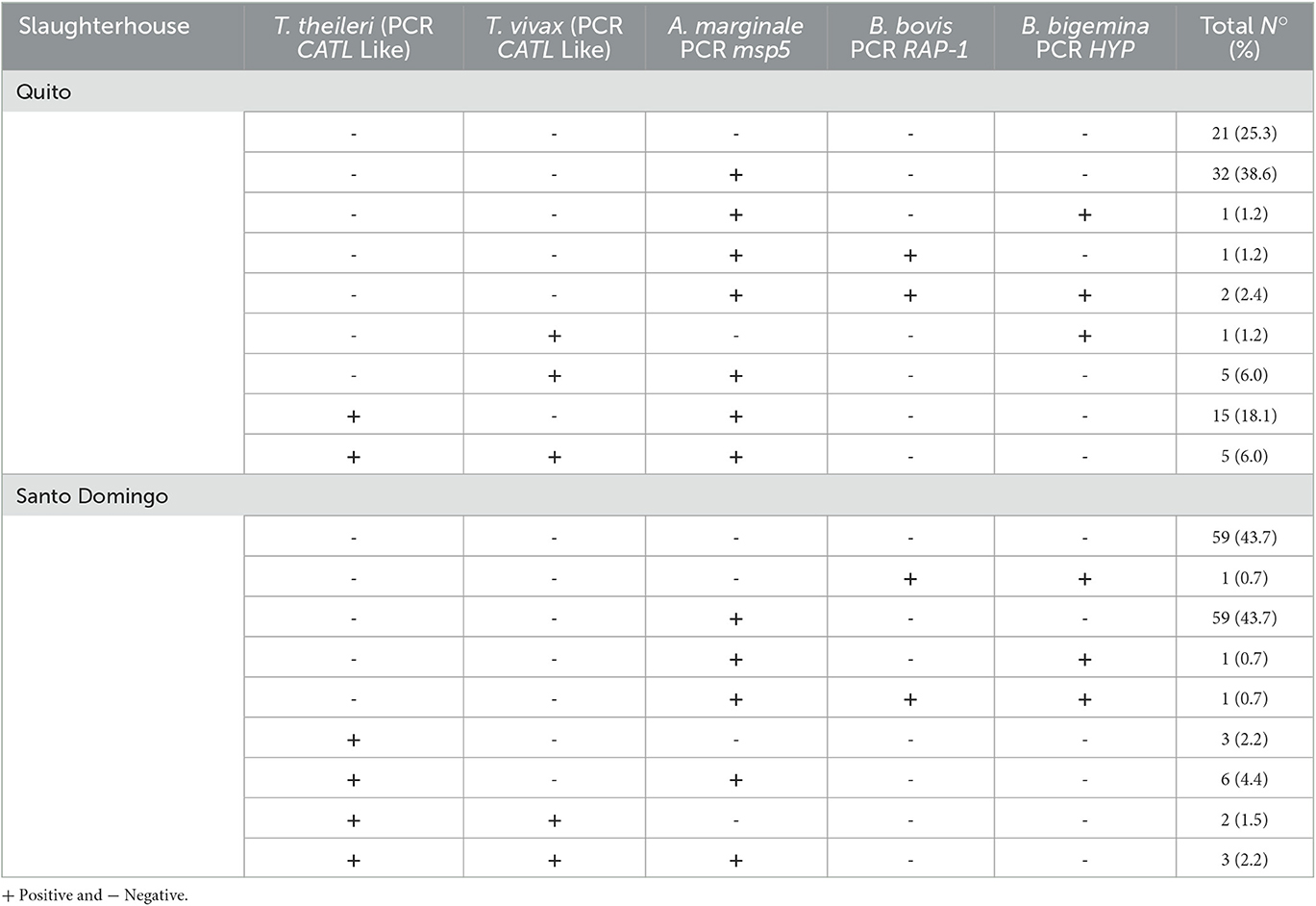
Table 3. Details of T. theileri coinfections with other haemopathogens in the slaughterhouses of Quito and Santo Domingo.
The phylogenetic tree constructed from the 13 best quality sequences (Figure 2) of the CATL-like and ITS PCR amplicons (n = 24), showed that the T. theileri isolates cluster within the ThI (n = 7) and ThII (n = 6) lineages. In the Quito slaughterhouse, two samples belong to the IB genotype and three to the IC genotype, within the TthI lineage, while five samples belonged to the IIB genotype in the TthII lineage. On the other hand, in the Santo Domingo slaughterhouse, two samples aligned with genotype IC of the TthI lineage, and one sample with the IIB genotype of the TthII lineage. The phylogeographic tree (Figure 2) showed that seven of the isolates, from Quito (n = 5) and Santo Domingo (n = 2), clustered within the Brazil clades; while the 6 remaining isolates, from Quito (n = 5) and Santo Domingo (n = 1), clustered with the Venezuelan and Colombia clades.

Figure 2. Maximum Parsimony Phylogram built with concatenated sequences of CatL gene and ITS region of Trypanosoma theileri, Lineages: TthI and TthII. Genotypes: IA, IIA, IB, IIB, IC, IIC. The DNAs obtained from cattle of two slaughterhouses in Ecuador in bold.
Out of the 13 samples that were analyzed for lineages and genotypes, 12 showed coinfection with T. vivax and/or A. marginale. Further details of the lineages-biotypes and coinfection with T. vivax, A. marginale, B. bovis and B. bigemina are presented in Table 3.
As for the origin of the animals from the Quito slaughterhouse, it was possible to determine that TthI-IB (n = 2) and TthI-IC (n = 2) originated from the province of Santo Domingo de los Tsachilas, while the animals with the genotype TthII-IIB (n = 5) originated from the province of Orellana.
This study evidenced the presence of T. theileri in bovines from two slaughterhouses in Ecuador, using a molecular test (CATL-like PCR). Significant prevalence values of 24.1 and 10.4% for T. theileri were determined for the Quito and the Santo Domingo abattoirs, located in the Andean and Coastal regions, respectively.
Prevalence values for T. theileri in South America are high, suggesting that this parasite is widely distributed in the cattle production systems in this region. The global prevalence values (15.6%) determined in the present study is congruent with reports from some farms in neighboring countries, including 5.0 and 30.4% in Venezuela and Brazil, respectively (39), 8.1% in Brazil (11), and 38.6 to 50.3% in Colombia (6, 9). The high prevalence values for T. theileri in other countries have been related to environmental conditions, host, and parasite factors (9). This report of the presence of T. theileri in cattle in Ecuador highlights the importance of these neglected diseases and the need to focus more attention and resources on haemopathogen agents that affect the cattle industry in this country and region.
Prevalence values obtained from the two slaughterhouses, in Quito and Santo Domingo, evidenced the presence of several haemopathogens, including in decreasing order: A. marginale (60.1%, the most predominant), T. theileri (15.6%), T. vivax (7.3%), Babesia bovis (3.2%), B. bigemina (2.3%) and T. evansi (0 %). Our study did not evidence T. evansi in any of the bovine samples from the two slaughterhouses. These values are compatible with previous studies in Ecuador, 31.0% for Trypanosoma spp and 65.5% for A. marginale in the Pastaza province (19) and 86.1% in the Santo Domingo de los Tsachilas province (22), 18.94 and 20.28% for Babesia spp in the Pichincha and Manabí provinces, respectively (25).
The implementation of molecular diagnostic tests such as the CATL-like PCR for T. theileri and T. vivax and msp5 PCR for A. marginale showed mixed infections of T. theileri with A. marginale, as well as T. theileri, T. vivax and A. marginale in the Quito (n = 20) and Santo Domingo (n = 14) abattoirs. Coinfections with other haemotropics have been reported in Colombia, where 53.9% of the bovines present mixed infections and 26.7% presented coinfections of T. theileri with A. marginale (6). In another study, 83.3% of the bovines positive to T. theileri showed coinfection with other haemopathogens (36). All the animals infected with the genotyped T. theileri carried mixed infections, which reinforces the hypothesis that we are in the presence of an opportunistic parasite (9).
Although this study did not present a clinical assessment of the animals in the abattoirs, PCV analysis showed normal levels, above 24%, in all the bovine specimens, including those in the animals positive to T. theileri. This is in agreement with previous reports that show that bovines infected with T. theileri do not suffer from anemia and have normal levels of erythrocytes (40, 41). However, in spite of this, T. theileri can be considered a potentially pathogenic parasite, in association with other haemopathogens, as evidenced in a livestock area in Colombia, where T. theileri infection was higher in bovines with signs of anemia (9). Furthermore, T. theileri infections may result in chronic and mild clinical signs, with low and persistent parasitemias (8), as could be the case for the bovines that were included in this study, whose hematocrit levels were above 24%. These different pieces of evidence highlight the need to further investigate the possible link between of coinfection with T. theileri and other haemopathogens and clinical disease.
The prevalence of T. theileri based on the HCT (14.5 %) and PCR CATL-like (24.1%) tests was higher in the Quito abattoir as compared to the Santo Domingo slaughterhouse, with 0.7 and 10.4% prevalence values, respectively. The bovines positive to T. theileri (20/83), in the Quito abattoir originated in the Santo Domingo province in the Coastal region and in the Orellana province in the Amazon region. The difference in the distribution of the positive results of HCT suggests that bovine trypanosomosis in the Coastal region is endemic, unlike the Amazon region where disease outbreaks can occur. This proposal is reinforced by the recent finding of a large percentage of Trypanosoma spp. positive farms (8/32) in the Amazon region, using the HCT (Chávez- Larrea et al. data not published). This could be related with the great diversity of animal reservoirs, both livestock and/or wildlife in the Amazon region.
Statistical analysis of the T. theileri positive animals from the Quito slaughterhouse showed no significant differences concerning bovine species, nonetheless significant differences were observed in relationship to sex, with a higher prevalence in males. Even though creole cattle breeds could be a risk factor associated with the presence of Trypanosoma spp., as suggested by Jaimes-Dueñez et al. (42) in Colombia studying creole races, additional research is needed to clarify the role of other risk factors, including sex and host species.
T. theileri has been reported in various animal species including bovines, buffaloes (Asia, South America), antelopes (Africa) (4), horse in Malaysia (43) and bats (44), with several lineages and genotypes. The implementation of molecular diagnostic techniques using CatL and ITS fragments, followed by DNA sequencing and analysis have been previously used to analyze T. theileri genotypes (39, 40, 45). The two T. theileri lineages, Tth I and TthII identified in this study have been reported in other South American countries, including Colombia (9), Brazil and Venezuela (29, 39, 46). Several genotypes have been identified within these lineages as specific for each host species, such is the case for bovines (IIA, IB, IIB, IC), buffaloes (IA) and deers (IIC) (9, 29, 46). Genotypes IB, IIB and IC, which have been previously described in Brazil, Venezuela and Colombia (9, 11, 39) were identified in this study. Genotype IIB was the most prevalent in the samples that were analyzed in this study (6/13), which is also consistent with the most prevalent genotype in South America (9, 39). Likewise, in the study carried out by De la Cadena et al. (21), in the Ecuadorian Amazon, they point out the presence of the TthI and TthII genotypes using the cathepsin L-like and 18S ribosomal DNA for the phylogenetic tree.
The T. theileri sequences from the Quito and Santo Domingo slaughterhouses were placed within the clade of the Colombian isolates. It is possible that cattle displacement between the two countries might be responsible for the introduction of T. theileri to Ecuador. Very little is known about the origin of the first bovines that arrived in Ecuador. According to a study on the indigenous breeds, the first bovines being introduced in the XV century during the Spanish colonization, with some were introduced from the Pacific coast of Colombia and Panama and some from Rio de la Plata (47). This mobilization of cattle appears to have influenced the distribution of genotype IIB, which is the most disseminated in South America from cattle of Iberian origin (9). The other genotypes found in our study, IB and IC, like those found in Brazil, maybe were introduced more recently (9).
In Ecuador, the regions below 1,000 masl are found both in the coastal and Amazon regions. These two zones are separated by the Sierra region (Los Andes), where the heights range from 2,000 to 4,000 meters above sea level. For this reason, in this work, only animals positive for T. theileri were found in the coastal region (Province Santo Domingo de los Tsachilas) and the Amazon region (Province Orellana) (Figure 1).
In America, the transmission of Trypanosoma spp in bovine cattle is mainly mechanical, through Tabanids and other blood-sucking flies (4). Tabanids also appear to play an essential role in the transmission of A. marginale too (48). In the present study, the estimated prevalence for A. marginale was 38.6% in the Quito slaughterhouse and 43.70% in the Santo Domingo slaughterhouse. Most of the animals were coinfected with this bacteria, possibly due to high prevalence of A. marginale all over the country. There have not been any studies in Ecuador on the importance of Tabanids and other blood-sucking flies as vectors for these diseases in livestock. The presence of the T. theileri genotypes has also been shown in Tabanids, which reinforces their role in the dissemination of Trypanosoma spp in America (46).
This study constitutes the first report of lineages TthI and TthII and genotypes IB, IC and IIB of T. theileri in bovine cattle in Ecuador (Figure 2), a finding that could be important to establish the genetic relationship between the variants present in this country and others in the region. On the other hand, existing coinfections between T. theileri, T. vivax, A. marginale, B. bovis and B. bigemina were also evaluated at the slaughterhouse level.
This study underscores the presence of T. theileri in livestock in Ecuador, with a prevalence of 15.6% (34/218). Even though, T. theileri can be considered non-pathogenic or mildly pathogenic, further studies are needed to evaluate the epidemiological situation in the region and its true impact on the health of bovines and other susceptible species, especially in case of coinfection with other haemopathogens.
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
Ethical review and approval was not required for the animal study because the work was carried out with animals from slaughterhouses, after their sacrifice. Written informed consent was obtained from the owners for the participation of their animals in this study.
Conceptualization: MC-L, CS, and AR-B. Methodology, data curation, and writing—original draft preparation: MC-L. Validation: MC-L, JC-V, AR-C, and CC-I. Formal analysis: MC-L, CC-I, and JR-R. Investigation: MC-L and AR-B. Writing—review and editing: AR-B, JR-R, and CS. Supervision: AR-C and CS. Project administration: JR-R and AR-B. Funding acquisition: CS, JR-R, and AR-B. All authors have read and agreed to the published version of the manuscript.
This work was funded by the Academy of Research and Higher Education (ARES) Belgica and Universidad de las Fuerzas Armadas ESPE-Ecuador through the project entitled Establishment of a platform to support training and awareness, diagnosis and development of a control strategy for brucellosis and trypanosomosis in Ecuador (acronym: BruTryp), Registered in the ESPE under the number CV-GNP-0056-2020.
We thank the managers of the Quito and Santo Domingo slaughterhouse, who allowed the collection of bovine blood. We are also thankful to the undergraduate students of Biotechnology of the ESPE University, in Quito and Santo Domingo, for their work collecting data in the slaughterhouses. Finally, we thank the Universidad de las Fuerzas Armadas, ESPE, for supporting and stimulating us in this work.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Instituto Nacional de Estadisticas y Censos Ecuador INEC. Encuesta de Superficie y Producción Agropecuaria Continua (ESPAC). Quito-Ecuador: INEC (2019). p. 33.
2. Galeas R, Guevara J, Medina Torres B, Chinchero MA, Herrera X. Sistema de Clasificacion de Ecosistemas del Ecuador Continental. 1re éd. Quito-Ecuador: Sistema Nacional de Información (2013). p. 239 (Ministerio del Ambiente del Ecuador. Subsecretaría de Patrimonio Natural, Quito; vol. 1).
3. Ron-Román J, Ron-Garrido L, Abatih E, Celi-Erazo M, Vizcaíno-Ordóñez L, Calva-Pacheco J, et al. Bayesian evaluation of three serological tests for detecting antibodies against Brucella spp. among humans in the northwestern part of Ecuador. Am J Trop Med Hyg. (2019) 100:1312–20. doi: 10.4269/ajtmh.18-0622
4. Desquesnes M. Livestock trypanosomoses and their vectors in Latin America. Paris: OIE (2004). p. 174.
5. Hoare CA. The trypanosomes of mammals. A zoological monograph. The trypanosomes of mammals A zoological monograph. (1972). Available online at: https://www.cabdirect.org/cabdirect/abstract/19722902365 (accessed September 2, 2022).
6. Jaimes-Dueñez J, Triana-Chávez O, Mejía-Jaramillo AM. Parasitological and molecular surveys reveal high rates of infection with vector-borne pathogens and clinical anemia signs associated with infection in cattle from two important livestock areas in Colombia. Tick Tick Borne Dis. (2017) 8:290–9. doi: 10.1016/j.ttbdis.2016.12.002
7. Hajihassani A, Maroufi S, Esmaeilnejad B, Khorram H, Tavassoli M, Dalir-Naghadeh B, et al. Hemolytic anemia associated with Trypanosoma theileri in a cow from Kurdistan province, West of Iran. Vet Res Forum. (2020) 11:191–3. doi: 10.30466/vrf.2019.103834.2465
8. Kelly S, Ivens A, Mott GA, O'Neill E, Emms D, Macleod O, et al. An alternative strategy for trypanosome survival in the mammalian bloodstream revealed through genome and transcriptome analysis of the ubiquitous bovine parasite Trypanosoma (megatrypanum) theileri. Genome Biol Evol. (2017) 9:2093–109. doi: 10.1093/gbe/evx152
9. Jaimes-Dueñez J, Triana-Chávez O, Mejía-Jaramillo AM. Spatial-temporal and phylogeographic characterization of Trypanosoma spp. in cattle (Bos taurus) and buffaloes (Bubalus bubalis) reveals transmission dynamics of these parasites in Colombia. Vet Parasitol. (2018) 249:30–42. doi: 10.1016/j.vetpar.2017.11.004
10. Calzolari M, Rugna G, Clementi E, Carra E, Pinna M, Bergamini F, et al. Isolation of a trypanosome related to Trypanosoma theileri (kinetoplastea: Trypanosomatidae) from Phlebotomus perfiliewi (diptera: Psychodidae). Biomed Res Int. (2018) 2018:2597074. doi: 10.1155/2018/2597074
11. Pacheco TDA, Marcili A, Costa AP da, Witter R, Melo ALT, Boas RV, et al. Genetic diversity and molecular survey of Trypanosoma (Megatrypanum) theileri in cattle in brazil's western amazon region. Rev Bras Parasitol Vet. (2018) 27:579–83. doi: 10.1590/s1984-296120180049
12. Böse R, Heister NC. Development of Trypanosoma (M.) theileri in Tabanids. J Eukaryot Microbiol. (1993) 40:788–92. doi: 10.1111/j.1550-7408.1993.tb04475.x
13. Werszko J, Szewczyk T, Steiner-Bogdaszewska Z, Wróblewski P, Karbowiak G, Laskowski Z. Molecular detection of Megatrypanum trypanosomes in tabanid flies. Med Vet Entomol. (2020) 34:69–73. doi: 10.1111/mve.12409
14. Cárdenas RE, Buestán J, Dangles O. Diversity and distribution models of horse Flyes (diptera: Tabanidae) from Ecuador. Int J Entomol. (2009) 45:511–28. doi: 10.1080/00379271.2009.10697633
15. Desquesnes M, Gonzatti M, Sazmand A, Thévenon S, Bossard G, Boulangé A, et al. A review on the diagnosis of animal trypanosomoses. Parasites Vectors déc. (2022) 15:64. doi: 10.1186/s13071-022-05190-1
16. Brotánková A, Fialová M, Cepička I, Brzonová J, Svobodová M. Trypanosomes of the Trypanosoma theileri group: phylogeny and new potential vectors. Microorganisms. (2022) 10:294. doi: 10.3390/microorganisms10020294
17. Rashid M, Rashid MI, Akbar H, Ahmad L, Hassan MA, Ashraf K, et al. A systematic review on modelling approaches for economic losses studies caused by parasites and their associated diseases in cattle. Parasitology. (2019) 146:129–41. doi: 10.1017/S0031182018001282
18. Coello-Peralta RD, Cabrera Solórzano NC, Salazar Mazamba MdL, Reyes Echeverria EO, Andrade Velásquez LL, Pazmiño Gómez BJ. Natural infection of Trypanosoma sp. in domestic sheep from Ecuador. Ciencia Rural. (2022) 9. doi: 10.1590/0103-8478cr20210141
19. Medina-Naranjo VL, Reyna-Bello A, Tavares-Marques LM, Campos AM. Diagnóstico de Los Hemotrópicos Anaplasma marginale, Trypanosoma Spp. y Babesia Spp. mediante Las Técnicas de ELISAi y PCR En Tres Fincas Ganaderas de La Provincia de Pastaza, Ecuador. Revista Científica. (2017) 27:162–71.
20. Chávez-Larrea MA, Medina-Pozo ML, Cholota-Iza CE, Jumbo-Moreira JR, Saegerman C, Proaño-Pérez F, et al. First report and molecular identification of Trypanosoma (Duttonella) vivax outbreak in cattle population from Ecuador. Transbound Emerg Dis. (2021) 68:2422–8. doi: 10.1111/tbed.13906
21. De la Cadena E, Camacho M, Vaca F, Enríquez S, Eleizalde MC, Arrivillaga-Henríquez J, et al. Molecular identification of Trypanosoma theileri in cattle from the Ecuadorian Amazon. Vet Parasitol Reg Stud Rep. (2023) 37:100824. doi: 10.1016/j.vprsr.2022.100824
22. Tana-Hernández L, Navarrete-Arroyo K, Ron-Román J, Reyna-Bello A, Chávez-Larrea MA. PCR-diagnosis of Anaplasma marginale in cattle populations of Ecuador and its molecular identification through sequencing of ribosomal 16S fragments. BMC Vet Res. (2017) 13:392. doi: 10.1186/s12917-017-1311-1
23. Guarnizo TRM, Alvarez DO, Díaz-Sánchez AA, Cabezas-Cruz A, Gutiérrez LZ, Marrero SM, et al. Epidemiology and genetic diversity of Anaplasma marginale in Zamora-Chinchipe, Ecuador. Ticks Tick Borne Dis. (2020) 11:101380. doi: 10.1016/j.ttbdis.2020.101380
24. Gioia GV, Vinueza RL, Marsot M, Devillers E, Cruz M, Petit E, et al. Bovine anaplasmosis and tick-borne pathogens in cattle of the Galapagos Islands. Transbound Emerg Dis. (2018) 65:1262–71
25. Chávez-Larrea MA, Cholota-Iza C, Medina-Naranjo V, Yugcha-Díaz M, Ron-Román J, Martin-Solano S, et al. Detection of Babesia spp. in high altitude cattle in ecuador, possible evidence of the adaptation of vectors and diseases to new climatic conditions. Pathogens. (2021) 10:1593. doi: 10.3390/pathogens10121593
26. Garcia HA, Blanco PA, Rodrigues AC, Rodrigues CMF, Takata CSA, Campaner M, et al. Pan-American Trypanosoma (Megatrypanum) trinaperronei n. sp. in the white-tailed deer odocoileus virginianus zimmermann and its deer ked lipoptena mazamae rondani, 1878: morphological, developmental and phylogeographical characterisation. Parasit Vectors. (2020) 13:308. doi: 10.1186/s13071-020-04169-0
27. Guillen MF. Informe de Rendicion de cuentas. Quito-Ecuador: Empresa Pública Metropolitana de Rastro Quito (2020). p. 39. Report No.: 1.
28. OIE. Trypanosomosis Animales (transmitidas por la mosca tse-tse, excepto la surra y la durina). In: Manual Terrestres de la OIE. OIE (2018). p. 15.
29. Rodrigues AC, Garcia HA, Ortiz PA, Cortez AP, Martinkovic F, Paiva F, et al. Cysteine proteases of Trypanosoma (Megatrypanum) theileri: cathepsin L-like gene sequences as targets for phylogenetic analysis, genotyping diagnosis. Parasitol Int. (2010) 59:318–25. doi: 10.1016/j.parint.2010.03.002
30. Cortez AP, Rodrigues AC, Garcia HA, Neves L, Batista JS, Bengaly Z, et al. Cathepsin L-like genes of Trypanosoma vivax from Africa and South America–characterization, relationships and diagnostic implications. Mol Cell Probes. (2009) 23:44–51. doi: 10.1016/j.mcp.2008.11.003
31. Cox A, Tilley A, McOdimba F, Fyfe J, Eisler M, Hide G, et al. A PCR based assay for detection and differentiation of African trypanosome species in blood. Exp Parasitol. (2005) 111:24–9. doi: 10.1016/j.exppara.2005.03.014
32. Holland WG, Claes F, My LN, Thanh NG, Tam PT, Verloo D, et al. A comparative evaluation of parasitological tests and a PCR for Trypanosoma evansi diagnosis in experimentally infected water buffaloes. Vet Parasitol. (2001) 97:23–33. doi: 10.1016/S0304-4017(01)00381-8
33. Reyna-Bello A, Cloeckaert A, Vizcaíno N, Gonzatti MI, Aso PM, Dubray G, et al. Evaluation of an enzyme-linked immunosorbent assay using recombinant major surface protein 5 for serological diagnosis of bovine anaplasmosis in Venezuela. Clin Diagn Lab Immunol. (1998) 5:259–62. doi: 10.1128/CDLI.5.2.259-262.1998
34. Figueroa JV, Chieves LP, Johnson GS, Buening GM. Multiplex polymerase chain reaction based assay for the detection of Babesia bigemina, Babesia bovis and Anaplasma marginale DNA in bovine blood. Vet Parasitol. (1993) 50:69–81. doi: 10.1016/0304-4017(93)90008-B
35. Figueroa JV, Chieves LP, Johnson GS, Buening GM. Detection of Babesia bigemina-infected carriers by polymerase chain reaction amplification. J Clin Microbiol. (1992) 30:2576–82. doi: 10.1128/jcm.30.10.2576-2582.1992
36. Yokoyama N, Sivakumar T, Fukushi S, Tattiyapong M, Tuvshintulga B, Kothalawala H, et al. Genetic diversity in Trypanosoma theileri from Sri Lankan cattle and water buffaloes. Vet Parasitol. (2015) 207:335–41. doi: 10.1016/j.vetpar.2014.12.006
37. Jackson AP, Berry A, Aslett M, Allison HC, Burton P, Vavrova-Anderson J, et al. Antigenic diversity is generated by distinct evolutionary mechanisms in African trypanosome species. Proc Natl Acad Sci U S A. (2012) 109:3416–21. doi: 10.1073/pnas.1117313109
38. Kumar S, Stecher G, Li M, Knyaz C, Tamura K. MEGA X: molecular evolutionary genetics analysis across computing platforms. Mol Biol Evol. (2018) 35:1547–9. doi: 10.1093/molbev/msy096
39. Garcia HA, Kamyingkird K, Rodrigues AC, Jittapalapong S, Teixeira MMG, Desquesnes M. High genetic diversity in field isolates of Trypanosoma theileri assessed by analysis of cathepsin L-like sequences disclosed multiple and new genotypes infecting cattle in Thailand. Vet Parasitol. (2011) 180:363–7. doi: 10.1016/j.vetpar.2011.03.017
40. Amato B, Mira F, Di Marco Lo Presti V, Guercio A, Russotto L, Gucciardi F, et al. A case of bovine trypanosomiasis caused by Trypanosoma theileri in Sicily, Italy. Parasitol Res. (2019) 118:2723–7. doi: 10.1007/s00436-019-06390-y
41. Bittner L, Krämer K, Wöckel A, Snedec T, Delling C, Böttcher D, et al. Malnutrition as the cause of recumbency in suckler cows associated with Trypanosoma theileri infection. Acta Vet Scand. (2021) 63:2. doi: 10.1186/s13028-020-00567-7
42. Jaimes-Dueñez J, Mogollón-Waltero E, Árias-Landazabal N, Rangel-Pachon D, Jimenez-Leaño A, Mejia-Jaramillo A, et al. Molecular surveillance of Trypanosoma spp. reveals different clinical and epidemiological characteristics associated with the infection in three creole cattle breeds from Colombia. Prev Vet Med. (2021) 193:105414. doi: 10.1016/j.prevetmed.2021.105414
43. Tan LP, Mohd Rajdi NZI, Mohamad MA, Mohamed M, Hamdan RH, Goriman Khan MAK, et al. First report of Trypanosoma theileri in equine host and tabanus sp. in Malaysia. J Equine Vet Sci. (2021) 108:103807. doi: 10.1016/j.jevs.2021.103807
44. Jaimes-Dueñez J, Cantillo-Barraza O, Triana-Chávez O, Mejia-Jaramillo AM. Molecular surveillance reveals bats from eastern Colombia infected with Trypanosoma theileri and Trypanosoma wauwau-like parasites. Prev Vet Med. (2020) 184:105159. doi: 10.1016/j.prevetmed.2020.105159
45. Suganuma K, Kondoh D, Sivakumar T, Mizushima D, Elata ATM, Thekisoe OMM, et al. Molecular characterization of a new Trypanosoma (Megatrypanum) theileri isolate supports the two main phylogenetic lineages of this species in Japanese cattle. Parasitol Res. (2019) 118:1927–35. doi: 10.1007/s00436-019-06313-x
46. Rodrigues AC, Paiva F, Campaner M, Stevens JR, Noyes HA, Teixeira MMG. Phylogeny of Trypanosoma (Megatrypanum) theileri and related trypanosomes reveals lineages of isolates associated with artiodactyl hosts diverging on SSU and ITS ribosomal sequences. Parasitology. (2005) 132:215. doi: 10.1017/S0031182005008929
47. Delgado Bermejo JV, Barba Capote CJ, Aguirre Riofrío EL, Cabezas Congo R, Cevallos Falquez OF, Rizzo Zamora LG, et al. Molecular inference in the colonization of cattle in Ecuador. Res Vet Sci. (2020) 132:357–68. doi: 10.1016/j.rvsc.2020.07.011
48. Hornok S, Földvári G, Elek V, Naranjo V, Farkas R, de la Fuente J. Molecular identification of Anaplasma marginale and rickettsial endosymbionts in blood-sucking flies (Diptera: Tabanidae, Muscidae) and hard ticks (Acari: Ixodidae). Vet Parasitol. (2008) 154:354–9. doi: 10.1016/j.vetpar.2008.03.019
Keywords: bovine trypanosomosis, Trypanosoma theileri, lineage (ThI-Th-II), cattle, Ecuador, Anaplasma marginale, Babesia, Trypanosoma vivax
Citation: Chávez-Larrea MA, Cholota-Iza C, Cueva-Villavicencio J, Yugcha-Díaz M, Ron-Román JW, Rodríguez-Cabezas A, Saegerman C and Reyna-Bello A (2023) Molecular identification of Trypanosoma theileri (Laveran, 1902) in cattle from two slaughterhouses in Ecuador and its relation with other haemotropic agents. Front. Vet. Sci. 10:1153069. doi: 10.3389/fvets.2023.1153069
Received: 28 January 2023; Accepted: 30 May 2023;
Published: 23 June 2023.
Edited by:
Mohamed Samir, The Pirbright Institute, United KingdomReviewed by:
Alireza Sazmand, Bu-Ali Sina University, IranCopyright © 2023 Chávez-Larrea, Cholota-Iza, Cueva-Villavicencio, Yugcha-Díaz, Ron-Román, Rodríguez-Cabezas, Saegerman and Reyna-Bello. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Armando Reyna-Bello, YWFyZXluYUBlc3BlLmVkdS5lYw==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.