
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Vet. Sci. , 05 July 2023
Sec. Animal Nutrition and Metabolism
Volume 10 - 2023 | https://doi.org/10.3389/fvets.2023.1149514
This article is part of the Research Topic New Advances in Functional Aquafeeds View all 6 articles
 Khang Wen Goh1
Khang Wen Goh1 Zulhisyam Abdul Kari2,3*
Zulhisyam Abdul Kari2,3* Wendy Wee4
Wendy Wee4 Nik Nur Azwanida Zakaria3,5
Nik Nur Azwanida Zakaria3,5 Mohammad Mijanur Rahman2,3Muhammad Anamul Kabir6
Mohammad Mijanur Rahman2,3Muhammad Anamul Kabir6 Noor Khalidah Abdul Hamid7
Noor Khalidah Abdul Hamid7 Albaris B. Tahiluddin8
Albaris B. Tahiluddin8 Ahmad Syazni Kamarudin9Guillermo Téllez–Isaías10
Ahmad Syazni Kamarudin9Guillermo Téllez–Isaías10 Lee Seong Wei2,3*
Lee Seong Wei2,3*Edwardsiellosis caused by Edwardsiella tarda resulted in significant economic losses in aquaculture operations worldwide. This disease could infect a wide range of hosts, including freshwater, brackish water, and marine aquatic animals. Currently, antibiotics and vaccines are being used as prophylactic agents to overcome Edwardsiellosis in aquaculture. However, application of antibiotics has led to antibiotic resistance among pathogenic bacteria, and the antibiotic residues pose a threat to public health. Meanwhile, the use of vaccines to combat Edwardsiellosis requires intensive labor work and high costs. Thus, phytobiotics were attempted to be used as antimicrobial agents to minimize the impact of Edwardsiellosis in aquaculture. These phytobiotics may also provide farmers with new options to manage aquaculture species' health. The impact of Edwardsiellosis in aquaculture worldwide was elaborated on and highlighted in this review study, as well as the recent application of phytobiotics in aquaculture and the status of vaccines to combat Edwardsiellosis. This review also focuses on the potential of phytobiotics in improving aquatic animal growth performance, enhancing immune system function, and stimulating disease resistance.
Nowadays, food security is a major concern throughout the world. Aquaculture can provide a reliable and affordable protein source for human consumption (1). This statement was supported by the data recorded that fish consumption per capita was 9 kg annually in 1961. The value increased rapidly to 20.5 kg in 2018 (2). The aquaculture industry is gearing up to fulfill the increasing demand for fish protein in the market. However, issues such as high stocking density and water quality have led to disease outbreaks (3), resulting in low production, poor growth performance, and a high mortality rate. Additionally, these issues also result in high operational costs and food insecurity and affect investors' income (4). Fish mortality rates due to disease outbreaks as high as 50% reported in developing countries have led some fish farmers to abandon their aquaculture operations. Based on the World Bank report, the disease outbreak caused approximately USD 6 billion in economic losses annually (5). The outbreaks may be due to pathogenic bacteria and stressful environmental conditions. Consequently, fish farmers had no option but to continue using antibiotics as a treatment against disease outbreaks (6).
Edwardsiella tarda was first reported in the literature by a Japanese scientist in 1962. This Gram-negative anaerobic facultative Brevibacterium infects a huge range of hosts, such as aquatic animals, amphibians, reptiles, and mammals throughout the world (7). E. tarda has infected various aquaculture species and has led to a huge economic loss (8, 9). Hemolysin is an important virulence factor of E. tarda that causes septicemia in the host (10). Other virulence factors that are responsible and involved in the infection process are catalase (11), Translocation and Assembly Module (Tam) (12), DNA-binding protein from starved cells (Dps) (13), undecaprenyl phosphate gylcosyltransferase (WcaJ) (14), and superoxide dismutase (15). The pathogenicity and virulence of E. tarda were reported due to the presence of virulence genes in the bacterium, namely, vibrioferrin synthesis (pvsA), sensor protein (qseC), chondroitinase (cds1), AHL-synthase (edwI), and DNA Gyrase (gyrB) (8, 16, 17).
The symptoms of Edwardsiellosis in infected fish species, such as hybrid snakehead (Channa maculate ♀ × Channa argus ♂) and grass carp (Ctenopharyngodon idella), are exophthalmia, hernia, internal organ damage (18), pigment loss, swollen anus, and enlarged kidney (19). Other symptoms reported in the literature are ascites and internal organ swelling (20). At present, antibiotics are used to lessen the impact of Edwardsiellosis on aquaculture species. However, the excessive use of antibiotics has led to an increment in antibiotic resistance cases against pathogenic E. tarda (21–24). For instance, E. tarda isolated from Siamese crocodile was found to be highly resistant to erythromycin, tetracycline, and oxytetracycline (25). Turbot farming in China was reported to rely on antibiotics and chemicals to combat Edwardsiellosis infection (26). In Korea, E. tarda isolated from farmed marine fishes was reported to be resistant to various antibiotics, such as streptomycin, cefaclor, lincomycin, penicillin, erythromycin, and rifampin (27). The application of antibiotics in aquaculture can control bacterial infection in the short term. However, adverse effects of using antibiotics as treatment include bioaccumulation of the antibiotic residues in aquatic animal tissues and organs, immunosuppression, and imbalance of gut microbiota (Figure 1) (28). The plasmid in E. tarda was found to carry antibiotic-resistance genes against multi-antibiotics (29). The application of antibiotics in aquaculture has led to the contamination of antibiotic residues in the human food chain (24). Furthermore, over usage of antibiotics in aquaculture can accelerate the emergence of multi-antibiotic-resistant pathogenic bacteria that can adversely affect public health (30). Therefore, there is a need to find alternative antimicrobial agents to control Edwardsiellosis in aquaculture to reduce overreliance on chemicals and antibiotics. This review discusses and summarizes the impact of Edwardsiellosis due to E. tarda, the application of phytobiotics in aquaculture, the status of the Edwardsiellosis vaccine, and the roles of phytobiotics in improving growth performance, enhancing the immune system, and stimulating disease resistance against E. tarda.

Figure 1. The impacts of antibiotics used in aquaculture.
Phytobiotics are plant-based derivatives that have beneficial effects on organisms. The bioactive compounds are responsible for the biological activities of phytobiotics, such as alkaloids, carotenoids, and phenolic compounds (31). The biological activities of phytobiotics can be anti-inflammatory, antimicrobial, antioxidant, and others. Generally, phytobiotics play an important role in promoting the growth of gut microbiota, increasing feed efficiency, and activating immune-related genes to enhance the immune system of fish (31). For example, Brown alga, Ecklonia cava, was found to promote the growth of probiotic lactic acid bacteria (LAB) in zebrafish and modulate the immune system of the fish against Edwardsiellosis infection (32).
E. tarda is an important disease-causing bacterium in aquaculture (33). This bacterium is under the genus of Edwardsiella. There are another four pathogenic bacteria under similar genera, namely, Edwardsiella. anguillarum (34), Edwardsiella piscicida (35), Edwardsiella ictaluri, and Edwardsiella hoshinae. E. tarda is a short rod-shaped Gram-negative bacterium with a diameter of 1–3 μm in length (36, 37). Based on the phenotypes, E. tarda can be divided into two groups, namely, typical and atypical (38). Typical and atypical groups are referred to as motile and non-motile E. tarda, respectively. The bacterium can be grouped into four serotypes (i.e., A, B, C, and D). The serotype grouping is based on the agglutination of the bacterium with specific antisera to identify variants of somatic (O) and flagella (H) antigens. This bacterium is responsible for Edwardsiellosis disease outbreaks in many fish farming. For instance, the Edwardsiellosis outbreak was reported in carp species, such as crucian carp in Japan (39) and grass carp in India (19, 40). Besides carp, Edwardsiellosis also infected Japanese eels in Fujian Province in China (41), giant mottled eels in China (42), and Japanese eels in South Korea (43). Edwardsiellosis has caused high mortality of hybrid snakeheads in China (18). Many cases of Edwardsiellosis outbreaks were recorded in olive flounder (Figure 2) and Japanese flounder farms in China (44–46). Furthermore, Edwardsiellosis has infected Chinook salmon in the US (47), Sharpsnout seabreams in Greece (48), Rainbow trout in Korea (49), Dabry sturgeon in China (50), Yellow catfish in China (51), Black rockfish in China (52), Chinese tongue sole (53) in China, Seahorse (54) in China, Siamese crocodile in Hainan and China (25), and Chinese soft-shelled turtle in China (55). E. tarda caused ascites disease in juvenile turbot (Scophtalmus maximus), which led to high mortality (30–50%) (56). In some cases, ascites in turbot have led to massive mortality, as high as 90% (41). As a result, Edwardsiellosis poses a significant threat to turbot farming, especially in producing seeds. Besides, E. tarda is also responsible for gangrene in fish, red disease in eels, emphysematous putrefactive disease in catfish (36), and fatal septicemia in newly hatched farmed crocodiles (57, 58). In the US, Edwardsiellosis is a major disease in farmed channel catfish, Ictalurus punctatus, and farmed barramundi, Lates calcarifer (59). Furthermore, Edwardsiellosis is a well-known disease in Egypt during summer that has caused huge mortality in Nile tilapia (60). The impact of Edwardsiellosis infection in fish organs is shown in Table 1 (61). This disease can spread through contaminated feed, water, or intestinal mucosa, and a poor environment, such as the presence of high organic, poor water quality, and high temperature can trigger Edwardsiellosis infection in fish (61). Overall, Edwardsiellosis is an important disease in aquaculture that has a huge economic impact. Edwardsiellosis-causing bacterium, E. tarda, can adapt to a wide range of environments and infect various hosts resulting in high mortality. Edwardsiellosis outbreak devastates many fish farmers causing them to sometimes end the farm's operation. In addition, investors also lose their income and many workers become jobless.

Figure 2. Edwardsiellosis infection in olive flounder, Paralichthys olivaceus (9).
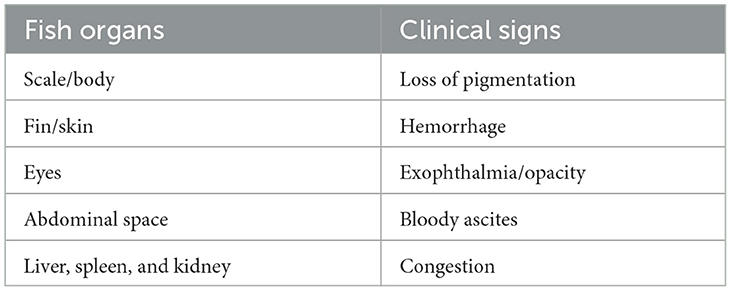
Table 1. The impacts of Edwardsiellosis on fish organs.
Phytobiotics are referred to as any plant-based products that have antimicrobial activity (31, 62) and antioxidant capacity, can promote growth performance (63), enhance the immune system (64), stimulate disease resistance, and mitigate stress. There are some phytobiotics have been approved by the U.S. Food and Drug Administration (FDA) to be used in animal production (65) such as cottonseed meal and rice hull. However, both phytobiotics cannot be applied more than 20% in the feed formulation. Different phytobiotics vary in their modes of action depending on the bioactive component. Generally, phytobiotics can modulate gut microbiota and change the membrane permeability of pathogenic microorganisms. However, the effectiveness of phytobiotics can be influenced by many factors, such as storage conditions, post-harvesting processing, geographical locations, and plant species (66, 67).
The potential of phytobiotics (e.g., essential oil, plant leaves, flowers, and alcoholic extracts) usage in aquaculture was widely documented (Table 2). Phytobiotics were used as feed additives in aquaculture, and some were used as a solution and medicated through bathing treatment. The exposure period of aquaculture species to phytobiotics ranges from 14 to 60 days. Generally, the modes of action of phytobiotics are to promote the growth of gut microbiota, increase feed utilization efficiency, and activate immune-related genes to produce antimicrobial production (68–72) (Figure 3). Based on the phytobiotics' mode of action, the beneficial effects of phytobiotics on aquaculture species include enhancing the immune system, increasing antioxidant activity, improving growth performance, and stimulating disease resistance of aquaculture species. Besides, phytobiotics can also be used as alternate commercial antibiotics, acting as antimicrobial agents and mitigating abiotic stress such as ammonia.
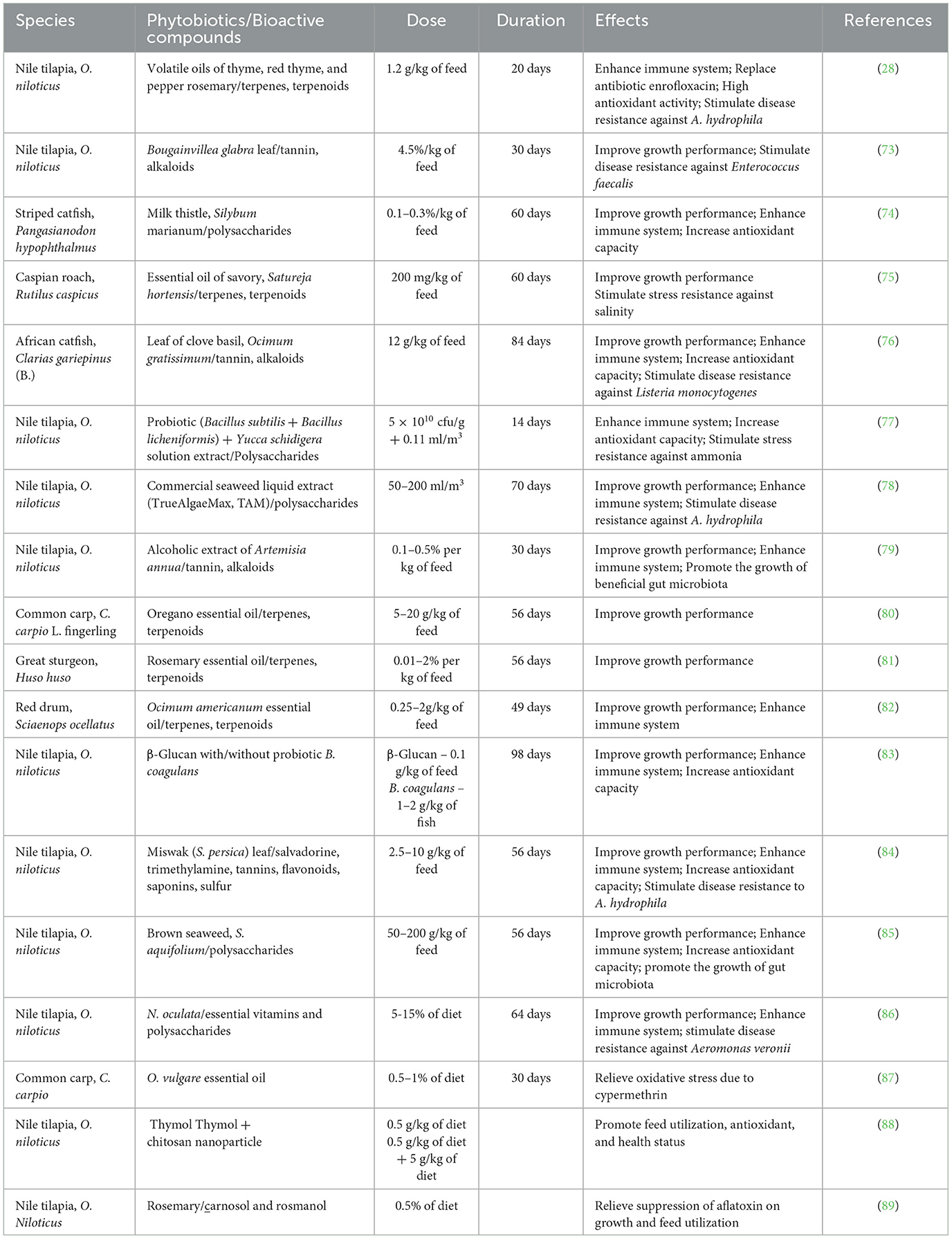
Table 2. Phytobiotics used in aquaculture.
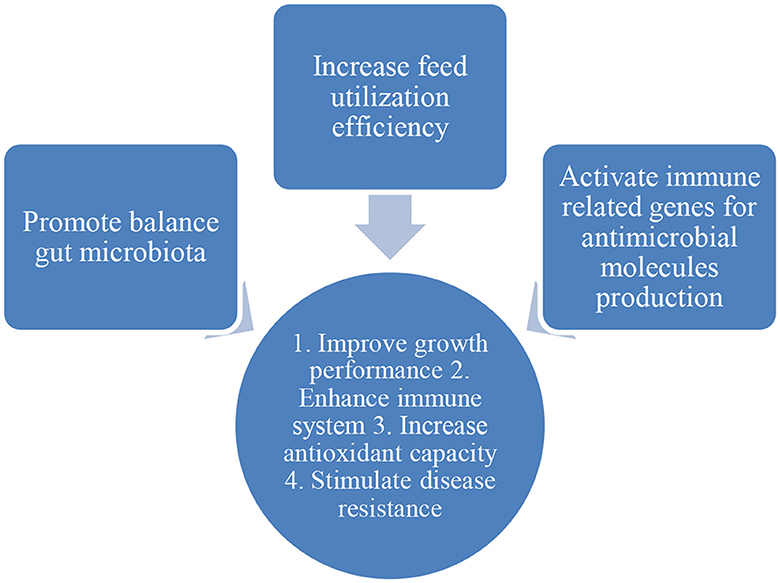
Figure 3. Mode of action of phytobiotics.
β-Glucan is a commercial polysaccharide that can be used as an immunostimulant. It was reported to increase the binding activity of receptors with natural killer cells and neutrophils (83). Hence, β-glucan can enhance the immune system. Many studies revealed that β-glucan could improve the immune system in aquatic animals, such as Oreochromis niloticus (83), Litopenaeus vannamei (90), Oncorhynchus mykiss (91), Lutjanus peru (92), Cyprinus carpio (93), and Trachinotus ovatus (94). In addition, a combination of probiotic Bacillus coagulans and β-glucan can perform a synergistic effect to enhance the immune system of O. niloticus (83). Dietary Miswak (Salvadora persica) leaf in Nile tilapia, O. niloticus, was found to have beneficial effects, such as growth performance improvement, immune system enhancement, antioxidant increment, and Aeromonas hydrophila disease resistance stimulation (84). The beneficial effects were linked to the bioactive compounds present in the phytobiotic, such as alkaloids comprising salvadorine, trimethylamine, tannins, flavonoids, saponins, and sulfur (95). These bioactive compounds have been stated to promote feed consumption, relieve stress, and act as immunostimulants (84). Besides, dietary of Miswak root was also reported to promote an immune system of common carp (96) and tilapia (97). Nile tilapia, O. niloticus that received brown seaweed Sargassum aquifolium in diet performed significantly better in growth and health (85). Polysaccharides, the bioactive compounds that were present in brown seaweed, were responsible for the positive response in Nile tilapia, O. niloticus. The polysaccharides can promote the growth of gut microbiota (32) and activate gene-related antimicrobial production molecules (98). Thus, dietary brown seaweed can promote feed utilization efficiency and the health status of Nile tilapia.
The potential of microalga, Nannochloropsis oculata, as a phytobiotic was revealed in the study of Abdelghany et al. (86). Dietary N. oculata at doses of 5–15% of the diet was found to promote the growth and health status of Nile tilapia, O. niloticus. Microalgae are widely used in aquaculture as they are rich in essential amino acids. Besides, they also carry bioactive compounds such as essential vitamins and polysaccharides that can fortify health status and promote growth performance (108). However, the application of N. oculata as a feed additive at higher doses may lead to disruption of nutrient digestion activity (109), and the presence of complexed non-starched polysaccharides, such as pectins, gums, cellulose, and hemicelluloses, can reduce nutrient absorption in the fish digestive system (110). Therefore, N. oculata must use in the optimal range to avoid adverse impacts on the fish. A diet of rosemary (Rosmarinus officinalis) was found to relieve aflatoxin B1-suppressed growth and feed utilization in Nile tilapia (89). Rosemary is a well-known herb for its high antioxidant activity (111). Polyphenol compounds are the main bioactive compounds in rosemary that are responsible for their antioxidant property (112). Carnosol and rosmanol are two bioactive compounds that are present in rosemary. These bioactive compounds can enhance nutrient digestibility and inhibit pathogenic bacteria in the intestine (113).
Dietary Origanum vulgare essential oil at the dose of 0.5–1% of the diet was reported to relieve oxidative stress due to the presence of insecticide, cypermethrin, in common carp, C. carpio (87). The bioactive compounds, which are present in the essential oil, such as carvacrol, thymol, cymene, and terpinene, are able to increase the antioxidative capacity of fish (80, 114). Besides, O. vulgare essential oil was also reported to mitigate oxidative stress due to carbon tetrachloride (115), gentamycin (116), and paraquat (117) in rats. Thymol is another phytobiotic that was reported to have a positive impact on aquaculture species. However, the application of thymol alone showed no significant impact on the growth performance in rainbow trout (118), channel catfish (119), and Nile tilapia (120). However, thymol, in combination with other prophylactic agents, was found to promote the growth performance of fish. For instance, a dietary combination of thymol and carvacrol can help to promote the growth performance of European sturgeons (121) and gilthead seabreams (122). In addition, a dietary combination of chitosan nanoparticles and thymol was found to promote the growth and health status of Nile tilapia (88).
Several studies have shown the potential use of vaccines in aquaculture against edwardsiellosis (Table 3). for instance, Castro et al. (123) and Lan et al. (124) reported that an effective edwardsiellosis vaccine has been developed for turbot, Sauertylenchus maxinus. inactivated E. tarda vaccine also has been shown to stimulate immune response effectively in zebrafish (125), flounder (46, 126), turbot (127), tilapia, and Oreochromis mossambicus (128), and japanese flounder, P. olivaceus (129). the studies have used different immunogens, such as whole cell, live cells extract, outer membrane protein (130), and attenuated E. tarda, to stimulate the immune response in fish against edwardsiellosis. although the application of vaccines can control edwardsiellosis effectively, issues, such as cost, huge labor requirements, and species-specific usage, have limited its use. besides, the application of live vaccines has legal objections in many countries. meanwhile, selective breeding has been proposed as a method to improve the genetics of aquaculture species to counter the disease resistance issue against E. tarda (13, 131, 132).
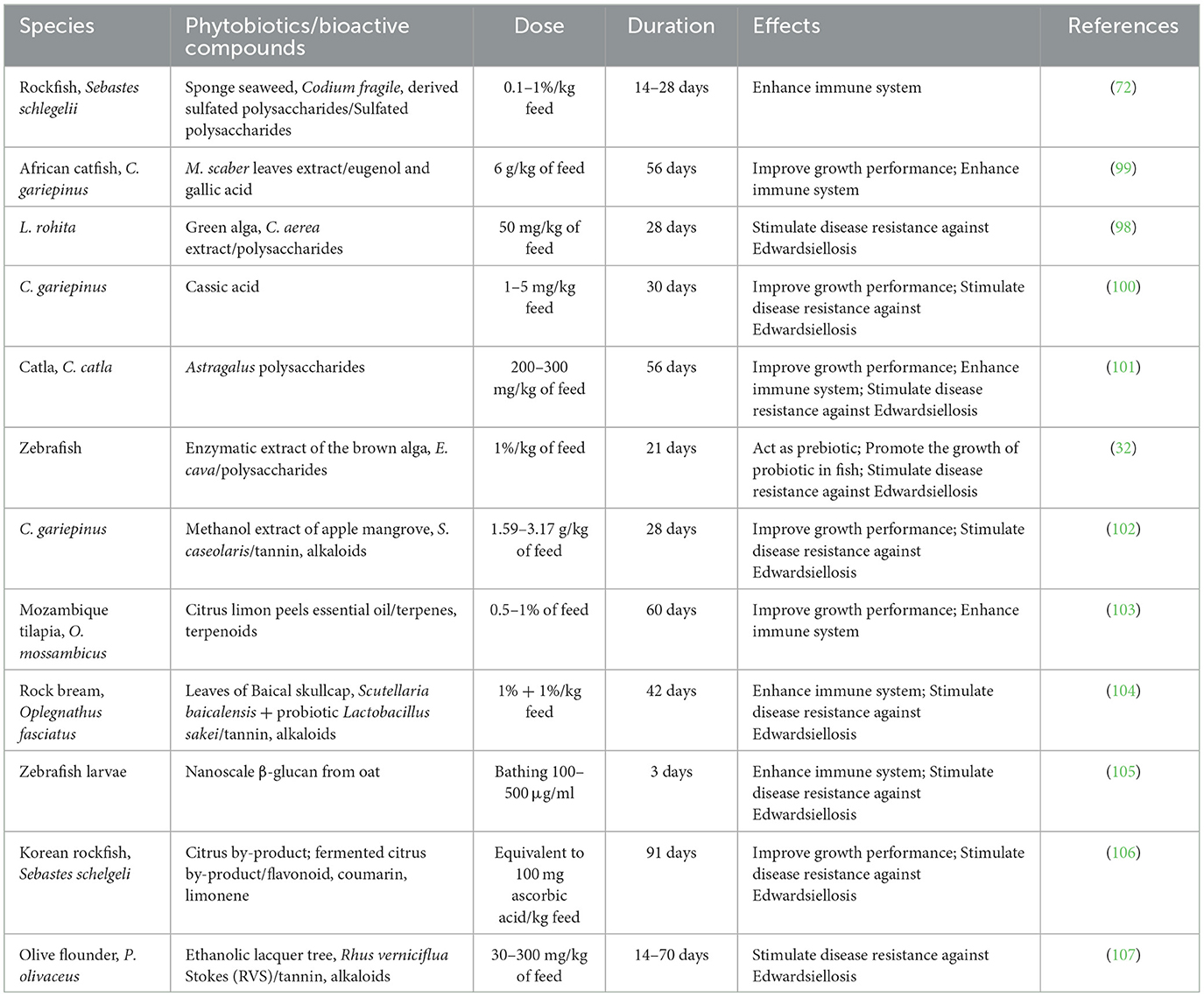
Table 3. Phytobiotics used to mitigate E. tarda impacts on aquatic animals.
Phytobiotics have been shown to stimulate disease resistance in various aquaculture species. For example, a recent study by Ahmadifar et al. (68) has claimed that Cornelian cherry (Cornus mas L.) fruit extract can stimulate disease resistance in common carp, C. carpio, against A. hydrophila. Meanwhile, studies found that phytobiotics can stimulate disease resistance in aquatic animals (69–71). Sulfated polysaccharides from sponge seaweed (72) and Astragalus (101) were found to enhance the disease resistance of both freshwater and marine aquaculture species against Edwardsiellosis (Table 2). Sulfated polysaccharides are referred to as anionic polysaccharides that carry sulfates (133). These sulfated polysaccharides have medicinal benefits, such as antibacterial, antiviral, anti-inflammatory, and rich antioxidant properties (134). Astragalus polysaccharides were also found to improve growth performance, enhance the immune system, and stimulate disease resistance to Edwardsiellosis in catla (Catla catla). Astragalus polysaccharides are bioactive compounds that are reported to possess rich antioxidant properties (135) and can play important roles in activating the immune system (101). Mitracarpus scaber leaf extract was reported to stimulate disease resistance of African catfish against Edwardsiellosis (99). This plant leaf extract possesses bioactive compounds, such as eugenol and gallic acid. These bioactive compounds are able to modulate gut microbiota, enhance feed utilization, and promote growth performance (99). Green alga, Chaetomorpha aerea extract may carry bioactive compounds that can play a role as an activator for genes related to antimicrobial molecules production (98). Therefore, this green alga extract was found to stimulate disease resistance in Labeo rohita against Edwardsiellosis (98). Cassic acid is a bioactive compound widely and commercially used in Chinese herb medicinal (100). This compound can be found in the root and leaf of plant species, such as Senna, Rheum, and Cassia (100). Cassic acid has medicinal values like antibacterial, antifungal, and antiviral properties and is rich in antioxidant properties (69, 136). These medicinal values were responsible for the growth performance improvement and disease resistance to Edwardsiellosis in African catfish, C. gariepinus that received cassic acid as a feed additive (69). Brown alga, E. cava was found to be used as a prebiotic in promoting the growth of probiotic, LAB (32). Further study on E. cava revealed that brown alga can be used as a feed additive in zebrafish. It can improve the growth of zebrafish and stimulate disease resistance to Edwardsiellosis (105). A recent report showed that apple mangrove Sonneratia caseolaris extract could be used feed additive in African catfish, C. gariepinus. The bioactive compounds in the plant extract can enhance the appetite of the fish, improve growth performance, and stimulate disease resistance to Edwardsiellosis (102). Besides that, agricultural waste, a citrus by-product, was claimed to help in improving the health status of aquaculture species against Edwardsiellosis (103). For example, essential oil from Citrus limon carries bioactive compounds, such as flavonoid, coumarin, and limonene, that are responsible for the antibacterial, antioxidant, and anticancer properties of the essential oil (103). Bioactive compounds in the phytobiotics can play an important role in activating the innate immune system in aquatic animals (104). One of the innate immune systems is serum lysozyme. Serum lysozyme catalyzes the pathogen cell wall and phagocytosis activities against pathogens, such as viruses, parasites, and bacteria, that invade the host (104). All phytobiotics administered as a feed additive have been shown to improve the health status of aquatic animals against Edwardsiellosis except for nanoscale β-glucan (NSBG), which can also be used to fish larvae via bathing treatment (105). In this context, NSBG acted as an immunostimulant to enhance the innate immune system in the fish larval before the larval adaptive immune system was well developed (105). β-Glucan is a commercial feed additive that is abundant and inexpensive. Therefore, this bioactive compound is widely used in animal feed to enhance animal production. The duration of phytobiotics served as a feed additive to aquatic animal range from 14 to 91 days. The benefits of phytobiotics used as feed additives are not only to stimulate disease resistance to Edwardsiellosis but also to improve growth performance and enhance the immune system of aquatic animals. Administration of phytobiotics orally is the most practical and non-stressful method, and can be used widely in aquaculture.
The application of phytobiotics in aquaculture was widely reported in the literature. Phytobiotics have beneficial effects on various aquatic animals, such as growth performance improvement, immune system enhancement, and disease resistance enhancement. Apart from their beneficial effects, phytobiotics were reported to have adverse impacts. For example, RVS has medicinal properties, such as anticancer (137), antiviral (138), antibacterial, and antioxidant (139) activities. This phytobiotic was reported to relieve the impacts of Edwardsiellosis infection in olive flounder (P. olivaceus) (33). In addition, the methanolic extract of RVS bark was also found to be significantly effective against E. tarda and Vibrio anguillarum (107). However, RVS possesses bioactive compounds known as urushiol congeners that can cause adverse effects, such as inflammation, blistering, and irritation (140). Thus, the adverse impacts limit the use of RSV in treatment. On the other hand, urushiol congeners were found absent in the RVS lignum (139). Furthermore, RVS lignum performed the highest antibacterial and antioxidant activities against E. tarda isolated from fish (141). Hence, RVS lignum has a high potential to be used as a phytobiotic in aquaculture. Some phytobiotics have low toxicity and few side effects on aquatic animals. For instance, Astragalus polysaccharides were widely used in Chinese medicine practice (101). These bioactive compounds were also shown to be promising as feed additives in chicken (142) and fish (101).
Edwardsiellosis due to E. tarda is an important disease in aquaculture that mainly affects carp fish, eels, flounder, turbot, channel catfish, and many other aquaculture species. This disease can devastate the whole fish farm and lead to huge economic loss. Traditionally, antibiotics and vaccines were used to combat Edwardsiellosis in aquaculture. However, antibiotics have an adverse impact on microbial communities in aquaculture sites, and their residues in aquaculture products can pose a threat to public health. On the other hand, the application of vaccines is expensive and requires high labor work. Therefore, these two issues became major constraints to the usage of vaccines in combating Edwardsiellosis in aquaculture. Therefore, phytobiotics can be an alternative option to fish farmers as a prophylactic agent against Edwardsiellosis in aquaculture. At present, phytobiotics are evidenced to have a high potential in controlling Edwardsiellosis. However, further studies should be carried out to investigate the effectiveness of phytobiotics against Edwardsiellosis in more important aquaculture species, such as eels, flounder, turbot, and channel catfish. Currently, there is a lack of information in the literature on the benefit of phytobiotics to the abovementioned aquaculture species, and many potential phytobiotics are waiting to be explored to relieve the impact of Edwardsiellosis in aquaculture.
Conceptualization: KG and NZ. Writing—original draft preparation: ZA and LW. Writing—review and editing: MR, MK, NA, AK, GT-I, and AT. All authors have read and agreed to the published version of the manuscript.
The project was mainly funded by Ministry of Higher Education, Malaysia, under the Fundamental Research Grant Scheme (FRGS) (FRGS/1/2022/STG03/UMK/03/1). Research was supported in part by funds provided by USDA-NIFA Sustainable Agriculture Systems, Grant No. 2019-69012-29905. Title of Project: Empowering US Broiler Production for Transformation and Sustainability USDA-NIFA (Sustainable Agriculture Systems): No. 2019-69012-29905 and Universiti Sultan Zainal Abidin (UniSZA/2021/PKP/02). The review article is from the collaboration between Universiti Malaysia Kelantan, Sylhet Agricultural University, INTI International University, Mindanao State University-Tawi-Tawi College of Technology and Oceanography, Universiti Sultan Zainal Abidin (UniSZA), University of Arkansas and Universiti Malaysia Terengganu. These collaborations are a part of the planning by the Advanced Livestock and Aquaculture Research Group - ALAReG under the Faculty of Agro-Based Industry, Universiti Malaysia Kelantan, Jeli Campus.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Muringai RT, Mafongoya P, Lottering RT, Mugandani R, Naidoo D. Unlocking the potential of fish to improve food and nutrition security in Sub-Saharan Africa. Sustainability. (2022) 14:318. doi: 10.3390/su14010318
2. Zakaria MK, Kari ZA, Van Doan H, Kabir MA, Che Harun H, Mohamad Sukri SA, et al. Fermented soybean meal (FSBM) in African Catfish (Clarias gariepinus) diets: effects on growth performance, fish gut microbiota analysis, blood haematology, and liver morphology. Life. (2022) 12:1851. doi: 10.3390/life12111851
3. Heikkinen J, Vielma J, Kemiläinen O, Tiirola M, Eskelinen P, Kiuru T, et al. Effects of soybean meal based diet on growth performance, gut histopathology and intestinal microbiota of juvenile rainbow trout (Oncorhynchus mykiss). Aquaculture. (2006) 261:259–68. doi: 10.1016/j.aquaculture.2006.07.012
4. Adeleke B, Robertson-Andersson D, Moodley G, Taylor S. Aquaculture in Africa: a comparative review of Egypt, Nigeria, and Uganda Vis-À-Vis South Africa. Rev Fisheries Sci Aquacult. (2021) 29:167–97. doi: 10.1080/23308249.2020.1795615
5. Cain K. The many challenges of disease management in aquaculture. J World Aquac Soc. (2022) 53:1080–3. doi: 10.1111/jwas.12936
6. Zhao Y, Yang QE, Zhou X, Wang F-H, Muurinen J, Virta MP, et al. Antibiotic resistome in the livestock and aquaculture industries: status and solutions. Crit Rev Environ Sci Technol. (2021) 51:2159–96. doi: 10.1080/10643389.2020.1777815
7. Buján N, Mohammed H, Balboa S, Romalde JL, Toranzo AE, Arias CR, et al. Genetic studies to re-affiliate Edwardsiella tarda fish isolates to Edwardsiella piscicida and Edwardsiella anguillarum species. Syst Appl Microbiol. (2018) 41:30–7. doi: 10.1016/j.syapm.2017.09.004
8. Mohanty BR, Sahoo PK. Edwardsiellosis in fish: a brief review. J Biosci. (2007) 32:1331–44. doi: 10.1007/s12038-007-0143-8
9. Park SB, Aoki T, Jung TS. Pathogenesis of and strategies for preventing Edwardsiella tarda infection in fish. Vet Res. (2012) 43:67. doi: 10.1186/1297-9716-43-67
10. Pirarat N, Kobayashi T, Katagiri T, Maita M, Endo M. Protective effects and mechanisms of a probiotic bacterium Lactobacillus rhamnosus against experimental Edwardsiella tarda infection in tilapia (Oreochromis niloticus). Vet Immunol Immunopathol. (2006) 113:339–47. doi: 10.1016/j.vetimm.2006.06.003
11. Srinivasa Rao PS, Yamada Y, Leung KY. A major catalase (KatB) that is required for resistance to H2O2 and phagocyte-mediated killing in Edwardsiella tarda. Microbiology (Reading). (2003) 149:2635–44. doi: 10.1099/mic.0.26478-0
12. Zheng W-j, Hu Y-h, Sun L. The two Dps of Edwardsiella tarda are involved in resistance against oxidative stress and host infection. Fish Shellfish Immunol. (2011) 31:985–92. doi: 10.1016/j.fsi.2011.08.018
13. Li M-f, Jia B-b, Sun Y-y, Sun L. The translocation and assembly module (TAM) of Edwardsiella tarda is essential for stress resistance and host infection. Front Microbiol. (2020) 11:1743. doi: 10.3389/fmicb.2020.01743
14. Yu M, Xu Y, Xu T, Wang B, Sheng A, Zhang X-H. WcaJ, the initiating enzyme for colanic acid synthesis, is required for lipopolysaccharide production, biofilm formation and virulence in Edwardsiella tarda. Aquaculture. (2015) 437:287–91. doi: 10.1016/j.aquaculture.2014.12.011
15. Cheng S, Zhang M, Sun L. The iron-cofactored superoxide dismutase of Edwardsiella tarda inhibits macrophage-mediated innate immune response. Fish Shellfish Immunol. (2010) 29:972–8. doi: 10.1016/j.fsi.2010.08.004
16. Algammal AM, Mabrok M, Ezzat M, Alfifi KJ, Esawy AM, Elmasry N, et al. Prevalence, antimicrobial resistance (AMR) pattern, virulence determinant and AMR genes of emerging multi-drug resistant Edwardsiella tarda in Nile tilapia and African catfish. Aquaculture. (2022) 548:737643. doi: 10.1016/j.aquaculture.2021.737643
17. Rodríguez Verjan C, Augusto V, Xie X, Buthion V. Economic comparison between Hospital at Home and traditional hospitalization using a simulation-based approach. J Enterp Inf Manag. (2013) 26:135–53. doi: 10.1108/17410391311289596
18. Guo M, Zhang L, Ye J, He X, Cao P, Zhou Z, et al. Characterization of the pathogenesis and immune response to a highly virulent Edwardsiella tarda strain responsible for mass mortality in the hybrid snakehead (Channa maculate ♀ × Channa argus ♂). Microb Pathog. (2022) 170:105689. doi: 10.1016/j.micpath.2022.105689
19. Pandey V, Hussain Bhat RA, Chandra S, Tandel RS, Dubey MK, Sharma P, et al. Clinical signs, lethal dose and histopathological lesions in grass carp, Ctenopharyngodon idella experimentally infected with Edwardsiella tarda. Microb Pathog. (2021) 161:105292. doi: 10.1016/j.micpath.2021.105292
20. Leung KY, Siame BA, Tenkink BJ, Noort RJ, Mok Y-K. Edwardsiella tarda – Virulence mechanisms of an emerging gastroenteritis pathogen. Microbes and Infection. (2012) 14:26–34. doi: 10.1016/j.micinf.2011.08.005
21. Sun K, Wang H-l, Zhang M, Xiao Z-z, Sun L. Genetic mechanisms of multi-antimicrobial resistance in a pathogenic Edwardsiella tarda strain. Aquaculture. (2009) 289:134–9. doi: 10.1016/j.aquaculture.2008.12.021
22. Venter H, Henningsen ML, Begg SL. Antimicrobial resistance in healthcare, agriculture and the environment: the biochemistry behind the headlines. Essays Biochem. (2017) 61:1–10. doi: 10.1042/EBC20160053
23. Abdul Kari Z, Kabir MA, Mat K, Rusli ND, Razab MKAA, Ariff NSNA, et al. The possibility of replacing fish meal with fermented soy pulp on the growth performance, blood biochemistry, liver, and intestinal morphology of African catfish (Clarias gariepinus). Aquacult Rep. (2021) 21:100815. doi: 10.1016/j.aqrep.2021.100815
24. Kari ZA, Kabir MA, Dawood MAO, Razab MKAA, Ariff NSNA, Sarkar T, et al. Effect of fish meal substitution with fermented soy pulp on growth performance, digestive enzyme, amino acid profile, and immune-related gene expression of African catfish (Clarias gariepinus). Aquaculture. (2022) 546:737418. doi: 10.1016/j.aquaculture.2021.737418
25. Rehman MNU, Wang Y, Pan J, Han Y, Yang N, Wang X, et al. Histological and molecular characterization of Edwardsiella tarda infection in Siamese crocodile (Crocodylus siamensis) hatchlings. Aquaculture. (2021) 535:736367. doi: 10.1016/j.aquaculture.2021.736367
26. Fu S, Tu J, Rahman MM, Tian H, Xiao P, Liu Y. Precise feeding of probiotics in the treatment of edwardsiellosis by accurate estimation of Edwardsiella tarda. Ann Microbiol. (2018) 68:645–54. doi: 10.1007/s13213-018-1371-x
27. Nikapitiya C, Chandrarathna HPSU, Dananjaya SHS, De Zoysa M, Lee J. Isolation and characterization of phage (ETP-1) specific to multidrug resistant pathogenic Edwardsiella tarda and its in vivo biocontrol efficacy in zebrafish (Danio rerio). Biologicals. (2020) 63:14–23. doi: 10.1016/j.biologicals.2019.12.006
28. Estaiano de Rezende RA, Soares MP, Sampaio FG, Cardoso IL, Ishikawa MM, Lima Dallago BS, et al. Phytobiotics blend as a dietary supplement for Nile tilapia health improvement. Fish Shellfish Immunol. (2021) 114:293–300. doi: 10.1016/j.fsi.2021.05.010
29. Yu JE, Cho MY, Kim JW, Kang HY. Large antibiotic-resistance plasmid of Edwardsiella tarda contributes to virulence in fish. Microb Pathog. (2012) 52:259–66. doi: 10.1016/j.micpath.2012.01.006
30. Abolghait SK, Fathi AG, Youssef FM, Algammal AM. Methicillin-resistant Staphylococcus aureus (MRSA) isolated from chicken meat and giblets often produces staphylococcal enterotoxin B (SEB) in non-refrigerated raw chicken livers. Int J Food Microbiol. (2020) 328:108669. doi: 10.1016/j.ijfoodmicro.2020.108669
31. Kari ZA, Wee W, Hamid NKA, Mat K, Rusli ND, Khalid HNM, et al. Recent advances of phytobiotic utilization in carp farming: a review. Aquacult Nutr. (2022) 2022:7626675. doi: 10.1155/2022/7626675
32. Lee W, Oh JY, Kim EA, Kang N, Kim KN, Ahn G, et al. A prebiotic role of Ecklonia cava improves the mortality of Edwardsiella tarda-infected zebrafish models via regulating the growth of lactic acid bacteria and pathogen bacteria. Fish Shellfish Immunol. (2016) 54:620–8. doi: 10.1016/j.fsi.2016.05.018
33. Wang L, Xu X, Zhang Z, Li K, Yang Y, Zheng W, et al. Transcriptome analysis and protein-protein interaction in resistant and susceptible families of Japanese flounder (Paralichthys olivaceus) to understand the mechanism against Edwardsiella tarda. Fish Shellfish Immunol. (2022) 123:265–81. doi: 10.1016/j.fsi.2022.02.055
34. Shao S, Lai Q, Liu Q, Wu H, Xiao J, Shao Z, et al. Phylogenomics characterization of a highly virulent Edwardsiella strain ET080813T encoding two distinct T3SS and three T6SS gene clusters: Propose a novel species as Edwardsiella anguillarum sp. nov. Syst Appl Microbiol. (2015) 38:36–47. doi: 10.1016/j.syapm.2014.10.008
35. Abayneh T, Colquhoun DJ, Sørum H. Edwardsiella piscicida sp. nov, a novel species pathogenic to fish. J Appl Microbiol. (2013) 114:644–54. doi: 10.1111/jam.12080
36. Evans J, Klesius P, Plumb J, Shoemaker C. Edwardsiella septicaemias: CABI International. Cambridge: CAB International (2011) p. 512–69. doi: 10.1079/9781845935542.0512
37. Kumar G, Sharma P, Rathore G, Bisht D, Sengupta U. Proteomic analysis of outer membrane proteins of Edwardsiella tarda. J Appl Microbiol. (2010) 108:2214–21. doi: 10.1111/j.1365-2672.2009.04627.x
38. Okuda J, Murayama F, Yamanoi E, Iwamoto E, Matsuoka S, Nishibuchi M, et al. Base changes in the fliC gene of Edwardsiella tarda: possible effects on flagellation and motility. Dis Aquat Organ. (2007) 76:113–21. doi: 10.3354/dao076113
39. Yamasaki M, Araki K, Nakanishi T, Nakayasu C, Yoshiura Y, Iida T, et al. Adaptive immune response to Edwardsiella tarda infection in ginbuna crucian carp, Carassius auratus langsdorfii. Vet Immunol Immunopathol. (2013) 153:83–90. doi: 10.1016/j.vetimm.2013.02.004
40. Kumar G, Rathore G, El-Matbouli M. Outer membrane protein assembly factor YaeT (omp85) and GroEL proteins of Edwardsiella tarda are immunogenic antigens for Labeo rohita (Hamilton). J Fish Dis. (2014) 37:1055–9. doi: 10.1111/jfd.12205
41. Xu T, Zhang X-H. Edwardsiella tarda: an intriguing problem in aquaculture. Aquaculture. (2014) 431:129–35. doi: 10.1016/j.aquaculture.2013.12.001
42. Mo Z-Q, Zhou L, Zhang X, Gan L, Liu L, Dan X-M. Outbreak of Edwardsiella tarda infection in farm-cultured giant mottled eel Anguilla marmorata in China. Fisheries Science. (2015) 81:899–905. doi: 10.1007/s12562-015-0906-3
43. Park SB, Nho SW, Jang HB, Cha IS, Lee J-H, Aoki T, et al. Phenotypic and genotypic analysis of Edwardsiella tarda isolated from olive founder (Paralichthys olivaceus) and Japanese eel (Anguilla japonica). Aquaculture. (2017) 473:449–55. doi: 10.1016/j.aquaculture.2017.03.015
44. Du Y, Tang X, Sheng X, Xing J, Zhan W. The influence of concentration of inactivated Edwardsiella tarda bacterin and immersion time on antigen uptake and expression of immune-related genes in Japanese flounder (Paralichthys olivaceus). Microb Pathog. (2017) 103:19–28. doi: 10.1016/j.micpath.2016.12.011
45. Li R, Qu J, Li H, Zhang Q. Genome-wide identification and analysis of scavenger receptors and their expression profiling in response to Edwardsiella tarda infection in Japanese flounder (Paralichthys olivaceus). Dev Comp Immunol. (2022) 132:104397. doi: 10.1016/j.dci.2022.104397
46. Wu X, Xing J, Tang X, Sheng X, Chi H, Zhan W. Protective cellular and humoral immune responses to Edwardsiella tarda in flounder (Paralichthys olivaceus) immunized by an inactivated vaccine. Mol Immunol. (2022) 149:77–86. doi: 10.1016/j.molimm.2022.06.008
47. Amandi A, Hiu SF, Rohovec JS, Fryer JL. Isolation and characterization of Edwardsiella tarda from fall chinook salmon (Oncorhynchus tshawytscha). Appl Environ Microbiol. (1982) 43:1380–4. doi: 10.1128/aem.43.6.1380-1384.1982
48. Katharios P, Kokkari C, Dourala N, Smyrli M. First report of Edwardsiellosis in cage-cultured sharpsnout sea bream, Diplodus puntazzo from the Mediterranean. BMC Vet Res. (2015) 11:155. doi: 10.1186/s12917-015-0482-x
49. Jang JH, Kim H, Cho JH. Molecular cloning and functional characterization of peptidoglycan recognition protein OmPGRP-L2 from the rainbow trout, Oncorhynchus mykiss. Vet Immunol Immunopathol. (2017) 192:28–32. doi: 10.1016/j.vetimm.2017.09.004
50. Yang R, Liu Y, Wang Y, Lei M, Pan G, Wen J, et al. Pathogenesis and pathological analysis of Edwardsiella tarda from Dabry's sturgeon (Acipenser dabryanus) in China. Aquaculture. (2018) 495:637–42. doi: 10.1016/j.aquaculture.2018.04.010
51. Liu H, Xie J-f, Yu H, Ma Z, Yu Y-Y, Yang Y. The early response expression profiles of miRNA-mRNA in farmed yellow catfish (Pelteobagrus fulvidraco) challenged with Edwardsiella tarda infection. Dev Comp Immunol. (2021) 119:104018. doi: 10.1016/j.dci.2021.104018
52. Cao M, Yan X, Li Q, Fu Q, Yang N, Song L, et al. Genome-wide identification and analysis of NOD-like receptors and their potential roles in response to Edwardsiella tarda infection in black rockfish (Sebastes schlegelii). Aquaculture. (2021) 541:736803. doi: 10.1016/j.aquaculture.2021.736803
53. Li M, Yang Y, Zheng W, Li Z, Cheng J, Li Y. Estimation of heritabilities of disease resistance to Edwardsiella tarda and genetic correlations between resistance and growth traits in Chinese tongue sole (Cynoglossus semilaevis). Aquaculture and Fisheries. (2020) 5:289–93. doi: 10.1016/j.aaf.2019.12.002
54. Wang X, Wang F, Chen G, Yang B, Chen J, Fang Y, et al. Edwardsiella tarda induces enteritis in farmed seahorses (Hippocampus erectus): An experimental model and its evaluation. Fish Shellfish Immunol. (2020) 98:391–400. doi: 10.1016/j.fsi.2020.01.049
55. Liang Q, Zhu N, Zheng X, Ding X, He R, Xu H, et al. Transcriptome analysis of immune responses and metabolic regulations of chinese soft-shelled turtle (Pelodiscus sinensis) against Edwardsiella tarda infection. Fishes. (2022) 7:79. doi: 10.3390/fishes7020079
56. Xu L, Che J, Xu Y, Chen Y, Li Y, Murtaza B, et al. Oral administration of microencapsulated egg yolk immunoglobulin (IgY) in turbot (Scophthalmus maximus) to combat against Edwardsiella tarda 2CDM001 infections. Fish Shellfish Immunol. (2020) 106:609–20. doi: 10.1016/j.fsi.2020.08.024
57. Buenviaje G, Ladds P, Melville L, Manolis S. Disease-husbandry associations in farmed crocodiles in Queensland and the Northern Territory. Aust Vet J. (1994) 71:165–73. doi: 10.1111/j.1751-0813.1994.tb03381.x
58. Guo G, Jiang J, Yang N, Wang P, Zhang L, Wang Y, et al. An investigation of sudden death in farmed infant Siamese crocodiles during winter and spring in Hainan, China. Indian J Anim Res. (2018) 52:1058–62.
59. Loch TP, Hawke JP, Reichley SR, Faisal M, Del Piero F, Griffin MJ. Outbreaks of edwardsiellosis caused by Edwardsiella piscicida and Edwardsiella tarda in farmed barramundi (Lates calcarifer). Aquaculture. (2017) 481:202–10. doi: 10.1016/j.aquaculture.2017.09.005
60. Elgendy MY, Sherif AH, Kenawy AM, Abdelsalam M. Phenotypic and molecular characterization of the causative agents of edwardsiellosis causing Nile tilapia (Oreochromis niloticus) summer mortalities. Microb Pathog. (2022) 169:105620. doi: 10.1016/j.micpath.2022.105620
61. Verma RK, Sankhla MS, Jadhav S, Parihar K, Gulliya S, Kumar R, et al. Chapter 8 - Global status of bacterial fish diseases in relation to aquatic pollution. In:Dar GH, Bhat RA, Qadri H, Al-Ghamdy KM, Hakeem KR, , editors. Bacterial Fish Diseases. Cambridge, MA: Academic Press. (2022) p. 155–82.
62. Van Hai N. The use of medicinal plants as immunostimulants in aquaculture: a review. Aquaculture. (2015) 446:88–96. doi: 10.1016/j.aquaculture.2015.03.014
63. Neamat-Allah ANF, El-Murr AeI, Abd El-Hakim Y. Dietary supplementation with low molecular weight sodium alginate improves growth, haematology, immune reactions and resistance against Aeromonas hydrophila in Clarias gariepinus. Aquacul Res. (2019) 50:1547–56. doi: 10.1111/are.14031
64. Newaj-Fyzul A, Austin B. Probiotics, immunostimulants, plant products and oral vaccines, and their role as feed supplements in the control of bacterial fish diseases. J Fish Dis. (2015) 38:937–55. doi: 10.1111/jfd.12313
65. Aljumaah MR, Alkhulaifi MM, Aljumaah RS, Abudabos AM, Abdullatif AA, Suliman GM, et al. Influence of sanguinarine-based phytobiotic supplementation on post necrotic enteritis challenge recovery. Heliyon. (2020) 6:e05361. doi: 10.1016/j.heliyon.2020.e05361
66. Brenes A, Roura E. Essential oils in poultry nutrition: main effects and modes of action. Anim Feed Sci Technol. (2010) 158:1–14. doi: 10.1016/j.anifeedsci.2010.03.007
67. Huyghebaert G, Ducatelle R, Immerseel FV. An update on alternatives to antimicrobial growth promoters for broilers. The Veterinary Journal. (2011) 187:182–8. doi: 10.1016/j.tvjl.2010.03.003
68. Ahmadifar E, Mohammadzadeh S, Kalhor N, Yousefi M, Moghadam MS, Naraballobh W, et al. Cornelian cherry (Cornus mas L.) fruit extract improves growth performance, disease resistance, and serum immune-and antioxidant-related gene expression of common carp (Cyprinus carpio). Aquaculture. (2022) 558:738372. doi: 10.1016/j.aquaculture.2022.738372
69. Abd El-Naby AS, El Asely AM, Hussein MN, Fawzy RM, Abdel-Tawwab M. Stimulatory effects of dietary chia (Salvia hispanica) seeds on performance, antioxidant-immune indices, histopathological architecture, and disease resistance of Nile tilapia. Aquaculture. (2023) 563:738889. doi: 10.1016/j.aquaculture.2022.738889
70. Abdel-Latif HMR, Dawood MAO, Alagawany M, Faggio C, Nowosad J, Kucharczyk D. Health benefits and potential applications of fucoidan (FCD) extracted from brown seaweeds in aquaculture: An updated review. Fish Shellfish Immunol. (2022) 122:115–30. doi: 10.1016/j.fsi.2022.01.039
71. Poolsawat L, Yu Y, Li X, Zhen X, Yao W, Wang P, et al. Efficacy of phytogenic extracts on growth performance and health of tilapia (Oreochromis niloticus × O. aureus). Aquacult Fisher. (2022) 7:411–9. doi: 10.1016/j.aaf.2020.08.009
72. Yang Y, Lim J, Li C, Lee S, Hong S. Effects of sulfated polysaccharides isolated from Codium fragile on inflammatory cytokine gene expression and Edwardsiella tarda infection in rockfish, Sebastes schlegelii. Fish Shellfish Immunol. (2021) 112:125–34. doi: 10.1016/j.fsi.2021.03.001
73. Uma A, Philominal P, Prabu E, Musthafa MS. Dietary Bougainvillea glabra leaf meal on growth, haemato-biochemical responses and disease resistance in Nile tilapia, Oreochromis niloticus against Enterococcus faecalis. Aquaculture. (2022) 549:737806. doi: 10.1016/j.aquaculture.2021.737806
74. Abdel-Latif HMR, Shukry M, Noreldin AE, Ahmed HA, El-Bahrawy A, Ghetas HA, et al. Milk thistle (Silybum marianum) extract improves growth, immunity, serum biochemical indices, antioxidant state, hepatic histoarchitecture, and intestinal histomorphometry of striped catfish, Pangasianodon hypophthalmus. Aquaculture. (2023) 562:738761. doi: 10.1016/j.aquaculture.2022.738761
75. Ghafarifarsani H, Hoseinifar SH, Aftabgard M, Van Doan H. The improving role of savory (Satureja hortensis) essential oil for Caspian roach (Rutilus caspicus) fry: Growth, haematological, immunological, and antioxidant parameters and resistance to salinity stress. Aquaculture. (2022) 548:737653. doi: 10.1016/j.aquaculture.2021.737653
76. Abdel-Tawwab M, Adeshina I, Jenyo-Oni A, Ajani EK, Emikpe BO. Growth, physiological, antioxidants, and immune response of African catfish, Clarias gariepinus (B.), to dietary clove basil, Ocimum gratissimum, leaf extract and its susceptibility to Listeria monocytogenes infection. Fish Shellfish Immunol. (2018) 78:346–54. doi: 10.1016/j.fsi.2018.04.057
77. Abdo SE, El-Nahas AF, Abdelmenam S, Elmadawy MA, Mohamed R, Helal MA, et al. The synergetic effect of Bacillus species and Yucca shidigera extract on water quality, histopathology, antioxidant, and innate immunity in response to acute ammonia exposure in Nile tilapia. Fish Shellfish Immunol. (2022) 128:123–35. doi: 10.1016/j.fsi.2022.07.058
78. Ashour M, Mabrouk MM, Abo-Taleb HA, Sharawy ZZ, Ayoub HF, Van Doan H, et al. A liquid seaweed extract (TAM®) improves aqueous rearing environment, diversity of zooplankton community, whilst enhancing growth and immune response of Nile tilapia, Oreochromis niloticus, challenged by Aeromonas hydrophila. Aquaculture. (2021) 543:736915. doi: 10.1016/j.aquaculture.2021.736915
79. Soares MP, Cardoso IL, Araújo FE, De Angelis CF, Mendes R, Mendes LW, et al. Influences of the alcoholic extract of Artemisia annua on gastrointestinal microbiota and performance of Nile tilapia. Aquaculture. (2022) 560:738521. doi: 10.1016/j.aquaculture.2022.738521
80. Abdel-Latif HMR, Abdel-Tawwab M, Khafaga AF, Dawood MAO. Dietary oregano essential oil improved the growth performance via enhancing the intestinal morphometry and hepato-renal functions of common carp (Cyprinus carpio L.) fingerlings. Aquacult. (2020) 526:735432. doi: 10.1016/j.aquaculture.2020.735432
81. Ebrahimi E, Haghjou M, Nematollahi A, Goudarzian F. Effects of rosemary essential oil on growth performance and hematological parameters of young great sturgeon (Huso huso). Aquaculture. (2020) 521:734909. doi: 10.1016/j.aquaculture.2019.734909
82. Sutili FJ, Velasquez A, Pinheiro CG, Heinzmann BM, Gatlin DM. 3rd, Baldisserotto B. Evaluation of Ocimum americanum essential oil as an additive in red drum (Sciaenops ocellatus) diets. Fish Shellfish Immunol. (2016) 56:155–61. doi: 10.1016/j.fsi.2016.07.008
83. Fath El-Bab AF, Majrashi KA, Sheikh HM, Shafi ME, El-Ratel IT, Neamat-Allah ANF, et al. Dietary supplementation of Nile tilapia (Oreochromis niloticus) with β-glucan and/or Bacillus coagulans: synergistic impacts on performance, immune responses, redox status and expression of some related genes. Front Vet Sci. (2022) 9:1011715. doi: 10.3389/fvets.2022.1011715
84. Naiel MAE, Khames MK, Abdel-Razek N, Gharib AA, El-Tarabily KA. The dietary administration of miswak leaf powder promotes performance, antioxidant, immune activity, and resistance against infectious diseases on Nile tilapia (Oreochromis niloticus). Aquaculture Rep. (2021) 20:100707. doi: 10.1016/j.aqrep.2021.100707
85. Negm SS, Ismael NEM, Ahmed AI, Asely AME, Naiel MAE. The efficiency of dietary Sargassum aquifolium on the performance, innate immune responses, antioxidant activity, and intestinal microbiota of Nile Tilapia (Oreochromis niloticus) raised at high stocking density. J Appl Phycol. (2021) 33:4067–82. doi: 10.1007/s10811-021-02572-3
86. Abdelghany MF, El-Sawy HB. Abd El-hameed SAA, Khames MK, Abdel-Latif HMR, Naiel MAE. Effects of dietary Nannochloropsis oculata on growth performance, serum biochemical parameters, immune responses, and resistance against Aeromonas veronii challenge in Nile tilapia (Oreochromis niloticus). Fish & Shellfish Immunology. (2020) 107:277–88. doi: 10.1016/j.fsi.2020.10.015
87. Khafaga AF, Naiel MAE, Dawood MAO, Abdel-Latif HMR. Dietary Origanum vulgare essential oil attenuates cypermethrin-induced biochemical changes, oxidative stress, histopathological alterations, apoptosis, and reduces DNA damage in Common carp (Cyprinus carpio). Aquatic Toxicol. (2020) 228:105624. doi: 10.1016/j.aquatox.2020.105624
88. Abd El-Naby AS, Al-Sagheer AA, Negm SS, Naiel MAE. Dietary combination of chitosan nanoparticle and thymol affects feed utilization, digestive enzymes, antioxidant status, and intestinal morphology of Oreochromis niloticus. Aquaculture. (2020) 515:734577. doi: 10.1016/j.aquaculture.2019.734577
89. Naiel MAE, Ismael NEM, Shehata SA. Ameliorative effect of diets supplemented with rosemary (R. officinalis) on aflatoxin B1 toxicity in terms of the performance, liver histopathology, immunity and antioxidant activity of Nile Tilapia (Oreochromis niloticus). Aquaculture. (2019) 511:734264. doi: 10.1016/j.aquaculture.2019.734264
90. Bai N, Gu M, Zhang W, Xu W, Mai K. Effects of β-glucan derivatives on the immunity of white shrimp Litopenaeus vannamei and its resistance against white spot syndrome virus infection. Aquaculture. (2014) 426–427:66–73. doi: 10.1016/j.aquaculture.2014.01.019
91. Ji L, Sun G, Li J, Wang Y, Du Y, Li X, et al. Effect of dietary β-glucan on growth, survival and regulation of immune processes in rainbow trout (Oncorhynchus mykiss) infected by Aeromonas salmonicida. Fish Shellfish Immunol. (2017) 64:56–67. doi: 10.1016/j.fsi.2017.03.015
92. Reyes-Becerril M, Guardiola FA, Sanchez V, Maldonado M, Angulo C. Sterigmatomyces halophilus β-glucan improves the immune response and bacterial resistance in Pacific red snapper (Lutjanus peru) peripheral blood leucocytes: In vitro study. Fish Shellfish Immunol. (2018) 78:392–403. doi: 10.1016/j.fsi.2018.04.043
93. Pionnier N, Falco A, Miest J, Frost P, Irnazarow I, Shrive A, et al. Dietary β-glucan stimulate complement and C-reactive protein acute phase responses in common carp (Cyprinus carpio) during an Aeromonas salmonicida infection. Fish Shellfish Immunol. (2013) 34:819–31. doi: 10.1016/j.fsi.2012.12.017
94. Souza FPd, Lima ECSd, Pandolfi VCF, Leite NG, Furlan-Murari PJ, Leal CNS, et al. Effect of β-glucan in water on growth performance, blood status and intestinal microbiota in tilapia under hypoxia. Aquaculture Rep. (2020) 17:100369. doi: 10.1016/j.aqrep.2020.100369
95. Halawany HS, A. review on miswak (Salvadora persica) and its effect on various aspects of oral health. Saudi Dental J. (2012) 24:63–9. doi: 10.1016/j.sdentj.2011.12.004
96. Syawal H, Syafriadiman S, Hidayah S. The use of miswak (Salvadora persica L.) extract to increase immune response of common carp (Cyprinus carpio L) in cage Biodiversitas. J Biol Divers. (2008) 9:44–7. doi: 10.13057/biodiv/d090110
97. Lebda MA, El-Hawarry WN, Shourbela RM, El-Far AH, Shewita RS, Mousa SA. Miswak (Salvadora persica) dietary supplementation improves antioxidant status and nonspecific immunity in Nile tilapia (Oreochromis niloticus). Fish Shellfish Immunol. (2019) 88:619–26. doi: 10.1016/j.fsi.2019.03.037
98. Sattanathan G, Tamizhazhagan V, Raza N, Shah SQA, Hussain MZ, Kim K-H. Effects of green alga, Chaetomorpha aerea extract on non-specific immune responses and disease resistance against Edwardsiella tarda infection in Labeo rohita. Applied Sci. (2021) 11:4325. doi: 10.3390/app11104325
99. Adeshina I, Tiamiyu LO, Abubakar MI, Ogundayomi OO, Adesanmi O. Effects of dietary Mitracarpus scaber leaves extract on growth, physiological, antioxidants, and mucosal immune profiles of North African catfish, Clarias gariepinus, and resistance against Edwardsiella tarda infection. Trop Anim Health Prod. (2021) 53:541. doi: 10.1007/s11250-021-02989-5
100. Harikrishnan R, Devi G, Paray BA, Al-Sadoon MK, Al-Mfarij AR, Van Doan H. Effect of cassic acid on immunity and immune-reproductive genes transcription in Clarias gariepinus against Edwardsiella tarda. Fish Shellfish Immunol. (2020) 99:331–41. doi: 10.1016/j.fsi.2020.02.037
101. Harikrishnan R, Devi G, Doan HV, Tapingkae W, Balasundaram C, Arockiaraj J, et al. Changes in immune genes expression, immune response, digestive enzymes -antioxidant status, and growth of catla (Catla catla) fed with Astragalus polysaccharides against edwardsiellosis disease. Fish Shellfish Immunol. (2022) 121:418–36. doi: 10.1016/j.fsi.2022.01.022
102. Aznan AS, Lee KL, Low CF, Iberahim NA, Wan Ibrahim WN, Musa N, et al. Protective effect of apple mangrove Sonneratia caseolaris extract in Edwardsiella tarda-infected African catfish, Clarias gariepinus. Fish Shellfish Immunol. (2018) 78:338–45. doi: 10.1016/j.fsi.2018.04.033
103. Baba E, Acar Ü, Öntaş C, Kesbiç OS, Yilmaz S. Evaluation of Citrus limon peels essential oil on growth performance, immune response of Mozambique tilapia Oreochromis mossambicus challenged with Edwardsiella tarda. Aquaculture. (2016) 465:13–8. doi: 10.1016/j.aquaculture.2016.08.023
104. Harikrishnan R, Kim MC, Kim JS, Balasundaram C, Heo MS. Protective effect of herbal and probiotics enriched diet on haematological and immunity status of Oplegnathus fasciatus (Temminck & Schlegel) against Edwardsiella tarda. Fish Shellfish Immunol. (2011) 30:886–93. doi: 10.1016/j.fsi.2011.01.013
105. Udayangani RMC, Dananjaya SHS, Fronte B, Kim C-H, Lee J, De Zoysa M. Feeding of nano scale oats β-glucan enhances the host resistance against Edwardsiella tarda and protective immune modulation in zebrafish larvae. Fish Shellfish Immunol. (2017) 60:72–7. doi: 10.1016/j.fsi.2016.11.035
106. Song JW, Lee K-J. Evaluation of citrus by-product on growth performance, innate immunity and disease resistance against Edwardsiella tarda of Korean rockfish Sebastes schelgeli. Fish Shellfish Immunol. (2013) 34:1738. doi: 10.1016/j.fsi.2013.03.314
107. Lim J-W, Shin S-M, Jung S-J, Lee MK, Kang SY. Optimization of antibacterial extract from lacquer tree (Rhus verniciflua Stokes) using response surface methodology and its efficacy in controlling edwardsiellosis of olive flounder (Paralichthys olivaceus). Aquaculture. (2019) 502:40–7. doi: 10.1016/j.aquaculture.2018.12.008
108. Borowitzka MA. Microalgae for aquaculture: opportunities and constraints. J Appl Phycol. (1997) 9:393–401. doi: 10.1023/A:1007921728300
109. Sarker PK, Kapuscinski AR, Bae AY, Donaldson E, Sitek AJ, Fitzgerald DS, et al. Towards sustainable aquafeeds: Evaluating substitution of fishmeal with lipid-extracted microalgal co-product (Nannochloropsis oculata) in diets of juvenile Nile tilapia (Oreochromis niloticus). PLoS ONE. (2018) 13:e0201315. doi: 10.1371/journal.pone.0201315
110. Norambuena F, Hermon K, Skrzypczyk V, Emery JA, Sharon Y, Beard A, et al. Algae in fish feed: performances and fatty acid metabolism in juvenile Atlantic salmon. PLoS ONE. (2015) 10:e0124042. doi: 10.1371/journal.pone.0124042
111. Caillet S, Yu H, Lessard S, Lamoureux G, Ajdukovic D, Lacroix M. Fenton reaction applied for screening natural antioxidants. Food Chem. (2007) 100:542–52. doi: 10.1016/j.foodchem.2005.10.009
112. El-Barbary MI, Mehrim AI. Protective effect of antioxidant medicinal herbs, rosemary and parsley, on subacute aflatoxicosis in Oreochromis niloticus. J Fish Aquat Sci. (2009) 4:178–90. doi: 10.3923/jfas.2009.178.190
113. Mansoori B, Acamovic T. Influence of tannic acid and polyethylene glycol on the excretion and digestibility of amino acids in gelatin-fed broilers. Br Poult Sci. (2009) 50:199–206. doi: 10.1080/00071660902736714
114. Alagawany M, Abd El-Hack ME, Farag MR, Shaheen HM, Abdel-Latif MA, Noreldin AE, et al. The applications of and its derivatives in human, ruminant and fish nutrition – a review. Annals Animal Sci. (2020) 20:389–407. doi: 10.2478/aoas-2020-0004
115. Botsoglou NA, Taitzoglou IA, Botsoglou E, Lavrentiadou SN, Kokoli AN, Roubies N. Effect of long-term dietary administration of oregano on the alleviation of carbon tetrachloride-induced oxidative stress in rats. J Agric Food Chem. (2008) 56:6287–93. doi: 10.1021/jf8003652
116. Sharifi-Rigi A, Heidarian E. Therapeutic potential of Origanum vulgare leaf hydroethanolic extract against renal oxidative stress and nephrotoxicity induced by paraquat in rats. Avicenna J Phytomed. (2019) 9:563–73.
117. Sharifi-Rigi A, Heidarian E, Amini SA. Protective and anti-inflammatory effects of hydroalcoholic leaf extract of Origanum vulgare on oxidative stress, TNF-α gene expression and liver histological changes in paraquat-induced hepatotoxicity in rats. Arch Physiol Biochem. (2019) 125:56–63. doi: 10.1080/13813455.2018.1437186
118. Giannenas I, Triantafillou E, Stavrakakis S, Margaroni M, Mavridis S, Steiner T, et al. Assessment of dietary supplementation with carvacrol or thymol containing feed additives on performance, intestinal microbiota and antioxidant status of rainbow trout (Oncorhynchus mykiss). Aquaculture. (2012) 350–353:26–32. doi: 10.1016/j.aquaculture.2012.04.027
119. Zheng ZL, Tan JYW, Liu HY, Zhou XH, Xiang X, Wang KY. Evaluation of oregano essential oil (Origanum heracleoticum L.) on growth, antioxidant effect and resistance against Aeromonas hydrophila in channel catfish (Ictalurus punctatus). Aquaculture. (2009) 292:214–8. doi: 10.1016/j.aquaculture.2009.04.025
120. Aanyu M, Betancor MB, Monroig O. Effects of dietary limonene and thymol on the growth and nutritional physiology of Nile tilapia (Oreochromis niloticus). Aquaculture. (2018) 488:217–26. doi: 10.1016/j.aquaculture.2018.01.036
121. Ahmadifar E, Razeghi Mansour M, Keramat Amirkolaie A, Fadaii Rayeni M. Growth efficiency, survival and haematological changes in great sturgeon (Huso huso Linnaeus, 1758) juveniles fed diets supplemented with different levels of thymol–carvacrol. Anim Feed Sci Technol. (2014) 198:304–8. doi: 10.1016/j.anifeedsci.2014.08.012
122. Pérez-Sánchez J, Benedito-Palos L, Estensoro I, Petropoulos Y, Calduch-Giner JA, Browdy CL, et al. Effects of dietary NEXT ENHANCE®150 on growth performance and expression of immune and intestinal integrity related genes in gilthead sea bream (Sparus aurata L. ). Fish Shellfish Immunology. (2015) 44:117–28. doi: 10.1016/j.fsi.2015.01.039
123. Castro N, Toranzo AE, Núñez S, Magariños B. Development of an effective Edwardsiella tarda vaccine for cultured turbot (Scophthalmus maximus). Fish Shellfish Immunol. (2008) 25:208–12. doi: 10.1016/j.fsi.2008.05.008
124. Lan MZ, Peng X, Xiang MY, Xia ZY, Bo W, Jie L, et al. Construction and characterization of a live, attenuated esrB mutant of Edwardsiella tarda and its potential as a vaccine against the haemorrhagic septicaemia in turbot, Scophthamus maximus (L.). Fish Shellfish Immunol. (2007) 23:521–30. doi: 10.1016/j.fsi.2006.11.002
125. Xu D, Wang J, Guo C, Peng XX Li H. Elevated biosynthesis of palmitic acid is required for zebrafish against Edwardsiella tarda infection. Fish Shellfish Immunol. (2019) 92:508–18. doi: 10.1016/j.fsi.2019.06.041
126. Tang X, Qin Y, Sheng X, Xing J, Zhan W. Characterization of CD3(+) T lymphocytes of Japanese flounder (Paralichthys olivaceus) and its response after immunization with formalin-inactivated Edwardsiella tarda. Fish Shellfish Immunol. (2017) 63:220–7. doi: 10.1016/j.fsi.2017.02.024
127. Liu X, Zhang H, Jiao C, Liu Q, Zhang Y, Xiao J. Flagellin enhances the immunoprotection of formalin-inactivated Edwardsiella tarda vaccine in turbot. Vaccine. (2017) 35:369–74. doi: 10.1016/j.vaccine.2016.11.031
128. Kwon SR, Nam YK, Kim SK, Kim KH. Protection of tilapia (Oreochromis mosambicus) from edwardsiellosis by vaccination with Edwardsiella tarda ghosts. Fish Shellfish Immunol. (2006) 20:621–6. doi: 10.1016/j.fsi.2005.08.005
129. Mekuchi T, Kiyokawa T, Honda K, Nakai T, Muroga K. Vaccination trials in the Japanese flounder against edwardsiellosis. Fish Pathol. (1995) 30:251–6. doi: 10.3147/jsfp.30.251
130. Jin C-d, Li M-f. Edwardsiella tarda outer membrane protein 1 is a virulence factor and a protective immunogen. Aquaculture. (2021) 539:736596. doi: 10.1016/j.aquaculture.2021.736596
131. Gjedrem T, Rye M. Selection response in fish and shellfish: a review. Rev Aquacult. (2018) 10:168–79. doi: 10.1111/raq.12154
132. Sun S, Lyu D, Hu Y, Wang W. Estimating genetic parameters of resistance against Edwardsiella tarda in turbot (Scophthalmus maximus). Aquaculture Rep. (2022) 26:101329. doi: 10.1016/j.aqrep.2022.101329
133. Necas J, Bartosikova L. Carrageenan: a review. Vet Med. (2013) 58:187–205. doi: 10.17221/6758-VETMED
134. Ngo D-H, Kim S-K. Sulfated polysaccharides as bioactive agents from marine algae. Int J Biol Macromol. (2013) 62:70–5. doi: 10.1016/j.ijbiomac.2013.08.036
135. Shahzad M, Small DM, Morais C, Wojcikowski K, Shabbir A, Gobe GC. Protection against oxidative stress-induced apoptosis in kidney epithelium by Angelica and Astragalus. J Ethnopharmacol. (2016) 179:412–9. doi: 10.1016/j.jep.2015.12.027
136. Antonisamy P, Agastian P, Kang C-W, Kim NS, Kim J-H. Anti-inflammatory activity of rhein isolated from the flowers of Cassia fistula L. and possible underlying mechanisms. Saudi J Biological Sci. (2019) 26:96–104. doi: 10.1016/j.sjbs.2017.04.011
137. Choi W, Jung H, Kim K, Lee S, Yoon S, Park J, et al. Rhus verniciflua Stokes against Advanced Cancer: A Perspective from the Korean Integrative Cancer Center. J Biomed Biotechnol. (2012) 2012:874276. doi: 10.1155/2012/874276
138. Kang SY, Kang J-Y, Oh M-J. Antiviral activities of flavonoids isolated from the bark of Rhus verniciflua stokes against fish pathogenic viruses in vitro. J Microbiol. (2012) 50:293–300. doi: 10.1007/s12275-012-2068-7
139. Kim J-S, Kwon Y-S, Chun W-J, Kim T-Y, Sun J, Yu C-Y, et al. Rhus verniciflua Stokes flavonoid extracts have anti-oxidant, anti-microbial and α-glucosidase inhibitory effect. Food Chem. (2010) 120:539–43. doi: 10.1016/j.foodchem.2009.10.051
140. Kim JE, Lee SY, Lee JS, Park YL, Whang KU. Clinical features of systemic contact dermatitis due to the ingestion of lacquer in the province of chungcheongnam-do. Ann Dermatol. (2012) 24:319–23. doi: 10.5021/ad.2012.24.3.319
141. Jang JY, Shin H, Lim J-W, Ahn JH, Jo YH, Lee KY, et al. Comparison of antibacterial activity and phenolic constituents of bark, lignum, leaves and fruit of Rhus verniciflua. PLoS ONE. (2018) 13:e0200257. doi: 10.1371/journal.pone.0200257
Keywords: phytobiotics, Edwardsiella tarda, medicinal herbs, disease tolerance, antibacterial, sustainable aquaculture
Citation: Goh KW, Abdul Kari Z, Wee W, Zakaria NNA, Rahman MM, Kabir MA, Abdul Hamid NK, Tahiluddin AB, Kamarudin AS, Téllez–Isaías G and Wei LS (2023) Exploring the roles of phytobiotics in relieving the impacts of Edwardsiella tarda infection on fish: a mini-review. Front. Vet. Sci. 10:1149514. doi: 10.3389/fvets.2023.1149514
Received: 22 January 2023; Accepted: 15 June 2023;
Published: 05 July 2023.
Edited by:
Xiuzhen Sheng, Ocean University of China, ChinaReviewed by:
Mohammed A. E. Naiel, Zagazig University, EgyptCopyright © 2023 Goh, Abdul Kari, Wee, Zakaria, Rahman, Kabir, Abdul Hamid, Tahiluddin, Kamarudin, Téllez–Isaías and Wei. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Zulhisyam Abdul Kari, enVsaGlzeWFtLmFAdW1rLmVkdS5teQ==; Lee Seong Wei, bGVlc2VvbmdAdW1rLmVkdS5teQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.