 Giovanni Buonaiuto1,2
Giovanni Buonaiuto1,2 Nicolas Lopez-Villalobos2
Nicolas Lopez-Villalobos2 Angela Costa1
Angela Costa1 Giovanni Niero3
Giovanni Niero3 Lorenzo Degano4
Lorenzo Degano4 Ludovica Maria Eugenia Mammi1*
Ludovica Maria Eugenia Mammi1* Damiano Cavallini1
Damiano Cavallini1 Alberto Palmonari1
Alberto Palmonari1 Andrea Formigoni1
Andrea Formigoni1 Giulio Visentin1
Giulio Visentin1- 1Department of Veterinary Medical Science, Alma Mater Studiorum – University of Bologna, Bologna, Italy
- 2School of Agriculture and Environment, Massey University, Palmerston North, New Zealand
- 3Department of Agronomy, Food, Natural Resources, Animals and Environment, University of Padova, Padova, Italy
- 4National Association of Italian Simmental Cattle Breeders (ANAPRI), Udine, Italy
The present study aimed to investigate the association between stayability (STAY) traits, muscularity, and body condition score (BCS) in the Italian Simmental dual-purpose cows. Data were collected from 2,656 cows linearly scored in their first lactation from 2002 to 2020 and reared in 324 herds. The binary trait STAY, which is the ability of a cow to stay in the herd, was obtained for each cow-lactation available up to parity 5 (from STAY1-2 to STAY4-5). Analysis of STAY was carried out using logistic regression, considering the fixed effect of energy corrected milk, conception rate, somatic cell score, and muscularity or BCS predicted at different time points. The herd of linear classification and residual error were the random effects. Primiparous cows with a medium BCS and muscularity in early lactation presented a more favorable STAY across life compared to thinner ones (P < 0.05). In fact, cows with an intermediate BCS/muscularity were more likely to stay in the herd after the third lactation (STAY3-4), compared to those presenting a lower BCS/muscularity (P < 0.01). However, cows whose muscularity was high were generally less likely to start the third lactation compared to the others. A potential explanation for this could be the willing to market cows with good conformation for meat purpose. Simmental is in fact a dual-purpose breed known for the good carcass yield and meat quality. This study demonstrates how muscularity and BCS available early in life can be associated with the ability of Simmental cows to stay in the herd.
Introduction
Herd-testing for daily milk yield and composition is one of the major sources of information for the genetic evaluation of dairy cows and quantification of herd productivity and profitability (1). For several decades, in European and Northern American countries, dairy breeding objectives included mainly traits related to milk production (2). Although being one the key drivers of profitable dairy farming, the genetic improvement for such characteristics has led to deterioration of functional traits, such as longevity and fertility, due to the antagonistic genetic correlations existing among these traits (3–5). For these reasons, nowadays breeding objectives of dairy cattle include a plethora of economically relevant traits not necessarily directly related to milk productivity (2, 6). In dairy farming, high culling rates indicate poor animal welfare, suboptimal farming conditions, and inefficient use of animal resources, impairing the sustainability of the dairy sector (7). According to Allendorf and Wettemann (8), high replacement rates cause a decrease in herd productivity followed by augmented replacement costs (9). Indeed, in Holstein-Friesian cows, milk production per lactation is maximized at the third lactation (10) and cows usually finish paying back their initial rearing cost at the end of their second lactation (11). Thus, culling cows before that moment has a detrimental impact on farmers' profitability (12, 13). Moreover, low culling rates may also improve the environmental footprint of dairy farms because of the lower number of heifers required in the herd (14).
In dairy cows, productive lifetime is defined as the period during which the animal is in production in the herd (15). Instead, longevity can be described in different manner in dairy cattle: e.g., by means of age at last calving, number of lactations started or completed, number of days from first calving to culling, age at culling, and survival at various ages or parities. Longevity combines all the characteristics that are directly associated with a cow's ability to successfully stay and perform in the herd (16). For this reason, some authors (17–19) have opted for the term “stayability” (STAY). This trait can be considered somehow equivalent to longevity, but it is usually expressed in a binary trait where 1 and 0 indicate if the animal remains in the herd and produces up to a specific moment or not, respectively (20). Cow's STAY is a key component of profitability in dairy production, as long-living cows allow to achieve the same herd production with a lower replacement rate. This implies that replacement costs can be reduced and that surplus newborn calves, preferably crossbred, can enter the beef market (21).
Conformation traits, or type traits, have been used for indirect selection to improve productive life due to their correlation with survival (22). Although collection of such phenotypes is consuming in terms of time and labor, the main advantage of type traits is that they are available early in life (2), and, indeed, several authors reported correlation of some type traits with longevity in different dairy cow breeds. Jovanovac and RaguŽ (23) reported that udder and body conformation traits, as well as muscularity and size traits, could be used as predictors of STAY and longevity in Croatian Simmental dairy cows. Schneider et al. (24) reported that udder and feet and legs traits had a strong relationship with functional herd life in Quebec Holsteins. In addition, Imbayarwo-Chikosi et al. (25) reported that chest width and rump angle were strongly associated with the risk of culling in South African Holstein dairy cows. To the best of our knowledge, no studies have attempted to identify factors associated with STAY in Italian Simmental cows, whose breeding objectives include both dairy and beef attitudes. This would be important to inform farmers in optimizing management and culling decisions based on some conformation traits, recorded within the national recording scheme.
Therefore, the aim of the present study was to retrospectively explore the variability of STAY in Italian Simmental cows and quantify its association with muscularity and BCS.
Materials and methods
Database
The present study was conducted using data retrieved from the National Association of Italian Simmental Cattle Breeders (ANAPRI, Udine, Italy) database that were collected between January 2002 and December 2020. Data was recorded on 2656 Italian Simmental dual-purpose cows reared in 324 dairy herds located in Emilia Romagna region, in North-eastern Italy. This region has a large number of dairy farms [3,519; BDN-Anagrafe Zootecnica Nazionale, (26)] that greatly contribute to the regional economy [60% of the regional gross saleable production; ISTAT, (27)]. The majority of the farms involved in the present research were multi-breed, i.e., the 97% of farms reared both Simmental and Holstein cows. Out of these, in 15 farms the number of Simmental cows was equal or above 50; only 2 of them had more than 130 heads. Only cows which were linearly classified in their first lactation were considered in the present study.
Data provided by ANAPRI included information regarding the cows' lactations estimated by the Italian Breeders Association (AIA, Rome, Italy), namely whole lactation milk and solids yield, and test-day milk records with the daily milk yield, gross composition, and somatic cell count (SCC, cells/mL). Linear type traits scores, measured once in life (in primiparous cows) by trained personnel were also present.
Phenotypes
Stayability
This trait was calculated in the lactation set, and it was defined for each cow-lactation up to the fifth, based on the presence or absence of the subsequent calving date (Table 1). Briefly, a STAY equal to 1 was assigned if a calving date was present after the previous lactation, otherwise STAY was considered equal to 0. The value was recursively set at 0 for all parities after the one incurring the culling date. This resulted in five different variables for each cow: STAY1-2, STAY2-3, STAY3-4, and STAY4-5.

Table 1. Definition and descriptive statistics of the stayability traits.
Milk traits
The energy corrected milk (ECM) was obtained from the actual lactation data according to the formula proposed by Dairy Records Management Systems (28):
where MY, FY, and PY indicate the kg of milk, fat, and protein produced within the lactation.
Milk SCC was converted to somatic cell score (SCS) according to the formula proposed by Ali and Shook (29) to achieve normal distribution:
Test-day SCS values were then averaged within each lactation, in order to be merged with STAY phenotypes and be used as an indicator of the cow's udder health.
Morphological characteristics
Linear-type traits can be scored on any days in milk (DIM) in primiparous cows. However, the morphological traits considered in the present study (muscularity and BCS) are known to vary within lactation, suggesting that observed differences among cows can be due also to the moment in which they were scored, i.e., stage of lactation. For this reason, following Buonaiuto et al. (30), individual muscularity and BCS lactation curves were adjusted through random regression analysis, allowing daily individual prediction of both traits to be present. In such a way, the differences in the expected muscularity and BCS among cows at the same DIM becomes independent from the number of days post-calving at linear type scoring. Subsequently, average lactation profiles were estimated for cows belonging to different classes of age at first calving (30), in order to evaluate the absolute growth rate (AGR) of both muscularity and BCS. The AGR was calculated according to the formula used by Handcock et al. (31):
where BTx and BTy are the predicted muscularity or BCS at xth and yth DIM, tx is the initial age of the cow (in days), and ty is the final age (in days) (30).
For further statistical analysis, only muscularity and BCS data predicted at four specific moments during the lactation were considered (30):
i. at the onset of lactation (5 DIM for both traits; Figure 1);
ii. at the nadir of muscle and/or fat reserves losses (first null AGR), i.e., the moment where the uptake from body reserves stops in Simmental (85 and 45 DIM for muscularity and BCS, respectively);
iii. at the maximum AGR, i.e., when the greatest recovery of muscle/fat reserves is observed in Simmental (180 and 160 DIM for muscularity and BCS, respectively);
iv. at the second null AGR, representing the moment from which cows start to lose again muscle/fat reserves (280 DIM in both traits);
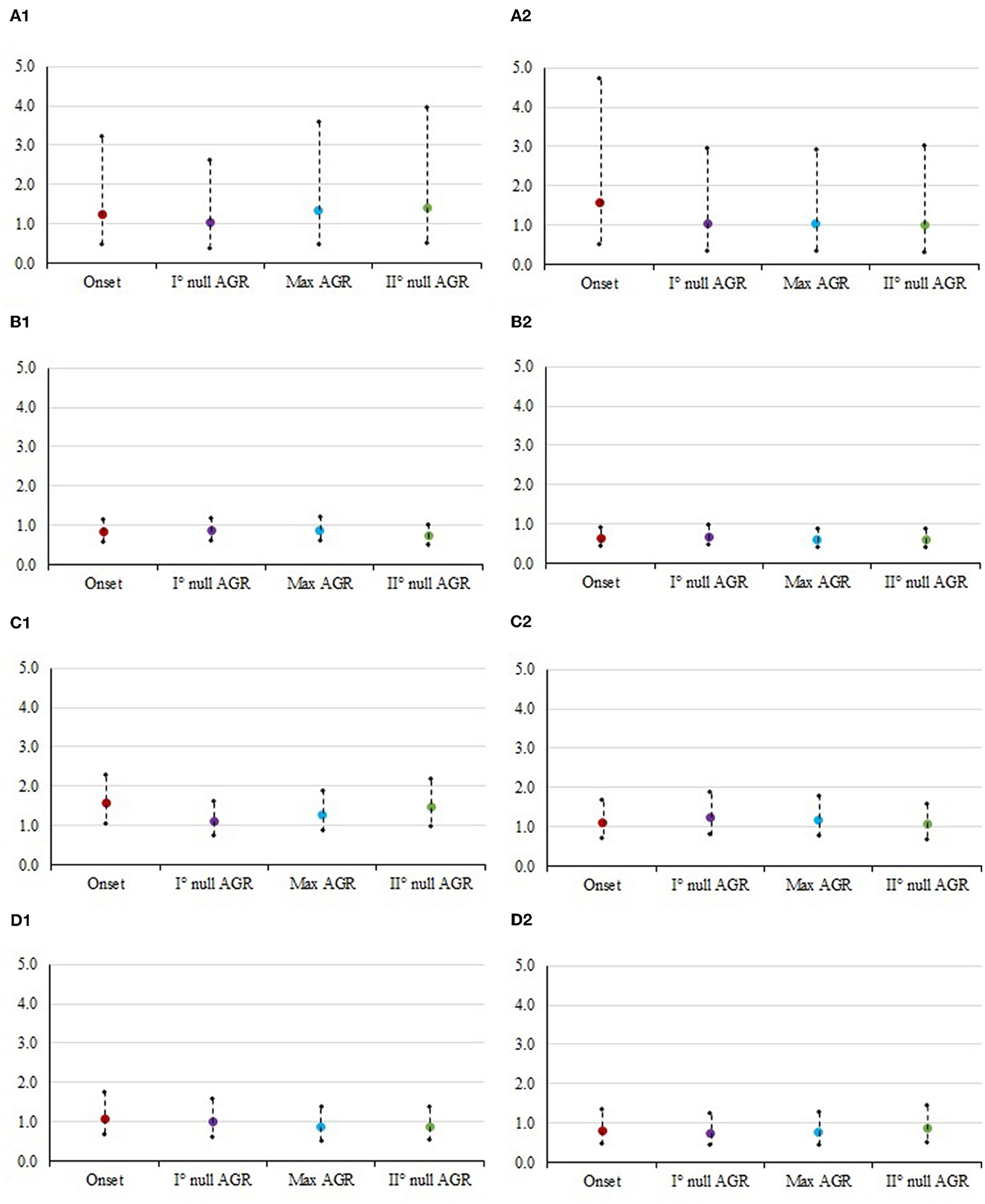
Figure 1. Odds ratios and 95% confidence interval for the risk of culling at each class of muscularity in different timepoints. The panels contain: (1) Mid vs. Low. (A1) STAY1-2, stayability as a primiparous cow, (B1) STAY2-3, stayability as a second-parity cow, (C1) STAY3-4, stayability as a third-parity cow, (D1) STAY4-5, stayability as a fourth-parity cow. (2) High vs. Low. (A2) STAY1-2, stayability as a primiparous cow, (B2) STAY2-3, stayability as a second-parity cow, (C2) STAY3-4, stayability as a third-parity cow, (D2) STAY4-5, stayability as a fourth-parity cow. For muscularity, timepoints were selected according to the absolute growth rates (AGR) trends reported by Buonaiuto et al. (30): onset of lactation = 5 DIM; I° null AGR = 85 DIM; Max AGR = 180 DIM; II° null AGR = 280 DIM.
Subsequently, cow-specific predictions of muscularity and BCS were merged to the lactation data.
Statistical analysis
Muscularity, BCS, ECM, and SCS (lactation average) were grouped into 5 classes based on quintile distribution for each individual variable, as: low, medium-low, medium, medium-high, and high. Records belonging to different lactations were analyzed separately, meaning that the effect of parity was not accounted for in the statistical models. Muscularity and BCS predicted at each given time point during the first lactation were included. Therefore, STAY1-2, STAY2-3, STAY3-4, and STAY4-5 were analyzed 4 times, by considering at each run classes of muscularity and BCS predicted at one out of the four different time points considered.
A logistic regression model was fitted with the GLIMMIX procedure using SAS software, version 9.4 (SAS Institute INC., Cary, NC):
where y is STAY1-2, STAY2-3, STAY3-4, or STAY4-5; μ is the overall intercept of the model; MUl is the fixed effect of the lth class (n = 5) of muscularity predicted at each specific aforementioned time point; BCSm is the fixed effect of the mth class (n = 5; defined according to quintiles) of BCS predicted at each specific aforementioned time point; ECMn is the fixed effect of the nth class (n = 5) of ECM; CRn is the fixed effect of the nth class (0 vs. 1) of conception rate at first insemination, where 1 indicates that only a single insemination is needed to achieve pregnancy and 0 otherwise; SCSo is the fixed effect of the 0th class (n = 5; defined according to quintiles) of milk SCS; Herdp is the random effect of the pth herd (n = 324) of linear classification, assumed to be distributed as where is the herd variance; and e is the random residual term, assumed to be distributed as where is the residual variance. For the fixed effects of muscularity and BCS the first class (low) was kept as the reference, and each odds ratio (OR) was considered significant when the 95% CI did not include 1.
Results and discussions
Overview of the studied population
The Italian Simmental cows included in the present research presented relatively high production levels compared to those reported by Cziszter et al. (32) for Fleckvieh (Austrian Simmental) cattle and by Erdem et al. (33) for Simmental cows reared in Turkey. The evolution of yield traits from parity 1 onwards in Italian Simmental cows are shown in Table 2. Descriptive statistics (mean ± standard deviation) indicated that the average MY in this study increased gradually from parity 1 to 3 (Table 2) and then decreased (parity 4: 6,660.04 ± 2,688.70 kg). The MY trend across parities was similar to that reported by different authors (34, 35) for different European populations of Simmental cows. Milk composition plays an important role in countries like Italy where approximately the 75% of the total national milk annually produced is used for cheese manufacturing (36–38). As a matter of fact, fat and protein content together with pH and acidity are essential factors during milk processing into finished dairy products (39).
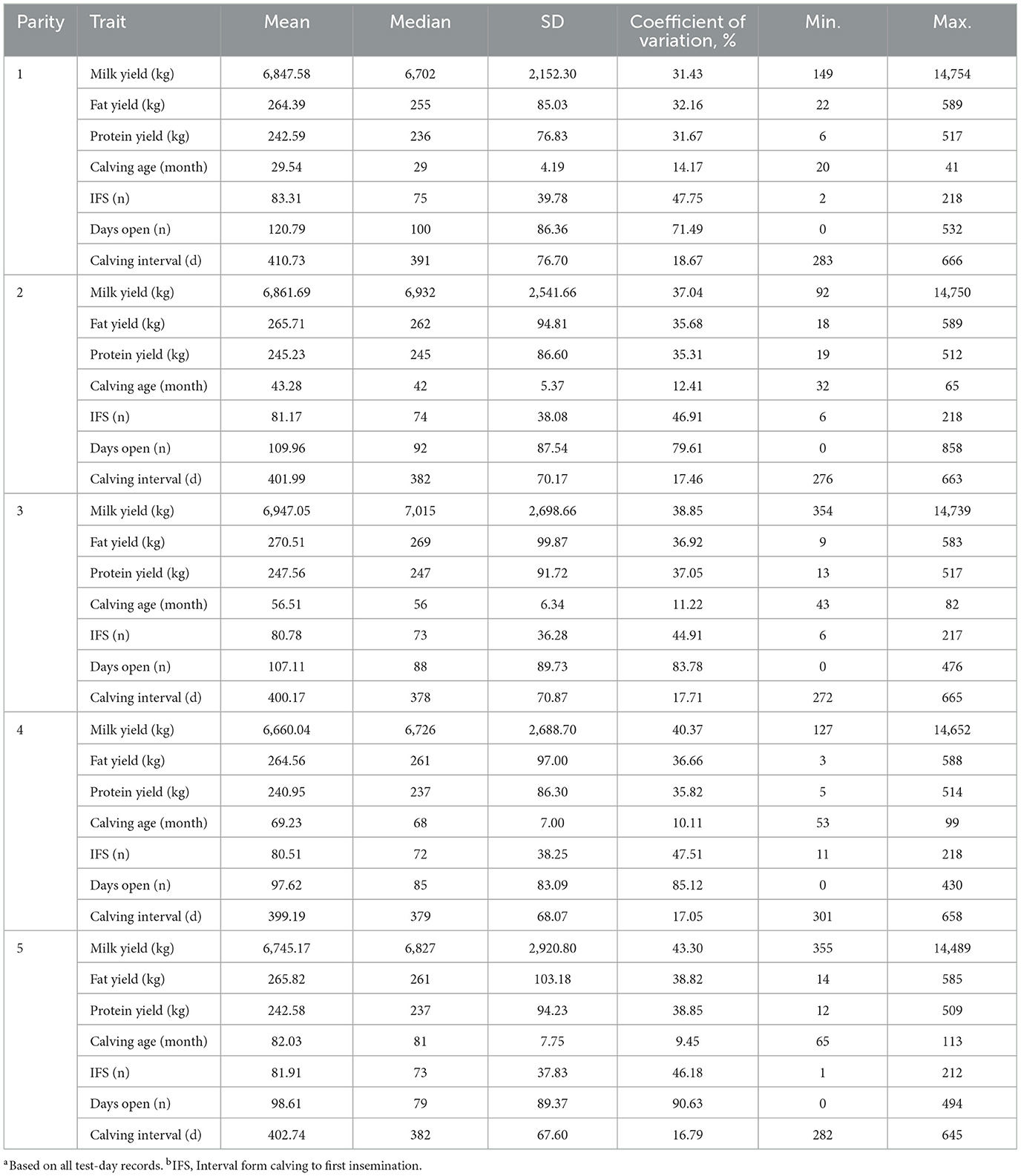
Table 2. Overview of Simmental cows' productivitya and fertilityb performance in different parities.
Along with the high productivity, the population studied was characterized by a composition comparable to that of specialized dairy breeds, with an average fat and protein content of 3.79 and 3.48%, respectively (data not shown). Parity-specific descriptive statistics of fat and protein yield, both used to calculate ECM, are reported in Table 2. Data observed are similar to that reported by other authors (40, 41) for Simmental dairy cows. By using test-day records of the first 150 DIM, Costa et al. (42) reported an average milk, fat, and protein yield of 4,157, 167.5, and 136.4 kg, respectively in Fleckvieh cows. In the same DIM window, these authors reported fat and protein content to average 4.03 and 3.28% (42). Numerous studies have investigated the effect of parity on yield traits using both test-day records or lactation data. According to the literature (43, 44) the positive correlation between MY and parity observed until a certain parity order could be attributed to the udder development/size, i.e., to the increasing number of functional secretory cells, and to the different requirements of primiparous and multiparous.
The cows' productive level and milk quality can be evaluated simultaneously by the means of ECM. The ECM can be considered as a key parameter for STAY in dairy cows as it directly affects the farm profit and, consequently, may have an effect on culling decisions (45). The culling of unproductive cows is necessary to keep the herd profitable and is thereby done on a voluntary basis. In the field, the real objective is to reduce the involuntary culling, e.g., elimination of cows—perhaps with a good MY—due to scarce fertility, severe disease, or acute inflammation (46). Well managed herds show high survival rates, thus a great proportion of mature cows and, consequently, a lower replacement rate (47).
In the present research the highest and the lowest CI value were observed in 1st (410.73 ± 76.70 d) and 4th lactation (399.19 ± 68.07 d) and, overall, the mean CI is similar to that reported in previous research in Italian Simmental cows (30). Dry period length averaged between 75.28 ± 29.22 (parity 1) to 81.29 ± 30.65 (parity 4). Across lactation the median of dry period ranged from 72 (for parity 1) to 77 (parity 2 and 4).
An overview of the investigated STAY traits and their definition is reported in Table 1. In particular, the survival rates were 98, 70, 62, and 57% (Table 1) and of the initial 2,656 cows present in parity 1 only 642 survived until parity 5 (24%; data not shown). Results are in general difficult to be compared with the literature due to scarce information available on such traits, especially for Simmental. In Holstein, Hardie et al. (19) reported that 84% (at parity 1) and 80% (at parity 2) of Holstein cows in US organic herds were able to survive and continue their productive career. Moreover, Garcia-Peniche et al. (47), report that from 38 to 43% of Holsteins stayed until 5 years of age, completing an average of 2.12 to 2.22 lactations. In the official annual report of Zuchtdata (48) the average number of calvings is equal to 3.83 for Austrian Fleckvieh while the average productive life is estimated at 3.66 years. The 24.17% of the Italian Simmental dual-purpose cows involved in this study achieved parity 5 (Table 1) which greatly differs from what has been reported for other breeds. As an example, Hare et al. (49) reported that US Holsteins dairy cows' population experienced a serious drop in survival to parity 5: from 24.2% recorded in 1980 to 14.3% in 1998. In Ireland, Williams et al. (50) reported that only 13% of the Holsteins cows involved in their study survived to the fourth lactation. Similar results are reported by Hardie et al. (19) that observe only 14% of US organic Holstein dairy cows reach parity 5. It is worth to highlight that, for the purpose of this study, data provided by ANAPRI belong exclusively to cows that were linearly classified during the first lactation, thus with a BCS and a muscularity score available. It derives that non-linear classified scored cow (e.g., for early culling in parity 1 before scoring) are not accounted for in this study. For this reason, results of this study may be interpreted in the light of absence of data from early culled cows.
According to Padilla et al. (51), a gradual age-related body deterioration is common to most animals, including dairy cows (52) and in livestock species this can affect both health and fitness of producing animals. In the past, the selection of high-producing dairy cows has favored larger more angular females, which resulted in skinny with poor carcass yield characteristics. Differently, dual-purpose cows as Simmental are characterized by a long and muscular body, that makes back and buttocks convex in most of the cases. This different body conformation was also observed by Knob et al. (53), who reported BCS of Simmental cows (and their crosses) to be approximately 1 point higher that of Holstein cows in all stages of lactation. Differences in body conformation can partly justify the survival rate observed in dairy vs. dual-purpose breeds (18). It is important to consider that, in the case of dual purpose breeds, culling could be influenced by external and economic factors, e.g., the market demand and price of milk and meat and the feed cost. Generally, when heifers of dual-purpose breeds are abundant and meat low-priced, farmers tend to cull more than usual, increasing the herd replacement rate.
Sources of variation of stayability
Results from the analysis of variance for STAY traits are summarized in Table 3. In the case of muscularity and BCS effect, the odds ratio estimates are depicted in Figures 1, 2, whereas estimates obtained for levels of SCS and CR are presented in Table 4. Overall, the effect of ECM, CR, and SCS were always significant indicating that some odds ratios differed (P < 0.001; Table 4), with the only exception of STAY1-2. Apart from severe reasons, in fact, Italian Simmental cows, whose average productive life is 3.3 lactations, are generally kept in the herd at least until second calving, i.e., regardless of the performance (26). This may partly explain why STAY1-2 was not affected by the above-mentioned fixed effects. Inclusion of ECM, CR and SCS allowed to account for that variability related to productivity level, udder health, and fertility. Odds ratio of these effects (Table 4) generally indicate that STAY is associated with SCS levels. In fact, as SCS increases, the risk of culling also increased; similar trends were observed for ECM. Regarding CR, we observed a higher risk of culling in cows with low CR (Table 4). However, although the odds ratio showed an association for all these effects, the significances was always >0.05.
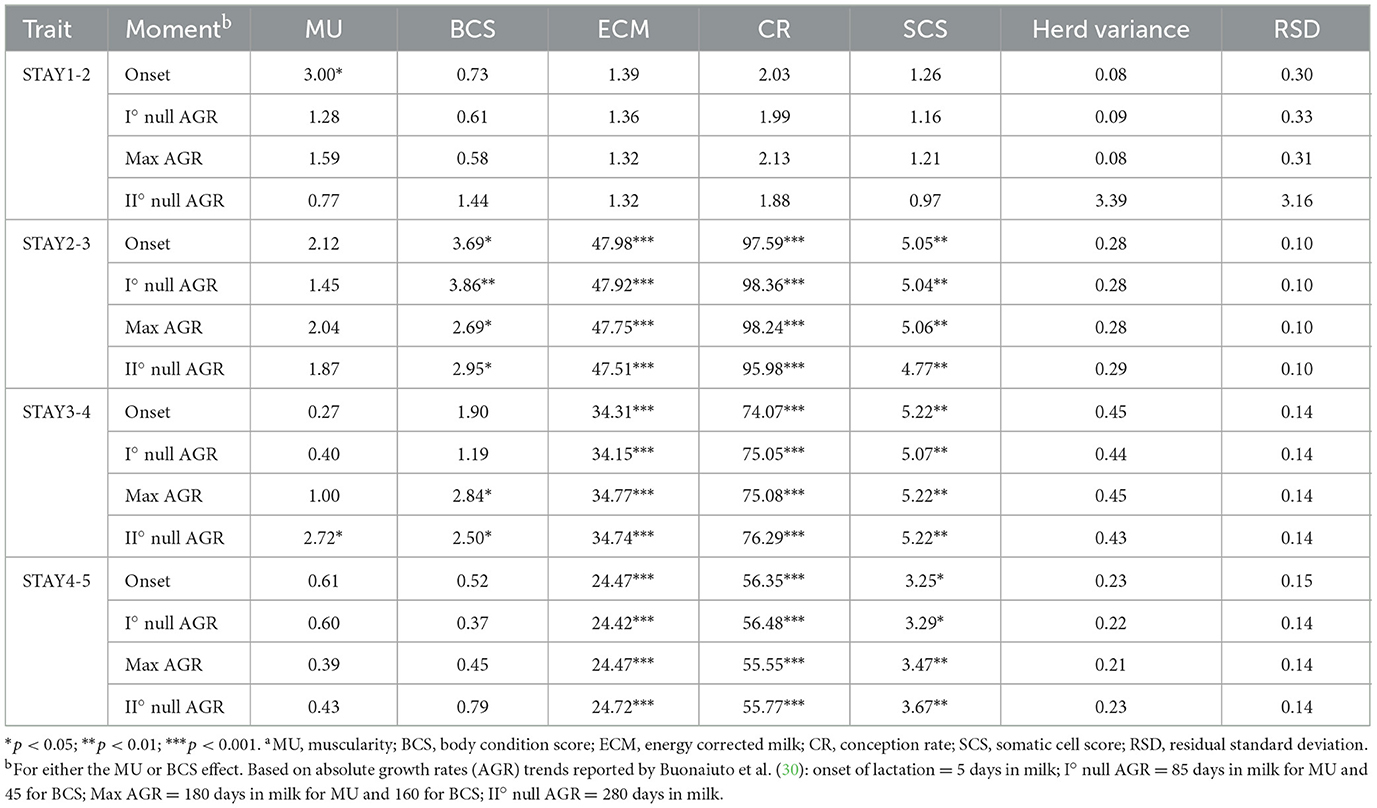
Table 3. F-values and significance of fixed effectsa included in the analysis of stayability traits.
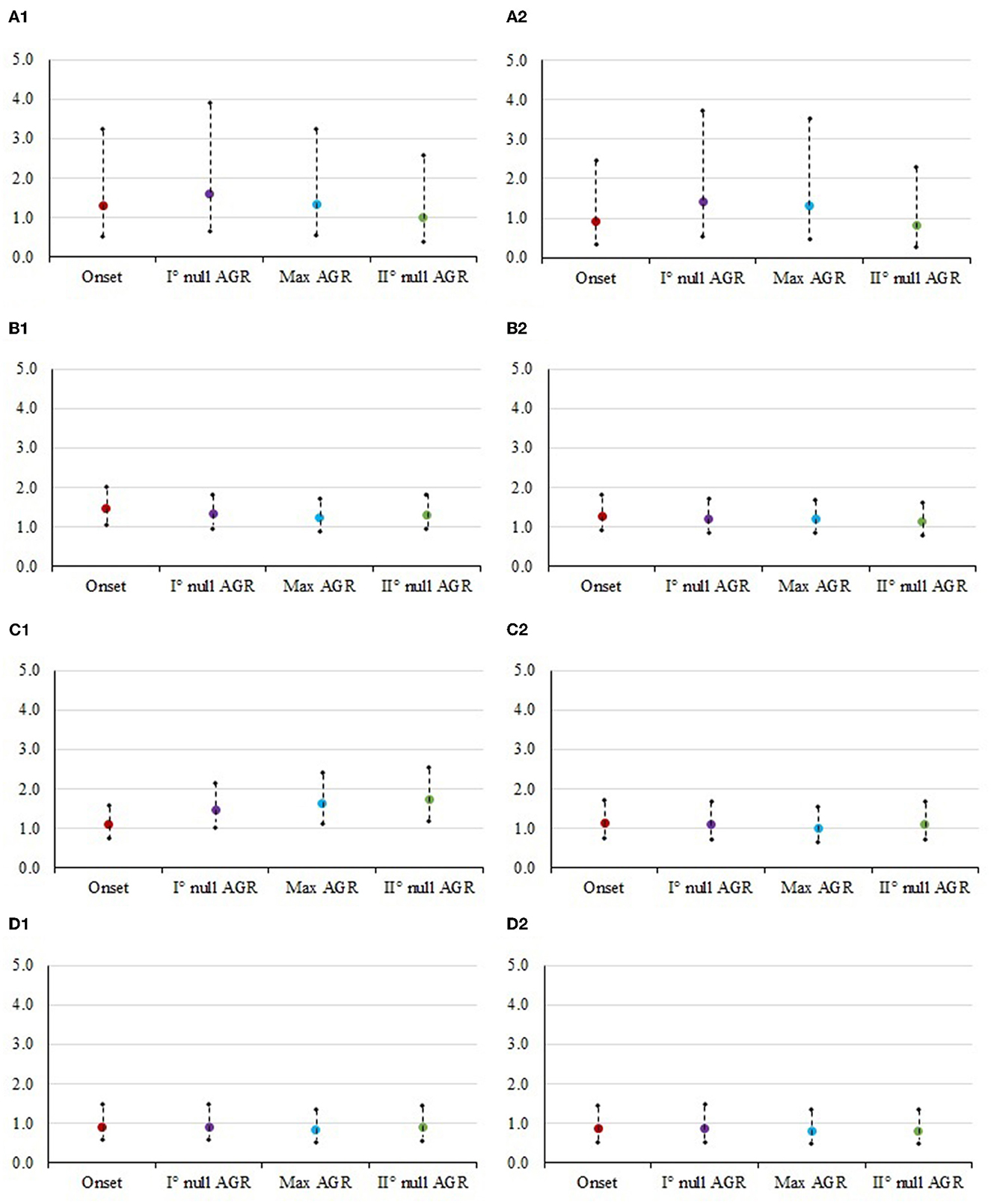
Figure 2. Odds ratios and 95% confidence interval for the risk of culling at each class of body condition score in different timepoints. The panels contain: (1) Mid vs. Low. (A1) STAY1-2, stayability as a primiparous cow, (B1) STAY2-3, stayability as a second-parity cow, (C1) STAY3-4, stayability as a third-parity cow, (D1) STAY4-5, stayability as a fourth-parity cow. (2) High vs. Low. (A2) STAY1-2, stayability as a primiparous cow, (B2) STAY2-3, stayability as a second-parity cow, (C2) STAY3-4, stayability as a third-parity cow, (D2) STAY4-5, stayability as a fourth-parity cow. For body condition score, timepoints were selected according to the absolute growth rates (AGR) trends reported by Buonaiuto et al. (30): onset of lactation = 5 DIM; I° null AGR = 45 DIM; Max AGR = 160 DIM; II° null AGR = 280 DIM.
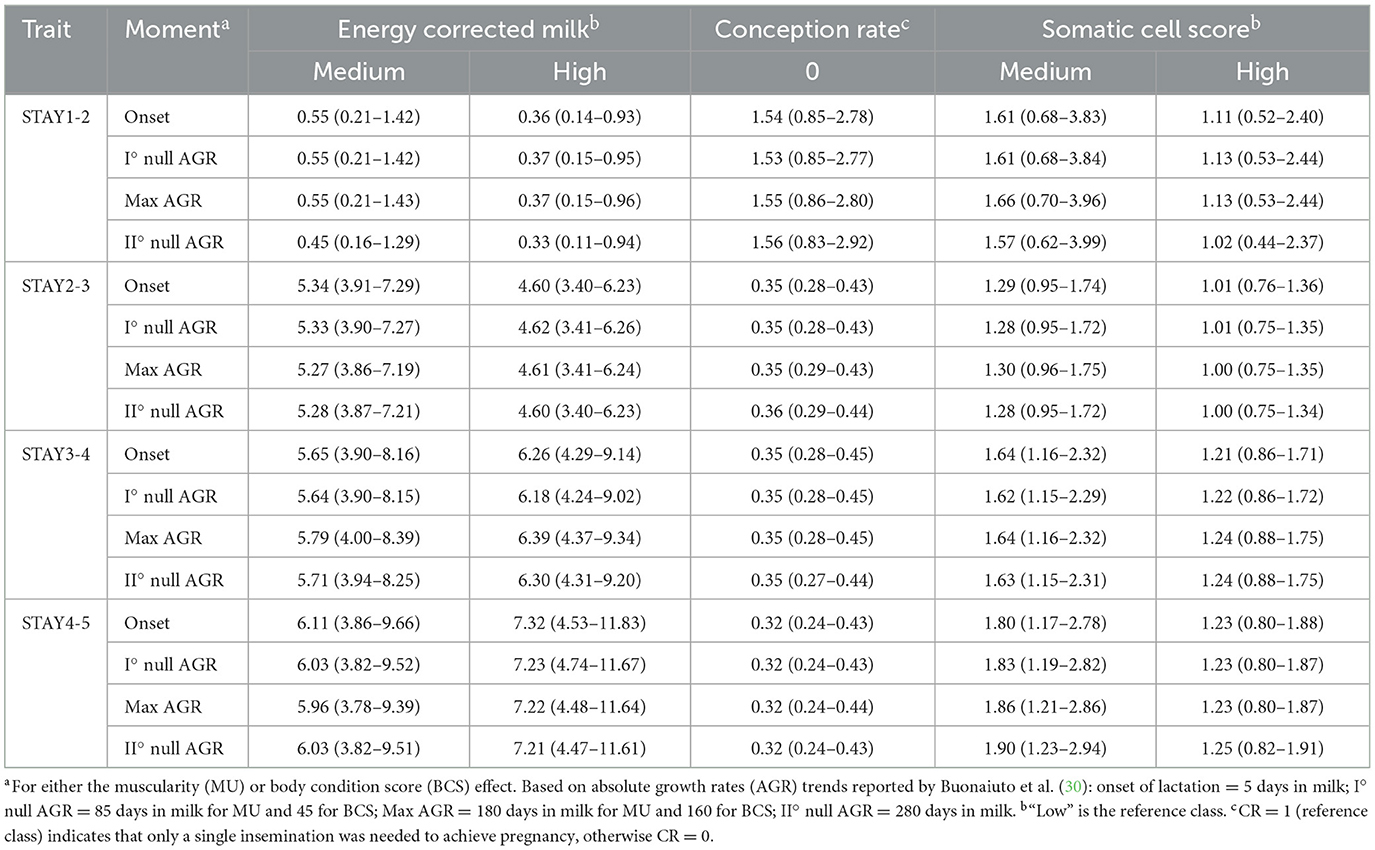
Table 4. Odds ratios (95% confidence interval) for the risk of culling at each class of energy corrected milk, somatic cell score, or conception rate estimated from the different time pointsa.
Muscularity was significant for STAY1-2 at the onset of lactation and for STAY3-4 in second null AGR and in late lactation. On the other hand, BCS was significant during all the phases considered for STAY2-3 (P < 0.05). STAY3-4 was significantly (P < 0.05) affected by cow's BCS during the max (around 160 DIM), and the second null AGR (around 280 DIM).
Figures 1, 2 show the odds ratio of the risk of culling from STAY1-2 to STAY 4-5 in cows showing different muscularity and BCS level. Although not significant in most of the cases (Table 3), the odds ratio generally indicates that animals with an average condition in terms of both muscularity and BCS are exposed to a lower risk of culling compared to cows with lower (sub-optimal) scores.
In particular, Italian Simmental dual-purpose cows with medium BCS at the beginning of lactation are more likely (1.3 times greater in the case of STAY1-2) to complete the lactation compared to those with lower condition (Figure 2A). From the moment of greatest recover of muscle and fat reserves (approximately at 180 DIM) to the moment after which animals lose again muscle and fat tissue (approximately at 280 DIM), dairy cows with medium conditions are more likely to stay in the herd, compared to cows with low condition (Figures 1, 2). An example is given by the odds ratios of STAY2-3 which are depicted in Figure 1B; in fact, the cows' muscularity had a strong impact on productive life, and therefore in the ability to stay in the herd. Indeed, at STAY2-3, cows whose muscularity was classified as high are less likely to stay in the herd compared to cows with a low muscularity, with odds ratio at DIM 5, 85, 180, and 280 being lower than unity and equal to 0.639, 0.690, 0.619, and 0.612. Conversely, at STAY2-3, cows with high BCS (Figure 2A) are more likely to stay in herd compared to cows with a low BCS, especially at 45 (odds ratio = 1.404) and 180 DIM (odds ratio = 1.310). The odds ratios (Figure 2C) show how, in parity 3, the BCS has a strong and significant impact on the cows' STAY (STAY3-4), with cows in the medium class being more likely to stay in the herd compared to those in the low class; the odds ratio at 45, 160, and 280 DIM was 1.470, 1.639, and 1.724, respectively. The same can be valid for cows with high BCS compared to those with low BCS (5 DIM: 1.129, 45 DIM: 1.102, 160 DIM: 1.005, 280 DIM: 1.100). During STAY3-4 cows with a medium muscularity condition are more likely to stay in the herd compared to cows with low muscularity, presenting an odds ratio of 1.569 at 5 DIM. At 280 DIM, the moment after which animals lose again muscle tissue, cows with medium BCS are significantly more likely (odds ratio = 1.724) to continue their career compared to those whose condition was classified as low. Similar result could be observed at STAY4-5 (Figure 1D1), in particular, cows with medium muscularity at onset of lactation (5 DIM) are more likely to stay in herd compared to cows with low conditions (odds ratio: 1.091). In general, the result depicts a fall in the probability to stay in the herd for cows with high muscularity and BCS conditions (Figures 1D2, 2D2). For example, cows with high muscularity conditions are less likely to stay in herd compared to cows with low conditions (odds ratio: 5 DIM: 0.806, 45 DIM: 0.784, 160 DIM: 0.775, 280 DIM: 0.872, Figure 1D2).
Potential reasons that can explain some of the results observed may be related to the status of negative energy balance (NEB) that commonly occurs in the periparturient period (54). Grummer et al. (55) estimated energy balance to be around−5.8 Mcal/d when cows are close to parturition, with peaks up to−20 Mcal/d during the 1st month of lactation. Plaizier et al. (56) reported that, in addition to NEB, cows can also experience a negative nitrogen balance in the 1st days after calving. During this phase, dairy cows, especially high-producing ones, cannot fulfill the energy deficit by increasing their feed intake (57). According to what has been reported by Straczek et al. (58), lactating dairy cows are characterized by high plasma levels of leptin, an anorectic hormone, directly related to a high loss of body condition caused by intensive lactogenesis. Therefore, cows are forced to mobilize body reserves, like fat and muscle tissue (33, 58–60). Indeed, even if body fat tissue is identified as the major body source of energy reserves, the catabolism of protein may also contribute to nutrient requirements especially in primiparous and/or early lactation (61). According to Komaragiri et al. (62), during early lactation a cow can lose around 20 kg of muscular tissue and between 8 to 57 kg of body fat. In particular, van der Drift et al. (54) found out that fat mobilization that starts immediately after calving continues up to the 8th week after parturition. Also Schäff et al. (63) observed that skeletal muscle mobilization takes place, starting immediately after calving, but the duration was shorter. In fact, it stopped at about 5 weeks postpartum, with a peak mobilization rate during the first 2 weeks of lactation (63). Findings by Megahed et al. (60) are in accordance with our results, particularly with the fact that body condition of cows facing up to the second calving is crucial to deciding their survival in the herd. Indeed, Megahed et al. (60) reported a greater periparturient mobilization of backfat and skeletal muscle in primiparous than in multiparous cows. The reasons for such greater mobilization in younger animals could be related to the reason they have not finished their growth yet (64) and that they have to cope also with growing requirements, in addition to production and maintenance. Straczek et al. (58) reported that Simmental cows have a greater capacity to adjust the NEB state compared to Holsteins, restoring earlier the BCS loss after the lactation peak. Consequently, cows with good conditions at the onset of second lactation are more prone to perform better along the lactation and to be more resilient to the different metabolic disorders and reduced fertility (65). It is worth considering that farmers rearing Simmentals may decide to cull cows with higher muscularity at a certain point for beef purposes in order to increase the herd profit. Although cows with high BCS are more likely to stay in herd compared to those with low BCS (Figure 2B), the odds ratios for cows with medium BCS were always the highest. Similar results were observed by Erdem et al. (33), who suggested that rearing cows with moderate BCS conditions can be considered an important approach for the herd management. This implies that farmers prefer to cull fat cows to leave space for animals with a medium condition. Probably, dairy farmers are interested to rear cows with appropriate BCS (around 3.0 on a 5-point scale) because these parameter plays an important role in maintaining the health status of lactating cows. As reported by Yasothai (66), dairy cows presenting a severe BCS loss during lactation are exposed to several reproductive problems resulting in longer intervals between first ovulation and estrus, more days open, and lower first-service conception rates. Moreover, literature demonstrates that dairy cows with BCS greater than 3.5 tend to exhibit several metabolic disorders, such as hypocalcaemia, fatty liver, oxidative stress and ketosis (67–70). In addition, fat or over-conditioned dairy cows are at higher risk of developing a combination of metabolic, digestive, infectious and reproductive conditions known as the “fat cow syndrome” (71, 72). Bahrami-Yekdangi et al. (73) reported that in over-conditioned cows (BCS > 3.75; odds ratio = 1.27) the incidence of dystocia was larger than in other cows. An excessive accumulation of body fat predisposes to more insulin resistance, especially during the prepartum, a metabolic disorder with characteristics similar to human type 2 diabetes (74, 75). A transitory phase state of insulin resistance is generally considered a homeorhetic adaptation during early lactation, which provides glucose supply to the mammary gland limiting glucose utilization by insulin-responsive peripheral tissues, such as skeletal muscle or adipose tissue (76). Furthermore, insulin resistance can increase lipolysis of adipose tissue, and the accumulation of non-esterified fatty acids leads in turn to increased insulin resistance. In addition, the high culling risk generally observed for fat cows could be related to the negative relationship between high BCS and milk production (77).
Conclusions
In the present study data of Italian Simmental cows were used to investigated the relationship between STAY and type traits, namely muscularity and BCS. The results indicate that cows characterized by a medium BCS/muscularity are more likely to stay in the herd compared to those with extreme body conditions, i.e., they are more likely to close the lactation and then start the subsequent one. Results of this study provide new insights into the survival and culling of Italian Simmental cattle population. Apart from productivity, in dual-purpose cows type traits and STAY are connected, being indicators of direct voluntary culling with a direct effect on farm's profitability. Further studies should disclose genetic architecture of STAY taking into account muscularity, BCS, and productive performance.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
This study did not require manipulation or modification of the usual handling of the animals, since we have worked directly with the routine records provided by the breeders' associations.
Author contributions
Conceptualization: AF, LD, and GV. Methodology: GV and LD. Software: AC and GV. Formal analysis and writing—original draft preparation: GB, GV, and AC. Data curation: LD, AC, GB, and GV. Writing—review and editing: GV, GN, AC, and NL-V. Visualization: AF, DC, AP, and LM. Supervision: GV, AF, and NL-V. Project administration: GV and AF. All authors have read and agreed to the published version of the manuscript.
Funding
This study was carried out within the Agritech National Research Center and received funding from the European Union Next-Generation EU [Piano Nazionale di Ripresa e Resilienza (PNRR)—Missione 4 Componente 2, Investimento 1.4 – D.D. 1032 17/06/2022, CN00000022]. This manuscript reflects only the authors' views and opinions, neither the European Union nor the European Commission can be considered responsible for them.
Acknowledgments
The authors are very grateful to the Italian Simmental Cattle Breeders Association (ANAPRI) for data supply.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Kristensen T, Jensen C, Østergaard S, Weisbjerg MR, Aaes O, Nielsen NI. Feeding, production, and efficiency of Holstein-Friesian, Jersey, and mixed-breed lactating dairy cows in commercial Danish herds. J Dairy Sci. (2015) 98:263–74. doi: 10.3168/jds.2014-8532
2. Miglior F, Fleming A, Malchiodi F, Brito LF, Martin P, Baes CF, et al. 100-Year Review: Identification and genetic selection of economically important traits in dairy cattle. J Dairy Sci. (2017) 100:10251–71. doi: 10.3168/jds.2017-12968
3. Wall E, White IMS, Coffey MP, Brotherstone S. The relationship between fertility, rump angle, and selected type information in Holstein-Friesian cows. J Dairy Sci. (2005) 88:1521–8. doi: 10.3168/jds.S0022-0302(05)72821-6
4. Dillon P, Berry DP, Evans RD, Buckley F, Horan B. Consequences of genetic selection for increased milk production in European seasonal pasture based systems of milk production. Livest Sci. (2006) 99:141–58. doi: 10.1016/j.livprodsci.2005.06.011
5. Oltenacu P, Broom D. The impact of genetic selection for increased milk yield on the welfare of dairy cows. Anim Welfare. (2010) 19:39–49. doi: 10.1017/S0962728600002220
6. Cole JB, VanRaden PM. Symposium review: Possibilities in an age of genomics: The future of selection indices1. J Dairy Sci. (2018) 101:3686–701. doi: 10.3168/jds.2017-13335
7. Ahlman T, Berglund B, Rydhmer L, Strandberg E. Culling reasons in organic and conventional dairy herds and genotype by environment interaction for longevity. J Dairy Sci. (2011) 94:1568–75. doi: 10.3168/jds.2010-3483
8. Allendorf JJ, Wettemann PJC. Does animal welfare influence dairy farm efficiency? A two-stage approach. J Dairy Sci. (2015) 98:7730–40. doi: 10.3168/jds.2015-9390
9. Fetrow J, Nordlund KV, Norman HD. Invited Review: Culling: Nomenclature, Definitions, and Recommendations. J Dairy Sci. (2006) 89:1896–905. doi: 10.3168/jds.S0022-0302(06)72257-3
10. Kuczyńska B, Puppel K, Gołebiewski M, Wiśniewski K, Przysucha T. Metabolic profile according to the parity and stage of lactation of high-performance Holstein-Friesian cows. Anim Biosci. (2021) 34:575–83. doi: 10.5713/ajas.20.0018
11. Boulton AC, Rushton J, Wathes DC. An empirical analysis of the cost of rearing dairy heifers from birth to first calving and the time taken to repay these costs. Animal. (2017) 11:1372–80. doi: 10.1017/S1751731117000064
12. Liang D, Cabrera VE. Optimizing productivity, herd structure, environmental performance, and profitability of dairy cattle herds. J Dairy Sci. (2015) 98:2812–23. doi: 10.3168/jds.2014-8856
13. Rilanto T, Reimus K, Orro T, Emanuelson U, Viltrop A, Mõtus K. Culling reasons and risk factors in Estonian dairy cows. BMC Vet Res. (2020) 16:173. doi: 10.1186/s12917-020-02384-6
14. Hristov AN, Ott T, Tricarico J, Rotz A, Waghorn G, Adesogan A, et al. SPECIAL TOPICS — Mitigation of methane and nitrous oxide emissions from animal operations: III. A review of animal management mitigation options1. J Anim Sci. (2013) 91:5095–113. doi: 10.2527/jas.2013-6585
15. Van Doormaal BJ, Schaeffer LR, Kennedy BW. Estimation of genetic parameters for stayability in Canadian Holsteins. J Dairy Sci. (1985) 68:1763–9. doi: 10.3168/jds.S0022-0302(85)81025-0
16. Tsuruta S, Misztal I, Lawlor TJ. Changing definition of productive life in us holsteins: effect on genetic correlations. J Dairy Sci. (2005) 88:1156–65. doi: 10.3168/jds.S0022-0302(05)72782-X
17. Hudson GFS, Van Vleck LD. Relationship between production and stayability in Holstein cattle. J Dairy Sci. (1981) 64:2246–50. doi: 10.3168/jds.S0022-0302(81)82836-6
18. Handcock RC, Lopez-Villalobos N, McNaughton LR, Back PJ, Edwards GR, Hickson RE. Body weight of dairy heifers is positively associated with reproduction and stayability. J Dairy Sci. (2020) 103:4466–74. doi: 10.3168/jds.2019-17545
19. Hardie LC, Heins BJ, Dechow CD. Genetic parameters for stayability of Holsteins in US organic herds. J Dairy Sci. (2021) 104:4507–15. doi: 10.3168/jds.2020-19399
20. Jamrozik J, McGrath S, Kemp RA, Miller SP. Estimates of genetic parameters for stayability to consecutive calvings of Canadian Simmentals by random regression models1. J Anim Sci. (2013) 91:3634–43. doi: 10.2527/jas.2012-6126
21. Pritchard T, Coffey M, Mrode R, Wall E. Understanding the genetics of survival in dairy cows. J Dairy Sci. (2013) 96:3296–309. doi: 10.3168/jds.2012-6219
22. Hu H, Mu T, Ma Y, Wang X, Ma Y. Analysis of longevity traits in Holstein cattle: a review. Front Genet. (2021) 12:695543. doi: 10.3389/fgene.2021.695543
23. Jovanovac S. RaguŽ Nikola. Analysis of the relationships between type traits and longevity in croatian simmental cattle using survival analysis. Agric Conspec Sci. (2011) 76:249–53.
24. Schneider MDP, Dürr JW, Cue RI, Monardes HG. Impact of type traits on functional herd life of Quebec Holsteins assessed by survival analysis. J Dairy Sci. (2003) 86:4083–9. doi: 10.3168/jds.S0022-0302(03)74021-1
25. Imbayarwo-Chikosi VE, Ducrocq V, Banga CB, Halimani TE, Wyk JBV, Maiwashe A, et al. Impact of conformation traits on functional longevity in South African Holstein cattle. Anim Prod Sci. (2016) 58:481–8. doi: 10.1071/AN16387
26. Banca Dati Nazionale (BDN). Anagrafe Zootecnica. Metadata. (2022). Available online at: https://www.vetinfo.it (accessed May 15, 2022).
27. ISTAT. Metadata. (2022). Available online at: https://www.istat.it/ (accessed May 15, 2022).
28. Dairy Records Management Systems. DHI Glossary. (2006). Available online at: http://www.drms.org/PDF/materials/glossary.pdf (accessed May 3, 2022).
29. Ali AKA, Shook GE. An optimum transformation for somatic cell concentration in milk1. J Dairy Sci. (1980) 63:487–90. doi: 10.3168/jds.S0022-0302(80)82959-6
30. Buonaiuto G, Lopez-Villalobos N, Niero G, Degano L, Dadati E, Formigoni A, et al. The application of Legendre Polynomials to model muscularity and body condition score in primiparous Italian Simmental cattle. Ital J Anim Sci. (2022) 21:350–60. doi: 10.1080/1828051X.2022.2032850
31. Handcock RC, Lopez-Villalobos N, McNaughton LR, Back PJ, Edwards GR, Hickson RE. Live weight and growth of Holstein-Friesian, Jersey and crossbred dairy heifers in New Zealand. N Z J Agric Res. (2019) 62:173–83. doi: 10.1080/00288233.2018.1465984
32. Cziszter L-T, Ilie D-E, Neamt R-I, Neciu F-C, Saplacan S-I, Gavojdian D. Comparative study on production, reproduction and functional traits between Fleckvieh and Braunvieh cattle. Asian-Australas J Anim Sci. (2016) 30:666–71. doi: 10.5713/ajas.16.0588
33. Erdem H, Atasever S, Kul E. Relations of body condition score with milk yield and reproduction traits in Simmental cows. Large Anim Rev. (2015) 21:231–4.
34. Perišić P, Skalicki Z, Petrovic MM, Bogdanović V, RuŽić-Muslić D. Simmental cattle breed in different production systems. Biotechnol Anim Husb. (2009) 25:315–26. doi: 10.2298/BAH0906315P
35. Shevhuzhev A, Belik N, Emelyanov E, Tokar A. Milk productivity of Simmental cows Austrian selection. In: Proceedings of the International Scientific Conference. Latvia University of Agriculture (2017). Available online at: http://tf.llu.lv/conference/proceedings2017/Papers/N304.pdf (accessed June 14, 2022).
36. CLAL. Metadata. (2021). Available online at: https://www.clal.it (accessed May 15, 2022).
37. Buonaiuto G, Cavallini D, Mammi LME, Ghiaccio F, Palmonari A, Formigoni A, et al. The accuracy of NIRS in predicting chemical composition and fibre digestibility of hay-based total mixed rations. Ital J Anim Sci. (2021) 20:1730–9. doi: 10.1080/1828051X.2021.1990804
38. Mammi LME, Palmonari A, Fustini M, Cavallini D, Canestrari G, Chapman JD, et al. Immunomodulant feed supplement to support dairy cows health and milk quality evaluated in Parmigiano Reggiano cheese production. Anim Feed Sci Technol. (2018) 242:21–30. doi: 10.1016/j.anifeedsci.2018.05.011
39. Cítek J, Brzáková M, Hanusová L, Hanuš O, Večerek L, Samková E, et al. Gene polymorphisms influencing yield, composition and technological properties of milk from Czech Simmental and Holstein cows. Anim Biosci. (2021) 34:2–11. doi: 10.5713/ajas.19.0520
40. Li X, Ma X, Xiao N, Long W, Wu Y, Tong P, et al. Change in chemical composition of Simmental crossbred cattle milk improved its physicochemical, nutritional, and processed properties. J Food Sci. (2019) 84:1322–30. doi: 10.1111/1750-3841.14631
41. Franzoi M, Manuelian CL, Penasa M, De Marchi M. Effects of somatic cell score on milk yield and mid-infrared predicted composition and technological traits of Brown Swiss, Holstein Friesian, and Simmental cattle breeds. J Dairy Sci. (2020) 103:791–804. doi: 10.3168/jds.2019-16916
42. Costa A, Egger-Danner C, Mészáros G, Fuerst C, Penasa M, Sölkner J, et al. Genetic associations of lactose and its ratios to other milk solids with health traits in Austrian Fleckvieh cows. J Dairy Sci. (2019) 102:4238–48. doi: 10.3168/jds.2018-15883
43. Sorensen MT, Nørgaard JV, Theil PK, Vestergaard M, Sejrsen K. Cell turnover and activity in mammary tissue during lactation and the dry period in dairy cows. J Dairy Sci. (2006) 89:4632–9. doi: 10.3168/jds.S0022-0302(06)72513-9
44. Wathes DC, Cheng Z, Bourne N, Taylor VJ, Coffey MP, Brotherstone S. Differences between primiparous and multiparous dairy cows in the inter-relationships between metabolic traits, milk yield and body condition score in the periparturient period. Domest Anim Endocrinol. (2007) 33:203–25. doi: 10.1016/j.domaniend.2006.05.004
45. De Vries A, Marcondes MI. Review: Overview of factors affecting productive lifespan of dairy cows. Animal. (2020) 14:s155–64. doi: 10.1017/S1751731119003264
46. Pinedo PJ, De Vries A, Webb DW. Dynamics of culling risk with disposal codes reported by Dairy Herd Improvement dairy herds. J Dairy Sci. (2010) 93:2250–61. doi: 10.3168/jds.2009-2572
47. Garcia-Peniche TB, Cassell BG, Misztal I. Effects of breed and region on longevity traits through five years of age in Brown Swiss, Holstein, and Jersey Cows in the United States. J Dairy Sci. (2006) 89:3672–80. doi: 10.3168/jds.S0022-0302(06)72407-9
48. Zuchtdata. Metadata. (2021). Available online at: https://www.rinderzucht.at (accessed July 21, 2022).
49. Hare E, Norman HD, Wright JR. Survival Rates and Productive Herd Life of Dairy Cattle in the United States. J Dairy Sci. (2006) 89:3713–20. doi: 10.3168/jds.S0022-0302(06)72412-2
50. Williams M, Sleator RD, Murphy CP, McCarthy J, Berry DP. Re-assessing the importance of linear type traits in predicting genetic merit for survival in an aging Holstein-Friesian dairy cow population. J Dairy Sci. (2022) 105:7550–63. doi: 10.3168/jds.2022-22026
51. Padilla CJ, Harrigan ME, Harris H, Schwab JM, Rutkove SB, Rich MM, et al. Profiling age-related muscle weakness and wasting: neuromuscular junction transmission as a driver of age-related physical decline. GeroScience. (2021) 43:1265–81. doi: 10.1007/s11357-021-00369-3
52. Costagliola A, Wojcik S, Pagano TB, De Biase D, Russo V, Iovane V, et al. Age-related changes in skeletal muscle of Cattle. Vet Pathol. (2016) 53:436–46. doi: 10.1177/0300985815624495
53. Knob DA, Thaler Neto A, Schweizer H, Weigand AC, Kappes R, Scholz AM. Energy balance indicators during the transition period and early lactation of purebred Holstein and Simmental cows and their crosses. Animals. (2021) 11:309. doi: 10.3390/ani11020309
54. van der Drift SGA, Houweling M, Schonewille JT, Tielens AGM, Jorritsma R. Protein and fat mobilization and associations with serum β-hydroxybutyrate concentrations in dairy cows. J Dairy Sci. (2012) 95:4911–20. doi: 10.3168/jds.2011-4771
55. Grummer RR, Mashek DG, Hayirli A. Dry matter intake and energy balance in the transition period. Vet Clin North Am Food Anim Pract. (2004) 20:447–70. doi: 10.1016/j.cvfa.2004.06.013
56. Plaizier JC, Martin A, Duffield T, Bagg R, Dick P, McBride BW. Effect of a prepartum administration of monensin in a controlled-release capsule on apparent digestibilities and nitrogen utilization in transition dairy cows. J Dairy Sci. (2000) 83:2918–25. doi: 10.3168/jds.S0022-0302(00)75192-7
57. Gáspárdy A, Schwartz Z, Zöldág L, Veresegyházy T, Fekete S. Changes in daily energy amounts of main milk components (lactose, protein and fat) during the lactation of high-yielding dairy cows. Acta Vet Hung. (2005) 52:457–67. doi: 10.1556/avet.52.2004.4.8
58. Straczek I, Młynek K, Danielewicz A. The Capacity of Holstein-Friesian and Simmental Cows to Correct a Negative Energy Balance in Relation to Their Performance Parameters, Course of Lactation, and Selected Milk Components. Animals. (2021) 11:1674. doi: 10.3390/ani11061674
59. Formigoni A, Pezzi P, Gramenzi A, Martino G, Neri E. Effect of body condition score variation on milk yield and fertility in post-partum dairy cows. Vet Res Commun. (2003) 27:647–9. doi: 10.1023/B:VERC.0000014238.09424.ee
60. Megahed AA, Hiew MWH, Ragland D, Constable PD. Changes in skeletal muscle thickness and echogenicity and plasma creatinine concentration as indicators of protein and intramuscular fat mobilization in periparturient dairy cows. J Dairy Sci. (2019) 102:5550–65. doi: 10.3168/jds.2018-15063
61. Chibisa GE, Gozho GN, Van Kessel AG, Olkowski AA, Mutsvangwa T. Effects of peripartum propylene glycol supplementation on nitrogen metabolism, body composition, and gene expression for the major protein degradation pathways in skeletal muscle in dairy cows. J Dairy Sci. (2008) 91:3512–27. doi: 10.3168/jds.2007-0920
62. Komaragiri MVS, Casper DP, Erdman RA. Factors Affecting Body Tissue Mobilization in Early Lactation Dairy Cows. 2 effect of dietary fat on mobilization of body fat and Protein1. J Dairy Sci. (1998) 81:169–75. doi: 10.3168/jds.S0022-0302(98)75564-X
63. Schäff C, Börner S, Hacke S, Kautzsch U, Sauerwein H, Spachmann SK, et al. Increased muscle fatty acid oxidation in dairy cows with intensive body fat mobilization during early lactation. J Dairy Sci. (2013) 96:6449–60. doi: 10.3168/jds.2013-6812
64. Wathes DC, Brickell JS, Bourne NE, Swali A, Cheng Z. Factors influencing heifer survival and fertility on commercial dairy farms. Animal. (2008) 2:1135–43. doi: 10.1017/S1751731108002322
65. Cavallini D, Mammi LME, Palmonari A, García-González R, Chapman JD, McLean DJ, et al. Effect of an immunomodulatory feed additive in mitigating the stress responses in lactating dairy cows to a high concentrate diet challenge. Animals. (2022) 12:2129. doi: 10.3390/ani12162129
66. Yasothai R. Importance of energy on reproduction in dairy cattle. Int J Sci Environ Technol. (2014) 3:2020–3.
67. Gillund P, Reksen O, Gröhn YT, Karlberg K. Body condition related to ketosis and reproductive performance in Norwegian dairy cows. J Dairy Sci. (2001) 84:1390–6. doi: 10.3168/jds.S0022-0302(01)70170-1
68. Bernabucci U, Ronchi B, Lacetera N, Nardone A. Influence of body condition score on relationships between metabolic status and oxidative stress in periparturient dairy cows. J Dairy Sci. (2005) 88:2017–26. doi: 10.3168/jds.S0022-0302(05)72878-2
69. Mahjoubi E, Amanlou H, Zahmatkesh D, Ghelich Khan M, Aghaziarati N. Use of beet pulp as a replacement for barley grain to manage body condition score in over-conditioned late lactation cows. Anim Feed Sci Technol. (2009) 153:60–7. doi: 10.1016/j.anifeedsci.2009.06.009
70. Roche JR, Friggens NC, Kay JK, Fisher MW, Stafford KJ, Berry DP. Invited review: Body condition score and its association with dairy cow productivity, health, and welfare. J Dairy Sci. (2009) 92:5769–801. doi: 10.3168/jds.2009-2431
71. Roche JR, Kay JK, Friggens NC, Loor JJ, Berry DP. Assessing and managing body condition score for the prevention of metabolic disease in dairy cows. Vet Clin N Am. (2013) 29:323–36. doi: 10.1016/j.cvfa.2013.03.003
72. Bogaert H, De Koster J, Van den Broeck W, Van Eetvelde M, Opsomer G. Effects of overconditioning on pancreatic insulin secretory capacity, fat infiltration, and the number and size of islets in dairy cows at the end of the dry period. J Dairy Sci. (2018) 101:11413–20. doi: 10.3168/jds.2018-14931
73. Bahrami-Yekdangi M, Ghorbani GR, Sadeghi-Sefidmazgi A, Mahnani A, Drackley JK, Ghaffari MH. Identification of cow-level risk factors and associations of selected blood macro-minerals at parturition with dystocia and stillbirth in Holstein dairy cows. Sci Rep. (2022) 12:5929. doi: 10.1038/s41598-022-09928-w
74. Wu J, Liu J, Wang D. Effects of body condition on the insulin resistance, lipid metabolism and oxidative stress of lactating dairy cows. Lipids Health Dis. (2020) 19:56. doi: 10.1186/s12944-020-01233-7
75. Prodanović R, Kirovski D, Vujanac I, Djordjevic A, Romić S, Pantelić M, et al. Obesity-related prepartal insulin resistance in dairy cows is associated with increased lipin 1 and decreased FATP 1 expression in skeletal muscle. Res Vet Sci. (2022) 150:189–94. doi: 10.1016/j.rvsc.2022.04.012
76. Contreras GA, Strieder-Barboza C, Raphael W. Adipose tissue lipolysis and remodeling during the transition period of dairy cows. J Anim Sci Biotechnol. (2017) 8:41. doi: 10.1186/s40104-017-0174-4
Keywords: culling, dual-purpose cows, functional longevity, resilience, survival rate
Citation: Buonaiuto G, Lopez-Villalobos N, Costa A, Niero G, Degano L, Mammi LME, Cavallini D, Palmonari A, Formigoni A and Visentin G (2023) Stayability in Simmental cattle as affected by muscularity and body condition score between calvings. Front. Vet. Sci. 10:1141286. doi: 10.3389/fvets.2023.1141286
Received: 10 January 2023; Accepted: 01 March 2023;
Published: 24 March 2023.
Edited by:
Nikola Čobanović, University of Belgrade, SerbiaReviewed by:
Alberto Sabbioni, University of Parma, ItalyAndras Gaspardy, University of Veterinary Medicine Budapest, Hungary
Copyright © 2023 Buonaiuto, Lopez-Villalobos, Costa, Niero, Degano, Mammi, Cavallini, Palmonari, Formigoni and Visentin. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ludovica Maria Eugenia Mammi, bHVkb3ZpY2EubWFtbWlAdW5pYm8uaXQ=