
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Vet. Sci. , 24 March 2023
Sec. Animal Nutrition and Metabolism
Volume 10 - 2023 | https://doi.org/10.3389/fvets.2023.1063216
 Mathilde Porato1*†
Mathilde Porato1*† Stéphanie Noël1†
Stéphanie Noël1† Joël Pincemail2
Joël Pincemail2 Adelin Albert3Jean-Paul Cheramy-Bien2Caroline Le Goff4
Adelin Albert3Jean-Paul Cheramy-Bien2Caroline Le Goff4 Annick Hamaide1
Annick Hamaide1Introduction: While oxidative stress has been studied in pathologic conditions in dogs, data in presumably healthy dogs and standardized protocols are lacking. This work purposed to bridge the gap by presenting provisional physiological ranges for oxidative stress biomarkers in a group of Beagle dogs.
Methods: Based on our long-standing clinical expertise in the field of oxidative stress, nine plasma biomarkers of oxidative stress were evaluated for their concentrations (mean ± SD) in 14 healthy adult Beagle dogs.
Results: Selected biomarkers were: vitamins C (7.90 ± 1.36 μg/mL) and E (34.1 ± 6.63 μg/mL), zinc (0.80 ± 0.17 mg/L), copper (0.54 ± 0.048 mg/L), selenium (256 ± 25.7 μg/L), total and oxidized glutathione (822 ± 108 μM and 3.56 ± 1.76 μM), myeloperoxidase (67.4 ± 56.2 ng/mL), and isoprostanes (340 ± 95.3 ng/mL). Glutathione peroxidase activity and superoxide anion production in whole blood were also measured. Glutathione peroxidase activity was 473 ± 34.0 IU/g of hemoglobin and superoxide anion production in whole blood was 18,930 ± 12,742 counts per 30 min. Reduced glutathione/oxidized glutathione and copper/zinc ratios were, respectively, 280 ± 139 and 0.70 ± 0.15. Sex-related differences were recorded for zinc (p = 0.0081), copper/zinc ratio (p = 0.0036) and plasma isoprostanes (p = 0.0045).
Conclusion: Provisional physiological norms covering 95% of our group were proposed for each biomarker and should be of interest for future studies of canine oxidative stress.
Oxidative stress has been defined as an imbalance between oxidants (e.g., free radical species derived from oxygen) and antioxidants in favor of oxidants, leading to a disruption of redox signaling and/or molecular damage to lipids, DNA and proteins (1). This definition takes into account both physiological and pathological effects of reactive oxygen species (ROS) (2). Produced in excess, ROS (including free radical (superoxide anion, hydroxyl radical) and non-free radical (hydrogen peroxide, hypochlorous acid) entities) induce irreversible oxidative damages to lipids, DNA and proteins that were associated with the development of diseases in humans (3). By contrast, ROS may also have important physiological actions when produced in low concentration, ROS may act as second messengers capable of regulating apoptosis, activating transcription factors and modulating the expression of various genes involved in the immune response (3). To regulate the excess of ROS production, both human and animal organisms respond with antioxidants (vitamins A, C and E, carotenoids, glutathione, ubiquinone and polyphenols), antioxidant enzymes (superoxide dismutase which catalyzes the dismutation of superoxide anions to hydrogen peroxide, glutathione peroxidase which catalyzes the reduction of several hydroperoxides to H2O via oxidation of reduced glutathione), trace elements (copper, zinc, and selenium), iron chelators and inhibitors of biological systems producing ROS (4).
Because ROS are short half-life molecules, the accurate detection of oxidative stress status (OSS) may reveal technically challenging. Moreover, it must be kept in mind that a single test does not reflect the presence of OS. Thus, in human studies, the first step to assess OS requires the use of a large battery of assays including determination of enzymes and low molecular weight antioxidants, analysis of trace elements, evidence of oxidative damages to lipids, proteins or DNA and identification of the sources responsible of high ROS production (5). The second step includes careful pre-analytical treatment of the sample since it may lead to artifactual values in the event of non-compliance with the protocol. This is of primordial importance to determine values for blood or urine OS biomarkers and, therefore, to correctly interpret the results (6). Such a rigorous methodology often lacks in canine research.
In dogs, several studies have evidenced increased OS in systemic diseases such as congestive heart failure, anemia, atopic dermatitis, mammary carcinoma, spinal cord injury or inflammatory bowel disease (7–16) when compared to healthy controls. More precisely, ill dogs have increased oxidant marker levels while endogenous antioxidants and biomarkers of the oxidant response concomitantly decrease in whole blood (12, 17–21), in serum (10, 13, 18, 20, 22), in plasma (12, 13, 17, 18, 20, 22), in urine (11) or in cerebrospinal fluid and spinal cord tissue (11). Other studies have described the values of several OS blood biomarkers in populations of healthy dogs from different breeds (17, 23) or from a single breed but at different ages (18), with the intent of addressing the effects of sex, age, and/or nutrition discrepancy. Despite these observations, basic information about physiological intervals of OS biomarkers in dogs is still lacking especially according to different breeds. Conversely, clinical studies rather defined a control population composed of healthy dogs of different environments, breeds, sex, reproductive status and ages that do not always match the study population. Consequently, and in spite of scientifically robust study designs, comparison between studies is limited whereas many of them tend to demonstrate that ill dogs experience an OS comparable to healthy subjects (8–11, 13, 24). Values obtained from homogeneous groups of Beagle dogs, with rigorous pre-analytical steps, would be beneficial to determine the baseline in a common breed, but to our knowledge, such data is lacking. This breed has already been chosen for the validation of OS biomarkers assays (erythrocyte osmotic fragility and detergent sensitivity to assess plasma alpha-tocopherol concentration) (19) and to determine the best storage conditions of serum for postponed measurement of OS biomarkers (20). The investigation of age-related and sex-related lipid peroxidation has also been made by comparing OS biomarkers in whole blood of different groups of Beagle dogs (22). The effect of vitamin C and E supplementation on blood OS biomarkers was also tested in this breed (21).
Reliable inter-individual physiological margins for OS biomarkers can be provided by thorough fundamental studies performed in a standardized population. Based on a highly-qualified expertise in humans, we initiated the present study to determine the OS biomarkers level in healthy Beagle dogs, respectively non-enzymatic anti-oxidants (glutathione, vitamins C and E), enzymatic antioxidant (glutathione peroxidase), trace elements (copper, zinc and selenium), inflammatory markers (myeloperoxidase and neutrophil activation) and specific markers of lipid peroxidation (isoprostanes). This allowed us to estimate mean and dispersion values and to propose provisional physiological norms for all selected OS biomarkers in our healthy dogs. Our results may serve as a reliable basis for future studies involving dogs, especially Beagle dogs.
Six healthy intact female Beagle dogs and eight healthy intact male Beagle dogs were included in the study. Females belonged to 4 different litters and males belonged to two different litters. Dogs were 4.1 ± 0.8 years old. They were born and housed at the local animal facilities and were kept in the same kennel, had unrestricted access to water and were fed with commercial dry food. Male and female dogs had different diets to provide a diet with a lower lipid content to female dogs (Light weight care medium, Royal Canin®) compared to males (Adult Beagle, Royal Canin®). Dogs did not receive any oral anti-oxidant supplementation. Diet composition in relation to requirement for dogs is presented in Table 1. Females were in the anestrus phase of the estrus cycle at the time of sample collection. The animal study was reviewed and approved by Commission d'Ethique Animale de l'Université de Liège (reference 2123, May 2019).
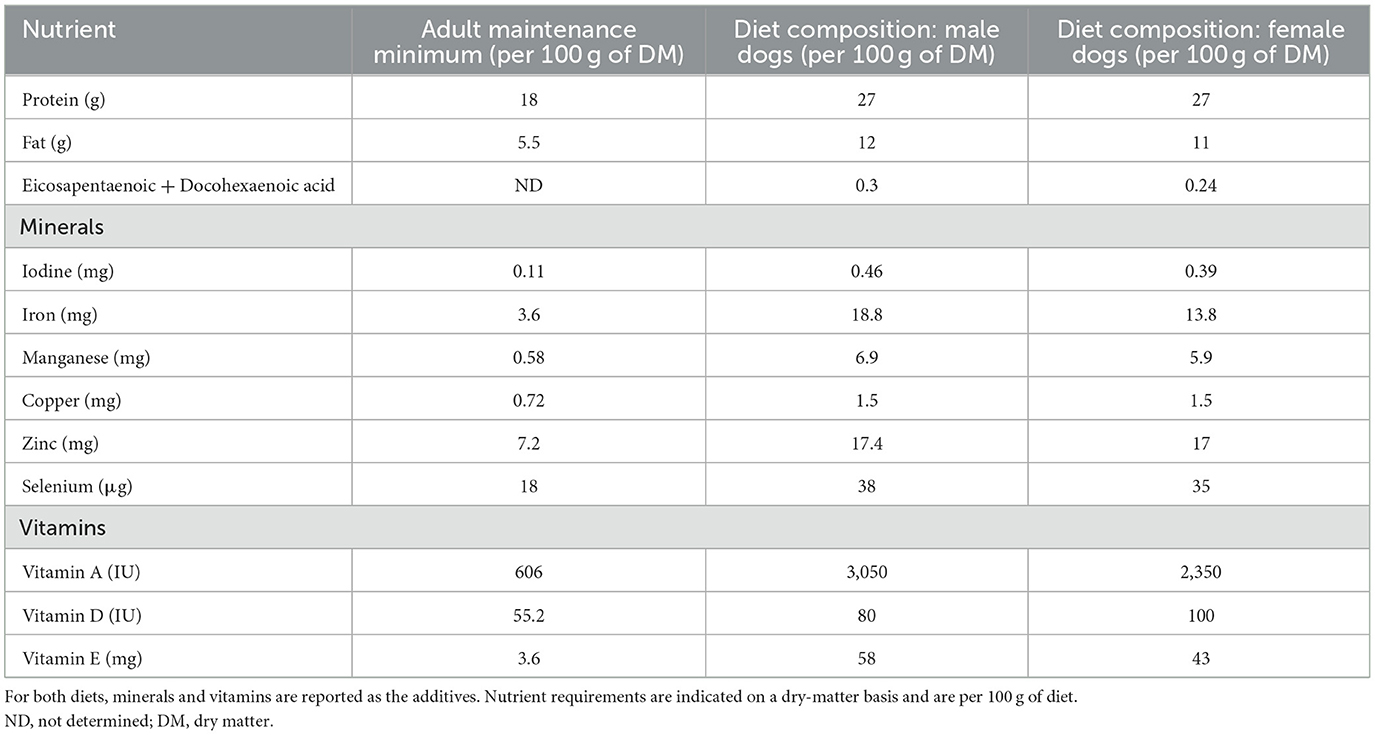
Table 1. Diet composition and recommended nutrient levels for dogs according to the European pet food industry federation guidelines (2020), based on metabolizable energy requirement of 110 kcal/kg0.75.
Venous blood samples were collected from 8 a.m. to 9 a.m. from fasted animals in vacutainer tubes containing Na heparin or EDTA according to the tested parameter. All dogs were fasted at least 12 h before blood sampling. External jugular venipuncture was performed with a 21-gauge needle, and blood samples were collected in the appropriate tubes. Part of whole blood was removed for reduced and oxidized glutathione and glutathione peroxidase determination. Then both tubes were centrifuged at 1,500 g during 10 min. 500 μL of plasma EDTA were immediately added to 500 μL metaphosphoric acid (10%) for vitamin C stabilization and then frozen on dry ice. Another aliquot of 1 mL plasma EDTA was added to 5 μL butylhydroxytoluene at a concentration of 2 mg/mL in ethanol for the measurement of isoprostanes and then frozen on dry ice. The rest of EDTA plasma and serum were then discarded into separated tubes for analysis of vitamin E, myeloperoxidase, copper, zinc and selenium and frozen on dry ice. All samples were then stored at −80°C until analysis of oxidative stress biomarkers.
Reduced (GSH) and oxidized (GSSG) levels were measured in whole blood using the BIOXYTECH® GSH/GSSG-412™ assay kit (OxisResearch™, Oxis Health Product, Inc., Portland, OR, USA). Briefly, 50 μL of whole blood were collected and immediately stored at −80°C for total glutathione (GSHt) measurement. For oxidized glutathione (GSSG) measurement, 100 μL whole blood was added to 10 μL 1-methyl-2-vinylpyridinium trifluoromethanesulfonate to avoid glutathione oxidation and then immediately stored at −80°C. The method employs Ellman's reagent (5,5'-dithiobis-2-nitrobenzoic acid), which reacts with GSH to form a spectrophotometrically detectable product at 412 nm. GSSG can be determined by the reduction of GSSG into GSH. The following formula was used to calculate the reduced/oxidized glutathione ratio: GSH/GSSG = ((GSHt −2GSSG)/GSSG).
Plasma vitamin C was determined by a spectrophotometric method (520 nm) using the reduction of 2,6-dichlorophenolindophenol (Perkin Elmer Lambda 40, Norwalk, CT, USA) (25).
Plasma samples added to butylhydroxytoluene were purified through an original solid-liquid extraction protocol (26). After evaporation to dryness and reconstitution, samples were directly injected into a Waters XevoTM TQ-S mass spectrometer (Milford, MA, USA) equipped with an electrospray ionization source operating in positive ion mode and interfaced with a Waters Acquity UPLC I-Class inlet system (Miflord, MA, USA). Data acquisition was achieved using MassLynx Version 4.2 software (Milford, MA, USA) and TargetLynx version 4.2 software to Tandem Mass Spectrometry (UHPLC/MS-MS) method (Miflord, MA, USA).
Analysis of vitamin E (alpha and gamma-tocopherol) was performed by HPLC procedure (Alliance Waters, Washington, DC, USA) coupled with a diode array detector (PDA 2996, Waters, Milford, MA, USA) using Chromsytems kits (32,000, 34,000, and 68,000) (Chromsystems, Gräfelfing-Munich, Germany).
Copper, zinc, and selenium were detected in serum by inductively coupled plasma-mass spectroscopy (27).
Glutathione peroxidase (GPx) activity was determined in 100 μL whole blood using Ransel kit (Randox Laboratories Ltd, County Antrim, New England). Briefly, GPx catalyzes the oxidation of GSH by cumene hydroperoxide. In presence of glutathione reductase and NADPH, the oxidized glutathione (GSSG) was converted to the reduced form with a concomitant oxidation of NADPH to NADP+. The decrease in absorbance at 340 nm was measured. Values expressed as International Unit (IU) were standardized to hemoglobin (IU/g Hb).
Plasma myeloperoxidase was assessed using the specific ELISA kit CSB-E12762C developed by Cusabio, Houston, TX, USA (detection range: 31.25–2,000 ng/mL; intra-assay precision: CV < 8%; inter-assay precision: CV < 10%).
Neutrophil count was measured with a commercial veterinary analyzer (Procyte Dx, IDEXX Laboratories, Inc, Westbrook, USA).
Determination of superoxide anion production by NADPH oxidase in blood was assessed by the protocol described by Baptista et al. (28). Using 96 well plate, superoxide anion production was determined in 200 μL whole blood, collected in tubes containing EDTA as anticoagulant and diluted in 820 μL Dulbecco's Modified Eagle Medium and 200 μL lucigenin (1.5 x 10 μM). After 20 min of incubation at 37°C under gentle agitation, whole blood was stimulated by using phorbol myristate acetate (at the concentration of 0.1 μM) as a specific activator of NADPH oxidase, leading to superoxide anion production. The luminescence was immediately recorded at 37°C during 30 min using a luminometer GloMax® Multi Microplate Multimode (Promega, Madison, WI, USA). Phorbol 12-myristate 13-acetate (PMA), N,N' Dimethyl-9,9'-biacridinium dinitrate (lucigenin) and fluorescein sodium salt were purchased from Sigma-Aldrich (St Louis, MI, USA).
Results were expressed as mean and standard deviation (SD). The Shapiro-Wilk (SW) test was used to assess the normality of the distribution of parameters. When significant, values were log-transformed to normalize their distribution. Mean values of male and female dogs were compared by the non-parametric Kruskal-Wallis (KW) test. Provisional physiological ranges containing 95% of the dogs were determined by computing 95%-content tolerance intervals at the 95% confidence level according to Guttman (29) and Eisenhart et al. (30). Specifically, given the mean m and the standard deviation SD of a sample of size N, the 95% confidence 95%-content tolerance interval is determined by computing the lower and upper limits m ± K.SD, where K values can be found in a published table (30). As an example, for N = 14, K = 3.012. Actually, as N tends to infinity, K gets closer and closer to the traditional 1.96 critical point of the standard Normal distribution. Results were considered significant at the 5% critical level (P < 0.05). Calculations were performed with R (version 4.1.0, Vienna, Austria).
The mean body weight of the dogs was 15.8 ± 1.2 kg in females and 17.3 ± 1.5 kg in males (p = 0.017). The body condition score (BCS, Body condition score chart, Ralston Purina Company, St Louis, MO) averaged 6.5 ± 1.2 in females and 5.3 ± 0.7 in males. All dogs, excepted females 9 and 11, had a BCS between 4 and 6. There was no significant difference in BCS between males and females.
The healthiness of the dogs was confirmed by the absence of illness history during the 2 weeks preceding the study, by a normal physical examination, normal complete blood count, normal serum biochemical analyses and normal urinalysis obtained during the week preceding the study. Specifically, plasma cholesterol concentration was similar to previously reported range (92–324 mg/dL) (31).
The diet was produced by a manufacturer according to the European Pet Food Industry Federation guidelines. These European guidelines are adapted from the guidelines of the National Research Council.
The distribution characteristics of all oxidative stress parameters are given in Table 2. Three parameters did not follow a Gaussian distribution, namely vitamin E (p = 0.0059), myeloperoxidase (p = 0.0004) and to a lesser extent superoxide anion production in whole blood (p = 0.012). A log-transform was applied to normalize their distribution. Of note, for myeloperoxidase, given the presence of 8 right censored values (< 31 ng/mL), the tolerance interval was based on the median (rather than the mean) and the SD calculated from the Tukey formula SD = 0.74 IQR, where IQR is the interquartile range.
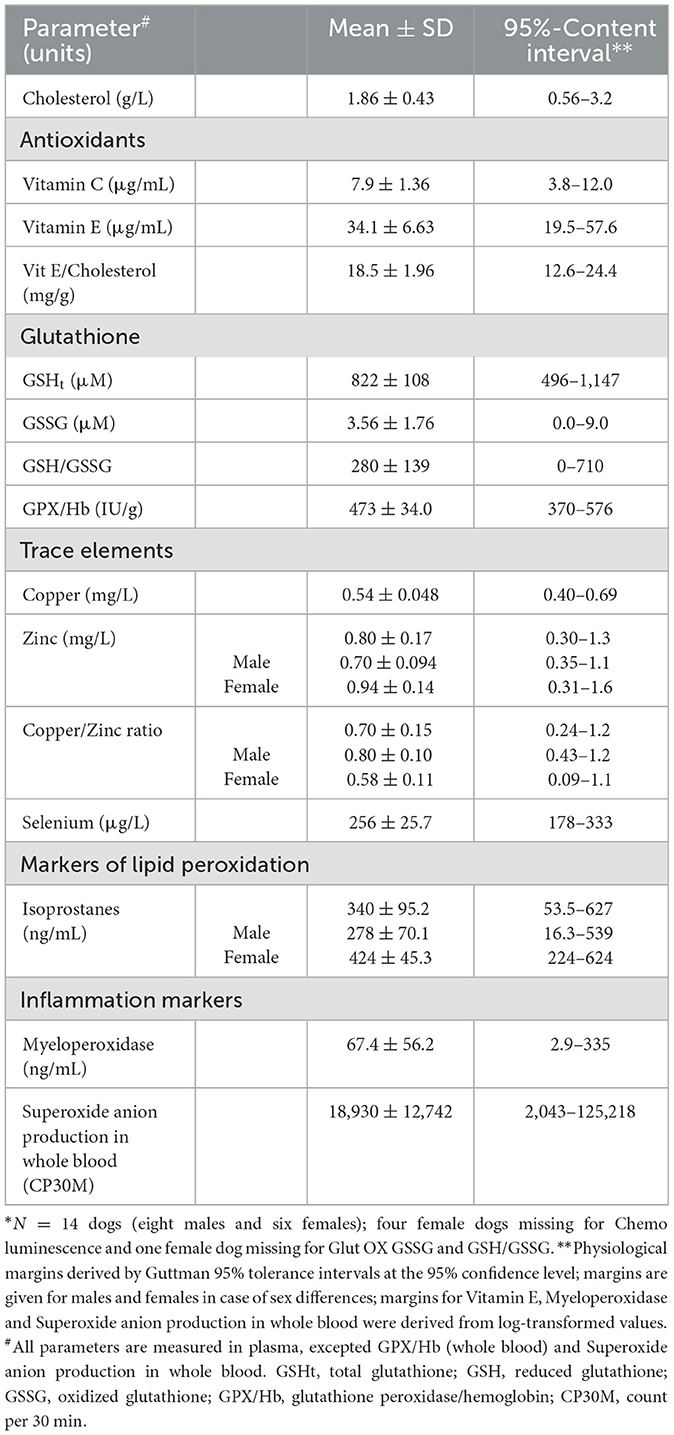
Table 2. Distribution characteristics and provisional 95%-content interindividual physiological intervals of oxidative stress parameters in presumably healthy Beagle dogs (N = 14)*.
Plasma concentrations in vitamins C and E, total glutathione, oxidized glutathione, copper, selenium, myeloperoxidase, the vitamin E/cholesterol ratio and glutathione peroxidase activity did not differ between males and females. Plasma concentrations in zinc were significantly higher in females than in males (p = 0.0081) and similarly for isoprostanes (p = 0.0045). By contrast, the plasma copper/zinc ratio was significantly higher in males compared to females (p = 0.0036). Superoxide anion production in PMA whole blood test did not differ between males and females.
Using mean and SD values, provisional physiological norms were determined for each OS biomarker by using the 95%-content tolerance intervals method. These norms are displayed in Table 2 not only globally but also according to sex when differences were observed between males and females.
Levels of OS biomarkers in canine plasma or urine may vary with age (22, 32), sex (22, 23, 33), diet (18) or with the amount of time spent by the dog in the veterinary facility before sampling (34). Details about pre-analytical sample collection and handling are not always mentioned so that doubt regarding potential artifactual oxidation can raise. Moreover, methodology varies widely between studies, further preventing comparisons within literature data.
The definition of a reference population is of major concern when reporting biological data, in particular when it comes to compare study results. Most studies are clinical studies, dealing with a heterogeneous population of healthy dogs as a control/baseline group, with subjects of different breeds, age, sex and sexual status, which induces a great variability within results. For example, Freeman et al. reported OS biomarkers concentrations in a population of dogs from several breeds (4 to 71 kg) and of different ages (8.8 ± 2.6 years old) to compose the healthy group (7). In other studies, Beagle (19, 20, 22) and Labrador (18) dogs were favored. Mostly, the groups of healthy dogs that serve as control groups include about 10 to 20 dogs of different breeds, ages and sexes (9, 11, 33, 35). We chose a group of intact young adult Beagle dogs, which represent a typical group of interest in veterinary canine fundamental research, with individuals of medium size and good temper. We assessed OS biomarkers from this homogenous group of dogs with a validated protocol and standardized pre-analytical steps. Under these conditions, we demonstrated (i) that the methodology is deeply involved in the consistency of the results, which should encourage future studies to apply such a protocol, and (ii) that our results are highly valuable in the field of fundamental research on canine physiopathological mechanisms, where reliable physiological margins for OS biomarkers are absent.
The values of OS biomarkers were similar in male and female dogs, except for zinc and isoprostanes plasma concentrations and for copper/zinc ratio. The difference in plasma zinc concentration between males and females was not attributable to the difference in diet since zinc represented, respectively 17.4 mg and 17 mg per 100 g of dry matter in the diet received by both groups. The females in the present study were in the anestrus phase of the reproductive cycle to reduce the potential hormonal influence on plasma zinc concentration, as observed in women. Indeed, in women, the changes in plasma zinc concentration are positively correlated with the changes in plasma estradiol concentration during the menstrual cycle (36). Despite the sex-related difference and despite lower copper and zinc plasma concentrations in our dogs, copper/zinc ratio was similar to the ratio obtained in a heterogenous population of dogs of different breeds (37). Copper and zinc are mineral micronutrients acting as cofactors of certain endogenous antioxidant enzymes (37, 38). In human studies, the copper/zinc ratio is related to the systemic oxidative status: the higher the ratio, the more lipidic peroxidation due to high copper concentration (39).
Selenium intake was 3 μg per 100 g of dry matter lower in females than in males. It was therefore important to check if this could impact the levels of the other OS biomarkers. Levels in selenium (p = 0.33), but also those of glutathione peroxidase (p = 0.25) requiring this trace element for its antioxidant activity, were perfectly similar in both groups. It is therefore unlikely that the slight difference in selenium intake between the two groups will have an impact on the concentration of the other OS biomarkers. A similar conclusion can be drawn for iron. Indeed, about 70% of body's iron is found as hemoglobin in the red blood cells. There was no difference in hemoglobin levels between males (17.7 ± 1.3 g/dL) and females (16.8 ± 1.5 g/dL) (p = 0.55). Therefore, we concluded that even if the diet was different between males and females regarding selenium and iron composition, this should have no impact on the concentration of the other OS biomarkers.
Isoprostanes, as lipid peroxidation products, have been measured in plasma (7, 40) and in urine of dogs (8, 9, 11, 12, 15, 35) with conflicting results, due to different methods of analysis and ways to express the results. Indeed, immunoassays have been reported to lack consistency and specificity both in human and veterinary studies (41). For example, plasma median values 10,000 times lower than ours have been reported by Freeman et al. (median 0.02529 ng/mL, range 0.0111–0.0804 ng/mL) (7). Such a discrepancy is due to assay inaccuracy and to pre-analytic methodology. Indeed, in that study, plasma isoprostanes were measured with a commercial ELISA kit and samples were processed within 15 min. Conversely, our results rely on immediate processing and standardized analytic methods (UHPLC/MS-MS) as used in human studies (26, 28). Recently, Putman et al. concluded that the measurement of isoprostanes in veterinary medicine should preferably be done by UHPLC/MS-MS method rather than using ELISA kits (41). The reasons are twofold: (1) consistent overestimation of isoprostanes concentrations in ELISA test due to cross-reactivity with related compounds such as alternative isoprostanes isomers and prostaglandins; (2) poor agreement between different ELISA tests but also with LC/LC-MS/MS. Although the measurement of isoprostanes by ELISA tests is less expensive than that by UHPLC/MS-MS and less time consuming, the latter should be favored especially if high precision is desired (definition of diagnostic cut-off values for example) or if dilutions are required with ELISA kits. It is therefore important to encourage researchers to seek for this possibility whenever possible, because it may represent a specific interest in the future. The hypothesis of methodology discrepancy can also be formulated to explain the extreme variations found in the literature for isoprostanes urinary concentration, with values of 0.00714 ng/mL (8) and 399 pg/mL (9) (median and mean, respectively). Moreover, isoprostanes are detectable in all fluids and tissues of mammals but they may not match with the systemic isoprostanes concentration (41). In our study, assays were also performed but were unsuccessful in determining isoprostanes in urine samples due to the presence of impurities in the matrix interfering with the elution peak. In human studies, a strong positive correlation has been reported between copper/zinc ratio and lipid peroxidation products (42, 43). In our dogs, this was not the case for unexplained reason (isoprostanes and copper/zinc ratio: r = −0.57, p = 0.033, data not shown).
Vitamin C, or ascorbate, is a hydro-soluble ubiquitous carbohydrate and a powerful antioxidant acting as a scavenger of several free radicals and ROS. It can regenerate other molecules from their oxidized form, such as vitamin E, hence inhibiting lipid peroxidation as well (44). Moreover, vitamin C synthesis is possible in all species except in higher-order primates, in guinea pigs and in some bat, fish, and bird species (44). While analytic methods were different, plasma concentrations in vitamin C in the present study were similar compared to the study of Freeman et al. (median 7.2 μg/mL, range 4.3–8.5 μg/mL) in which dogs were fed with commercial food (7). Vitamin E, represented by eight molecules (four tocopherols and four tocotrienols) is a potent antioxidant, made by plants, with lipoperoxyl radical scavenging activities (45). We observed higher plasma concentrations in vitamin E compared to the study of Freeman et al. (median 0.0282 μg/mL, range 0.0174–0.0378 μg/mL) (7). This difference could be attributable to the dosage of alpha-tocopherol only by Freeman et al. It may also be due to differences in the pre-analytical steps, since dosage method was similar to ours. This is further highlighted by the variability between other studies in which control groups are, respectively composed of 14 and 10 adult dogs of different breeds and sexes: median 55 μg/mL, range 18–88 μg/mL by Viviano and VanderWielen (9) and median 24.1 μg/mL, range 16.8–31.2 μg/mL by Verk et al. (33). As a labile compound, dosage of vitamin E relies on sample process and delay during the pre-analytical steps may compromise the quantity of vitamin E in the sample. Moreover, food type and modalities of food preservation might have influenced vitamin E intake by the dogs, contrary to the dogs of our study who were fed the same food. However, the major reason explaining such a high variability in all these studies (7, 9, 33) is due to the fact that the authors do not take in account the cholesterol concentration as we did in the present work. Indeed, we found a highly positive correlation (r = 0.91, p < 0.0001) between both plasma vitamin E and cholesterol concentrations (data not shown), which was expected as it had already been reported in human (46).
Among OS biomarkers, glutathione is a thiol tripeptide synthesized in all mammalian cells. The blood concentrations in oxidized glutathione (median 0.08 μM, range 0.01–0.29 μM) reported by Freeman et al. (7) were lower than ours which is surprising. Indeed, the population of dogs in the study of Freeman et al. was older (8.8 ± 2.2 years old) compared to our subjects. Consequently, higher blood concentrations in oxidized glutathione would have been expected, as it has been described in elderly (47). Again, this illustrates the divergences of the results when obtained from studies built on different protocols and conducted on different populations. The cellular balance of the redox couple formed by glutathione and its oxidized disulfide form provides a dynamic indicator of OS. During acute OS, reduced glutathione concentration decreases and the concentration of its disulfide form increases (24, 48). Glutathione peroxidase is a selenium-dependent enzyme responsible for glutathione oxidation coupled with organic peroxides reduction. Its value in our study was similar to values previously reported in a population of 30 healthy male and female dogs aged 7 months to 11 years old (454 (349–705) IU/g Hb) (17) and was not affected by the difference in diet composition between males and females. Similarly, in the study of Tomsič et al. glutathione peroxidase activity did not differ between males and females despite the different diet provided by the owners (17). A comparable range (394 ± 44 IU/g Hb) was also found in 10 healthy adult dogs (4.4 ± 2.5 years of age) of different breeds and sexes (33).
Myeloperoxidase is an inflammatory marker which illustrates the capacity of a system in producing ROS, such as hypochlorous acid. It is produced in both neutrophils and monocytes. Our plasma levels in myeloperoxidase were higher than those previously reported (0.2733 ± 0.1796 ng/mL) in a population of healthy small-breed dogs, aged from 7 to 14 years old (49). In the same study, myeloperoxidase levels were higher in healthy dogs than in dogs with a cardiac disease, which contrasts with the trends observed in humans with cardiovascular diseases (50).
To our knowledge, canine neutrophil activation has never been reported in a standardized group of dogs and with the protocol we used. In the study of Almeida et al. neutrophil activation was determined through superoxide production using the nitroblue tetrazolium reduction test in isolated neutrophils in the presence or absence of phorbol 12-myristate 13-acetate (PMA) (51). Silva et al. estimated neutrophil activation by measurement of the fluorescence in a capillary flow cytometer of isolated neutrophils in the presence or absence of PMA (52). These different protocols do not allow comparison of neutrophil activation between studies.
The small number of dogs (N = 14) included in this study is a limitation, in particular when it comes to establish reference norms according to the recommendations of the American Society of Veterinary Clinical Pathology for de novo reference interval definition (53). The method of 95%-content tolerance intervals at specific confidence levels provide an interesting alternative to conventional reference intervals; they have been widely used in industry and can be determined regardless of the sample size. Thus, they can operate as provisionary working reference norms and may be regularly updated as more animals become available. When a population of < 40 individuals is studied, the American Society of Veterinary Clinical Pathology advises to emphasize on the subjects and to adhere to standardized collection techniques and well-controlled methods of analysis (53). This study purposed to determine the OS status in healthy Beagle dogs and to establish working interindividual physiological margins of OS biomarkers. Consequently, we selected a standard and homogeneous group of dogs to minimize the variation between animals and adhered to a methodology identical to human studies. Our results allow comparison with a similar population of healthy adult Beagle dogs but they preclude extrapolation to growing dogs or older dogs, as well as dogs with specific condition such as pregnant bitches, athletic dogs or specific breeds (such as brachycephalic dogs, Greyhounds).
The 95%-content physiological intervals we proposed for plasma concentrations in vitamin C, vitamin E, zinc, copper, copper/zinc ratio, glutathione, myeloperoxidase and isoprostanes have been defined in a standardized group of healthy Beagle dogs using highly specialized techniques of human medicine research. They represent provisional inter-individual physiological margins for future studies involving OS in dogs. Sex-related differences observed for zinc, copper/zinc ratio, and plasma isoprostanes should be interpreted with caution.
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
The animal study was reviewed and approved by Commission d'Ethique Animale de l'Université de Liège.
Conceptualization: AH, JP, SN, and MP. Investigation: MP, JP, J-PC-B, SN, and CL. Writing—original draft preparation: MP. Writing—review and editing: JP, AA, SN, and AH. Funding acquisition: AH and SN. All the authors have read and agreed to the published version of the manuscript.
This research was supported by the Fonds Spéciaux pour la Recherche Facultaires (reference 3328, October 2019).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Jones DP. Redefining oxidative stress. Antioxid Redox Signal. (2006) 8:1865–79. doi: 10.1089/ars.2006.8.1865
2. Di Meo S, Reed TT, Venditti P, Victor VM. Role of ROS and RNS sources in physiological and pathological conditions. Oxid Med Cell Longev. (2016) 2016:1–44. doi: 10.1155/2016/1245049
3. Egea J, Fabregat I, Frapart YM, Ghezzi P, Görlach A, Kietzmann T, et al. European contribution to the study of ROS: a summary of the findings and prospects for the future from the COST action BM1203 (EU-ROS). Redox Biol. (2017) 13:94–162. doi: 10.1016/j.redox.2017.05.007
4. Vertuani S, Angusti A, Manfredini S. The antioxidants and pro-antioxidants network: an overview. Curr Pharm Des. (2004) 10:1677–94. doi: 10.2174/1381612043384655
5. Sánchez-Rodríguez MA, Mendoza-Núñez VM. Oxidative stress indexes for diagnosis of health or disease in humans. Oxid Med Cell Longe. (2019) 2019:8152. doi: 10.1155/2019/4128152
6. Pincemail J, Le Goff C, Charlier C, Gillion P, Cheramy-Bien JP, Van Honaker E, et al. Evaluation biologique du stress oxydant. Application en routine clinique. Nutr Endocrinol. (2009) 5:16–31. doi: 10.2268/40285
7. Freeman LM, Rush JE, Milbury PE, Blumberg JB. Antioxidant status and biomarkers of oxidative stress in dogs with congestive heart failure. J Vet Intern Med. (2005) 19:537–41. doi: 10.1111/j.1939-1676.2005.tb02724.x
8. McMichael MA, Ruaux CG, Baltzer WI, Kerwin SC, Hosgood GL, Steiner JM, et al. Concentrations of 15F 2t isoprostane in urine of dogs with intervertebral disk disease. Am J Vet Res. (2006) 67:1226–31. doi: 10.2460/ajvr.67.7.1226
9. Viviano KR, VanderWielen B. Effect of N-acetylcysteine supplementation on intracellular glutathione, urine isoprostanes, clinical score, and survival in hospitalized ill dogs. J Vet Intern Med. (2013) 27:250–8. doi: 10.1111/jvim.12048
10. Machado VS, Crivellenti LZ, Bottari NB, Tonin AA, Pelinson LP, Borin-Crivellenti S, et al. Oxidative stress and inflammatory response biomarkers in dogs with mammary carcinoma. Pathol - Res Pract. (2015) 211:677–81. doi: 10.1016/j.prp.2015.06.011
11. Marquis A, Packer RA, Borgens RB, Duerstock BS. Increase in oxidative stress biomarkers in dogs with ascending–descending myelomalacia following spinal cord injury. J Neurol Sci. (2015) 353:63–9. doi: 10.1016/j.jns.2015.04.003
12. Kendall A, Woolcock A, Brooks A, Moore GE. Glutathione peroxidase activity, plasma total antioxidant capacity, and urinary F2-isoprostanes as markers of oxidative stress in anemic dogs. J Vet Intern Med. (2017) 31:1700–7. doi: 10.1111/jvim.14847
13. Rubio CP, Martínez-Subiela S, Hernández-Ruiz J, Tvarijonaviciute A, Cerón JJ, Allenspach K. Serum biomarkers of oxidative stress in dogs with idiopathic inflammatory bowel disease. Vet J. (2017) 221:56–61. doi: 10.1016/j.tvjl.2017.02.003
14. Almela RM, Rubio CP, Cerón JJ, Ansón A, Tichy A, Mayer U. Selected serum oxidative stress biomarkers in dogs with non-food-induced and food-induced atopic dermatitis. Vet Dermatol. (2018) 29:229–e82. doi: 10.1111/vde.12525
15. Hagen DM, Ekena JL, Geesaman BM, Viviano KR. Antioxidant supplementation during illness in dogs: effect on oxidative stress and outcome, an exploratory study. J Small Anim Pract. (2019) 60:543–50. doi: 10.1111/jsap.13050
16. Kapun AP, Salobir J, Levart A, Kotnik T, Svete AN. Oxidative stress markers in canine atopic dermatitis. Res Vet Sci. (2012) 92:469–70. doi: 10.1016/j.rvsc.2011.04.014
17. Tomsič K, Seliškar A, Lukanc B, Nemec Svete A. Plasma total antioxidant capacity and activities of blood glutathione peroxidase and superoxide dismutase determined in healthy dogs by using commercially available kits. Acta Vet. (2016) 66:534–48. doi: 10.1515/acve-2016-0046
18. Stowe HD, Lawler DF, Kealy RD. Antioxidant status of pair-fed labrador retrievers is affected by diet restriction and aging1,2. J Nutr. (2006) 136:1844–8. doi: 10.1093/jn/136.7.1844
19. Pillai SR, Steiss JE, Traber MG, Kayden HJ, Wright JC. Comparison of four erythrocyte fragility tests as indicators of vitamin E status in adult dogs. J Comp Pathol. (1992) 107:399–410. doi: 10.1016/0021-9975(92)90014-L
20. Rubio CP, Tvarijonaviciute A, Caldin M, Hernández-Ruiz J, Cerón JJ, Martínez-Subiela S, et al. Stability of biomarkers of oxidative stress in canine serum. Res Vet Sci. (2018) 121:85–93. doi: 10.1016/j.rvsc.2018.09.007
21. Hesta M, Ottermans C, Krammer-Lukas S, Zentek J, Hellweg P, Buyse J, et al. The effect of vitamin C supplementation in healthy dogs on antioxidative capacity and immune parameters. J Anim Physiol Anim Nutr. (2009) 93:26–34. doi: 10.1111/j.1439-0396.2007.00774.x
22. Vajdovich P. Changes in some red blood cell and clinical laboratory parameters in young and old beagle dogs. Vet Res Commun. (1997) 21:463–70. doi: 10.1023/a:1005929801735
23. Todorova I, Simeonova G, Kyuchukova D, Dinev D, Gadjeva V. Reference values of oxidative stress parameters (MDA, SOD, CAT) in dogs and cats. Comp Clin Pathol. (2005) 13:190–4. doi: 10.1007/s00580-005-0547-5
24. Viviano KR, Lavergne SN, Goodman L, VanderWielen B, Grundahl L, Padilla M, et al. Glutathione, cysteine, and ascorbate concentrations in clinically ill dogs and cats. J Vet Intern Med. (2009) 23:250–7. doi: 10.1111/j.1939-1676.2008.0238.x
25. Omaye ST, Turnbull JD, Sauberlich HE. Selected methods for the determination of ascorbic acid in animal cells, tissues, and fluids. Methods Enzymol. (1979) 62:3–11. doi: 10.1016/0076-6879(79)62181-x
26. Pincemail J, Dubrowski T, Durand T, Chiap P, Charlier C, Donneau AF, et al. Validated routine-ready UHPLC/MS-MS method for the reference range determination in human plasma of 15-f2t-isoprostane, biomarker of the oxidative stress. Free Radic Biol Med. (2017) 108:S42. doi: 10.1016/j.freeradbiomed.2017.04.158
27. Stürup S, Hayes RB, Peters U. Development and application of a simple routine method for the determination of selenium in serum by octopole reaction system ICPMS. Anal Bioanal Chem. (2005) 381:686–94. doi: 10.1007/s00216-004-2946-x
28. Baptista G, Dupuy A-M, Jaussent A, Durant R, Ventura E, Sauguet P, et al. Low-grade chronic inflammation and superoxide anion production by NADPH oxidase are the main determinants of physical frailty in older adults. Free Radic Res. (2012) 46:1108–14. doi: 10.3109/10715762.2012.692784
29. Guttman I. Statistical Tolerance Regions: Classical and Bayesian. London: Griffin (1970). 150 p.
30. Eisenhart C, Hastay MW, Wallis WA. Selected Techniques of Statistical Analysis for Scientific and Industrial Research and Production and Management Engineering. New York: McGraw-Hill (1947). 473 p.
31. Klaassen JK. Reference values in veterinary medicine. Lab Med. (1999) 30:194–7. doi: 10.1093/labmed/30.3.194
32. Alexander JE, Colyer A, Haydock RM, Hayek MG, Park J. Understanding how dogs age: longitudinal analysis of markers of inflammation, immune function, and oxidative stress. J Gerontol Ser A. (2018) 73:720–8. doi: 10.1093/gerona/glx182
33. Verk B, Nemec Svete A, Salobir J, Rezar V, Domanjko Petrič A. Markers of oxidative stress in dogs with heart failure. J Vet Diagn Invest. (2017) 29:636–44. doi: 10.1177/1040638717711995
34. JuodŽente D, Karveliene B, Riškevičiene V. The influence of the duration of the preoperative time spent in the veterinary clinic without the owner on the psychogenic and oxidative stress in dogs. J Vet Med Sci. (2018) 80:1129–33. doi: 10.1292/jvms.18-0066
35. Barry-Heffernan C, Ekena J, Dowling S, Pinkerton ME, Viviano K. Biomarkers of oxidative stress as an assessment of the redox status of the liver in dogs. J Vet Intern Med. (2019) 33:611–7. doi: 10.1111/jvim.15443
36. Michos C, Kalfakakou V, Karkabounas S, Kiortsis D, Evangelou A. Changes in copper and zinc plasma concentrations during the normal menstrual cycle in women. Gynecol Endocrinol Off J Int Soc Gynecol Endocrinol. (2010) 26:250–5. doi: 10.3109/09513590903247857
37. Dimri U, Ranjan R, Kumar N, Sharma MC, Swarup D, Sharma B, et al. Changes in oxidative stress indices, zinc and copper concentrations in blood in canine demodicosis. Vet Parasitol. (2008) 154:98–102. doi: 10.1016/j.vetpar.2008.03.001
38. Panda D, Patra RC, Nandi S, Swarup D. Oxidative stress indices in gastroenteritis in dogs with canine parvoviral infection. Res Vet Sci. (2009) 86:36–42. doi: 10.1016/j.rvsc.2008.05.008
39. Mezzetti A, Pierdomenico SD, Costantini F, Romano F, De Cesare D, Cuccurullo F, et al. Copper/zinc ratio and systemic oxidant load: effect of aging and aging-related degenerative diseases. Free Radic Biol Med. (1998) 25:676–81. doi: 10.1016/S0891-5849(98)00109-9
40. Hinchcliff KW, Reinhart GA, DiSilvestro R, Reynolds A, Blostein-Fujii A, Swenson RA. Oxidant stress in sled dogs subjected to repetitive endurance exercise. Am J Vet Res. (2000) 61:512–7. doi: 10.2460/ajvr.2000.61.512
41. Putman AK, Contreras GA, Sordillo LM. Isoprostanes in veterinary medicine: beyond a biomarker. Antioxidants. (2021) 10:145–60. doi: 10.3390/antiox10020145
42. Guo CH, Chen PC, Yeh MS, Hsiung DY, Wang CL. Cu/Zn ratios are associated with nutritional status, oxidative stress, inflammation, and immune abnormalities in patients on peritoneal dialysis. Clin Biochem. (2011) 44:275–80. doi: 10.1016/j.clinbiochem.2010.12.017
43. Ozturk P, Belge Kurutas E, Ataseven A. Copper/zinc and copper/selenium ratios, and oxidative stress as biochemical markers in recurrent aphthous stomatitis. J Trace Elem Med Biol. (2013) 27:312–6. doi: 10.1016/j.jtemb.2013.04.002
44. Lykkesfeldt J, Michels AJ, Frei B. Vitamin C1. Adv Nutr. (2014) 5:16–8. doi: 10.3945/an.113.005157
45. Jiang Q. Natural forms of vitamin E: metabolism, antioxidant and anti-inflammatory activities and the role in disease prevention and therapy. Free Radic Biol Med. (2014) 72:76–90. doi: 10.1016/j.freeradbiomed.2014.03.035
46. Vatassary GT, Eckfeldt, J. H Vitamin E concentrations plasma and platelets. Am J Clin Nutr. (1983) 37:1020–4. doi: 10.1093/ajcn/37.6.1020
47. Samiec PS, Drews-Botsch C, Flagg EW, Kurtz JC, Sternberg P, Reed RL, et al. Glutathione in human plasma: decline in association with aging, age-related macular degeneration, and diabetes. Free Radic Biol Med. (1998) 24:699–704. doi: 10.1016/S0891-5849(97)00286-4
48. Jones DP. “[11] Redox potential of GSH/GSSG couple: Assay and biological significance.” In:Sies H, Packer L, , editors. Methods in Enzymology. Protein Sensors and Reactive Oxygen Species - Part B: Thiol Enzymes and Proteins. Academic Press (2002). p. 93–112 doi: 10.1016/S0076-6879(02)48630-2
49. Park JI, Suh SI, Hyun C. Evaluation of serum myeloperoxidase concentration in dogs with heart failure due to chronic mitral valvular insufficiency. Can J Vet Res. (2017) 81:37–40.
50. Nussbaum C, Klinke A, Adam M, Baldus S, Sperandio M. Myeloperoxidase: a leukocyte-derived protagonist of inflammation and cardiovascular disease. Antioxid Redox Signal. (2013) 18:692–713. doi: 10.1089/ars.2012.4783
51. Almeida BFM, Narciso LG, Bosco AM, Pereira PP, Braga ET, Avanço SV, et al. Neutrophil dysfunction varies with the stage of canine visceral leishmaniosis. Vet Parasitol. (2013) 196:6–12. doi: 10.1016/j.vetpar.2013.02.016
52. Silva ACRA, de Almeida BFM, Soeiro CS, Ferreira WL, de Lima VMF, Ciarlini PC. Oxidative stress, superoxide production, and apoptosis of neutrophils in dogs with chronic kidney disease. Can J Vet Res. (2013) 77:136–41.
53. Friedrichs KR, Harr KE, Freeman KP, Szladovits B, Walton RM, Barnhart KF, et al. American society for veterinary clinical pathology reference interval guidelines: determination of de novo reference intervals in veterinary species and other related topics. Vet Clin Pathol. (2012) 41:441–53. doi: 10.1111/vcp.12006
Keywords: dog, oxidative stress, isoprostanes, biomarkers, Beagle
Citation: Porato M, Noël S, Pincemail J, Albert A, Cheramy-Bien J-P, Le Goff C and Hamaide A (2023) Selected biomarkers of oxidative stress in healthy Beagle dogs: A preliminary study. Front. Vet. Sci. 10:1063216. doi: 10.3389/fvets.2023.1063216
Received: 06 October 2022; Accepted: 09 March 2023;
Published: 24 March 2023.
Edited by:
Fazul Nabi, Lasbela University of Agriculture, Water and Marine Sciences, PakistanReviewed by:
Josiane Cillard, University of Rennes 1, FranceCopyright © 2023 Porato, Noël, Pincemail, Albert, Cheramy-Bien, Le Goff and Hamaide. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mathilde Porato, bXBvcmF0b0B1bGllZ2UuYmU=
†These authors share first authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.