
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Vet. Sci., 28 October 2022
Sec. Veterinary Infectious Diseases
Volume 9 - 2022 | https://doi.org/10.3389/fvets.2022.981207
This article is part of the Research TopicWomen in Veterinary Infectious Diseases: 2022View all 6 articles
 Joana Castro1*
Joana Castro1* Maria Margarida Barros1
Maria Margarida Barros1 Daniela Araújo1
Daniela Araújo1 Ana Maria Campos1
Ana Maria Campos1 Ricardo Oliveira1,2,3
Ricardo Oliveira1,2,3 Sónia Silva1,4,5
Sónia Silva1,4,5 Carina Almeida1,2,3,4,5*
Carina Almeida1,2,3,4,5*Enteric colibacillosis is a common disease in nursing and weanling pigs. It is caused by the colonization of the small intestine by enterotoxigenic strains of Escherichia coli (ETEC) that make use of specific fimbria or pili to adhere to the absorptive epithelial cells of the jejunum and ileum. Once attached, and when both the immunological systems and the gut microbiota are poorly developed, ETEC produce one or more enterotoxins that can have local and, further on, systemic effects. These enterotoxins cause fluid and electrolytes to be secreted into the intestinal lumen of animals, which results in diarrhea, dehydration, and acidosis. From the diversity of control strategies, antibiotics and zinc oxide are the ones that have contributed more significantly to mitigating post-weaning diarrhea (PWD) economic losses. However, concerns about antibiotic resistance determined the restriction on the use of critically important antimicrobials in food-producing animals and the prohibition of their use as growth promoters. As such, it is important now to begin the transition from these preventive/control measures to other, more sustainable, approaches. This review provides a quick synopsis of the currently approved and available therapies for PWD treatment while presenting an overview of novel antimicrobial strategies that are being explored for the control and treatment of this infection, including, prebiotics, probiotics, synbiotics, organic acids, bacteriophages, spray-dried plasma, antibodies, phytogenic substances, antisense oligonucleotides, and aptamers.
Post-weaning period is a critical phase in the pig's life because the immune system is immature, and the sow milk removal, and consequent interruption of nutritive intake of immunoglobulin present in the milk, increases pigs' susceptibility to microbial infections, especially in the current lines of hyperprolific sows (1). As such, piglets are highly susceptible to Post-Weaning Diarrhea (PWD), a disorder that often affects pigs during the first 2 weeks of weaning and is characterized by sudden death or diarrhea and growth retardation in surviving piglets (2). PWD is one of the most serious threats to the swine industry worldwide, with episodes reaching mortality rates of 20 to 30% (2) and is associated with the proliferation of enterotoxigenic Escherichia coli (ETEC) in the pig intestine (3).
ETEC are gram-negative, flagellated bacilli and most pathogenic strains form smooth to mucoid colonies. Virulence factors include fimbriae, enterotoxins, endotoxins, and capsules. Fimbriae are the small hair-like structures on the bacterial surface that allow attachment to specific receptors on the surface of mucosal enterocytes of the small intestine, being crucial in the colonization process. Pathogenic strains also produce one or more enterotoxins, which are exotoxins elaborated locally in the small intestine that can have either local or systemic effects (1). There are 5 common, antigenically different fimbriae types found in pigs: F4 (K88), F5 (K99), F41, F6 (987P), and F18. The first 4 fimbriae types are responsible for mediating adhesion in neonates, while F18 is not associated with neonatal colibacillosis but is common in postweaning colibacillosis as is F4. It is also important to note that hemolysis is a common trait for pathogenic F4 and F18 isolates. Furthermore, the virulence of ETEC can also be characterized by the production of heat-labile toxins (LT), heat-stable toxin A (StA), heat-stable toxin B (StB), and verotoxin (shiga-like toxin, STX). The first three act locally interfering with electrolytes fluid increasing the fluid secretion to the lumen leading to diarrhea while verotoxin is responsible for the systemic vascular effects of edema disease (4).
As an attempt to promote health and growth performance, different approaches have been used to prevent PWD, including mostly dietary supplementation, such as prebiotics and probiotics; genetic breeding for ETEC-resistant herds; administration of growth promotors; and vaccines (5, 6). It is important to highlight that growth promoters and zinc oxide (7), have been (or are being) banned from animal husbandry. Also, antibiotics use is now more restricted because of antimicrobial resistance observed in ETEC and potential consequences for human health (1). These restrictions are strongly affecting the control of PWD (8). This makes room for innovative, antibiotic-free, intervention/control strategies that can control ETEC infections.
Antibiotic therapy is required in many cases of enteric colibacillosis, being the main antimicrobials used for the treatment of enteric colibacillosis listed in Supplementary Table 1. According to Luppi, the choice of antibiotics for the treatment of PWD must consider several aspects (1): (i) the local of infection that must be located mainly in the small intestine; (ii) the empiric treatments based on the knowledge of the individual herd and local data on the resistance pattern; and (iii) the evaluation of the isolated strain's antimicrobial susceptibility. However, as an outbreak of colibacillosis frequently requires quick actions, the use of antibiotics almost always precedes the results of the resistance pattern (1, 9).
Also, it has been established that the antibiotic should be administered to all animals exhibiting signs referable to colibacillosis and sick pigs must be treated parenterally since they eat and drink very little. In practice when mortality occurs, a metaphylactic approach is applied wherein all animals in the pens are treated (1, 9–11). Of note that guidelines for the prudent use of antimicrobials in veterinary medicine (2015/C/ 299/04) published in the official journal of the European Union considered the use of metaphylaxis and stated that antimicrobial metaphylaxis should be prescribed only when there is a real need for treatment and that the veterinarian should justify and document the treatment on the basis of clinical findings on the development of a disease in herd or flock (10).
As described above, alternatives to antibiotic therapy are some preventive approaches that have been applied to avoid the development of colibacillosis illness. Those therapies are essentially based on genetic breeding, vaccines, and zinc oxide administration, as illustrated in Figure 1.
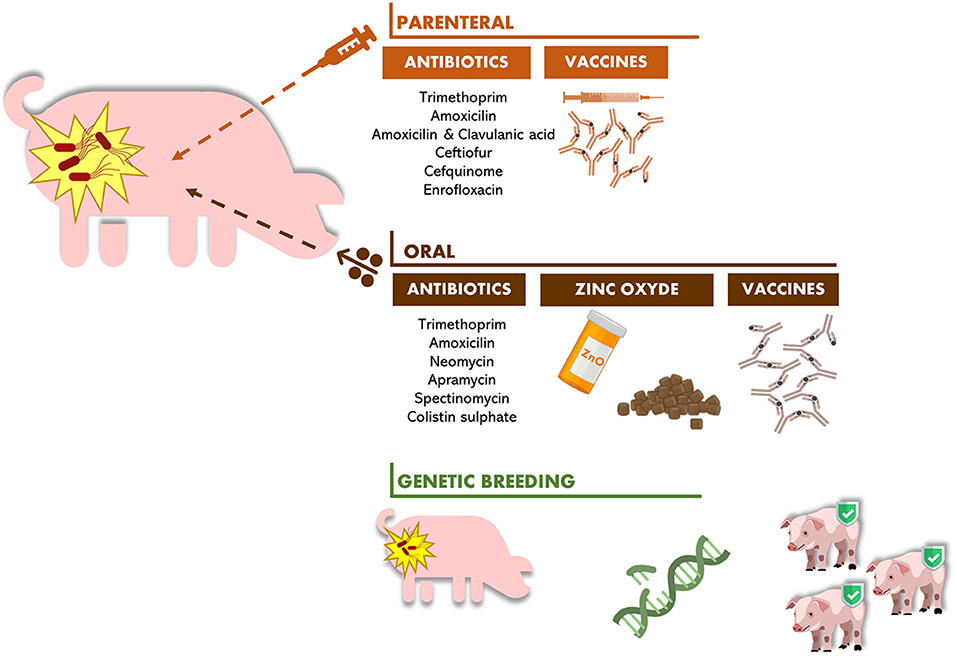
Figure 1. Current therapies used to control PWD in piglets.
The advances in molecular approaches sparked the interest of the scientific community in obtaining genetic breeding for ETEC-resistant herds. As mentioned above, the pig population presents specific receptors on the intestinal epithelial cells for the F4 and F18 fimbriae, which are the primary virulence factors carried by E. coli responsible for PWD. However, certain pigs do not have receptors for F4 and F18 adhesins and are thus resistant to infection by these fimbriae. Therefore, an attractive approach to preventing PWD is increasing the presence of both the F18 and F4 resistance loci in the pig population through breeding (12). Despite all the efforts, there are more insights about F18 resistance loci than about F4 resistance loci. In this sense, it was employed a molecular test for the large-scale selection of resistant pigs for breeding, using the polymerase chain reaction-restriction fragment length polymorphism (PCR-RFLP) methodology. In brief, using this approach, we can detect polymorphisms in the FUT1 gene in a piglet, that encodes an α-(1,2) fucosyltransferase protein. It is important to highlight that FUT1 polymorphisms are responsible for controlling the expression of the receptor for F18 fimbriated E. coli in the intestinal mucosa. Regarding the F4 resistance loci, by using PCR-RFLP approach (13), it is also possible to detect polymorphisms in the MUC4 gene in a piglet, that encodes for a membrane-bound-O-glycoprotein that is extensively expressed on the surface of gastrointestinal epithelial cells in which protects and lubricates the epithelial surfaces (14, 15). As such, using this approach, it was possible to select the piglets that do not present these receptors for the E. coli F18 and F4 adhesion (12, 16, 17).
It is known that newborn pigs can establish immune reactions against mucosal antigens (18, 19). However, the newly weaned pigs need active intestinal mucosal immunization due to the lack of passive lactogenic immunity. Therefore, these vaccines can induce this protective effect by activating the mucosal immune system and antigen-specific immunoglobulins (A and M) responses (20, 21). Thus, 3 main types of vaccines have been used on pigs. Firstly, intramuscular injectable vaccines stimulate systemic immunity by increasing circulating antibodies to keep intestinal bacteria levels low enough to be non-pathogenic (21). Secondly, an oral administration of live attenuated or live wild-type non-enterotoxigenic E. coli strains with fimbrial adhesins can stimulate intestinal colonization by this E. coli by inducing the secretion of intestinal antibodies, and finally blocks the adherence of ETEC (22). For example, a single dose of a commercial vaccine (Coliprotec® F4) showed a significant reduction in the incidence of diarrhea, ileum colonization by F4-ETEC, and fecal shedding of F4-ETEC after the heterologous challenge at 7 and 21 days post-vaccination (23). Lastly, the oral administration of purified fimbria results in a specific mucosal immune response in the intestines and may cause a significant decrease in fecal excretion of the pathogenic E. coli (21, 24). Supplementary Table 2 shows the current vaccines available in the market.
Pig producers have been successful with weaning at an early age with limited signs of gastrointestinal disease shortly after weaning (25, 26). In fact, it has been described that feed containing between 2,400 and 3,000 ppm of zinc reduces diarrhea and mortality, and improves the growth of piglets. However, in the last decade, several studies have centered attention on heavy metals used in animal farming and possible mechanisms that could promote the spread of antibiotic resistance via co-selection (1). Furthermore, the use of zinc oxide possesses an environmental hazard, as most of the zinc is excreted with the feces and through manuring, it accumulates in soil (27).
Recently, the Committee for Medicinal Products for Veterinary Use (CVMP) of the European Medicines Agency set the referral procedure for veterinary medicinal products containing zinc oxide to be administered orally to food-producing species. The Committee adopted an opinion by consensus determining that the benefits of zinc oxide for the prevention of diarrhea in pigs do not outweigh the risks to the environment. The CVMP emphasized that there is a risk of co-selection for resistance related to the use of zinc oxide, but at the present time, that risk is not quantifiable. Accordingly, medicinal zinc oxide has been prohibited in the European Union effectively coming into force no later than June 2022 (28, 29). Consequently, there is an urgent need for adopting novel strategies for the control of PWD.
The increasing evidence that the current approaches to treat/prevent PWD raise problems associated with antimicrobial resistance or the risks to the environment sparked the interest of the scientific community in exploring new eco-friendly prophylactic or antimicrobial tools. Thus, in recent years, studies of anti-enteric colibacillosis agents started to include prebiotics, probiotics, synbiotics, organic acids, phytogenic substances, bacteriophages, spray-dried plasma, antibodies, antisense oligonucleotides, and aptamers (see Table 1).

Table 1. Benefits and limitations of the major alternative agents for the control of post-weaning diarrhea (PWD) based on studies published since 2010.
Maintaining a healthy gut is crucial for a pig to digest and absorb dietary nutrients efficiently (76). As such, prebiotics, which are non-digestible feed ingredients positively affecting the host by stimulating the growth and/or the activity of a limited number of bacteria in the gastrointestinal microbiota (5), can be used to modulate the gut microbiota. Probiotics are defined as viable microorganisms that when ingested in sufficient amounts reach the intestine in an active state where they can exert positive effects on the host's health status (77, 78). In last, synbiotics are defined as a mixture comprising live microorganisms (probiotics) and substrate(s) (prebiotics) selectively utilized by host microorganisms that confer a health benefit on the host (79). According to Girard and colleagues, this combination improves the survival rate and favors the growth and activity of beneficial microorganisms in the gut of the piglets (51). Among the prebiotics and probiotics evaluated for the control of ETEC colonization in pigs, inulin and Lactobacillus plantarum are the ones that have been more systematically assessed, either individually or in combination (see Table 1). Overall, the results of the different studies indicate that these strategies can reduce ETEC colonization and relieve some symptoms. However, a few studies also have found no significant differences between the tested probiotics and the respective controls (41). This is not surprising as the effect of these approaches is most probably highly dependent on the strains, concentrations applied, animal age, and the strategies used for the delivery in the gut. Further studies are necessary to systematically evaluate the real potential of those approaches for controlling ETEC.
Organic acids (OA) are considered powerful antimicrobials, which are extensively used as feed additives in pig nutrition, in particular the short-chain fatty acids (SCFAs) and medium-chain fatty acids (MCFAs) (80). Because of their antimicrobial effects, OA are able to prevent the colonization and proliferation of pathogenic bacteria (52) and their antimicrobial effects are related to their action on cellular metabolism and their ability to deplete cellular energy. Normally, a reduction in pH of the diet can happen because of the dissociation of OA, which is favorable for the growth of lactic acid bacteria (55). In contrast, the non-dissociated OA can act as an antimicrobial agent, since it can penetrate the cell wall of bacteria and interfere with normal physiology (81). In the pigs with a diet containing OA, gastrointestinal health is improved, reducing the incidence to develop diarrhea as described in Table 1.
Plant-derived compounds are also studied as alternative growth promoters in pigs, as is the case of phytogenic feed additives (82–84). These compounds are added to the pigs' diet, due to their positive effects on animal growth and health, since they present antibacterial, anti-inflammatory, and antioxidant properties (59). Examples of substances used as phytogenic feed additives include essential oils, herbs, and spices (see Table 1). Recent studies reported the positive results of these compounds in pig health, such as growth performance and nutrient digestibility (82). Moreover, to improve its performance, the combination of different phytogenic substances has been explored in order to study the synergistic or complementary activity on pig health.
Bacteriophages are non-hazardous self-replicating agents that increase their numbers as they destroy specific target bacteria (85). It is noteworthy, that groundbreaking work in bacteriophage therapy addressing colibacillosis in swine was performed in the 1980s by Smith and Huggings (86–88). After 2 decades, efforts conducted by Jamalludeen and colleagues showed that the 6 tested bacteriophages exhibited prophylactic activity against diarrhea and shedding of the challenge ETEC following experimental oral infection of pigs (89). More recently, some studies on dietary supplementation of bacteriophages for E. coli have been conducted aiming to improve the performance, feed efficiency, and fecal microbiota in pigs as mentioned in Table 1.
Spray-dried plasma (SDP) is a protein-rich product from the industrial fractionation of blood from healthy animals (90). To obtain SDP, blood is collected with an anticoagulant and centrifuged to separate the blood cells. Plasma is then concentrated and spray-dried under high pressure to achieve a minimum of 80°C throughout its substance. Of note that through this procedure, proteins preserve most of their biological activity (91). Multiple modes of action of SDP have been described, including either directly influencing the immune inflammatory response locally or systemically, and/or through the indirect modification of beneficial microbial populations (90). As such, in the past decades, several studies have been conducted to assess the role of SDP in swine production and calf milk replacers to improve performance, feed efficiency, and swine health (92, 93). Furthermore, some evidence suggests that SDP supplementation can modify the composition of the intestinal microbiota (92). More recently, few researchers have also conducted experiments in this field, as depicted in Table 1.
Chicken IgY, known as the immunoglobulin Y (IgY), is an antigen-specific antibody produced by B lymphocytes and accumulated in the yolk of chicken eggs (94). Due to its high specificity, this approach has attracted considerable attention for the treatment of gastrointestinal infections. Importantly, Wang and colleagues proved that IgY was able to significantly inhibit the growth of E. coli K88 by blocking the binding of E. coli to small intestinal mucus (73). Nevertheless, the activity of orally administered IgY can reduce rapidly under gastric conditions. Hence several nanotechnology applications, such as chitosan-alginate microcapsules, methacrylic acid copolymers, liposomes, polymeric microspheres multiple emulsification and more recently hydrogel-carbon nanotubes composites, have been used to protect IgY from hydrolysis by gastric enzymes (72), as described in Table 1.
Lactoferrin (LF), an iron-binding glycoprotein, is found in the colostrum, milk, and other secretions of many mammalian species. Lactoferrin is indicated to have several physiologic functions, including regulation of iron absorption, and it has an important role in host defense, including broad-spectrum antibacterial effects (74, 95). Recently, efforts have been conducted to assess if lactoferrin can be used to control PWD, as shown in Table 1.
An alternative antibacterial strategy to treat colibacillosis is the development of antisense oligonucleotides therapy against E. coli (75, 96–98). Antisense oligonucleotides (ASOs) are short, single-stranded nucleic acid sequences that are complementary to a target mRNA. They can down-regulate gene expression by binding to their target mRNA and inhibiting its translation through the creation of a steric block to ribosome binding and/or by facilitating RNase H recruitment and RNA cleavage. ASOs have been proposed to enter into cells through high-and low-binding plasma protein receptors on the cell surface, resulting in ASO compartmentalization into lysosomes and endosomes. Through a largely unknown mechanism, ASOs are released from the vesicles into the cytoplasm where they can freely move in and out of the nucleus. Upon entry into the nucleus, ASOs can bind directly to mRNA structures and prevent the formation of the 5'-mRNA cap, modulate alternative splicing, and recruit RNaseH to induce cleavage. ASOs in the cytoplasm can also bind directly to the target mRNA and sterically block the ribosomal subunits from attaching and/or running along the mRNA transcript during translation. ASOs can also be designed to directly bind to microRNA (miRNA) sequences and natural antisense transcripts (NATs), thereby prohibiting miRNAs and NATs from inhibiting their own specific mRNA targets (99). Typically, ASOs are synthesized from nucleotide analogs in order to enhance the affinity for RNA and decrease the susceptibility to cellular nucleases. A key advantage of the antisense approach is that it should be possible to rationally design an antisense oligonucleotide to target any mRNA. If the target mRNA encodes an essential protein, then the ASOs may have antibacterial properties. A number of ASOs, that target a variety of mRNAs E. coli genes (Supplementary Table 3), have been reported to have antibacterial activity in vitro and provide a proof of concept for further exploitation to test in vivo to control colibacillosis.
Aptamers are quickly gaining significance as alternative biorecognition molecules to antibodies in targeted therapy (100). They are able to bind (i.e., diagnosis tool) and inhibit the function (i.e., therapeutic tool) of other biomolecules such as virulence factors normally associated with ETEC (i.e., toxins and fimbriae), as well as being used as carriers for other biochemical compounds (i.e., drug-delivery tool) (101). Sometimes referred to as “synthetic antibodies”, aptamers are typically single-stranded DNA or RNA oligonucleotides with defined secondary motifs (e.g., loop, stem, or G-quadruplex) that confer them complex three-dimensional structures and the ability to recognize and bind targets with exceptional binding affinity and specificity to their target (102). The aptamer-target interaction results mainly from the compatibility of physical structures and/or the stacking of chemical groups that are stabilized by hydrogen bonds, electrostatic interactions, hydrophobic effects, π-π stacking, Van der Waals forces, or combinations of these different forces (103). As a diagnostic or drug-delivery tool, aptamers can be chemically linked to other compounds (e.g., fluorophores or drugs) that will have a direct action on the target when the binding complexes are formed. As a therapeutic tool, the physicochemical binding can block receptor-binding domains of the target from interacting with receptors and, thus, neutralize their action by blocking vital proteins on the cell membrane and/or cytoplasmic proteins such as virulence factors normally associated with ETEC (i.e., toxins and fimbriae) (104). Aptamers are isolated by a repetitive interactive selection procedure known as Systematic Evolution of Ligands by EXponential enrichment (SELEX). This methodology follows a standard pattern that comprises a large pool of random sequences subjected to successive steps of (1) binding, (2) partition and (3) amplification to obtain a pool of molecules enriched for those with high affinity to the target (105). Some aptamers have been reported to be specific for E. coli and their virulence factors (Supplementary Table 4) providing a proof of concept for further exploration of specific aptamers for the diagnosis and treatment of colibacillosis. Aptamers for E. coli K88 fimbriae have been reported (106) and are of particular relevance as this is a high prevalent ETEC fimbria-type that allows adhesion to the animal gut cells. However, until now the potential of aptamers to block fimbriae-mediated adhesion in animal models has not been evaluated.
Swine colibacillosis prevention and control therapies are facing a huge challenge because of the recent policies on the pharmacological dose of zinc oxide and antibiotic use. While these policies are inevitable and necessary to deal with the problem of antimicrobial resistance and environmental sustainability; the need for effective strategies to control infectious disease in pigs' production is a pressing matter. By properly addressing the virulence factors of ETEC, novel strategies will hopefully arise with more eco-friendly approaches to control PWD and more in line with the public health concerns regarding antimicrobial resistance.
JC, MB, DA, AC, RO, and SS prepared the first draft of the manuscript. JC and CA defined the content of the manuscript. All authors critically reviewed and approved the final version of the article.
Research in swine colibacillosis in CA Laboratory was supported by funding from the Fundação para a Ciência e a Tecnologia (FCT) Strategic Project Unit PTDC/CVT-CVT/4620/2021. It was also partially funded by LA/P/0045/2020 (ALiCE), UIDB/00511/2020 and UIDP/00511/2020 (LEPABE), funded by national funds through FCT/MCTES (PIDDAC), by FCT under the scope of the strategic funding of UIDB/04469/2020 unit (CEB), and by LABBELS-Associate Laboratory in Biotechnology, Bioengineering, and Microelectromechnaical Systems, LA/P/0029/2020.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fvets.2022.981207/full#supplementary-material
1. Luppi A. Swine enteric colibacillosis: diagnosis, therapy and antimicrobial resistance. Porc Heal Manag. (2017) 3:1–18. doi: 10.1186/s40813-017-0063-4
2. Rhouma M, Fairbrother JM, Beaudry F, Letellier A. Post weaning diarrhea in pigs: Risk factors and non-colistin-based control strategies. Acta Vet Scand. (2017) 59:1–19. doi: 10.1186/s13028-017-0299-7
3. Bin P, Tang Z, Liu S, Chen S, Xia Y, Liu J, et al. Intestinal microbiota mediates Enterotoxigenic Escherichia coli-induced diarrhea in piglets. BMC Vet Res. (2018) 14:1–13. doi: 10.1186/s12917-018-1704-9
4. Hartadi EB, Effendi MH, Plumeriastuti H, Sofiana ED, Wibisono FM, Hidayatullah AR. A review of enterotoxigenic Escherichia coli infection in piglets: public health importance. Syst Rev Pharm. (2020) 11:687–98. doi: 10.31838/srp.2020.9.100
5. Dubreuil JD. Enterotoxigenic Escherichia coli and probiotics in swine: what the bleep do we know? Biosci. Microbiota, Food Heal. (2017) 36:75–90. doi: 10.12938/bmfh.16-030
6. Liu Y, Espinosa CD, Abelilla JJ, Casas GA, Lagos LV, Lee SA, et al. Non-antibiotic feed additives in diets for pigs: a review. Anim Nutr. (2018) 4:113–25. doi: 10.1016/j.aninu.2018.01.007
7. D?bski B. Supplementation of pigs diet with zinc and copper as alternative to conventional antimicrobials. Pol J Vet Sci. (2016) 19:917–24. doi: 10.1515/pjvs-2016-0113
8. Bonetti A, Tugnoli B, Piva A, Grilli E. Towards Zero Zinc Oxide : feeding strategies to manage post-weaning diarrhea in piglets. Animals. (2021) 1–24. doi: 10.3390/ani11030642
9. Burch DGS. Antimicrobial drug use in swine. In: Antimicrobial Therapy in Veterinary Medicine. 5th, ed. New Jersey, NJ: John Wiley & Sons, Ltd (2013). p. 553–68. doi: 10.1002/9781118675014.CH33
10. Burch DGS, Oliver DC, Aarestrup FM. COMMISSION NOTICE. Guidelines for the prudent use of antimicrobials in veterinary medicine (2015/C 299/04). Off J Eur Union. (2009).
11. Fairbrother JM, Nadeau É. Colibacillosis. In Zimmerman JJ, Karriker LA, Ramirez A, Schwartz KJ, Stevenson GW, Zhang J, editors. Diseases of Swine. (2019). p. 807–834.
12. Fairbrother JM, Nadeau É, Gyles CL. Escherichia coli in postweaning diarrhea in pigs: an update on bacterial types, pathogenesis, and prevention strategies. Anim Heal Res Rev. (2005) 6:17–39. doi: 10.1079/AHR2005105
13. Goetstouwers T, Van Poucke M, Nguyen VU, Melkebeek V, Coddens A, Deforce D, et al. F4-related mutation and expression analysis of the aminopeptidase N gene in pigs. J Anim Sci. (2014) 92:1866–73. doi: 10.2527/jas.2013-7307
14. Rasschaert K, Verdonck F, Goddeeris BM, Duchateau L, Cox E. Screening of pigs resistant to F4 enterotoxigenic Escherichia coli (ETEC) infection. Vet Microbiol. (2007) 123:249–53. doi: 10.1016/j.vetmic.2007.02.017
15. Fontanesi L, Bertolini F, Dall'Olio S, Buttazzoni L, Gallo M, Russo V. Analysis of Association between the MUC4 g.8227C>G polymorphism and production traits in italian heavy pigs using a selective genotyping approach. Anim. Biotechnol. (2012) 23:147–155. doi: 10.1080/10495398.2011.653462
16. Meijerink E, Neuenschwander S, Fries R, Dinter A, Bertschinger HU, Stranzinger G, et al. A DNA polymorphism influencing α(1,2)fucosyltransferase activity of the pig FUT1 enzyme determines susceptibility of small intestinal epithelium to Escherichia coli F18 adhesion. Immunogenetics. (2000) 52:129–36. doi: 10.1007/s002510000263
17. Frydendahl K, Jensen TK, Andersen JS, Fredholm M, Evans G. Association between the porcine Escherichia coli F18 receptor genotype and phenotype and susceptibility to colonisation and postweaning diarrhoea caused by E. coli O138:F18. Vet Microbiol 93. (2003) 93:39–51. doi: 10.1016/S0378-1135(02)00348-6
18. Rothkötter HJ, Sowa E, Pabst R. The pig as a model of developmental immunology. Hum Exp Toxicol. (2002) 21:533–6. doi: 10.1191/0960327102ht293oa
19. Sun Y, Kim SW. Intestinal challenge with enterotoxigenic Escherichia coli in pigs, and nutritional intervention to prevent postweaning diarrhea. Anim Nutr. (2017) 3:322–30. doi: 10.1016/j.aninu.2017.10.001
20. Bianchi ATJ, Scholten JW, Van Zijderveld AM, Van Zijderveld FG, Bokhout BA. Parenteral vaccination of mice and piglets with F4+ Escherichia coli suppresses the enteric anti-F4 response upon oral infection. Vaccine. (1996) 14:199–206. doi: 10.1016/0264-410X(95)00192-4
21. Van Den Broeck W, Cox E, Goddeeris BM. Induction of immune responses in pigs following oral administration of purified F4 fimbriae. Vaccine. (1999) 17:2020–9. doi: 10.1016/S0264-410X(98)00406-X
22. Van Den Broeck W, Cox E, Goddeeris BM. Receptor-dependent immune responses in pigs after oral immunization with F4 fimbriae. Infect Immun. (1999) 67:520–6. doi: 10.1128/IAI.67.2.520-526.1999
23. Fairbrother JM, Nadeau É, Bélanger L, Tremblay CL, Tremblay D, Brunelle M, et al. Immunogenicity and protective efficacy of a single-dose live non-pathogenic Escherichia coli oral vaccine against F4-positive enterotoxigenic Escherichia coli challenge in pigs. Vaccine. (2017) 35:353–60. doi: 10.1016/j.vaccine.2016.11.045
24. Verdonck F, Cox E, Van Der Stede Y, Goddeeris BM. Oral immunization of piglets with recombinant F4 fimbrial adhesin FaeG monomers induces a mucosal and systemic F4-specific immune response. Vaccine. (2004) 22:4291–9. doi: 10.1016/j.vaccine.2004.04.016
25. Eriksen EØ, Kudirkiene E, Christensen AE, Agerlin MV, Weber NR, Nødtvedt A, et al. Post-weaning diarrhea in pigs weaned without medicinal zinc: risk factors, pathogen dynamics, and association to growth rate. Porc Heal Manag. (2021) 7:1–19. doi: 10.1186/s40813-021-00232-z
26. Nielsen CL, Kongsted H, Tind J, Krogh MA. Antibiotic and medical zinc oxide usage in Danish conventional and welfare-label pig herds in 2016 – 2018. Prev Vet Med. (2021) 189:105283. doi: 10.1016/j.prevetmed.2021.105283
27. Jensen J, Kyvsgaard NC, Battisti A, Baptiste KE. Environmental and public health related risk of veterinary zinc in pig production - using Denmark as an example. Environ Int. (2018) 114:181–90. doi: 10.1016/j.envint.2018.02.007
28. European Medicines Agency (EMA). Committee for Medicinal Products for Veterinary Use (CVMP) meeting of 13-15 June 2017 | European Medicines Agency. Available online at: https://www.ema.europa.eu/en/news/committee-medicinal-products-veterinary-use-cvmp-meeting-13-15-june-2017 (accessed October 14, 2022).
29. EMA and CVMP. EMA—Zinc Oxide—Annex II—Scientific Conclusions and Grounds for the Refusal of the Marketing Authorisation and for Withdrawal of the Existing Marketing Authorisations. (2017). p. 1–47. Available online at: https://www.ema.europa.eu/en/medicines/veterinary/referrals/zinc-oxide (accessed October 14, 2022).
30. Hermes RG, Manzanilla EG, Martín-Orúe SM, Pérez JF, Klasing KC. Influence of dietary ingredients on in vitro inflammatory response of intestinal porcine epithelial cells challenged by an enterotoxigenic Escherichia coli (K88). Comp Immunol Microbiol Infect Dis. (2011) 34:479–88. doi: 10.1016/j.cimid.2011.08.006
31. Sarabia-Sainz HM, Armenta-Ruiz C, Sarabia-Sainz JAI, Guzmán-Partida AM, Ledesma-Osuna AI, Vázquez-Moreno L, et al. Adhesion of enterotoxigenic Escherichia coli strains to neoglycans synthesised with prebiotic galactooligosaccharides. Food Chem. (2013) 141:2727–34. doi: 10.1016/j.foodchem.2013.05.040
32. Liu L, Chen D, Yu B, Yin H, Huang Z, Luo Y, et al. Fructooligosaccharides improve growth performance and intestinal epithelium function in weaned pigs exposed to enterotoxigenic: Escherichia coli. Food Funct. (2020) 11:9599–612. doi: 10.1039/D0FO01998D
33. Yu E, Chen D, Yu B, Luo Y, Zheng P, Yin H, et al. Amelioration of enterotoxigenic Escherichia coli-induced disruption of intestinal epithelium by manno-oligosaccharide in weaned pigs. J Funct Foods. (2021) 82:104492. doi: 10.1016/j.jff.2021.104492
34. Wang J, Zeng Y, Wang S, Liu H, Zhang D, Zhang W, et al. Swine-derived probiotic Lactobacillus plantarum inhibits growth and adhesion of enterotoxigenic Escherichia coli and mediates host defense. Front Microbiol. (2018) 9:1–11. doi: 10.3389/fmicb.2018.01364
35. Yin H, Ye P, Lei Q, Cheng Y, Yu H, Du J, et al. In vitro probiotic properties of Pediococcus pentosaceus L1 and its effects on enterotoxigenic Escherichia coli-induced inflammatory responses in porcine intestinal epithelial cells. Microb Pathog. (2020) 144:104163. doi: 10.1016/j.micpath.2020.104163
36. Hansen LHB, Cieplak T, Nielsen B, Zhang Y, Lauridsen C, Canibe N. Screening of probiotic candidates in a simulated piglet small intestine in vitro model. FEMS Microbiol Lett. (2021) 368:1–8. doi: 10.1093/femsle/fnab045
37. Sudan S, Zhan X, Li J. A novel probiotic bacillus subtilis strain confers cytoprotection to host pig intestinal epithelial cells during enterotoxic Escherichia coli infection. Microbiol Spectr. (2022) 10:e0125721. doi: 10.1128/spectrum.01257-21
38. Pézsa NP, Kovács D, Gálfi P, Rácz B, Farkas O. Effect of Enterococcus faecium NCIMB 10415 on gut barrier function, internal redox state, proinflammatory response and pathogen inhibition properties in porcine intestinal epithelial cells. Nutrients. (2022) 14:1486. doi: 10.3390/nu14071486
39. Kaewchomphunuch T, Charoenpichitnunt T, Thongbaiyai V, Ngamwongsatit N, Kaeoket K. Cell-free culture supernatants of Lactobacillus spp. and Pediococcus spp inhibit growth of pathogenic Escherichia coli isolated from pigs in Thailand. BMC Vet Res. (2022) 18:1–13. doi: 10.1186/s12917-022-03140-8
40. Daudelin JF, Lessard M, Beaudoin F, Nadeau É, Bissonnette N, Boutin Y, et al. Administration of probiotics influences F4 (K88)-positive enterotoxigenic Escherichia coli attachment and intestinal cytokine expression in weaned pigs. Vet Res. (2011) 42:69. doi: 10.1186/1297-9716-42-69
41. Czyzewska-Dors E, Kwit K, Stasiak E, Rachubik J, Slizewska K, Pomorska-Mól M. Effects of newly developed synbiotic and commercial probiotic products on the haematological indices, serum cytokines, acute phase proteins concentration, and serum immunoglobulins amount in sows and growing pigs- A pilot study. J Vet Res. (2018) 62:317–28. doi: 10.2478/jvetres-2018-0046
42. Li HH, Li YP, Zhu Q, Qiao JY, Wang WJ. Dietary supplementation with Clostridium butyricum helps to improve the intestinal barrier function of weaned piglets challenged with enterotoxigenic Escherichia coli K88. J Appl Microbiol. (2018) 125:964–75. doi: 10.1111/jam.13936
43. Hrala M, Bosák J, Micenková L, Krenová J, Lexa M, Pirková V, et al. Escherichia coli strains producing selected bacteriocins inhibit porcine enterotoxigenic Escherichia coli (ETEC) under both In Vitro and In Vivo Conditions. Appl Environ Microbiol. (2021) 87:1–12. doi: 10.1128/AEM.03121-20
44. Sun Y, Duarte ME, Kim SW. Dietary inclusion of multispecies probiotics to reduce the severity of post-weaning diarrhea caused by Escherichia coli F18+ in pigs. Anim Nutr. (2021) 7:326–33. doi: 10.1016/j.aninu.2020.08.012
45. Apiwatsiri P, Pupa P, Sirichokchatchawan W, Sawaswong V, Nimsamer P, Payungporn S, et al. Metagenomic analysis of the gut microbiota in piglets either challenged or not with enterotoxigenic Escherichia coli reveals beneficial effects of probiotics on microbiome composition, resistome, digestive function and oxidative stress responses. PLoS ONE. (2022) 17:1–23. doi: 10.1371/journal.pone.0269959
46. Qin D, Bai Y, Li Y, Huang Y, Li L, Wang G, et al. Changes in Gut Microbiota by the Lactobacillus casei Anchoring the K88 Fimbrial Protein Prevented Newborn Piglets From Clinical Diarrhea. Front Cell Infect Microbiol. (2022) 12:1–14. doi: 10.3389/fcimb.2022.842007
47. Zhong Y, Fu D, Deng Z, Tang W, Mao J, Zhu T, et al. Lactic acid bacteria mixture isolated from wild pig alleviated the gut inflammation of mice challenged by Escherichia coli. Front Immunol. (2022) 13:1–13. doi: 10.3389/fimmu.2022.822754
48. Krause DO, Bhandari SK, House JD, Nyachoti CM. Response of nursery pigs to a synbiotic preparation of starch and an anti-Escherichia coli K88 Probiotic. Appl Environ Microbio. (2010) 76:8192–200. doi: 10.1128/AEM.01427-10
49. Guerra-Ordaz AA, González-Ortiz G, La Ragione RM, Woodward MJ, Collins JW, Pérez JF, et al. Lactulose and Lactobacillus plantarum, a potential complementary synbiotic to control postweaning colibacillosis in piglets. Appl Environ Microbiol. (2014) 80:4879–86. doi: 10.1128/AEM.00770-14
50. Wang W, Wang Y, Hao X, Duan Y, Meng Z, An X, et al. Dietary fermented soybean meal replacement alleviates diarrhea in weaned piglets challenged with enterotoxigenic Escherichia coli K88 by modulating inflammatory cytokine levels and cecal microbiota composition. BMC Veterinary Research, (2020) 16:1–11. doi: 10.1186/s12917-020-02466-5
51. Girard M, Tretola M, Bee G. A single dose of synbiotics and vitamins at birth affects piglet microbiota before weaning and modifies post-weaning performance. Animals. (2021) 11:1–14. doi: 10.3390/ani11010084
52. Bonetti A, Tugnoli B, Rossi B, Giovagnoni G, Piva A, Grilli E. Nature-identical compounds and organic acids reduce E. coli K88 growth and virulence gene expression in vitro. Toxins. (2020) 12:1–12. doi: 10.3390/toxins12080468
53. Wang J, Lu JX, Xie XW, Xiong J, Huang N, Wei H, et al. Blend of organic acids and medium chain fatty acids prevents the inflammatory response and intestinal barrier dysfunction in mice challenged with enterohemorrhagic Escherichia coli O157:H7. Int Immunopharmacol. (2018) 58:64–71. doi: 10.1016/j.intimp.2018.03.014
54. Ren C, Zhou Q, Guan W, Lin X, Wang Y, Song H, et al. Immune response of piglets receiving mixture of formic and propionic acid alone or with either capric acid or bacillus licheniformis after Escherichia coli challenge. Biomed Res Int. (2019) 2019:6416187. doi: 10.1155/2019/6416187
55. Jiménez MJ, Berrios R, Stelzhammer S, Bracarense APFRL. Ingestion of organic acids and cinnamaldehyde improves tissue homeostasis of piglets exposed to enterotoxic Escherichia coli (ETEC). J Anim Sci. (2020) 98:1–11. doi: 10.1093/jas/skaa238
56. Choi J, Wang L, Liu S, Lu P, Zhao X, Liu H, et al. Effects of a microencapsulated formula of organic acids and essential oils on nutrient absorption, immunity, gut barrier function, and abundance of enterotoxigenic Escherichia coli F4 in weaned piglets challenged with E. coli F4. J Anim Sci. (2020) 98:1–17. doi: 10.1093/jas/skaa259
57. Fratini F, Forzan M, Turchi B, Mancini S, Alcamo G, Pedonese F, et al. In vitro antibacterial activity of manuka (Leptospermum scoparium J.R. et G. Forst) and winter savory (satureja montana l.) essential oils and their blends against pathogenic E. coli isolates from pigs. Animals. (2020) 10:1–8. doi: 10.3390/ani10122202
58. Mohana Devi S, Lee SI, Kim IH. Effect of phytogenics on growth performance, fecal score, blood profiles, fecal noxious gas emission, digestibility, and intestinal morphology of weanling pigs challenged with Escherichia coli K88. Pol J Vet Sci. (2015) 18:557–64. doi: 10.1515/pjvs-2015-0072
59. Zeng Z, Xu X, Zhang Q, Li P, Zhao P, Li Q, et al. Effects of essential oil supplementation of a low-energy diet on performance, intestinal morphology and microflora, immune properties and antioxidant activities in weaned pigs. Anim Sci J. (2015) 86:279–85. doi: 10.1111/asj.12277
60. Montoya D, D'angelo M, Martín-orúe SM, Rodríguez-sorrento A, Saladrigas-García M, Araujo C. Effectiveness of two plant-based in-feed additives against an Escherichia coli F4 oral challenge in weaned piglets. Animals. (2021) 11:1–28. doi: 10.3390/ani11072024
61. Maito CD, Diego Brandão Melo A, da Fonseca de Oliveira AC, Genova JL, Filho JRE, de Macedo REF, et al. Simultaneous feeding of calcium butyrate and tannin extract decreased the incidence of diarrhea and proinflammatory markers in weaned piglets. Anim Biosci. (2022) 35:87–95. doi: 10.5713/ab.21.0011
62. Caprarulo V, Turin L, Reggi S, Dell'anno M, Riccaboni P, Trevisi P, et al. Protective effect of phytogenic based additives in enterotoxigenic Escherichia coli challenged piglets. Vet Res Commun. (2022). doi: 10.21203/rs.3.rs-1207181/v1. [Epub ahead of print].
63. Chang SY, Song MH, Lee JH, Oh HJ, Kim YJ, An JW, et al. Phytogenic feed additives alleviate pathogenic Escherichia coli-induced intestinal damage through improving barrier integrity and inhibiting inflammation in weaned pigs. J Anim Sci Biotechnol. (2022) 13:1–12. doi: 10.1186/s40104-022-00750-y
64. Kim N, Gu MJ, Kye YC, Ju YJ, Hong R, Ju D, et al. Bacteriophage EK99P-1 alleviates enterotoxigenic Escherichia coli K99-induced barrier dysfunction and inflammation. Sci Rep. (2022) 12:1–10. doi: 10.1038/s41598-022-04861-4
65. Yan L, Hong SM, Kim IH. Effect of bacteriophage supplementation on the growth performance, nutrient digestibility, blood characteristics, and fecal Microbial Shedding in Growing Pigs. Asian-Australasian J Anim Sci. (2012) 25:1451–6. doi: 10.5713/ajas.2012.12253
66. Cha S, Bin Yoo AN, Lee WJ, Shin MK, Jung MH, Shin SW, et al. Effect of bacteriophage in enterotoxigenic Escherichia coli (ETEC) infected pigs. J Vet Med Sci. (2012) 74:1037–9. doi: 10.1292/jvms.11-0556
67. Kim KH, Ingale SL, Kim JS, Lee SH, Lee JH, Kwon IK, et al. Bacteriophage and probiotics both enhance the performance of growing pigs but bacteriophage are more effective. Anim Feed Sci Technol. (2014) 196:88–95. doi: 10.1016/j.anifeedsci.2014.06.012
68. Lee S, Hosseindoust A, Goel A, Choi Y, Kwon IK, Chae BJ. Effects of dietary supplementation of bacteriophage with or without zinc oxide on the performance and gut development of weanling pigs. Ital J Anim Sci. (2016) 15:412–8. doi: 10.1080/1828051X.2016.1188676
69. Torrallardona D, Polo J. Effect of spray-dried porcine plasma protein and egg antibodies in diets for weaned pigs Under environmental challenge conditions. J Swine Heal Prod. (2016) 24:21–8.
70. Che L, Hu L, Zhou Q, Peng X, Liu Y, Luo Y, et al. Microbial insight into dietary protein source affects intestinal function of pigs with intrauterine growth retardation. Eur J Nutr. (2020) 59:327–44. doi: 10.1007/s00394-019-01910-z
71. Crenshaw J, del Río LLSanjoaquin L, Tibble S, González-Solé F, Solà-Oriol D, et al. Effect of spray-dried porcine plasma in peripartum sow feed on subsequent litter size. Porc Heal Manag. (2021) 7:1–16. doi: 10.1186/s40813-020-00180-0
72. Alustiza F, Bellingeri R, Picco N, Motta C, Grosso MC, Barbero CA, et al. IgY against enterotoxigenic Escherichia coli administered by hydrogel-carbon nanotubes composites to prevent neonatal diarrhoea in experimentally challenged piglets. Vaccine. (2016) 34:3291–7. doi: 10.1016/j.vaccine.2016.05.004
73. Wang Z, Li J, Li J, Li Y, Wang L, Wang Q, et al. Protective effect of chicken egg yolk immunoglobulins (IgY) against enterotoxigenic Escherichia coli K88 adhesion in weaned piglets. BMC Vet Res. (2019) 15:1–12. doi: 10.1186/s12917-019-1958-x
74. Dierick M, Weken H, Van der Rybarczyk J, Vanrompay D, Devriendt B, Cox E. Porcine and bovine forms of lactoferrin inhibit growth of porcine enterotoxigenic Escherichia coli and degrade its virulence factors. Appl Environ Microbiol. (2020) 86:1–15. doi: 10.1128/AEM.00524-20
75. Gelgel KTP, Sudipa PH, Mufa RMD. Lactoferrin effect on piglets in terms of body weight and number of bacteria. EAS J Vet Med Sci. (2021) 3:58–61. doi: 10.36349/easjvms.2021.v03i06.001
76. Liao SF, Nyachoti M. Using probiotics to improve swine gut health and nutrient utilization. Anim Nutr. (2017) 3:331–43. doi: 10.1016/j.aninu.2017.06.007
77. Timmerman HM, Koning CJM, Mulder L, Rombouts FM, Beynen AC. Monostrain, multistrain and multispecies probiotics —a comparison of functionality and efficacy. Int J Food Microbiol. (2004) 96:219–33. doi: 10.1016/j.ijfoodmicro.2004.05.012
78. Su W, Gong T, Jiang Z, Lu Z, Wang Y. The role of probiotics in alleviating postweaning diarrhea in piglets from the perspective of intestinal barriers. Front Cell Infect Microbiol. (2022) 12:1–12. doi: 10.3389/fcimb.2022.883107
79. Swanson KS, Gibson GR, Hutkins R, Reimer RA, Reid G, Verbeke K, et al. The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of synbiotics. Nat Rev Gastroenterol Hepatol. (2020) 17:687–701. doi: 10.1038/s41575-020-0344-2
80. Ferronato G, Prandini A. Dietary supplementation of inorganic, organic, and fatty acids in pig: a review. Animals. (2020) 10:1740. doi: 10.3390/ani10101740
81. Papatsiros VG, Billinis C. The prophylactic use of acidifiers as antibacterial agents in Swine. In:Bobbarala V, , editor. Antimicrobial Agents. London: IntechOpen (2012). doi: 10.5772/32278
82. Upadhaya SD, Kim IH. Efficacy of phytogenic feed additive on performance, production and health status of monogastric animals - A review. Ann Anim Sci. (2017) 17:929–48. doi: 10.1515/aoas-2016-0079
83. Lillehoj H, Liu Y, Calsamiglia S, Fernandez-Miyakawa ME, Chi F, Cravens RL, et al. Phytochemicals as antibiotic alternatives to promote growth and enhance host health. Vet Res. (2018) 49:1–18. doi: 10.1186/s13567-018-0562-6
84. Reyes-Camacho D, Vinyeta E, Pérez JF, Aumiller T, Criado L, Palade LM, et al. Phytogenic actives supplemented in hyperprolific sows: Effects on maternal transfer of phytogenic compounds, colostrum and milk features, performance and antioxidant status of sows and their offspring, and piglet intestinal gene expression. J Anim Sci. (2020) 98:1–13. doi: 10.1093/jas/skz390
85. Jamal M, Bukhari SMAUS, Andleeb S, Ali M, Raza S, Nawaz MA, et al. Bacteriophages: an overview of the control strategies against multiple bacterial infections in different fields. J Basic Microbiol. (2019) 59:123–33. doi: 10.1002/jobm.201800412
86. Smith HW, Huggins MB. Successful treatment of experimental Escherichia coli infections in mice using phage: its general superiority over antibiotics. J Gen Microbiol. (1982) 128:307–18. doi: 10.1099/00221287-128-2-307
87. Smith HW, Huggins MB. Effectiveness of phages in treating experimental Escherichia coli diarhoea in calves, piglets and lambs. J Gen Microbiol. (1983) 129:2659–75. doi: 10.1099/00221287-129-8-2659
88. Smith HW, Huggins MB, Shaw KM. The control of experimental Escherichia coli diarrhoea in calves by means of bacteriophages. J Gen Microbiol. (1987) 133:1111–26. doi: 10.1099/00221287-133-5-1111
89. Jamalludeen N, Johnson RP, Shewen PE, Gyles CL. Evaluation of bacteriophages for prevention and treatment of diarrhea due to experimental enterotoxigenic Escherichia coli O149 infection of pigs. Vet Microbiol. (2009) 136:135–41. doi: 10.1016/j.vetmic.2008.10.021
90. Campbell JM, Crenshaw JD, González-Esquerra R, Polo J. Impact of spray-dried plasma on intestinal health and broiler performance. Microorganisms. (2019) 7:6–8. doi: 10.3390/microorganisms7080219
91. Pérez-Bosque A, Polo J, Torrallardona D. Spray dried plasma as an alternative to antibiotics in piglet feeds, mode of action and biosafety. Porc Heal Manag. (2016) 2:1–10. doi: 10.1186/s40813-016-0034-1
92. Torrallardona D, Conde MR, Badiola I, Polo J, Brufau J. Effect of fishmeal replacement with spray-dried animal plasma and colistin on intestinal structure, intestinal microbiology, and performance of weanling pigs challenged with Escherichia coli K99. J Anim Sci. (2003) 81:1220–6. doi: 10.2527/2003.8151220x
93. Nofrarías M, Manzanilla EG, Pujols J, Gibert X, Majó N, Segalés J, et al. Effects of spray-dried porcine plasma and plant extracts on intestinal morphology and on leukocyte cell subsets of weaned pigs. J Anim Sci. (2006) 84:2735–42. doi: 10.2527/jas.2005-414
94. Han S, Wen Y, Yang F, He P. Chicken egg yolk antibody (IgY) protects mice against enterotoxigenic Escherichia coli infection through improving intestinal health and immune response. Front Cell Infect Microbiol. (2021) 11:1–14. doi: 10.3389/fcimb.2021.662710
95. Baker EN, Baker HM. Molecular structure, binding properties and dynamics of lactoferrin. Cell Mol Life Sci. (2005) 62:2531–9. doi: 10.1007/s00018-005-5368-9
96. Bai H, Xue X, Hou Z, Zhou Y, Meng J, Luo X. Antisense antibiotics: a brief review of novel target discovery and delivery. Curr Drug Discov Technol. (2010) 7:76–85. doi: 10.2174/157016310793180594
97. Sully EK, Geller BL. Antisense antimicrobial therapeutic. Curr Opin Microbiol. (2016) 33:45–55. doi: 10.1016/j.mib.2016.05.017
98. Hegarty JP, Stewart DB. Advances in therapeutic bacterial antisense biotechnology. Appl Microbiol Biotechnol. (2018) 102:1055–65. doi: 10.1007/s00253-017-8671-0
99. Scoles DR, Minikel EV, Pulst SM. Antisense oligonucleotides: a primer. Neurol Genet. (2019) 5:1–8. doi: 10.1212/NXG.0000000000000323
100. Zamay TN, Zamay GS, Belyanina IV, Zamay SS, Denisenko VV, Kolovskaya OS, et al. Noninvasive microsurgery using aptamer-functionalized magnetic microdisks for tumor cell eradication. Nucleic Acid Ther. (2017) 27:105–14. doi: 10.1089/nat.2016.0634
101. Zhu G, Chen X. Aptamer-based targeted therapy. Adv Drug Deliv Rev. (2018) 134:65–78. doi: 10.1016/j.addr.2018.08.005
102. Blind M, Blank M. Aptamer selection technology and recent advances. Mol Ther Nucleic Acids. (2015) 4:e223. doi: 10.1038/mtna.2014.74
103. Dunn M, Jimenez RM, Chaput J. Analysis of aptamer discovery and technology. Nat Rev Chem. (2017) 1:1–16. doi: 10.1038/s41570-017-0076
104. Keefe AD, Pai S, Ellington A. Aptamers as therapeutics. Nat Rev Drug Discov. (2010) 9:537–50. doi: 10.1038/nrd3141
105. Stoltenburg R, Reinemann C, Strehlitz B. SELEX-A (r)evolutionary method to generate high-affinity nucleic acid ligands. Biomol Eng. (2007) 24:381–403. doi: 10.1016/j.bioeng.2007.06.001
Keywords: enteric colibacillosis, post-weaning diarrhea, gut microbiome, antibiotic therapy, zinc oxide, novel antimicrobial approaches
Citation: Castro J, Barros MM, Araújo D, Campos AM, Oliveira R, Silva S and Almeida C (2022) Swine enteric colibacillosis: Current treatment avenues and future directions. Front. Vet. Sci. 9:981207. doi: 10.3389/fvets.2022.981207
Received: 29 June 2022; Accepted: 10 October 2022;
Published: 28 October 2022.
Edited by:
Mariangela Caroprese, University of Foggia, ItalyReviewed by:
Eric R. Burrough, Iowa State University, United StatesCopyright © 2022 Castro, Barros, Araújo, Campos, Oliveira, Silva and Almeida. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Joana Castro, am9hbmEuY2FzdHJvQGluaWF2LnB0; Carina Almeida, Y2FyaW5hLmFsbWVpZGFAaW5pYXYucHQ=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.