 Qi Sun1†
Qi Sun1† Hu Xu
Hu Xu Hongliang Zhang
Hongliang Zhang
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
BRIEF RESEARCH REPORT article
Front. Vet. Sci. , 09 September 2022
Sec. Veterinary Epidemiology and Economics
Volume 9 - 2022 | https://doi.org/10.3389/fvets.2022.974743
This article is part of the Research Topic Rising Stars in Veterinary Epidemiology and Economics 2022: Porcine Reproductive and Respiratory Syndrome Virus: Epidemiology, Immunology and Virology View all 5 articles
Porcine reproductive and respiratory syndrome virus 1 (PRRSV-1) is one of the main pathogens causing porcine reproductive and respiratory syndrome (PRRS). In recent years, the rate of PRRSV-1 detection in China has gradually increased, and the PRRSV-1 strains reported in China belong to subtype I (Global; Clade A-L). In the present study, a novel PRRSV-1 strain, TZJ2134, was found during epidemiological surveillance of PRRSV-1 in Shandong Province in China. We obtained two fragments of the TZJ2134 genome: TZJ2134-L12 (located at nt 1672-nt 2112 in the partial Nsp2 gene) and TZJ2134-(A+B) (located at nt 7463-nt 11272 in the partial Nsp9, complete Nsp10 and partial Nsp11 genes). Phylogenetic and recombination analyses based on the two sequences showed that TZJ2134 is a recombinant strain derived from two commercial PRRSV-1 modified live vaccine (MLV) strains (the Amervac vaccine and DV vaccine strains) that formed a new recombinant subgroup of DV+Amervac-like isolates with other strains. However, PRRSV-1 MLV is not currently allowed for use in China. This study is the first to detected recombinant PRRSV-1 MLV strain in China and provides new data for the epidemiological study of PRRSV-1 in China. The existence of the TZJ2134 strain is a reminder that the swine surveillance at the Chinese customs should be strengthened.
Porcine reproductive and respiratory syndrome (PRRS) is a highly contagious disease causing substantial economic losses in the swine industry worldwide. It is mainly characterized by widespread reproductive failure in pregnant sows and respiratory symptoms in pigs of all ages (1). PRRSV is divided into two species, namely, Betaarterivirus suid 1 (PRRSV-1) and Betaarterivirus suid 2 (PRRSV-2) (https://talk.ictvonline.org/taxonomy/p/taxonomy-history?taxnode_id=20171832), which share only 50–60% nucleotide sequence identity (2) and have attracted increasing attention due to the high incidence of PRRSV mutation and recombination. Based on the phylogenetic analysis of ORF5 nucleotide sequences and the global PRRSV classification system, PRRSV-1 is divided into four subtypes: subtype I (Global; Clade A-L), subtype I (Russian), subtype II, and subtype III. PRRSV-2 can be divided into nine subtypes: lineages 1~9 (3).
PRRSV-1 and PRRSV-2 have significant differences in geographical distribution. PRRSV-2 was isolated in 1992 in America (ATCC-VR2332, the North American prototypic strain) (4) and is mainly prevalent in North America and Asia (5). PRRSV-1 was isolated in 1991 in the Netherlands (Lelystad virus, the European prototypic strain) (6) and is mainly prevalent in Europe. However, only subtype I (Global; Clade A-L) has spread to continents other than Europe. The remaining subtypes have been reported only in Eastern European countries and Russia (7). In 1997, the isolated PRRSV-1 strain B13 (GenBank: AY633973) was first detected by customs in Mainland China. In 2011, Chen et al. isolated PRRSV-1 strains (BJEU06-1, NMEU09-1) in China, which was the first report of wild PRRSV-1 isolates on a pig farm in mainland China (8). To date, PRRSV-1 has been prevalent in at least 20 provinces in China (9–13). In recent years, the rate of PRRSV-1 detection in China has gradually increased. The PRRSV-1 strains reported in China all belong to subtype I (Global) and can be divided into four subgroups (NMEU09-1-like, Amervac-like, HKEU16-like, and BJEU06-1-like isolates) (14). However, the number of PRRSV-1 infections detected in China was found to be lower than that of PRRSV-2 infections (15, 16). We speculated that the reason might be that PRRSV-1 is not given much attention because it is not the main epidemic strain in China, and there is evidence that most PRRSV-1-infected pigs in China exhibit mild clinical symptoms (10, 17). Even so, there is still a risk of the recombination of vaccine strains to create new ones, so the effect of PRRSV-1 on pigs should not be ignored.
Vaccination is a key strategy for PRRSV-1 prevention and control. In the late 1990s, PRRSV-1 MLV was used in Europe (18). Nevertheless, the particularly high variability of PRRSV and the possibility of vaccine revertant PRRSV emerging in pigs vaccinated with PRRSV MLV could result in recombination between different MLV strains (19, 20) or recombination between MLV strains and wild-type PRRSV strains (14, 21, 22). This potential indicates the importance of the rational use of vaccines. In this study, a novel PRRSV-1 strain was identified in an epidemiological investigation of PRRSV in China; this strain was derived from the recombination of two commercial PRRSV-1 MLV strains (Amervac vaccine strain and DV vaccine strain).
In 2021, a clinical (lung) sample, TZJ2134, was detected in an aborted sow from a very small backyard farm in Shandong Province of China that did not utilize vaccination. Tissue sample processing, RNA extraction, cDNA preparation, RT–PCR and genome sequencing were performed as described previously (23, 24). The TZJ2134 strain was identified as PRRSV-1 by RT–PCR with the primer L12 (primer for specific detection of PRRSV-1) (Supplementary Table S1). Based on the complete genomic sequence of PRRSV-1, eight primer pairs were designed for RT–PCR amplification and sequencing (Supplementary Table S1), and two overlapping fragments of TZJ2134 were amplified by RT–PCR. Each PCR product was purified with the E.Z.N.A.® Gel Extraction Kit-Omega Bio-Tek, cloned into the pMD18-T vector according to the manufacturer's instructions, and then submitted to Comate Bioscience Co., Ltd. (Changchun, China) for sequencing.
Sequence analysis was performed with DNASTAR (version 7.1) software. The resulting sequences were assembled using SeqMan. Phylogenetic trees were generated and molecular evolutionary analyses were performed by using the neighbor-joining method in MEGA 7.0. with 1,000 bootstrap replicates for each node (25). The generated phylogenetic tree was annotated using iTOL online software (https://itol.embl.de/) (26). Sixty-five representative PRRSV-1 strains from different subtypes by referring to the following articles (9, 11–14, 27) available in GenBank were used for comparative sequence analyses in this study.
To analyze the recombination events, RDP4 and the NCBI BLAST results were used to identify the major and minor parental strains (28). Seven different algorithms (RDP, GeneConv, BootScan, MaxChi, Chimera, SiScan, and 3 Seq) embedded in the RDP4 software package with Bonferroni correction were utilized to detect recombination events and breakpoints. Detection using four or more of the seven methods implemented in RDP4 was taken as significant evidence for recombination. Recombination events were further confirmed by SimPlot 3.5.1 (29), and boot scanning analysis was performed with a 200-bp window, sliding along the genome alignment with a step size of 20 bp. Other strains in the same group as TZJ2134-(A+B) (PRRS-FR-2014-56-11-1, DK-2011-05-23-9 and OLot/91 strain) were analyzed by SimPlot 3.5.1 (29) using the same method.
In 2021, the TZJ2134 strain was isolated in Shandong Province in China and showed positivity for PRRSV-1 by detection using primer L12 (Supplementary Table S1). Subsequently, a sequence of TZJ2134 (TZJ2134-L12) was obtained by amplification with the detection primer L12, located in the partial Nsp2 gene (nt 1672-nt 2112 of DV) (Figure 1C). Phylogenetic analysis showed that all Chinese PRRSV-1 isolates belonged to subtype I and could be divided into four subgroups (Amervac-like, BJEU06-1-like, HKEU16-like, and NMEU09-1-like isolates) (Figure 1). TZJ2134-L12 belonged to DV-like isolates and shared the highest sequence identity (99.54%) with the DV vaccine strain (Table 1).
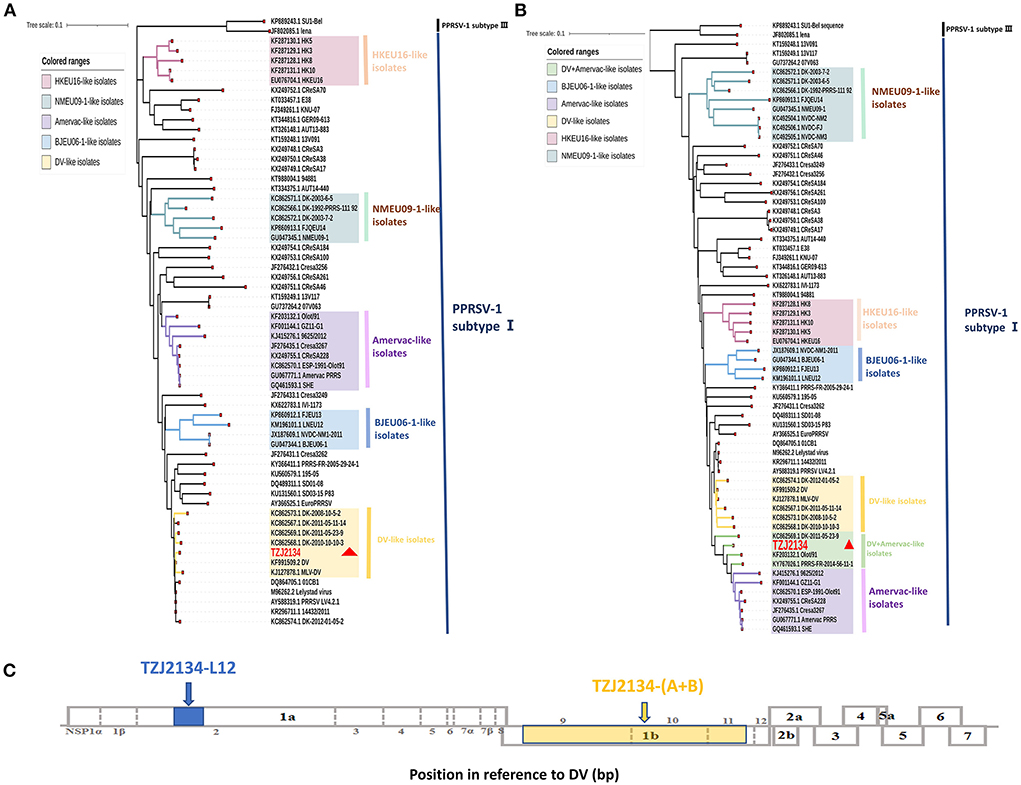
Figure 1. Phylogenetic analysis based on the TZJ2134-L12 and TZJ2134-(A+B) gene sequences and positions of TZJ2134-L12 and TZJ2134-(A+B) in the whole genome structure. (A) Phylogenetic tree based on TZJ2134-L12 of 60 PRRSV-1 strains. The TZJ2134 and DV strains belong to DV-like isolates, which are shown in yellow. (B) Phylogenetic tree based on TZJ2134-(A+B) of 65 PRRSV-1 strains. A novel subgroup DV+Amervac-like isolate is shown in green. (C) The whole genome structure of PRRSV, which shows the positions of 10 open reading frames. TZJ2134-L12 is located at nt 1672-nt 2112 in partial Nsp2, shown in blue; TZJ2134-(A+B) is located at nt 7463-nt 11272 in partial Nsp9, complete Nsp10 and partial Nsp11, shown in yellow. TZJ2134 is indicated with a red triangle in phylogenetic trees.
Table 1. Nucleotide similarity between different gene fragments of TZJ2134 and vaccine strains of PRRSV-1.
To obtain the complete genome of TZJ2134, we designed eight pairs of primers for amplification. Unfortunately, the whole-genome sequence could not be obtained after repeated attempts, and only two overlapping fragments of the TZJ2134 genome were amplified by using the primers Ly-E and Ly-F (Supplementary Table S1). Our primers have amplified multiple PRRSV-1 whole genome sequences in other studies (30). Lung sample was kept frozen during transport, and we designed multiple pairs of primers for amplification using the obtained TZJ2134 sequence, but no more PRRSV-1 nucleotide sequences were obtained in the end. We speculate that there may be three reasons. Firstly, TZJ2134 may have a low viral load in the host. Secondly, although we received a frozen sample, but it is not clear how the samples were stored before transport, which may also contribute to the degradation of viral nucleic acid. Thirdly, there may be high level structure (such as hairpin structure) in some parts of the genome after virus recombination, which leads to the inability or low binding efficiency of primers. Therefore, we did not obtain the full genome sequence of TZJ2134. Subsequently, the resulting sequences of the two overlapping fragments were assembled into a contig [named TZJ2134-(A+B)]. Further genetic evolution and homology analyses showed that TZJ2134-(A+B) located at nt 7463-nt 11272 (3810 nt in length) in partial Nsp9, complete Nsp10 and partial Nsp11 (Figure 1C) shared 97.1% nucleotide identity with the DV vaccine strain and 97.4% nucleotide identity with the Amervac vaccine strain (Table 1). To establish a genetic relationship between TZJ2134-(A+B) and other PRRSV-1 isolates, we generated a phylogenetic tree based on 65 PRRSV-1 strains in GenBank that are prevalent in China and the world (Supplementary Table S2). Phylogenetic analysis showed that TZJ2134-(A+B) was an intermediate between Amervac-like isolates and DV-like isolates and formed a separate subgroup (DV+Amervac-like isolates) with PRRS-FR-2014-56-11-1, DK-2011-05-23-9 and OLot/91 strains (Figure 1B).
BLAST analysis showed that the 5' end and 3' end of the TZJ2134-(A+B) sequence had high homology with DV and Amervac strains, respectively (Data not shown). RDP4 and SimPlot (version 3.5.1) were used to test for recombination of TZJ2134-(A+B). The RDP4 analysis results showed that TZJ2134-(A+B) was a recombinant strain from Amervac and DV vaccine strains with a potential crossover event spanning Nsp10. Additionally, the recombination event was further confirmed by SimPlot 3.5.1, which showed that the recombination breakpoint was located in Nsp10 (nt 9423) (Figure 2A). The recombination break point is not located at the splicing of the two nucleotide sequences, thus TZJ2134 is a natural recombinant virus. Based on the putative recombination breakpoint (nt 9243), we divided TZJ2134-(A+B) into two fragments, TZJ2134-A (nt 7463-nt 9423) and TZJ2134-B (nt 9423-nt 11272), for phylogenetic and homology analyses. The results revealed that the homology between the two fragments and the corresponding parent viruses showed high similarity (Table 1). TZJ2134-A shared the highest nucleotide identity (99.17%) with the DV vaccine strain (Table 1) and belonged to DV-like isolates (Figure 2B). TZJ2134-B shared the highest nucleotide identity (99.73%) with the Amervac vaccine strain (Table 1) and belonged to Amervac-like isolates (Figure 2C). Both the DV and Amervac vaccine strains were PRRSV-1 MLV strains. To the best of our knowledge, only two reports from France and Denmark have described recombination events between two PRRSV-1 MLV strains (20, 31). One of them, PRRS-FR-2014-56-11-1, was the first recombinant strain derived from the Amervac vaccine strain and the DV vaccine strain described previously, with recombination events occurring at nt 500 to nt 1370, nt 3646 to nt 4272 and nt 4972 to nt 8430 in ORF1, as determined using RDP4 (31). Homology analysis showed that TZJ2134-(A+B) has the highest nucleotide identity (97.6%) with PRRS-FR-2014-56-11-1. PRRS-FR-2014-56-11-1 and TZJ2134-(A+B) are intermediates between Amervac-like isolates and DV-like isolates with DK-2011-05-23-9 and OLot/91 strains in the phylogenetic tree (Figure 1B). The recombinant and phylogenetic analysis results showed that all three viruses were recombinant strains derived from the Amervac vaccine strain and DV vaccine strain but with different recombinant patterns (Supplementary Figure S1) and formed a novel subgroup (DV+Amervac-like isolates) in the phylogenetic tree (Figure 1B).
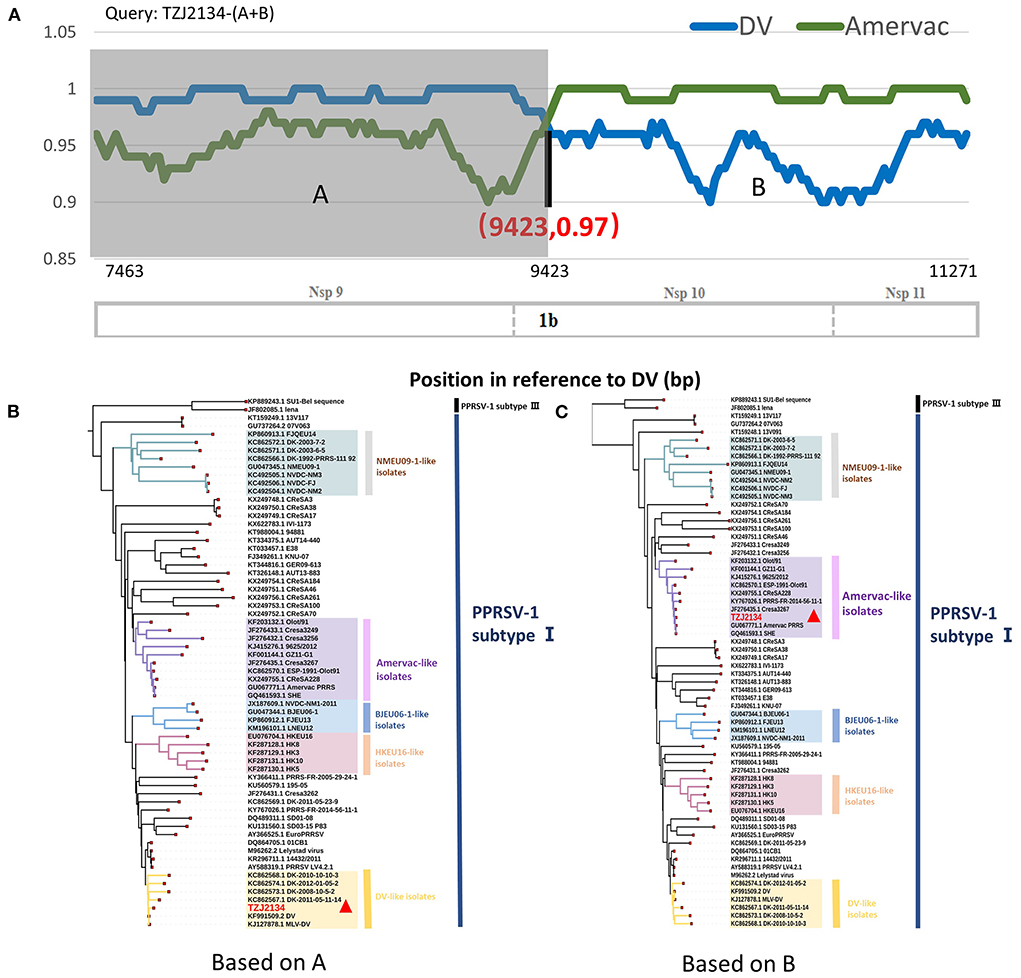
Figure 2. Recombination analysis of TZJ2134-(A+B) and phylogenetic analysis of TZJ2134-A and TZJ2134-B. (A) Recombination analysis of TZJ2134-(A+B). Comparison was performed using TZJ2134-(A+B) as the query sequence and DV (blue) and Amervac (green) as the parent strains. The background color of the DV parental region is gray, whereas that of the Amervac parental region is white. Below the similarity plots is the partial genome structure of PRRSV, which shows the positions of three nonstructural proteins (Nsp9, Nsp10, and Nsp11). (B) Phylogenetic analysis of TZJ2134-A. The parental group (DV-like isolates) is shown in yellow. (C) Phylogenetic analysis of TZJ2134-B. The parental group (Amervac-like isolates) is shown in purple, and the query strain TZJ2134 is indicated with a red triangle.
In the late 1990s, PRRSV-1 MLVs were usually used to control PRRSV-1 infection in Europe (18). PRRSV-1 MLVs used worldwide include Porcilis PRRS (Merck), Amervac PRRS (Laboratories Hipra S.A.), ReproCyc PRRS EU (Boehringer Ingelheim), Pyrsvac-183 (SYVA Laboratories), and Ingelvac PRRSFLEX® EU (Boehringer Ingelheim) (32). They are used in Vietnam, Korea, Russia and Denmark and Taiwan in China. However, PRRSV-1 MLVs are not approved for use in mainland China. In this study we found a strain that is a product of recombination between vaccine strains (Amervac vaccine strain with the DV vaccine strain) and that these vaccines are not approved for use in China.
Since China is the largest pork importer in the world, the swine industry in China is vulnerable to the influence of the foreign swine industry. The existence of the TZJ2134 strain is a reminder that the swine surveillance at the Chinese customs should be strengthened.
In summary, this study reports the first recombination event of the Amervac vaccine strain with the DV vaccine strain in China. These findings suggest that the surveillance against the introduction of foreign animal diseases should be strengthened.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found at: www.ncbi.nlm.nih.gov/nuccore/, ON983961, ON974983.
The animal study was reviewed and approved by the Animal Ethics Committee of the School of Harbin Veterinary Research Institute of the Chinese Academy of Agricultural Sciences. Sampling procedures were performed in accordance with the guidelines of said committee The Animal Ethics Committee Approval Number was SYXK(Hei) 2011022.
Conceived and designed the experiments: Z-JT and HZ. Performed the experiments: QS and HX. Contributed reagents or materials and assisted in some experiments: ZL, CL, and BG. Analyzed the data: QS, HX, and HZ. Contributed to the writing of the manuscript: QS, HX, CL, BG, HZ, and Z-JT. All authors contributed to the article and approved the submitted version.
This study was supported by grants from the National Natural Science Foundation of China (Grant Nos. 32002315 and 32172890) and the China Postdoctoral Fund (Grant No. 2020M680788).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fvets.2022.974743/full#supplementary-material
1. Butler JE, Lager KM, Golde W, Faaberg KS, Sinkora M, Loving C, et al. Porcine reproductive and respiratory syndrome (PRRS): an immune dysregulatory pandemic. Immunol Res. (2014) 59:81–108. doi: 10.1007/s12026-014-8549-5
2. Murtaugh MP, Faaberg KS, Laber J, Elam M, Kapur V. Genetic variation in the PRRS virus. Adv Exp Med Biol. (1998) 440:787–94. doi: 10.1007/978-1-4615-5331-1_102
3. Shi M, Lam TT, Hon CC, Hui RK, Faaberg KS, Wennblom T, et al. Molecular epidemiology of PRRSV: a phylogenetic perspective. Virus Res. (2010) 154:7–17. doi: 10.1016/j.virusres.2010.08.014
4. Wensvoort G, de Kluyver EP, Pol JM, Wagenaar F, Moormann RJ, Hulst MM, et al. Lelystad virus, the cause of porcine epidemic abortion and respiratory syndrome: a review of mystery swine disease research at Lelystad. Vet Microbiol. (1992) 33:185–93. doi: 10.1016/0378-1135(92)90046-V
5. Brar MS, Shi M, Murtaugh MP, Leung FC. Evolutionary diversification of type 2 porcine reproductive and respiratory syndrome virus. J Gen Virol. (2015) 96:1570–80. doi: 10.1099/vir.0.000104
6. Collins JE, Benfield DA, Christianson WT, Harris L, Hennings JC, Shaw DP, et al. Isolation of swine infertility and respiratory syndrome virus (isolate ATCC VR-2332) in North America and experimental reproduction of the disease in gnotobiotic pigs. J Vet Diagn Invest. (1992) 4:117–26. doi: 10.1177/104063879200400201
7. Stadejek T, Stankevicius A, Murtaugh MP, Oleksiewicz MB. Molecular evolution of PRRSV in Europe: current state of play. Vet Microbiol. (2013) 165:21–8. doi: 10.1016/j.vetmic.2013.02.029
8. Chen N, Cao Z, Yu X, Deng X, Zhao T, Wang L, et al. Emergence of novel European genotype porcine reproductive and respiratory syndrome virus in mainland China. J Gen Virol. (2011) 92:880–92. doi: 10.1099/vir.0.027995-0
9. Zhou Z, Liu Q, Hu D, Zhang Q, Han T, Ma Y, et al. Complete genomic characterization and genetic diversity of four European genotype porcine reproductive and respiratory syndrome virus isolates from China in 2011. Virus Genes. (2015) 51:375–84. doi: 10.1007/s11262-015-1256-z
10. Wang X, Yang X, Zhou R, Zhou L, Ge X, Guo X, et al. Genomic characterization and pathogenicity of a strain of type 1 porcine reproductive and respiratory syndrome virus. Virus Res. (2016) 225:40–9. doi: 10.1016/j.virusres.2016.09.006
11. Gao JC, Xiong JY, Ye C, Chang XB, Guo JC, Jiang CG, et al. Genotypic and geographical distribution of porcine reproductive and respiratory syndrome viruses in mainland China in 1996-2016. Vet Microbiol. (2017) 208:164–72. doi: 10.1016/j.vetmic.2017.08.003
12. Liu JK, Wei CH, Dai AL, Fan KW, Yang BH, Huang CF, et al. Complete genomic characterization of two European-genotype porcine reproductive and respiratory syndrome virus isolates in Fujian province of China. Arch Virol. (2017) 162:823–33. doi: 10.1007/s00705-016-3136-9
13. Zhang Q, Song Z, Yu Y, Huang J, Jiang P, Shan H. Genetic analysis of a porcine reproductive and respiratory syndrome virus 1 strain in China with new patterns of amino acid deletions in nsp2, GP3 and GP4. Microb Pathog. (2020) 149:104531. doi: 10.1016/j.micpath.2020.104531
14. Chen N, Liu Q, Qiao M, Deng X, Chen X, Sun M. Whole genome characterization of a novel porcine reproductive and respiratory syndrome virus 1 isolate: genetic evidence for recombination between Amervac vaccine and circulating strains in mainland China. Infect Genet Evol. (2017) 54:308–13. doi: 10.1016/j.meegid.2017.07.024
15. Zhai SL, Lin T, Zhou X, Pei ZF, Wei ZZ, Zhang H, et al. Phylogeographic analysis of porcine reproductive and respiratory syndrome virus 1 in Guangdong province, Southern China. Arch Virol. (2018) 163:2443–9. doi: 10.1007/s00705-018-3873-z
16. Lin WH, Kaewprom K, Wang SY, Lin CF, Yang CY, Chiou MT, et al. Outbreak of Porcine Reproductive and Respiratory Syndrome Virus 1 in Taiwan. Viruses. (2020) 12:316. doi: 10.3390/v12030316
17. Ming S, Yongying M, Bohua L, Huiying L, Xiaoyu D, Qiaorong L, et al. Pathogenic characterization of European genotype porcine reproductive and respiratory syndrome virus recently isolated in Mainland China. Open Virol J. (2017) 11:83–9. doi: 10.2174/1874357901711010083
18. Chae C. Commercial PRRS modified-live virus vaccines. Vaccines. (2021) 9:185. doi: 10.3390/vaccines9020185
19. Eclercy J, Renson P, Lebret A, Hirchaud E, Normand V, Andraud M, et al. A Field recombinant strain derived from two type 1 porcine reproductive and respiratory syndrome virus (PRRSV-1) modified live vaccines shows increased viremia and transmission in SPF Pigs. Viruses. (2019) 11:296. doi: 10.3390/v11030296
20. Kvisgaard LK, Kristensen CS, Ryt-Hansen P, Pedersen K, Stadejek T, Trebbien R, et al. A recombination between two Type 1 porcine reproductive and respiratory syndrome virus (PRRSV-1) vaccine strains has caused severe outbreaks in Danish pigs. Transbound Emerg Dis. (2020) 67:1786–96. doi: 10.1111/tbed.13555
21. Marton S, Szalay D, Kecskemeti S, Forro B, Olasz F, Zadori Z, et al. Coding-complete sequence of a vaccine-derived recombinant porcine reproductive and respiratory syndrome virus strain isolated in Hungary. Arch Virol. (2019) 164:2605–8. doi: 10.1007/s00705-019-04338-2
22. Wang Chen Q, Wang LY, Madson D, Harmon K, Gauger P. Recombination between vaccine and field strains of porcine reproductive and respiratory syndrome virus. Emerg Infect Dis. (2019) 25:2335–7. doi: 10.3201/eid2512.191111
23. Leng CL, Tian ZJ, Zhang WC, Zhang HL, Zhai HY, An TQ, et al. Characterization of two newly emerged isolates of porcine reproductive and respiratory syndrome virus from Northeast China in 2013. Vet Microbiol. (2014) 171:41–52. doi: 10.1016/j.vetmic.2014.03.005
24. Zhang H, Leng C, Feng L, Zhai H, Chen J, Liu C, et al. A new subgenotype 2.1d isolates of classical swine fever virus in China, 2014. Infect Genet Evol. (2015) 34:94–105. doi: 10.1016/j.meegid.2015.05.031
25. Kumar S, Stecher G, Tamura K. MEGA7: molecular evolutionary genetics analysis version 70 for bigger datasets. Mol Biol Evol. (2016) 33:1870–4. doi: 10.1093/molbev/msw054
26. Letunic I, Bork P. Interactive Tree Of Life (iTOL) v5: an online tool for phylogenetic tree display and annotation. Nucleic Acids Res. (2021) 49:W293–6. doi: 10.1093/nar/gkab301
27. Karniychuk UU, Geldhof M, Vanhee M, Van Doorsselaere J, Saveleva TA, Nauwynck HJ. Pathogenesis and antigenic characterization of a new East European subtype 3 porcine reproductive and respiratory syndrome virus isolate. BMC Vet Res. (2010) 6:30. doi: 10.1186/1746-6148-6-30
28. Xiang L, Xu H, Li C, Tang YD, An TQ, Li Z, et al. Long-term genome monitoring retraces the evolution of novel emerging porcine reproductive and respiratory syndrome viruses. Front Microbiol. (2022) 13:885015. doi: 10.3389/fmicb.2022.885015
29. Lole KS, Bollinger RC, Paranjape RS, Gadkari D, Kulkarni SS, Novak NG, et al. Full-length human immunodeficiency virus type 1 genomes from subtype C-infected seroconverters in India, with evidence of intersubtype recombination. J Virol. (1999) 73:152–60. doi: 10.1128/JVI.73.1.152-160.1999
30. Zhang H, Zhang J, Zhang W, Xiang L, Song S, Liu C, et al. Complete genomic characterization of three European genotype porcine reproductive and respiratory syndrome viruses from China in 2016. Zhongguo Yufang Shouyi Xuebao/Chinese Journal of Preventive Veterinary Medicine. (2017) 39:513–7. doi: 10.3969/j.issn.1008-0425.2017.07.01
31. Renson P, Touzain F, Lebret A, Le Dimna M, Quenault H, Normand V, et al. Complete genome sequence of a recombinant porcine reproductive and respiratory syndrome virus strain from two genotype 1 modified live virus vaccine strains. Genome Announc. (2017) 5:e00454–17. doi: 10.1128/genomeA.00454-17
Keywords: PRRSV-1, recombination, novel strains, vaccine, DV, Amervac
Citation: Sun Q, Xu H, Li C, Gong B, Li Z, Tian Z-J and Zhang H (2022) Emergence of a novel PRRSV-1 strain in mainland China: A recombinant strain derived from the two commercial modified live viruses Amervac and DV. Front. Vet. Sci. 9:974743. doi: 10.3389/fvets.2022.974743
Received: 21 June 2022; Accepted: 04 August 2022;
Published: 09 September 2022.
Edited by:
Igor Paploski, University of Minnesota Twin Cities, United StatesReviewed by:
Tongling Shan, Shanghai Veterinary Research Institute (CAAS), ChinaCopyright © 2022 Sun, Xu, Li, Gong, Li, Tian and Zhang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Hongliang Zhang, emhhbmdob25nbGlhbmcwMUBjYWFzLmNu
†These authors have contributed equally to this work and share first authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.