
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Vet. Sci. , 05 October 2022
Sec. Veterinary Infectious Diseases
Volume 9 - 2022 | https://doi.org/10.3389/fvets.2022.946545
This article is part of the Research Topic Current Knowledge In Control And Immunoprophylaxis Of Tick-Borne Diseases Of Veterinary Importance: Special Focus In Diseases Of Cattle View all 6 articles
 Elizabeth Salinas-Estrella
Elizabeth Salinas-Estrella Itzel Amaro-Estrada
Itzel Amaro-Estrada Mayra E. Cobaxin-Cárdenas
Mayra E. Cobaxin-Cárdenas Jesús F. Preciado de la Torre
Jesús F. Preciado de la Torre Sergio D. Rodríguez*
Sergio D. Rodríguez*Bovine anaplasmosis is a tick-borne bacterial disease with a worldwide distribution and the cause of severe economic losses in the livestock industry in many countries, including México. In the present work, we first review the elements of the immune response of the bovine, which allows ameliorating the clinical signs while eliminating the majority of the blood forms and generating an immunologic memory such that future confrontations with the pathogen will not end in disease. On the other hand, many vaccine candidates have been evaluated for the control of bovine anaplasmosis yet without no commercial worldwide effective vaccine. Lastly, the diversity of the pathogen and how this diversity has impaired the many efforts to control the disease are reviewed.
Bovine anaplasmosis is a tick-borne rickettsial disease with worldwide distribution. It causes severe economic losses in the livestock industry in many countries, including México (1, 2). Anaplasma marginale, the causative agent, is a vector-borne, Gram-negative bacterium that replicates in mature erythrocytes of cattle and other ruminants (3). The disease is more notorious in cattle older than 2 years old and is rarely apparent in younger animals (1). Clinical signs include anorexia, jaundice, abortion, weight loss, decreased meat and milk production, and, potentially, death (2). Losses due to bovine anaplasmosis reach billions of dollars worldwide (4).
Cattle affected with any clinical form of anaplasmosis may recover when antibiotics are administered promptly. Yet, the pathogen may not be completely eliminated; thus, some cattle may be lifelong carriers (5, 6), acting as reservoirs for susceptible or healthy animals under inappropriate veterinary practices (7).
A. marginale has a small genome composed roughly of 1.2 mega pair bases, with two gene superfamilies and a highly diverse genetic composition among geographical isolates (8, 9). This genetic composition provides the capability to produce different variants of major surface proteins (MSPs) that help the pathogen evade the immune response and remain within the animal throughout its life span (10).
Ticks are an important A. marginale reservoir and maintain the pathogen in nature. While one-host female ticks (e.g., Rhipicephalus microplus) do not readily move from one animal to another, their male counterparts do so when cattle are held nearby, enabling transmission from an infected to a non-infected host (11, 12). Furthermore, unfed larvae from infected female R. microplus ticks can transmit A. marginale when they feed for the first time on a susceptible host (13).
Vaccination is considered the best option to control infectious diseases. For some diseases, there may be several vaccine options for different species, ages, or sexes (14), but bovine anaplasmosis is not such a case. The problem of bovine anaplasmosis is that several factors allow the pathogen to remain in the host for its entire lifetime (1). There is still no practical solution to eradicate the disease because the tick vector would also have to be eradicated. The absence of immunity in a large proportion of the herd in the presence of ixodicide failure can cause disasters in the form of massive outbreaks (15). Conversely, epidemiological stability, which refers to a minimum of carriers of both ticks and Anaplasma, may not be practical when new susceptible animals or a new strain is introduced into an infected herd (16).
Here, we first reviewed the protective immune response of bovine, which allows controlling A. marginale infections. Then, we reviewed some of the most relevant efforts to develop vaccines, their drawbacks, and their potential for success. Finally, we question the possibility of a vaccine that could protect cattle worldwide, even in the presence of controlled infection.
Anaplasma marginale enters the vascular system through a vector bite. Initial bodies contact bovine erythrocytes through adhesins present on the outer surface of the bacteria (17) and their (still uncharacterized) cognate receptors on the bovine erythrocyte. This inclusion body releases the initial bodies, which infect other erythrocytes, multiply, and promote further infection. Erythrocytes are not destroyed by penetration or release of the initial bodies from the bacteria (3).
The most accepted model for the protective immune response indicates that IgG2 is necessary for the clearance of A. marginale through neutralization of outer membrane B-cell epitopes. At the same time, CD4+ T-cell-mediated macrophage activation is essential for opsonization and microbial killing (18).
Many studies showed that specific antibodies directed to relevant A. marginale protein epitopes correlate with the protection against two of the most important clinical indicators of disease: acute rickettsemia and anemia (19). In this model of immunity, antibodies alone are insufficient for protection, but they are required for macrophage opsonophagocytosis (19).
CD4+ T cells expressing interferon γ (IFN-γ) are central to this model of protective immunity against A. marginale (Figure 1). IFN-γ enhances IgG2 production in cattle (20), and bovine CD4+ T cells expressing IFN-γ have been shown to induce IgG2 secretion in B cells. Similarly, IFN-γ activates macrophages to enhance Fc receptor expression, phagocytosis, phagolysosomal fusion, and the production of rickettsiacidal nitric oxide (21). This model provides the basis for specific cell-mediated immunity against a pathogen limited strictly to intraerythrocytic parasitism, which thus cannot be directly targeted by major histocompatibility complex (MHC) class I-restricted cytotoxic lymphocytes.
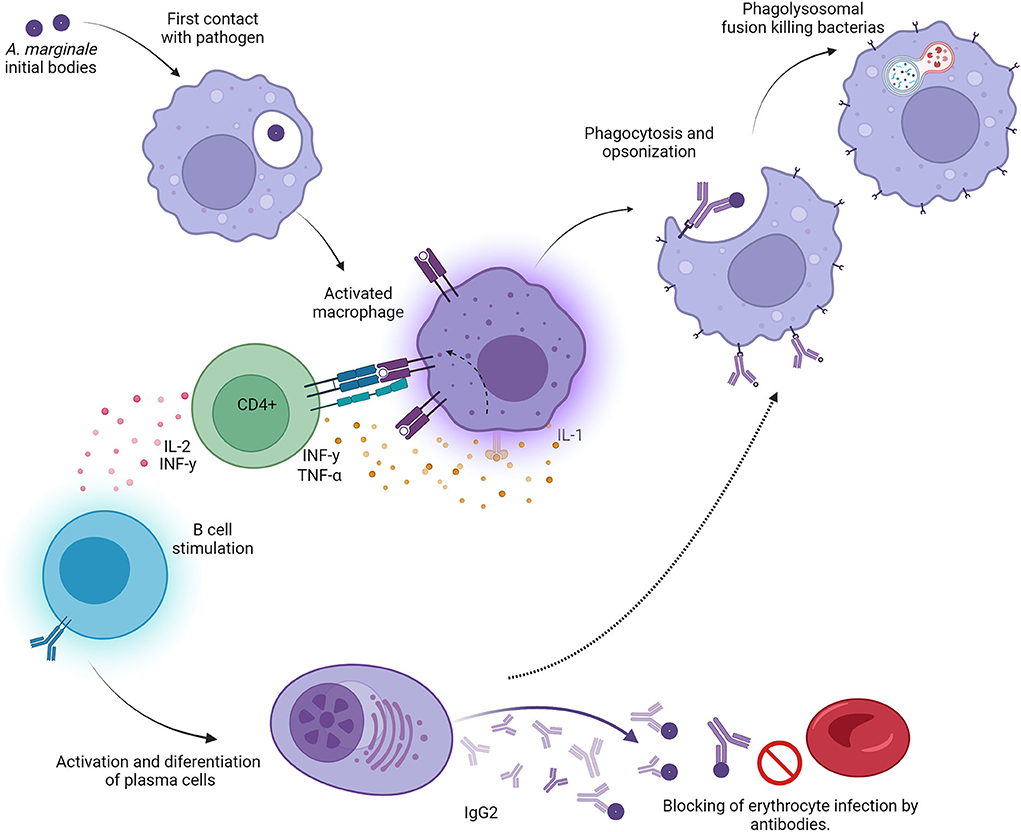
Figure 1. Graphic representation of the “desired” Th1-type immune response in animals protected against bovine anaplasmosis. Initial bodies released from the bovine erythrocyte encounter antigen-presenting cells (macrophages and/or dendritic cells), which engulf, process, and present epitopes to both B and T cells. CD4+ T cells are activated when they encounter the appropriate epitopes in the context of class II major histocompatibility receptors on antigen-presenting cells. These activated T cells secrete specific lymphokines, including IFNγ and IL-2. B cells are activated and stimulated by CD4+ T-cell cytokines and become IgG2-producing plasma cells. The large quantities of IgG2 can neutralize initial bodies, rendering them incapable of infecting normal erythrocytes. At the same time, activated macrophages destroy opsonized bacteria faster in the presence of IFNγ, leading to recovery from the clinical syndrome.
Immunization with Anaplasma outer membrane fractions that have been cross-linked and consist of native proteins induced a protective immune response dependent on Th1 lymphocytes (22, 23). This response is represented by the high production of IgG2, IFN-γ, and IL-2 (21, 24). Unlike IgG1, IgG2 is an opsonizing immunoglobulin; once it binds with its epitope, the immunoglobulin Fc region activates local macrophages and neutrophils, which become more efficient in removing free initial bodies. In turn, the secretion of IFN-γ by specific Th1 lymphocytes augments local phagocyte activity and eliminates opsonized initial bodies [(21, 24); Figure 1].
This model of protective immune response was mostly based on immunization and challenge experiments carried out in calves, mainly 6-month-olds. However, cattle younger than 12 months do not usually develop clinical disease, despite being infected with A. marginale (1, 21). This model of protective immunity was validated by inoculating heifers older than 1 year with an inactivated vaccine incorporated with Quil-A saponin. This adjuvant is known to induce a Th1-type immune response in humans (25). In this experiment, the vaccinated animals showed a Th1-type immune response before and after a challenge with a virulent Mexican strain. Most vaccinated animals had higher IgG2 than IgG1 titers, a higher proportion of CD4+ than CD8+ T cells, and produced IFN-γ. Moreover, few animals that presented the inverse proportions of IgG and T cells (IgG2 < IgG1 and CD4+ < CD8+) suffered from acute clinical signs of disease and required treatment to avoid death (26).
Although the mechanisms of CD4+ preference over CD8+ are still unknown, it is clear that the CD4+-lymphocyte immune response is necessary for protection against natural infection or under conditions of artificial immunization (26). Thus, recent studies on potential native or recombinant antigens for vaccine use include CD4+ T-cell epitopes. Here, we reviewed some of the different efforts to produce an effective vaccine that will stimulate the ideal type of immune response and solidly protect cattle against anaplasmosis with the support of other practices.
Early studies hypothesized great diversity among A. marginale strains from different geographical locations. Molecular analyses confirmed this diversity (27); for example, the Msp1a gene, encoding for a major surface protein (Msp), includes up to 11 highly similar tandem repeat sequences (28). To date, there are over 700 accession numbers (NCBI) for complete or partial amino acid sequences for this protein alone (29–32). By analyzing isolates from different regions and even among organisms isolated in the same location or same animal, it has been shown that, while some proteins are conserved, others are not (33). Furthermore, proteins encoded by multigene families are even more diverse than proteins encoded by a single gene. Msp1b, for instance, is encoded in at least two complete and three partial genes that recombine to produce variants of the same protein (28, 34). Other major surface proteins like Msp4 or Msp5 are coded in single genes and are highly conserved. Unfortunately, these proteins are not good vaccine candidates; Msp4 immunization does not always produce specific antibodies (35), whereas Msp5 induces large amounts of antibodies that are not protective (36, 37).
Conversely, Msp2 and Msp3 proteins are each encoded in multigene families composed of a main gene and a variable number of (5–7) partial genes. The main gene recombines with each partial gene fully or in segments through a mechanism known as segmental gene conversion. Variants emerge at cycles of 6–8 weeks throughout the host's lifetime (38, 39). This information indicates that none of these proteins are suitable vaccine candidates despite their vital role in erythrocyte infection. Other bacterial proteins, like type 4 secretion system (TFSS) proteins, react with IgG2 in vivo; however, they fail to induce protective immunity when inoculated in cattle (40). Future proposals of proteins as vaccine candidates should consider their molecular and antigenic diversities.
Live heterologous A. marginale vaccines have been used since the early 20th century. Theiler reported the presence of Anaplasma centrale as a different organism in South Africa (41), which was less virulent and could be used for the immunization of naive cattle against the more virulent A. marginale (42). A. centrale is still being produced as a live—very often as trivalent (Babesia bovis, B. bigemina, and A. centrale)—vaccine in South Africa, Australia, Argentina, Brazil, Uruguay, Israel (43–47), among others. Thousands of doses of these vaccines are distributed every day, enabling more affordable cattle production. However, a few reports identified A. centrale as the cause of outbreaks with fatalities (48) or failure to induce immunity against A. marginale challenge (49, 50). Potgieter (42), citing Theiler, stated that immunization with A. centrale attenuates the clinical presentation of A. marginale infection but does not prevent the infection from occurring. Thus, live agents should be used with caution.
Using live virulent A. marginale as an immunogen involves the sub-inoculation of blood from infected carriers to susceptible animals paired with treatment—premunition—to avoid acute or fatal anaplasmosis (49–52). In turn, using infected blood from a carrier or a patent animal implies injecting an unknown number of infected erythrocytes; thus, the monitoring period varies from one inoculation to another (53). The results of these types of trials show the unreliability of the method: in some animals, 10 μl of infected blood can induce an infection with a delayed incubation period, while other animals showed no clinical signs (53).
Some virulent strains have been attenuated by passaging organisms in “unnatural” hosts. A. marginale has been passaged in splenectomized sheep and deer (53, 54). These organisms were partially successful in several countries, including Mexico, Perú, and Colombia (55–57). The efficacy of virulent, sheep-attenuated, and deer-attenuated vaccines is different; the deer-passaged strain failed to induce solid protection (56), and live virulent and sheep-attenuated strains induced similar protection (53). However, calves administered with virulent strains showed less severity of the signs after an artificial or natural inoculation challenge 6 months later than those previously administered with the sheep-attenuated strain (54). The difference in the presentation of clinical signs in the immunized animals has been attributed to the lack of cross-protection between strains or to the magnitude of the antibody response between the immunizing and the challenge strains (57).
Anaplasma marginale strains of naturally low virulence have also been tested as potential vaccines. One of the earliest efforts (53) used the Florida strain. Further efforts using local strains have been reported in Australia, Mexico, Brazil, to mention a few (58–61). All these efforts have one thing in common: fresh or frozen infected erythrocytes were inoculated to induce a mild clinical syndrome (premunition). As occurs when using infected blood, there is always the risk of transferring other blood-borne pathogens, including Babesia, Ehrlichia, or virus. For example, in Australia, the bovine leucosis virus (BLV) was transmitted in BLV-free cattle with dire consequences (62).
One of the expected advances in bovine anaplasmosis is the in vitro cultivation of the pathogen. This has been explored for a long time in mammalian systems without success. Early reports claimed that the pathogen could be grown in several conditions, including rabbit bone marrow tissue cultures (63), bovine erythrocytes (64, 65), bovine erythrocytes co-cultured with endothelial cells (66), and endothelial cells (67). Most of these efforts have not been replicated by others or even the same authors.
More recently, A. marginale has been cultured in several tick cell lines (68, 69). Some of these reports showed that major surface proteins expressed in erythrocytic stages in A. marginale and A. centrale are also expressed when cultured in vitro in tick cell lines (70–72). Immunization of cattle with in vitro-cultured A. marginale did induce an antibody immune response but not to the expected protection level (68, 71). The protection level was, at best, comparable to immunization with crude antigens derived from initial bodies against a homologous challenge (72). However, tick cell lines are still a tool to isolate A. marginale from field outbreaks or even carriers, providing material for further characterization (73, 74).
Facing the impossibility of using A. centrale, early vaccine efforts used lyophilized blood with high numbers of infected erythrocytes that was reconstituted with an oil adjuvant and inoculated into susceptible hosts (53). Earlier inactivated vaccines were unpractical due to the high content of erythrocyte stroma that was probably related to cases of neonatal isoerythrolysis in calves born to vaccinated dams (4).
Inactivated preparations of purified initial bodies from bovine erythrocytes have also been used. A study immunized adult cattle with the preparation of initial bodies solubilized with detergent and incorporated with Quil-A saponin. After immunization, the animals were challenge-inoculated with 1 × 109 infected erythrocytes of a heterologous strain, but the vaccine was not effective (75). Similar observations were also reported with other inactivated vaccines (76, 77). In an effort to cover a wider antigenic spectrum, a Mexican vaccine essay used three strains that shared major surface proteins Msp1a and Msp4; only the animals (yearlings) challenged with the combination that included the homologous strain resisted the challenge, while all other groups had to receive chemotherapy to prevent death under stall conditions (78). By contrast, a similar preparation [in adjuvant and antigen composition and applied by the same route as in the study mentioned in Rodríguez et al. (78)] was used to immunize yearlings under the same regime as the previous experiment under ranch conditions; the immunized animals produced specific antibodies (as determined by iELISA) against all three strains and were protected against naturally tick-transmitted anaplasmosis a year after a two-dose vaccine application (79). In both of these studies, high antibody titers were observed, regardless of the vaccination settings. Although the results of these two experiments seem contradictory, one major difference was the challenge dose and settings: the first essay used 108 freshly reactivated infected erythrocytes as inoculum under stall conditions, whereas in the second experiment, animals acquired the infection gradually as they were infested by ticks in the ranch where the experiment was carried out.
University Products LLC (Louisiana, USA) currently offers the only inactivated commercial vaccine against bovine anaplasmosis (https://www.prnewswire.com/news/university-products-llc/). The producer claims that, while the vaccine does not prevent infection with virulent A. marginale, it induces enough immunity to protect cattle against the clinical syndrome. This vaccine has been used in thousands of animals for over two decades since its development. The methodology for extracting the initial bodies is referred to in the study mentioned in Orozco-Vega et al. (80), McCorkle et al. (81).
The search for immunodominant and subdominant proteins in the published A. marginale genomes has yielded lists of potential vaccine candidates, including outer membrane proteins (OMPs), major surface proteins (MSPs), and several type 4 secretion system (TFSS) proteins (82). Early reports explored Msp1a, Msp1b, Msp2, Msp3, Msp4, and Msp5 (83) recombinant proteins for the immunization of calves. Table 1 shows a list of MSPs and other membrane proteins that have been studied as potential candidates for immunization in the form of DNA, plasmids, recombinant proteins, or even synthetic peptides.

Table 1. A summary of some of the many vaccine candidates, animal models, the stage of the study and adjuvants used for immunization against bovine anaplasmosis.
One of the first attempts to produce a recombinant DNA vaccine included the msp1a gene coupled to promoters of different origins into a vaccinia virus vector (84). After inoculating it into mice, this immunogen induced limited production of specific antibodies in experimental subjects.
Another DNA vaccine was constructed by fusing a sequence encoding B- and T-cell antigens from the A. marginale msp1a gene with a BVP22 domain and an invariant-chain MHC class II-targeting motif (fetal liver tyrosine kinase) capable of enhancing dendritic cell antigen uptake and presentation (85). This vaccine was inoculated in 6-month-old calves; it induced proliferative responses and expansion of gamma interferon-positive CD4+ T cells and immunoglobulin G responses against the linked B-cell epitope. However, no challenge with the live agent was reported.
A similar experiment used msp1a, msp1b, and msp5 in the form of recombinant plasmids pET102-msp1α, pET101-msp1β, and pRSET-msp5. Inoculating a mixture of these plasmids primarily induced a Th2-type immune response in mice, and inoculating pET102-msp1α only induced an immunoglobulin response slightly higher than that of negative controls (86).
Using recombinant proteins is the next best choice. A mixture of rMSP1a, rMSP1b, rMSP4, and rMSP5 incorporated into ISCOM and ISCOMATRIX adjuvants induced the production of IgG1 and IgG2 in mice (86). A similar complex of the same recombinant MSPs stimulated the production of IgG, IgG1, and IgG2 when inoculated in mice (87, 93). Recombinant VirB2, VirB4-1, VirB4-2, VirB6-1, VirB7, VirB8-2, VirB9-1, VirB9-2, VirB10, VirB11, and VirD4 of the TFSS proteins were linked to major histocompatibility complex class II DRB3 antigens and were shown to induce IgG and stimulate CD4+ T cells from A. marginale membrane-immunized cattle. In these experiments, not all immunized animals responded to all TFSS proteins, yet most responded to recombinant VirB9-1, VirB9-2, and VirB10 both in antibody production and Th cell lines (96, 97). Inoculation of mice with recombinant VirB9-1 and VirB10 expressed in Pichia pastoris formulated with the self-adjuvanting silica vesicles, SV-100, and 200 μg of VirB9-1 and VirB10 induced higher antibody responses than a similar Quil-A saponin formulation (98). This same preparation induced a strong T-cell reaction in cells from calves previously immunized with A. marginale outer membranes (88).
Many studies used various options for immunizing mice, rabbits, or calves, from vector DNA vaccines to recombinant proteins with different adjuvants (Table 1). These studies include MSPs, TFSS proteins, outer membrane proteins, and other subdominant proteins, exemplifying many attempts for an immunoprophylactic solution to bovine anaplasmosis. Recombinant proteins have been tested in mice or calves for the type of immune response (Th1 or 2) (89, 110). In one case, steers immunized with a mixture of VirB9.1, VirB9.2, VirB10, VirB11, and EfTu produced the desired Th1-type immune response, but this response did not correlate with protection (40).
In addition to the “usual suspects,” other subdominant proteins with putative functions (at least in A. marginale) have been expressed in bacterial systems and used for immunization. Inoculation of AM854 (OmpA equivalent) and AM936 (Asp14 equivalent) incorporated with saponin induced both antigen-specific IgG1 and IgG2 in steers. Vaccinated animals developed higher levels of rickettsemia and greater packed cell volume (PCV) losses than controls immunized with A. marginale membrane fragments or negative controls when challenged with 10 Anaplasma–Dermacentor andersoni-infected ticks (102). In this study, E. coli-expressed recombinant antigens caused more severe bacteremia and clinical syndrome after a challenge. By contrast, the animals inoculated with membrane fractions, which included other proteins in addition to the native proteins of interest, did develop the desired Th-1 immune response.
Finally, a proof-of-concept study used an eight-branched multiple antigenic peptide (R10K-MAP) derived from Msp1a tandem repeat K;S as the antigen (90). The vaccination scheme comprised three doses (a prime boost-like scheme). The first dose consisted of a soluble inoculum applied subcutaneously on one side of the neck. The second dose was an ear implant that included (1) the antigen and one of two adjuvants, DEAE-dextran or Quil A (saponin), or (2) the same antigen with the two adjuvants. The third dose was either a second implant of the antigen with the same adjuvant or the antigen with the alternative adjuvant. After immunization, the experimental animals were challenged with a dose of 109 recently thawed erythrocytes infected with a heterologous strain.
According to the clinical signs developed upon the challenge, the authors suggested that all (three) animals receiving a single adjuvant and one from each group receiving the two adjuvants were not protected from the disease (90). On the one hand, the results of this study are relatively consistent with the use of several adjuvants to increase the antigenicity of the chosen protein (105). On the other hand, this study only included three animals per group, so it is difficult to predict the effects on a larger group of animals. While initially promising, many questions should be answered before such a vaccine can be commercially released. For instance, what is the shortest time between implant application and exposure to a natural challenge in order to ensure adequate protection? Is this antigen the best option for immunization? Can other antigens be included in the implant? Finally, is the implant affordable?
Recombinant organisms are used for the immunoprophylaxis of many diseases (111). Transformation of A. marginale was only recently generated by transposon mutagenesis of the A. marginale Virginia strain. This mutant has extremely reduced expression of outer membrane protein (Omp)9, Omp8, Omp7, and Omp6 genes (108); it can be transmitted by ticks and shows reduced infectivity in both intact and splenectomized cattle (109). The authors did not test for protection against the Virginia wild type or a heterologous challenge. Another vaccine candidate was generated from a St. Maries strain with a transposon-mediated insertion of a 4.5-kb construct containing antibiotic resistance genes for selection and Turbo GFP as a marker. This strain, called AmStM-GFP, grows more slowly than the parent strain in culture (106). It induces immunity and similar clinical parameters for immunization with A. centrale but a lower maximum percentage of infected erythrocytes, a smaller drop in packed cell volume, and a longer time to reach peak bacteremia than wild-type AmStM (107). These new developments represent efforts to control the disease by means of live genetically modified organisms. However, their condition of live agents enables them to transmit other organisms like mycoplasmas and even viruses present in the culture systems.
More than a century has passed since Theiler (41) described both A. marginale and A. centrale as independent causal agents of bovine anaplasmosis and used the latter as a live vaccine. Until now, none of the options for vaccination against anaplasmosis prevent infection. Furthermore, vaccination is only expected to ameliorate the clinical signs should the animal get infected with A. marginale (42). This fact may be advantageous since, once an animal gets infected, it acquires a degree of permanent (concomitant) immunity that lasts throughout the animal's life span.
Since the early use of the A. centrale live vaccine, vaccination has not progressed beyond the use of an inactivated vaccine (in the United States). However, new advances include live attenuated A. marginale, recombinant proteins, synthetic peptides, genetically modified organisms, and an array of adjuvants and delivery systems like IL-2 and antigens linked to MHC class II molecules.
Vaccine design entails many obstacles, including, but is not restricted to, the induction of the desired immune response, the search for the ideal antigen(s) in terms of their contribution to virulence or metabolism, the expression of the antigens of interest at the key moment of pathogen development, and the feasibility of vaccine production, application, and commercialization. Furthermore, saponin has been the most widely used adjuvant for A. marginale vaccine production, followed by water-in-oil (w/o) and water-in-oil-in-water (w/o/w) emulsions, dextrans, and other commercially available mixtures. However, the results of the two trials mentioned previously showed that more than one adjuvant may be needed to stimulate several types of immune cells (90, 105).
In the case of recombinant or synthetic peptides or proteins, one should consider whether the protein is immunogenic and if the relevant epitopes are in the right conformation and within reach of the antibodies. Native proteins embedded within the membrane are immunogenic enough to induce protective immunity, but their recombinant counterparts might not be enough (22, 23). Furthermore, the fact that an antigen triggers the right immune response (IgG2) does not imply that the antigen is immunogenic enough to induce the same response alone or even that it is immunoprotective (40, 102). For example, subdominant proteins such as TFSS proteins appear as suitable vaccine candidates based on their location and function, but they did not induce immunoprotective immunity when tested.
The bleak panorama seems to indicate that we will never achieve cattle immunization against this formidable pathogen. However, the development of inactivated and mRNA vaccines against SARS-CoV-2 is a reminder of humanity's ability to confront health problems.
Live vaccines (A. centrale) have been an alternative for mass vaccination in several countries; vaccination with live cultivable GMOs may be a future alternative. New mutagenic techniques should be considered while designing live vaccines.
Furthermore, while it may seem that we are far from achieving the goal, the publication of more than 20 genome sequences of A. marginale at the NCBI should aid in the search for better vaccine candidates. Many outer membrane proteins and membrane-associated proteins have been tested as vaccine candidates and are yet to be developed into commercial vaccines. Small genomes of A. marginale comprise ~1,000 genes, with many of them (≥60%) still not described (8, 112). This fact should bring hope that other proteins involved in replication or metabolic or signaling pathways may be conserved among strains or may perform vital functions for the parasite survival, providing antigen candidates. New forms of antigen delivery (microtubules, nanoparticles, MAP, etc.) (88, 89, 98, 110) should also be tested to provide promising antigens with the best chance to induce the desired immune response. Finally, the experience of live and whole-cell immunogens suggests that a single antigen hardly induces the appropriate immune response or protects against deliberate or natural challenges. Thus, vaccination experiments with recombinant or synthetic proteins/peptides should include many candidates to mimic the immune response of the whole-cell inactivated or live vaccines.
Thus, we expect bovine anaplasmosis immunization to be realized in the near future, contributing to safer and more profitable cattle production.
SR: conceptualization, review, and editing. ES-E: design of Figure 1. SR, ES-E, MC-C, IA-E, and JP: investigation and original draft preparation and writing. All authors have read and agreed to the published version of the manuscript.
This work was supported by the INIFAP project (SIGI Number 11231734788).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Aubry P, Geale DW. A review of bovine anaplasmosis. Transbound Emerg Dis. (2011) 58:1–30. doi: 10.1111/j.1865-1682.2010.01173.x
2. Rodríguez SD, García Ortiz MA, Jiménez Ocampo R, Vega y Murguía CA. Molecular epidemiology of bovine anaplasmosis with a particular focus in Mexico. Infect Genet Evol. (2009) 9:1092–101. doi: 10.1016/j.meegid.2009.09.007
3. Kocan KM, de La Fuente J, Blouin EF, Garcia-Garcia JC. Adaptations of the tick-borne pathogen, Anaplasma marginale, for survival in cattle and ticks. Exp Appl Acarol. (2002) 28:9–25. doi: 10.1023/A:1025329728269
4. Rodríguez-Vivas RI, Grisi L, Pérez de León AA, Silva-Villela H, Torres-Acosta JFJ, Fragoso-Sánchez H, et al. Potential economic impact assessment for cattle parasites in Mexico. Rev Mex Cienc Pecu. (2017) 8:61–74. doi: 10.22319/rmcp.v8i1.4305
5. Reinbold JB, Coetzee JF, Hollis LC, Nickell JS, Riegel C, Olson KC, et al. The efficacy of three chlortetracycline regimens in the treatment of persistent Anaplasma marginale infection. Vet Microbiol. (2010) 145:69–75. doi: 10.1016/j.vetmic.2010.02.031
6. Primo ME, Sarli M, Thompson CS, Torioni SM, Echaide IE. Sensitivity of Anaplasma marginale genotypes to oxytetracycline assessed by analyzing the msp1α gene in experimentally infected cattle. Ticks Tick Borne Dis. (2021) 12:101787. doi: 10.1016/j.ttbdis.2021.101787
7. Reinbold JB, Coetzee JF, Hollis LC, Nickell JS, Riegel CM, Christopher JA, et al. Comparison of iatrogenic transmission of Anaplasma marginale in Holstein steers via needle and needle-free injection techniques. Am J Vet Res. (2010) 71:1178–88. doi: 10.2460/ajvr.71.10.1178
8. Brayton KA, Kappmeyer LS, Herndon DR, Dark MJ, Tibbals DL, Palmer GH, et al. Complete genome sequencing of Anaplasma marginale reveals that the surface is skewed to two superfamilies of outer membrane proteins. Proc Natl Acad Sci USA. (2005) 102:844–49. doi: 10.1073/pnas.0406656102
9. Dark MJ, Herndon DR, Kappmeyer LS, Gonzales MP, Nordeen E, Palmer GH, et al. Conservation in the face of diversity: multistrain analysis of an intracellular bacterium. BMC Genomics. (2009) 10:16. doi: 10.1186/1471-2164-10-16
10. Palmer GH, Bankhead T, Lukehart SA. 'Nothing is permanent but change'- antigenic variation in persistent bacterial pathogens. Cell Microbiol. (2009) 11:1697–705. doi: 10.1111/j.1462-5822.2009.01366.x
11. Callow LL. Some aspects of the epidemiology and control of bovine babesiosis in Australia. J S Afr Vet Assoc. (1979) 50:353–6.
12. Futse JE, Ueti MW, Knowles DP Jr, Palmer GH. Transmission of Anaplasma marginale by Boophilus microplus: retention of vector competence in the absence of vector-pathogen interaction. J Clin Microbiol. (2003) 41:3829–34. doi: 10.1128/JCM.41.8.3829-3834.2003
13. Amaro-Estrada I, García-Ortiz MA, Preciado de la Torre JF, Rojas-Ramírez EE, Hernández-Ortiz R, Alpírez-Mendoza F, et al. Transmission of Anaplasma marginale by unfed Rhipicephalus microplus tick larvae under experimental conditions. Rev Mex Cienc Pecu, (2020) 11:116–31. doi: 10.22319/rmcp.v11i1.5018
14. Dorneles EM, Sriranganathan N, Lage AP. Recent advances in Brucella abortus vaccines. Vet Res. (2015) 46:76. doi: 10.1186/s13567-015-0199-7
15. Rodriguez-Martins KR, Garcia MV, Bonatte-Junior P, Duarte PO, de Higa LOS, Csordas BG, et al. Correlation between Rhipicephalus microplus ticks and Anaplasma marginale infection in various cattle breeds in Brazil. Exp Appl Acarol. (2020) 81:585–98. doi: 10.1007/s10493-020-00514-1
16. Jonsson NN, Bock RE, Jorgensen WK, Morton JM, Stear MJ. Is endemic stability of tick-borne disease in cattle a useful concept? Trends Parasitol. (2012) 28:85–9. doi: 10.1016/j.pt.2011.12.002
17. McGarey DJ, Allred DR. Characterization of hemagglutinating components on the Anaplasma marginale initial body surface and identification of possible adhesins. Infect Immun. (1994) 62:4587–93. doi: 10.1128/iai.62.10.4587-4593.1994
18. Palmer GH, Rurangirwa FR, Kocan KM, Brown WC. Molecular basis for vaccine development against the ehrlichial pathogen Anaplasma marginale. Parasitol Today. (1999) 15:281–6. doi: 10.1016/S0169-4758(99)01469-6
19. Cantor GH, Pontzer CH, Palmer GH. Opsonization of Anaplasma marginale mediated by bovine antibody against surface protein MSP-1. Vet Immunol Immunopathol. (1993) 37:343–50. doi: 10.1016/0165-2427(93)90206-J
20. Estes DM, Closser NM, Allen GK. IFN-gamma stimulates IgG2 production from bovine B cells costimulated with anti-mu and mitogen. Cell Immunol. (1994) 154:287–95. doi: 10.1006/cimm.1994.1078
21. Palmer GH, McGuire TC. Immune serum against Anaplasma marginale initial bodies neutralizes infectivity for cattle. J Immunol. (1984) 133:1010–5.
22. Brown WC, Zhu D, Shkap V, McGuire TC, Blouin EF, Kocan KM, et al. The repertoire of Anaplasma marginale antigens recognized by CD4(+) T-lymphocyte clones from protectively immunized cattle is diverse and includes major surface protein 2 (MSP-2) and MSP-3. Infect Immun. (1998) 66:5414–22. doi: 10.1128/IAI.66.11.5414-5422.1998
23. Lopez JE, Palmer GH, Brayton KA, Dark MJ, Leach SE, Brown WC. Immunogenicity of Anaplasma marginale type IV secretion system proteins in a protective outer membrane vaccine. Infect Immun. (2007) 75:2333–42. doi: 10.1128/IAI.00061-07
24. Macmillan H, Norimine J, Brayton KA, Palmer GH, Brown WC. Physical linkage of naturally complexed bacterial outer membrane proteins enhances immunogenicity. Infect Immun. (2008) 76:1223–9. doi: 10.1128/IAI.01356-07
25. Brown WC, Shkap V, Zhu D, McGuire TC, Tuo W, McElwain TF, et al. CD4(+) T-lymphocyte and immunoglobulin G2 responses in calves immunized with Anaplasma marginale outer membranes and protected against homologous challenge. Infect Immun. (1998) 66:5406–13. doi: 10.1128/IAI.66.11.5406-5413.1998
26. Barigye R, Garcia-Ortiz MA, Rojas Ramírez EE, Rodriguez SD. Identification of IgG2 specific antigens in three Mexican strains of Anaplasma marginale. Ann N Y Acad Sci. (2004) 1026:84–94. doi: 10.1196/annals.1307.011
27. Brown WC, Palmer GH, Lewin HA, McGuire TC. CD4(+) T lymphocytes from calves immunized with Anaplasma marginale major surface protein 1 (MSP1), a heteromeric complex of MSP1a and MSP1b, preferentially recognize the MSP1a carboxyl terminus that is conserved among strains. Infect Immun. (2001) 69:6853–62. doi: 10.1128/IAI.69.11.6853-6862.2001
28. Battilani M, De Arcangeli S, Balboni A, Dondi F. Genetic diversity and molecular epidemiology of Anaplasma. Infect Genet Evol. (2017) 49:195–211. doi: 10.1016/j.meegid.2017.01.021
29. Allred DR, McGuire TC, Palmer GH, Leib SR, Harkins TM, McElwain TF, et al. Molecular basis for surface antigen size polymorphisms and conservation of a neutralization-sensitive epitope in Anaplasma marginale. Proc Natl Acad Sci USA. (1990) 87:3220–24. doi: 10.1073/pnas.87.8.3220
30. Aktas M, Özübek S. Outbreak of anaplasmosis associated with novel genetic variants of Anaplasma marginale in a dairy cattle. Comp Immunol Microbiol Infect Dis. (2017) 54:20–6. doi: 10.1016/j.cimid.2017.07.008
31. Hove P, Chaisi ME, Brayton KA, Ganesan H, Catanese HN, Mtshali MS, et al. Co-infections with multiple genotypes of Anaplasma marginale in cattle indicate pathogen diversity. Parasit Vectors. (2018) 11:5. doi: 10.1186/s13071-017-2595-5
32. Yang J, Han R, Liu Z, Niu Q, Guan G, Liu G, Luo J, Yin H. Insight into the genetic diversity of Anaplasma marginale in cattle from ten provinces of China. Parasit Vectors. (2017) 10:565. doi: 10.1186/s13071-017-2485-x
33. Garcia AB, Jusi MMG, Freschi CR, Ramos IAS, Mendes NS, Bressianini do Amaral R, et al. High genetic diversity and superinfection by Anaplasma marginale strains in naturally infected Angus beef cattle during a clinical anaplasmosis outbreak in southeastern Brazil. Ticks Tick Borne Dis. (2021) 13:101829. doi: 10.1016/j.ttbdis.2021.101829
34. Castañeda-Ortiz EJ, Ueti MW, Camacho-Nuez M, Mosqueda JJ, Mousel MR, Johnson WC, et al. Association of Anaplasma marginale strain superinfection with infection prevalence within tropical regions. PLoS ONE. (2015) 10:e0120748. doi: 10.1371/journal.pone.0120748
35. Barbet AF, Allred DR. The msp1 beta multigene family of Anaplasma marginale: nucleotide sequence analysis of an expressed copy. Infect Immun. (1991) 59:971–6. doi: 10.1128/iai.59.3.971-976.1991
36. Oberle SM, Palmer GH, Barbet AF. Expression and immune recognition of the conserved MSP4 outer membrane protein of Anaplasma marginale. Infect Immun. (1993) 61:5245–51. doi: 10.1128/iai.61.12.5245-5251.1993
37. Visser ES, McGuire TC, Palmer GH, Davis WC, Shkap V, Pipano E, et al. The Anaplasma marginale msp5 gene encodes a 19-kilodalton protein conserved in all recognized Anaplasma species. Infect Immun. (1992) 60:5139–44. doi: 10.1128/iai.60.12.5139-5144.1992
38. Munodzana D, McElwain TF, Knowles DP, Palmer GH. Conformational dependence of Anaplasma marginale major surface protein 5 surface-exposed B-cell epitopes. Infect Immun. (1998) 66:2619–424. doi: 10.1128/IAI.66.6.2619-2624.1998
39. Alleman AR, Palmer GH, McGuire TC, McElwain TF, Perryman LE, Barbet AF. Anaplasma marginale major surface protein 3 is encoded by a polymorphic, multigene family. Infect Immun. (1997) 65:156–63. doi: 10.1128/iai.65.1.156-163.1997
40. French DM, McElwain TF, McGuire TC, Palmer GH. Expression of Anaplasma marginale major surface protein 2 variants during persistent cyclic rickettsemia. Infect Immun. (1998) 66:1200–7. doi: 10.1128/IAI.66.5.2400-2400.1998
41. Sarli M, Novoa MB, Mazzucco MN, Signorini ML, Echaide IE, de Echaide ST, et al. A vaccine using Anaplasma marginale subdominant type IV secretion system recombinant proteins was not protective against a virulent challenge. PLoS ONE. (2020) 15:e0229301. doi: 10.1371/journal.pone.0229301
42. Theiler A. Anaplasma marginale (Gen. and spec. nov.). The marginal points in the blood of cattle suffering from a specific disease. 1908 Rep Gov Vet Bact Transv. 1908 – 1909. p 7–29.
43. Potgieter FT. Epizootiology and control of anaplasmosis in south Africa. J S Afr Vet Assoc. (1979) 50:367–72.
44. de Wall DT. Anaplasmosis control and diagnosis in South Africa. Ann N Y Acad Sci. (2000) 916:474–83. doi: 10.1111/j.1749-6632.2000.tb05327.x
45. Rogers RJ, Shiels IA. Epidemiology and control of anaplasmosis in Australia. J S Afr Vet Assoc. (1979) 50:363–6.
46. Abdala AA, Pipano E, Aguirre DH, Gaido AB, Zurbriggen MA, Mangold AJ, et al. Frozen and fresh Anaplasma centrale vaccines in the protection of cattle against Anaplasma marginale infection. Rev Elev Med Vet Pays Trop. (1990) 43:155–8. doi: 10.19182/remvt.8834
47. Vidotto O, Barbosa CS, Andrade GM, Machado RZ, da Rocha MA, Silva SS. Evaluation of a frozen trivalent attenuated vaccine against Babesiosis and anaplasmosis in Brazil. Ann N Y Acad Sci. (1998) 849:420–3. doi: 10.1111/j.1749-6632.1998.tb11086.x
48. Miraballes C, Aráoz V, Riet-Correa F. Rhipicephalus microplus, babesiosis and anaplasmosis in Uruguay: current situation and control or elimination programs on farms. Exp Appl Acarol. (2019) 78:579–93. doi: 10.1007/s10493-019-00405-0
49. Carelli G, Decaro N, Lorusso E, Paradies P, Elia G, Martella V, et al. First report of bovine anaplasmosis caused by Anaplasma centrale in Europe. Ann N Y Acad Sci. (2008) 1149:107–10. doi: 10.1196/annals.1428.069
50. Todorovic RA, Lopez LA, Lopez AG, Gonzalez EF. Bovine babesiosis and anaplasmosis: control by premunition and chemoprophylaxis. Exp Parasitol. (1975) 37:92–104. doi: 10.1016/0014-4894(75)90056-9
51. Uilenberg G. Notes sur les babésioses et l'anaplasmose des bovins á Madagascar. VI. Prémunition artificielle [Notes on bovine babesiosis and anaplasmosis in Madagascar. VI. Artificial premunization]. Rev Elev Med Vet Pays Trop. (1971) 24:23–35. French. doi: 10.19182/remvt.7762
52. Kuttler KL. Current anaplasmosis control techniques in the United States. J S Afr Vet Assoc. (1979) 50:314–20.
53. Franklin TE, Huff JW. A Proposed method of premunizing cattle with minimum inocula of Anaplasma marginale. Res Vet Sci. (1967) 8: 415–18. doi: 10.1016/S0034-5288(18)34600-9
54. Ristic M, Carson CA. Methods of immunoprophylaxis against bovine anaplasmosis with emphasis on use of the attenuated Anaplasma marginale vaccine. Adv Exp Med Biol. (1977) 93:151–88. doi: 10.1007/978-1-4615-8855-9_10
55. Kuttler KL, Zaugg JL. Characteristics of an attenuated Anaplasma marginale of deer origin as an anaplasmosis vaccine. Trop Anim Health Prod May. (1988) 20:85–91.
56. Osorno BM, Solana P, Ristic M, Serrano E. Estudio de una vacuna atenuada de Anaplasma marginale en México. Tec Pecu Mex. (1973) 24:57–63.
57. Lora CA, Koechlin A. An attenuated Anaplasma marginale vaccine in Peru. Am J Vet Res. (1969) 30:1993–8.
58. Zaraza H, Kuttler KL. Comparative efficacy of different immunization systems against anaplasmosis. Trop Anim Health Prod. (1971) 3:77–82. doi: 10.1007/BF02356479
59. Kenneil R, Shkap V, Leibovich B, Zweygarth E, Pfister K, Ribeiro MF, et al. Cross-protection between geographically distinct Anaplasma marginale isolates appears to be constrained by limited antibody responses. Transbound Emerg Dis. (2013) 60(Suppl. 2):97–104. doi: 10.1111/tbed.12125
60. Bock RE, deVos AJ, Kingston TG, Carter PD. Assessment of a low virulence Australian isolate of Anaplasma marginale for pathogenicity, immunogenicity and transmissibility by Boophilus microplus. Vet Parasitol. (2003) 118:121–31. doi: 10.1016/j.vetpar.2003.08.011
61. Rodríguez Camarillo SD, García Ortiz MA, Rojas Ramírez EE, Cantó Alarcón GJ, Preciado de la Torre JF, Rosario Cruz R, et al. Anaplasma marginale Yucatan (Mexico) Strain. Assessment of low virulence and potential use as a live vaccine. Ann N Y Acad Sci. (2008) 1149:98–102. doi: 10.1196/annals.1428.067
62. Silvestre BT, Silveira JAG, Meneses RM, Facury-Filho EJ, Carvalho AU, Ribeiro MFB. Identification of a vertically transmitted strain from Anaplasma marginale (UFMG3): molecular and phylogenetic characterization, and evaluation of virulence. Ticks Tick Borne Dis. (2016) 7:80–4. doi: 10.1016/j.ttbdis.2015.09.001
63. Rogers RJ, Dimmock CK, de Vos AJ, Rodwell BJ. Bovine leucosis virus contamination of a vaccine produced in vivo against bovine babesiosis and anaplasmosis. Aust Vet J. (1988) 65:285–7. doi: 10.1111/j.1751-0813.1988.tb16144.x
64. Marble DW, Hanks MA. A tissue culture method for Anaplasma marginale. Cornell Vet. (1972) 62:196–205.
65. Kessler RH, Ristic M. In vitro cultivation of Anaplasma marginale: invasion of and development in noninfected erythrocytes. Am J Vet Res. (1979) 40:1774–6.
66. Blouin EF, Kocan KM, Ewing SA. Preliminary attempts to infect bovine erythrocytes in vitro with a tick-derived stage of Anaplasma marginale. Ann N Y Acad Sci. (1992) 653:72–7. doi: 10.1111/j.1749-6632.1992.tb19631.x
67. Waghela SD, Cruz D, Droleskey RE, DeLoach JR, Wagner GG. In vitro cultivation of Anaplasma marginale in bovine erythrocytes co-cultured with endothelial cells. Vet Parasitol. (1997) 73:43–52. doi: 10.1016/S0304-4017(97)00045-9
68. Munderloh UG, Lynch MJ, Herron MJ, Palmer AT, Kurtti TJ, Nelson RD, Goodman JL. Infection of endothelial cells with Anaplasma marginale and A. phagocytophilum. Vet Microbiol. (2004) 101:53–64. doi: 10.1016/j.vetmic.2004.02.011
69. Barbet AF, Blentlinger R, Yi J, Lundgren AM, Blouin EF, Kocan KM. Comparison of surface proteins of Anaplasma marginale grown in tick cell culture, tick salivary glands, and cattle. Infect Immun. (1999) 67:102–7. doi: 10.1128/IAI.67.1.102-107.1999
70. Lis K, Fernández de Mera IG, Popara M, Cabezas-Cruz A, Ayllón N, Zweygarth E, et al. Molecular and immunological characterization of three strains of Anaplasma marginale grown in cultured tick cells. Ticks Tick Borne Dis. (2015) 6:522–9. doi: 10.1016/j.ttbdis.2015.04.006
71. Bell-Sakyi L, Palomar AM, Bradford EL, Shkap V. Propagation of the Israeli vaccine strain of Anaplasma centrale in tick cell lines. Vet Microbiol. (2015) 179:270–6. doi: 10.1016/j.vetmic.2015.07.008
72. Lasmar PV, Carvalho AÚ, Facury Filho EJ, Bastos CV, Ribeiro MF. Evaluating the effectiveness of an inactivated vaccine from Anaplasma marginale derived from tick cell culture. Rev Bras Parasitol Vet. (2012) 21:112–7. doi: 10.1590/S1984-29612012000200008
73. de la Fuente J, Kocan KM, Garcia-Garcia JC, Blouin EF, Claypool PL, Saliki JT. Vaccination of cattle with Anaplasma marginale derived from tick cell culture and bovine erythrocytes followed by challenge-exposure with infected ticks. Vet Microbiol. (2002) 89:239–51. doi: 10.1016/S0378-1135(02)00206-7
74. Baêta BA, Ribeiro CC, Teixeira RC, Cabezas-Cruz A, Passos LM, Zweygarth E, et al. Characterization of two strains of Anaplasma marginale isolated from cattle in Rio de Janeiro, Brazil, after propagation in tick cell culture. Ticks Tick Borne Dis. (2015) 6:141–5. doi: 10.1016/j.ttbdis.2014.11.003
75. Cobaxin-Cárdenas M, Aguilar Díaz H, Olivares Avelino P, Salinas-Estrella E, Preciado-de la Torre J, Quiroz-Castañeda R, et al. Primer abordaje para la propagación de Anaplasma marginale (MEX-31–096) en células de garrapata Rm-sus. Rev Cent Investig Univ Salle. (2019) 13:67–80. doi: 10.26457/recein.v13i51.2200
76. Montenegro-James S, James MA, Benitez MT, Leon E, Baek BK, Guillen AT. Efficacy of purified Anaplasma marginale initial bodies as a vaccine against anaplasmosis. Parasitol Res. (1991) 77:93–101. doi: 10.1007/BF00935421
77. Wilson JS, Trace JC. A new inactivated vaccine for bovine anaplasmosis. Bulletin. (1966) 66:897–902.
78. Rodríguez SD, García Ortiz MA, Hernández Salgado G, Santos Cerda NA, Aboytes Torre R, Cantó Alarcón GJ. Anaplasma marginale inactivated vaccine: dose titration against a homologous challenge. Comp Immunol Microbiol Infect Dis. (2000) 23:239–52. doi: 10.1016/S0147-9571(99)00076-4
79. Ocampo-Espinoza V, Salazar Vázquez JE, Durán Aguilar M, García Ortiz MA, Cantó Alarcón GJ, Rodríguez SD. Anaplasma marginale: lack of cross-protection between strains that share MSP1a variable region and MSP4. Vet Micro. (2006) 114:34–40. doi: 10.1016/j.vetmic.2005.11.058
80. Orozco-Vega LE, Rodriguez SD, Alarcon GJ, Flores RL, Ocampo RJ, Ortiz MA, et al. Anaplasma marginale field challenge: protection by an inactivated immunogen that shares partial sequence of msp1alpha variable region with the challenge strain. Vaccine. (2007) 25:519–25. doi: 10.1016/j.vaccine.2006.07.049
81. McCorkle-Shirley S, Hart LT, Larson AD, Todd WJ, Myhand JD. High-yield preparation of purified Anaplasma marginale from infected bovine red blood cells. Am J Vet Res. (1985) 46:1745–7.
82. Luther DG, Hart LT, Todd WJ, Morris NG, Taylor ND, McRae BS. Field study of an experimental anaplasmosis vaccine on pregnant cows and neonatal isoerythrolysis. In: Proc, Eighth Natl Vet Hemoparasite Dis Conf. St. Louis, MO: Anaplasmosis, Babesiosis Network. US Agency for International Development. (1989). p. 559–62.
83. Dark MJ, Lundgren AM, Barbet AF. Determining the repertoire of immunodominant proteins via whole-genome amplification of intracellular pathogens. PLoS ONE. (2012) 7:e36456. doi: 10.1371/journal.pone.0036456
84. McGuire TC, Palmer GH, Goff WL, Johnson MI, Davis WC. Common and isolate-restricted antigens of Anaplasma marginale detected with monoclonal antibodies. Infect Immun. (1984) 45:697–700. doi: 10.1128/iai.45.3.697-700.1984
85. McGuire TC, Stephens EB, Palmer GH, McElwain TF, Lichtensteiger CA, Leib SR, Barbet AF. Recombinant vaccinia virus expression of Anaplasma marginale surface protein MSP-1a: effect of promoters, leader sequences and GPI anchor sequence on antibody response. Vaccine. (1994) 12:465–71. doi: 10.1016/0264-410X(94)90126-0
86. Mwangi W, Brown WC, Splitter GA, Davies CJ, Howard CJ, Hope JC, et al. DNA vaccine construct incorporating intercellular trafficking and intracellular targeting motifs effectively primes and induces memory B- and T-cell responses in outbred animals. Clin Vaccine Immunol. (2007) 14:304–11. doi: 10.1128/CVI.00363-06
87. Kano FS, Tamekuni K, Coelho AL, Garcia JL, Vidotto O, Itano EN, et al. Induced immune response of DNA vaccine encoding an association MSP1a, MSP1b, and MSP5 antigens of Anaplasma marginale. Vaccine. (2008) 26:3522–7. doi: 10.1016/j.vaccine.2008.04.047
88. Zhang B, Cavallaro AS, Mody KT, Zhang J, Deringer JR, Brown WC, Mahony TJ, Yu C, Mitter N. Nanoparticle-based delivery of Anaplasma marginale membrane proteins; VirB9–1 and VirB10 produced in the Pichia pastoris expression system. Nanomaterials. (2016) 6:201. doi: 10.3390/nano6110201
89. Zhao L, Mahony D, Cavallaro AS, Zhang B, Zhang J, Deringer JR, et al. Immunogenicity of outer membrane proteins VirB9–1 and VirB9–2, a novel nanovaccine against Anaplasma marginale. PLoS ONE. (2016) 11:e0154295. doi: 10.1371/journal.pone.0154295
90. Ducken DR, Brown WC, Alperin DC, Brayton KA, Reif KE, Turse JE, et al. Subdominant outer membrane antigens in Anaplasma marginale: conservation, antigenicity, and protective capacity using recombinant protein. PLoS ONE. (2015) 10:e0129309. doi: 10.1371/journal.pone.0129309
91. Hammac GK, Ku PS, Galletti MF, Noh SM, Scoles GA, Palmer GH, et al. Protective immunity induced by immunization with a live, cultured Anaplasma marginale strain. Vaccine. (2013) 31:3617–22. doi: 10.1016/j.vaccine.2013.04.069
92. de Andrade GM, Machado RZ, Vidotto MC, Vidotto O. Immunization of bovines using a DNA vaccine (pcDNA3.1/MSP1b) prepared from the Jaboticabal strain of Anaplasma marginale. Ann N Y Acad Sci. (2004) 1026:257–66. doi: 10.1196/annals.1307.040
93. Watthanadirek A, Chawengkirttikul R, Poolsawat N, Junsiri W, Boonmekam D, Reamtong O, et al. Recombinant expression and characterization of major surface protein 4 from Anaplasma marginale. Acta Trop. (2019) 197:105047. doi: 10.1016/j.actatropica.2019.105047
94. Palacios C, Torioni de Echaide S, Mattion N. Evaluation of the immune response to Anaplasma marginale MSP5 protein using a HSV-1 amplicon vector system or recombinant protein. Res Vet Sci. (2014) 97:514–20. doi: 10.1016/j.rvsc.2014.10.005
95. Watthanadirek A, Junsiri W, Minsakorn S, Poolsawat N, Srionrod N, Khumpim P, et al. Molecular and recombinant characterization of major surface protein 5 from Anaplasma marginale. Acta Trop. (2021) 220:105933. doi: 10.1016/j.actatropica.2021.105933
96. Kawasaki PM, Kano FS, Tamekuni K, Garcia JL, Marana ER, Vidotto O, et al. Immune response of BALB/c mouse immunized with recombinant MSPs proteins of Anaplasma marginale binding to immunostimulant complex (ISCOM). Res Vet Sci. (2007) 83:347–54. doi: 10.1016/j.rvsc.2007.02.002
97. Morse K, Norimine J, Palmer GH, Sutten EL, Baszler TV, Brown WC. Association and evidence for linked recognition of type IV secretion system proteins VirB9–1, VirB9–2, and VirB10 in Anaplasma marginale. Infect Immun. (2012) 80:215–27. doi: 10.1128/IAI.05798-11
98. Morse K, Norimine J, Hope JC, Brown WC. Breadth of the CD4+ T cell response to Anaplasma marginale VirB9–1, VirB9–2 and VirB10 and MHC class II DR and DQ restriction elements. Immunogenetics. (2012) 64:507–23. doi: 10.1007/s00251-012-0606-4
99. Hebert KS, Seidman D, Oki AT, Izac J, Emani S, Oliver LD Jr, et al. Anaplasma marginale Outer Membrane Protein A Is an adhesin that recognizes sialylated and fucosylated glycans and functionally depends on an essential binding domain. Infect Immun. (2017) 85:e00968-16. doi: 10.1128/IAI.00968-16
100. Futse JE, Buami G, Kayang BB, Koku R, Palmer GH, Graça T, et al. Sequence and immunologic conservation of Anaplasma marginale OmpA within strains from Ghana as compared to the predominant OmpA variant. PLoS ONE. (2019) 14:e0217661. doi: 10.1371/journal.pone.0217661
101. Belkahia H, Ben Abdallah M, Andolsi R, Selmi R, Zamiti S, Kratou M, et al. Screening and Analysis of Anaplasma marginale tunisian isolates reveal the diversity of lipA phylogeographic marker and the conservation of OmpA protein vaccine candidate. Front Vet Sci. (2021) 8:731200. doi: 10.3389/fvets.2021.731200
102. Silvestre BT, Silveira JAGD, Facury-Filho EJ, Carvalho AÚ, Versiani AF, Estevam LGTM, Araújo MSS, Martins-Filho OA, Negrão-Corrêa DA, Ribeiro MFB. Immune response and biochemistry of calves immunized with rMSP1a (Anaplasma marginale) using carbon nanotubes as carrier molecules. Rev Bras Parasitol Vet. (2018) 27:191–202. doi: 10.1590/s1984-296120180029
103. Hove P, Brayton KA, Liebenberg J, Pretorius A, Oosthuizen MC, Noh SM, et al. Anaplasma marginale outer membrane protein vaccine candidates are conserved in North American and South African strains. Ticks Tick Borne Dis. (2020) 11:101444. doi: 10.1016/j.ttbdis.2020.101444
104. Albarrak SM, Brown WC, Noh SM, Reif KE, Scoles GA, Turse JE, et al. Subdominant antigens in bacterial vaccines: AM779 is subdominant in the Anaplasma marginale outer membrane vaccine but does not associate with protective immunity. PLoS ONE. (2012) 7:e46372. doi: 10.1371/journal.pone.0046372
105. Curtis AK, Reif KE, Kleinhenz MD, Martin MS, Skinner B, Kelly SM, et al. Development of a subcutaneous ear implant to deliver an anaplasmosis vaccine to dairy steers. J Anim Sci. (2020) 98:skz392. doi: 10.1093/jas/skaa054.275
106. Crosby FL, Brayton KA, Magunda F, Munderloh UG, Kelley KL, Barbet AF. Reduced Infectivity in cattle for an outer membrane protein mutant of Anaplasma marginale. Appl Environ Microbiol. (2015) 81:2206–14. doi: 10.1128/AEM.03241-14
107. Felsheim RF, Chávez AS, Palmer GH, Crosby L, Barbet AF, Kurtti TJ, et al. Transformation of Anaplasma marginale. Vet Parasitol. (2010) 167:167–74. doi: 10.1016/j.vetpar.2009.09.018
108. Kauffmann F, Van Damme P, Leroux-Roels G, Vandermeulen C, Berthels N, Beuneu C, et al. Clinical trials with GMO-containing vaccines in Europe: status and regulatory framework. Vaccine. (2019) 37:6144–53. doi: 10.1016/j.vaccine.2019.08.018
109. Crosby FL, Wamsley HL, Pate MG, Lundgren AM, Noh SM, Munderloh UG, et al. Knockout of an outer membrane protein operon of Anaplasma marginale by transposon mutagenesis. BMC Genomics. (2014) 15:278. doi: 10.1186/1471-2164-15-278
110. Silvestre BT, Rabelo ÉM, Versiani AF, da Fonseca FG, Silveira JA, Bueno LL, et al. Evaluation of humoral and cellular immune response of BALB/c mice immunized with a recombinant fragment of MSP1a from Anaplasma marginale using carbon nanotubes as a carrier molecule. Vaccine. (2014) 32:2160–6. doi: 10.1016/j.vaccine.2014.02.062
111. Hope M, Riding G, Menzies M, Willadsen P. A novel antigen from Anaplasma marginale: characterization, expression and preliminary evaluation of the recombinant protein. Vaccine. (2004) 22:407–15. doi: 10.1016/j.vaccine.2003.07.006
Keywords: Anaplasma marginale, immunity, bovine anaplasmosis, inactivated vaccine, live vaccines
Citation: Salinas-Estrella E, Amaro-Estrada I, Cobaxin-Cárdenas ME, Preciado de la Torre JF and Rodríguez SD (2022) Bovine Anaplasmosis: Will there ever be an almighty effective vaccine? Front. Vet. Sci. 9:946545. doi: 10.3389/fvets.2022.946545
Received: 17 May 2022; Accepted: 11 August 2022;
Published: 05 October 2022.
Edited by:
Hua-Ji Qiu, Harbin Veterinary Research Institute (CAAS), ChinaReviewed by:
Alejandro Cabezas-Cruz, Institut national de recherche pour l'agriculture, l'alimentation et l'environnement (INRAE), FranceCopyright © 2022 Salinas-Estrella, Amaro-Estrada, Cobaxin-Cárdenas, Preciado de la Torre and Rodríguez. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sergio D. Rodríguez, cm9kcmlndWV6LnNlcmdpb0BpbmlmYXAuZ29iLm14; c2VyZ2VpeWV2c2t5QHlhaG9vLmNvbQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.