 Carolina Menezes Suassuna de Souza1,2*
Carolina Menezes Suassuna de Souza1,2* Ewerton de Souza Lima1,2
Ewerton de Souza Lima1,2 Raphael Ferreira Ordonho1,2Bianca Rafaella Rodrigues dos Santos Oliveira2Rebeca Cordeiro Rodrigues2Marquiliano Farias de Moura3Daniel Magalhães Lima4
Raphael Ferreira Ordonho1,2Bianca Rafaella Rodrigues dos Santos Oliveira2Rebeca Cordeiro Rodrigues2Marquiliano Farias de Moura3Daniel Magalhães Lima4 Maiara Garcia Blagitz5,6
Maiara Garcia Blagitz5,6 Eduardo Milton Ramos Sanchez1,7,8
Eduardo Milton Ramos Sanchez1,7,8 Isac Almeida de Medeiros9
Isac Almeida de Medeiros9 Fernando Nogueira Souza1,2,6†
Fernando Nogueira Souza1,2,6† Artur Cezar de Carvalho Fernandes1,2†
Artur Cezar de Carvalho Fernandes1,2†- 1Programa de Pós-Graduação em Ciência Animal, Centro de Ciências Agrárias, Universidade Federal da Paraíba, Areia, Brazil
- 2Núcleo Aplicado à Produção e Sanidade da Glândula Mamária (NAPROSA), Departamento de Ciências Veterinárias, Centro de Ciências Agrárias, Universidade Federal da Paraíba, Areia, Brazil
- 3Departamento de Ciências Veterinárias, Centro de Ciências Agrárias, Universidade Federal da Paraíba, Areia, Brazil
- 4Departamento de Medicina Veterinária Preventiva e Saúde Animal, Faculdade de Medicina Veterinária e Zootecnia, Universidade de São Paulo, São Paulo, Brazil
- 5Programa de Pós-Graduação em Saúde, Bem-Estar e Produção Animal Sustentável na Fronteira Sul, Universidade Federal da Fronteira Sul, Realeza, Brazil
- 6Veterinary Clinical Immunology Research Group, Departamento de Clínica Médica, Faculdade de Medicina Veterinária e Zootecnia, Universidade de S3ão Paulo, São Paulo, Brazil
- 7Department of Public Health, School of Health Sciences, National University Toribio Rodriguez de Mendoza of Amazonas, Chachapoyas, Peru
- 8Laboratório de Sorologia e Imunobiologia, Instituto de Medicina Tropical, Universidade de São Paulo, São Paulo, Brazil
- 9Departamento de Ciências Farmacêuticas, Centro de Ciências da Saúde, Universidade Federal da Paraíba, João Pessoa, Brazil
The present study sought to evaluate the expression of PD-1 and CTLA-4 in blood T lymphocytes during the periparturient period and their relationship with uterine health in dairy cows, as determined by endometrial cytology and serum concentrations of β-hydroxybutyrate (BHB) and non-esterified fatty acids (NEFAs), which are indicators of a negative energy balance. The second objective of this study was to investigate whether the expression of PD-1 and CTLA-4 in T lymphocytes is associated with the serum acute phase-protein haptoglobin concentration during the periparturient period. To address these objectives, 26 clinically healthy dairy cows were used. Peripheral blood was collected 14 days prepartum (T-14), at calving (T0), and 30 days postpartum (T30) to measure the expression of PD-1 and CTLA-4 in blood T lymphocytes by flow cytometry. In addition, we collected blood at T0, 10 days after parturition (T10), and T30 to obtain serum and determine the serum concentrations of NEFA, BHB, and Hp. Endometrial cytology was performed at T10, 20 days after parturition (T20), and T30. In the present study, we observed higher expression of CTLA-4 and PD-1 in T lymphocytes at parturition and in the prepartum period, which could indicate a relationship between these immune checkpoints and immunological tolerance during gestation in dairy cattle. In addition, a negative association between the expression of these immune checkpoints prepartum or at parturition and endometrial cytology at T20 and T30 was observed, indicating the negative implications of these immune response regulators in susceptibility to infections. This finding was further corroborated by the relationship between the serum concentration of haptoglobin and the expression of CTLA-4 and PD-1 by T lymphocytes. However, we did not observe a relationship between the indicators of negative energy balance, evaluated by the serum concentrations of BHB and NEFA, and the expression of the immune checkpoint markers studied. Thus, our findings represent an initial step that paves the way for the development of new therapeutic alternatives directed by the host with the objective of increasing the resistance of dairy cattle to infections in this critical period of life.
Introduction
The transition period in dairy cows between the 3 weeks preceding calving and 3 weeks after calving is characterized by drastic metabolic and immune response changes. During this period, dairy cows are usually in a state of negative energy balance that results in substantial lipid mobilization, a drop in glucose levels, and a dramatic rise in nonesterified fatty acid (NEFA) and β-hydroxybutyrate (BHBA) levels, which impairs immune cell functions (1, 2). Thus, dairy cows face the highest incidence of infectious diseases and metabolic disorders during this period (3, 4). For instance, dairy cows are often affected by endometritis during the postpartum period, which has a substantial negative impact on subsequent reproductive performance (5, 6). This condition is typically detected through endometrial cytology assessment and is referred to as cytological endometritis (6).
Although most dairy cows are subject to bacterial contamination of the uterus during the postpartum period, only a fraction of these animals develop the disease. However, the mechanisms that determine the process by which some animals resolve uterine infection, while others resist it, are not yet fully understood (7). In this context, the investigation of immune checkpoint inhibitors, particularly programmed cell death protein 1 (PD-1) and cytotoxic T lymphocyte-associated antigen-4 (CTLA-4), has gained prominence in recent years, as these molecules have been linked to immunosuppression, disease progression, and a poor prognosis (8, 9), including in bovine infectious diseases (10–12). These molecules, PD1 and CTLA-4, are vital in maintaining self-tolerance and modulating the magnitude and duration of effector immune responses to prevent collateral tissue damage. Signaling by these molecules may induce 'exhaustion' in effector immunological cells (particularly T lymphocytes). T-cell exhaustion is characterized by decreased effector function, persistent expression of immunological checkpoint molecules, poor recall responses, and a transcriptional state that differs from that of functional effector or memory T cells. Thus, there is emerging evidence that several pathogens promote inhibitory interactions between immune cells through immune checkpoint proteins to evade the host immune system by exploiting immunological tolerance and restricting immune-mediated pathogen clearance (8). Investigations into these immunosuppressive interactions have the potential to develop clinical strategies for treating and controlling veterinary infectious diseases by boosting the body's natural immune response to fight diseases, similar to how antibodies that block these immune checkpoints have been used to treat and control many types of malignancies (8, 13).
However, the relationships between these immune checkpoints and the transition period in dairy cows and uterine health in the postpartum period in dairy cows have not yet been studied. Thus, the present study sought to evaluate the expression of PD-1 and CTLA-4 in blood T lymphocytes during the periparturient period and their relationship with uterine health, as determined by endometrial cytology, and serum concentrations of β-hydroxybutyrate (BHB) and non-esterified fatty acids (NEFAs), widely used indicators of negative energy balance.
Furthermore, the measurement of acute-phase proteins is widely used in the diagnosis of diseases in ruminants and is an important indicator of inflammatory processes in cattle (14), in which haptoglobin (Hp) stands out for its high sensitivity and specificity and ability to discriminate between acute and chronic infections (15). Therefore, Hp is the most studied acute-phase protein and is used to detect cows with a high risk of systemic disease and severe postpartum inflammation (16). Hence, the second objective of the present study was to investigate whether the expression of PD-1 and CTLA-4 in blood T lymphocytes is associated with the serum Hp concentration during the periparturient period.
Materials and Methods
Animals and Experimental Design
For the present study, 26 clinically healthy dairy cows, including 19 multiparous cows (between their 2nd and 5th lactation; 3.26 ± 0.21) and 7 primiparous cows from two dairy farms, were used. Among these animals, 16 animals of the Guzerá breed (Farm A) and 10 animals of the Girolando breed, composed of the crossbreeding of Holstein and Gir cattle (Farm B), were used. At the first sampling, only healthy cows were enrolled, with no detectable clinical illness. Peripheral blood was collected 14 days prepartum (T-14), at calving (T0), and 30 days postpartum (T30) to measure the expression of PD-1 and CTLA-4 in blood T lymphocytes. In addition, we collected blood at T0, 10 days after parturition (T10), and T30 to obtain serum and determine the serum concentrations of NEFA, BHB, and the acute-phase protein haptoglobin. Endometrial cytology was performed at T10, 20, days after parturition (T20) and T30.
In herd A, the Guzerá cows were kept on Massai grass (a spontaneous hybrid between Panicum maximum and Panicum infestum) pasture and received soybean meal, corn meal, and cottonseed meal and cake as the concentrates according to their milk production, as well as vitamins and mineral supplements. These zebu dairy cows were milked once a day by manual milking with the calf at foot and yielded an average of 20 kg milk/day, and the calves were maintained half of the day with their dams, as the high weight of the calves was prioritized due their high economic value. Both artificial insemination and natural mating were used in this herd. In herd B, the dairy cows were maintained part of the time on Mombaça grass (Panicum maximum) pasture (in the morning) and received palm as the roughage and soybean meal, corn meal, and cottonseed meal as the concentrates according to their milk production, as well as vitamins and mineral supplements. These dairy cows were milked twice daily using machine milking and yielded an average of 27 kg milk/day, and all cows were artificially inseminated in this herd.
Collection of Peripheral Blood Samples
The peripheral blood samples were collected aseptically by venipuncture of the jugular vein in vacutainer® tubes containing sodium heparin (T-14, T0, and T30; two tubes with 4 mL per sampling time; cat. no. 367871, BD Biosciences, New Jersey, USA) for isolation of peripheral venous blood mononuclear cells (PBMCs) and without anticoagulant (T0, T10, and T30; one tube with 4 mL per sampling time; cat. no. 367812, BD Biosciences, New Jersey, USA) to obtain blood serum. Blood serum was obtained by centrifugation for 10 min at 2,500 x g at room temperature.
Serum Concentrations of NEFA, BHB, and Haptoglobin
The serum concentrations of BHB and NEFA were analyzed with an automatic analyser (Randox Rx Daytona Chemistry Analyser™, Crumlin, UK) using Randox® commercial kits (Randox Laboratories, Crumlin, UK) for BHB (Randox, cat. n. RB 1007) and NEFA (Randox, cat. n. FA115). The cutoff value established to determine ketosis was 1.2 mmol/L for BHB and 0.8 mmol/L for NEFA, as previously described by Roberts et al. (17).
Furthermore, the serum concentrations of Hp were measured by means of a colorimetric procedure that measures the formation of haptoglobin–hemoglobin complexes and estimates the differences in peroxidase activity, as previously described (18).
Endometrial Cytology
To assess uterine health, endometrial cytology (EC) was performed with the aid of the Cytobrush® cervical brush (Kolplast, Itupeva, Brazil). Collection of uterine contents was performed as described by Martins et al. (19), which consisted of collecting samples using a human gynecological brush coupled to a universal semen applicator with a stainless steel rod, with protection by a French sheath and sanitary shirt. Then, after passing through the cervix and reaching the base of the uterine body, the sanitary liner was broken, and the applicator plunger was pushed forwards, exposing the brush, which was in contact with the endometrium, was rotated clockwise, and then removed as described by Moura et al. (20). Subsequently, smears with light circular movements were performed on slides with the uterine content obtained with the Cytobrush® cervical brush (Kolplast, Itupeva, Brazil), and subsequently, the prepared slides were stained with panoptic fast stain for evaluation of cellularity by optical microscopy (Primo Star 1, Carl Zeiss, São Paulo, Brazil). The EC was determined by evaluating the percentage of neutrophils from 100 cells at 100X magnification. For the present study, animals with >18% neutrophils in the uterine cytology of the samples collected at T20 and T30 were considered to have a diagnosis of endometritis, as previously described based on examinations of EC in dairy cows between 20 and 33 days postpartum (21–23).
Expression of PD-1 and CTLA-4 in the T Lymphocyte Population
Initially, PBMCs were isolated by a Ficoll-Paque™ PLUS density gradient (GE Healthcare, Darmstadt, Germany) according to the manufacturer's recommendations. The PBMCs of each sample were transferred to two 5-mL round-bottom polypropylene tubes (A and B), 12 × 75 mm, suitable for flow cytometry, in which they were incubated for 30 min at room temperature with the primary antibodies. Tube A contained 1 μL of the primary mouse anti-bovine IgG1 antibody CD3 (clone MM1A, cat. N. BOV 2009, Washington State University Monoclonal Antibody Center, USA) and goat anti-human PD-1 with crossreaction with cattle (diluted 1/10 in PBS with 1% heat sterile inactivated fetal bovine serum and 0.09% sodium azide; cat. No. LS-C55247-100, LSBIO, USA), and tube B contained 1 μL of the primary mouse anti-bovine IgG1 CD3 (clone MM1A, cat. no. BOV 2009, Washington State University Monoclonal Antibody Center, USA) and goat anti-human CTLA-4 crossreactive with cattle (diluted 1/10 in PBS with 1% heat-inactivated sterile-filtered fetal bovine serum and 0.09% sodium azide; cat. n. AF-386-PB, R&D Systems, USA). After the incubation period, the cells were washed with PBS and centrifuged at 250 × g for 8 min at 4 °C, the supernatant was discarded, the cells were resuspended in 100 μL of PBS, and goat anti-mouse IgG1-conjugated secondary antibodies conjugated with PE-Texas Red (cat. No. M32017, Thermo Fisher, USA) and crossadsorbed donkey IgG conjugated with Alexa Fluor 488 (cat. No. A11055, Thermo Fisher, Carlsbad, USA) were added. The samples were incubated at room temperature for 30 min. After the incubation period, the cells were washed with PBS and centrifuged at 250 × g at 4 °C for 8 min. The supernatant was discarded, and the cells were resuspended in 300 μL of PBS with 1% heat-inactivated fetal bovine serum. The sample readings were performed using a BD FACSCanto™ II flow cytometer (BD Biosciences, New Jersey, USA). For this assay, 10,000 cells from each sample were examined. An unstained control, secondary antibody control, isotype control, and single-stained PBMC samples were also prepared as compensation controls. Flow Jo Tree Star software (FlowJo - Treestar 10.5.3 for Windows, Tree Star Inc., Ashland, OR, USA) was used to analyse the data. An example of our gate strategy for the determination of PD-1 expression in T lymphocytes is shown in Supplementary Figure 1. The relative geometric mean fluorescence intensity (GMFI) was chosen because it was far more discriminatory than the percentage of positive cells. The GMFI accurately measures the brightness of the stained cells and consequently indicates the number of receptors per cell (24).
Statistical Analysis
Data distribution was initially assessed using the Shapiro–Wilk test. The parametric distribution data were subjected to ANOVA followed by the Student–Newman–Keuls test to compare each variable analyzed at different time points. The variables with a non-parametric distribution were compared by the Kruskal–Wallis test, followed by Dunn's test, to verify significance (P ≤ 0.05), except when indicated. The correlations between the variables analyzed were examined by means of the Pearson and Spearman correlation tests for parametric and non-parametric data, respectively. The statistical software GraphPad Prism 9.0 was used (GraphPad software, Inc., San Diego, USA).
To investigate the association between the expression of CLTA-4 and PD-1 in T lymphocytes, with endometritis indicated as the percentage of neutrophils > 18% in EC at T20 and T30, generalized linear regression was used, with the dependent variable being disease (percentage of neutrophils > 18%) and explanatory variables being the sampling time, parity, and the farm/breed. All of these analyses were performed in the R program [R (25)] and the tidyverse (26) and broom (27) packages. The results are expressed as the mean ± standard error, and α = 5% was used for all analyses, except when indicated.
Results
Descriptive Analysis
In the present study, no significant differences were observed in the generalized linear regression model between multiparous and primiparous cows, with no effect of dairy farms/breeds on the variables analyzed. Out of 26 enrolled dairy cows, 16 (61.54%) and six (23.08%) dairy cows developed cytological endometritis at T20 and T30, respectively, i.e., showed a high percentage of neutrophils (>18%) in the endometrial cytology. The diagnosis of ketosis determined by the serum concentrations of BHB and NEFA was detected at T0 in one (3.85%) and seven (26.92%) dairy cows, at T10 in four (15.38%) and three (11.54%) dairy cows, and at T30 in two (7.69%) and two (7.69%) dairy cows, respectively.
Diagnosis of Ketosis and Its Relationship With the Expression of PD-1 and CTLA-4 by Blood T Lymphocytes
The analyzed time points did not affect the serum concentration of BHB (P = 0.45); however, the serum NEFA concentration was higher at T0 than T10 (P = 0.02; Supplementary Figure 2). In addition, high concentrations of BHB (≥ 1.2 mmol/L) and NEFA (≥ 0.8 mmol/L) were not associated with CTLA-4 or PD-1 expression in blood T lymphocytes in the generalized linear regression model.
Diagnosis of Cytological Endometritis and Its Relationship With the Expression of PD-1 and CTLA-4 by Blood T Lymphocytes
One notable result was the higher expression of CTLA-4 in T lymphocytes at T-14 (P = 0.0007) and T0 (P = 0.0004) than at T30 (Figure 1A). The expression of PD-1 in T lymphocytes at T-14 was also higher than that at T30 (P = 0.003; Figure 1B). The percentage of neutrophils in the EC at T10 (27.50 ± 5.90), T20 (30.31 ± 5.26), and T30 (14.65 ± 4.00) also did not differ among the analyzed time points (P = 0.12).
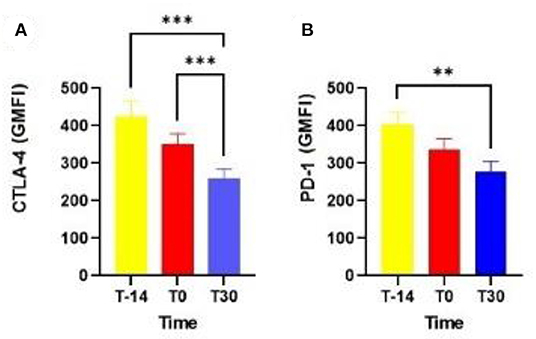
Figure 1. Expression of CTLA-4 (A) and PD-1 by blood T lymphocytes in dairy cows at T-14, T0, and T30 (B). CTLA-4: cytotoxic T lymphocyte-associated antigen-4; PD-1, programmed cell death protein 1; T-14 = 14 days before delivery; T0 = day of delivery; T30 = 30 days postpartum; GMFI, geometric mean fluorescence intensity. **P < 0.01; ***P < 0.0001.
In the generalized linear regression model, the diagnosis of cytological endometritis at T20 showed an association with the expression of PD-1 in T lymphocytes (P = 0.02) and a trend with the expression of CTLA-4 in T lymphocytes (P = 0.086) at parturition. In addition, dairy cows diagnosed with cytological endometritis (neutrophil percentage > 18 determined by EC) at T30 showed a higher expression of PD-1 by T lymphocytes at T0 (P = 0.009) as well as a tendency for a higher expression of PD-1 at T30 (P = 0.09; Figure 2).
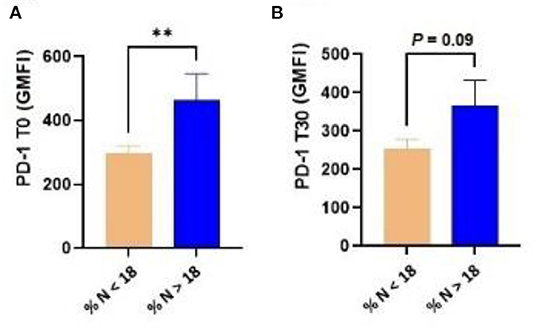
Figure 2. Dairy cows diagnosed with cytological endometritis [i.e., percentage of neutrophils higher than 18 (% N > 18)] at T30 had a higher expression of PD-1 in blood T lymphocytes at both T0 (A) and T30 (B). PD-1, programmed cell death protein 1; T0 = day of delivery; T30 = 30 days postpartum; GMFI, geometric mean fluorescence intensity. **P < 0.01.
The expression of CTLA-4 in blood T lymphocytes at T-14 was correlated with the percentage of neutrophils at T30 (r = 0.56; P = 0.01). Furthermore, at T0, the expression of CTLA-4 in T lymphocytes was again correlated with the percentage of neutrophils T30 (r = 0.50; P = 0.03). Similarly, at T0, the expression of PD-1 in T lymphocytes showed a significant correlation with the percentage of neutrophils at T30 (r = 0.62; P = 0.006).
Serum Concentration of Haptoglobin and Its Relationship With the Expression of PD-1 and CTLA-4 by Blood T Lymphocytes
The analyzed time points did not affect the serum concentration of haptoglobin (P = 0.38). The expression of CTLA-4 in blood T lymphocytes at T-14 was correlated with the serum concentration of haptoglobin at T30 (r = 0.68; P = 0.002). Furthermore, at T0, the expression of CTLA-4 in T lymphocytes was again correlated with the serum concentration of haptoglobin T30 (r = 0.66; P = 0.003). Additionally, at T-14, the expression of PD-1 in T lymphocytes was correlated with the serum concentration of T30 haptoglobin (r = 0.62; P = 0.006). Similarly, at T0, the expression of PD-1 in T lymphocytes showed a significant correlation with the serum concentration of haptoglobin T30 (r = 0.72; P = 0.0007).
In addition, the serum concentration of haptoglobin at T10 was correlated with the expression of CTLA-4 in T lymphocytes at T30 (r = 0.74; P = 0.0005) and the expression of PD-1 in T lymphocytes at T30 (r = 0.63; P = 0.005). The serum concentration of haptoglobin at T30 was correlated with CTLA-4 expression in T lymphocytes at T30 (r = 0.65; P = 0.41) and PD-1 expression in T lymphocytes at T30 (r = 0, 65; P = 0.004). The predictive values of haptoglobin at distinct time points from diagnosed endometrial cytology are shown in Supplementary Table 1.
Discussion
The postpartum period in dairy cows represents the successful completion of a long gestation period in which there is a challenge to uterine immunity. For instance, during pregnancy, host immunity must be suppressed to limit the immune response against the halogen concept, in which the differentiation of regulatory T cells is crucial to inhibit T-cell responses and the inflammatory process. On the other hand, the immune response must act against infections (28, 29). In this scenario, the immune checkpoints CTLA-4 and PD-1 are widely recognized as negative regulators of T-cell immunity (30). Thus, the higher expression of CTLA-4 and PD-1 in T lymphocytes during the prepartum period in dairy cows in the present study indicates its important role in the maintenance of maternal immune tolerance that occurs during pregnancy, as previously reported in humans (31–34) and rats (35). In fact, T cells play a key role in the induction and maintenance of maternal–fetal tolerance in dairy cows (26) because they are able to differentiate into distinct subsets, including Th1, Th2, Th17, and Treg cells, and during normal pregnancy, there is a dominance of Th2 and Treg-type cytokines, considered the main mechanism of induction of tolerance of the fetus; however, in spontaneous abortion, Th1-type cytokines are dominant (36, 37). At this point, progesterone, although not measured in the experiment, seems to play an important role in the maternal immune tolerance process in dairy cows (38). Although no study has investigated the link between progesterone and the expression of immune checkpoints in T cells in dairy cows, the induction of the expression of CTLA-4 and PD-1 immune checkpoints in T lymphocytes by progesterone has been previously demonstrated in mice (39).
In addition, it is clear that both the intensity of uterine immunosuppression that occurs during the prepartum period and the rapid restoration of defense mechanisms determine whether the animal will develop metritis or endometritis in the postpartum period (38). This may explain, at least in part, the relationship between the expression of CTLA-4 and PD-1 immune checkpoints in T lymphocytes in the prepartum and at partum with endometrial cytology at T20 and T30, an important tool used in the diagnosis of bovine endometritis. Thus, we should highlight that our study showed that the levels of PD-1 and CTLA-4 expression in T cells at parturition could predict the occurrence of cytological endometritis in dairy cows, paving the way for the early identification and diagnosis of susceptible cows. Furthermore, the increased expression of CTLA-4 and PD-1 immune checkpoint markers in T lymphocytes may explain the susceptibility of dairy cattle to infection in the prepartum and calving periods, especially when considering the importance of T cells for mucosal immunity (40). Unsurprisingly, the prevalence of new intramammary infections in the prepartum period and at partum is also very high (41) and may be explained at least in part by the findings of the present study; this possibility deserves further research. Additionally, the aforementioned finding is supported by the correlation between the expression of the immune checkpoint markers CTLA-4 and PD-1 in T lymphocytes during the prepartum period and with the increase in the serum concentration of Hp, which is the most important acute-phase protein in cattle (14, 42) and is usually associated with inflammatory processes resulting from infections.
The negative energy balance gains prominence in the postpartum period, leading to the occurrence of metabolic disorders and infectious diseases such as hypocalcaemia, abomasal displacement, mastitis, and metritis (43, 44) due at least in part to negative implications for the immune response (28, 45). However, the present study did not find a relationship between the occurrence of ketosis and the expression of CTLA-4 and PD-1 in blood T lymphocytes, suggesting that the recognized immunosuppression related to the negative energy balance is not related to the expression of the immune checkpoints studied here in T lymphocytes. Therefore, considering the limited number of dairy cows diagnosed with ketosis in our study, we should be careful when interpreting our findings.
We should also note that the inhibition of T lymphocytes by these immune checkpoints is reversible, for example, by the use of anti-programmed death-ligand 1 (PD-L1), anti-PD-1, or CTLA-4 monoclonal antibodies, opening new frontiers for the treatment and prevention of diseases. Thus, the functional inhibition of immunosuppressive factors represents a viable tool to reactivate immune cells in a state of tolerance, providing a new therapeutic strategy for chronic infections and tumor diseases or as an approach to increase the efficacy of vaccines (46).
Conclusion
The present study pointed to the important role of the expression of PD-1 and CTLA-4 in T lymphocytes during the periparturient period and its effect on uterine health. Thus, our findings suggest that immune checkpoints play a key role in immunological tolerance during bovine pregnancy, which in turn, if they are highly expressed in T lymphocytes, may inhibit the immune response to such an extent that they lead to negative implications for uterine health. Thus, the present study represents an initial step that paves the way for the development of new therapeutic alternatives directed by the host with the objective of increasing the resistance of dairy cattle to infections in this critical period of their life.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author/s.
Ethics Statement
The animal study was reviewed and approved by Universidade Federal da Paraíba. Written informed consent was obtained from the owners for the participation of their animals in this study.
Author Contributions
CS performed all the experiments and wrote and edited the manuscript. EL, RO, BO, RR, and MM performed the experiments. DL performed all statistical analyses. MB, ER, IM, FS, and AF designed and supervised the studies and drafted and edited the manuscript. All authors have read and agreed to the published version of this manuscript.
Funding
This work was financed by Public Call n. 01/2021 Produtividade em Pesquisa PROPESQ/PRPG/UFPB, project code: 13303-2020.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
CS is grateful to Coordenação de Aperfeiçoamento de Pessoal de Nível Superior-CAPES for her scholarship. The authors would also like to thank Empresa Paraibana de Pesquisa, Extensão Rural e Regularização Fundiária-Estação Experimental de Alagoinha, Paraíba, for kindly providing the animals for this study.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fvets.2022.928521/full#supplementary-material
References
1. Lacasse P, Vanacker N, Ollier S, Ster C. Innovative dairy cow management to improve resistance to metabolic and infectious diseases during the transition period. Res Vet Sci. (2018) 116:40–6. doi: 10.1016/j.rvsc.2017.06.020
2. Santos KR, Souza FN, Blagitz MG, Batista CF, Bertagnon HG, Gomes RC, et al. Mastitis in the transition period: identification of potential blood markers. Arquivo Brasileiro de Medicina Veterinária e Zootecnia. (2018) 70:1120–8. doi: 10.1590/1678-4162-10069
3. Leblanc S. Monitoring metabolic health of dairy cattle in the transition period. J. Reprod. Develop. (2010) 56:29–35. doi: 10.1262/jrd.1056S29
4. Mezzetti M, Cattaneo L, Passamonti MM, Lopreiato V, Minuti A, Trevisi E, et al. The transition period updated: a review of the new insights into the adaptation of dairy cows to the new lactation. Dairy. (2021) 2:617–36. doi: 10.3390/dairy2040048
5. Sheldon IM, Cronin J, Goetze L, Donofrio G, Schuberth HJ. Defining postpartum uterine disease and the mechanisms of infection and immunity in the female reproductive tract in cattle. Biol Reprod. (2009) 81:1025–32. doi: 10.1095/biolreprod.109.077370
6. Druker SA, Sicsic R, Straten M, Goshen T, Kedmi M, Raz T, et al. Cytological endometritis diagnosis in primiparous versus multiparous dairy cows. J Dairy Sci. (2022) 105:665–83. doi: 10.3168/jds.2020-20064
7. Sheldon IM, Molinari PCC, Ormsbya TJR, Bromfieldb JJ. Preventing postpartum uterine disease in dairy cattle depends on avoiding, tolerating and resisting pathogenic bacteria. Theriogenology. (2020) 150:158–65. doi: 10.1016/j.theriogenology.2020.01.017
8. Wykes MN, Sharon RL. Immune checkpoint blockade in infectious diseases. Nat Rev Immunol. (2018) 18:91–104. doi: 10.1038/nri.2017.112
9. Watari K, Konnai S, Maekawa N, Okagawa T, Suzuki Y, Murata S, et al. Immune inhibitory function of bovine CTLA-4 and the effects of its blockade in IFN-γ production. BMC Vet Res. (2019) 15:380. doi: 10.1186/s12917-019-2082-7
10. Ikebuchi R, Konnai S, Okagawa T, Yokoyama K, Nakajima C, Suzuki Y, et al. Blockade of bovine PD-1 increases T cell function and inhibits bovine leukemia virus expression in B cells in vitro. Vet Res. (2013) 44:59. doi: 10.1186/1297-9716-44-59
11. Goto S, Konnai S, Okagawa T, Nishimori A, Maekawa N, Gondaira S, et al. Increase of cells expressing PD-1 and PD-L1 and enhancement of IFN-γ production via PD-1/PD-L1 blockade in Bovine mycoplasmosis. Immunity, Inflamm Dis. (2017) 5:355–63. doi: 10.1002/iid3.173
12. Okagawa T, Konnai S, Nishimori A, Maekawa N, Goto S, Ikebuchi R, et al. Cooperation of PD-1 and LAG-3 in the exhaustion of CD4[[sp]]+[[/sp]] and CD8[[sp]]+[[/sp]] T cells during bovine leukemia virus infection. Vet Res. (2018) 49:50. doi: 10.1186/s13567-018-0543-9
13. Okagawa T, Konnai S, Nishimori A, Maekawa N, Ikebuchi R, Goto S, et al. Anti-bovine programmed death-1 rat-bovine chimeric antibody for immunotherapy of bovine leukemia virus infection in cattle. Front Immunol. (2017) 8:650. doi: 10.3389/fimmu.2017.00650
14. Eckersall PD, Bell R. Acute phase proteins: Biomarkers of infection and inflammation in veterinary medicine. The Veterinary Journal. (2010) 185:23–7. doi: 10.1016/j.tvjl.2010.04.009
15. Horadagoda NU, Knox KMG, Gibbs HA, Reid SWJ, Horadagoda A, Edwards SER, et al. Acute phase proteins in cattle: discrimination between acute and chronic inflammation. Veterinary Record. (1999) 144:437–41. doi: 10.1136/vr.144.16.437
16. Schmitt R, Pieper L, Gonzalez-Grajales LA, Swinkels J, Gelfert CC, Staufenbiel R, et al. Evaluation of different acute-phase proteins for herd health diagnostics in early postpartum Holstein Friesian dairy cows. J. Dairy Research. (2021) 88:33–7. doi: 10.1017/S0022029921000078
17. Roberts T, Chapinal N, LeBlanc SJ, Kelton DF, Dubuc J, Duffield TF. Metabolic parameters in transition cows as indicators for early-lactation culling risk. J. Dairy Sci. (2012) 95:3057–63. doi: 10.3168/jds.2011-4937
18. Makimura S, Suzuki N. Quantitative determination of bovine serum haptoglobin and its elevation in some inflammatory diseases. Japanese J. Veter Sci. (1982) 44:15–21. doi: 10.1292/jvms1939.44.15
19. Martins TM, Santos RL, Paixão TA, Muniz CS, Borges ÁM. Imunidade inata uterina em vacas após o parto. Revista Brasileira de Reprodução Animal-Belo Horizonte. (2014) 38:214–9.
20. Moura ARF, Tsuruta SA, Oliveira PM, Nasciutti NR, Santos RM, Saut JPE, et al. Endometrite subclínica após o tratamento de vacas com endometrite clínica. Arch Veter Sci. (2012) 17:32–41. doi: 10.5380/avs.v17i3.25529
21. Kasimanickam R, Duffield TF, Foster RA, Gartley CJ, Leslie KE, Walton JS, et al. Endometrial cytology and ultrasonography for the detection of subclinical endometritis in postpartum dairy cows. Theriogenology. (2004) 62:9–23. doi: 10.1016/j.theriogenology.2003.03.001
22. Kim IH, Kang HG, Jeong JK, Hur TY, Jung YH. Inflammatory cytokine concentrations in uterine flush and serum samples from dairy cows with clinical or subclinical endometritis. Theriogenology. (2014) 82:427–32. doi: 10.1016/j.theriogenology.2014.04.022
23. Sheldon IM. Endometritis in cattle: pathogenesis, consequences for fertility, diagnosis and therapeutic recommendations. Partners Reprod. (2006) 2:1–5.
24. Della Libera AMMP, Souza FN, Batista CF, Santos BP, Azevedo LFF, Sanchez EMR, et al. Effects of bovine leukemia virus infection on milk neutrophil function and the milk lymphocyte profile. Vet Res. (2015) 46:2. doi: 10.1186/s13567-014-0125-4
25. Core Team R. R: A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing. (2021).
26. Wickham H, Averick M, Bryan J, Chang W, Mcgowan LA, François R, et al. Welcome to the tidyverse. J. Open Source Software. (2019) 4:1686. doi: 10.21105/joss.01686
27. Robinson D, Hayes A, Couch S. Convert Statistical Objects into Tidy Tibbles. R Package Version. 0.7,6. (2021).
28. Esposito G, Irons PC, Webb EC, Chapwanya A. Interactions between negative energy balance, metabolic diseases, uterine health and immune response in transition dairy cows. Anim Reprod Sci. (2014) 144:60–71. doi: 10.1016/j.anireprosci.2013.11.007
29. Meggyes M, Miko E, Szigeti B, Farkas N, Szereday L. The importance of the PD-1/PD-L1 pathway at the maternal-fetal interface. BMC Pregnancy Childbirth. (2019) 19:1–12. doi: 10.1186/s12884-019-2218-6
30. Buchbinder EI, Desai A. CTLA-4 and PD-1 pathways: similarities, differences, and implications of their inhibition. Am J Clin Oncol. (2016) 39:98. doi: 10.1097/COC.0000000000000239
31. Andrikopoulou A, Korakitik AM, Apostolidou K, Dimopoulos MA, Zagouri F. Immune checkpoint inhibitor administration during pregnancy: a case series. ESMO Open. (2021) 6:100–262. doi: 10.1016/j.esmoop.2021.100262
32. Enninga EAL, Harrington SM, Creedon DJ, Ruano R, Markovic SN, Dong H, et al. Immune checkpoint molecules soluble program death ligand 1 and galectin-9 are increased in pregnancy. Am J Reprod Immunol. (2018) 79:e12795. doi: 10.1111/aji.12795
33. Miko E, Meggyes M, Doba K, Barakonyi A, Szereday L. Immune checkpoint molecules in reproductive immunology. Front Immunol. (2019) 10:846. doi: 10.3389/fimmu.2019.00846
34. Wang S, Chen C, Li M, Qian J, Sun F, Li Y, et al. Blockade of CTLA-4 and Tim-3 pathways induces fetal loss with altered cytokine profiles by decidual CD4+ T cells. Cell Death Dis. (2019) 10:1–10. doi: 10.1038/s41419-018-1251-0
35. Tian M, Zhang Y, Liu Z, Sun G, Mor G, Liao A, et al. The PD-1/PD-L1 inhibitory pathway is altered in pre-eclampsia and regulates T cell responses in pre-eclamptic rats. Sci Rep. (2016) 6:1–14. doi: 10.1038/srep27683
36. Piccinni MP, Beloni L, Livi C, Maggi E, Scarselli G, Romagnani S, et al. Defective production of both leukemia inhibitory factor and type 2 T-helper cytokines by decidual T cells in unexplained recurrent abortions. Nat Med. (1998) 4:1020–4. doi: 10.1038/2006
37. Arck PC, Hecher K. Fetomaternal immune cross-talk and its consequences for maternal and offspring's health. Nat Med. (2013) 19:548–56. doi: 10.1038/nm.3160
38. Hansen PJ. Maternal immunological adjustments to pregnancy and parturition in ruminants and possible implications for postpartum uterine health: Is there a prepartum–postpartum nexus? J. Animal Sci. (2013) 91:1639–1649. doi: 10.2527/jas.2012-5934
39. D'Alotto-Moreno T, Cerliani J, Croci D, Salatino M, Rabinovich GA. Progesterone links. tumor-associated immunosuppression and metastasis in triple-negative breast cancer. In: Front. Immunol. Conference Abstract: IMMUNOCOLOMBIA 2015-11[[sp]]th[[/sp]] Congress of the Latin American Association of Immunology-10o. Congreso de la Asociación Colombiana de Alergia, Asma e Inmunología. (2015).
40. Chase C, Kaushik RS. Mucosal immune system of cattle: all immune responses begin here. Veter Clin. (2019) 35:431–51. doi: 10.1016/j.cvfa.2019.08.006
41. De Vliegher S, Fox LK, Piepers S, McDougall S, Barkerma HW. Invited review: Mastitis in dairy heifers: Nature of the disease, potential impact, prevention, and control. J Dairy Sci. (2012) 95:1025–40. doi: 10.3168/jds.2010-4074
42. Brandão AP, Cooke RF, Corrá FN, Piccolo MB, Gennari R, Leiva T, et al. Physiologic, health, and production responses of dairy cows supplemented with an immunomodulatory feed ingredient during the transition period. J Dairy Sci. (2016) 99:5562–5572. doi: 10.3168/jds.2015-10621
43. Ospina PA, Nydam DV, Stokol T, Overton TR. Association between the proportion of sampled transition cows with increased nonesterified fatty acids and β-hydroxybutyrate and disease incidence, pregnancy rate, and milk production at the herd level. J Dairy Sci. (2010) 93:3595–601. doi: 10.3168/jds.2010-3074
44. Chapinal N, Carson M, Duffield TF, Capel M, Godden S, Overton M, et al. The association of serum metabolites with clinical disease during the transition period. J Dairy Sci. (2011) 94:4897–903. doi: 10.3168/jds.2010-4075
45. Wathes DC, Clempson AM, Pollott GE. Associations between lipid metabolism and fertility in the dairy cow. Reprod Fertility Develop. (2012) 25:48–61. doi: 10.1071/RD12272
Keywords: immune checkpoints, endometrial cytology, transition period, endometritis, dairy cow
Citation: Souza CMS, Lima ES, Ordonho RF, Oliveira BRRS, Rodrigues RC, Moura MFD, Lima DM, Blagitz MG, Ramos Sanchez EM, Medeiros IA, Souza FN and Fernandes ACC (2022) Brief Research Report: Expression of PD-1 and CTLA-4 in T Lymphocytes and Their Relationship With the Periparturient Period and the Endometrial Cytology of Dairy Cows During the Postpartum Period. Front. Vet. Sci. 9:928521. doi: 10.3389/fvets.2022.928521
Received: 25 April 2022; Accepted: 17 June 2022;
Published: 22 July 2022.
Edited by:
Massimo Amadori, Italian Network of Veterinary Immunology, ItalyReviewed by:
Federica Riva, University of Milan, ItalyAspinas Chapwanya, Ross University School of Veterinary Medicine, Saint Kitts and Nevis
Copyright © 2022 Souza, Lima, Ordonho, Oliveira, Rodrigues, Moura, Lima, Blagitz, Ramos Sanchez, Medeiros, Souza and Fernandes. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Carolina Menezes Suassuna de Souza, Y2Fyb2xpbmEuc3Vhc3N1bmFAZ21haWwuY29t
†These authors have contributed equally to this work and share last authorship