
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Vet. Sci. , 23 June 2022
Sec. Animal Nutrition and Metabolism
Volume 9 - 2022 | https://doi.org/10.3389/fvets.2022.882754
 Hui Huang1†
Hui Huang1† Xia Wang1,2†Ling Yang1Wenxiang He1
Xia Wang1,2†Ling Yang1Wenxiang He1 Tiantian Meng1Ke Zheng1Xin Xia1Yingjun Zhou3,4
Tiantian Meng1Ke Zheng1Xin Xia1Yingjun Zhou3,4 Jianhua He1Chunming Liu1Shengwen Zou5
Jianhua He1Chunming Liu1Shengwen Zou5 Dingfu Xiao1*
Dingfu Xiao1*In order to study the regulation of Fenugreek seed extract (FSE) on the immunity of broilers, and explore the appropriate amount of FSE in broilers' production, 1-day-old yellow feather broilers with a total of 420 birds were randomly allocated into seven treatments. Each treatment had six replicates, with 10 birds per replicate. The two control groups were the basic fodder group fed with basal diet and the bacitracin zinc group added 30 mg/kg bacitracin zinc to the basal diet. Experimental groups included five levels of FSE (50, 100, 200, 400, and 800 mg/kg FSE to the basal diet, respectively). The pre-test period was 7 days and the formal test lasted for 56 days. The results showed that the average daily gain (ADG) of 50 and 800 mg/kg FSE groups was significantly increased (P < 0.01), and the feed to gain ratio (F/G) of FSE groups was significantly decreased (P < 0.01) compared with the basic fodder and the bacitracin zinc groups. Compared with the basic fodder group, the serum total cholesterol (TC) content in the FSE groups was significantly decreased (P < 0.05), the serum low density lipoprotein cholesterol (LDL-C) content of 50, 100, and 800 mg/kg FSE groups was significantly lower than that of the basic fodder group (P < 0.05). Compared with the basic fodder and bacitracin zinc groups, the serum immunoglobulins (IgG, IgM, IgA) content of 100 and 200 mg/kg FSE groups were significantly increased (P < 0.05). Compared with the bacitracin zinc group, the serum interleukins (IL-1, IL-10) content of 400 mg/kg FSE group were significantly increased (P ≤ 0.05), and the serum interferon-γ (IFN-γ) content of 100 and 200 mg/kg FSE groups was significantly increased (P < 0.05). Compared with the basic fodder group, the lower doses (0–400 mg/kg) of FSE had no significant effect on the mRNA expression of toll-like receptors 4/ myeloid differentiation factor 88/ nuclear factor-κB (TLR4/MyD88/NF-κB) signaling pathways (P > 0.05). The 800 mg/kg FSE treatment group significantly increased the expression levels of nuclear factor-κB (NF-κB) mRNA in the spleen of broilers (P < 0.05). The zinc bacitracin group significantly increased the expression levels of myeloid differentiation factor 88 (MyD88) and nuclear factor-κB (NF-κB) mRNA (P ≤ 0.05). The results showed that FSE could promote the secretion of immunoglobulins, regulate the body's cytokines, and have a positive effect on immunity in broilers. Furthermore, the recommended supplement of FSE is 100 mg/kg in the broiler diet.
Antibiotics are widely added in feed because they are beneficial to the development and disease resistance of livestock and poultry. However, the long-term use of antibiotics makes animals resistant to antibiotics, while leaving antibiotics in animal products, adversely affecting human health (1, 2). Since July 1, 2020, banning the use of antibiotics in livestock and poultry nutrition in China, plant secondary metabolites and their derivatives have attracted a lot of attention for their potential role as alternatives for antibiotics. The research of plant extracts replacing antibiotics has become a hot spot (3). Fenugreek is a famous medicinal plant. The plant, about 60 cm tall, has light yellow triangular-shaped flowers and trilobal-shaped leaves. It is widely used in medicine as one type of plant extracts that have been frequently studied in recent years. Fenugreek is a multifunctional herb belonging to the family of fabacecae which is known for its antifungal, antiviral, anticarcinogenic, antidiabetic and antimicrobial properties (4). Fenugreek extract mainly refers to plant extracts from fenugreek seeds that have a certain beneficial effect on the animal body. It is mainly rich in polyphenols, flavonoids, saponins, polysaccharides and alkaloids, and other nutrients and chemical components, which have positive effects on livestock and poultry such as promoting growth and improving immunity (5). Some researchers have reported that fenugreek can stimulate humoral and cellular immune mechanisms (6, 7) and regulate body immunity (8–11). However, others have noticed that the use of fenugreek in layer diets in amounts of 1.00 and 2.00% has a negative influence on the egg production. The objective of the present study was to confirm previous work on dietary supplementation of fenugreek extract on growth performance, immune organ indexes, blood parameters and NF-κB signaling pathway in broilers, and explore whether fenugreek extract can replace antibiotics as a new feed additive and be applied to broiler production. Moreover, the study is helpful for finding out the appropriate concentration of fenugreek extract that is beneficial to the growth of broiler and the health of broiler production. It provides new ideas and methods for improving the growth of broiler and the health of broilers by means of nutritional regulation in production and also provides a reference for the research of FSE in improving the quality and health of chickens.
The fenugreek extract used in the experiment contained 50% polysaccharides, 10% flavonoids, 15% saponins, 2% alkaloids and other ingredients, and was provided by Hunan Jinhan Pharmaceutical Co., Ltd. The fenugreek was extracted by water, concentrating the filtrate after two times of reflux extraction and then spray-drying into powder.
The animal experiment was approved by the Institutional Animal Care and Use Committee of Hunan Agricultural University, Hunan, China.
In this study, a total of 420 1-day-old yellow feather broilers with similar body weight (BW)(55.30 ± 0.10 g) were randomly allocated into seven treatments. Each treatment had six replicates, with 10 birds per replicate. Five concentrations of FSE experimental groups were added 50, 100, 200, 400, and 800 mg/kg FSE to the basal diet, respectively. The trial period lasted 63 days. The first 7 days were a pre-test period lasted for the broiler to adapt to the diets, and the formal trial period lasted for 56 days. According to the standard nutritional requirement of broilers (Agricultural industry standard of the people's Republic of China—chicken breeding standard NY/T33-2004), the basic diet formula of the formal trial period was divided into two periods (1–28, 29–56 days). The composition and nutritional level of basic diet and premix were shown in Tables 1, 2, respectively. The experiment was conducted by a 3-layer cage with natural ventilation, each cage was a replicate of 10 chickens, and the 6 replicates of each treatment were divided into two 1–3 layers, which were evenly distributed in each position of the chicken house. Birds received 24 h of light until day 4; then, light duration was decreased to 18 h for the remainder of the trial. The temperature gradually decreased from 32°C on day 1 to 20°C on day 27, and the humidity was maintained at 50–65%. The experimental chickens had free diets and water, and the experiment routine immunization and disinfection.
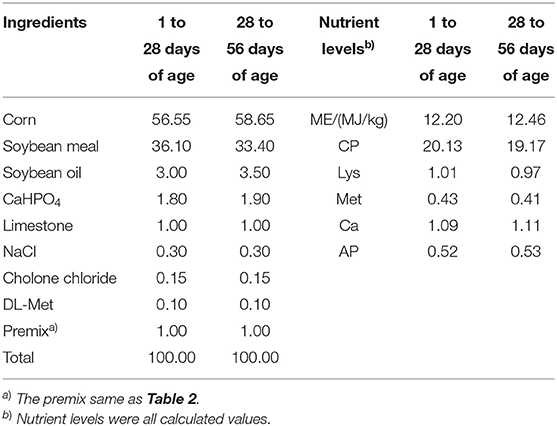
Table 1. Composition and nutrient levels of basal diets (DM basis) %.
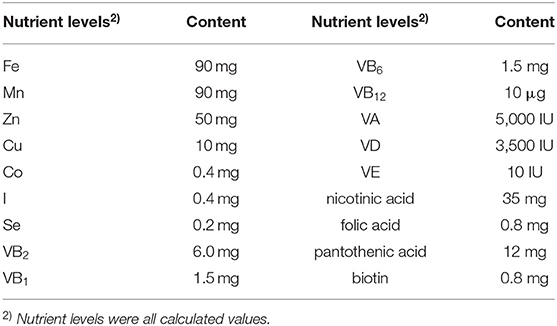
Table 2. Nutrient level of premix (provided per kg of diet).
At day 56 of the experiment, broilers were weighed on an empty stomach (fasting for 12 h). The average daily gain (ADG), average daily feed intake (ADFI) and feed conversion ratio (Feed/Gain, F/G) of the birds were determined.
At day 56 of the experiment, healthy broilers weighing close to the average weight from each repetition were randomly selected and weighed after 12 h feed withdrawal and immediately slaughtered after exsanguination. The thymus, spleen and bursa of Fabricius were separated after dissection and weighed. Immune organ indexes were calculated according to the change of the immune organ weight. The spleen, thymus, and bursa of Fabricius indexes were calculated.
Blood samples (20 mL) were taken from the wing vein and centrifuged at 3,500 r/min for 10 min at 4°C after standing at 37°C for 1 h to prepare serum. The separated serum was frozen at −20°C pending analysis. The content of total protein (TP), albumin (ALB), triglycerides (TG), total cholesterol (TC), high density lipoprotein cholesterol (HDL-C), low density lipoprotein cholesterol (LDL-C), glucose (GLU), alanine transaminase (ALT), aspartate transaminase (AST), urea nitrogen (BUN) and creatinine (CREA) were measured with commercial kits (Jiancheng Bioengineering Institute) using an Mindray BS200 automatic biochemical analyzer (12).
Individual serum samples were detected for the levels of serum immunoglobulin (IgA, IgG, IgM) and cytokines (IL-1, IL-2, IL-6, IL-10, INF-γ, and TNF-α) by ELISA using test kits purchased from Jiangsu Yutong Biological Technology Co., Ltd. (Yancheng, China), following the manufacturer's instructions (13).
The mRNA expression of nuclear factor-κB (NF-κB), toll-like receptor 4 (TLR4), myeloid differentiation factor 88 (MyD88), interferon-γ (IFN-γ), interleukin-1 (IL-1), interleukin-10 (IL-10), and TNF receptor associated factor 6 (TRAF6) in the broilers' spleen was quantified by quantitative real-time PCR. GAPDH was used as the reference gene to normalize the gene expression data. According to the gene sequence retrieved by Gene in NCBI, the primers were designed and synthesized by Shenggong Bioengineering (Shanghai) Co., Ltd. The sequence is shown in Table 3.
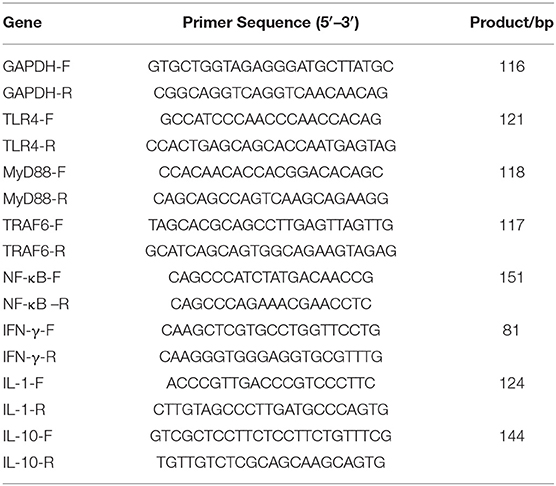
Table 3. Primer sequences for quantitative real-time PCR analysis.
The statistical methods were used to analyze the data in Tables 4–8; Figure 1. All of the data were expressed as the mean ± standard deviation (SD) and were analyzed statistically by one-way ANOVA and Duncan multiple comparison of variance using SPSS 21.0.The linear and quadratic effects of dietary FSE supplementation levels were determined by polynomial contrasts. The differences were considered to be significant at P ≤ 0.05.
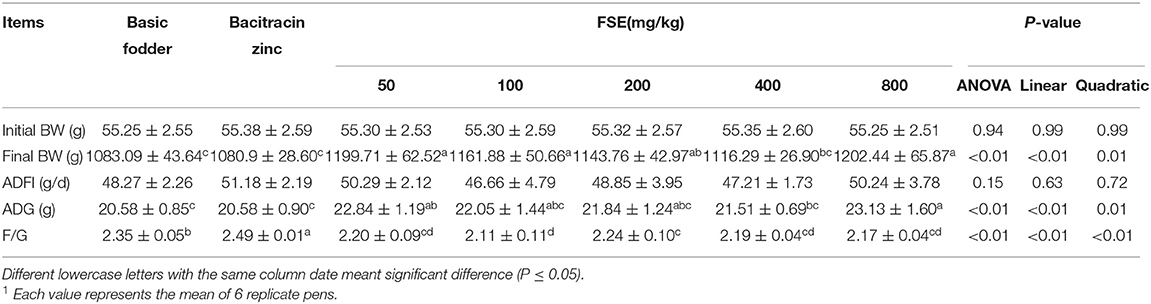
Table 4. Effect of fenugreek seed extract on growth performance of broilers1.
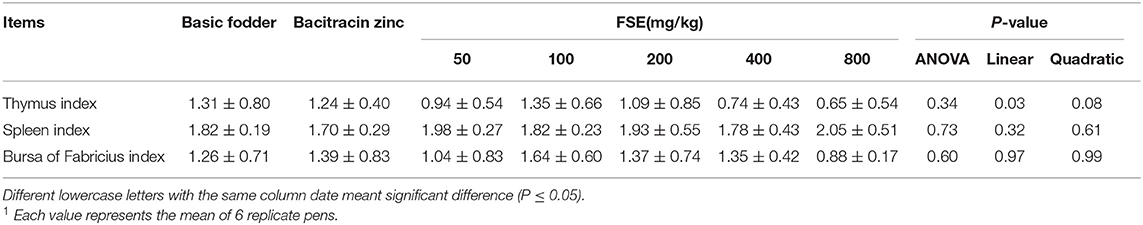
Table 5. Effect of fenugreek seed extract on immune organ index of broilers (%)1.
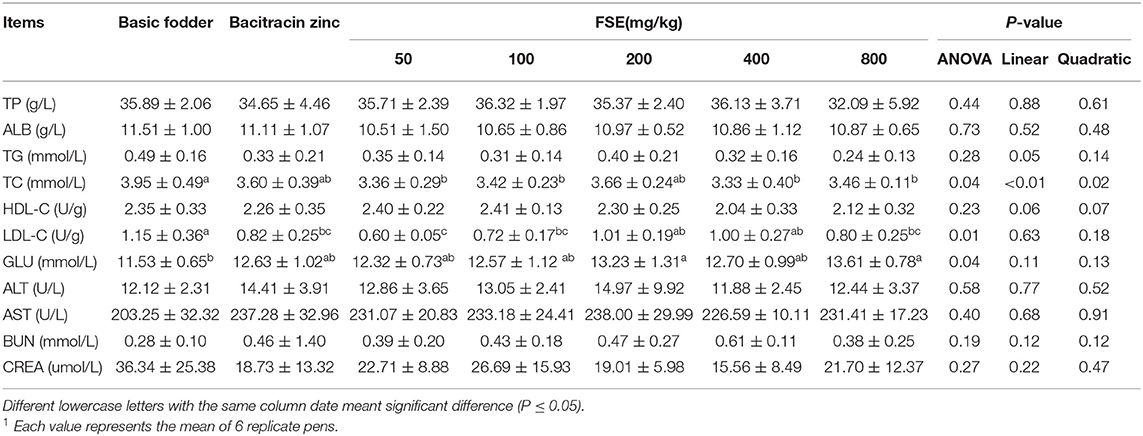
Table 6. Effects of fenugreek seed extract on serum biochemistry in broilers1.
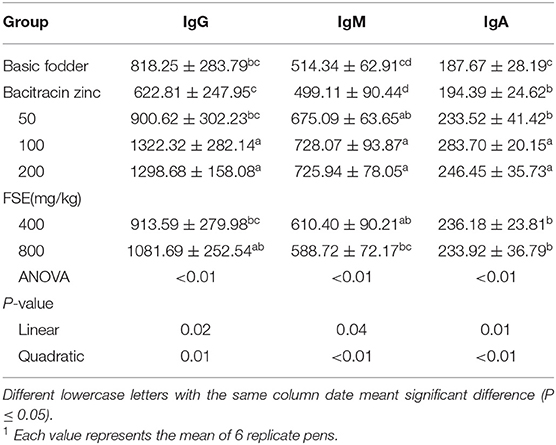
Table 7. Effects of fenugreek seed extract on serum immunoglobulin in broilers1.
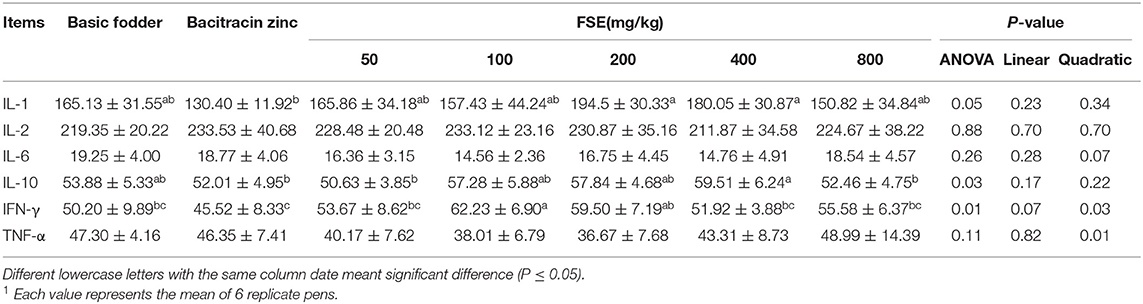
Table 8. Effect of fenugreek seed extract on serum cytokine content in broilers1.
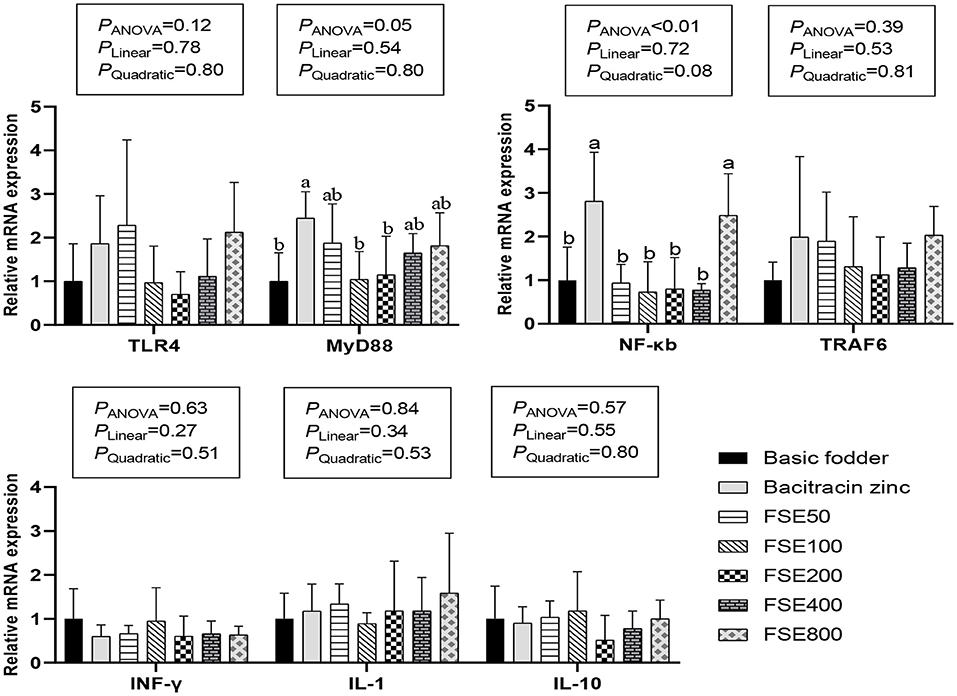
Figure 1. Effects of fenugreek seed extract on gene expression of TLR4/MyD88/NF-κB signaling pathway in spleen of broilers.
As shown in Table 4, dietary FSE levels did not significantly affect ADFI (P > 0.05). The ADG linearly and quadratically increased (P < 0.01) and F/G decreased linearly and quadratically (P < 0.01) with increasing FSE supplementation in broilers. The ADG of 50 and 800 mg/kg FSE groups was significantly higher than that of the basic fodder group and bacitracin zinc group (P < 0.01). The F/G of 50, 100, 200, 400, 800 mg/kg FSE groups was significantly lower than that of the basic fodder group and bacitracin zinc group (P < 0.01).
As shown in Table 5, detary FSE levels did not significantly affect spleen index and bursa of Fabricius index (P > 0.05), but the thymus index decreased linearly (P < 0.05) with increasing FSE supplementation in broilers.
As shown in Table 6, dietary FSE levels did not significantly affect the serum TP,ALB,TG,HDL-C,ALT,AST,BUN,CREA contents (P > 0.05). The serum TC content linearly and quadratically decreased with increasing FSE supplementation in broilers (P < 0.05). The serum TC content of 50, 100, 400, and 800 mg/kg FSE groups was significantly lower than that of the basic fodder group (P < 0.05), and no significant difference from the bacitracin zinc group (P > 0.05). The serum LDL-C content of 50, 100, and 800 mg/kg FSE groups was significantly lower than that of the basic fodder group (P < 0.05), and no significant difference compared with the bacitracin zinc group (P > 0.05). The serum GLU content of 200 and 800 mg/kg FSE groups was significantly higher than that of the basic fodder group (P < 0.05).
As shown in Table 7, the serum IgG, IgM, IgA contents linearly and quadratically increased with increasing FSE supplementation in broilers (P < 0.05). The serum IgG content of 100 and 200 mg/kg FSE groups was significantly higher than that of the basic fodder group and bacitracin zinc group (P < 0.05). The serum IgG content of 800 mg/kg FSE group was significantly higher than that of the bacitracin zinc group (P < 0.05), but no significant difference from the basic fodder group (P > 0.05). The serum IgM content of 50, 100, 200, and 400 mg/kg FSE groups was significantly higher than that the basic fodder group and bacitracin zinc group (P < 0.05). The serum IgM content of 800 mg/kg FSE group was significantly higher than that of bacitracin zinc group (P < 0.05). The serum IgA content of 50, 100, 200, 400, and 800 mg/kg FSE groups was significantly higher than that of the basic fodder group (P < 0.05). The serum IgA content of 100 and 200 mg/kg FSE groups was significantly higher than that of the bacitracin zinc group (P < 0.05).
As shown in Table 8, there was no significant difference in the serum IL-2, IL-6, TNF-α content in each group (P > 0.05). Contrasted with the bacitracin zinc group, the serum IL-1 content of 200 and 400 mg/kg FSE groups was significantly increased (P = 0.05), but not significantly different from the basic fodder group (P > 0.05). The serum IL-10 content of 50, 100, 200, and 800 mg/kg FSE groups was not significantly different between that the basic fodder group and bacitracin zinc group (P > 0.05). The serum IL-10 content of 400 mg/kg FSE group was significantly higher than that of the bacitracin zinc group (P < 0.05), but not significantly different from the basic fodder group (P > 0.05). The serum IFN-γ content quadratically increased with increasing FSE supplementation in broilers (P < 0.05). The serum TNF-α content quadratically decreased with increasing FSE supplementation in broilers (P < 0.05). The serum IFN-γ content of 50, 400, and 800 mg/kg FSE groups was not significantly different compared with the basic fodder group and bacitracin zinc group (P > 0.05). The serum IFN-γ content of 100 and 200 mg/kg FSE groups was significantly higher than that of the bacitracin zinc group (P < 0.05). The serum IFN-γ content of 100 mg/kg FSE group was significantly higher than that of the basic fodder group (P < 0.05).
As shown in Figure 1, there were no significant differences in the expression levels of TLR4,TRAF6,IFN-γ, IL-1, and IL-10 mRNA in the spleen TLR4/MyD88/ NF-κB signaling pathway of each group (P > 0.05). The expression of MyD88 mRNA in the spleen of 50, 400, and 800 mg/kg FSE groups was not significantly different between that of the basic fodder group and bacitracin zinc group (P > 0.05). The expression of MyD88 mRNA in the spleen of 100 and 200 mg/kg FSE groups was not significantly different from the basic fodder group (P > 0.05), but was significantly lower than that in the bacitracin zinc group (P = 0.05). The expression of MyD88 mRNA in the spleen of the bacitracin zinc group was significantly higher than that of the basic fodder group (P = 0.05). The expression of NF-κB mRNA in the spleen of 50, 100, 200, and 400 mg/kg FSE groups was not significantly different from the basic fodder group (P > 0.05), but was significantly lower than that in the bacitracin zinc group (P < 0.01). The expression of NF-κB mRNA in the spleen of 800 mg/kg FSE group was significantly higher than that of the basic fodder group (P < 0.05), and was not significantly different with that of bacitracin zinc group (P > 0.05). The expression of NF-κB mRNA in the spleen of the bacitracin zinc group was significantly higher than that of the basic fodder group (P < 0.01).
Fenugreek contains carbohydrates and fatty acids, which can improve the growth performance of livestock and poultry. Alloui et al. (14) showed that broilers fed a diet containing 3,000 mg/kg FSE could increase broiler feed intake and reduce F/G. Adding 2% fenugreek seed powder to the diet could increase the ADFI of broilers, while adding 3% fenugreek seed powder to the diet reduces the ADFI of broilers (15). Amein et al. (16) and Laudadio et al. (17) found that the powdered seeds of fenugreek and the 0.5% fenugreek in the diet could reduce F/G of broilers. There was a research result showing that greater dietary concentrations of fenugreek reduced BW and increase F/G (18). In this study, we observed that the 50 and 800 mg/kg FSE groups significantly increased ADG, and except 400 mg/kg FSE, other doses of FSE (50–800 mg/kg) significantly increased the final BW of broilers. This may be affected by the number of samples. Because from the numerical point of view, 400 mg/kg FSE also has a tendency to increase the final BW of broilers, and 100–400 mg/kg FSE has a tendency to increase the ADG of broilers. And the final BW and ADG increased linearly and quadratically with increasing FSE supplementation in broilers. Moreover, dietary FSE supplementation significantly decreased F/G. The F/G decreased linearly and quadratically with increasing FSE supplementation in broilers. These results were similar to Amein et al. (16) and Laudadio et al. (17). FSE can improve broiler production, ensuring the health of chickens without toxic side effects to humans.
The main immune organs of the poultry body are the thymus, spleen and bursa of Fabricius. The bursa of Fabricius is extremely important for the maturation of plasma cells and B lymphocytes that can synthesize and secrete antibodies (19). The spleen of poultry is one of the most important peripheral lymphatic organs that respond to foreign antigen stimulation (20). The development of immune organs greatly affects the immune function and disease resistance of the body. Its weight is caused by the growth and division of its own cells. The immune organ index can reflect the development of immune organs and the functional status of immune cells, indirectly reflect the body's immune level (21). Raju and Bird (22) and Bin-Hafeez et al. (8) found that aqueous extracts of fenugreek could increase thymus index. In this experiment, FSE as a feed additive added to broiler feed show no effect on immune organ index, which is inconsistent with the report of Raju and Bin-Hafeez. The variation in the results from different studies might be due to the difference in the rearing environment and species used (22).
Immunoglobulin (Ig) is a kind of macromolecular protein that can bind with antigen and is rapidly produced after the body stimulated by foreign pathogens. It is the most commonly used detection method for the performance of the body's humoral immune function (23). IgM is the first-line defense against a broad range of infections (24). The presence of immunoglobulins especially IgA and IgG in chicken sera enhances the immunity of the birds (25). According to the research reports, FSE can promote Ig secretion in animals. Hossain et al. (26) found that FSE significantly increased IgG levels in growing pigs. In this study, we observed that every level of FSE dietary supplementation had a positive regulation effect on the serum IgG, IgM, and IgA of broilers, and among them 100 mg/kg is the best. It shows that FSE could significantly increase the level of immunoglobulin in broilers, which is beneficial to enhance the immunity of broilers. The main reason why FSE affect immunoglobulin secretion is that FSE contains 50% polysaccharides, and polysaccharide compounds can enhance the immunomodulatory activity of macrophages in animal bodies (27, 28), have the ability to stimulate the production of serum immunoglobulins, and have the potential to regulate innate and adaptive immunity (29, 30).
Serum biochemical indicators can reflect the body's metabolism and some disease states as an effective indicator for checking the health of animal's body. The concentration of TC and TG in serum can be used as indicators of lipid metabolism. Under normal conditions, serum triglycerides maintain dynamic balance. The increase in the concentration of TC and TG in serum indicates that the body's lipid metabolism is abnormal, or the accumulation of fat in the animal's body increases, resulting in an increase in blood lipid contents in the serum (31). LDL-C and HDL-C are lipoproteins associated with cholesterol transport (32). Both are related to atherosclerosis and cardiovascular disease. The former (LDL-C) is positively correlated with the occurrence of the disease, and the latter (HDL-C) is negatively correlated with the occurrence of the disease. FSE can control blood lipids and lower serum total cholesterol. The results of Belguith-Hadriche et al. (33) showed that the use of fenugreek ethyl acetate extracts significantly reduced the levels of TC, TG, and LDL-C in plasma while increasing the plasma levels of HDL-C in plasma. Begum et al. (34) found that FSE has a significant increase in serum HDL-C concentration. In this study, we observed that dietary FSE supplementation decreased the serum TC content in the broiler, and the 50, 100, and 800 mg/kg FSE decreased the serum LDL-C content, and the 200 and 800 mg/kg FSE increased the serum GLU content. Also, FSE had no effect on serum TG and HDL-C. The test results are similar to those of Begum et al. (34) and Belguith-Hadriche et al. (33). The results showed that FSE has the effect on lowering blood lipids, which is beneficial to the body lipid metabolism of broilers. The reason why FSE reduced the content of TC and LDL-C may be related to the alkaloids contained in fenugreek extract (35). According to research, alkaloids can reduce blood TC and LDL-C levels, while beneficially increasing HDL-C levels or no effect on HDL-C levels (36). Trigonelline in FSE may control the absorption of intestinal cholesterol and affect the LDL cholesterol clearance mediated by LDL receptors, thereby controlling the serum cholesterol of broilers, affecting the lipid metabolism of broilers, and playing the role of lowering blood lipids (37–40).
Cytokines are small molecular proteins produced by exogenous or endogenous stimulants induced by immune cells, endothelial cells, and fat cells in the body. They have various physiological functions such as regulating immunity, stimulating endothelial growth, and repairing damage. According to their function and source, cytokines are divided into interferon (IFN), interleukin (IL), tumor necrosis factor (TNF), etc. (41). TNF-α mainly causes white blood cells to accumulate in the inflammation site and has a dual biological role. On the one hand, it is an important medium for the immune protection of the body; on the other hand, it can participate in the immune pathological damage of the body. It plays a very important role in the pathogenesis of tumors and immune diseases (42). IFN-γ has the ability to interfere with virus infection and replication, and its main function is immune regulation (43). The increase of serum IFN-γ is beneficial to activate the body's immunity (44, 45). IL-1, IL-2, and IL-6 are pro-inflammatory factors in the body. Their reduced expression in the body is beneficial to improve the body's anti-inflammatory response and regulate the body's immune function. On the contrary, it can reflect that the body has corresponding inflammation. The reaction will also affect the body's immunity and make the body's immune function imbalance (46, 47). As an anti-inflammatory factor, IL-10 expression in the body is beneficial to improve or activate the body's immune function (48). Studies have shown that FSE could stimulate the regulation of cytokines and significantly reduce the levels of IL-1α, IL-1β, IL-2, IL-6, and TNF-α (49–52). In this study, compared with the bacitracin zinc group, the 200 and 400 mg/kg FSE dietary supplementation increased the serum IL-1 content, the 400 mg/kg FSE increased the serum IL-10 content, the 100 and 200 mg/kg FSE increased the serum IFN-γ content. The serum IFN-γ content quadratically increased with increasing FSE supplementation in broilers. The serum TNF-α content quadratically decreased with increasing FSE supplementation in broilers. The results of this test showed that FSE can increase the secretion of anti-inflammatory factors in broilers and inhibit the synthesis of pro-inflammatory factors, which is basically similar with the studies of Tripathi et al. (10) and Ghosh et al. (51). Fenugreek contains a number of important, beneficial flavonoids, and polyphenol compounds. Nagulapalli et al. (5) have reported the presence of a wide range of flavonoids, namely quercetin, luteolin, vitexin, and 7, 4′-dimethoxy flavanones in the alcoholic extracts of the whole plant. Benayad et al. (53) have reported similar findings of the presence of aglycones kaempferol, quercetin, tricin, and naringenin. The reason why FSE increases serum IFN-γ and IL-10 may be due to the anti-inflammatory active substances contained in FSE. For example, fenugreek flavonoids are the main anti-inflammatory and antioxidant bioactive components (54, 55), and fenugreek polysaccharides are also have anti-inflammatory properties (56). FSE increased the serum IFN-γ value of broiler chickens, may regulate Th17 and Th1 cells (57), inhibit the secretion of pro-inflammatory factors (IL-1, IL-2, IL-6, TNF-α), and promote the anti-inflammatory factor IL-10 secretion, thereby activating the immune function of broilers. In addition, this test found that the serum levels of IL-1, IFN-γ, and IL-10 in the zinc bacitracin group were reduced, which inhibited the activation of inflammatory and immune responses to a certain extent.
Toll-like receptors 4 (TLR4) is the earliest discovered protein among TLR-related proteins. It is distributed in almost all cell lines and is expressed in cells involved in host defense functions (58). The mechanism of its pathway is that the TLR4 receptor binds to the corresponding ligand, the signal is transduced to the TLR4 region, and then the NF-κB and MAPK signaling pathways are further activated to promote the expression and activation of various inflammatory cytokine genes (59). MyD88 is both a key downstream signaling ligand of TLR4 and an important protein in the NF-κB signaling pathway (60, 61). Their mechanism of action is that some extracellular inflammatory signals are presented to MyD88 through the TLR domain, allowing MyD88 to interact with interleukin-1 receptor-associated kinase 4 (IRAK4) to phosphorylate IRAK4 (62). Then phosphorylate IRAK1, and then act on ubiquitin ligase (TRAF6). After MyD88 is activated, TAK-1 and IκB kinase (IKK) are activated under the action of trans-growth factor-β activated kinase 1 binding protein (TAB-1/TAB-2) to activate transcription factors, and then initiate signal transduction through the nucleus (63). Research has shown that plant polysaccharides and plant flavonoids can activate the immune system and play an immunomodulatory effect (64, 65). Hou et al. (64) found that G. frondosa polysaccharide activated macrophages through TLR4-MyD88-IKKβ-NF-κBp65 signaling pathways. Juglan, a flavonol derivative, in LPS-induced C57B/L6 mice significantly reduced pro-inflammatory cytokines and blocked TLR4/NF-κB pathway (66). However, in the present study, the mRNA expression of TLR4/MyD88/NF-κB signaling pathways was not significantly affected by lower doses (0–400 mg/kg) of FSE containing 50% polysaccharides and 10% flavonoids. The variation in the results from different studies might be due to the different species used and the way of adding active substances (64, 66). Furthermore, in the present study, the 800 mg/kg FSE treatment group up-regulated NF-κB expression. The zinc bacitracin group significantly increased the expression levels of MyD88 and NF-κB mRNA in the spleen of broilers. It showed that bacitracin zinc and high doses (800 mg/kg) of FSE have some negative effects on body immunity by the TLR4/MyD88/ NF-κB signaling pathway.
Dietary FSE could further improve growth performance, promote the secretion of broiler immunoglobulins, regulate the body's cytokines and have a positive effect on immunity. FSE is expected to replace antibiotics to improve broiler health and growth and increase chicken production while ensuring chicken quality and health. The optimal concentration of 100 mg/kg for FSE has been initially found.
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author/s.
The animal study was reviewed and approved by the Institutional Animal Care and Use Committee of Hunan Agricultural University, Hunan, China.
All authors listed have made a substantial, direct, and intellectual contribution to the work and approved it for publication.
This project was funded by National Natural Science Foundation of China (31872991), National Natural Science Foundation of Hunan Province-China (2020JJ4364), and Project of Science and Technology Department of Hunan Province (2020NK4247). Cooperation project between Hunan Agricultural University and Hunan Jinhan Pharmaceutical Co., Ltd. (2019xny-js065).
YZ was employed by Geneham Pharmaceutical Co., Ltd. SZ was employed by Hunan Wenfeng Agricultural and Animal Husbandry Technology Co.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fvets.2022.882754/full#supplementary-material
FSE, Fenugreek seed extract; ADG, Average daily gain; ADFI, Average daily feed intake; F/G, Feed/Gain; TP, Total protein; ALB, Albumin; TG, Triglyceride; TC, Total cholesterol; HDL-C, High density lipoprotein cholesterol; LDL-C, Low density lipoprotein-cholesterol; GLU, Glucose; ALT, Alanine aminotransferase; AST, Aspartate aminotransferase; BUN, Blood urea nitrogen; CREA, Creatinine; IL-1, Interleukin-1; IL-2, Interleukin-2; IL-6, Interleukin-6; IL-10, Interleukin-10; IFN-γ, Interferon; TNF-α, Tumor necrosis factor; IgG, Immunoglobulin G; IgM, Immunoglobulin M; IgA, Immunoglobulin A; TLR4, Toll-like receptors 4; GAPDH, Glyceraldehyde-3-phosphate dehydrogenase; MyD88, Myeloid differentiation factor 88; TRAF6, TNF receptor associated factor 6; NF-κB, Nuclear factor-κb; IL-1β, Interleukin-1β; MAPK, Mitogen-activated protein kinase; IRAK4, Interleukin-1 receptor-associated kinase 4; TAK-1, Transforming growth factor-β activated protein kinase; IKK, IκB kinase; IRAK4, Interleukin-1 receptor-associated kinase 4; ELISA, Enzyme-linked immunosorbent assay; TAB-1, Transforming growth factor-β activated protein kinase binding protein.
1. Bacanli M, Başaran N. Importance of antibiotic residues in animal food. Food Chem Toxicol. (2019) 125:462–6. doi: 10.1016/j.fct.2019.01.033
2. Chen J, Ying GG, Deng WJ. Antibiotic residues in food: extraction, analysis, and human health concerns. J Agric Food Chem. (2019) 67:7569–86. doi: 10.1021/acs.jafc.9b01334
3. Cheng G, Hao H, Xie S, Wang X, Dai M, Huang L, et al. Antibiotic alternatives: the substitution of antibiotics in animal husbandry? Front Microbiol. (2014) 5:217. doi: 10.3389/fmicb.2014.00217
4. Alloui MN, Agabou A, Alloui N. Application of herbs and phytogenic feed additives in poultry production. Glob J Anim Sci Res. (2014) 2:234–43.
5. Nagulapalli VK, Swaroop A, Bagchi D, Bishayee A. A small plant with big benefits: fenugreek (Trigonella foenum-graecum Linn.) for disease prevention and health promotion. Mol Nutr Food Res. (2017) 61:1600950. doi: 10.1002/mnfr.201600950
6. Anarthe SJ, Sunitha D, Sandhya R, Ganga M. Immunomodulatory activity for methanolic extract of Trigonella foenum graecum whole plant in Wistar Albino rats. Am J Phytomed Clin Ther. (2019) 2:1081–092.
7. Guardiola FA, Bahi A, Esteban MA. Effects of dietary administration of fenugreek seeds on metabolic parameters and immune status of gilthead seabream (Sparus aurata L.). Fish Shellfish Immunol. (2018) 74:372–9. doi: 10.1016/j.fsi.2018.01.010
8. Bin-Hafeez B, Haque R, Parvez S, Pandey S, Sayeed I, Raisuddin S. Immunomodulatory effects of fenugreek (Trigonella foenum graecum L.) extract in mice. Int Immunopharmacol. (2003) 3:257–65. doi: 10.1016/S1567-5769(02)00292-8
9. Meghwal M, Goswami TK. A Review on the functional properties, nutritional content, medicinal utilization and potential application of fenugreek. Food Process Technol. (2012) 3:1–10. doi: 10.4172/2157-7110.1000181
10. Tripathi S, Maurya AK, Kahrana M, Kaul A, Sahu RK. Immunomodulatory property of ethanolic extract of Trigonella foenum- graeceum leaves on mice. Der Pharmacia Lettre. (2012) 4:708–13.
11. Wani SA, Kumar P. Fenugreek: a review on its nutraceutical properties and utilization in various food products. J Saudi Soc Agric Sci. (2018) 17:97–106. doi: 10.1016/j.jssas.2016.01.007
12. Hu H, Bai X, Xu K, Zhang C, Chen L. Effect of phloretin on growth performance, serum biochemical parameters and antioxidant profile in heat-stressed broilers. Poult Sci. (2021) 100:101217. doi: 10.1016/j.psj.2021.101217
13. Das Q, Tang J, Yin X, Ross K, Warriner K, Marcone MF, et al. Organic cranberry pomace and its ethanolic extractives as feed supplement in broiler: impacts on serum Ig titers, liver and bursal immunity. Poult Sci. (2021) 100:517–26. doi: 10.1016/j.psj.2020.09.044
14. Alloui N, Aksa SB, Alloui MN, Ibrir F. Utilization of fenugreek (Trigonella foenum-graecum) as growth promoter for broiler chickens. J World s Poultry Res. (2012) 2:25–7.
15. Toaha SM, Mollah R, Ahammad MU. Use of dietary fenugreek (Trigonella foenum-graecum L.) seed for the production of safe broiler lean meat. Res Agric Livestock Fish. (2016) 3:305–14. doi: 10.3329/ralf.v3i2.29356
16. Amein SM, Mosaad GMM, Hussein MK. Effect of some medicinal plants as feed additives on growth performance, blood constituents and carcass characteristics of broilers. J Adv Vet Res. (2019) 9:170–7.
17. Laudadio V, Nasiri-Dehbaneh M, Bilal RM, Qotbi A, Javandel F, Ebrahimi A, et al. Effects of different levels of dietary black cumin (Nigella sativa L.) and fenugreek (Trigonella foenum-graecum L.) and their combination on productive traits, selected blood constituents, microbiota and immunity of broilers. Anim Biotechnol. (2020) 16:1–14. doi: 10.1080/10495398.2020.1853138
18. Al-Homidan IH, Ebeid TA, Al-Muzaini A, Abou-Emera OK, Fathi MM. Impact of dietary fenugreek, mung bean, and garden cress on growth performance, carcass traits, blood measurements, and immune response in broiler chickens. Livestock Sci. (2020) 242:104318. doi: 10.1016/j.livsci.2020.104318
19. Liu XD, Song J, Liu X, Shan H. Research note: circular RNA expressing in different developmental stages of the chicken bursa of fabricius. Poult Sci. (2020) 99:3846–52. doi: 10.1016/j.psj.2020.04.026
20. Stutz MW, Johnson SL, Judith FR. Effects of diet, bacitracin, and body weight restrictions on the intestine of broiler chicks. Poult Sci. (1983) 62:1626–32. doi: 10.3382/ps.0621626
21. He S, Yin Q, Xiong Y, Liu D, Hu H. Effects of dietary fumaric acid on the growth performance, immune response, relative weight and antioxidant status of immune organs in broilers exposed to chronic heat stress. Czech J Anim Sci. (2020) 65:104–13. doi: 10.17221/13/2020-CJAS
22. Raju J, Bird RP. Alleviation of hepatic steatosis accompanied by modulation of plasma and liver TNF-alpha levels by Trigonella foenum graecum (fenugreek) seeds in Zucker obese (fa/fa) rats. Int J Obes. (2006) 30:1298–307. doi: 10.1038/sj.ijo.0803254
23. Adeyemi K, Sola-Ojo F, Ishola J, Ahmed M, Lawal M. Influence of Anacardium occidentale leaf supplementation in broiler chicken diet on performance, caecal microbiota, blood chemistry, immune status, carcass, and meat quality. Br Poult Sci. (2021) 62:552–61. doi: 10.1080/00071668.2021.1894321
24. Shi H, Luo Y, Li Y, Zhang F, Liu N. Tetramethylpyrazine supplementation improves performance, digestion, blood and immune state of broilers exposure to oxidative stress. J Anim Physiol Anim Nutr. (2021) 106:132–8. doi: 10.1111/jpn.13566
25. Alyileili SR, El-Tarabily K, Ibrahim WH, Sulaiman M, Hussein AS. Effect of Trichoderma reesei degraded date pits supplementation on growth performance, immunoglobulin levels and intestinal barrier functions of broiler chickens. Recent Pat Food Nutr Agric. (2020) 11:168–81. doi: 10.2174/2212798410666190716163009
26. Hossain MM, Begum M, Nyachoti CM, Hancock JD, Kim IH. Dietary fenugreek seed extract improves performance and reduces fecal E. coli counts and fecal gas emission in lactating sows and suckling piglets. Can J Anim Sci. (2015) 95:561–8. doi: 10.4141/cjas-2014-154
27. Chen F, Huang G. Preparation and immunological activity of polysaccharides and their derivatives. Int J Boil Macromol. (2018) 112:211–16. doi: 10.1016/j.ijbiomac.2018.01.169
28. Tang C, Sun J, Liu J, Jin C, Wu X, Zhang X, et al. Immune-enhancing effects of polysaccharides from purple sweet potato. Int J Boil Macromol. (2019) 123:923–30. doi: 10.1016/j.ijbiomac.2018.11.187
29. Zuo T, Zhao R, Lu S, Zhang N, Zhang Q, Xue C. Novel dietary polysaccharide SIP promotes intestinal secretory immunoglobulin A secretion in mice under chemotherapy. J Funct Foods. (2017) 37:379–89. doi: 10.1016/j.jff.2017.08.012
30. Ramesh HP, Yamaki K, Tsushida T. Effect of fenugreek (Trigonella foenum-graecum L.) galactomannan fractions on phagocytosis in rat macrophages and on proliferation and IgM secretion in HB4C5 cells. Carbohydr Polym. (2002) 50:79–83. doi: 10.1016/S0144-8617(01)00377-0
31. Tan L, Song A, Ren L, Wang C, Song G. Effect of pioglitazone on skeletal muscle lipid deposition in the insulin resistance rat model induced by high fructose diet under AMPK signaling pathway. Saudi J Boil Sci. (2020) 27:1317–23. doi: 10.1016/j.sjbs.2020.03.014
32. Pizzini A, Kurz K, Orth-Hoeller D, Fille M, Rabensteiner J, Lunger F, et al. The impact of bacteremia on lipoprotein concentrations and patient's outcome: a retrospective analysis. Eur J Clin Microbiol Infect Dis. (2019)38:1279–86. doi: 10.1007/s10096-019-03543-w
33. Belguith-Hadriche O, Bouaziz M, Jamoussi K, El Feki A, Sayadi S, Makni-Ayedi F. Lipid-lowering and antioxidant effects of an ethyl acetate extract of fenugreek seeds in high-cholesterol-fed rats. J Agric Food Chem. (2010) 58:2116–22. doi: 10.1021/jf903186w
34. Begum M, Hossain MM, Kim IH. Effects of fenugreek seed extract supplementation on growth performance, nutrient digestibility, diarrhoea scores, blood profiles, faecal microflora and faecal noxious gas emission in weanling piglets. J Anim Physiol Anim Nutr. (2016) 100:1121–9. doi: 10.1111/jpn.12496
35. Cheng C, Li Z, Zhao X, Liao C, Quan J, Bode AM, et al. Natural alkaloid and polyphenol compounds targeting lipid metabolism: treatment implications in metabolic diseases. Eur J Pharmacol. (2020) 870:172922. doi: 10.1016/j.ejphar.2020.172922
36. Wang Y, Zidichouski JA. Update on the benefits and mechanisms of action of the bioactive vegetal alkaloid berberine on lipid metabolism and homeostasis. Cholesterol. (2018) 2018:7173920. doi: 10.1155/2018/7173920
37. Cicero A, Alessandro C, Gani B, Descamps O, Djuric DM, Ezhov M, et al. Lipid lowering nutraceuticals in clinical practice: position paper from an International Lipid Expert Panel. Arch Med Sci. (2017) 13:965–1005. doi: 10.5114/aoms.2017.69326
38. Johnston TP, Korolenko TA, Pirro M, Sahebkar A. Preventing cardiovascular heart disease: promising nutraceutical and non-nutraceutical treatments for cholesterol management. Pharmacol Res. (2017) 120:219–25. doi: 10.1016/j.phrs.2017.04.008
39. Koppen LM, Whitaker A, Rosene A, Beckett RD. Efficacy of berberine alone and in combination for the treatment of hyperlipidemia: a systematic review. J Evid Based Complement Altern Me. (2017) 22:956–68. doi: 10.1177/2156587216687695
40. Wei X, Wang C, Hao S, Song H, Yang L. The therapeutic effect of berberine in the treatment of nonalcoholic fatty liver disease: a meta-analysis. Evid Based Complement Altern Med. (2016) 2016:3593951. doi: 10.1155/2016/3593951
41. Banerjee M, Saxena M. Genetic polymorphisms of cytokine genes in type 2 diabetes mellitus. World J Diabetes. (2014) 5:493–504. doi: 10.4239/wjd.v5.i4.493
42. Lai WY, Wang JW, Huang BT, Lin EP, Yang PC. A Novel TNF–targeting aptamer for TNF–mediated acute lung injury and acute liver failure. Theranostics. (2019) 9:1741–51. doi: 10.7150/thno.30972
43. Shcheglovitova ON, Boldyreva NV, Sklyankina NN, Babayants AA, Frolova IS. IFN-α, IFN-β, and IFN-γ have different effect on the production of proinflammatory factors deposited in weibel-palade bodies of endothelial cells infected with herpes simplex virus type 1. Bull Exp Boil Med. (2016) 161:270–5. doi: 10.1007/s10517-016-3393-3
44. Cruz M, Maldonado-Bernal C, Mondragón-Gonzalez R, Sanchez-Barrera R, Wacher NH, Carvajal-Sandoval G. Glycine treatment decreases proinflammatory cytokines and increases interferon-gamma in patients with type 2 diabetes. J Endocrinol Invest. (2008) 31:694–9. doi: 10.1007/BF03346417
45. Regad T, Chelbi-Alix MK. Role and fate of PML nuclear bodies in response to interferon and viral infections. Oncogene. (2001) 20:7274–86. doi: 10.1038/sj.onc.1204854
46. Dinarello CA. Interleukin-1 in the pathogenesis and treatment of inflammatory diseases. Blood. (2011) 117:3720–32. doi: 10.1182/blood-2010-07-273417
47. Wei L, Lin JX, Leonard WJ. IL-2 family cytokines: new insights into the complex roles of IL-2 as a broad regulator of T helper cell differentiation. Curr Opin Immunol. (2011) 23:598–604. doi: 10.1016/j.coi.2011.08.003
48. Moore KW, De W, Coffman RL, O'Garra A. Interleukin-10 and the interleukin-10 receptor. Annu Rev Immunol. (2001) 19:683–765. doi: 10.1146/annurev.immunol.19.1.683
49. Abedinzade M, Nasri S, Jamal OM, Ghasemi E, Ghorbani A. Efficacy of Trigonella foenum-graecum seed extract in reducing metabolic and inflammatory alterations associated with menopause. Iran Red Crescent Med J. (2015) 17:e26685. doi: 10.5812/ircmj.26685
50. Joshi DV, Patil RR, Naik SR. Hydroalcohol extract of Trigonella foenum-graecum seed attenuates markers of inflammation and oxidative stress while improving exocrine function in diabetic rats. Pharm Biol. (2015) 53:201–11. doi: 10.3109/13880209.2014.913296
51. Ghosh S, Gomes A. Hepatoprotective efficacy of gold nanoparticle synthesized by green method using trigonella foenum-graecum seed extract. Transl Med. (2016) 6:1–5. doi: 10.4172/2161-1025.1000190
52. Suresh P, Kavitha C, Babu SM, Reddy VP, Latha AK. Effect of ethanol extract of Trigonella foenum graecum (Fenugreek) seeds on Freund's adjuvant-induced arthritis in albino rats. Inflammation. (2012) 35:1314–21. doi: 10.1007/s10753-012-9444-7
53. Benayad Z, Gómez-Cordovés C, Es-Safi NE. Characterization of flavonoid glycosides from fenugreek (Trigonella foenum-graecum) crude seeds by HPLC-DAD-ESI/MS analysis. Int J Mol Sci. (2014) 15:20668–85. doi: 10.3390/ijms151120668
54. Li X, Lu S, Wang H, Li G, He YF, Liu XY, et al. Effects of the fenugreek extracts on high-fat diet-fed and streptozotocin-induced type 2 diabetic mice. Anim Model Exp Med. (2018) 1:68–73. doi: 10.1002/ame2.12004
55. Pournamdari M, Mandegary A, Sharififar F, Zarei G, Zareshahi R, Asadi A, et al. Anti-inflammatory subfractions separated from acidified chloroform fraction of fenugreek seeds (Trigonella foenum-graecum L.). J Diet Suppl. (2018) 15:98–107. doi: 10.1080/19390211.2017.1326431
56. Ktari N, Trabelsi I, Bardaa S, Triki M, Bkhairia I, Ben Slama-Ben Salem R, et al. Antioxidant and hemolytic activities, and effects in rat cutaneous wound healing of a novel polysaccharide from fenugreek (Trigonella foenum-graecum) seeds. Int J Boil Macromol. (2017) 95:625–34. doi: 10.1016/j.ijbiomac.2016.11.091
57. Veldhoen M, Hocking RJ, Atkins CJ, Locksley RM, Stockinger B. TGFbeta in the context of an inflammatory cytokine milieu supports de novo differentiation of IL-17-producing T cells. Immunity. (2006) 24:179–89. doi: 10.1016/j.immuni.2006.01.001
58. Akira S, Takeda K. Toll-like receptor signalling. Nat Rev Immunol. (2004) 4:499–511. doi: 10.1038/nri1391
59. Hwang D. Modulation of the expression of cyclooxygenase-2 by fatty acids mediated through toll-like receptor 4-derived signaling pathways. FASEB J. (2001) 15:2556–64. doi: 10.1096/fj.01-0432com
60. Azam S, Jakaria M, Kim IS, Kim J, Haque ME, Choi DK. Regulation of Toll-Like Receptor (TLR) signaling pathway by polyphenols in the treatment of age-linked neurodegenerative diseases: focus on TLR4 signaling. Front Immunol. (2019) 10:1000. doi: 10.3389/fimmu.2019.01000
61. Duan D, Zhang S, Li X, Guo H, Chen M, Zhang Y, et al. Activation of the TLR/MyD88/NF-κB signal pathway contributes to changes in IL-4 and IL-12 production in piglet lymphocytes infected with porcine circovirus type 2 in vitro. PLoS ONE. (2014) 9:e97653. doi: 10.1371/journal.pone.0097653
62. West AP, Koblansky AA, Ghosh S. Recognition and signaling by toll-like receptors. Annu Rev Cell Dev Biol. (2006) 22:409–37. doi: 10.1146/annurev.cellbio.21.122303.115827
63. Kawai T, Akira S. Signaling to NF-kappaB by Toll-like receptors. Trends Mol Med. (2007) 13:460–9. doi: 10.1016/j.molmed.2007.09.002
64. Hou L, Meng M, Chen Y, Wang C. A water-soluble polysaccharide from Grifola frondosa induced macrophages activation via TLR4-MyD88-IKKβ-NF-κB p65 pathways. Oncotarget. (2017) 8:86604–14. doi: 10.18632/oncotarget.21252
65. Ji W, Peng X, Lou T, Wang J, Qiu W. Total flavonoids from Tetrastigma hemsleyanum ameliorates inflammatory stress in concanavalin A-induced autoimmune hepatitis mice by regulating Treg/Th17 immune homeostasis. Inflammopharmacology. (2019) 27:1297–307. doi: 10.1007/s10787-019-00599-0
Keywords: fenugreek seed extract, broiler, immunity, production performance, NF-κB signaling pathway
Citation: Huang H, Wang X, Yang L, He W, Meng T, Zheng K, Xia X, Zhou Y, He J, Liu C, Zou S and Xiao D (2022) The Effects of Fenugreek Extract on Growth Performance, Serum Biochemical Indexes, Immunity and NF-κB Signaling Pathway in Broiler. Front. Vet. Sci. 9:882754. doi: 10.3389/fvets.2022.882754
Received: 25 March 2022; Accepted: 25 April 2022;
Published: 23 June 2022.
Edited by:
Bing Dong, China Agricultural University, ChinaReviewed by:
Zhigang Song, Shandong Agricultural University, ChinaCopyright © 2022 Huang, Wang, Yang, He, Meng, Zheng, Xia, Zhou, He, Liu, Zou and Xiao. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Dingfu Xiao, eGlhb2RpbmdmdTIwMDFAMTYzLmNvbQ==
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.