
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Vet. Sci. , 28 April 2022
Sec. Veterinary Infectious Diseases
Volume 9 - 2022 | https://doi.org/10.3389/fvets.2022.880152
This article is part of the Research Topic Antimicrobials Alternatives for the Prevention and Treatment of Veterinary Infectious Diseases View all 16 articles
 Liheng Zhang1,2
Liheng Zhang1,2 Xianghui Li2Xinghui Song1Chuanzhou Bian2Xiangtao Kang3Junqiang Zhao4
Xianghui Li2Xinghui Song1Chuanzhou Bian2Xiangtao Kang3Junqiang Zhao4 Hongxing Qiao2*
Hongxing Qiao2* Yanzhang Gong1*
Yanzhang Gong1*Endophytic fungus represents microorganisms existing within the healthy plant organs, which can significantly influence metabolic product production in plants, a process with great research value and broad prospects for development. To investigate the effect of fermentation with probiotic cultures on the endophytic fungal diversity and composition of Astragalus membranaceus, we used single-molecular, real-time sequencing (Pacific Biosciences) for 18S ribosomal RNA (rRNA) sequencing. The results showed that the endophytic fungi of A. membranaceus mainly belonged to Aspergillus, Penicillium, Cystofilobasidium, Candida, Guehomyces, and Wallemia. Furthermore, the endophytic fungal diversity and abundance of A. membranaceus were more variable after fermentation with Enterococcus faecium and/or Lactobacillus plantarum. Our data lays a solid and comprehensive foundation for further exploration of endophytic fungi from A. membranaceus as potential sources of functional compounds.
Astragalus membranaceus (Fisch.)Bge. as part of one of the largest genera in the Leguminosae family, has been used as a traditional tonic for promoting digestion and metabolism, enhancing immunity, and accelerating injury/wound healing (1, 2).
Astragalus is a traditional Chinese herbal medicine that has been widely used by humans for hundreds of years. There are few studies on the use of A. membranaceus in livestock. With the rapid development of intensive livestock production, there is an urgent need for sustainable and environmentally friendly practices in this area. A. membranaceus is also utilized to be the supplementary agent in anti-tumor treatment, which exhibits diverse effects like anti-bacterium, anti-virus, antioxidation, anti-inflammation, and hidroschesis. However, the extraction of A. membranaceus' functional ingredients is limited because of plant cell wall recalcitrance, while microbial fermentation offers a possibility to improve A. membranaceus utilization efficiency. Previous studies show that A. membranaceus fermentation by Lactobacillus plantarum (L. plantarum) and/or Enterococcus faecium (E. faecium) can increase Astragalus polysaccharides, flavonoids, and saponins contents, whereas A. membranaceus after fermentation has high microbial abundance and diversity (3). It can be speculated that the fermentation of A. membranaceus elevates the content of active components by affecting its endophytic fungi.
Microorganisms exist in almost all living and non-living niches on the earth, like in thermal vents, deep rocks, sediments, or under extreme conditions like marine habitats and deserts (4). Endophytes are microorganisms (mostly bacteria and fungi) that reside in plants during one part or the entire life cycle with no instigation of distinct infection symptoms or visible manifestation of diseases in their hosts (5, 6). Endophytic fungi are meiosporic or mitosporic ascomycetes and present great biological diversity, and at least one species exists in every plant (7). In addition, microorganisms also produce metabolites to promote development, repellents for pests and insects, protectors, and antibacterial agents to resist plant pathogens under conditions like stress (8, 9). Moreover, investigations have clearly demonstrated that plant survival and health are strongly dependent on their endophytes (10, 11). Endophytic fungi may generate specific secondary metabolites, and these may be utilized in fields like pharmaceutics and agriculture (8). Paclitaxel (Taxol), a commonly used antitumor agent, can be generated by Taxus brevifolia and subsequently via Taxomyces andreanae (also an endophytic fungus), which accounts for a distinct example for the production of endophyte-plant metabolites (4).
Recently, research mainly aimed to isolate endophytic fungi from A. membranaceus in China and examine their antimicrobial activities. Several reports have stated that A. membranaceus endophytic fungal species can generate diverse secondary metabolites with bioactivity (12). However, endophytic fungal species in fermented A. membranaceus have never been studied. Here, we aimed to detect and compare endophytic fungi in A. membranaceus under fermentation via L. plantarum and E. faecium by' single-molecular, real-time (SMRT) sequencing (Pacific Biosciences), the third next-generation sequencing platform.
Astragalus membranaceus dried roots were provided by Gansu Huisen Pharmaceutical Co., Ltd. (Minxian County, Gansu Province), whereas the isolation and preservation of E. faecium (CGMCC 1.130) and L. plantarum (CGMCC 1.557) were completed at the China General Microbial Species Preservation Center (CGMCC, Beijing, China). The preparation of fermented A. membranaceus was as previously reported (3). In short, A. membranaceus was first ground into powder and passed through the 100-mesh filter. Thereafter, we classified 10,000 g dried A. membranaceus powders into 4 groups: A, B, C, and D. This work inoculated group A by 106 colony-forming units (CFU)/g E. faecium, group B by 106 CFU/g L. plantarum, group C with 106 CFU/g E. faecium combined with L. plantarum, and group D by lactobacillus selector (agar) medium as a blank control. Each group had three replicates. Then, fermentation was carried out in plastic bags (35 × 45 mm, Zhejiang Jinhu Company, China), followed by vacuum pumping and sealing by vacuum packaging of the bags. Thereafter, the mixture was fermented under 37°C and anaerobic situation for a 6-day period for producing the fermented A. membranaceus. On the 6th day of fermentation, the four groups were sampled. Three repetitive samples were collected in each group and mixed into one sample for sequencing. This study labeled the samples as A, B, C, and D.
Four samples were collected from each group (200 mg each) after fermentation, followed by immediate freezing under −196°C to extract the DNA subsequently. Afterward, this study utilized the Qubit 2.0 fluorometer (Invitrogen, Carlsbad, CA, USA) for quantifying the DNA samples. We evaluated the quality of the extracted DNA through 0.8% agarose gel as well as 260/280 mm spectrophotometry. Before further analysis, the DNA extracted was preserved under −20°C. PCR was performed to amplify 18S ribosomal RNA (rRNA) of fungi, followed by being sequenced by SMRT barcodes using the primers below, 27F (5′-CCGTGTTTCAAGACGG-3′, forward) and 1492R (5′-CTTGGTCATATA GAGTAA-3′, reverse). The 25-μl amplification volume was prepared, containing 5 μl of 5 × GC buffer, 5 μl of 5 × reaction buffer, 2 μl of 2.5 mm dNTP, 2 μl DNA template, 1 μl of 10 μm reverse primer, 1 μl of 10 μm forward primer, 0.25 μl Q5 DNA polymerase, and 8.75 μLddH2O. The PCR was implemented as follows: 2-min predenaturation step under 98°C; followed by 15-s under 98°C, 10-s under 55°C and 30-s under 72°C for 30 cycles; and eventual 5-min elongation under 72°C. The whole 18S rRNA length was sequenced by the Shanghai Passenger Company.
The original reads generated by cyclic consensus sequencing were used to render the accuracy of prediction up to 90%. We extracted high-quality sequences using the QIIME (Shanghai Personalbio Technology Co., Ltd, Shanghai, China) package (Quantitative Insights into Microbial Ecology, v1.8, https://docs.qiime2.org/2019.4/tutorials/) (13). In addition, we utilized USEARCH (Shanghai Personalbio Technology Co., Ltd, Shanghai, China) (v5.2.236, http://www.drive5.com/usearch/) for excluding chimeras and clustering the clean sequence data into operational taxonomic units (OTUs) at the similarity degree of 97% (14). Later, this work adopted UNITE fungal ITS database (https://unite.ut.ee/) for sequence comparison through blast search (15).
Moreover, ACE, Chao 1, Simpson, and Shannon indicators were utilized to estimate the alpha-diversity. The R software (Bell Lab, Lucent Technologies, Beijing, China) was also employed to classify data from 50 species with the highest abundances, later, the heat map was generated. This study also carried out principal component analysis (PCA) with R software (16).
The original sequence readings were imported into the National Center for Biotechnology Information Sequence Read Archive with accession No. SAMN07411593-SAMN07411601.
Full-length 18S rRNA gene was subject to SMRT sequencing for obtaining the precise fungal profiles from all four samples. We acquired altogether 99,923 sequence readings in the four samples, resulting in 24,980 readings per sample on average. Thereafter, this study determined Chao 1, ACE, Simpson, and Shannon indicators, together with the diversity of fungal species in each group is shown in Table 1. According to Table 1, Chao1, ACE, and Shannon indicators of group B were the highest, while the Simpson index showed little difference among the four groups.
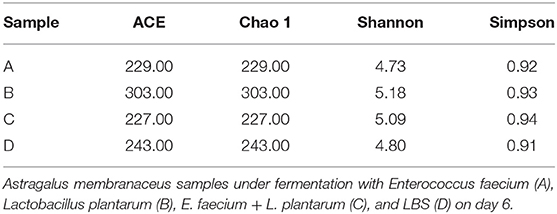
Table 1. Estimated alpha-diversity indices of 18S ribosomal RNA (rRNA) gene libraries for four samples used for sequencing.
The comparative analysis of the total OTUs of groups A, B, C, and D was carried out (Figure 1). Altogether, 1,002 OTU types were detected from all groups, and 85 OTU types were common across them. Eighty-eight OTUs were common among groups A, B, and C, whereas 107, 160, and 163 OTU types of group D were specifically associated with groups A, B, and C, respectively.
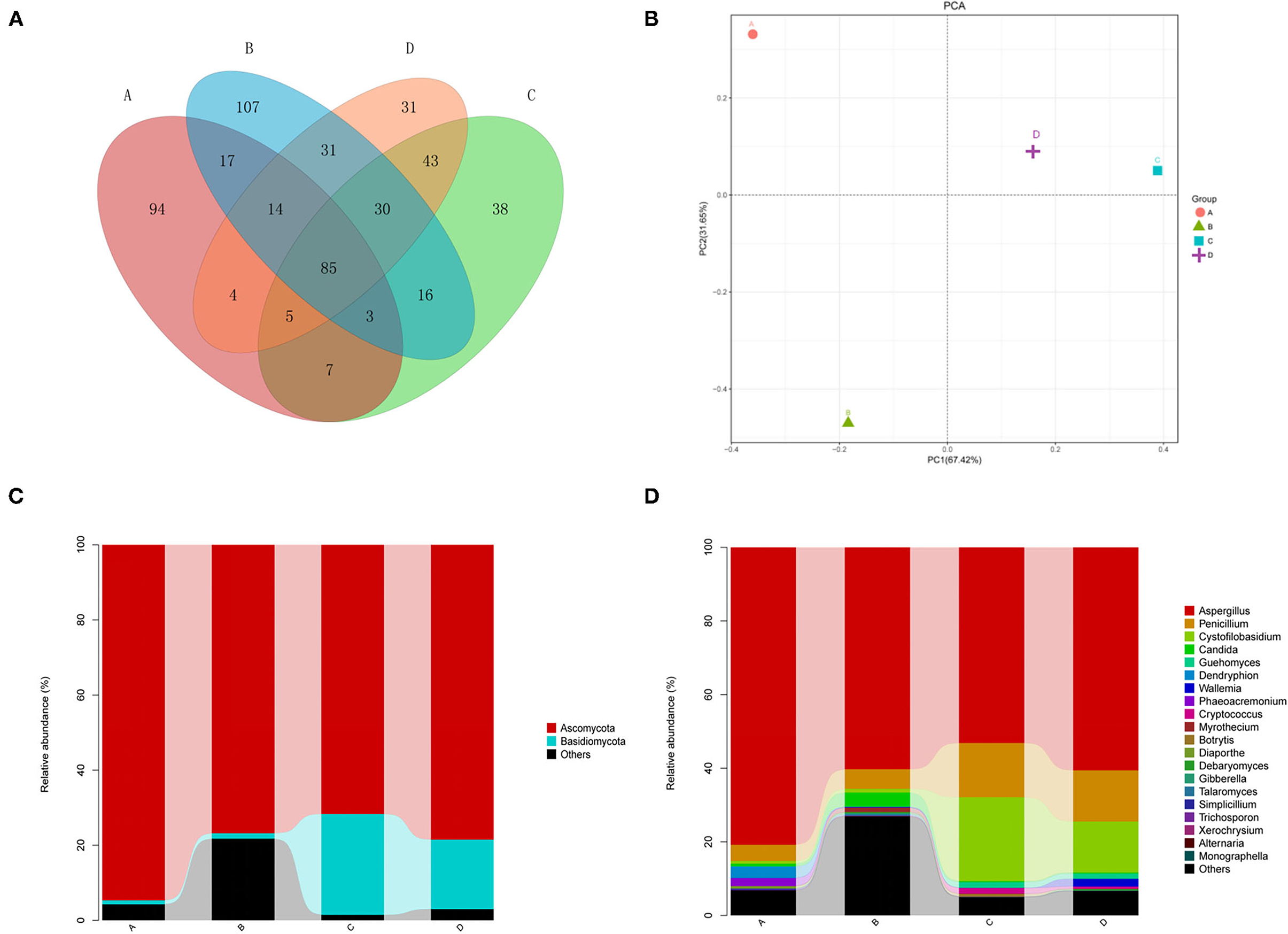
Figure 1. (A) Venn diagram showing the distributions of operational taxonomic units (OTUs) across 4 groups. Astragalus membranaceus samples under fermentation with Enterococcus faecium. E. faecium (A), Lactobacillus plantarum (B), E. faecium + L. plantarum (C), and LBS (D) on day 6. (B) Two-dimensional principal component analysis plot of the samples. A. membranaceus samples under fermentation with E. faecium (A), L. plantarum (B), E. faecium + L. plantarum (C), and LBS (D) on day 6. (C) Phylum levels in samples. A. membranaceus samples under fermentation with E. faecium (A), L. plantarum (B), E. faecium + L. plantarum (C), and LBS (D) on day 6. (D) Genus levels in samples. membranaceus samples under fermentation with E. faecium (A), L. plantarum (B), E. faecium + L. plantarum (C), and LBS (D) on day 6.
Two-dimensional PCA at the genus level was carried out for evaluating different fungal community structures among 4 groups (Figure 1). PC1/PC2 of the PCA at the genus level explained 67.42 and 31.65% of the total variation, separately. As revealed by PCA, all four groups were grouped separated, and groups C and D were closer than the others, indicating that the fungal communities of groups C and D were similar.
Our results showed that the dominant fungal phyla of A. membranaceus were Ascomycota and Basidiomycota (Figure 1). The abundances of Ascomycota in 4 groups were 94.7, 76.8, 71.7, and 78.5%. The abundances of Basidiomycota in the 4 groups were 1.1, 1.5, 26.9, and 18.5%. The abundances of unclassified phyla in the 4 groups were 4.2, 21.7, 1.4, and 2.9%. These results indicated that the fermentation of A. membranaceus by L. plantarum or E. faecium had a greater impact on endophytic fungi of A. membranaceus, with a significantly decreased abundance of Basidiomycota. Furthermore, L. plantarum-fermented A. membranaceus significantly increased the abundance of unclassified phyla.
According to Figure 1, the dominant fungal genera of A. membranaceus were Aspergillus, Penicillium, Cystofilobasidium, Candida, Guehomyces, and Wallemia. The abundances of Aspergillus in 4 groups were 80.8, 60.3, 53.1, and 60.6%. The abundances of Penicillium in the 4 groups were 4.4, 5.3, 14.7, and 13.9%. The abundances of Cystofilobasidium were.7, 1.0, 22.8, and 13.8%. The abundances of Candida were 0.7, 3.8, 0.3, and 0.3%. The abundances of Guehomyces were 0.1, 0.1, 1.5, and 1.4%. The abundances of Wallemia were 0.0, 0.2, 0.1, and 2.1%. The abundances of unclassified genera were 4.9, 26.1, 2.9, and 5.1%. The above results suggested that the fermentation of A. membranaceus by E. faecium increased the abundance of Aspergillus, while the fermentation of A. membranaceus by E. faecium or L. plantarum reduced the abundance of Penicillium and Cystofilobasidium. Moreover, L. plantarum-fermentation of A. membranaceus significantly increased the abundance of unclassified genera.
Endophytic fungal species in A. membranaceus were mainly Aspergillus cibarius, Aspergillus piperis, Penicillium polonicum, Cystofilobasidium infirmominiatum, and Plantae sp (Figure 2). The abundances of Aspergillus cibarius in the 4 groups were 41.7, 49.3, 40.3, and 54.0%. The abundances of Aspergillus piperis were 39.1, 11.0, 12.8, and 6.5%. The abundances of Penicillium polonicum were 4.4, 5.3, 14.4, and 13.6%. The abundances of Cystofilobasidium infirmominiatum were 0.6, 1.0, 20.4, and 12.5%. The abundances of Plantae were 4.2, 21.7, 1.4, and 2.9%, respectively. The above results show that the fermentation of A. membranaceus by E. faecium mainly increased the abundance of Aspergillus piperis and reduced that of Penicillium polonicum and Cystofilobasidium infirmominiatum. Meanwhile, fermentation of A. membranaceus by L. plantarum reduced the abundance of Penicillium polonicum and Cystofilobasidium infirmominiatum but increased that of Plantae.

Figure 2. Species levels in samples. membranaceus samples under fermentation with E. faecium (A), L. plantarum (B), E. faecium + L. plantarum (C), and LBS (D) on day 6.
This study clustered those 50 species with the highest abundances by R software. As revealed by the heat map (Figure 3), the highly abundant species of fungi in groups A, B, and C were much better than those in group D. Group A had a higher abundance of Trichosporon cutaneum, Simplicillium obclavatum, Aspergillus.piperis, Cordycipitaceae sp., Sordariomycetes sp., Dendryphion nanum, Exobasidiomycetes sp., Hypocreales sp., Microascaceae sp., Phaeoacremonium hungaricum, and Diaporthe citri. Group B had a higher abundance of Myrothecium verrucaria, Talaromyces marneffei, Xerochrysium xerophilum, Plantae sp., Candida glaebosa, Torulaspora delbrueckii, Ascomycota sp., and Gibberella tricincta. In group C, Cryptococcus albidus, Penicillium cvjetkovicii, Cryptococcus magnus, Monographella cucumerina, Leucosporidium sp., Botrytis caroliniana, and Myrothecium roridum were the most abundant species. However, only Wallemia sebiand and Aspergillus cibarius were particularly abundant species in group D. The above results demonstrated that the endophytic fungal abundance of A. membranaceus can be changed by fermentation, and the change was closely related to the bacteria used for fermentation.
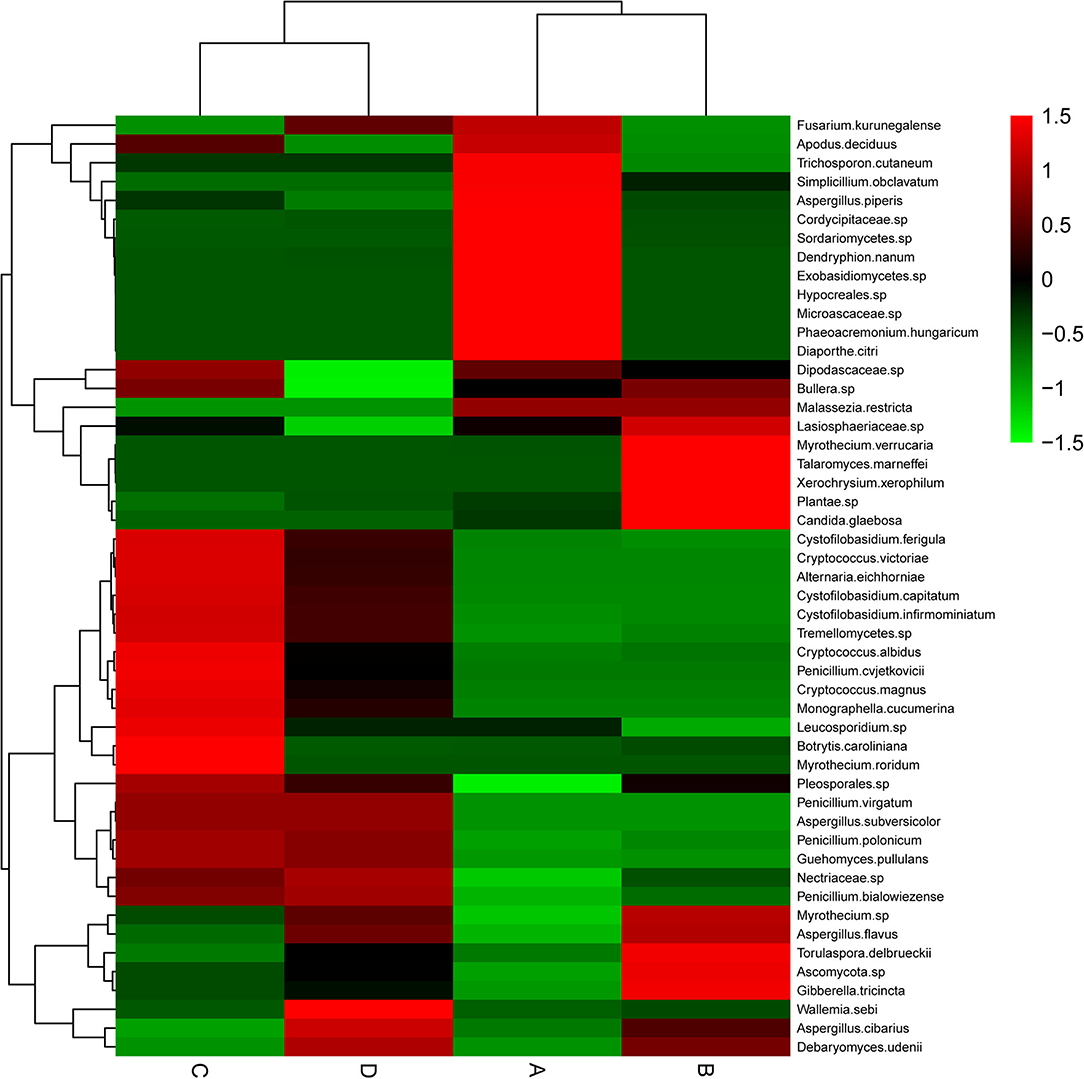
Figure 3. Heat map of endophytic fungal community composition with cluster analysis. A. membranaceus samples under fermentation with E. faecium (A), L. plantarum (B), E. faecium + L. plantarum (C), and LBS (D) on day 6. Green and red colors stand for low and high abundances, respectively.
Endophytic fungi are extensively distributed within healthy plant organs, which are the critical parts in the plant micro-ecosystems during the long-term evolution, significantly influencing plant metabolite production and affecting medicinal plants-derived crude medicine quantity and quality (7). At present, the cultivation method is primarily adopted for investigating endophytic fungal diversity, but this method is limited by the low authenticity of endophytic fungal community composition. With the rapid development of sequencing technology, SMRT sequencing can more accurately describe microbial diversity (17). Previously, 16S rRNA gene-SMRT sequencing was conducted for analyzing A. membranaceus diversity fermented by L. plantarum and E. faecium. As a result, it was appropriate to assess fermented A. membranaceus quality using the SMRT sequencing system (3). For exploring how E. faecium and L. plantarum fermentation affected the endophytic fungi of A. membranaceus, 18S rRNA gene-SMRT sequencing was used for analyzing A. membranaceus fermented endophytic fungal composition and diversity.
Based on the ACE, Chao, and Shannon indices, A. membranaceus fermented by L. plantarum possessed the highest fungal community diversity. Furthermore, A. membranaceus fermented by L. plantarum exhibited the highest number of OTUs by sequencing, suggesting that L. plantarum fermentation increased the abundance of endophytic fungal diversity of A. membranaceus. In general, the endophytic fungal diversity of A. membranaceus was relatively low, independent of fermentation. Composition analysis of the endophytic fungal community showed that the dominant fungal phyla of A. membranaceus were Ascomycota and Basidiomycota, and the abundance of Ascomycota was increased by E. faecium fermentation and decreased by E. faecium and L. plantarum fermentation. However, E. faecium and L. plantarum fermentation increased the abundance of Basidiomycota. Aspergillus was the genus with the greatest relative abundance among the endophytic fungi of A. membranaceus. In Asian cuisines, Aspergilli represent the critical microorganisms that can enhance the flavors, nutrients, and textures of fermented foods (18). Moreover, the use of Aspergillus fungus for fermenting the native A. membranaceus significantly elevated phenolic levels of A. membranaceus; besides, A. membranaceus after fermentation displayed significantly increased antioxidation capacity (19). According to our results, the fermented E. faecium had markedly increased Aspergillus abundance. Overall, the results showed that fermentation can change the proportions of endophytic fungi of A. membranaceus, which might change the metabolite contents of A. membranaceus processed by fermentation.
A recent study on the saponins of Dipsacus asperoides showed that endophytic fungi from taproots generated metabolites with bioactivity close to plant hosts, and their quantity showed positive relation to Dipsacus saponin content (20).
Some studies used high-throughput sequencing technology to analyze and identify the endophytic fungal community of A. membranaceus, explored the diversity information of endophytic fungi of A. membranaceus, solved the limitations of non-culturable and low-abundance fungi that could not be detected in traditional methods, and provided a reference for the use of endophytic fungi in A. mongholicus for pure fermentation or mixed fermentation to transform glycosides in Astragalus.
High-throughput sequencing in this study may provide a partial answer to previous research demonstrating that the active substances of A. membranaceus, such as Astragalus polysaccharides, flavonoids, saponins, and organic acids, are changed by L. plantarum and E. faecium fermentation (3). Recently, endophytic fungi originating from Astragalus species have been used for the biotransformation of Astragalus sapogenins (21). A. chinensis-derived endophytic fungi are verified to be bactericides and fungicides (22). It is exciting to speculate that Astragalus polysaccharides or flavonoids may be obtained from the endophytic fungi that colonize A. membranaceus. Our findings provide evidence for this possibility.
In summary, 18S rRNA gene-SMRT sequencing was used for analyzing endophytic fungal composition and diversity from fermented A. membranaceus. Our study presents the most comprehensive data for a more extensive follow-up study.
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author/s.
YG and XK shouldered the responsibility of studying conception and design. LZ and XL were in charge of experiment implementation. XS and HQ were responsible for statistical analysis and were in charge of manuscript writing. JZ and CB isolated and preserved microbial strains. All authors revised the study and approved the final version for submission.
This work was supported by the Natural Science Foundation of Henan Province of China (202300410189), the Research and Innovation Team of Henan University of Animal Husbandry and Economy (2018KYTD13), and the Key Discipline of Preventive Veterinary Medicine of Henan University of Animal Husbandry and Economy (mxk2016102 and 2021HNUAHEDF004).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
A. membranaceus, Astragalus membranaceus; L. plantarum, Lactobacillus plantarum; E. faecium, Enterococcus faecium; SMRT, single-molecule, real-time; CFU, colony forming units; OTUs, operational taxonomic units; PCA, Principal component analysis.
1. Kim YB, Thwe AA Li X, Tuan PA, Lee S, Lee JW, et al. Accumulation of astragalosides and related gene expression in different organs of astragalus membranaceus Bge. var Mongholicus (Bge). Molecules. (2014) 19:10922–35. doi: 10.3390/molecules190810922
2. Wang X, Li Y, Shen J, Wang S, Yao J, Yang X. Effect of Astragalus polysaccharide and its sulfated derivative on growth performance and immune condition of lipopolysaccharide-treated broilers. Int J Biol Macromol. (2015) 76:188–94. doi: 10.1016/j.ijbiomac.2015.02.040
3. Qiao H, Zhang X, Shi H, Song Y, Bian C, Guo A. Assessment of the physicochemical properties and bacterial composition of Lactobacillus plantarum and Enterococcus faecium-fermented Astragalus membranaceus using single molecule, real-time sequencing technology. Sci Rep. (2018) 8:11862. doi: 10.1038/s41598-018-30288-x
4. Tanvir R, Javeed A, Bajwa AG. Endophyte bioprospecting in South Asian medicinal plants: an attractive resource for biopharmaceuticals. Appl Microbiol Biotechnol. (2017) 101:1831–44. doi: 10.1007/s00253-017-8115-x
5. Kouipou Toghueo RM, Boyom FF. Endophytic fungi from terminalia species: a comprehensive review. J Fungi. (2019) 5:43. doi: 10.3390/jof5020043
6. Strobel Gary A. Endophytes as sources of bioactive products. Microbes and Infection. (2003) 5:535–44. doi: 10.1016/S1286-4579(03)00073-X
7. Min J, Ling C, Hai-Liang X, Cheng-Jian Z, Khalid R, Ting H, et al. A friendly relationship between endophytic fungi and medicinal plants: a systematic review. Front Microbiol. (2016) 7:906. doi: 10.3389/fmicb.2016.00906
8. Golinska P, Wypij M, Agarkar G, Rathod D, Dahm H, Rai M. Endophytic actinobacteria of medicinal plants: diversity and bioactivity. Antonie Van Leeuwenhoek. (2015) 108:267–89. doi: 10.1007/s10482-015-0502-7
9. Ryan RP, Germaine K, Franks A, Ryan DJ. Dowling DNBacterial endophytes: recent developments and applications. FEMS Microbiol Lett. (2008) 278:1–9. doi: 10.1111/j.1574-6968.2007.00918.x
10. Hardoim PR, Van Overbeek LS, Berg G, Pirttilä AM, Compant S, Campisano A, et al. The hidden world within plants: ecological and evolutionary considerations for defining functioning of microbial endophytes. Microbiol Molec Biol Rev. (2015) 79:293–320. doi: 10.1128/MMBR.00050-14
11. Potshangbam M, Devi SI, Sahoo D, Strobel GA. Functional Characterization of Endophytic Fungal Community Associated with Oryza sativa L. and Zea mays L. Front Microbiol. (2017) 8:325. doi: 10.3389/fmicb.2017.00325
12. Xu YM, Espinosa-Artiles P, Liu MX, Arnold AE, Gunatilaka AL, Secoemestrin D. A Cytotoxic Epitetrathiodioxopiperizine and Emericellenes A–E, Five Sesterterpenoids from Emericella sp. AST0036, a Fungal Endophyte of Astragalus lentiginosus. J Nat Prod. (2013) 76:2330–2336. doi: 10.1021/np400762k
13. Caporaso JG, Kuczynski J, Stombaugh J, Bittinger K, Bushman FD, Costello EK, et al. QIIME allows analysis of high-throughput community sequencing data. Nat Methods. (2010) 7:335–6. doi: 10.1038/nmeth.f.303
14. Edgar RC. Search and clustering orders of magnitude faster than BLAST. Bioinformatics. (2010) 26:2460–1. doi: 10.1093/bioinformatics/btq461
15. Spouge JL. Towards a unified paradigm for sequence-based identification of fungi. Molec Ecol. (2013) 22:5271–7. doi: 10.1111/mec.12481
16. Lozupone C, Knight RJ. UniFrac: a new phylogenetic method for comparing microbial communities. Appl Environ Microbiol. (2005) 71:8228–35. doi: 10.1128/AEM.71.12.8228-8235.2005
17. Hestand MS, Ameur AJ. The versatility of SMRT sequencing. Genes. (2019) 10:24. doi: 10.3390/genes10010024
18. Park HS, Jun SC, Han KH, Hong SB Yu JH. Diversity, application, and synthetic biology of industrially important aspergillus fungi. Adv Appl Microbiol. (2017) 100:161–202. doi: 10.1016/bs.aambs.2017.03.001
19. Sheih IC, Fang TJ, Wu TK, Chang CH, Chen RY. Purification and properties of a novel phenolic antioxidant from radix astragali fermented by aspergillus oryzae M29. J Agric Food Chem. (2011) 59:6520–5. doi: 10.1021/jf2011547
20. Gong A, Zhou T, Xiao C, Jiang W, Zhou Y, Zhang J, et al. Association between dipsacus saponin VI level and diversity of endophytic fungi in roots of Dipsacus asperoides. J Microbiol Biotechn. (2019) 35:42. doi: 10.1007/s11274-019-2616-y
21. Ekiz G, Yilmaz S, Yusufoglu H, Ballar P, Bedir E. Microbial transformation of cycloastragenol and astragenol by endophytic fungi isolated from astragalus species. Natural Products. (2019) 82:2979–85. doi: 10.1021/acs.jnatprod.9b00336
Keywords: Astragalus membranaceus, fermentation, endophytic fungi, SMRT sequencing, 18S rRNA
Citation: Zhang L, Li X, Song X, Bian C, Kang X, Zhao J, Qiao H and Gong Y (2022) Assessment of the Endophytic Fungal Composition of Lactobacillus plantarum and Enterococcus faecalis-Fermented Astragalus membranaceus Using Single-Molecule, Real-Time Sequencing Technology. Front. Vet. Sci. 9:880152. doi: 10.3389/fvets.2022.880152
Received: 21 February 2022; Accepted: 03 March 2022;
Published: 28 April 2022.
Edited by:
Kun Li, Nanjing Agricultural University, ChinaReviewed by:
Junqing Guo, Henan Academy of Agricultural Sciences (HNAAS), ChinaCopyright © 2022 Zhang, Li, Song, Bian, Kang, Zhao, Qiao and Gong. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Hongxing Qiao, enptenFoeEAxNjMuY29t; Yanzhang Gong, cG91bHRyeUBtYWlsLmh6YXUuZWR1LmNu
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.