 Yicheng Wu
Yicheng Wu Ao Ren1,3†
Ao Ren1,3† Xiaokang Lv
Xiaokang Lv Tao Ran
Tao Ran Chuanshe Zhou
Chuanshe Zhou Zhiliang Tan
Zhiliang Tan
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Vet. Sci., 24 June 2022
Sec. Animal Nutrition and Metabolism
Volume 9 - 2022 | https://doi.org/10.3389/fvets.2022.852251
This article is part of the Research TopicWomen in Animal Nutrition and Metabolism: 2021View all 6 articles
This study was conducted to investigate the protective effects of mycotoxin adsorbent galactomannan oligosaccharides (GMOS) on growth performance, fermentation parameters, mycotoxins residues, serum biochemistry and oxidative stress parameters of the goats. The in vitro test indicated that 0.05% GMOS outperformed yeast cell wall (YCW) and montmorillonite (MMT) in aflatoxins absorption. Then 20 3-month-old Xiangdong black goats (15.0 ± 1.9 kg) were randomly divided into two dietary treatments for the animal test. The control group (CON group) was fed a multi-mycotoxins contaminated diet, whereas the experimental group (GMOS group) received multi-mycotoxins contaminated diet plus 0.05% GMOS. The trail lasted for 60 days, with 12 days of adaptation period and 48 days of formal experiment period. There were no treatment effects (P > 0.10) on growth performance, serum antioxidant capacity and activities of serum aspartate aminotransferase (AST), alanine aminotransferase (ALT), and alkaline phosphatase (ALP). The concentrations of zearalenone in the rumen were lower (P < 0.05) in the GMOS group. GMOS significantly reduced (P < 0.05) propionate concentration in the cecum, resulting in a rise (P < 0.01) in acetate/propionate ratio in GMOS as compared to CON. Goats of GMOS exhibited considerably greater (P < 0.05) levels of creatine kinase but lower (P = 0.02) levels of creatinine than CON. Compared with CON, GMOS supplementation significantly increased (P < 0.05) platelet count (PLT), platelet volume distribution width (PDW), and platelet hematocrit (PCT), while decreased (P < 0.05) albumin content (ALB). The 0.05% GMOS protected goats in ruminal fermentation parameters, mycotoxins residues and serum biochemistry. Moreover, GMOS had no adverse effect on goat health. To our knowledge, this is the first report of GMOS in small ruminants. These findings suggested the feasibility of dietary GMOS as a health-maintaining addictive in goat diets.
The prevalence of mycotoxins in feed is one of the major factors affecting feed quality, particularly in the humid and rainy areas of southern China (1). Mycotoxins can affect animal health, with poultry being the most sensitive, followed by pigs and ruminants (2). Toxins such as aflatoxins B1 (AFB1), ochratoxin A (OTA), zearalenone (ZEN) and deoxynivalenol (DON) have been detected throughout the food chain, contaminating animal products (1). Therefore, mycotoxins of feed origin are regarded as a significant threat to human and animal health worldwide. Bentonite, Zeolite and activated charcoal are widely used for mycotoxin elimination to reduce their impact (3). However, they bind to minerals and vitamins in the diet (4). As a result, researchers have concentrated their efforts on developing effective methods for removing mycotoxins from the diets without substantial nutrient loss.
Functional oligosaccharides are widely used as feed additives due to their green and non-toxic qualities (5). Both galactomannan oligosaccharides (GMOS) and mannan oligosaccharides (MOS) are functional oligosaccharides (6) and the use of GMOS and MOS as feed additives have been approved in China (7). Previous report has shown that MOS can dramatically promote the proliferation of beneficial bacteria in the gastrointestinal tract (GIT), thereby enhancing animal immune function, and regulating animal growth and metabolism (8). For ruminants, MOS maintained the overall health and performance of crossbred calves (9) and dairy calves (10). In addition, the capacity of MOS to adsorb mycotoxins such as ZEN and OTA had been well established using static in vitro models (11, 12).
Due to the special GIT structure of ruminants, it is commonly considered that rumen microbes could degrade oligosaccharides and weaken their activity (13). However, based on the positive effects of MOS on growth performance in sheep and MOS is an effective organic adsorbent to bind various mycotoxins while posing no toxicity to essential dietary components (6). We hypothesized that adding GMOS to goat diets would be beneficial to the adsorption of mycotoxins in the GIT, hence alleviating the deleterious impact of mycotoxins on animal health. To our knowledge, this is the first report of GMOS on the efficiency of toxin decontamination in small ruminants.
Therefore, the current study sought to evaluate the effects of GMOS supplementation in mycotoxin-contaminated diets on the growth and health of goats and establish a theoretical basis for the use of GMOS as a mycotoxin adsorbent in goat feeds.
The present study included an in vitro mycotoxin adsorption experiment and a goat feeding trial. The in vitro experiment was carried out at the Institute of Subtropical Agriculture, Chinese Academy of Sciences, Changsha, China in May 2019; while the animal study was conducted at the Xiangdong Black Goat Reproduction Center, Liuyang, Hunan province, China from 29 April 2019 to 27 June 2019.
The adsorbents used in the in vitro adsorption test included GMOS, yeast cell wall (YCW) and montmorillonite (MMT). The GMOS was kindly provided by Jiangsu Kangwei Biologic Co., Ltd. (Jiangsu, China); yeast cell wall (YCW) and montmorillonite (MMT), provided by Xuzhou Saifu Biological Co., Ltd. (Xuzhou, China), were used as controls to determine the toxin adsorption efficiencies of GMOS. All other reagents were of analytical grade or higher and purchased from Jiangsu Kangwei Biologic Co., Ltd. (Jiangsu, China). Feed materials were stored at 37°C and 15% humidity for a week to let them mildew (Supplementary Figure 1). Feed samples were collected, dried, pooled, and ground through a 1 mm screen for AFB1 extraction. Five grams of feed powder was weighed into a 50 mL conical tube (Thermo Fisher Scientific, Waltham, USA) that contained 20 mL of a mixture of methanol: water (1:1 v/v). The mixture was vigorously mixed on a multitube vortexer for 10 min, followed by centrifugation at 4,000 rpm for 5 min. The supernatants were collected and filtered through 0.45 μm filters (Thermo Fisher Scientific, Waltham, USA), used as testing samples. Adsorbent solutions of different concentrations (0.00, 0.01, 0.02, 0.03, 0.04, 0.05, 0.06, 0.07, 0.08, and 0.09%) were prepared by diluting GMOS with ultra-pure water. The in vitro adsorption experiment was then performed as described by Saleemi et al. (12). Then the concentration of GMOS with the best toxin adsorption efficiency was selected for further comparision with conventional adsorbents such as YCW (0.05%), MMT (0.05%) following the methods described by Marroquin et al. (14). The concentrations of AFB1 extracted from feed samples treated with or without adsorbents were measured using an enzyme-linked immunosorbent assays (ELISA) kit (Neogen Co., Shanghai, China) according to the description of Aazami et al. (15).
Twenty Xiangdong goats (3-month-old, 15.0 ± 0.1 kg) were randomly divided into two feeding treatments. The control group (CON) was fed a naturally contaminated diet, while the GMOS group (GMOS) was fed the control diet supplemented with 0.05% GMOS, which was added to the naturally contaminated diet in the mixer as the last step in mixing. The ingredients and nutrient compositions, and the initial mycotoxin concentrations of the basal experimental diets are illustrated in Table 1.
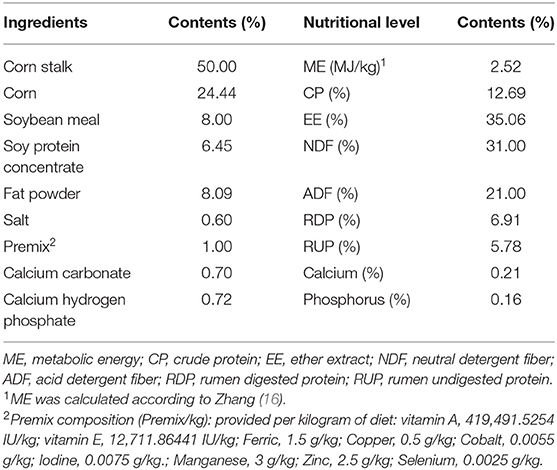
Table 1. Ingredients and nutrient compositions of the experimental diet (DM basis).
The animal study lasted 60 days, with 12 days of pre-feeding and 48 days of formal experiment. All goats were housed individually in stainless steel metabolic cages (150 × 60 × 80 cm) with plastic slatted floors. Goats were individually fed a naturally contaminated diet twice daily (08:00 and 18:00) with the same amount of diet. All goats had free access to clean water. During the feeding trial, body weight was recorded weekly before morning feeding. Feed offered and refusals for each goat were recorded daily throughout the experiment period to calculate the dry matter intake (DMI), average daily gain (ADG), and feed to gain ratio (F/G).
Refusals and experimental diets were sampled for chemical analysis. Feed samples were oven-dried at 65°C for 72 h, ground to pass through a 1-mm sieve and sealed in bags before chemical analysis. Blood samples were obtained by jugular venipuncture before morning feeding on the 27th and 28th of the formal experimental period. A pair of 5 ml blood samples were collected into test tubes (Guangzhou Improve Medical, China), among which one contained EDTA for the routine hematological parameters and the other tube contained no anticoagulants for the serum biochemical analysis. After 30 min still standing, samples were centrifuged at 3,500 rpm for 15 min, 1 ml plasma was taken for antioxidant capacity and biochemical parameters analysis. At the end of the experiment, all goats were fasted for 24 h and water was withheld for 12 h. The animals were weighed, electrically stunned, and exsanguinated under commercial procedures. One hundred milliliter rumen content and 20 g cecum content were extracted upon euthanasia with an alcohol-cleaned spatula and stored at −80°C. Cecal digesta was diluted with deionized water. After vortexing, ~10 ml rumen content and cecum content were centrifuged at 10,000 rpm for 15 min at 4°C, 1 ml supernatant was transferred into tubes containing 0.1 ml of 25% (w/v) metaphosphoric acid, and then frozen at −20°C for subsequent determination of volatile fatty acids (VFAs) and ammonia nitrogen (NH3-N) concentration. Frozen feed materials, ruminal and cecal content samples were thawed, dried, pooled, and ground through a 1 mm screen for AFB1, ZEN and DON extraction.
Feed samples and feces were used for nutrient composition analysis. The dry matter (DM) (method 934.01), ash (method 927.02), ether extract (EE, method 920.39), crude protein (CP, method 976.06), calcium (Ca, method 978.02), total phosphorus (TP, method 946.06) were determined according to AOAC (17). Neutral detergent fiber (NDF) and acid detergent fiber (ADF) contents were determined using a Fibretherm Fiber Analyzer with F57 filter bags (ANKOM A200, ANKOM Technology Corp., Fairport, NY, USA) according to Van Soest et al. (18). Rumen degradable protein (RDP), rumen undegradable protein (RUP) were calculated according to the description of Kekana et al. (19). The AFB1, ZEN and DON residues in the ruminal and cecal contents of the two treatments and the basal diet were carried out according to the description of Guo et al. (20).
Volatile fatty acids and NH3-N concentrations were measured according to Chen et al. (21). Hematological examination of blood samples was performed following the standard method reported by Bafti and Mozaffari (22) using a CELL-DYN 3700 analyzer. Routine hematological parameters include white blood cell (WBC), mononuclear cells (MON), eosinophil granulocyte counts (EOS), basophil granulocyte counts (BAS), neutrophils (NEU), lymphocyte (LYM), red blood cell counts (RBC), platelet count (PLT), platelet volume distribution width (PDW), and platelet hematocrit (PCT) were tested. The detection of serum antioxidant capacity, including total antioxidant capacity (T-AOC), superoxide dismutase (SOD) and malondialdehyde (MDA) levels were performed using commercial kits according to the manufacturer's protocols (Nanjing Jiancheng Bioengineering Institute, Nanjing, China). Serum activities of alanine aminotransferase (ALT), aspartate aminotransferase (AST), alkaline phosphatase (ALP) and creatine kinase (CK) and concentrations of albumin (ALB), blood urea nitrogen (BUN) and creatinine (CREA) were determined using an automatic biochemistry analyzer (Sekisui Medical Co. Ltd., Tokyo, Japan) using commercially available kits (Nanjing Jiancheng Bioengineering Institute, Nanjing, China).
All data were processed preliminarily by Excel. Then the data of the growth performance, toxins concentration, fermentation parameters, serum biochemistry, and hematology indexes of the two groups were analyzed using the Shapiro–Wilk test for normal distribution and followed by student t-test to compare means using SPSS 19.0 (SPSS Inc., Chicago, USA) (23). Statistical significance was defined as P < 0.05, and tendencies at 0.05 ≤ P < 0.10.
The results of GMOS on adsorbing aflatoxins in a naturally contaminated diet are presented in Figure 1. The added level of 0.05% GMOS showed the lowest AFB1 concentration (Figure 1A), suggesting the best efficacy in adsorbing AFB1. Furthermore, the concentration of AFB1 in 0.05% GMOS treat was significantly lower (P < 0.05) than 0.05% YCW and 0.05% MMT treats (Figure 1B), indicating that 0.05% GMOS outperformed traditional adsorbents like YCW and MMT in absorption of aflatoxins.
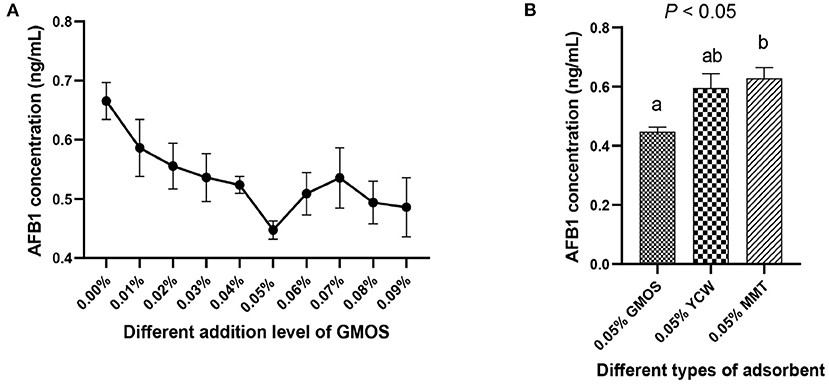
Figure 1. The ability of GMOS to adsorb mycotoxins (Aflatoxins B1, AFB1), which were measured in vitro using ELISA. (A) Effects of different addition levels of GMOS (0.00, 0.01, 0.02, 0.03, 0.04, 0.05, 0.06, 0.07, 0.08, and 0.09%) on AFB1 concentration. (B) Effects of 0.05% GMOS, 0.05% Yeast cell wall (YCM), 0.05% Montesserstone (MMT) on AFB1 concentration. a,b Mean values within a row with different superscript letters were significantly different (P < 0.05).
The DMI, ADG and F/G of goats from GMOS treatment were numerically more remarkable than that of CON goats in Table 2; however, not reach statistically significant differences (P > 0.10).
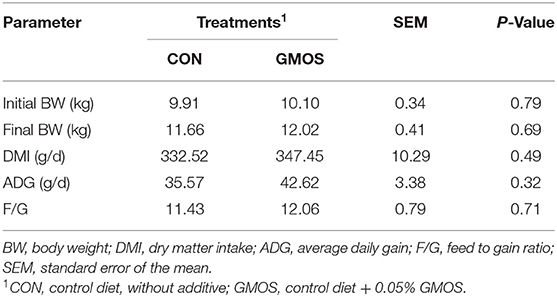
Table 2. Effects of dietary supplementation of GMOS on growth performance in goats.
The concentrations of mycotoxins including AFB1 and DON did not differ between CON and GMOS treatments either in ruminal or cecal digesta; whereas, the concentration of ZEN in rumen was significantly lower (P < 0.01) in GMOS treatment than that in CON (Table 3). Notably, the concentration of ZEN in cecal digesta was greater than that in ruminal digesta.
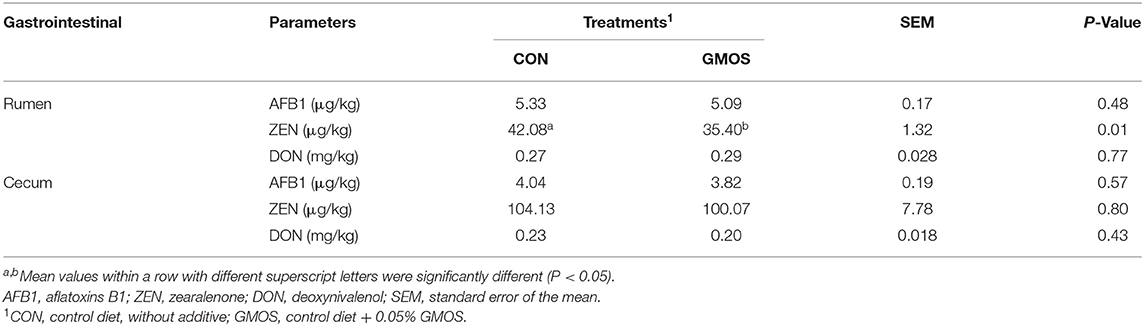
Table 3. Effects of dietary supplementation of GMOS on concentrations of toxins in the gastrointestinal digesta in goats.
The results of NH3-N concentration, total and individual VFA, and A/P ratio was presented in Table 4. The amounts of NH3-N, TVFA, and the molar proportions of butyric acid, isobutyric acid, valeric acid and isovaleric acid in the rumen or cecum did not differ (P > 0.10) between CON and GMOS treatments. In the rumen, the molar proportion of propionic acid was greater (P < 0.05) in GMOS than that in CON, and the molar proportion of acetic acid tended to be lower (P = 0.08) in GMOS than that in CON, thus leading to a tendency of lower (P = 0.07) A/P ratio in GMOS. On the contrary, the molar proportion of acetic acid in the cecum did not differ between CON and GMOS treatments, but the molar proportion of propionic acid was lower (P < 0.05) in GMOS than in CON, resulting in a more significant (P < 0.01) A/P ratio in GMOS.
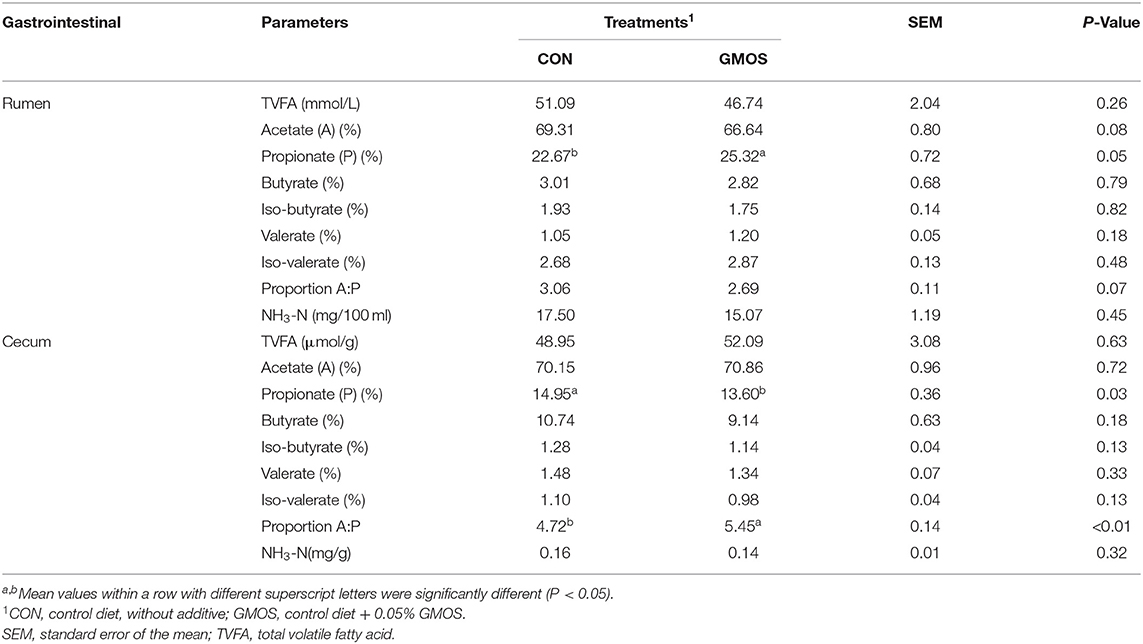
Table 4. Effects of dietary supplementation of GMOS on ruminal and cecal fermentation parameters in goats.
The serum antioxidant indexes including SOD, T-AOC and MDA were not (P > 0.10) different between CON and GMOS treatments (Table 5). Table 6 shows the impact of GMOS supplementation on hematologic parameters and serum biochemical indexes of goats. Compared to CON, the percentage of MON, PDW and PCT and counting of PLT were greater (P < 0.05) in the GMOS supplemented group; whereas, the other indexes did not differ between the two treatments. As for the biochemical parameters, the Albumin (ALB) and creatinine (CREA) levels were significantly lower (P < 0.05) in the GMOS group compared to the CON group. In contrast, the serum creatine kinase (CK) concentrations were markedly higher (P < 0.05) in goats fed GMOS than those fed only a control diet.
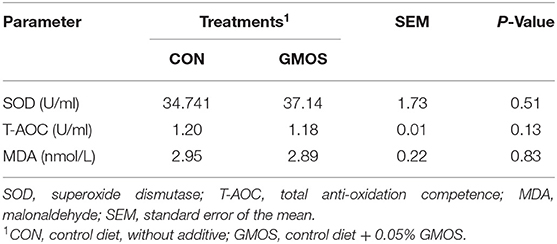
Table 5. Effects of dietary supplementation of GMOS on serum antioxidant indexes of goats.
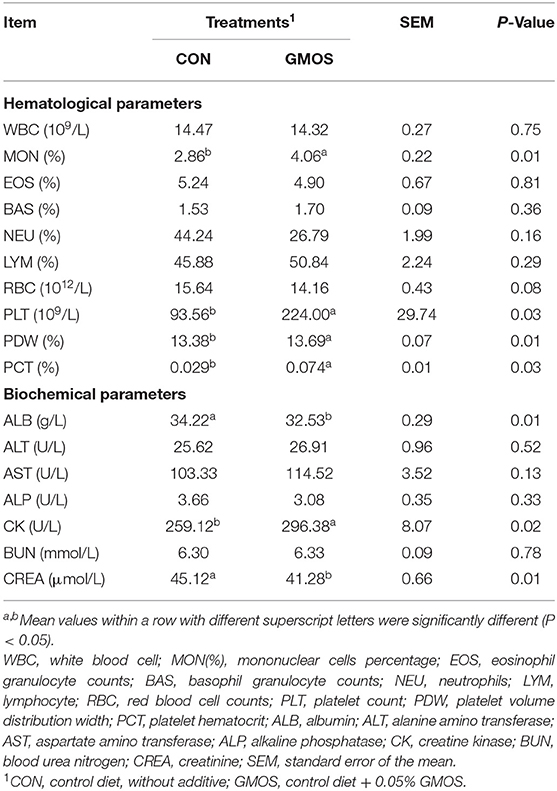
Table 6. Effect of dietary supplementation of GMOS on hematological and serum biochemical parameters in goats.
In the current study, the in vitro test results indicated that 0.05% GMOS outperformed traditional adsorbents like YCW and MMT in the absorption of aflatoxins. However, there was no difference in DMI, F/G, or ADG between the GMOS and the CON treatments. Previous studies have reported that dietary supplementation of MOS has been proven to enhance the performance of monogastric animals, including BW gain and feed conversion efficiencies (5, 24, 25). However, investigations on MOS in ruminants have yielded inconsistent results. Hill et al. (26) observed no differences in DMI and ADG when 3 g/day of a MOS was added to the diet of young Jersey calves. Similarly, Westland et al. (27) reported that dietary supplementation of 2 g of MOS to each calf daily did not improve the performance of calves. However, some other studies demonstrated enhanced ADG, AFDI, and feed efficiency in calves when a greater amount of MOS (4 g/calf/day) was supplemented in the diet (9) or milk replacer (10). Therefore, the growth performance of goats was not well improved as expected in the present study, likely attributed to the low dose of GMOS.
The in vivo test revealed that toxin concentrations in the GIT were much lower than those detected in the diet (refer to Suppplementary Table 1). These were likely due to two reasons: on the one hand, the toxins were adsorbed by mannan products (24); on the other hand, the complex GIT bacteria can degrade mycotoxins to different degrees (28). Although GMOS effectively adsorbed AFB1 in in vitro test, the concentrations of AFB1 did not differ between CON and GMOS treatments either in ruminal or cecal digesta, suggesting the importance of carrying out in vivo animal study. DON and AFB1 are metabolized into less toxic substances by rumen microorganisms (29), while ZEN is commonly transferred to metabolites like α-zearalenol (α-ZEL) and β-zearalenol (β-ZEL), which are significantly more hazardous than ZEN (30). Currently, it is challenging to separate ZEN and its metabolites using ELISA assay due to their similar structures. Moreover, rumen microbes can also selectively utilize oligosaccharides as fermentation substrates (31). Fewer GMOS reached the cecum, which is insufficient to change the mycotoxin concentration in the cecum. Therefore, the greater concentration of ZEN in cecal digesta than that in ruminal digesta was reasonable. Better analysis methods are needed to determine ZEN and its metabolites in the future. Our results suggested that GMOS could decrease the concentration of ZEN in the rumen; however, an insufficient amount of GMOS could adsorb ZEN in the cecum.
Then we investigated how GMOS affects the animal's fermentation parameters. GMOS did not change the TVAF concentration and the ruminal ammonia concentration. It significantly enhanced the molar proportion of propionate in the rumen but decreased that in the cecum. Wang et al. (31) have reported that mycotoxins negatively impact ruminal and intestinal fermentation. In contrast, MOS can attenuate the adverse effects of mycotoxins by promoting the growth of beneficial bacteria such as Lactobacillus in the rumen, and Lactobacillus is essential for the production of butyric and propionic acids (8). So we speculate that GMOS could also repair the detrimental effects of mycotoxins by promoting the growth of beneficial rumen bacteria. Furthermore, the small amount of GMOS reached the cecum, and the accumulation of toxins, especially ZEN, inhibited the growth of beneficial bacteria in the cecum.
Mycotoxins cause liver lesions and kidney damage with subsequent changes of some enzymatic parameters in goats (20). Previous studies have shown that MOS have antioxidative properties in sheep and chickens, including increasing the serum levels of SOD and T-AOC while lowing serum MDA levels (32, 33). In the current study, dietary supplementation GMOS had no effect on antioxidant indices in goats, including SOD, T-AOC and MDA. This finding suggested that GMOS supplementation would not cause any oxidative stress to small ruminants. Unaffected serum EOS and BAS levels used as stress indicators could also be robust support (34), suggesting that GMOS supplementation did not cause stress to goats. Similarly, dietary supplementation MOS did not affect the WBC count, EOS or BAS of dairy cows (35). Zheng et al. (36) reported that MOS did not significantly influence basal hematological parameters because their effects on the animal body are slight. In the current study, dietary supplementation of GMOS significantly increased routine blood indicators like MON, PCT, PLT and PDW. Previous study has shown a decrease in PLT and PDW (37), MON and PCT (8) in pigs exposed to mycotoxins such as ZEN and DON. Generally, an increase in MON indicates enhanced resistance to pathogens (38). In addition, platelet-related parameters such as PCT, PLT and PDW are associated with inflammatory reactions and immune responses (39). The rising level of PCT, PLT and PDW indicated the potential of GMOS to affect hematological parameters. We speculate that GMOS could prevent mycotoxin attacks on the goats' immune system to some extent.
In general, mycotoxins reduces the concentrations of CK and the decrease in serum levels of this substance may reflect renal cell damage with leakage of the contents into the blood (40). The current experiment showed an elevated CK level in the GMOS group, suggesting that GMOS may mitigate the damage caused by mycotoxins to the kidneys. Nargeskhani et al. (41) indicated that animals challenged by mycotoxins showed lower ALB level, the dropping level of serum ALB content indicated liver damage in animals. Although the current experiment showed decreased ALB content in the GMOS group, the ALB level was within the normal range (42). This is the first time we have found similar results. Furthermore, the serum levels of ALT, AST, and ALP were widely used to evaluate liver functions (41). The serum levels of ALT, AST, and ALP showed no differences between two treatments. Our results suggested that the addition of GMOS did not impair the serum biochemistry of goats. The similar result showed that the addition of mannan in diets had no deleterious effects on serum biochemistry and hematological characteristics of laying hen (43). These facts suggested that addition GMOS to goats' diet had no negative effects on their liver function. The presence of an increase in serum CREA level indicates the damage of kidney function (44). In our study, CREA level was significantly decreased (P < 0.05) in the GMOS group, suggesting that feeding a naturally contaminated diet to goats might impair kidney function and supplementation of GMOS could mitigate the nephrotoxicity. The decreased CREA level observed in our study was similar to the result reported by Raja et al. (45).
It was concluded that GMOS at 0.05% level was beneficial in promoting the proliferation of beneficial rumen bacteria. GMOS could also alleviate mycotoxin injury on the liver and kidney. Further research is needed to evaluate the degradation of mycotoxins by rumen microorganisms. And more in vivo experiments should be conducted to adjust the amount of GMOS in the diet for the adsorption effect of various mycotoxins. However, we can affirm the ability of GMOS as a green mycotoxin adsorbent. GMOS could serve as an economical solution to mycotoxicosis and make livestock feeding more cost-effective.
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding authors.
The animal study was reviewed and approved by Animal Care and Use Guidelines of the Animal Care and the Use Committee, Institute of Subtropical Agriculture, Chinese Academy of Sciences, Changsha, PR China (Approval Number: ISA-2019-0115).
YW, AR, CZ, and ZT designed the experiment. YW, AR, and XL conducted the experiment. YW, AR, XL, and GZ collected and analyzed data. YW prepared the tables, prepared the figures, and wrote the manuscript. CZ acquired funding. TR and CZ revised the manuscript. All authors contributed to the article and approved the submitted version.
This work was jointly supported by the National Natural Science Foundation of China (U20A2057 and 31730092) and National Basic Research and Development Program of China (2018YFD0501903).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We would like to thank Teketay Wassie (Institute of Subtropical Agriculture, Chinese Academy of Sciences, Changsha, China) for his linguistic assistance of this manuscript.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fvets.2022.852251/full#supplementary-material
1. Lee HJ, Ryu D. Worldwide occurrence of mycotoxins in cereals and cereal-derived food products: public health perspectives of their co-occurrence. J Agr Food Chem. (2017) 65:7034–51. doi: 10.1021/acs.jafc.6b04847
2. Horky P, Skalickova S, Baholet D, Skladanka J. Nanoparticles as a solution for eliminating the risk of mycotoxins. Nanomaterials. (2018) 8:727–47. doi: 10.3390/nano8090727
3. Mgbeahuruike AC, Ejioffor TE, Christian OC, Shoyinka VC, Karlsson M, et al. Detoxification of aflatoxin-contaminated poultry feeds by 3 adsorbents, bentonite, activated charcoal, and fuller's earth. J Appl Poult Res. (2018) 27:461–71. doi: 10.3382/japr/pfy054
4. Bhatti SA, Khan MZ, Ul Hassan Z, Saleemi MK, Saqib M, et al. Comparative efficacy of bentonite clay, activated charcoal and trichosporon mycotoxinivorans in regulating the feed-to-tissue transfer of mycotoxins. J Sci Food Agric. (2018) 98:884–90. doi: 10.1002/jsfa.8533
5. Tiwari UP, Fleming SA, Rasheed M, Jha R, Dilger RN. The role of oligosaccharides and polysaccharides of xylan and mannan in gut health of monogastric animals. J Nutr Sci. (2020) 9:E21. doi: 10.1017/jns.2020.14
6. Hang SQ, Mao SY, Huang RH, Su Y, Zhu WY. Effect of mannan-oligosaccharide on bacterial community of piglet after weaning. J Agric Biotechnol. (2006) 5:701–5.
7. Ministry of Agriculture of the People's Republic of China Announcement (No. 1126) [EB/OL]. Available online at: http://www.moa.gov.cn/nybgb/2014/dyq/201712/t20171219_6104350.htm (accessed February 01, 2014).
8. Reddy KE, Kim M, Kim KH, Baek JiSY, Chun YC, et al. Effect of commercially purified deoxynivalenol and zearalenone mycotoxins on microbial diversity of pig cecum contents. Anim Biosci. (2021) 34:243–55. doi: 10.5713/ajas.20.0137
9. Ghosh S, Mehla RK. Influence of dietary supplementation of prebiotics (mannan oligosaccharide) on the performance of crossbred calves. Trop Anim Health Prod. (2012) 44:617–22. doi: 10.1007/s11250-011-9944-8
10. Heinrichs AJ, Jones CM, Heinrichs BS. Effects of mannan oligosaccharide or antibiotics in neonatal diets on health and growth of dairy calves. J Dairy Sci. (2003) 86:4064–9. doi: 10.3168/jds.S0022-0302(03)74018-1
11. Joannis-Cassan C, Tozlovanu M, Hadjeba-Medjdoub K, Ballet N, Pfohl-Leszkowic A. Binding of Zearalenone, Aflatoxin B1, and Ochratoxin A by yeast-based products: a method for quantification of adsorption performance. J Food Protect. (2011) 74:1175. doi: 10.4315/0362-028X.JFP-11-023
12. Saleemi MK, Ashraf K, Gul ST, Naseem MN, Khan A. Toxicopathological effects of feeding aflatoxins B1 in broilers and its ameliosration with indigenous mycotoxin binder. Ecotox Environ Safe. (2019) 187:109712. doi: 10.1016/j.ecoenv.2019.109712
13. Swelum AA, Hashem NM, Abdelnour SA, Taha AE, Ei-hack MEA. Effects of phytogenic feed additives on the reproductive performance of animals. Saudi J Biol Sci. (2021) 28:5816–22. doi: 10.1016/j.sjbs.2021.06.045
14. Marroquin CA, Deng Y, Taylor JF, Hallmark CT, Johnson NM, et al. In vitro and in vivo characterization of mycotoxin-binding additives used for animal feeds in Mexico. Food Addit Contam A. (2009) 26:733–43. doi: 10.1080/02652030802641872
15. Aazami MH, Nasri MHF, Mojtahedi M, Battacone G. Preparation and characterization of yeast cell wall beta-glucan encapsulated humic acid nanoparticles as an enhanced aflatoxin B1 binder. Carbohyd Polym. (2018) 203:185–92. doi: 10.1016/j.carbpol.2018.08.047
16. Zhang H (2010). Animal nutrition parameter and feeding standard. Beijing: China Agriculture Press (in Chinese).
17. AOAC. Official Methods of Analysis. 17th ed. Arlington, VA: Association of Official Analytical Chemists (2012).
18. Van Soest PJ, Robertson JB, Lewis BA. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J Dairy Sci. (1991) 74:3583–97. doi: 10.3168/jds.S0022-0302(91)78551-2
19. Kekana TW, Marume U, Muya MC, Nherera-Chokuda FV. Periparturient antioxidant enzymes, haematological profile and milk production of dairy cows supplemented with moringa oleifera leaf meal. Anim Feed Sci Technol. (2020) 268:114606. doi: 10.1016/j.anifeedsci.2020.114606
20. Guo YP, Huo XT, Zhao LH, Ma QG, Zand JY, et al. Protective effects of Bacillus subtilis ANSB060, Bacillus subtilis ANSB01G, and Devosia sp. ANSB714-based mycotoxin biodegradation agent on mice fed with naturally moldy diets. Probiotics Antimicrob Proteins. (2020) 12:994–1001. doi: 10.1007/s12602-019-09606-w
21. Chen L, Ren A, Zhou CS, Tan ZL. Effects of Lactobacillus acid ophilus supplementation for improving in vitro rumen fermentation characteristics of cereal straws. Ital J Anim Sci. (2016) 16:52–60. doi: 10.1080/1828051X.2016.1262753
22. Bafti MS, Mozaffari AA. Normal hematological parameters in iranian cashmere (raini) goats. Comp Clin Pathol. (2012) 21:1653–5. doi: 10.1007/s00580-011-1343-z
24. Khatke PA, Singh R, Mandal AB, Tyagi PK. Evaluation of the ability of Saccharomyces cerevisiae and mannan oliosaccharides to ameliorate the adverse effects of aflatoxin B1 in broiler chickens. Indian J Poul Sci. (2012) 47:176–82.
25. Soumeh EA, Mohebodini H, Toghyani M, Shabani A, Ashayerizadeh A, et al. Synergistic effects of fermented soybean meal and mannan-oligosaccharide on growth performance, digestive functions, and hepatic gene expression in broiler chickens. Poul Sci. (2019) 98:6797–807. doi: 10.3382/ps/pez409
26. Hill SR, Hopkins BA, Davidson S, Bolt SM, Diaz DE, et al. The addition of cottonseed hulls to the starter and supplementation of live yeast or mannan oligosaccharide in the milk for young calves. J Dairy Sci. (2009) 92:790–8. doi: 10.3168/jds.2008-1320
27. Westland A, Martin R, White R, Martin JH. Mannan oligosaccharide prepartum supplementation: effects on dairy cow colostrum quality and quantity. Animal. (2017) 11:1779–82. doi: 10.1017/S1751731117000672
28. Morgavi DP, Boudra H, Jouany JP, Graviou D. Prevention of patulin toxicity on rumen microbial fermentation by SH-containing reducing agents. J Agric Food Chem. (2003) 51:6906–10. doi: 10.1021/jf034505v
29. Dänicke S, Brezina U. Kinetics and metabolism of the Fusarium toxin deoxynivalenol in farm animals: consequences for diagnosis of exposure and intoxication and carry over. Food Chem Toxicol. (2013) 60:58–75. doi: 10.1016/j.fct.2013.07.017
30. Dänicke S, Valenta H, Klobasa F, Döll S, Ganter M, et al. Effects of graded levels of Fusarium toxin contaminated wheat in diets for fattening pigs on growth performance, nutrient digestibility, deoxynivalenol balance and clinical serum characteristics. Arch Anim Nutr. (2004) 58:1–17. doi: 10.1080/0003942031000161045
31. Wang LJ, Zhang GN, Xu HJ, Xin HS, Zhang YG. Metagenomic analyses of microbial and carbohydrate-active enzymes in the rumen of Holstein cows fed different forage-to-concentrate ratios. Front Microbiol. (2019) 10:649. doi: 10.3389/fmicb.2019.00649
32. Zheng C, Li F, Hao Z, Liu T. Effects of adding mannan oligosaccharides on digestibility and metabolism of nutrients, ruminal fermentation parameters, immunity, and antioxidant capacity of sheep. J Anim Sci. (2018) 96:284–92. doi: 10.1093/jas/skx040
33. Zhou M, Tao Y, Lai C, Huang C, Zhou Y, et al. Effects of mannanoligosaccharide supplementation on the growth performance, immunity, and oxidative status of partridge shank chickens. Animals. (2019) 9:817. doi: 10.3390/ani9100817
34. Akrami R, Mansour MR, Ghobadi S, Ahmadifar E, Khoshroudi MS, et al. Effect of prebiotic mannan oligosaccharide on hematological and blood serum biochemical parameters of cultured juvenile great sturgeon. J Appl Ichthyol. (2013) 29:1214–8. doi: 10.1111/jai.12245
35. Franklin ST, Newman MC, Newman KE, Meek KI. Immune parameters of dry cows fed mannan oligosaccharide and subsequent transfer of immunity to calves. J Dairy Sci. (2005) 88:766–75. doi: 10.3168/jds.S0022-0302(05)72740-5
36. Zheng C, Zhou JW, Zeng YQ, Liu T. Effects of mannan oligosaccharides on growth performance, nutrient digestibility, ruminal fermentation and hematological parameters in sheep. PeerJ. (2021) 9:e11631. doi: 10.7717/peerj.11631
37. Reddy KE, Song J, Hyun JL, Minseok K, Dong WK, Bumseok K, et al. Effects of high levels of deoxynivalenol and zearalenone on growth performance, and hematological and immunological parameters in pigs. Toxins. (2018) 10:114. doi: 10.3390/toxins10030114
38. Chen SK, Wang XF, Guo LiS, Zhou F, Luo HS, et al. Diagnostic method of pig blood routine index for pig health. J Anim Sci. (2018) 54:117–20.
39. Xiong L, Zhang WH, Kong XF, Wang XD, You JM. Effects of intrauterine growth retardation on blood routine indices of growing pigs. J Anim Nutr. (2020) 32:417–22.
40. Andretta I. Meta-analysis of the relationship of mycotoxins with biochemical and hematological parameters in broilers. Poult Sci. (2012). doi: 10.3382/ps.2011-01813
41. Nargeskhani A, Dabiri N, Esmaeilkhanian S, Alipour MM, Bojarpour M. Effects of mannanoligosaccharide–β glucan or antibiotics on health and performance of dairy calves. Anim Nutr Feed Tech. (2010) 10:29–36.
42. Tian XG, Zhu HG, Zhu X, Pan SB, Wu F, et al. Determination and analysis of partial blood physiological indexes of guizhou black female goat. Southwest China J Agri Sci. (2011) 24:366–8.
43. Maziar MA, Seyed AH, Houshang L, Farid S. Effect of probiotics, yeast, vitamin e and vitamin c supplements on performance and immune response of laying hen during high environmental temperature. Int J Poult Sci. (2007) 6:895–900. doi: 10.3923/ijps.2007.895.900
44. Ogunade IM, Martinez TC, Queiroz OCM, Jiang Y, Drouin P, et al. Silage review: mycotoxins in silage: occurrence, effects, prevention, and mitigation. J Dairy Sci. (2018) 101:4034–59. doi: 10.3168/jds.2017-13788
Keywords: galactomannan oligosaccharides, mycotoxins, biochemical parameters, goat, detoxification
Citation: Wu Y, Ren A, Lv X, Ran T, Zhang G, Zhou C and Tan Z (2022) Effects of Galactomannan Oligosaccharides on Growth Performance, Mycotoxin Detoxification, Serum Biochemistry, and Hematology of Goats Fed Mycotoxins-Contaminated Diets. Front. Vet. Sci. 9:852251. doi: 10.3389/fvets.2022.852251
Received: 11 January 2022; Accepted: 19 May 2022;
Published: 24 June 2022.
Edited by:
Elsa Lamy, University of Évora, PortugalReviewed by:
Yangchun Cao, Northwest A&F University, ChinaCopyright © 2022 Wu, Ren, Lv, Ran, Zhang, Zhou and Tan. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Tao Ran, cmFudEBsenUuZWR1LmNu; Chuanshe Zhou, emNzQGlzYS5hYy5jbg==
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.