
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Vet. Sci. , 13 January 2023
Sec. Veterinary clinical, anatomical, and comparative pathology
Volume 9 - 2022 | https://doi.org/10.3389/fvets.2022.1080994
This article is part of the Research Topic Biology and Pathology of Tumor Viruses in Animals, Volume II View all 6 articles
 José Gomes Pereira*
José Gomes Pereira* Cândida de Assunção SilvaLucas Diniz SilvaCristian Alex Aquino Lima
Cândida de Assunção SilvaLucas Diniz SilvaCristian Alex Aquino Lima Carla Janaina Rebouças Marques do RosárioEllainy Maria Conceição SilvaMaria do Socorro Costa OliveiraLarissa Sarmento dos Santos RibeiroHamilton Pereira Santos
Carla Janaina Rebouças Marques do RosárioEllainy Maria Conceição SilvaMaria do Socorro Costa OliveiraLarissa Sarmento dos Santos RibeiroHamilton Pereira Santos Ana Lucia Abreu-SilvaFerdinan Almeida Melo
Ana Lucia Abreu-SilvaFerdinan Almeida MeloEnzootic bovine leukosis (EBL) is a chronic viral disease of wide distribution in cattle herds and may take several years for the first manifestation of clinical signs. Most animals do not present clinical signs. However, the economic losses are underestimated due to this disease. Thus, this work aimed to detect and characterize BLV in dairy cattle in the Maranhão state, northeastern Brazil. Blood samples were collected from 176 animals from 8 municipalities in the southeastern state of Maranhão. Bovine blood samples were subjected to DNA extraction and molecular diagnosis using nested PCR assays for BLV, targeting gp51 gene. Positive samples were then sequenced and then subjected to phylogenetic inferences. BLV DNA was detected in 16 cattle (16/176, 9.09%) in 4 municipalities. Phylogenetic analyzes showed that the sequence obtained clustered in a clade containing BLV sequences classified as genotype 6, with a high degree of support. Our data shows BLV occurrence in the Northeast of Brazil and the identification of genotype 6 in this region. These findings contribute to the molecular epidemiology of this agent in Brazil.
Enzootic Bovine Leukosis (EBL) is a chronic viral disease of wide distribution in bovine herds, caused by the bovine leukosis virus (BLV) belonging to the Retroviridae family and Orthoretrovirinae subfamily (1, 2).
About 30–70% of infected animals may develop persistent lymphocytosis (PL), which observes an increase in circulating B lymphocytes. In addition, 0.1–10% of infected animals may develop lymphosarcomas, which has a fatal course (3, 4). Furthermore, once infected, animals remain carriers and are sources of lifelong shedding of the virus (5).
According to normative instruction 50 of September 24, 2013 of MAPA (Ministry of Agriculture, Livestock and Supply/Ministry of Agriculture, Livestock and Supply) (6), EBL is one of the diseases that must be notified monthly. However, the disease remains unknown to most farmers, and a strict sanitary policy needs to be. These factors contribute to the wide dissemination in the Brazilian herd.
Clinical signs include digestive disturbances, inappetence, and weight loss. In addition, lymphosarcoma is reported in animals aged 4–8 years. It is most frequent in peripheral and internal lymph nodes but, it was also observed abomasum, heart, uterus, retrobulbar space, and epidural region of the central nervous system (3, 7–9).
Blood containing contaminated lymphocytes transferred from animal to animal by iatrogenic means is considered the main route of disease transmission; however, it is reported that vertical transmission of BVL can occur by the uterine route or even orally, through the ingestion of colostrum and milk-containing lymphocytes contaminated with BLV (10, 11).
The BLV genome is constituted by identical copies of single-stranded RNA, which is transcribed into a double-stranded DNA called a provirus. It encodes capsid structural genes (gag), the RNA-dependent DNA polymerase (the reverse transcriptase) (pol) and viral envelope proteins (env) (12). For purposes of phylogenetic analysis, env genes are widely used. Furthermore, this gene encodes two surface glycoproteins, GP51 and GP30, which are essential for infectivity and immune evasion. In addition, these genes made it possible to perform BLV genotyping and characterize at most minuscule 10 virus genotypes worldwide (13–16).
Many serological studies carried out in Brazil demonstrate the exposure of animals to BLV in all regions of the country (17–23). Studies carried out mainly in the country's southeastern region evidenced the agent's circulation and the characterization of genotypes 1, 2, 5, and 6 (24–27).
In the state of Maranhão, only one serological survey showed exposure of dairy cattle to the virus, with a prevalence of 53.8% (28). Thus, this study aimed to investigate, using a molecular diagnostic technique the occurrence of BLV in cattle herds and to characterize the phylogenetic relationships of the sequence described for the first time in Maranhão, a state located in the Northeast of the country.
The research project was approved by the Committee on Ethics and Animal Experimentation (CEEA) of the Veterinary Medicine Course at UEMA, Protocol No. 044/2017 CEEA/UEMA).
The study was conducted in semi-intensive dairy cattle farms located in the rural zone of 8 municipalities in the southeastern region of the state of Maranhão: Imperatriz, Davinópolis, Porto Franco, Governador Edson Lobão, Amarante, Senador lá Roque, João Lisboa e São João do Paraíso (Figure 1).
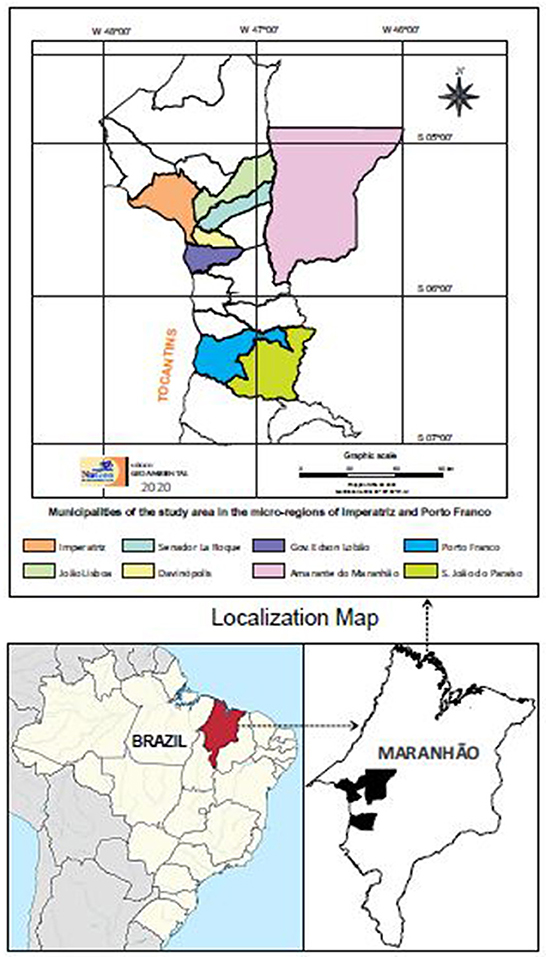
Figure 1. Map showing the location of municipalities where the sample collection was carried out. Locations are indicated by color.
The climate of this area is classified as hot and humid, with two well-defined seasons, rainy and dry. The average annual temperature in the region is about 26°C, with an annual amplitude of < 5°C, which characterizes the region's thermal regime with slight seasonal variation. This region is part of the cerrado biome (29).
Blood collection was performed between August and September of 2018, during the dry season, from 176 randomly selected cattle. The animals' age ranged from 1 to 4 years, and dairy cattle, including females and calves, were all raised in a semi-intensive system.
Blood was obtained by venipuncture using vacutainer tubes with an anticoagulant. Then the samples were centrifuged at 1610 xG for 10 min to separate the buffy coat, which was removed and transferred to microtubes and stored at −20°C for other molecular biology techniques. No clinical sign was observed during the sample collection.
For extraction of genomic DNA from the buffy coat, the commercial kit InstaGene™ Matrix (Bio-Rad Laboratories, Inc., USA), was used, following the manufacturer's instructions. To assess the quality, and the presence of extraction inhibitors and eliminate false negative results, all extracted DNA was subjected to PCR for the conserved cyt b of the mitochondrial DNA of vertebrates (mtDNA), according to the protocol described by Steuber et al. (30), which amplifies a 359 base pairs (bp) product.
The amplification reaction was performed using a final volume of 25 μL, using a mastermix solution containing 5.1 μL of Green GoTaq® Flexi Buffer 5x (Promega, USA), 1.9 μL of each primer (5 pmol/μL–Sigma), 0.5 μL of dNTPmix (10 mM–Promega, USA) 1.5 μL of MgCl2 (10 mM–Promega, USA), 0.26 μL of Go TaqFlexi DNA Polymerase (500 U-Promega, USA), 2 μL of DNA template and DNase-free ultrapure water and RNase (Invitrogen-Life Technologies®, USA). The PCR cycles were programmed as follows: initial denaturation at 95°C for 5 min, 35 cycles composed of denaturation at 95°C for 15 s, annealing at 50°C for 30 s and extension at 72°C for 30 s, and final extension at 72°C for 5 min.
The samples positive in the cyt b gene were then subjected to n-PCR for detection of BLV, using primers described by Asfaw et al. (31), which amplifies a region of the env gene (gp51), of 444 bp. The first reaction had a final volume of 25 ul, with 3.0 μl of 10 mM of each primer, 0.4 mM DNTPs, 1X PCR buffer (Thermo Scientific®), 3 mM MgCl2 and 1U of Taq DNA polymerase. In the second PCR reaction, 5 μL DNA of the product of the first amplification was used as a template, with the same concentrations of the other reagents and the primers env 5099 (5' CCCACAAGGGCGGCGCCGGTTT-3') and env 5521 (5' GCGAGGCCGGGTCCAGAGCTGG-3'), in a final volume of 30 μL.
Both reactions (first and second) were carried out according to the following program: initial denaturation at 94 °C for 5 min, followed by 40 cycles of 94°C for 30 s, 60°C for 30 s and 72°C for 1 min, and final extension of at 72°C for 5 min. A BVL positive sample was used as a positive control and ultrapure water was used as a negative control in all reactions.
The amplified products were subjected to horizontal electrophoresis on a 1.5% agarose gel stained with ethidium bromide (0.5 μL/ml) in TAE buffer pH 8.3 (Tris Acetate EDTA). Electrophoresis was performed at 100 V and a current of 400 mA for 40 min. The results were visualized and analyzed by an ultraviolet light transilluminator coupled to a computer program for image analysis (L-PIX gels Loccus®).
Amplified products positive for all genes tested were purified using the commercial Wizard® SV Gel and PCR Clean-Up System (Promega®) kit, according to the manufacturers' recommendations. The quantification of purified DNA was performed using a NanoDrop 2000 spectrophotometer (Thermo Scientific, San Jose CA, USA). Sanger sequencing was performed according to the dideoxynucleotide chain termination method (32) on the ABI PRISM 3,500 Genetic Analyzer (Applied Biosystem, Foster City, CA, USA). The sequencing was performed at the ACTGene Molecular Analysis, Rio Grande do Sul, Brazil.
The nucleotide sequence obtained was inserted into the FinchTV 1.4 software (Geospiza, Inc., Seattle, WA, USA) for evaluating the quality of electropherograms. In the Basic Local Alignment Search Tool program (33) the inserted sequence was compared with other sequences deposited in GenBank.
For phylogenetic analyses, gp51 sequences were aligned with other homologous sequences of the same sequenced gene, taken from the Genbank database, using the Clustal/W tool via Bioedit 7.2 (34). The best-fit model of sequence evolution was selected based on the Corrected Akaike Information Criterion (cAIC), using MEGAX (35). The phylogenetic trees were reconstructed based on the neighbor-joining (NJ) and maximum likelihood (ML) methods, with 1,000 replicates for bootstrap analysis. The sequences generated in this search were submitted to GenBank and assigned accession number MT135126.
All samples amplified the endogenous cyt b gene, which was used for extraction evaluation. Thus, they were tested in nested PCR based on the env gene and in total 9.09% (16/176) of animals were positive. In total, 8 municipalities in the southeastern region of the state of Maranhão were included in this research, and of these, 4 had nPCR positive animals (Table 1).

Table 1. Percentage of animals positive for BLV by cities in the Southwest region of Maranhão.
A sequence was obtained and used for phylogenetic analysis from positive samples. The genetic identity ranged from 96.85 to 100% with different BLV isolates from various locations in Brazil (JN254639, DQ059415, AY185360), South American countries such as Bolivia (LC075576), Paraguay (LC080656), and others countries like India (MH341525), Pakistan (MW926787), Russia (KC886631), Myanmar (LC466592), Thailand (KU233536).
Phylogenetic analysis was performed using ML and NJ methodologies, and both analyzes showed similar topology, so only the NJ is shown. For phylogenetic reconstruction, sequences of isolates from genotypes 1–12, deposited in the Genbank were selected. The analyzed sequence was grouped in a clade formed by BLV sequences belonging to genotype 6, with strong clade support. In this group, sequences from different countries were observed, such as Brazil, in São Paulo state, and other countries such Peru and Argentina all isolated from bovine blood (Figure 2).
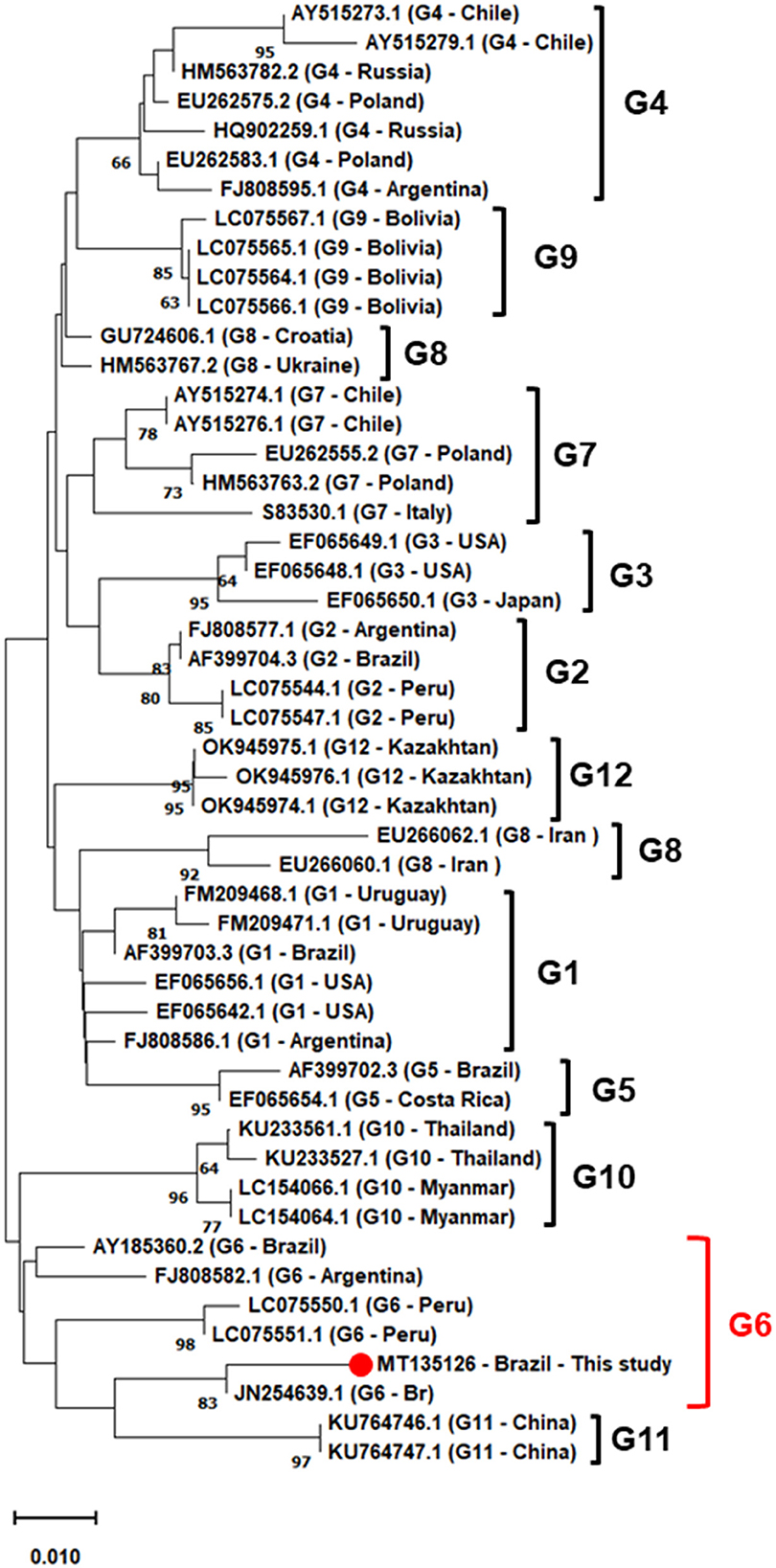
Figure 2. Phylogenetic inferences using the neighbor-joining (NJ) method based on the Kimura 2-parameter model, with gamma distribution (k2+G), the numbers at the branches show bootstrap support (1.000 replicates). The analysis involved 50 nucleotide env gene sequences from Brazil and the world, based on 464 bp fragment. The sequence described in this study is indicated by filled red circle. Evolutionary analyses were conducted in MEGAX.
This study presents data on the occurrence and phylogenetic characterization of BLV in cattle herds in the southeastern state of Maranhão. This area has a critical livestock activity because it is the second largest cattle herd in the northeastern region of Brazil since it is a significant exporter of live cattle, ~1,000 heads/yearly (36).
According to the Ministry of Agriculture, Livestock, and Supply (Ministério da Agricultura, Pecuária e Abastecimento) (MAPA), enzootic bovine leukosis is mandatory monthly notification of confirmed cases (category 4). However, no specific public policies exist to control or eradicate the disease (6). Serological rates have varied between 11.78 and 45% in states in the northeast region (Pernambuco and Sergipe), in the midwest region (Mato Grosso do Sul) and in the southern region (Santa Catarina) (19–21, 37). In the state of Maranhão, there is only one survey showing the exposure of animals to BLV, and it showed high rates of 53.8% of antibodies (28).
On the other hand, surveys using molecular diagnostic techniques targeting the identification of viral DNA are even more limited. In southeastern and southern Brazil, respectively, D'Angelino et al. (26) reported 4.8% of CNS tissues from BLV-positive cattle in the state of São Paulo, and Rodakiewicz et al. (37), found a high rate of 80.5% (62/77) of positive animals in the State of Santa Catarina. Although serological studies are observed in the northeastern region, direct detection of virus DNA by molecular techniques has not been observed. This is the first molecular study, to our knowledge, conducted in the state of Maranhão.
Surveillance activities for the occurrence of BLV are very important, considering two aspects: the damage caused to infected animals and the zoonotic potential of BLV. The impacts caused by leukosis are expressive in the affected herds, as these animals maintain low production rates. Direct losses have been estimated due to reduced milk production, reproduction and premature slaughter of animals, and restrictions on international trade from countries where the virus is circulating (38–41). Because of this, it is crucial to have a current plan for the occurrence of BLV in Brazil, as it will allow a better understanding of the virus and the possible economic impacts in different regions.
It is important to mention that previous studies carried out in recent years have indicated the zoonotic potential of BLV. Although it has not yet been clearly determined how humans can be infected by the virus or its clinical implications, it has been associated with breast cancer in women (42, 43). Olaya-Galan et al. (44) reported the occurrence of BLV DNA in milk and meat products, demonstrating that human infection can occur orally. This fact reinforces the need to create BLV eradication programs in Brazil.
Regarding phylogenetic analyses, it was shown, using the NJ algorithm, that the sequence isolated from bovine in this research is clustered in a strongly supported clade, with isolates from bovines from São Paulo (JN254639) and Mato Grosso do Sul (AY185360) (100% bootstrap), in addition to sequences from Peru (LC05551 and LC05550) and Argentina (FJ808582). These results are similar to those already indicated in previous studies conducted in Brazil and Latin America, where genotype 6 seems to be very frequent. In addition to the South American continent, genotype 6 has been reported in Asian countries such as China (45, 46), Thailand (47), Myanmar (48), Vietnam (49), demonstrating the distribution of this genotype around the world. Sultanov et al. (50) point out that the introduction of infected cattle into herds is the main form of transmission of BLV, considering that the genotype 6 characterized in this locality is phylogenetically related to that found in the central-west and southeastern regions of Brazil, geographically distant, but that carry out the cattle trade with the state of Maranhão (36). This may have been an important route for the dissemination of genotype 6 in the country and also to other locations around the world. Brazil is one of the largest exporters of beef and lives cattle in the world; countries such as China, United States of America, Egypt, and Chile are the largest importers of Brazilian meat (51). Therefore, concentrated efforts to develop public policies to control enzootic bovine leukosis are essential to contain the spread of specific BLV genotypes.
BLV circulation was evidenced in dairy cattle in the state of Maranhão, Northeastern Brazil. For the first time, phylogenetic inferences showed genotype 6 in this region related to sequences previously identified in central-western and southeastern Brazil.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found at: https://www.ncbi.nlm.nih.gov/genbank/, MT13512.
The research project was approved by the Committee on Ethics and Animal Experimentation (CEEA) of the Veterinary Medicine Course at UEMA, Protocol No. 044/2017 CEEA/UEMA). Written informed consent was obtained from the owners for the participation of their animals in this study.
JP, FM, AA-S, MO, CR, LR, and HS: project idealization, experimental data interpretation, and writing. CS and LS: sample collection, hematological analyzes, data interpretation, and writing. CL and ES: helped molecular and experimental conduction. All authors read and approved the final manuscript.
This research was funded by FAPEMA—Fundação de Amparo à Pesquisa e ao Desenvolvimento Científico e Tecnológico do Maranhão FAPEMA; CAPES-Coordenação de Aperfeiçoamento de Pessoal de Nível Superior—Finance Code 001, and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) grant number 313348/2021-9.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Radostits OM, Gay CC, Blood DC, Hinchcliff KW. Doenças causadas por virus e Chlamydia. In:Clínica Veterinária: Um tratado de doenças dos bovinos ovinos suínoscaprinos e equinos. 9, ed. Rio de Janeiro. Guanabara Koogan (2002). p. 940–51.
2. Ravazzolo AP, Costa UM. Retroviridae. In:Eduardo Furtado Flores (Org) Virologia Veterinária 1ª, ed. Santa Maria (2007). p. 809–37.
4. Barez PY, De Brogniez A, Carpentier A, Gazon H, Gillet N, Gutiérrez G, et al. Recent advances in BLV research. Viruses. (2015) 7:6080–8. doi: 10.3390/v7112929
5. Shirley JE, Smith JF, Stokka GL, Scoby R, Van Anne T. The compendium on continuing education for the practicing veterinarian. Princeton. (1997) 19:651–4.
6. BRASIL. Ministério da Agricultura, Pecuária e Abastecimento (MAPA). Instrução Normativa MAPA n° 50, de 23 de setembro de. Lista das doenças animais de notificação obrigatória ao Serviço Veterinário Oficial brasileiro. (2013). Available online at: https://www.gov.br/agricultura/pt-br/ (accessed September 10, 2022).
7. Mekata H, Sekiguchi S, Konnai S, Kirino Y, Honkawa K, Nonaka N, et al. Evaluation of the natural perinatal transmission of bovine leukaemia virus. Vet Rec. (2015) 176:254–254. doi: 10.1136/vr.102464
8. Panei CJ, Serena MS, Metz GE, Bravi ME, González ET, Echeverría MG. Analysis of the pX region of bovine leukemia virus in different clinical stages of Enzootic Bovine Leukemia in Argentine Holstein cattle. Virus Res. (2013) 171:97–102. doi: 10.1016/j.virusres.2012.08.001
9. Mekata H, Sekiguchi S, Konnai S, Kirino Y, Horii Y, Norimine J. Horizontal transmission and phylogenetic analysis of bovine leukemia virus in two districts of Miyazaki, Japan. J Vet Med Sci. (2015) 77:1115–20. doi: 10.1292/jvms.14-0624
10. Juliarena MA, Barrios CN, Ceriani MC, Esteban EN. Hot topic: Bovine leukemia virus (BLV)-infected cows with low proviral load are not a source of infection for BLV-free cattle. J Dairy Sci. (2016) 99:4586–9. doi: 10.3168/jds.2015-10480
11. Takeshima SN, Ohno A, Aida Y. Bovine leukemia virus proviral load is more strongly associated with bovine major histocompatibility complex class II DRB3 polymorphism than with DQA1 polymorphism in Holstein cow in Japan. Retrovirology. (2019) 16:1–6. doi: 10.1186/s12977-019-0476-z
12. Gillet N, Kettmann R, Willems L. Homologies entre les deux rétrovirus BLV et HTLV-1 et développement d'une nouvelle approche thérapeutique basée sur la levée de la latence virale. BASE. (2007) 11:151–7.
13. Zhao X, Buehring GC. Natural genetic variations in bovine leukemia virus envelope gene: possible effects of selection and escape. Virology. (2007) 366:150–65. doi: 10.1016/j.virol.2007.03.058
14. Polat M, Takeshima SN, Hosomichi K, Kim J, Miyasaka T, Yamada K, et al. A new genotype of bovine leukemia virus in South America identified by NGS-based whole genome sequencing and molecular evolutionary genetic analysis. Retrovirology. (2016) 13:1–23. doi: 10.1186/s12977-016-0239-z
15. Polat M, Moe HH, Shimogiri T, Moe KK, Takeshima SN, Aida Y. The molecular epidemiological study of bovine leukemia virus infection in Myanmar cattle. Arch Virol. (2017) 162:425–37. doi: 10.1007/s00705-016-3118-y
16. Polat M, Takeshima SN, Aida Y. Epidemiology and genetic diversity of bovine leukemia virus. Virol J. (2017) 14:1–16. doi: 10.1186/s12985-017-0876-4
17. Fernandes CHC, de Melo LEH, Tenório TDS, Mendes EI, Fernandes ADC, Ramalho TRR, et al. Soroprevalência e fatores de risco da infecção pelo vírus da leucose dos bovinos em rebanhos leiteiros da região norte do estado do Tocantins, Brasil. Arq Inst Biol. (2021) 76:327–334. doi: 10.1590/1808-1657v76p3272009
18. de Barros Filho IR, Guimarães AK, Sponchiado D, Krüger ER, Wammes EV, Ollhoff RD, et al. Soroprevalência de anticorpos para o vírus da leucose enzoótica em bovinos criados na região metropolitana de Curitiba, Paraná. Arq Inst Biol. (2020) 77:511–5. doi: 10.1590/1808-1657v77p5112010
19. Heidman M, Fischer VL, de Castro BG. Enzootic bovine leukosis antibodies in cattle from mid-northern Mato Grosso State, Brazil. Arq Inst Biol. (2015). doi: 10.1590/1808-1657000792013
20. Batista JM, Batista DM, Costa JN, Costa Neto AO, Lima CV, Souza TS, et al. Inquérito soroepidemiológico da leucose enzoótica bovina em rebanhos bovinos do estado de Sergipe. Cienc Anim Bras. (2018). doi: 10.1590/1809-6891v19e-23196
21. Baptista L, Fernandes A, Silva T, Ramos T, Melo L. Performance assessment of imported ELISA in the serodiagnosis of the enzootic bovine leukosis in herds of Pernambuco state, Brazil. Arq Inst Biol. (2019). doi: 10.1590/1808-1657000662018
22. Mendes EI, Melo LEH, Tenório TGS, Sá LM, Souto RJC, Fernandes ACC, et al. Intercorrência entre leucose enzoótica e tuberculose em bovinos leiteiros do estado de Pernambuco. Arq Inst Biol. (2020) 78:1–8. doi: 10.1590/1808-1657v78p0012011
23. Ramalho GC, Silva ML, Falcão BMR, Limeira CH, Nogueira DB, Dos Santos AM, et al. High herd-level seroprevalence and associated factors for bovine leukemia virus in the semi-arid Paraíba state, Northeast Region of Brazil. Prev Vet Med. (2021) 190:105324. doi: 10.1016/j.prevetmed.2021.105324
24. Camargos MF, Pereda A, Stancek D, Rocha MA, Reis JKPD, Greiser-Wilke I, et al. Molecular characterization of the env gene from Brazilian field isolates of Bovine leukemia virus. Virus Genes. (2007) 34:343–50. doi: 10.1007/s11262-006-0011-x
25. Dias NL, Fonseca AA, Rodrigues DS, Camargos MF. PCR em tempo real para diagnóstico da leucose enzoótica bovina. Cienc Rural. (2012) 42:1434–9. doi: 10.1590/S0103-84782012005000053
26. D'Angelino RHR, Pituco EM, Villalobos EMC, Harakava R, Gregori F, Del Fava C. Detection of bovine leukemia virus in brains of cattle with a neurological syndrome: pathological and molecular studies. Biomed Res Int. (2013) 2013:425646. doi: 10.1155/2013/425646
27. Gregory L, Gaeta NC, Araújo J, Thomazelli LM, Harakawa R, Ikuno AA, et al. Bovine leukaemia virus genotypes 5 and 6 are circulating in cattle from the state of São Paulo, Brazil. J Med Microbiol. (2017) 66:1790–7. doi: 10.1099/jmm.0.000639
28. Santos HP, Pereira HDM, Nascimento SA, Coutinho LCA, Teixeira WC, Arruda RCN, et al. Frequência de anticorpos e fatores de risco associados á Leucose Enzoótica Bovina em rebanhos da bacia leiteira do estado do Maranhão. Arq Inst Biol. (2020) 78:351–8. doi: 10.1590/1808-1657v78p3512011
29. Spinelli-Araujo L, da Silva GBS, Torresan FE, Victoria DDC, Vicente LE, et al. Conservação da biodiversidade do Estado do Maranhão: cenário Atual em dados geoespaciais (2016).
30. Steuber S, Abdel-Rady A, Clausen PH. PCR-RFLP analysis: a promising technique for host species identification of blood meals from tsetse flies (Diptera: Glossinidae). Parasitol Res. (2005) 97:247–54. doi: 10.1007/s00436-005-1410-y
31. Asfaw Y, Tsuduku S, Konishi M, Murakami K, Tsuboi T, Wu D, et al. Distribution and superinfection of bovine leukemia virus genotypes in Japan. Arch. Virol. (2005) 150:493–505. doi: 10.1007/s00705-004-0433-5
32. Sanger F, Nicklen S, Coulson AR. DNA sequencing with chain-terminating inhibitors. Proc Natl Acad Sci USA. (1977) 74:5463–7. doi: 10.1073/pnas.74.12.5463
33. Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignment search tool. J Mol Biol. (1990) 215:403–10. doi: 10.1016/S0022-2836(05)80360-2
34. Hall TA. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. In: Nucleic Acids Symposium Series. London: Information Retrieval Ltd (1999). p. c1979–2000.
35. Kumar S, Stecher G, Li M, Knyaz C, Tamura K. MEGA X: molecular evolutionary genetics analysis across computing platforms. Mol Biol Evol. (2018) 35:1547–9. doi: 10.1093/molbev/msy096
36. Fundação de Desenvolvimento da Pecuária do Estado do Maranhão (FUNDEPEC-MA). Bovinocultura de corte. (2018). Available online at: https://fundepecma.org.br/bovinocultura-de-corte/ (accessed September 10, 2022).
37. Rodakiewicz SM, Fernandez ML, Munhoz ML, Yamakawa FHS, Urio M, Forell F, et al. Heterogeneity determination of bovine leukemia virus genome in Santa Catarina state, Brazil. Arq Inst Biol. (2018). doi: 10.1590/1808-1657000742016
38. Erskine RJ, Bartlett PC, Byrem TM, Render CL, Febvay C, Houseman JT. Association between bovine leukemia virus, production, and population age in Michigan dairy herds. J Dairy Sci. (2012) 95:727–34. doi: 10.3168/jds.2011-4760
39. Norby B, Bartlett PC, Byrem TM, Erskine RJ. Effect of infection with bovine leukemia virus on milk production in Michigan dairy cows. J Dairy Sci. (2016) 99:2043–52. doi: 10.3168/jds.2015-10089
40. Nakada S, Fujimoto Y, Kohara J, Adachi Y, Makita K. Estimation of economic loss by carcass weight reduction of Japanese dairy cows due to infection with bovine leukemia virus. Prev Vet Med. (2022) 198:105528. doi: 10.1016/j.prevetmed.2021.105528
41. World Organisation for Animal Health (WOAH). Enzootic bovine leukosis. Manual of Diagnostic Tests and Vaccines for Terrestrial Animals. (2018). Available online at: https://www.woah.org/en/what-we-do/standards/codes-and-manuals/terrestrial-manual-online-access (accessed October 10, 2022).
42. Buehring GC, Shen HM, Jensen HM, Choi KY, Sun D, Nuovo G. Bovine leukemia virus DNA in human breast tissue. Emerg Infect Dis. (2014) 20:772. doi: 10.3201/eid2005.131298
43. Olaya-Galán NN, Salas-Cárdenas SP, Rodriguez-Sarmiento JL, Ibáñez-Pinilla M, Monroy R, Corredor-Figueroa AP, et al. Risk factor for breast cancer development under exposure to bovine leukemia virus in Colombian women: a case-control study. PLoS ONE. (2021) 16:e0257492. doi: 10.1371/journal.pone.0257492
44. Olaya-Galán NN, Corredor-Figueroa AP, Guzmán-Garzón TC, Ríos-Hernandez KS, Salas-Cárdenas SP, Patarroyo MA, et al. Bovine leukaemia virus DNA in fresh milk and raw beef for human consumption. Epidemiol Infect. (2017) 145:3125–30. doi: 10.1017/S0950268817002229
45. Gautam S, Mishra N, Kalaiyarasu S, Jhade SK, Sood R. Molecular characterization of bovine leukaemia virus (BLV) strains reveals existence of genotype 6 in cattle in India with evidence of a new subgenotype. Transbound Emerg Dis. (2018) 65:1968–78. doi: 10.1111/tbed.12979
46. Yang Y, Chu S, Shang S, Yang Z, Wang C. Genotyping and single nucleotide polymorphism analysis of bovine leukemia virus in Chinese dairy cattle. J Dairy Sci. (2019) 102:3469–73. doi: 10.3168/jds.2018-15481
47. Lee E, Kim EJ, Ratthanophart J, Vitoonpong R, Kim BH, Cho IS, et al. Molecular epidemiological and serological studies of bovine leukemia virus (BLV) infection in Thailand cattle. Infect Genet Evol. (2016) 41:245–54. doi: 10.1016/j.meegid.2016.04.010
48. Moe KK, Polat M, Borjigin L, Matsuura R, Hein ST, Moe HH, et al. New evidence of bovine leukemia virus circulating in Myanmar cattle through epidemiological and molecular characterization. PLoS ONE. (2020) 15:e0229126. doi: 10.1371/journal.pone.0229126
49. Le DT, Yamashita-Kawanishi N, Okamoto M, Nguyen SV, Nguyen NH, Sugiura K, et al. Detection and genotyping of bovine leukemia virus (BLV) in Vietnamese cattle. J Vet Med Sci. (2020). doi: 10.1292/jvms.20-0094
50. Sultanov A, Rola-Łuszczak M, Mamanova S, Ryło A, Osiński Z, Saduakassova MA, et al. Molecular characterization of bovine leukemia virus with the evidence of a new genotype circulating in cattle from Kazakhstan. Pathogens. (2022) 11:180. doi: 10.3390/pathogens11020180
51. Associação Brasileira das Indústrias Exportadoras de Carnes (ABIEC). Beef Report. Perfil da pecuária no Brasil. (2022). Available online at: https://abiec.com.br/wp-content/uploads/Beef-Report-2022_atualizado_jun2022.pdf (acessed October 01, 2022).
Keywords: EBL, bovine, PCR, viral diseases, genetic characterization
Citation: Pereira JG, Silva CdA, Silva LD, Lima CAA, do Rosário CJRM, Silva EMC, Oliveira MdSC, Ribeiro LSdS, Santos HP, Abreu-Silva AL and Melo FA (2023) Diagnosis and phylogenetic analysis of bovine leukemia virus in dairy cattle in northeastern Brazil. Front. Vet. Sci. 9:1080994. doi: 10.3389/fvets.2022.1080994
Received: 26 October 2022; Accepted: 28 December 2022;
Published: 13 January 2023.
Edited by:
Yi Yang, Yangzhou University, ChinaReviewed by:
Dipak Deka, Assam Agricultural University, IndiaCopyright © 2023 Pereira, Silva, Silva, Lima, do Rosário, Silva, Oliveira, Ribeiro, Santos, Abreu-Silva and Melo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: José Gomes Pereira,  am9zZWdvbWVzbmV0b0Bob3RtYWlsLmNvbQ==
am9zZWdvbWVzbmV0b0Bob3RtYWlsLmNvbQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.