 Hatem A. Hamdon1
Hatem A. Hamdon1 Ayman Y. Kassab1
Ayman Y. Kassab1 Einar Vargas-Bello-Pérez2*Galal A. Abdel Hafez3Talaat A. Sayed1Mohsen M. Farghaly3
Einar Vargas-Bello-Pérez2*Galal A. Abdel Hafez3Talaat A. Sayed1Mohsen M. Farghaly3 Ahmed E. Kholif4*
Ahmed E. Kholif4*- 1Department of Animal Production, Faculty of Agriculture, New Valley University, Kharga, Egypt
- 2Department of Animal Sciences, School of Agriculture, Policy and Development, University of Reading, Reading, United Kingdom
- 3Department of Animal Production, Faculty of Agriculture, Assiut University, Assiut, Egypt
- 4Department of Dairy Science, National Research Centre, Giza, Egypt
The study determined the ability of three probiotics to improve the nutritional value of date palm leaves in diets of growing lambs. Twenty male Farafra lambs (26 ± 0.33 kg) were randomly allocated to one of four treatments (n = 6) and fed: a control or basal diet (C; 70% concentrate + 30% date palm leaves without additives) and supplemented with Bacillofort containing 2 × 1011 CFU of Bacillus subtilis/g (BAC treatment), Lacotpro containing 1 × 1012 CFU of Lactobacillus acidophilus/g (LAC treatment) or ZAD containing 6 × 108 CFU of R. albus/g (ZAD treatment) at 4 g of all additives for 150 days. As a result of this study, LAC improved (P < 0.05) growth performance and feed efficiency compared to control. Additives increased (P = 0.001) concentrations of albumin, triiodothyronine, and thyroxine, hemoglobin concentration and red blood cells and decreased (P = 0.001) globulin and urea-N. Additives increased hot carcass (P = 0.040) while BAC increased Longissimus dorsi, meat and fat without affecting water holding capacity compared to other treatments. In the metabolism experiment, BAC increased the digestibility of crude protein, while BAC and ZAD increased the digestibility of dry matter, organic matter, and neutral detergent fiber. Additives did not affect nitrogen (N) intake and urinary N; however, decreased fecal N and increased N balance compared to the control. BAC and ZAD increased ruminal volatile fatty acids concentration compared to the control. Based on our results, Lacotpro could be used to improve growth performance and feed efficiency, while Bacillofort could be used to improve meat quality of in lambs.
Introduction
Semi-arid and arid areas have difficult conditions including low rainfall, resulting in low livestock production with low quality forages (1) making trees and shrubs as sources of small ruminants feed (2). Moreover, in many countries, including Egypt, there is a shortage of feed availability, making the use of unusual feeds and agricultural byproducts as critical options for animal feed (3). The main challenge of using such resources is their low nutritive value (4). Thus, enhancing the nutritive value of fibrous feed is essential (5).
Date palm (Phoenix dactilifera) is an important crop in Egypt, with roughly 650,000 tons of dry matter (DM) annually (6). Approximately 20 kg DM are produced from each palm tree, without a significant utilization. The major challenges of feeding date palm byproducts (i.e., leaves) are the high levels of neutral detergent fiber (NDF; 58%), low levels of crude protein (CP; 5%) and low digestibility (< 50%) compared to other forage feeds (7, 8). Recently, Kholif et al. (7) evaluated the nutritive value of date palm leaves and reported its potential as a sustainable feed for ruminant. With lactating Farafra animals, Kholif et al. (1, 5) observed improved feed utilization and lactation performance when fed ewes on a diet based on date palm leaves and treated with organic acids, fibrolytic enzymes and multi-species probiotics. Feeding some microbes proved to be a beneficial strategy to improve utilization of low-quality feeds and animal performance (1).
In growing animal diets, probiotic additives may offer alternatives to ionophores (9). Probiotics alter rumen microflora populations and fermentation end-products (10–12). Reducing animal health problems is associated with improvements in immunological responses which reduces the need for the use of antimicrobials, and in turn boost growth performance and productive efficiency (11–13). The main problem with probiotic administration in animal diets is the high variability between experiments and, its efficacy depends on the specific product and concentration (14, 15). In many experiments, probiotics improved nutrient absorption and reduced pathogens counts (16), as well as enhanced immune system and competed with invading microbes without leaving any residual toxic effects in an eco-friendly manner (17). Sallam et al. (18) reported that feeding microbial feed additives to growing Barki lambs fed diets that included peanut hay, resulted in improved fiber digestibility, N intake and N retention and enhanced final body weight gain without affecting feed efficiency. Moreover, Hamdon et al. (2) fed Farafra lambs on date palm-based diet supplemented with Bacillus subtilis and Phanerochaete chrysosporium and observed improved growth performance and feed utilization.
However, many experiments evaluated the inclusion of probiotics in diets of animals, the present study evaluated three local developed products with an ingredient (i.e., date palm leaves) that has gained increasing interests in recent years. Therefore, we hypothesized that feeding bacterial feed additives would affect ruminal environment and improve feed digestion resulting in improved performance of growing Farafra lambs. Accordingly, the present study aimed to evaluate the effects of three bacterium containing products in the diet of Farafra lambs fed a control diet based on palm leaf hay. Two experiments were performed: (i) to determine the effects of diets on feed intake, performance efficiency, blood metabolites and carcass characteristics of the growing Farafra lambs, and (ii) to determine the effects of diets on nutrient digestibility of adult Farafra rams.
Materials and methods
This study was performed at the Department of Animal Production, Faculty of Agriculture, New Valley University, New Valley, Egypt (25°26′N and 30°32′E). The area has a desert climate with rare rains (0.18 rainy days, 0.05% of the time), 0.08 millimeters of precipitation and high temperature (23°C during January and 41°C during June to August).
Microbial products
Three probiotics were used in this study: (i) Bacillofort (Bactizad, Al Sharqia, Egypt) is a microbial feed additive that contains 2 × 1011 CFU of B. subtilis, 100 g mannan oligosaccharides, 150 g of β-glucan and corn starch as a carrier. (ii) Lacotpro (Bactizad, Al Sharqia, Egypt) is a microbial feed additive that contains 1 × 1012 CFU of Lactobacillus acidophilus, 100 g mannan oligosaccharides, 150 g β-glucan and corn starch as a carrier. (iii) ZAD (Bactizad, Al Sharqia, Egypt) is microbial feed additive that contains 6 × 108 CFU of Ruminococcus albus, 100 g mannan oligosaccharides, 150 g of β-glucan and corn starch as a carrier.
Experiment 1 (growth performance)
Twenty Farafra lambs with an average body weight of 26 ± 0.33 kg and 5.5 months of age were randomly assigned to four experimental groups (5 lambs per treatment) and fed according to their average initial weights. The experimental period lasted 150 d, consisting of a 15-d adaptation period followed by a 150-d sampling period. Lambs were offered a basal diet based on concentrate and ground wheat straw at 70:30 without additives (control treatment) or supplemented with Bacillofort (BAC treatment), Lacotpro (LAC treatment) or ZAD (ZAD treatment) at 4 g/lamb daily. The concentrate feed mixture contained 500 g corn, 320 g wheat bran, 150 g soybean meal, 20 g limestone, 5 g sodium chloride, and 5 g minerals and vitamins mixture per kg DM. Diets were prepared to meet the nutrient needs of lambs according to the NRC (19) combined with a 10% margin to ensure collection of orts. Ingredients were mixed and the whole concentrate was analyzed (Table 1). Lamb's needs were corrected once every 2 weeks based on changes in body weight. Diets were offered to each lamb individually at 08:00 and 16:00 h in two equal portions. Diets were prepared, mixed 1 day before feeding. Daily allocations of probiotics for each lamb were manually provided individually in 100 g of concentrate before morning feeding at 08:00 h to assure intake. Samples of feed ingredients were obtained daily, composited weekly, and dried at 60 °C in a forced-air oven for 48 h.
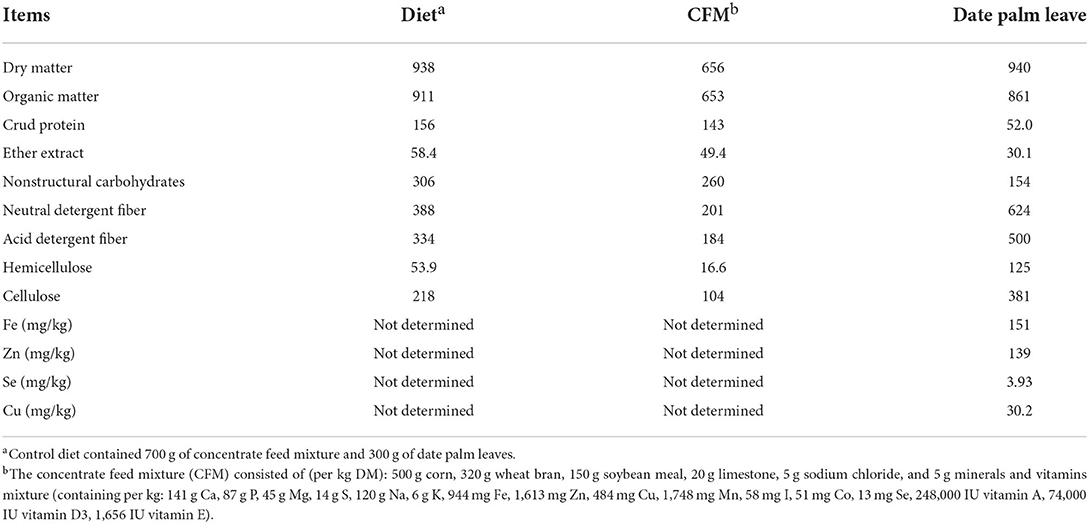
Table 1. Chemical composition (g/kg DM) of date palm leaves, the concentrate feed mixture and basal diet used in experiment 1 and 2.
During sampling, feed intake was recorded as the difference between feed offered and refusals from the previous day. Lambs were biweekly weighed, and feed efficiency was calculated as g of feed per g of weight gain.
As previously reported by Kholif et al. (20), blood samples (about 10 mL) were collected from all the lambs from the jugular vein on days 30, 60, 90, 120 and 150 after the morning feeding at 10:00 h. The last sample was obtained at the time of slaughter at 150 d. Blood samples were immediately centrifuged at 3,000 × g for 15 min, and serum samples were stored at −20 °C until analysis. Stored serum samples were analyzed for total protein, albumin, glucose, urea-N, creatinine, total cholesterol, alanine aminotransferase (ALT), aspartate aminotransferase (AST) cholesterol, triglyceride, triiodothyronine (T3), thyroxine (T4) and globulin as reported previously (2).
As previously reported by Rivero et al. (21), the hematological parameters were assessed by using automatic, fully Digital Hematology Analyzer (Shenzhen Mind ray Auto Hematology Analyzer (Model Bc-3200, Shenzhen Mind ray Biomedical Electronics Co. Hamburg 20,537, Germany). These parameters included a total count of white blood cells (WBC), the total count of red blood cells (RBC), hemoglobin, hematocrit (HCT), mean corpuscular hemoglobin (MCH), mean corpuscular volume (MCV), and mean corpuscular hemoglobin concentration (MCHC).
At the end of the experimental period, three random animals from each group were slaughtered. Fasting body weight was recorded and immediately after slaughter, the hot carcass weight was recorded, and the weights of the head, pelt, liver, lungs, heart, spleen, kidneys, kidney fat, omental fat, testes, trachea, and heart were recorded. Full methodological details for charctass traits have been reported in a companion paper (2).
Water holding capacity (WHC) was measured by weighing about 0.3 g (W1) of meat in a filter paper (Whatman No.1) and subjected to pressure of 1,000 gm for 10 min, then it was weighed again (W2). WHC was estimated as a percentage according to the following formula: WHC % = [W2/W1] × 100. WHC was expressed as the percentage of loss related to the initial weight (22).
Experiment 2 (digestibility experiment)
As in Experiment 1, 12 adult rams weighing 50 ± 0.25 kg were randomly allocated into four treatment groups to determine the nutrient digestibility using the fecal bag technique. Animals received the same diets as in Experiment 1. Rams were housed individually in metabolic cages (70 cm width × 120 cm high × 150 cm length). The experiment lasted 22 d, consisting of a 15-d adaptation period followed by 7 d of sample collection. Feeds and orts also were dried before grinding through a 1-ml screen for chemical analysis. Feces were collected daily, and 10% of the fecal matter (by weight) was dried at 70 °C for 24 h, combined and ground through a 1-mm mill screen for subsequent chemical analysis. Apparent digestion coefficients of the nutrients were calculated by expressing the difference between the nutrient content in both consumed feed and feces as a percentage of intakes.
During the collection period, urine was also collected in large polyethylene bags containing 20 ml of 10% H2SO4 to prevent losses of ammonia N. Daily urine sample of 10% was collected for each ram and stored in a glass bottle in a refrigerator until it was used for nitrogen (N) determination to calculate the N balance.
Ruminal contents (about 150 mL) were monthly sampled at 3 h after the morning feeding to determine the pH and fermentation end-products. After initial discarding of 50 ml ruminal fluid to prevent contamination with saliva, 100 ml ruminal fluid were collected by utilizing a stomach tube, and samples removed from each lamb were strained through 4 layers of cheesecloth. Ruminal fluid pH was determined using a pH meter (Beckman, model 45, USA).
A subsample of 5 mL ruminal fluid was preserved with 5 mL of 0.2 M HCl for ammonia-N analysis (23), and 0.8 mL of ruminal fluid was mixed with 0.2 mL of a solution containing 250 g of metaphosphoric acid/L for total volatile fatty acids (VFA) analysis. Samples were preserved at −20°C until analyses (24).
Samples of fermented fluid (4 mL) were individually combined with 4 mL of methyl green-formalin-saline solution and stored in a refrigerator at 4°C until analysis of bacterial and protozoal count following methods from Dehority (25). Total bacteria concentration was determined using a Petroff-Hausser counting chamber (Hausser Scientific®, 3900, Horsham, PA) and a phase contrast microscope at a magnification of 100 ×.
The nutritive value of diets as total digestible nutrients (TDN), digestible energy (DE), metabolizable energy (ME) were calculated according to NRC (26) equations, while the unité fourragère du lait (UFL; net energy requirements for lactation equivalent to 1 kg of standard air-dry barley) was used according to INRA (27) equation.
Feed, orts, and fecal samples were ground through a 1-mm screen using a Wiley mill and analyzed for DM (ID 930·15), ash (ID 942·05), N (ID 954·01), and ether extract (EE) (ID 920·39), according to the official methods of the AOAC (23). NDF levels were determined by the procedure of Van Soest et al. (28) with sodium sulphite. Acid detergent fiber (ADF) concentrations (method ID 973·18) were analyzed as described by the AOAC (23). Non-structural carbohydrate [NSC = 1,000 – (NDF + CP + EE + ash)] and organic matter (OM = 1,000 – ash) concentrations were determined.
Statistical analysis
Data for total gain, carcass characteristics and composition, and nutrient digestibility were analyzed using the PROC GLM procedure of SAS, online version (SAS Inst. Inc. Cary, NC, USA) for a complete randomized design with individual animal as the experimental unit. The model included the effect of treatment. Measurements (body weight changes by difference, feed intake and conversion, and blood measurements) recorded daily or weekly were analyzed as repeated measures. When the treatment F-test was significant at P < 0.05, means were compared by applying the probability of difference option of the least squares means statement. The treatment × week interaction was non-significant (i.e., P > 0.05) for most of the measurements; thus, only the main effects of treatments were reported.
Results
Feed intake, growth performance and feed efficiency
Probiotics did not impact feed intake (Table 2). Initial weight did not vary between treatments. However, LAC improved final body weight (P = 0.120), total weight gain (P = 0.042) and daily gain (P = 0.033), and feed efficiency (P = 0.011) while ZAD decreased these parameters compared to the control.
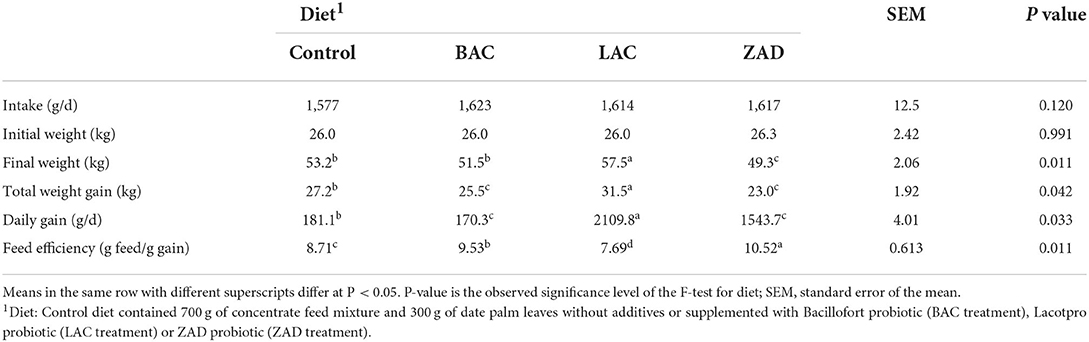
Table 2. Feed intake, growth performance, and feed efficiency of growing Farafra lambs fed diets containing date palm leaves treated with different probiotics.
Blood measurements
Compared to control, additives did not impact blood glucose, AST, ALT, and triglyceride; however, additives increased (P = 0.001) concentrations of albumin, T3 and T4 and decreased (P = 0.001) globulin and urea-N (Table 3). BAC and ZAD increased concentrations of blood total proteins (P = 0.001) and decreased blood creatinine (P = 0.024). BAC and ZAD lowered (P = 0.031) cholesterol compared to control.
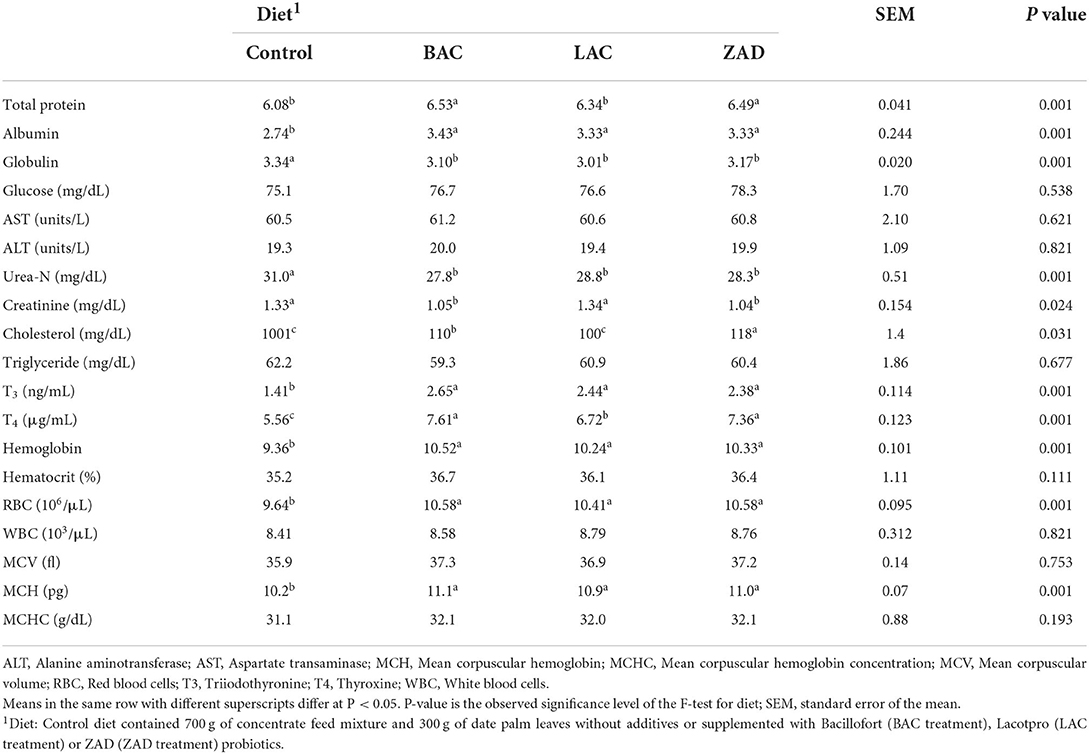
Table 3. Blood metabolites (g/dL) and hematological parameters of growing Farafra lambs fed diets containing date palm leaves treated with different probiotics.
Additives did not alter blood HCT, WBC, MCV or MCHC (Table 3). Raised (P = 0.001) blood hemoglobin and RBC were observed with the additive treatments.
Carcass characteristics and meat composition
Increased fasting weight (P = 0.011) and legs weight (P = 0.022), and lowered brisket (P = 0.01) and flank weights (P = 0.016) were observed with BAC and ZAD; however, all additives increased hot carcass (P = 0.040), shoulder weight (P = 0.021), loin weight (P = 0.017) and rack weight (P = 0.021) compared to the control (Table 4). The heaviest Longissimus dorsi muscle was observed with BAC.
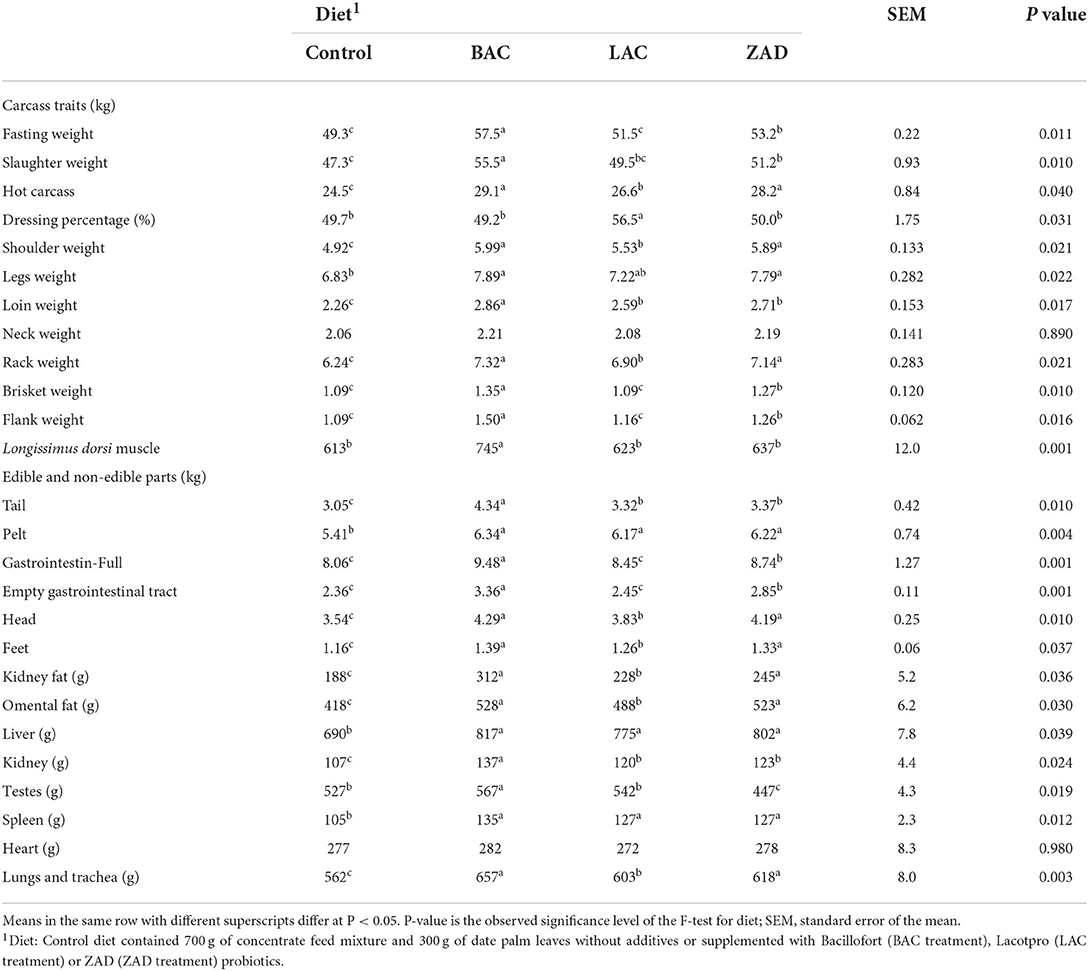
Table 4. Carcass traits and edible and non-edible parts of growing Farafra lambs fed diets containing date palm leaves treated with different probiotics.
Additives increased the weights of tail (P = 0.010), pelt (P = 0.004), head (P = 0.01), feet (P = 0.037), kidney fat (P = 0.036), omental fat (P = 0.03), liver (P = 0.039), kidney (P = 0.024), spleen (P = 0.012), lungs and trachea (P = 0.003) compared to the control (Table 4). BAC increased the weight of testes compared to other treatments.
BAC increased the weights of Longissimus dorsi muscle (P = 0.022), meat (P = 0.003) and fat (P = 0.039) without affecting bone weight or WHC (Table 5). LAC and ZAD decreased the proportion of crude fat (P = 0.02) without influencing crude protein or ash.

Table 5. Gross and chemical compositions, and physicochemical properties of carcass of growing Farafra lambs fed diets containing date palm leaves treated with different probiotics.
Nutrient digestibility and nitrogen balance
However, BAC increased the digestibility of CP, while BAC and ZAD increased the digestibility of DM, OM and NDF (Table 6). All additives increased the digestibility of NSC; however, LAC did not change digestibility of DM, CP, or NDF compared to the control. Both of BAC and ZAD increased the digestible CP, TDN, DE, ME, NEL and UFL compared to the control treatment.
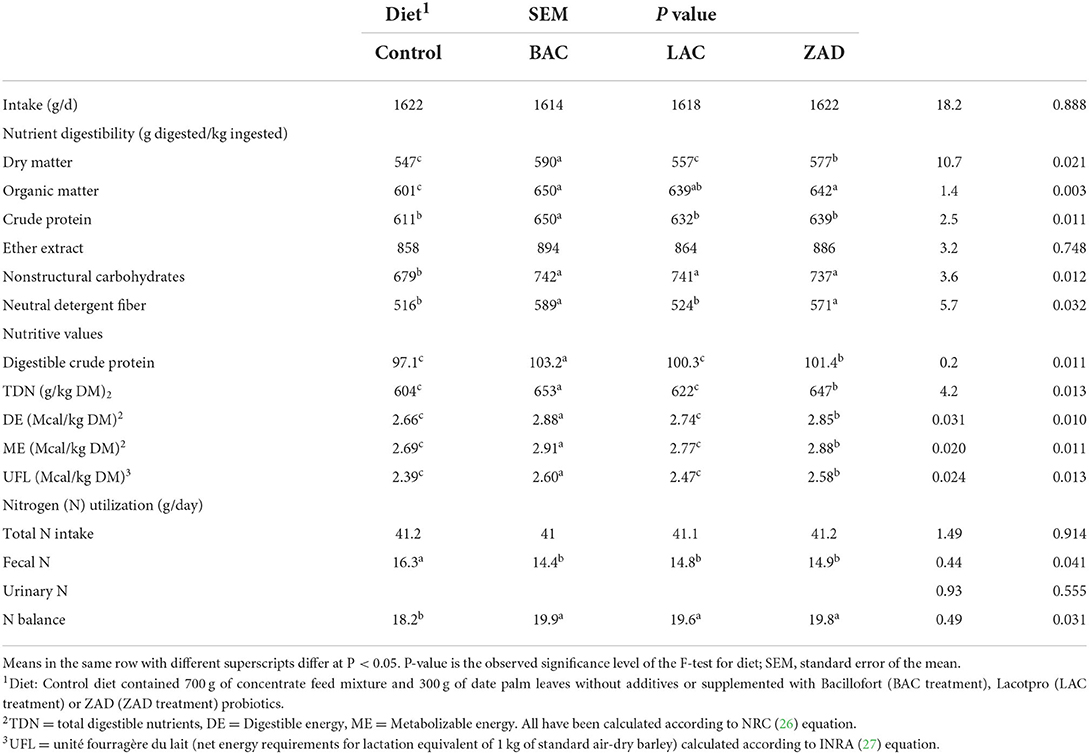
Table 6. Feed intake, nutrient digestibility, diet nutritive values and nitrogen (N) balance of adult Farafra rams fed with diets containing date palm leaves treated with different probiotics.
Compared to control, additives did not influence N intake or urinary N; however, decreased fecal N and increased N balance (Table 6).
Ruminal fermentation
Additives did not change ruminal pH or ammonia-N (Table 7). Both of BAC and ZAD increased ruminal VFA compared to the control.
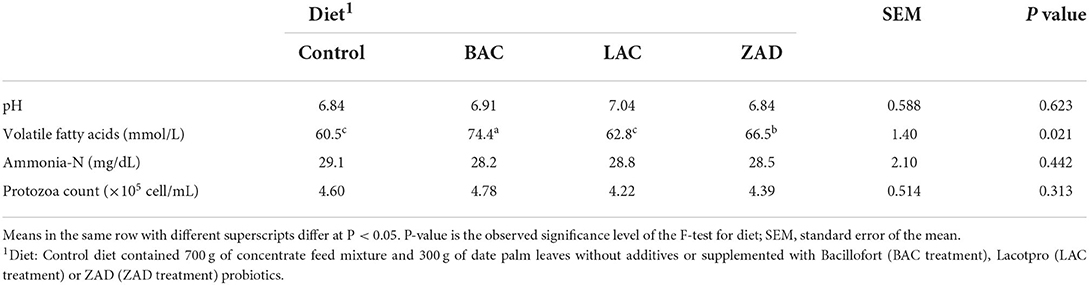
Table 7. Ruminal fermentation of adult Farafra rams fed with diets containing date palm leaves treated with different probiotics.
Discussion
The inclusion of probiotics in animal diets has been evaluated in many experiments. The novelty in the present study is the evaluation of three local developed probiotics added to diets containing date palm leaves that recently gained increasing interests in the sustainable sheep nutrition under desert and tropical conditions. More experiments are required to explore the effects of these probiotics in animal diets to explore their modes of action in the gastrointestinal tract of ruminants.
Feed intake, growth performance and feed efficiency
Microbial feed additives did not affect feed intake and that somehow explains that there was unchanged feed palatability. In a previous experiment under the same environmental conditions, Hamdon et al. (2) observed that feeding B. subtilis and P. chrysosporium to growing Farafra lambs had weak effects on feed intake. However, others (18) observed increased feed intake when lambs were fed diets based on peanut hay supplemented with probiotics. Animal used, growth stage, environmental condition, dose, strains, duration and frequency or compositions of animal diets may partially explain the discrepancy between experiments (29).
The insignificant differences between initial weights of lambs in all treatments reflect the random distribution of animal before feeding the experimental treatments, and that any changes in the measured parameters are mainly due to the additives. One of the main objectives of the feedlot industry is to boost animal growth performance. The LAC treatment improved final body weight by 8.1%, total weight gain by 15.8% and daily gain by 15.8% compared to the control, which confirm the positive relationship observed between probiotic supplementation and animal growth performance (18, 30). As observed in the digestibility trial, LAC did not affect nutrient digestibility, which may indicate that the digestibility is not the main reason for the observed growth improvements. However, it may indicate that LAC could be recommended for growing animals while BAC, could be recommended for adult animals to improve their performance and meat quality and these will be discussed later.
The improved daily gain with probiotics supplementation to diets of ruminants are mainly due to improvement in nutrient absorption and reduction in the counts of pathogens (16). Using microbial strains improves immune system without leaving residual toxic effects (17), and this results in an improved daily gain and final body weight (18). Using additives throughout their first months of life improves gut microbiota and growth performance (11). LAC contains L. acidophilus which prevents gastrointestinal infection by pathogens or by producing antimicrobials (31, 32), and may be improved the microbial balance in the rumen of lambs compared to B. subtilis or R. albus (11), especially when be fed to animals consuming a fibrous diet (18). Increasing microbial protein synthesis with LAC additive may have increased amino acid supply at the post-ruminal level resulting in increasing daily gain (18, 33). Moreover, the results indicate that LAC increased the antagonism of pathogenic organisms through antimicrobial effects such as stimulation of host defines mechanisms and inhibition of bacterial toxins (10, 33). Additionally, probiotics increase the release of different endogenous substances, such as antibacterial substances, antioxidants, growth factors and coagulating agents, causing enhanced growth (9, 33, 34), and thereby lamb's daily gains.
The ZAD treatment decreased growth performance, which confirm the importance of selecting the suitable strain or product of probiotics. Different probiotics mean different composition, different specific activity of the probiotic strains and even different strains may have distinct effects based on enzymatic activities depending on host species (35).
Increasing growth rate without affecting feed intake in the LAC treatment was reflected as it increased feed efficiency by 11.7%, while the decreased growth rate in ZAD treatment was reflected as it decreased feed efficiency by 20.8% compared to the control. Improving feed efficiency results in improved nutrient utilization. The improved feed efficiency may be related to improved efficiency in nutrient utilization, and thereby improving N retention while reducing the excretion of essential nutrients (32). Hamdon et al. (2) observed that feeding date palm leaves treated with probiotic improved feed efficiency between 15.3 to 23.6%.
Blood measurements
As the experiment evaluates new feed additives, blood analysis gives the opportunity to study several metabolites and other constituents that helps to detect nutritional, environmental, or physical stress. Blood components were within the ranges reported for healthy animals (21). It was expected that increasing nutrient digestibility and ruminal VFA production will increase glucose concentration, but this was not observed which may indicate that the absorption rate of glucose from treatments was not enough to be reflected in blood (36). The minor impacts of treatments on blood glucose are consistent with those observed by Abd El-Wahab et al. (16) who reported insignificantly differed blood glucose, total protein, and albumin with feeding probiotic to growing calves. However, (37) El-Mehanna et al. (37) observed an increase of 13% in blood glucose of lambs fed with probiotics. The minor effects of treatments on the concentration of blood AST and ALT, which were within the normal physiological range (38), indicate that additives did not affect liver function (39).
Additives increased the concentrations of albumin b while decreasing globulin and urea-N suggesting that they improved nutrient utilization without impairing kidney performance and protein catabolism in muscles (40). BAC and ZAD increased blood total proteins by 7.4 and 6.7%, respectively and decreased blood creatinine by 21.1 and 21.8%, respectively indicating an enhanced nutritional status of lambs and improved kidney function (38). Increased serum total protein and albumin can be related to higher nutrient supply in ewes fed with additives.
BAC, LAC and ZAD increased the concentrations of thyroid hormones (T3 by about 87.9, 73.0 and 68.8%, respectively and T4 by about 36.9, 20.9 and 32.4%, respectively). Mousa et al. (41) observed increased T3 and T4 hormones with probiotic supplementation to lambs. T3 and T4 work together to regulate energy usage and are key for controlling weight, body temperature, muscle strength, and nervous system. Thyroid hormones play important roles in the regulation of triglycerides, cholesterol metabolism, lipoprotein homeostasis, and the induction of the genes involved in glycolysis and gluconeogenesis (42). Probiotics positively affect thyroid-stimulating hormone-releasing in the hypothalamus (42).
BAC and ZAD lowered cholesterol concentration by 9.2 and 16.9%; however, El-Mehanna et al. (37) observed that probiotics supplementation to growing lambs did not affect cholesterol or insulin but increased serum glucose levels. The lowered cholesterol concentration may be due to the inhibition of cholesterol synthesis or the direct absorption of cholesterol (43). Probiotics decrease cholesterol through the deconjugation of bile acids, and the increase in the degradation of cholesterol throughout the gastrointestinal tract (44).
The increased blood hemoglobin (by 12.4, 9.4, and 10.4%, respectively), RBC (by 9.8, 8.0, and 9.8%, respectively), and MCH (by 9.4, 7.2, and 8.7%, respectively) with for BAC, LAC and ZAD treatments indicated that there was improved homeostatic impacts on the treated animals. The is scarce research on the effects of probiotics on hematological traits of animals and makes the explanation of our results challenge. These effects may be related to an increased synthesis of vitamin B12 and enhanced iron salt absorption by the small intestine, resulting in greater hematopoiesis (41). The unchanged WBC numbers with probiotic supplementation somehow indicate that animals were not under stress during the experiment (36). El-Mehanna et al. (37) observed unaffected RBC, hemoglobin levels and HCT value, while increased numbers of WBC with probiotic supplementation to growing lambs. Moreover, Sarwar et al. (45) observed increased Hb, PCV and RBC in Kajli lambs supplemented with probiotics.
Carcass traits and meat composition
Scarce data concerning the effect of feeding B. subtilis, L. acidophilus or R. albus on carcass characteristics is available. BAC and ZAD increased fasting weight indicating anabolic effects of these additives. Such results may be due to probable increased microbial protein synthesis. Additives increased hot carcass weight, which is similar to Hamdon et al. (2) who observed improved hot carcass weight and dressing with B. subtilis and Phanerochaete chrysosporium administration to growing lambs. Increasing daily gain and body weight gain of lambs fed LAC can partially explain these results found for LAC but not in the other treatments. El-Mehanna et al. (37) observed that feeding probiotics to growing lambs did not affect carcass weight, dressing percentage and offal weights; however, increased chest depth, eye muscle area and breast weight.
The BAC treatment increased the weight of Longissimus dorsi muscle by 21.5%. The eye muscle area is considered as an indicator of tissue growth. Lahiri et al. (46) showed that gut microbiota impacts muscle cell metabolism across gut microbiota–skeletal muscle axis generating favorable effects in animals. Final body weight of lambs largely explains the changes in Longissimus dorsi characteristics (47).
BAC increased meat weight by 18.4%, while ZAD increased it by 12%. Moreover, BAC increased fat weight by 42.4%, indicating a differed ruminal fermentation and energy and fat metabolism pathways with the additives, which has been discussed earlier in this section. Increasing acetate-to-propionate ratio (48) and acetate contributes to increasing fat deposition in ruminants (49). Increased internal fat may also reflect the greater energy retention observed for those lambs that received probiotics. Crude fat is important to ensure the flavor, aroma, and juiciness of the muscle (50). Increasing the portion of fat is a result of intramuscular fat deposition, which benefits marbling, carcass quality, and isensorial properties (47).
Nutrient digestibility and nitrogen balance
In the performance experiment, growing animals were used while adult rams were used for the digestibility experiment were and that brought differential effects that could be attributed to growth stages. Hence, data from the digestibility trial somewhat explain what was noted in the lamb trial.
The BAC treatment increased the digestibility of CP by 6.3%; however, Hamdon et al. (2) observed lowered CP digestibility with feeding lambs fed on a diet containing date palm leaves supplemented with microbial feed additives. They explained that results due to the presence of a substantial amount of CP related to the fiber fraction, which lowered disposal of these proteins for the animal (51). Probiotic type and grinding degree of date palm leaves may have been different between the one used in their experiment contrasted to the one applied in the present study, and this may partially explain the difference between their experiment and ours.
It was expected that the presence of high concentrations of carotenoids, isoflavones, lignans, flavonoids, tannins, and sterols in ate palm leaves can negatively affect nutrient digestibility (52); however, these effects were not observed as the values of nutrient digestibility were within the ranges reported for ruminants. Such results indicate the ability of the evaluated additives to cover the negative effects of bioactive components on nutrient digestion. The BAC and ZAD improved digestibility of DM (by 7.8 and 5.5%, respectively), OM (by 8.2 and 6.9%, respectively) and NDF (by 14.1 and 10.7%, respectively), while BAC, LAC and ZAD increased digestibility of NSC by 9.2, 9.1 and 8.5%, respectively. Many modes of action were proposed to explain these results (11, 12). In the present experiment, the improved CP and fiber digestibility indicate that feeding microbial feed additives may loosen the association among the fiber bundles and between protein and fiber fractions in date palm leaves (2). Although activity of ruminal cellulolytic microbial populations was not measured in the present experiment, microbial supplementation could produce a tonic level of lactate, which will then boost a basal abundance of lactate utilizing bacteria in the rumen, thus stabilization of pH as observed in the metabolism trial in the present experiment (8, 32). The improved ruminal environment may increase the fiber degrading microbial communities in the rumen resulting in improved nutrient digestion and synthesis of microbial proteins (15). Furthermore, improved digestibility may enhance the interaction of microbes in the additives with the ruminal microbial flora (53, 54) and increase enzyme activity in the gastrointestinal tract resulting in improved nutrient digestibility (55, 56). Another reason for the improved nutrient digestibility is the probable beneficial inhibitory effects of probiotics on the gastrointestinal infection by pathogens (32). Additionally, experiments (57) showed that probiotic supplementation improves ruminal cellulolytic microbial populations and buffers rumen pH, leading to enhanced nutrient digestion (33) and synthesis of microbial proteins (15).
Additives decreased fecal N by 11.7, 9.2, and 8.6% and increased N balance by 9.3, 7.7, and 8.8% for BAC, LAC and ZAD, respectively indicating improved N utilization by animals. Sallam et al. (18) observed improved N balance by 22% when animals were fed a diet supplemented with probiotics. Moreover, Kawauchi et al. (58) observed that feeding cows on a diet supplemented with B. subtilis did not affect the amounts of N loss in the feces or urine or retained N. Increased N retention in body tissues may explain the increased daily gain in animals fed additives. Lack of differences in ruminal ammonia-N, blood urea-N concentrations and N excreted revealed an improved utilization of nutrients (59). These effects are desirable from an environmental point of view as N is rapidly mineralized from NH3/NH4+ to nitrous oxide, which has a huge global warming potential of about 265–298 compared with 21 for methane.
Ruminal fermentation
Rumen pH (60) and ammonia-N (61) were greater than rates required for fiber digestion and microbial protein synthesis. Probiotics did not impact rumen pH, and this is important to avoid changes of rumen microbial population from fibrolytic to amylolytic microbes (62). Probiotics work effectively to balance ruminal pH (11, 12). In the present experiment, the values of ruminal pH indicate that lambs kept their rumen near neutrality (6.84–7.04), which may be related to feeding lambs on a fibrous diet based on date palm leaves. Abd El-Wahab et al. (16) observed no effects on ruminal pH of growing calves fed probiotics.
Probiotics did not impact ammonia N, indicating minimal effects on dietary protein breakdown and digestion, and the uptake of ammonia by bacteria (58). However, BAC increased CP digestibility indicating increased uptake of ammonia by bacteria and increased microbial protein in the rumen. Such assumption may be partially explaining the increased meat proportion in the carcass of lambs fed BAC. The rumen microflora starts to convert a part of ammonia-N into microbial proteins, which represent an essential source of N for the growing animal, and another part is recycled as urea (63). Similarly, Kawauchi et al. (58) observed minor effects on ruminal ammonia-N concentration when feeding B. subtilis to cows.
The BAC and ZAD increased ruminal VFA concentration by 33 and 9.9%, respectively, which may be related to an increased digestibility of OM, NSC and NDF. Increasing microbial population can result in increased VFA with positive impacts on energy supply to the animal (32). Increasing the production of VFA ensures enough energy formation and stimulates the growth in the surface, length, and width of rumen papillae (64). Increased total VFA concentration likely resulted from the increased fermentable carbohydrate availability, ruminal microbial activity, and fermentation rate.
Conclusions
In arid environments, date palm leaves can be utilized as forage source for lambs. However, improving its utilization with probiotic additives is recommended. The responses to different products of probiotics containing different types and proportions of microbes differ. Probiotics positively affected blood chemistry and improved N balance and carcass characteristics. Bacillofort increased ruminal volatile fatty acids production and improve meat quality in lambs as increasing the weight of Longissimus dorsi muscle, meat, and fat. However, Lacotpro could be used to improve growth performance and feed efficiency.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The protocol of the experiment was reviewed and approved by the Institutional Animal Care and Use Committee of the Faculty of Agriculture, New Valley University, New Valley, Egypt.
Author contributions
Conceptualization: HH, AYK, GA, TS, MF, and AEK. Methodology, investigation, and resources: HH, AYK, GA, TS, and MF. Validation and data curation: HH and AEK. Project administration: HH and AYK. Formal analysis: HH, TS, and AEK. Visualization: HH, TS, and AYK. Supervision: HH, AYK, GA, and MF. Writing—original draft preparation: AEK. Writing—review and editing: EV-B-P and AEK. All authors have read and agreed to the published version of the manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Kholif AE, Hamdon HA, Gouda GA, Kassab AY, Morsy TA, Patra AK. Feeding date-palm leaves ensiled with fibrolytic enzymes or multi-species probiotics to Farafra ewes: intake, digestibility, ruminal fermentation, blood chemistry, milk production and milk fatty acid profile. Animals. (2022) 12:1107. doi: 10.3390/ani12091107
2. Hamdon HA, Kholif AE, Mahmoud GB, Khalifa AMA, Abdel Ati MNM. Enhancing the utilization of palm leaf hay using Bacillus subtilis and Phanerochaete chrysosporium in the diet of lambs under desert conditions. Ann Anim Sci. (2020) 20:1395–409. doi: 10.2478/aoas-2020-0052
3. Mahrous AA, El-Tahan AAH, Hafez YH, El-Shora MA, Olafadehan OA. Hamdon H. Effect of date palm (Phoenix dactylifera L) leaves on productive performance of growing lambs. Trop Anim Health Prod. (2021) 53:1–8. doi: 10.1007/s11250-020-02493-2
4. Kholif AE, Gouda GA, Morsy TA, Patra AK. The effects of replacement of berseem hay in total mixed rations with date palm leaves ensiled with malic or lactic acids at different levels on the nutritive value, ruminal in vitro biogas production and fermentation. Biomass Convers Biorefin. (2022). doi: 10.1007/s13399-022-02508-y
5. Gouda GA, Kholif AE, Hamdon HA, Kassab AY, Patra AK. Utilization of waste date palm leaves biomass ensiled with malic or lactic acids in diets of Farafra ewes under tropical conditions. Animals. (2022) 12:1432. doi: 10.3390/ani12111432
6. Shawky AB, El-Sharabasy SF. Date palm status and perspective in Egypt, date palm genetic resources and utilization. In:Al-Khayri J, Jain SM, Johnson D, , editors. Netherlands: Springer (2015) p. 75–123.
7. Kholif AE, Gouda GA, Morsy TA, Matloup OH, Fahmy M, Gomaa AS, et al. Dietary date palm leaves ensiled with fibrolytic enzymes decreased methane production, and improved feed degradability and fermentation kinetics in a ruminal in vitro system. Waste Biomass Valorization. (2022) 13:3475–88. doi: 10.1007/s12649-022-01752-7
8. Kholif AE, Gouda GA, Patra AK. The sustainable mitigation of in vitro ruminal biogas emissions by ensiling date palm leaves and rice straw with lactic acid bacteria and Pleurotus ostreatus for cleaner livestock production. J Appl Microbiol. (2022) 132:2925–39. doi: 10.1111/jam.15432
9. Hall JB, Bloomsburg MR, Goddard SA. Effect of a lactobacillus fermentation product on postweaning heifer performance. Transl Anim Sci. (2022) 6:1. doi: 10.1093/tas/txac015
10. Elghandour MMY, Salem AZM, Castañeda JSM, Camacho LM, Kholif AE, Chagoyán JCV. Direct-fed microbes: A tool for improving the utilization of low quality roughages in ruminants. J Integr Agric. (2015) 14:526–33. doi: 10.1016/S2095-3119(14)60834-0
11. Kober AKMH, Riaz Rajoka MS, Mehwish HM, Villena J, Kitazawa H. Immunomodulation potential of probiotics: a novel strategy for improving livestock health, immunity, and productivity. Microorganisms. (2022) 10:388. doi: 10.3390/microorganisms10020388
12. Nalla K, Manda NK, Dhillon HS, Kanade SR, Rokana N, Hess M, et al. Impact of probiotics on dairy production efficiency. Front Microbiol. (2022) 13:1519. doi: 10.3389/fmicb.2022.805963
13. Broadway PR, Carroll JA, Burdick Sanchez NC, Callaway TR, Lawhon SD, Gart E, et al. Bacillus subtilis PB6 supplementation in weaned Holstein steers during an experimental Salmonella challenge. Foodborne Pathog Dis. (2020) 17:521–8. doi: 10.1089/fpd.2019.2757
14. Markowiak P, Slizewska K. The role of probiotics, prebiotics and synbiotics in animal nutrition. Gut Pathog. (2018) 10:21. doi: 10.1186/s13099-018-0250-0
15. Uyeno Y, Shigemori S, Shimosato T. Effect of probiotics/prebiotics on cattle health and productivity. Microbes Environ. (2015) 30:126–32. doi: 10.1264/jsme2.ME14176
16. Abd El-Wahab A, Abbas T, Orma AE-H, Ibrahim T. Effects of dietary probiotic supplementation on growth, rumen development and selected blood metabolites of growing calves. Mansoura Vete Med J. (2021) 22:166–71. doi: 10.21608/mvmj.2021.47831.1012
17. Yeoman CJ, White BA. Gastrointestinal tract microbiota and probiotics in production animals. Annu Rev Anim Biosci. (2014) 2:469–86. doi: 10.1146/annurev-animal-022513-114149
18. Sallam SMA, Kholif AE, Amin KA, Nour El-Din ANM, Attia MFA, Matloup OH, et al. Effects of microbial feed additives on feed utilization and growth performance in growing Barki lambs fed diet based on peanut hay. Anim Biotechnol. (2020) 31:447–54. doi: 10.1080/10495398.2019.1616554
19. NRC. Nutrient Requirements of Small Ruminants: Sheep, Goats, Cervids, and New World Camelids. Washington, DC, USA: National Academy Press. (2007).
20. Kholif AE, Morsy TA, Abd El Tawab AM, Anele UY, Galyean ML. Effect of supplementing diets of Anglo-Nubian goats with soybean and flaxseed oils on lactational performance. J Agric Food Chem. (2016) 64:6163–70. doi: 10.1021/acs.jafc.6b02625
21. Rivero N, Salem AZM, Ayala M, Elghandour MMY, Kholif AE, Barbabosa A, et al. Influence of Salix babylonica extract, exogenous enzyme of xylanase and their combination on blood haematological and biochemical profile in sheep and goats. Indian J Animal Sci. (2016) 86:1140–4.
22. Sami AS, El-Asheen AK, Radwan MA. Carcass merits, chemical composition and physical properties of beef under unconventional fattening regimen. J Anim Vet Adv. (2012) 11:3732–7. doi: 10.3923/javaa.2012.3732.3737
23. AOAC. Official Methods of Analysis of AOAC International. 18th ed. In: Latimer GW, editor. Washington DC: Oxford University Press Oxford. (2019). doi: 10.1093/9780197610138.001.0001
24. Ahmed MH, Elghandour MMY, Salem AZM, Zeweil HS, Kholif AE, Klieve AV, et al. Influence of Trichoderma reesei or Saccharomyces cerevisiae on performance, ruminal fermentation, carcass characteristics and blood biochemistry of lambs fed Atriplex nummularia and Acacia saligna mixture. Livest Sci. (2015) 180:90–7. doi: 10.1016/j.livsci.2015.06.019
25. Dehor¡ty BA. Laboratory Manual for Classification and Morphology of Rumen Ciliate Protozoa. 1st ed. Boca Raton, FL, USA: CRC Press (1993). doi: 10.1201/9781351073912
26. NRC. Nutrient Requirements of Dairy Cattle. 7th ed. Washington, D.C., D.C., USA: National Academies Press (2001). doi: 10.17226/9825
27. INRA. INRA Feeding System for Ruminants. The Netherlands: Wageningen Academic Publishers. (2018). doi: 10.3920/978-90-8686-292-4
28. Van Soest PJ, Robertson JB, Lewis BA. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J Dairy Sci. (1991) 74:3583–97. doi: 10.3168/jds.s0022-0302(91)78551-2
29. Khan RU, Naz S, Dhama K, Karthik K, Tiwari R, Abdelrahma MM, et al. Direct-fed microbial: beneficial applications, modes of action and prospects as a safe tool for enhancing ruminant production and safeguarding health. Int J Pharmacol. (2016) 12:220–31.
30. Abd El Tawab AM, Kholif AE, Hassan AM, Matloup OH, Abo El-Nor SA, Olafadehan OA, et al. Feed utilization and lactational performance of Friesian cows fed beet tops silage treated with lactic acid bacteria as a replacement for corn silage. Anim Biotechnol. (2020) 31:473–82. doi: 10.1080/10495398.2019.1622556
31. Kritas SK, Govaris A, Christodoulopoulos G, Burriel AR. Effect of Bacillus licheniformis and Bacillus subtilis supplementation of ewe's feed on sheep milk production and young lamb mortality. J Vet Med Series A. (2006) 53:170–3. doi: 10.1111/j.1439-0442.2006.00815.x
32. Arowolo MA, He J. Use of probiotics and botanical extracts to improve ruminant production in the tropics: a review. Animal Nutr. (2018) 4:241–9. doi: 10.1016/j.aninu.2018.04.010
33. Retta KS. Role of probiotics in rumen fermentation and animal performance: A review. Int J Livestock Product. (2016) 7:24–32. doi: 10.5897/ijlp2016.0285
34. Pant B, Lohani V, Mishra A, Trakroo MD, Tewari H. Effect of probiotic supplementation on growth of carp fingerlings. Natl Acad Sci Lett. (2019) 42:215–20. doi: 10.1007/s40009-018-0733-z
35. Adjei-Fremah S, Ekwemalor K, Worku M, Ibrahim S. Probiotics and ruminant health. In:Enany S, , editor. Probiotics - Current Knowledge and Future Prospects. London, UK: InTech (2018). p. 133–150 doi: 10.5772/intechopen.72846
36. Mallaki M, Hosseinkhani A, Taghizadeh A, Hamidian G, Paya H. The effect of Bovine Lactoferrin and probiotic on performance and health status of ghezel lambs in preweaning phase. Iran J Appl Anim Sci. (2021) 11:101–10.
37. El-Mehanna SF, Abdelsalam MM, Hashem NM, El-Azrak KEM, Mansour MM, Zeitoun MM. Relevance of probiotic, prebiotic and synbiotic supplementations on hemato-biochemical parameters, metabolic hormones, biometric measurements and carcass characteristics of sub-tropical Noemi lambs. Int J Animal Res. (2017) 1:1–12. doi: 10.28933/ijar-2017-09-3001
38. Zaitsev SY, Bogolyubova N, Zhang X, Brenig B. Biochemical parameters, dynamic tensiometry and circulating nucleic acids for cattle blood analysis: A review. PeerJ. (2020) 8:e8997. doi: 10.7717/peerj.8997
39. Pettersson J, Hindorf U, Persson P, Bengtsson T, Malmqvist U, Werkström V, et al. Muscular exercise can cause highly pathological liver function tests in healthy men. Br J Clin Pharmacol. (2008) 65:253–9. doi: 10.1111/j.1365-2125.2007.03001.x
40. Hosten AO. BUN and Creatinine. In:Walker HK, Hall WD, Hurst JW, , editors. Clinical Methods: The History, Physical, and Laboratory Examinations. Boston, MA: Butterworths. (1990). p. 874–878
41. Mousa GA, Allak MA, Shehata MG, Hashem NM, Hassan OGA. Dietary supplementation with a combination of fibrolytic enzymes and probiotics improves digestibility, growth performance, blood metabolites, and economics of fattening lambs. Animals. (2022) 12:476. doi: 10.3390/ani12040476
42. Oetting A, Yen PM. New insights into thyroid hormone action. Best Pract Res Clin. Endocrinol Metab. (2007) 21:193–208. doi: 10.1016/j.beem.2007.04.004
43. Brown AJ, Coates HW, Sharpe LJ. Cholesterol synthesis. In: Biochemistry of Lipids, Lipoproteins and Membranes. Elsevier. (2021). p. 317–355 doi: 10.1016/b978-0-12-824048-9.00005-5
44. Yen PM, Feng X, Flamant F, Chen Y, Walker RL, Weiss RE, et al. Effects of ligand and thyroid hormone receptor isoforms on hepatic gene expression profiles of thyroid hormone receptor knockout mice. EMBO Rep. (2003) 4:581–7. doi: 10.1038/sj.embor.embor862
45. Sarwar M, Shahzad MA, Farooq MK, Nisa M. Performance of growing lambs receiving altered plant protein sources with or without probiotics. In: International Conference on Asia Agriculture and Animal. Singapoore: IACSIT Press (2011). p. 139–144
46. Lahiri S, Kim H, Garcia-Perez I, Reza MM, Martin KA, Kundu P, et al. The gut microbiota influences skeletal muscle mass and function in mice. Sci Transl Med. (2019) 11: doi: 10.1126/scitranslmed.aan5662
47. Gloria-Trujillo A, Hernández-Sánchez D, Crosby-Galván MM, Hernández-Mendo O, Mata-Espinosa MÁ, Pinto-Ruiz R, et al. Performance and carcass characteristics of lambs fed diets supplemented with different levels of Saccharomyces cerevisiae. Revista Brasileira de Zootecnia. (2022) 51. doi: 10.37496/RBZ5120200281
48. Estrada-Angulo A, Zapata-Ramírez O, Castro-Pérez BI, Urías-Estrada JD, Gaxiola-Camacho S, Angulo-Montoya C, et al. The effects of single or combined supplementation of probiotics and prebiotics on growth performance, dietary energetics, carcass traits, and visceral mass in lambs finished under subtropical climate conditions. Biology (Basel). (2021) 10:1137. doi: 10.3390/biology10111137
49. Ladeira MM, Schoonmaker JP, Swanson KC, Duckett SK, Gionbelli MP, Rodrigues LM, et al. Review: Nutrigenomics of marbling and fatty acid profile in ruminant meat. Animal. (2018) 12:S282–94. doi: 10.1017/S1751731118001933
50. Hwang Y-H, Joo S-T. Fatty acid profiles, meat quality, and sensory palatability of grain-fed and grass-fed beef from Hanwoo, American, and Australian crossbred cattle. Korean J Food Sci Anim Resour. (2017) 37:153–61. doi: 10.5851/kosfa.2017.37.2.153
51. Ziaei N, Hosseini SMMS. Feeding value and in vitro digestibility of date-palm leaves supplemented with different supplementary energy. Pakistan J Biol Sci. (2009) 12:817–20.
52. Sharifi M, Bashtani M, Naserian AA, Farhangfar H. The effect of feeding low quality date palm (Phoenix dactylifera L) on the performance, antioxidant status and ruminal fermentation of mid-lactating Saanen dairy goats. Small Ruminant Res. (2015) 130:95–100. doi: 10.1016/j.smallrumres.2015.07.031
53. Maake TW, Adeleke M, Aiyegoro OA. Effect of lactic acid bacteria administered as feed supplement on the weight gain and ruminal pH in two South African goat breeds. Trans R Soc South Africa. (2021) 76:35–40. doi: 10.1080/0035919X.2020.1870018
54. Wang Y, Xia K, Wang XN, Lin X, Liu J, Li YJ, et al. Improvement of feed intake, digestibility, plasma metabolites, and lactation performance of dairy cows fed mixed silage of sugar beet pulp and rice straw inoculated with lactic acid bacteria. J Dairy Sci. (2022) 105:269–80. doi: 10.3168/jds.2021-20494
55. Sharma AN, Kumar S, Tyagi AK. Effects of mannan-oligosaccharides and Lactobacillus acidophilus supplementation on growth performance, nutrient utilization and faecal characteristics in Murrah buffalo calves. J Anim Physiol Anim Nutr (Berl). (2018) 102:679–89. doi: 10.1111/jpn.12878
56. Anee IJ, Alam S, Begum RA, Shahjahan RM, Khandaker AM. The role of probiotics on animal health and nutrition. J Basic Appl Zool. (2021) 82:52. doi: 10.1186/s41936-021-00250-x
57. Ghazanfar S, Anjum MI, Azim A, Ahmed I. Effects of dietary supplementation of yeast (Saccharomyces cerevisiae) culture on growth performance, blood parameters, nutrient digestibility and fecal flora of dairy heifers. J Anim Plant Sci. (2015) 25:53–9.
58. Kawauchi D, Angthong W, Keaokliang O, Ishida T, Takahashi T, Kawashima T. Effect of feeding Bacillus subtilis on rumen fermentation, blood metabolites, nutrient digestibility, and energy and nitrogen balances in non-lactating crossbred cows. Animal Sci J. (2021) 92:e13531. doi: 10.1111/asj.13531
59. Elizondo SJ. El nitrógeno en los sistemas ganaderos de leche. Agronomía Mesoamericana. (2006) 17:69–77.
60. Ørskov ERR, Ryle M. Energy nutrition in ruminants. Switzerland AG: Springer Netherlands (1990). 149 p.
61. Dewhurst RJ, Newbold JR. Effect of ammonia concentration on rumen microbial protein production in vitro. Br J Nutr. (2022) 127:847–9. doi: 10.1017/S000711452100458X
62. Tajima K, Aminov RI, Nagamine T, Matsui H, Nakamura M, Benno Y. Diet-dependent shifts in the bacterial population of the rumen revealed with real-time PCR. Appl Environ Microbiol. (2001) 67:2766–74.
63. Bach A, Giménez A, Juaristi JL, Ahedo J. Effects of physical form of a starter for dairy replacement calves on feed intake and performance. J Dairy Sci. (2007) 90:3028–33. doi: 10.3168/jds.2006-761
Keywords: blood metabolites, date palm leaves, digestibility, performance, wheat straw
Citation: Hamdon HA, Kassab AY, Vargas-Bello-Pérez E, Abdel Hafez GA, Sayed TA, Farghaly MM and Kholif AE (2022) Using probiotics to improve the utilization of chopped dried date palm leaves as a feed in diets of growing Farafra lambs. Front. Vet. Sci. 9:1048409. doi: 10.3389/fvets.2022.1048409
Received: 19 September 2022; Accepted: 11 October 2022;
Published: 26 October 2022.
Edited by:
Ilias Giannenas, Aristotle University of Thessaloniki, GreeceReviewed by:
Valiollah Palangi, Atatürk University, TurkeyTugay Ayasan, Osmaniye Korkut Ata University, Turkey
Alireza Seidavi, Islamic Azad University, Iran
Copyright © 2022 Hamdon, Kassab, Vargas-Bello-Pérez, Abdel Hafez, Sayed, Farghaly and Kholif. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Einar Vargas-Bello-Pérez, ZS52YXJnYXNiZWxsb3BlcmV6QHJlYWRpbmcuYWMudWs=; Ahmed E. Kholif, YWVfa2hvbGlmQGxpdmUuY29t