
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Vet. Sci. , 07 November 2022
Sec. Livestock Genomics
Volume 9 - 2022 | https://doi.org/10.3389/fvets.2022.1038218
 Laura Montoro-Dasi1
Laura Montoro-Dasi1 Laura Lorenzo-Rebenaque1
Laura Lorenzo-Rebenaque1 Adrian Ramon-Moragues2
Adrian Ramon-Moragues2 Maria Teresa Pérez-Gracia3
Maria Teresa Pérez-Gracia3 María de Toro4
María de Toro4 Clara Marin1*Arantxa Villagra2
Clara Marin1*Arantxa Villagra2Antimicrobial resistance is an important threat to public health worldwide, being one of the main death causes in 2050. Moreover, global health is currently underpinned by the “One Health” concept, whereby livestock is strictly related to human and environmental health. However, in the case of the meat rabbit industry, antibiotic additives are still added to prevent gastrointestinal diseases. Current food and consumer awareness require the implementation of sustainable production systems, where robustness and resilience are increasingly important. Hence, the aim of this study was to evaluate the effect of antibiotic feed supplementation on microbiota, and productive performance during the rabbit growing period in a robust genetic line. For this purpose, a total of 432 weaned rabbits were randomly housed, cecum samples were taken on the weaning day and at the end of the growing period (28 and 61 days of age, respectively), and 16S rRNA sequencing analysis was performed. Results showed a higher microbiota complexity at the end of growing in both experimental groups. Firmicutes represented the dominant phylum of the cecal community, followed by Bacteroidota in both groups. Moreover, Victivallis and Escherichia-Shigella genera were only identified in the experimental group without antibiotic supplementation at the end of the growing period. In conclusion, antibiotic feed supplementation had no effect on microbiota composition and productive performance in the robust genetic line reared. These results evidence the importance of the development of rabbit robust genetic lines as an alternative tool to antibiotic administration in epizootic enteropathy control.
Antimicrobial resistance (AMR) is an important threat to public health worldwide (1). In fact, 700,000 people die annually as a result of resistant bacteria around the world. The World Health Organization published that by 2050, AMR and consequent failed treatments will cause 10 million deaths and economic losses of $100 trillion annually (2). Moreover, global health is currently underpinned by the “One Health” concept, whereby livestock and agri-food systems are at the crossroads of human, animal and environmental health (3). In fact, a close association has been demonstrated between antibiotic (AB) use in animal production and the emergence of AMR in humans and environment, making it mandatory to reduce AB administration in farms (4, 5). However, in the case of the meat rabbit industry, although the use of AB as growth promoters has been banned in the European Union (EU) since 2006 (6), AB additives are still added to rabbit diets to prevent gastrointestinal disease (rabbit epizootic enteropathy) (7). In this sense, although humankind depends on agriculture and livestock for its food, more than 20% of current losses in animal production are still linked to animal diseases (8).
Moreover, as the world population is set to increase to 10 billion people by 2050, the animal protein required will be more than 70% of current yields (9). In this context, the rabbit has many interesting aspects for which it could be considered an ideal meat-producing animal: it has a short life cycle, a short gestation period and a high feed conversion ratio (10). This production sector is important in the EU, which is the world's second largest meat rabbit producer, accounting for 93% of the world's imports and exports, with Spain as one of the major exporting countries (11).
Metagenomic studies are therefore becoming increasingly relevant, due to the important role of microbiota balance in animal health, welfare and meat production (6). The gut microbiota is the complex microbial communities (bacteria, fungi, archaea, protozoa, and virus) that live together in a harmonic and dynamic equilibrium interacting with the host, playing an important role in metabolic, and immunologic functions (12). However, when this equilibrium is disturbed and beneficial bacteria cannot control detrimental bacteria, leads to in a dysbiosis status (13). Dysbiosis status has been related to pathologies and reduced production parameters. For that reason, many therapeutic strategies aimed at restoring the equilibrium of the intestinal microbiota (13). It is demonstrated that the presence of AB could affect the caecal microbial environment by modulating the microbiota composition and enhancing metabolic capacities by improving digestion and absorption of nutrients (7, 14, 15). Traditionally, antibiotics such as neomycin, tiamulin, valnemulin, chlortetracycline or bacitracin, have been used in rabbit production in sub-lethal doses to control pathogens and dysbiosis processes throughout the growing period (14, 16). Whitin them, neomycin and valnemulin are commonly used to prevent ERE, because it is demonstrated that increase rabbit's immune response, and modulate intestinal microbiota composition being specifically active against Clostridium perfringes, a pathogen strictly related to rabbit gastrointestinal disorders (17, 18). Nowadays, in rabbit production these molecules are still widely administered, especially after weaning, to control mortality peaks as a result of the onset of gastrointestinal symptoms (19). However, to be able to reduce AMR transmission throughout the food chain, new strategies should be developed to favor the establishing of a correct microbiota balance that increases the digestible efficacy of nutrients and maintains animal health and welfare (15).
Traditionally, strategies to meet protein demands were based on genetic selection focused on improving growth rate and muscle mass, and the intensification and automation of farm facilities (11, 20, 21). However, current food and consumer awareness require the implementation of sustainable production systems, respectful with animal welfare and efficiently facing environmental concerns (11). This objective is closely linked with genetic selection, where more rustic genetic lineages, which bear greater resilience against diseases, and therefore less AB requirement, will meet such concerns. Robustness is defined as the capacity to maintain adequate production levels, supporting all body functions at the highest performance, under different environmental conditions and production systems (22, 23). In accordance, the EFSA Panel on Animal Health and Welfare concluded that more emphasis should be placed on genetic selection traits such as disease resistance and stress resistance (11).
Nevertheless, to be able to assess the effectiveness of these alternatives it is necessary to have better knowledge of the development of microbiota composition with and without AB administration in broiler rabbits under animal production conditions (7). For this purpose, the cecum is commonly chosen to evaluate microbial composition and development, as it is the main site of fermentation and hosts the most diverse bacterial species of the gastrointestinal tract (7, 15, 19, 24).
Hence, the aim of this study was to evaluate the effect of AB feed supplementation on microbiota development (using 16S rRNA sequencing analysis) and productive performance during the rabbit growing period in a robust genetic line.
In this experiment, all animals were handled according to the principles of animal care published by Spanish Royal Decree 53/2013 (25). All protocols were approved by the Ethical Review Panel of the Directorate-General for Agriculture, Fisheries and Livestock of the Valencian Community under code 2018/VSC/PEA/0067.
The study was carried out in the experimental farm of the Polytechnic University of Valencia (UPV, in its Spanish acronym, Valencia, Spain). All the animals were provided from the same center (UPV), from a robust line (called LP) created and developed there (26), selected for its resilience and its ability to make use of available resources (22, 27).
A total of 432 weaned rabbits (28 days of age) were randomly housed in 12 collective cages (36 animals/cage) of 100 x 75 x 50 cm in size until the end of the growing period (61 days of age). Moreover, two management conditions were evaluated: animals fed with AB supplementation (ABs group, 216 rabbits located in 6 collective cages), and animals fed without AB supplementation (NoABs group, 216 rabbits located in 6 collective cages). The house was supplied with programmable electrical lights 12L:12D, automated electric heating and forced ventilation, in line with common practice in rabbit production. The experimental pelleted diets were commercial feed according to standard diets for rabbits, and the only difference between them was the presence of AB (Table 1). Nutritional and product analysis was assessed before the arrival of animals. Feed was weighed, manually distributed and added ad libitum. Finally, the mortality and the presence of diarrhea were recorded daily.

Table 1. Analytical feed dry matter composition (%).
In this experiment, two different sampling times were established: the weaning day (28 days of age) and the end of the growing period (61 days of age). At each sampling time, 4 animals per cage of each experimental group (n = 24 samples/ experimental group/ sampling time) were randomly selected and cecal samples were collected. Ceca were taken individually, placed in sterile jars and processed within 24 h after collection.
In first place, cecal content was removed and homogenized. Then, pools of four animals from the same cage were prepared (n = 6 pools/ experimental group/ sampling time); the DNA of pools content was extracted (QIAamp Power Fecal DNA kit, Werfen, Barcelona, Spain) and frozen at −80°C for shipment to the Center for Biomedical Research of La Rioja (CIBIR, in its Spanish acronym, Logroño, Spain), according to the manufacturer's instructions.
Once there, 16S rRNA gene amplification and MiSeq sequencing was performed according to Montoro-Dasi et al. (28) in the Centre for Biomedical Research of La Rioja (CIBIR, in its Spanish acronym, Logroño, Spain).
Briefly, primer sequences cover the V3–V4 regions of the 16S rRNA gene, and the following primers included the Illumina adapters: 16S Amplicon PCR Forward Primer = 50 (TCGTCGGCAGCGTCAGATGTGTATAAGAGACAGCCTACGGGNGGCWGCAG) and 16S Amplicon PCRReverse Primer = 50 (GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAGGACTACHVGGGTATCTAATCC). Finally, the sequencing run was performed in a MiSeq (Illumina) system in 2 x 300 bp format. After evaluating the quality of the raw unprocessed reads, the adapters were removed, and the reads were re-evaluated. Finally, the V3–V4 region of the 16S rRNA gene was partially reconstructed into fragments of approximately 550–580 bp. The OTU (Operational Taxonomic Unit) picking, and analysis was performed with QIIME (v1.9.1) pipeline.
To record performance data, animals and feed consumption were weighed per cage at weekly intervals. Thus, mean daily feed intake (ADFI), mean daily gain (ADG) and feed conversion ratio (FCR) were evaluated each week during the growing period.
First, to evaluate performance parameters for all the observed variables, a descriptive analysis of each sample was carried out to detect out-of-range data (outliers), proceeding to eliminate those records that the program indicated as such. Then, a Kolmogorov-Smirnov test was performed to ensure data followed normal distribution. Finally, a multi-factor two ways Analysis of Variance (ANOVA) test was used to compare the performance results obtained for both experimental groups. Statistical Analysis was performed using the Statgraphics XVII Centurión® program.
Furthermore, to perform the statistical analysis of bioinformatics results, demultiplexed paired FASTQ sequences were imported into the QIIME2 v2021.4. The DADA2 pipeline incorporated into QIIME2 was used for the denoising, filtering and chimera removal of the sequences and assigned reads into Amplicon Sequence Variants (ASVs). Then, taxonomic annotation was obtained using the SILVA v138 database (29) and sequences not assigned to any taxa or classified as Eukaryote or Archaea were filtered out. Sequencing statistical analyses were done using QIIME2 v2021.4. Moreover, to compare diversity and richness between cecal communities, Chao1, Shannon, and observed number of OTUs indexes were computed after OTUs rarefication at 17 658 contigs. The statistical method used for comparison of the communities was a paired samples analysis of variance that included the following factors: sampling time (weaning day vs. end of the growing period), AB treatment (ABs vs. NoABs). The significance of differences among different groups was evaluated by Kruskal–Wallis test. Box-and-whisker plots for species richness and evenness were generated using Graphad Prims 8. Finally, a Venn diagram was drawn up to show the shared and unique features among groups, based on the occurrence of features in a sample group regardless of their relative abundance, by using InteractiVenn software for Venn diagram construction (30). Finally, to quantitatively measure beta diversities, the Bray-Curtis distance and unweighted Unifrac and weighted Unifrac values were calculated, and Principle Coordinate Analysis (PCoA) plots were generated for the origin of the sample from Bray-Curtis distances using ClustVist software (31). Differences in microbial mean taxa abundance according to group were detected using ANCOM (Analysis of Composition of Microbiomes), with W value corresponding to the number of times an ASV abundance is significantly different for a group (32).
A total of 24 cecal pools were collected and processed: six cecal pools for each experimental group (ABs and NoABs) at the weaning day, and six cecal pools for each experimental group (ABs and NoABs) at the end of the growing period.
The total of sequencing reads of the 24 cecal pools samples was 5 409 112 (mean 225 379.7 reads/sample), with a mean read length of 444.3 ± 13.88 pb. A total of 524,841 sequences and 5 091 ASVs were generated. A total of 6,001 chimeric sequences were removed from the dataset with a total of 518,840 sequences left for ASVs table generation and database alignment. After filtering, a total of 4,956 unique sequences were left for taxonomic assignment. The datasets generated and analyzed are available at NCBI's BioProject PRJNA612272 and BioSample SAMN30645692.
The diversity study was performed after rarefying 17,658 reads, with the samples from the NoABs group at weaning day as the limit of the rarefaction. Diversity analysis for the different sampling times revealed no significant differences in richness (Chao 1 index), or other alpha-diversity indices (Shannon and Observed OTUs) between animals on ABs and NoABs treatment at the different sampling times (p-value > 0.05, Supplementary Figure 1). However, the alpha-diversity indexes reveal a notable difference between the cecal microbiota diversity depending on the time of sampling (weaning day vs. end of the growing period) (p-value < 0.05, Supplementary Figure 1).
Beta-diversity was measured by PERMANOVA test using Bray-Curtis dissimilarity and revealed a similar pattern to that of alpha-diversity, with no significant differences between the animals from ABs and NoABs groups (p-value > 0.05, Supplementary Figure 2). Results of the PERMANOVA indicated that there were significant differences in the microbial community composition in relation to sampling time (weaning day vs. end of the growing period) (p-value < 0.05, Supplementary Figure 2).
To better understand how the microbial community composition changes between the different sampling times (weaning day vs. end of the growing period) and the management conditions (ABs vs. NoABs), we examined which organisms were present at different taxonomic levels and their relative abundance. Alignment of ASVs against the SILVA database resulted in identification of 9 bacterial phyla and 134 bacterial genera. While the majority of OTUs were identified at the genus level, some were only classified at the phylum, class, order or family.
At phylum level, Firmicutes represented the dominant phylum of the cecal community, followed by Bacteroidota in both groups at the two sampling times (Table 2; Figure 1).
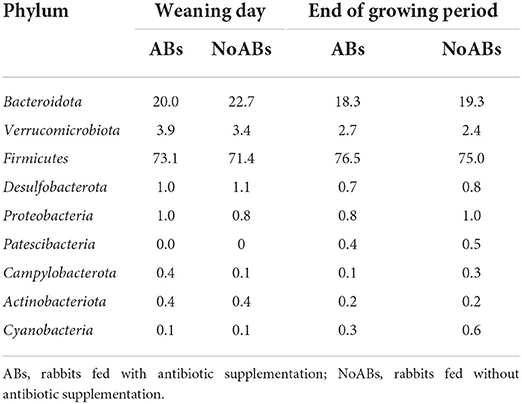
Table 2. Relative abundance (%) of the taxonomic profiles in caecal samples at phylum level according to management conditions (ABs vs. NoABs) and sampling time (weaning day vs. end of the growing period).
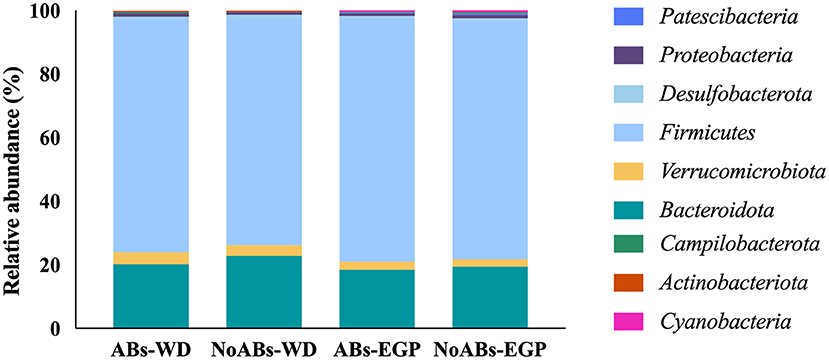
Figure 1. Taxonomic analysis at phylum level according to management conditions (ABs vs. NoABs) and sampling time (weaning day vs. end of the growing period). ABs-WD, animals fed with AB supplementation at weaning day; NoABs-WD, animals fed without AB supplementation at weaning day; ABs-EGP, animals fed with AB supplementation at the end of the growing period; NoABs-EGP, animals fed without AB supplementation at the end of the growing period.
Among the 134 genera detected, 95 and 88 genera were detected in ABs group for the samples of the weaning day and the end of the growing period, respectively (Figure 2). Meanwhile, 101 and 97 genera were detected in NoABs group for the samples of the weaning day and the end of the growing period, respectively (Figure 2). Moreover, 78 genera were shared by groups (Figure 2). Finally, identified genera present only in NoABs group at the end of the growing period were Victivallis (0.01% of relative abundance), and Escherichia-Shigella (0.002% of relative abundance).
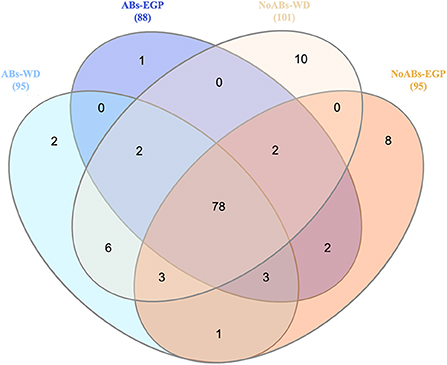
Figure 2. Venn diagram showing unique and shared taxa at genus level between samples according to antibiotic treatment (ABs vs. NoABs) and sampling time (weaning day vs. end of the growing period). Selection criteria were based on presence or absence regardless of abundance. Not drawn to scale. Value in brackets represent the genera found in each origin. Abs, rabbits fed with antibiotic supplementation; NoABs, rabbits fed without antibiotic supplementation; WD, weaning day; EGP, end of the growing period.
The 45 genera with a relative abundance of more than 0.5% in at least one sample group are presented in Supplementary Table 1 and Figure 3 (28, 33). In weaning day rabbits, for both experimental groups the most prevalent genera were unclassified members (U.m.) of Lachnospiraceae family (14.6 and 14.2%, respectively), Clostridia_vadinBB60_group spp. (10.7 and 9.8%, respectively), Bacteroides spp. (6.3 and 8.7%, respectively), Clostridia_UCG-014 spp. (6.3 and 8.7%, respectively), Muribaculaceae spp. (5.0 and 4.5%, respectively) and Ruminococcus spp. (4.5 and 5.1%, respectively). Finally, at the end of the growing period, for both experimental groups the most common genera were Muribaculaceae spp. (12.4 and 14.1%, respectively), Clostridia_UCG-014 spp. (11.6 and 11.4%, respectively), U.m. of Lachnospiraceae family (10.5 and 10.0%, respectively), Clostridia_vadinBB60_group spp. (19.2 and 9.7%, respectively), Ruminococcus spp. (5.2 and 4.0%, respectively) and U.m. of Eubacteriaceae family (4.4 and 5.0%, respectively).
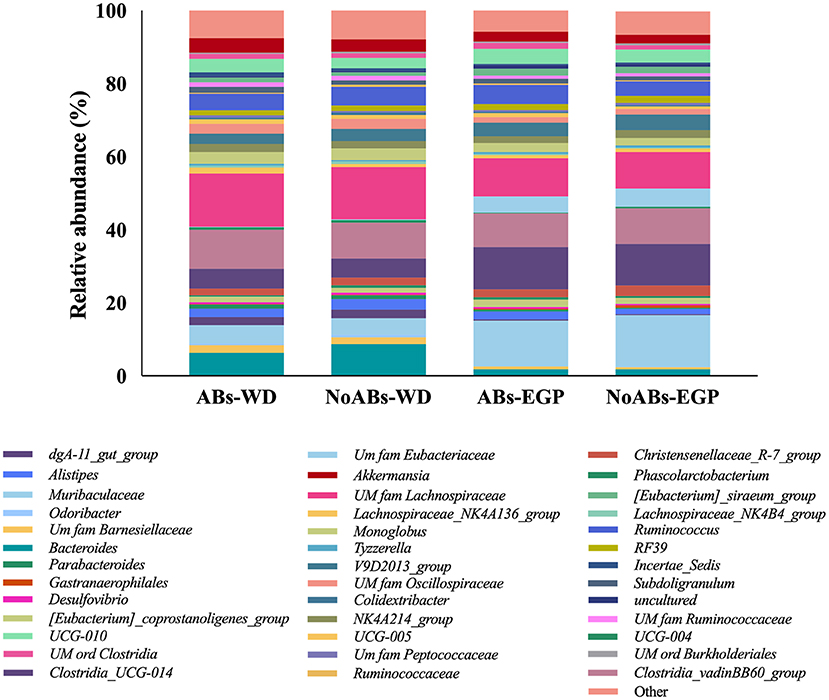
Figure 3. Taxonomic analysis at genus level according to management conditions (ABs vs. NoABs) and sampling time (weaning day vs. end of the growing period). ABs-WD, animals fed with AB supplementation at weaning day; NoABs-WD, animals fed without AB supplementation at weaning day; ABs-EGP, animals fed with AB supplementation at the end of the growing period; NoABs-EGP, animals fed without AB supplementation at the end of the growing period.
ANCOM was performed between samples according to antibiotic treatment (ABs vs. NoABs) and sampling time (weaning day vs. end of the growing period). Weaning day had three differentially abundant taxa against end of the growing period: Clostridia_UCG-014 spp. (W = 4,292), Muribaculaceae spp. (W = 4,143), and U.m. of Eubacteriaceae family (W = 4,048). In ABs and NoABs in sampling time did not yield differentially abundant taxa.
The productive parameters obtained were in accordance with the breed standards, without significant differences for any of the productive parameters measured between management systems (ABs vs. NoABs) (p-value > 0.05).
Regarding health status of the rabbits and diarrhea presence, significant differences were observed during the second week of the growing period, when NoABs group showed 6.20% of animals with diarrhea, while the ABs group only presented 1.05% (Table 3).
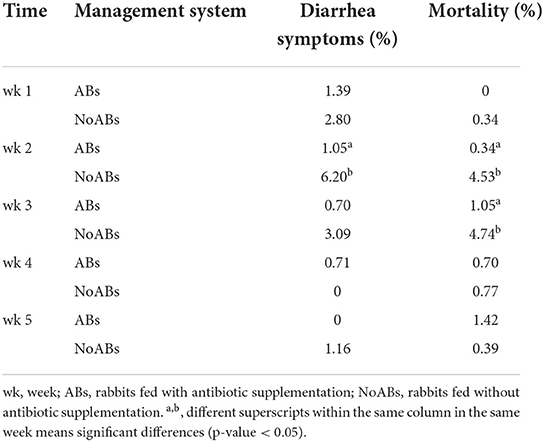
Table 3. Presence of diarrhea symptoms and mortality rates during the growing period for both experimental groups.
Finally, mortality rates showed significant differences between groups (p-value < 0.05) at the second (ABs group: 0.34%, and NoABs group: 4.53%) and third (ABs group: 1.05%, and NoABs group: 4.74%) weeks of growth (p-value < 0.05) (Table 3).
The present study assessed the cecal microbiota development and performance parameters in two different rabbit management systems during the growing period: animals fed with AB supplementation, and animals fed without AB supplementation. As reported above, although the increase in multidrug-resistant zoonotic pathogens in the food chain is one of the main concerns of public health, AB are still added in rabbit diets to prevent epizootic enteropathy (6). However, society and European health authorities are forcing the agri-food sector to find cost-effective, animal and environment friendly alternatives to AB.
In this sense, knowing the development of rabbit microbiota composition from weaning to the end of the growing period and how management practices, such as AB supplementation, influence its modulation could help in decision-making at farm level, and to evaluate the efficacy of alternatives proposed, such as robust genetic lines. For that reason, it might be interesting to consider microbiota composition as a biomarker of rabbit health and productive performance (15).
It is well demonstrated that a greater complexity of the cecal microbiota is observed as animals grow (34). Our findings showed that there is an important change in microbiota diversity from the weaning day to the end of the growing period, regardless of the administration of antibiotics at sub-therapeutic doses during the growing period, in agreement with previous studies (16, 35). Previous authors reported that microbial diversity is positively related to gut health (15, 16). In this sense, the results of this study showed that the withdrawal of ABs from the diet has no negative implications for intestinal health.
Regarding microbiota composition, Firmicutes and Bacteroidota were the predominant phyla for all groups, according to previous studies (15). Firmicutes has a fundamental role in rabbits' digestion, as it was considered the most efficient cellulose degrader (15, 36). Thus, the fact that no differences were found in this phylum between groups may mean that the absence of ABs in these animals would not affect the intestinal function of rabbits. Within this phylum, Ruminococcus spp. is the most relevant genus, being dominant in healthy rabbits and decreasing in the presence of disease (15, 37). Members of the Lachnospiraceae family are also significantly abundant in the microbiota of rabbits, as concurred with the findings of previous authors (35, 38, 39). High prevalence of this group has been observed in healthy young rabbits, related to the stimulation of cecotrophic behavior, which was also associated with a reduction in mortality (40). Moreover, Bacteroidetes was one of the major commensal phyla in the gut microbiota of rabbits, without statistical differences between groups. This phylum is proven to stimulate the development of gut-associated immune tissue (15, 36, 41). Indeed, members of this phylum have been related to degradation of vegetal polysaccharides and amino acid fermentation (16). Specifically, the presence of Muribaculaceae, the most relative abundant genus, has an important impact on host development and health (42). In this study, the removal of ABs in the feed had no impact on their abundance, maintaining a correct balance in cecal microbiota for both groups. On the other hand, the family Clostridiaceae plays an important role, being responsible for cellulose degradation, although the presence of some species in the cecum could lead in a decrease in butyrate yield, which has been related to rabbit epizootic enteropathy (16, 43). Although antibiotics have been used to control these bacteria (35), no differences were observed between groups.
However, at the end of the growing period it is important to highlight that Victivallis and Escherichia-Shigella genera, although in low relative abundances, were only present in the NoABs group, and both are associated with rabbit epizootic enteropathy (44, 45). For that reason, they could be considered as biomarkers of this disease, and alterative tools to AB should be developed and evaluated against these genera.
Regarding productive parameters, large group sizes during the growing period have been associated with negative effects on the mean daily gain, the mean daily intake, and the final body weight (46). Conversely, AB used as growth promoters have been related to better productive profitability (47). However, in this study, probably due to the use of a robust genetic line (LP), with greater resilience and better management of available resources (23), productive performance was according to the breed standards in both experimental groups. Thus, its development should be considered as an interesting alternative tool to reduce AB administration in rabbit production.
Finally, regarding health status of the rabbits and diarrhea presence, significant differences were observed during the second week of the growing period, when the NoABs group showed a higher percentage of diarrhea and mortality rate, probably associated with the effects of rabbit epizootic enteropathy (48, 49). This fact could be related to pathological disorders without a microbiota impact (50). However, it would be necessary to obtain global approaches to microbiability through the omic sciences (metagenomics, metabolomics, transcriptomics, genomics and epigenomics, etc.) throughout the growing period.
In conclusion, AB feed supplementation had no effect on microbiota diversity and phyla composition when a robust genetic line LP was reared. However, some differences appeared at genera level when antibiotics has been removed, probably related to rabbit epizootic enteropathy. For that reason, further studies are needed to pinpoint the specific causes of this disease and be able to develop complementary, effective, sustainable and animal-friendly alternatives applicable during the rabbit growing period to avoid antibiotic use on rabbit farms.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: https://www.ncbi.nlm.nih.gov/, SAMN30645692.
The animal study was reviewed and approved by Ethical Review Panel of the Directorate-General for Agriculture, Fisheries and Livestock of the Valencian Community under code 2018/VSC/PEA/0067.
Data curation: LM-D, LL-R, AR-M, MP-G, CM, and AV. Formal analysis: LM-D, LL-R, and MT. Funding acquisition: AV. Investigation: LM-D and CM. Methodology: AV and CM. Writing–original draft: LM-D, LL-R, and CM. Writing–review and editing: MP-G, CM, and AV. All authors contributed to the article and approved the submitted version.
This research was funded by the National Institute for Agricultural Research and Experimentation and the Ministry of Economy, Industry and Competitiveness (RTA 2017-00013, Programme: MINECO, Ministerio de Economía y Competitividad). LL-R was supported by a research grant from the Generalitat Valenciana-Fondo SocialEuropeo (ACIF/2020/376).
The authors wish to thank the Improvement of Production System-related Food Safety and End Products research group (Veterinary Faculty, University CEU-Cardenal Herrera) for their technical support. English text version was revised by N. Macowan English Language Service.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fvets.2022.1038218/full#supplementary-material
1. WHO (World Health Organization). Critically Important Antimicrobials for Human Medicine, 6th revision. World Health Organization (2019). Available online at: https://www.who.int/publications/i/item/9789241515528 (accessed September 5, 2020).
2. WHO (World Health Organization). New Report Calls for Urgent Action to Avert Antimicrobial Resistance Crisis. (2019). Available online at: https://www.who.int/news/item/29-04-2019-new-report-calls-for-urgent-action-to-avert-antimicrobial-resistance-crisis (accessed February 15, 2022).
3. Mackenzie JS, Jeggo M. The one health approach-why is it so important? Trop Med Infect Dis. (2019) 4:88. doi: 10.3390/tropicalmed4020088
4. Sharma C, Rokana N, Chandra M, Singh BP, Gulhane RD, Gill JPS, et al. Antimicrobial resistance: its surveillance, impact, and alternative management strategies in dairy animals. Front Vet Sci. (2018) 4:237. doi: 10.3389/fvets.2017.00237
5. CDC (Centers for Disease Control Prevention) NCEZID (National Center for Emerging Zoonotic Infectious Diseases) and DHQP (Division of Healthcare Quality Promotion). About Antibiotic Resistance. (2020). Available online at: https://www.cdc.gov/drugresistance/about.html (accessed February 7, 2021).
6. EC (European Commission),. Regulation (EC) No 1831/2003 of the European Parliament of the Council of 22 September 2003 on additives for use in animal nutrition (Text with EEA relevance). (2003). Available online at: https://eur-lex.europa.eu/legal-content/EN/ALL/?uri=celex%3A32003R1831 (accessed February 24, 2021).
7. Zou F, Zeng D, Wen B, Sun H, Zhou Y, Yang M, et al. Illumina Miseq platform analysis caecum bacterial communities of rex rabbits fed with different antibiotics. AMB Express. (2016) 6:1–11. doi: 10.1186/s13568-016-0273-1
8. Van Boeckel TP, Brower C, Gilbert M, Grenfell BT, Levin SA, Robinson TP, et al. Global trends in antimicrobial use in food animals. Proc Natl Acad Sci U S A. (2015) 112:5649–54. doi: 10.1073/pnas.1503141112
9. FAO (Food and Agriculture Organization of the United Nations). OECD-FAO Agricultural Outlook 2021-2030. Paris (2021).
10. Cullere M, Dalle Zotte A. Rabbit meat production and consumption: state of knowledge and future perspectives. Meat Sci. (2018) 143:137–46. doi: 10.1016/j.meatsci.2018.04.029
11. EFSA (European Food Safety Authority). Health and welfare of rabbits farmed in different production systems. EFSA J. (2020) 18:5944. doi: 10.2903/j.efsa.2020.5944
12. Berg GD, Rybakova D, Fischer T, Cernava MCC, Vergès T, Charles X, et al. Microbiome definition re-visited: old concepts and new challenges. Microbiome. (2020) 8:103. doi: 10.1186/s40168-020-00875-0
13. Gagliardi A, Totino V, Cacciotti F, Iebba V, Neroni B, Bonfiglio G, et al. Rebuilding the gut microbiota ecosystem. Int J Environ Res Public Health. (2018) 15:1679. doi: 10.3390/ijerph15081679
14. Abecia L, Fondevila M, Balcells J, Lobley GE, McEwan NR. The effect of medicated diets and level of feeding on caecal microbiota of lactating rabbit does. J Appl Microbiol. (2006) 103:787–93. doi: 10.1111/j.1365-2672.2007.03309.x
15. Cotozzolo E, Cremonesi P, Curone G, Menchetti L, Riva F, Biscarini F, et al. Characterization of bacterial microbiota composition along the gastrointestinal tract in rabbits. Animals. (2020) 11:31. doi: 10.3390/ani11010031
16. Velasco-Galilea M, Guivernau M, Piles M, Viñas M, Rafel O, Sánchez A, et al. Breeding farm, level of feeding and presence of antibiotics in the feed influence rabbit cecal microbiota. Anim Microbiome. (2020) 2:1–16. doi: 10.1186/s42523-020-00059-z
17. Djukovic A, Garcia-Garcera M, Martínez-Paredes E, Isaac S, Artacho A, Martínez J, et al. Gut colonization by a novel clostridium species is associated with the onset of epizootic rabbit enteropathy. Vet Res. (2018) 49:123. doi: 10.1186/s13567-018-0617-8
18. Aggag SA, Abdelkader M-AE, Yacout MM. Neomycin's immunomodulatory effect on the gene expression, some hematologic parameters, and intestinal histology in two rabbit lines. Animal Gene. (2022) 25:200130. doi: 10.1016/j.angen.2022.200130
19. Velasco-Galilea M, Guivernau M, Piles M, Viñas M, Rafel O, Sánchez A, et al. Housing Conditions, Level of Feeding and Presence of Antibiotics in The Feed Shape Rabbit Cecal Microbiota. (2020). doi: 10.21203/rs.3.rs-38388/v1
20. FAO (Food Agriculture Organization of the United Nations). Meat Market Review. (2020). Available online at: http://www.fao.org/3/ca3880en/ca3880en.pdf
21. MAPA (Ministerio de Agricultura, Pesca y, Alimentación),. Dirección General De Producciones Y Mercados Agrarios. Minist. Agric. Pesca y Aliment. (2020). p. 77. Available online at: http://publicacionesoficiales.boe.es/
22. Savietto D, Cervera C, Blas E, Baselga M, Larsen T, Friggens NC, et al. Environmental sensitivity differs between rabbit lines selected for reproductive intensity and longevity. Animal. (2013) 7:1969–77. doi: 10.1017/S175173111300178X
23. Savietto D, Friggens NC, Pascual JJ. Reproductive robustness differs between generalist and specialist maternal rabbit lines: the role of acquisition and allocation of resources. Genet Sel Evol. (2015) 47:1–12. doi: 10.1186/s12711-014-0073-5
24. Velasco-Galilea M, Piles M, Viñas M, Rafel O, González-Rodríguez O, Guivernau M, et al. Rabbit microbiota changes throughout the intestinal tract. Front Microbiol. (2018) 9:1–14. doi: 10.3389/fmicb.2018.02144
25. MAPA (Ministerio de Agricultura, Pesca y Alimentación). Real Decreto 53/2013, de 1 de febrero, por el que se establecen las normas básicas aplicables para la protección de los animales utilizados en experimentación y otros fines científicos, incluyendo la docencia (2013).
26. Marco-Jiménez F, Baselga M, Vicente JS. Successful re-establishment of a rabbit population from embryos vitrified 15 years ago: the importance of biobanks in livestock conservation. PLoS ONE. (2018) 13:e0199234. doi: 10.1371/journal.pone.0199234
27. El Nagar AG, Sánchez JP, Ragab M, Mínguez C, Baselga M. Genetic variability of functional longevity in five rabbit lines. Animal. (2020) 14:1111–9. doi: 10.1017/S1751731119003434
28. Montoro-Dasi L, Villagra A, de Toro M, Pérez-Gracia MT, Vega S, Marin C. Fast and slow-growing management systems: characterisation of broiler caecal microbiota development throughout the growing period. Animals. (2020) 10:1401. doi: 10.3390/ani10081401
29. Quast C, Pruesse E, Yilmaz P, Gerken J, Schweer T, Yarza P, et al. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. (2013) 41:590–6. doi: 10.1093/nar/gks1219
30. Heberle H, Meirelles VG, da Silva FR, Telles GP, Minghim R. InteractiVenn: a web-based tool for the analysis of sets through Venn diagrams. BMC Bioinform. (2015) 16. doi: 10.1186/s12859-015-0611-3
31. Metsalu T, Vilo J. ClustVis: a web tool for visualizing clustering of multivariate data using principal component analysis and heatmap. Nucleic Acids Res. (2015) 43:W566–70. doi: 10.1093/nar/gkv468
32. Dhariwal A, Chong J, Habib S, King IL, Agellon LB, Xia J. Microbiome analyst: a web-based tool for comprehensive statistical, visual and meta-analysis of microbiome data. Nucleic Acids Res. (2017) 45:W180–8. doi: 10.1093/nar/gkx295
33. Montoro-Dasi L, Villagra A, de Toro M, Pérez-Gracia MT, Vega S, Marin C. Assessment of microbiota modulation in poultry to combat infectious diseases. Animals. (2021) 11:1–9. doi: 10.3390/ani11030615
34. Puón-peláez XHD, McEwan NR, Gómez-soto JG, Álvarez-martínez RC, Olvera-ramírez AM. Metataxonomic and histopathological study of rabbit epizootic enteropathy in Mexico. Anim. (2020) 10:936. doi: 10.3390/ani10060936
35. Bäuerl C, Collado MC, Zúñiga M, Blas E, Martínez GP. Changes in cecal microbiota and mucosal gene expression revealed new aspects of epizootic rabbit enteropathy. PLoS ONE. (2014) 9:e105707. doi: 10.1371/journal.pone.0105707
36. Abecia L, Rodríguez-Romero N, Yañez-Ruiz DR, Fondevila M. Biodiversity and fermentative activity of caecal microbial communities in wild and farm rabbits from Spain. Anaerobe. (2012) 18:344–9. doi: 10.1016/j.anaerobe.2012.04.004
37. Combes S, Gidenne T, Cauquil L, Bouchez O, Fortun-Lamothe L. Coprophagous behavior of rabbit pups affects implantation of cecal microbiota and health status. J Anim Sci. (2014) 92:652–65. doi: 10.2527/jas.2013-6394
38. Massip K, Combes S, Cauquil L, Zemb O, Gidenne T. High throughput 16S-DNA sequencing for phylogenetic affiliation of the caecal bacterial community in the rabbit - Impact of the hygiene of housing and of the intake level. In: Proceeding of Symposium on Gut Microbiology. Clermont-Ferrand (2012).
39. Zhu Y, Wang C, Li F. Impact of dietary fiber/starch ratio in shaping caecal microbiota in rabbits. Can J Microbiol. (2015) 61:771–84. doi: 10.1139/cjm-2015-0201
40. Mattioli S, Dal Bosco A, Combes S, Moscati L, Crotti S, Mancinelli AC, et al. Dehydrated alfalfa and fresh grass supply in young rabbits: effect on performance and caecal microbiota diversity. Animals. (2019) 9:341. doi: 10.3390/ani9060341
41. Monteils V, Cauquil L, Combes S, Godon JJ, Gidenne T. Potential core species and satellite species in the bacterial community within the rabbit caecum. FEMS Microbiol Ecol. (2008) 66:620–9. doi: 10.1111/j.1574-6941.2008.00611.x
42. Li Z, He H, Ni M, Wang Z, Guo C, Niu Y, et al. Microbiome-Metabolome Analyze The Immune Microenvironment of Cecal Contents, Soft and Hard Feces of Hyplus Rabbits. Research Square Preprints (2021). doi: 10.21203/rs.3.rs-1047430/v1
43. Jin DX, Zou HW, Liu SQ, Wang LZ, Xue B, Wu D, et al. The underlying microbial mechanism of epizootic rabbit enteropathy triggered by a low fiber diet. Sci Rep. (2018) 8:12489. doi: 10.1038/s41598-018-30178-2
44. Shanmuganandam S, Hu Y, Strive T. Uncovering the microbiome of invasive sympatric European brown hares and European rabbits in Australia. PeerJ. (2020) 8:1–21. doi: 10.7717/peerj.9564
45. Shen XM, Cui HX, Xu XR. Orally administered Lactobacillus casei exhibited several probiotic properties in artificially suckling rabbits. Asian-Australas J Anim Sci. (2020) 33:1352–9. doi: 10.5713/ajas.18.0973
46. Szendro Z, Dalle Zotte A. Effect of housing conditions on production and behaviour of growing meat rabbits: a review. Livest Sci. (2011) 137:296–303. doi: 10.1016/j.livsci.2010.11.012
47. Falcão-e-Cunha L, Solla LC, Maertens L, Marounek M, Pinheiro V, Freire J, et al. Alternatives to antibiotic growth promoters in rabbit feeding: a review. World Rabbit Sci. (2007) 15:127–40. doi: 10.4995/wrs.2007.597
48. Licois D, Wyers M, Coudert P. Epizootic rabbit enteropathy: experimental transmission and clinical characterization. Vet Res. (2005) 36:601–13. doi: 10.1051/vetres:2005021
49. Allen HK, Levine UY, Looft T, Bandrick M, Casey TA. Treatment, promotion, commotion: antibiotic alternatives in food-producing animals. Trends Microbiol. (2013) 21:114–9. doi: 10.1016/j.tim.2012.11.001
Keywords: rabbit production, antibiotic supplementation, farm management, microbiota, 16S rRNA analysis, livestock
Citation: Montoro-Dasi L, Lorenzo-Rebenaque L, Ramon-Moragues A, Pérez-Gracia MT, de Toro M, Marin C and Villagra A (2022) Antibiotic removal does not affect cecal microbiota balance and productive parameters in LP robust rabbit line. Front. Vet. Sci. 9:1038218. doi: 10.3389/fvets.2022.1038218
Received: 06 September 2022; Accepted: 21 October 2022;
Published: 07 November 2022.
Edited by:
Shailendra Kumar Mishra, National Institute for Research in Reproductive Health (ICMR), IndiaReviewed by:
Irida Palamidi, Agricultural University of Athens, GreeceCopyright © 2022 Montoro-Dasi, Lorenzo-Rebenaque, Ramon-Moragues, Pérez-Gracia, de Toro, Marin and Villagra. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Clara Marin, Y2xhcmEubWFyaW5AdWNoY2V1LmVz
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.