
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Vet. Sci. , 18 January 2022
Sec. Parasitology
Volume 8 - 2021 | https://doi.org/10.3389/fvets.2021.798942
This article is part of the Research Topic Antiparasitic Activity and the Modes of Action of Natural Products and Traditional Medicines View all 10 articles
 Hornblenda Joaquina Silva Bello1,2*
Hornblenda Joaquina Silva Bello1,2* José Gabriel Gonçalves Lins1,2
José Gabriel Gonçalves Lins1,2 Ana Cláudia Alexandre de Albuquerque2Gabriel Badial Ferreira1Mônica Regina Vendrame Amarante2Alessandro Francisco Talamini do Amarante2
Ana Cláudia Alexandre de Albuquerque2Gabriel Badial Ferreira1Mônica Regina Vendrame Amarante2Alessandro Francisco Talamini do Amarante2The sheep nasal bots Oestrus ovis is parasite of the nasal cavities and sinuses of small ruminants causing oestrosis, one of the most frequent parasitic diseases in sheep and goats. The widely use of ivermectin and closantel by the sheep breeders in the treatment and prophylaxis of gastrointestinal nematodes resulted in widespread cases of anthelmintic resistance. However, there is no report about cases of O. ovis with drug-resistance. In this study, we evaluated the prophylactics and therapeutic effects of both antiparasitics in sheep with O. ovis natural infestation. The trial was carried out from early December 2019 to March 2020, with 30 crossbred males lambs allocated into three groups of 10 animals each: control (without treatment), treated with ivermectin (0.2 mg/kg subcutaneously) and treated with closantel (10 mg/kg orally). The animals were kept together grazing the same pasture area. The treatment groups were drenched in two occasions 70 days apart: on 5th December 2019 and on 13th February 2020. On 19th March 2020, all lambs were slaughtered. The lamb heads were removed and sectioned along their longitudinal and sagittal axis to search for larvae. Recovered O. ovis larvae were counted and identified according to their developmental stage (L1, L2, and L3). Seven of the control lambs were infested with O. ovis larvae ranging from six to 17 larvae (11.6 mean infestation intensity). All recovered larvae from control group were intact and active. Three animals treated with ivermectin had O. ovis larvae (1–3 larvae), however they were dead and in degeneration. The animals treated with closantel did not have any larvae. The clinical suggestive signs of oestrosis were scarce over the experimental period. The averages of daily weight gain were similar (p > 0.05) among groups. Closantel and ivermectin had high efficacy against oestrosis and O. ovis parasitism did not hinder the performance of lambs.
Oestrus ovis L. (Diptera: Oestridae) is a cosmopolitan parasite with a higher prevalence in tropical regions. It causes cavitary myiasis in small ruminants, once the larvae are an obligate parasite of the nasal and sinus cavities of sheep and goats (1).
In endemic areas, the O. ovis larvae have been found in humans' eyes and nasopharyngeal airway. Cases of ocular affections are likely to occur more frequently than currently notified, because many cases without complications are unreported (2).
The female fly flies around the head of its host to deposits larvae at a few centimeters from sheep nostrils (3). In case of the fly presence, the animals hide the muzzle on the soil or in the others sheep's wool, swing head and sneezing, which leads to distress. The larvae hooks and spine irritate the nasal mucosal, provoking inflammation and mucous nasal discharge. At the same time, this irritating action secures the production of an inflammatory exudate, which count for larva's feeding (4). Besides frequent sneezes, sheep infested with O. ovis larvae may also present difficulty in breathing, hyporexia, and weight loss. In heavy infestation, animals may have related secondary bacterial infection in the lungs (5).
In a trial conducted in Brazil for three consecutive years, the prevalence of O. ovis was 50% with the occurrence of the parasite in all seasons. Nevertheless, the highest prevalence (61.1%) were observed in the spring and summer. The climatic conditions of this region during this period are optimal for fly activity, and consequently for a high rate of sheep infestation (6).
The oestrosis therapy is based on the use of antiparasitic drugs that target the larvae. Many systemic antiparasitic drugs may be used for the oestrosis treatment. The most widely used antiparasitics to control O. ovis are closantel and the macrocyclic lactones, such as orally or injectable ivermectin (7, 8). Sheep treated with closantel, injectable and oral ivermectin had 100, 100, and 98% of efficacy, respectively considering all instars (7). In a survey conducted by Oliveira et al. (8) the injectable doramectin was 100% effective against all larval stages. Nevertheless, the first instar larvae (L1) is considered less susceptible to these drugs in comparison with the second instar larvae (L2) and third instar larvae (L3) (9) because the L1 larvae feed less thus they are less prone to ingest the systemic parasiticides (10). Martínez-Valladares et al. (11) observed that the long-acting injectable moxidectin had 90.2% of efficacy against L1.
The intensification of sheep farming and emergence of drug-resistant parasites have brought new challenges regarding the prophylaxis and treatment of small ruminant parasitism. Frequent cases of anthelmintic resistance have been reported (12–14) as well as ectoparasites, such as Dermatobia hominis (15, 16), Rhipicephalus microplus (17) and Cochliomyia hominivorax (18), with resistance to the treatments with macrocyclic lactones. For this reason, the aim of this work was to verify whether closantel and ivermectin that have been used for decades are still effective in sheep naturally infested with O. ovis. Because animals rarely die due to oestrosis, the economic losses caused by the parasitism may be underestimated. Therefore, the present study aimed also to assess the prophylactic effects of closantel and ivermectin on the productive performance of lambs.
The study was carried out according to the standards established by the local Ethics Committee on Animal Use (FMVZ/UNESP protocol number 0159/2019).
The experiment was conducted in the experimental area of the São Paulo State University (UNESP), Department of Biostatistics, Plant Biology, Parasitology and Zoology of the Bioscience Institute, Botucatu, SP, Brazil.
The lambs were kept in an area of 3.840 m2 divided in 5 paddocks, in rotational grazing on Urochloa decumbens, exposed to natural infestation with O. ovis, and infection with gastrointestinal nematodes. The area had been grazed previously by sheep with natural infestation by those parasites. The animals had free access to tap water and mineral salt (Supre Ovinos - Coopermota®) and were fed daily with supplement with 16.5% of crude protein (Ração Ovinos Confinamento - Coopermota®) with an amount corresponding to 2% of their body weight.
The experiment was conducted from December 2019 to March 2020, during the rainy season. Thirty 2 to 3-month-old uncastrated male lambs crossbred Santa Ines x White Dorper were purchased from a local commercial farm. The lambs arrived in the experimental area on October 29th, and underwent an adaptation period of 37 days, before the beginning of trial on December 5th. Upon the arrival, blood and fecal samples from all the animals were collected for hematological and parasitological procedures, and in addition, all lambs were vaccinated against clostridiosis (Sintoxan Polivalente® - Merial), and were treated with Toltrazuril (Farmacox® - Farmabase, 20 mg/kg, orally) in order to prevent coccidiosis.
The animals were randomly distributed into three groups that were as homogenous as possible regarding body weight: group 1 (n = 10), control animals that did not receive any antiparasitic treatment against oestrosis; group 2 (n = 10), animals treated with ivermectin (Ivomec® - Merial), 0.2 mg/kg, subcutaneous injection; and group 3 (n = 10), animals treated with closantel (Diantel® - Merial), 10 mg/kg, orally. Treatments against oestrosis with closantel or ivermectin were performed on 5th December 2019 and on 13th February 2020.
Additionally, all experimental animals were treated with an anthelmintic combination with monepantel (2.5 mg/kg, Zolvix® - Novartis), albendazole (10 mg/kg Endazol® - Hipra) and levamisole phosphate (9.4 mg/kg Ripercol – Zoetis) for three consecutive days, which were administered in three different occasions: on 12th December, 16th January and 13th February. These anthelminitic treatments were performed to prevent losses due to nematode infections, keeping helminth infection degree similar among groups.
Weekly, the lambs were weighed and examined looking for clinical signs of infestation by O. ovis, which included their nasal discharge degree (Figure 1; Table 1), recorded according to Dorchies et al. (7).
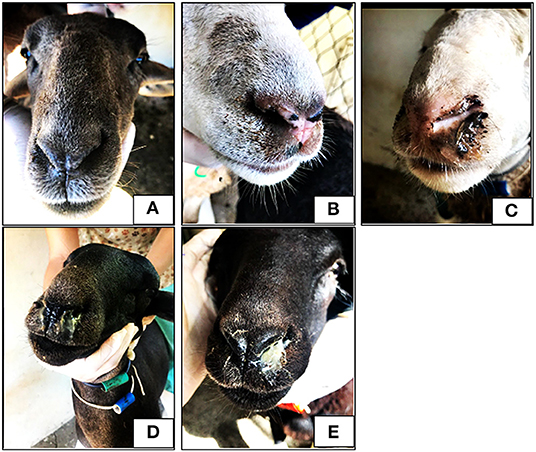
Figure 1. Photos of lambs with different nasal discharge score. No discharge (A), serous discharge (B), sero-mucous discharge (C), thick mucous discharge (D) and mucopurulent thick discharge (E).

Table 1. Nasal discharge score.
Feces samples were obtained directly from the rectum of each animal weekly to perform fecal egg counts by modified McMaster technique, in which each worm egg counted represented 100 eggs per gram (EPG) (19). In addition, fecal cultures were performed for each group to obtain infective larvae (L3) of gastrointestinal nematodes, which were identified according to descriptions of Ueno and Gonçalves (19).
The animals were slaughtered 35 days after the last oestrosis treatment. They had their heads removed and then sectioned along their longitudinal and sagittal axis. Nasal cavity (nasal passage, septum, middle meatus, and conchae) (Figure 2A) and frontal sinus (Figure 2B) were carefully examined, and all larvae found were collected, counted, and identified with regards to their stage of development (1, 20).
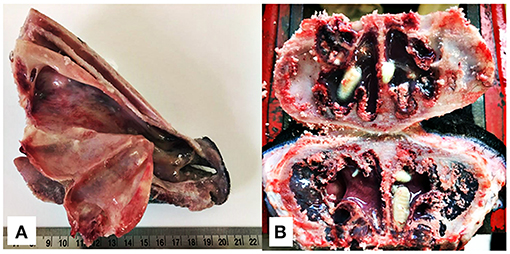
Figure 2. Longitudinal section of the rostral portion of the head without the mandible (A) and cross section of the nasal sinuses for the recovery of O. ovis larvae from naturally infested lambs (B).
One lamb from each group, with the highest EPG in the last sampling, was chosen for worm counts. After slaughter, the lambs had their abomasum and small intestine removed and opened longitudinally for content recovery. Their gastrointestinal contents were placed in graduated buckets to obtain aliquots of 10% of the total contents from each organ. The aliquots of 10% were stored in identified plastic flasks and preserved with 5% formaldehyde. All nematodes present in the 10% preserved material were morphologically identified and quantified according to their developmental stages (19).
Blood samples (5 ml) were obtained every 14 days by jugular vein puncture into Vacutainer® tubes containing anticoagulant (K2 EDTA 7.2 mg, BD, Brazil). Packed cell volume (PCV) was determined by microhematocrit centrifugation (5 min/15,000 g, Fanem 211 microhematocrit centrifuge), and Total Protein Plasmatic (TPP) concentrations were estimated using a handheld refractometer (Refractometer SPR-N, Atago) (21, 22). Eosinophil counts were performed in a Neubauer's chamber after staining with Carpentier's solution, and the counts were expressed as the number of eosinophil cells per μL of blood (23).
Plasma samples collected at 8 time-points were used to determine IgG levels against L2-soluble extract of O. ovis. The L2 extract was prepared as described by Silva et al. (24). A previously described protocol was applied to determine the parasite-specific plasma IgG levels (24) with some modifications: the plates were coated with 2 μg of antigen/mL, each wash was done six times which were incubated with peroxidase-conjugated rabbit anti-sheep IgG diluted at 1:5,000 then the plates were read at 450 nm by using an automated ELISA reader (Biotrak II; Amersham Biosciences, Little Chalfont, UK). For the negative control, plasma samples were obtained from young animals that were kept indoors without any contact with adult bot flies as previously described by Silva et al. (24). The plasma positive control sample used was from a sheep naturally infested with Oestrus ovis. Results were expressed as the percentage of plasma sample optical density (OD) value divided by OD of positive control.
The therapeutic efficacy of each antiparasitic was based on the mean number of O. ovis larvae recovered from lambs of each group at necropsy and calculated according to the following Abbot's formula (25).
For a therapeutic claim a reduction of larval counts should be at least 90% (25).
The meteorological data was registered daily (temperatures, precipitation and relative humidity). The climatic conditions during the experiment period were similar to those reported in the last years, except for February with total rainfall of 552.2 mm. This high value occurred because on the 9th it rained 299 mm, more than expected for the whole month. Data was obtained from the Lageado Experimental Farm weather station belonging to Unesp - Campus Botucatu at a distance of 8 km from the experimental area.
All data were submitted to the normality test Shapiro-Wilk and transformed using log10 (x + 1) when necessary, which was the case of EPG, O. ovis larvae stages, eosinophils and IgG values. Data were analyzed by one-way analysis of variance (ANOVA) in the case of variables measured only once (carcass weight, daily BWG and larvae stages of O. ovis) and by ANOVA with repeated measures in the case of variables measured at several time points (body weight, PCV, TPP, OD and eosinophils) using the General Linear Model (GLM) of the Statistical Analysis System, version 9.2 (SAS Institute, Inc., Cary, NC, USA). The group means were compared by Tukey's test and chi-square test was used for categorical variable (nasal discharge). Values of p < 0.05 were considered statistically significant.
Descriptive statistical analyses were used to summarize the data on larval burden, in agreement with Bush et al. (26), using the following terms:
Prevalence: the number of hosts infested with O. ovis larvae, divided by the number of hosts examined;
Intensity of infestation: the number of O. ovis larvae in a single infested host;
Mean intensity of infestation: the total number of O. ovis larvae found divided by the number of hosts infected with that parasite.
During the study none of sheep showed adverse effects as a consequence of the treatments. Seven animals of the control group were infested with O. ovis larvae, which were alive. The control group average, considering also three animals with zero value, was 8.1 larvae per sheep (Table 2), which were 1.4 L1, 4.3 L2 e 3.0 L3 per head. An animal from control group with the highest intensity of infestation had 17 larvae (four L2 and 13 L3).
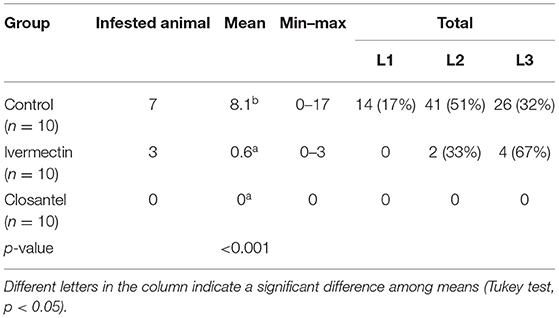
Table 2. First (L1), second (L2) and third stage (L3) larvae of Oestrus ovis in naturally infested lambs of the control, ivermectin and closantel groups.
The treatment with closantel was 100% effective against oestrosis, once there were no larvae on animals' heads. The treatment with ivermectin was 93% effective with three lambs presenting two larvae each in total six O. ovis larvae (Table 2). However, the larvae found were dead and had altered morphology, suggestive of the beginning of larva degeneration process. Differently from the lambs treated with ivermectin, the larvae identified in animals from the control group were active and without any morphological changes. Therefore, considering only alive larvae counting, the efficacy of ivermectin was 100%.
All the L1 observed in control group were in nasal conchae, 56% of the L2 were in ethmoidal conchae, and 44% in frontal sinus, while 23% of the L3 were in ethmoidal conchae and 77% in frontal sinus. Regarding the six dead larvae found in lambs treated with ivermectin, one L2 were in ethmoidal conchae, two L3 were in ethmoidal conchae and two L3 were in frontal sinus.
The clinical suggestive signs of oestrosis were scarce over the experimental period (Table 3) and there was no influence of treatment in the characterization of the lamb nasal discharge (p = 0.060). Each animal was examined 15 times over the trial. From the 450 nasal discharge exams conducted, 88% of them were serous, 9% were sero-mucous, 1% were very tick mucous, and 2% were mucopurulent thick (Table 3). This last score was observed in six animals, two from each group. Two weeks post the second oestrosis treatment, one animal from control group and one from group treated with closantel had muco-purulent discharges. Because lambs treated with closantel did not present larvae, it was possible to deduce that such discharge had no relation with O. ovis infestation.

Table 3. Total and percentage of nasal discharge classification of lambs naturally infested with Oestrus ovis of the control, ivermectin and closantel groups, in 15 evaluations from December 2019 to March 2020.
At the beginning of the trial, the average (±standard error) body weight were 23.1 (±1.3) kg, 22.8 (±1.4) kg and 22.9 (±1.6) kg, for non-treated control, ivermectin and closantel groups, respectively. In the last sampling, lambs were about 7–8 months old, and before slaughter, they had the following weight averages: 40.8 (±1.7), 40.5 (±2.0) kg and 40.6 (±1.6) kg for non-treated control, ivermectin and closantel groups, respectively. There was no significant difference among groups (p = 0.971) and also no significant group × time interaction (p = 0.615). The control group had daily gain mean (0.193 ± 0.016 kg), similar to ivermectin group (0.192 kg ± 0.01) and closantel group (0.192 kg ±0.02) (p = 0.994). In the same way, the means of carcass weight were similar (p = 0.999) among the different groups: 19.16 kg (±0.80), 19.11 kg (±1.01), 19.13 kg (±0.92), for control, ivermectin and closantel groups, respectively (Table 4).

Table 4. Initial and final body weight, daily body weight gain and carcass weight averages (±S.E.) of lambs naturally infested with Oestrus ovis of the control, ivermectin and closantel groups.
Because the animals had been kept in feedlot on their commercial farm of origin, EPG values were low and relatively homogeneous with averages similar between groups (p = 0.611) at the arrival in the experimental facilities. Lambs from control group had 70 (±39.58) EPG, lambs from ivermectin group had 740 (±322.90) EPG, and lambs from closantel group had 370 (±247.68) EPG.
After the first treatment for oestrosis, closantel group had significantly lower (p < 0.001) EPG mean (300 ± 84) than the control (2,220 ± 424) and ivermectin (2,730 ± 925) groups. In order to avoid interference of helminthiasis in the animals' performance, it was chosen to drench all the animals with the combination of three anthelmintics (levamisole + albendazole + monepantel) on three occasions as previously described. By adopting this procedure, the EPG counts remained similar in all groups until the day of slaughter (Figure 3). Despite the low EPG values, group mean differences were observed on 19th December 2019, when the EPG average of the control group (390 ± 55) was higher (p < 0.001) than the means of the ivermectin (110 ± 48) and closantel (40 ± 22) groups; and on the last evaluation (12th March 2020), when once again the control group presented EPG mean (170 ± 49) significantly higher (p = 0.024) than the closantel (40 ± 40) and ivermectin (70 ± 30) groups. There was no significant interaction between the group and time (p = 0.281), but there was significant effect of time (p < 0.001) in the EPG counts.
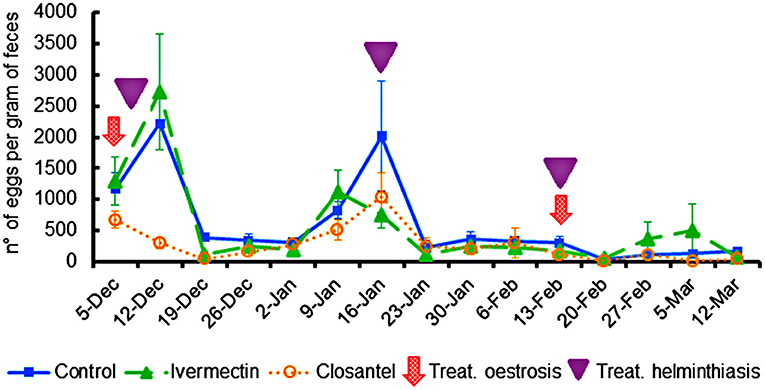
Figure 3. Averages of eggs per gram feces (±standard error) of lambs naturally infected with gastrointestinal nematodes and infested with O. ovis. The moments of treatments against oestrosis (with ivermectin or closantel) are indicated by arrow and treatments against helminthiasis (with a combination of levamisole + albendazole + monepantel) are indicated by triangle.
It was also observed eggs of Strongyloides spp. and Moniezia spp. and oocysts of Eimeria spp. in fecal exams, however in small amounts. None of lambs had symptoms of parasitic gastroenteritis during the trial.
In fecal cultures, larvae of Haemonchus spp., Cooperia spp., and Trichostrongylus spp. were detected. However, Haemonchus spp. predominated during the experiment, being the only genus recorded from February until the end of experiment period (Table 5).
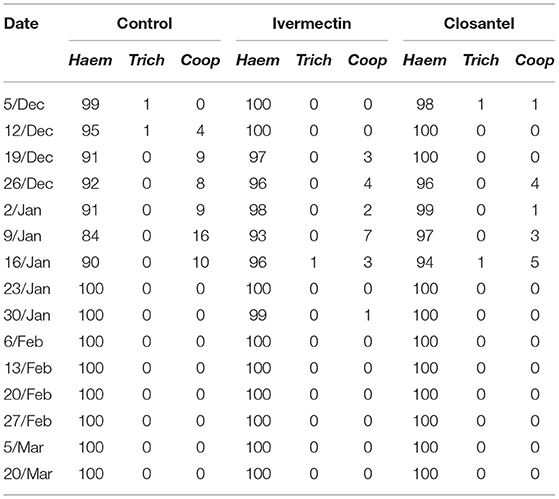
Table 5. Percentage of infective larvae of Haemonchus spp. (Haem), Trichostrongylus spp. (Trich) e Cooperia spp. (Coop) in fecal cultures from control, ivermectin and closantel groups.
In the examination of the gastrointestinal contents, Haemonchus spp. was the major genus found, with a high number of immatures stages (Table 6). Additionally, ten adult females of Trichostrongylus spp. were recovered from a lamb treated with ivermectin and ten in an animal of the control group (Table 6).
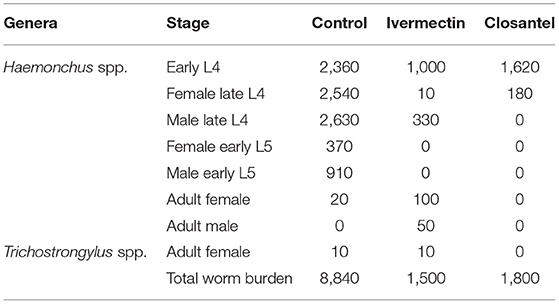
Table 6. Total Haemonchus contortus and Trichostrongylus spp. worm burden from one lamb of each group naturally infected with gastrointestinal nematodes.
The values of eosinophils, PVC and TPP (Figure 4) were similar among the three groups throughout the trial, and there were no significant interactions between group and time for these variables. The only exception occurred on 19th December 2019, at the beginning of the experiment, when the TTP mean of ivermectin group (6.02 ± 0.109) was significantly higher than the closantel group (5.60 ± 0.126) (p = 0.017). The lowest means of PCV (Figure 4B) were registered on 16th January 2020 (p = 0.097), that were 21.7% (±3.65), 25.2% (±4.10) and 24.2% (±2.86), for the control, ivermectin and closantel groups, respectively. This reduction in the values of PCV coincided with the increase in the EPG means (Figure 3). Additionally, in the same date, it was observed the highest means of blood eosinophils (Figure 4A).
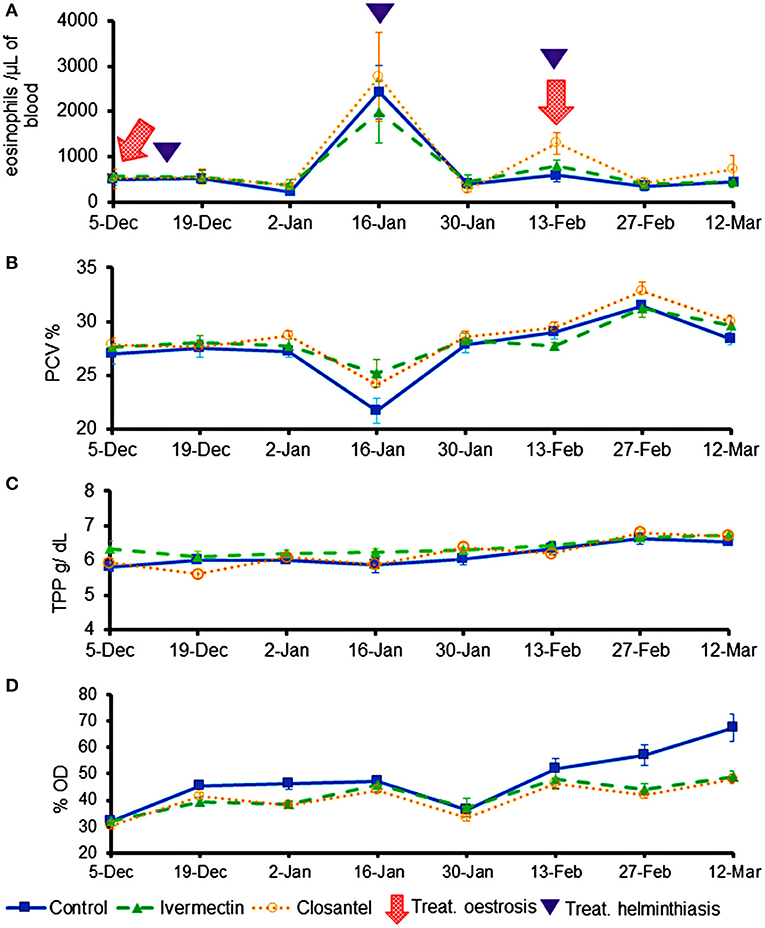
Figure 4. Averages (±standard error) of eosinophils (cells/μL) (A), packed cell volume (%) (B), total plasma protein (g/dL) (C), and optical density (%OD) of IgG anti-O. ovis L2 (D) of lambs treated with ivermectin or closantel. The moments of treatments against oestrosis (with ivermectin or closantel) are indicated by arrow and treatments against helminthiasis (with a combination of levamisole + albendazole + monepantel) are indicated by triangle.
The mean levels of anti-O. ovis L2 IgG in plasma showed changes over time, resulting in significant interaction between time and treatment (p = 0.007). The antibody levels increased in the control group on the last 2 weeks of the trial, being significantly higher than values of the treated groups (p = 0.004) on 27th February and 12th March (Figure 4D). The three animals from control group that did not have any larvae of O. ovis presented the lowest averages of antibodies in their group (lamb 1: 49%; lamb 2: 49%; and lamb 4: 45%).
The results of this trial confirmed the high effectiveness of closantel in animals naturally infested by O. ovis, with the absence of larvae 35 days after the last treatment. Regarding the ivermectin, some treated animals still hosted larvae of second and third stages. Nevertheless, the larvae recovered from these animals were already dead presumably because of ivermectin action. However, it was not possible to determine when the larvae died. It is possible that the larvae have died soon after the treatment, but, somehow, they remained in the nasal cavities. The dead larvae are possibly expelled by sneezes that combined with the force of gravity cause nasal discharge.
Therefore, the results of the present study were in accordance with Dorchies et al. (7) that reported 100% of efficacy of oral closantel and injectable ivermectin against oestrosis, in sheep treated twice with an interval of 60 days. Likewise, oral ivermectin resulted in 100% efficacy against all stages 12 days after the treatment (9). Different of other Diptera that cause myiasis (D. hominis and C. hominivorax) with cases of drug-resistance (15-16-18), to the best of our knowledge, there is no report of populations of O. ovis with resistance to ivermectin or closantel.
The typical clinical signs of oestrosis were uncommon over trial and during the examination performed at necropsy there were no macroscopic morphological changes in the nasal cavities. There was only a small amount of secretion at the site where parasites were located and around the larvae. The absence of clinical sings in most of the evaluations may be a consequence of the low infestation rate. Oestrus ovis clinical manifestation is induced partially by spines on the larval cuticle and oral hooks used by L1 to move and progress toward nasal cavities and to anchor to the mucous surface. In addition, the L3 has large hooks, stout spines and dorsal plates which serve to support its gradual descend in the nasal cavities to the outside environment (4), whereupon this mechanical action provokes irritation in nasal mucosa (27). The pathogenesis of O. ovis is also induced by biomolecules (enzymes and antigens) excreted or secreted by the larvae onto the mucosa. These biomolecules comprise a complex array of enzymes necessary to degrade mucus and plasma proteins for larval feeding and nutrition (28). In agreement with our results, Silva et al. (6) observed that most tracer lambs in their study did not show any clinical signs typical of oestrosis after exposed to infestation during 4 weeks, although they hosted among 12 and 66 larvae, mainly L1 and L2. Like in the trial conducted by Silva et al. (6), possibly in our trial the infestation was recent, without enough time to manifestation of the immunopathological changes typical of the disease. Although not all infested sheep show clinical signs of oestrosis, the major symptoms of infestation (nasal discharge and frequent sneezing) are immune-mediated depending on acquisition of an immune response against the parasite (6, 29). Infestation induces a great recruitment of inflammatory cells such as mast cells and eosinophils, to site of host-parasite interaction and also increases the immunoglobulin production (29).
If the trial had a longer duration, perhaps the typical clinical signs would occur, because the ability of sheep to become resistant to oestrosis is limited, different from what usually happens in infection by helminths (24). Also drew attention the fact that the treatment against oestrosis did not influence the weight gain of the animals, demonstrating the benign character of the disease in lambs. The treatment with anthelminthics, including in those lambs from control group, avoided the interference of helminthiasis in the body weight gain of the lambs, which were around 190 g per day, within the expected for grass fed male lambs supplemented with concentrate (30). However, more studies are necessary to evaluate the influence of oestrosis on sheep productivity in our environmental conditions, especially in older animals continuously exposed to the parasitism.
Lopes et al. (31) reported a case of poisoning by closantel in three animals from a group of 15 sheep treated with closantel twice with interval of 28 days. These lambs showed apathy, anorexia, diarrhea and blindness after the second treatment with 7.5 mg/kg orally. In our trial, lambs did not show any adverse effects post treatment with closantel or ivermectin. Some studies have shown that well-nourished animal, which was the case of our lambs, has less chances of poisoning because the closantel binds to plasmatic proteins, mainly albumin and healthy animals tend to have higher plasmatic protein level that reduce the amount of drug available in tissues (32, 33).
There was an increase in EPG means and a reduction in PCV values caused by Haemonchus spp. in all groups on 16th January 2020. After the anthelmintic treatment, PCV means increased and remained within the reference interval for sheep until the end of the study. Concomitantly, it was observed a significant increase in blood eosinophils counts on 16th January. Eosinophilia has been reported as one of the defense mechanisms against the gastrointestinal nematode parasitism (34, 35) and against O. ovis infestation (36).
While IgG-anti O. ovis means remained low in treated groups, the control lambs showed an increase in averages at the end of the trial, thus indicating that the animals were responding to parasite infestation. Previous studies demonstrated that detection of O. ovis antibodies in sheep is a valid diagnosis technique (24, 37–39).
Closantel oral drench at a dose rate 10 mg/kg and ivermectin subcutaneously at 0.2 mg/kg remain effective in the prophylaxis of oestrosis. These drugs are effective at the same concentrations used in the previous studies and there is no evidence of increasing trends of resistance. At the early stage of infestation, O. ovis parasitism did not cause a negative impact on the performance of lambs.
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author/s.
The animal study was reviewed and approved by Ethics Committee on Animal Use (FMVZ/UNESP Protocol Number 0159/2019).
HB performed all experiment, collected and analyzed the data, and completed the manuscript preparation. JL, ACA, and GF helped in the collection of data. AFA, JL, ACA, and MA participated in its design and coordination and helped to interpret the results. All authors interpreted the results and substantively revised the manuscript, read, and approved the final manuscript.
HB and JL received scholarship from Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), ACA was in receipt of a fellowship from Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP) (grant #2021/03479-1), GF received scholarship from PIBIC-CNPq, and AFA was in receipt of a fellowship from Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) (grant #305187/2017-1). This study was funded by Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP) (grant #2019/25185-0).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Zumpt P. Myiasis in man and animals in the old world. Bull Wildlife Dis Assoc. (1967) 3:40. doi: 10.7589/0090-3558-3.1.1.a
2. Panadero-Fontán R, Otranto D. Arthropods affecting the human eye. Vet Parasitol. (2015) 208:84–93. doi: 10.1016/j.vetpar.2014.12.022
3. Hall M, Wall R. Myiasis of humans and domestic animals. Adv Parasitol. (1995) 35:257–334. doi: 10.1016/S0065-308X(08)60073-1
4. Giannetto S, Santoro V, Pampiglione S. Scanning electron microscopy of Oestrus ovis larvae (Diptera: Oestridae): skin armour and posterior spiracles. Parasite. (1999) 6:73–7. doi: 10.1051/parasite/1999061073
6. Silva BF, Bassetto CC, Amarante AFT. Epidemiology of Oestrus ovis (Diptera: Oestridae) in sheep in Botucatu, State of São Paulo. Rev Bras Parasitol Vet. (2012) 21:386–90. doi: 10.1590/S1984-29612012000400008
7. Dorchies Ph, Alzieu JP, Cadiergues MC. Comparative curative and preventive efficacies of ivermectin and closantel on Oestrus ovis (Linn 1758) in naturally infected sheep. Vet Parasitol. (1997) 72:179–84. doi: 10.1016/S0304-4017(97)00052-6
8. Oliveira CMB, Moro E, Caproni JL, Gonçalves LCB, Umehara O, Oliveira LO. Efficacy of doramectin in the treatment of sheep naturally infested by Oestrus ovis. Rev Bras Parasitol Vet. (2000) 9:61–4. doi: 10.1016/s0304-4017(00)00429-5
9. Lucientes J, Castillo JA, Ferrer LM, Peribáñez MA, Ferrer-Dufol M, Gracia-Salinas MJ. Efficacy of orally administered ivermectin against larval stages of Oestrus ovis in sheep. Vet Parasitol. (1998) 75:255–9. doi: 10.1016/S0304-4017(97)00168-4
10. Rugg R, Ferrer LM, Sarasola P, Figueras L, Lacasta D, Liu B, et al. Persistent efficacy of a long acting injectable formulation of moxidectin against natural infestations of the sheep nasal bot (Oestrus ovis) in Spain. Vet Parasitol. (2012) 188:330–6. doi: 10.1016/j.vetpar.2012.03.057
11. Martínez-Valladares M, Valcárcel F, Álvarez-Sánchez MA, Cordero-Pérez C, Fernández-Pato N, Frontera E, et al. Efficacy of moxidectin long-acting injectable formulation (1 mg/kg bodyweight) against first instar larvae of Oestrus ovis in naturally infected sheep. Vet Parasitol. (2013) 193:320–4. doi: 10.1016/j.vetpar.2012.10.006
12. Almeida FA, Garcia KCOD, Torgerson PR, Amarante AFT. Multiple resistance to anthelmintics by Haemonchus contortus and Trichostrongylus colubriformis in sheep in Brazil. Parasitol Int. (2010) 59:622–5. doi: 10.1016/j.parint.2010.09.006
13. Neves JH, Carvalho N, Rinaldi L, Cringoli G, Amarante AF. Diagnosis of anthelmintic resistance in cattle in Brazil: a comparison of different methodologies. Vet Parasitol. (2014) 206:216–26. doi: 10.1016/j.vetpar.2014.10.015
14. Albuquerque ACA, Bassetto CC, Almeida F A, Amarante AF. Development of Haemonchus contortus resistance in sheep under suppressive or targeted selective treatment with monepantel. Vet Parasitol. (2017) 246:112–7. doi: 10.1016/j.vetpar.2017.09.010
15. Neves JH, Carvalho N, Amarante AF. Dermatobia hominis: potencial risk of resistance to macrocyclic lactones. Vet Parasitol. (2015) 212:483–6. doi: 10.1016/j.vetpar.2015.06.029
16. Conde MH, Borges DGL, Freitas MG, Silva MC, Almeida BF. First report of Dermatobia hominis resistant to doramectin in cattle. Vet Parasitol. (2020) 289:109–335. doi: 10.1016/j.vetpar.2020.109335
17. Reck J, Klafke GM, Webster A, Dall'agnol B, Scheffer R, Souza UA, et al. First report of fluazuron resistance in Rhipicephalus microplus: a field tick population resistant to six classes of acaricides. Vet Parasitol. (2014) 201:128–36. doi: 10.1016/j.vetpar.2014.01.012
18. Lopes WDZ, Teixeira WFP, Felippelli G, Cruz BC, Maciel WG, Matos LVSD, et al. Ivermectina e abamectina em diferentes doses e vias de aplicação contra larvas de Cochliomyia hominivorax em bolsas escrotais de bovinos recém-castrados, provenientes da região sudeste do Brasil. Cienc Rural. (2013) 43:2195–201. doi: 10.1590/S0103-84782013001200013
19. Ueno H, Gonçalves PC. Manual para diagnóstico das helmintoses de ruminantes. 4th ed. Tokyo: Japan International Cooperation Agency (1998). 143 p.
20. Capelle KJ. The occurrence of Oestrus ovis L. (Diptera: Oestridae) in the bighorn sheep from Wyoming and Montana. J Parasitol. (1966) 52:618–21. doi: 10.2307/3276337
21. Weiser G. Laboratory technology for veterinary medicine. In: Thrall MA, editor. Veterinary Hematology and Clinical Chemistry. 2th ed. Iowa: Wiley-Blackwell (2012). 775 p.
22. Weiss DJ, Wardrop KJ. Schalm's Veterinary Hemathology. 6th ed. Iowa: Wiley-Blackwell (2010). 1206 p.
23. Dawkins HJS, Windon RG, Eagleston GK. Eosinophil response in sheep selected for high and low responsiveness to Trichostrongylus colubriformis. Int J Parasitol. (1989) 19:199–205. doi: 10.1016/0020-7519(89)90008-8
24. Silva BF, Bassetto CC, Amarante AFT. Immune responses in sheep naturally infected with Oestrus ovis (Diptera: Oestridae) and gastrointestinal nematodes. Vet Parasitol. (2012) 190:20–126. doi: 10.1016/j.vetpar.2012.06.004
25. Holdsworth P, Rehbein S, Jonsson NN, Peter R, Vercruysse J, Fourie J. World association for the advancement of veterinary parasitology (WAAVP): guideline for evaluating the efficacy of parasiticides against ectoparasites of ruminants. Vet Parasitol. (2021). doi: 10.1016/j.vetpar.2021.109613. [Epub ahead of print].
26. Bush AO, Lafferty KD, Lotz JM, Shostak AW. Parasitology meets ecology on its own terms: Margolis et al. revisited. J Parasitol. (1997) 83:575–83. doi: 10.2307/3284227
27. Angulo-Valadez CE, Cepeda-Palacios R, Ascencio F, Jacquiet P, Dorchies P, Romero MJ, et al. Proteolytic activity in salivary gland products of sheep bot fly (Oestrus ovis) larvae. Vet Parasitol. (2007) 149:117–25. doi: 10.1016/j.vetpar.2007.06.040
28. Frugère S, Leon AC, Prévot F, Palacios RC, Tabouret G, Bergeaud JP, et al. Immunisation of lambs with excretory secretory products of Oestrus ovis third instar larvae and subsequent experimental challenge. Vet Res. (2000) 31:527–35. doi: 10.1051/vetres:2000105
29. Angulo-Valadez CE, Ascencio F, Jacquiet P, Dorchies P, Cepeda-Palacios R. Sheep and goat immune responses to nose bot infestation: a review. Med Vet Entomol. (2011) 25:117–25. doi: 10.1111/j.1365-2915.2010.00911.x
30. Starling RZC, Almeida FA, Viana MVG, Castilhos AM, Amarante AFT. Losses caused by gastrointestinal nematode infections in Dorper lambs under two nutritional status. Rev Bras Parasitol Vet. (2019) 28:652–60. doi: 10.1590/s1984-29612019084
31. Lopes WDZ, Carvalho RS, Oliveira PV, Pereira V, Martinez AC, Mazzucatto BC, et al. Intoxicação de ovinos que receberam duas aplicações de closantel, na dosagem terapêutica (7,5mg/kg), com intervalo de 28 dias. Pesq Vet Bras. (2014) 34:1162–6. doi: 10.1590/S0100-736X2014001200003
32. Henessy DR, Sangster NC, Steel JW. Comparative pharmacokinetic disposition of closantel sheep and goats. J Vet Pharmacol Ther. (1993) 16:254–60. doi: 10.1111/j.1365-2885.1993.tb00172.x
33. Barlow AM, Sharpe JAE, Kincaid EA. Blindness in lambs due to inadvertent closantel overdose. Vet Res. (2002) 151:25–6. doi: 10.1136/vr.151.1.25
34. Meeusen EN, Balic A. Do eosinophils have a role in the killing of helminth parasites? Parasitol Today. (2000) 16:95–101. doi: 10.1016/S0169-4758(99)01607-5
36. Dorchies Ph, Bergeaud J, Khanh N, Morand S. Reduced egg counts in mixed infections with Oestrus ovis and Haemonchus contortus : influence of eosinophils? Parasitol Res. (1997) 83:727–30. doi: 10.1007/s004360050328
37. Alcaide M, Reina D, Sánchez-López J, Frontera E, Navarrete I. Seroprevalence of Oestrus ovis (Diptera, Oestridae) infestation and associated risk factors in ovine livestock from Southwestern Spain. J Med Entomol. (2005) 42:327–31. doi: 10.1603/0022-2585(2005)042[0327:SOOODO]2.0.CO;2
38. Angulo-Valadez CE, Scala A, Grisez C, Prévot F, Bergeaud JP, Carta A, et al. Specific IgG antibody responses in Oestrus ovis L. (Diptera: Oestridae) infected sheep: associations with intensity of infection and larval development. Vet Parasitol. (2008) 155:257–63. doi: 10.1016/j.vetpar.2008.05.015
Keywords: myiasis, Ovis aires, lambs, bot fly, Oestridae
Citation: Bello HJS, Lins JGG, Albuquerque ACA, Ferreira GB, Amarante MRV and Amarante AFT (2022) Prophylactic Effects of Ivermectin and Closantel Treatment in the Control of Oestrus ovis Infestation in Sheep. Front. Vet. Sci. 8:798942. doi: 10.3389/fvets.2021.798942
Received: 20 October 2021; Accepted: 22 December 2021;
Published: 18 January 2022.
Edited by:
Annamaria Passantino, University of Messina, ItalyReviewed by:
Smaragda Sotiraki, Veterinary Research Institute Greek Agricultural Organization Demeter, GreeceCopyright © 2022 Bello, Lins, Albuquerque, Ferreira, Amarante and Amarante. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Hornblenda Joaquina Silva Bello, aG9ybmJsZW5kYS5iZWxsb0B1bmVzcC5icg==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.