 Zhongle Li
Zhongle Li Qi Wang2†
Qi Wang2† Keping Sun
Keping Sun Jiang Feng
Jiang Feng
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
SYSTEMATIC REVIEW article
Front. Vet. Sci. , 17 December 2021
Sec. Veterinary Epidemiology and Economics
Volume 8 - 2021 | https://doi.org/10.3389/fvets.2021.791237
Chytridiomycosis is an amphibian fungal disease caused by Batrachochytrium dendrobatidis (Bd), which has caused large-scale death and population declines on several continents around the world. To determine the current status of Bd infection in amphibians, we conducted a global meta-analysis. Using PubMed, ScienceDirect, SpringerLink, China National Knowledge Infrastructure (CNKI) and Wanfang database searches, we retrieved a total of 111 articles from 2000 to 2021. Based on these, we estimated the Bd prevalence to be 18.54% (95% CI: 13.76–20.52) in current extent amphibians. Among these populations, the prevalence of Bd in Asia was the lowest at 7.88% (95% CI: 1.92–8.71). Further, no Bd infection was found in Vietnam. However, the prevalence of Bd in Oceania was the highest at 36.34% (95% CI: 11.31–46.52). The Bd prevalence in Venezuela was as high as 49.77% (95% CI: 45.92–53.62). After 2009, the global Bd prevalence decreased to 18.91% (95% CI: 13.23–21.56). The prevalence of Bd in epizootic populations was significantly higher than enzootic populations. The highest prevalence of Bd was detected with real-time PCR at 20.11% (95% CI: 13.12–21.38). The prevalence of Bd in frogs was the highest at 20.04% (95% CI: 13.52–21.71), and this different host was statistically significant (P < 0.05). At the same time, we analyzed the geographic factors (longitude, latitude, elevation, rainfall and temperature) that impacted the fungal prevalence in amphibians. Our meta-analysis revealed that factors including region, disease dynamic, detection method, host and climate may be sources of the observed heterogeneity. These results indicate that chytridiomycosis was a consistent threat to amphibians from 2000 to 2021. Based on different habitat types and geographical conditions, we recommend formulating corresponding control plans and adopting reasonable and efficient biological or chemical methods to reduce the severity of such diseases.
- The highest prevalence of Bd was in Oceania, followed by South America.
- The prevalence of amphibian chytridiomycosis has decreased over the past decade.
- Climate and geographical conditions may be the main risk factors for amphibian chytridiomycosis.
Over the last four decades, ~43% of amphibian species have become threatened with extinction worldwide (1, 2). Habitat destruction (3), natural environment pollution (4), climate change (5) and emerging infectious diseases (6) are all potential causes of morbidity and mortality. Chytridiomycosis is the disease caused by Batrachochytrium dendrobatidis (Bd) and has become a major menace to amphibians, leading to the decline and extinction of amphibians worldwide (7, 8).
Bd invades epidermal cells and grows the outer keratinized layers, causing skin thickening, erosions and ulcerations, disrupting the transport of water, oxygen and ions, and eventually leading to death (9). This pathogen has been detected in ~700 amphibian species (10) and occurs in different regions around the world, including Europe, Oceania, Africa, North America and South America (11, 12), particularly in the Neotropics and Australia (13, 14). Bd may be one of the most seriously infectious diseases currently threatening biodiversity (8) and is now listed as an important epidemic disease by the World Organization for Animal Health (15). Currently, there are several methods to control the spread of Bd, such as use of fungicides, bioaugmentation (microorganisms that inhibit the growth of Bd) and vaccination. The first two methods may be considered risky for other species due to their broad spectrum, and the effectiveness and safety of vaccination still need be further validation for controlling chytridiomycosis in the field (16, 17).
Bd is regarded as a generalist pathogen (18) and is distributed across different ecosystems (19, 20). The prevalence and infection intensity of Bd are associated with abiotic factors (21) and may vary according to host species, age and individual susceptibility (22). Currently, there are several Bd lineages, mainly divided into enzootic and epizootic lineages, where the enzootic lineages has a lower virulence probably due to its smaller zoosporangium size, and more limited geographic distributions vs. epizootic lineages (23–25). Meanwhile, the inherent characteristics of amphibians (behavior and immunity) and environmental features (vegetation coverage, temperature, rainfall, seasonality, and geographical conditions) can predict dynamic changes of diseases in natural populations (26, 27).
Amphibians are a critical links in ecosystem food chains, as they are able to prey on pests, effectively protect crops and play an important role in maintaining the ecosystem balance (18, 28). In addition, some amphibians are edible while others have high medicinal and scientific research value (29, 30). Considering the serious impact of chytridiomycosis on the health of amphibians, there are still limited articles on the overall assessment of the factors that potentially influence the prevalence of Bd. Thus, we conducted a global systematic review and meta-analysis on Bd infection over the last 21 years. The sampling year, geographic location, disease dynamic, detection method, season, host, age, International Union for the Conservation of Nature (IUCN) classification, sampling source and other potential risk factors along with several geographic factors were collected and analyzed to determine which were related to Bd prevalence in amphibians. This study could support ongoing conservation efforts for amphibian chytridiomycosis.
We used the preferred reporting items for systematic reviews and meta-analysis (PRISMA) algorithm to select reasonable study reports (31). In this study, we searched for articles with the prevalence of Bd in amphibians worldwide available from five literature databases, including PubMed, ScienceDirect, SpringerLink, China National Knowledge Infrastructure (CNKI) and Wanfang Databases (the retrieval time was from Jan 1, 2000, to August 18, 2021). In PubMed, we used the MeSH terms “Batrachochytrium dendrobatidis,” “Prevalence,” and “Amphibians” for our searches. Then we queried the corresponding MeSH term “Batrachochytrium dendrobatidis” using the free words “Chytridiomycosis” and “Chytrid.” Simultaneously, the Boolean operator “AND” to connect MeSH terms and “OR” to connect the free words were used. Finally, the search formula “(Batrachochytrium dendrobatidis OR Chytridiomycosis OR Chytrid) AND (Amphibians) AND (Prevalence)” was used. In the ScienceDirect and SpringerLink databases, we used the same search formula. Meanwhile, we used the keywords “(Batrachochytrium dendrobatidis) AND (Amphibians)” for advanced searches and added a fuzzy search and synonym expansion in two Chinese databases. Moreover, we did not include unpublished data and did not seek additional information from the authors of published articles. Endnote X9 (version 9.3.1) was used to sort out the retrieved citations.
We adopted the following inclusion criteria: (1) the study was aimed to estimate the prevalence of Bd from amphibians inhabiting the natural environment; (2) the study presented the number of examined species and the number of Bd positive species to evaluate the prevalence (the number of individuals positive to Bd/total number of individuals examined); (3) the sample size of the study was more than 60 (32); (4) the study design was cross-sectional for estimating the prevalence in disease studies. Moreover, the following exclusion criteria were used: (1) the study had internal data conflicts (the data described in the conclusion not corresponding to the actual number of tests); (2) the study included museum samples or specimen (unclear source of samples); (3) the study used experimental group data; (4) the study was conducted earlier than 2000; or (5) the study presented dead samples.
Based on the acquired studies, we extracted the information using standardized data collection forms: first author, publication year, sampling date, continent of the study, country of the study, disease dynamic (the process of the spread of diseases from epizootic phase to enzootic phase), detection method, host classification (order), geographical factors, collection season, age, International Union for Conservation of Nature (IUCN) category, sample source, total number of amphibian samples and the number of Bd positive samples. The geographic factor data were based on weather stations collected from the NOAA National Centers for Environmental Information (https://gis.ncdc.noaa.gov/maps/ncei/cdo/monthly), including longitude, latitude, elevation, monthly average rainfall and monthly average temperature.
The quality of the publications was scored with reference to a previously published method from the Grading of Recommendations Assessment, Development and Evaluation method (GRADE) (33, 34). The scoring criteria had the following conditions: clear detection method, definite random sampling, definite sampling time, clear sampling method and four or more risk factors evaluated in collection articles. Each condition was scored with one point. High-quality articles were valued to have 4–5 points, and medium-quality articles had 2–3 points. Low-quality articles had only 1 or 0 points (34). The scoring of an article did not represent the level of its research content, but was used to assess risk bias.
All statistical computing was conducted in R version 4.0.2 (35). Based on previous studies, we chose arcsine transformation (PFT) data to be closer to the normal distribution before our meta-analysis (W = 0.96961; P < 0.05; Supplementary Table 1) (36).
The formulas for PFT were as follows: t = arcsin {sqrt [r/(n + 1)]} + arcsin {sqrt [(r + 1)/(n + 1)]}, note: t = transformed prevalence; n = sample size; r = positive number; se = standard error (37). We calculated the heterogeneity between studies using Cochran's Q, I2 statistics and the χ2 test, where I2>50% indicates a significant degree of heterogeneity. Based on the obvious heterogeneity from the selected articles, we chose a random model for meta-analysis (38). In order to examine the potential sources of heterogeneity, we analyzed the research data using subgroup analyses and univariate regression analysis to identify potential risk factors predictive of heterogeneity. Forest plots were used to visually evaluate the overall results of metaanalysis. The studies of publication bias were explained with funnel plots and an Egger's test. Finally, we used sensitivity analysis to estimate the stability of our results (39).
We further studied the potential sources of heterogeneity from each subgroup (40), including the geographic region (Africa, Asia, Europe, North America, Oceania and South America), the sampling year (2000–2008 and after 2009), disease dynamic (enzootic and epizootic), detection method (conventional PCR, nested PCR, real-time PCR, histopathology examination and LAMP), host (caecilian, frog, salamander, toad, newt), season (spring, summer, autumn and winter), age (adult, subadult, juvenile and tadpole), IUCN category (least concern, near threatened, vulnerable, endangered, and critically endangered), sample source (forest, park, lake, pond, stream, and other sources), and quality level (middle quality and high quality).
We also analyzed the subgroup of geographical risk factors that influenced the heterogeneity (41), based on latitude (north latitude 0–15°, north latitude 15–30°, north latitude 30–60°, south latitude 0–20°, and south latitude 20–40°), longitude (east longitude 0–80°, east longitude 80–120°, east longitude 120–160°, west longitude 0–100°, and west longitude 100–160°), elevation (0–100 m, 100–500 m, 500–1700 m, and 1700–5000 m), monthly average rainfall (0–50 mm, 50–100 mm, 100–150 mm, and >150 mm) and monthly average temperature (5–16°C, 17–25°C, and >26°C). In addition, each subgroup was jointly analyzed with regions to trace the source of heterogeneity. The heterogeneity of the covariates is explained in R2.
According to the inclusion criteria for each article, we finally selected 111 studies from the five databases for meta-analysis (Figure 1), including 20 publications of medium quality (2–3 points) and 91 publications of high quality (4 or 5 points; Supplementary Table 4).
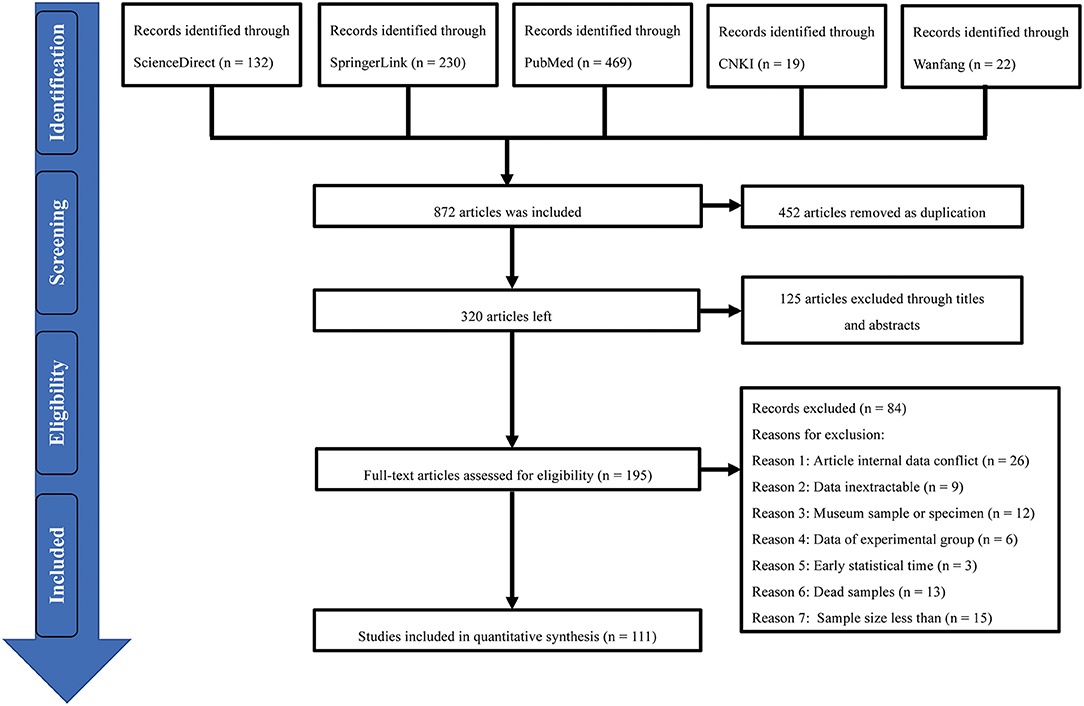
Figure 1. Flow diagram of the study screening process of inclusion and exclusion of studies.
The results of our funnel plots could not directly judge whether there was publication bias in the included studies (Figure 2), but the result of an Egger's test demonstrated that there was no publication bias (P > 0.05; Supplementary Table 2). A forest plot illustrated the prevalence estimates of Bd in amphibians worldwide, showing an obvious heterogeneity between studies (χ2 = 11, 374.69, P = 0; I2 = 99.00%, 95% CI: 13.76–20.52; Supplementary Figure 1). Finally, the sensitivity analysis clarified the data reliability, when excluding any one study, the estimate values laid within the 95% CI of the respective overall prevalence, and had no significant influence (Supplementary Figure 2).
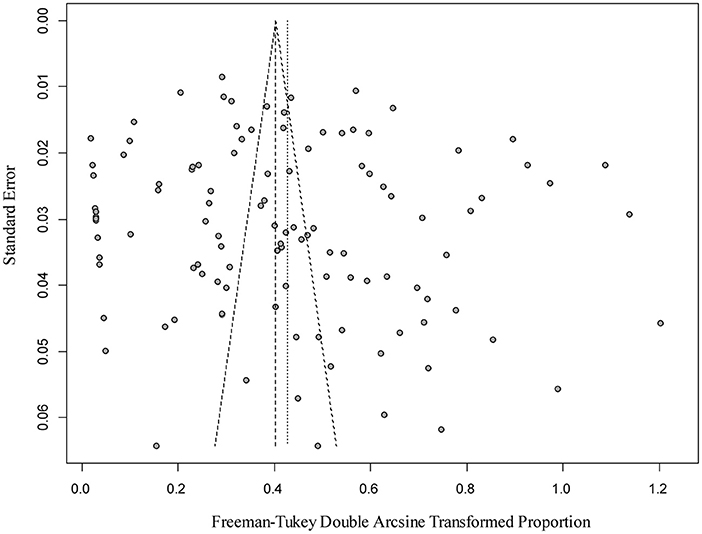
Figure 2. Funnel plot of the analysis publications bias of studies.
A total of 50,985 amphibians were surveyed, and the prevalence of Bd in amphibians worldwide was 18.54% (95% CI: 13.76–20.52; Supplementary Table 3). The detailed Bd prevalence in amphibians from different regions ranged from 7.88% (95% CI: 1.92–8.71) to 36.34% (95% CI: 11.31–46.52; Table 1). The prevalence was lower in Asia than in other continents (P < 0.05; Table 1). Furthermore, the prevalence of Bd varied among countries (Supplementary Table 3). Venezuela had the highest prevalence at 49.77% (95% CI: 45.92–53.62), and the lowest prevalence was Benin, Burkina Faso, Cameroon, Côte d'Ivoire, Ghana, Guinea, Liberia, Madagascar, Papua New Guinea, Ireland, Sierra Leone and Vietnam at 0.00% (Supplementary Table 3; Figure 3).
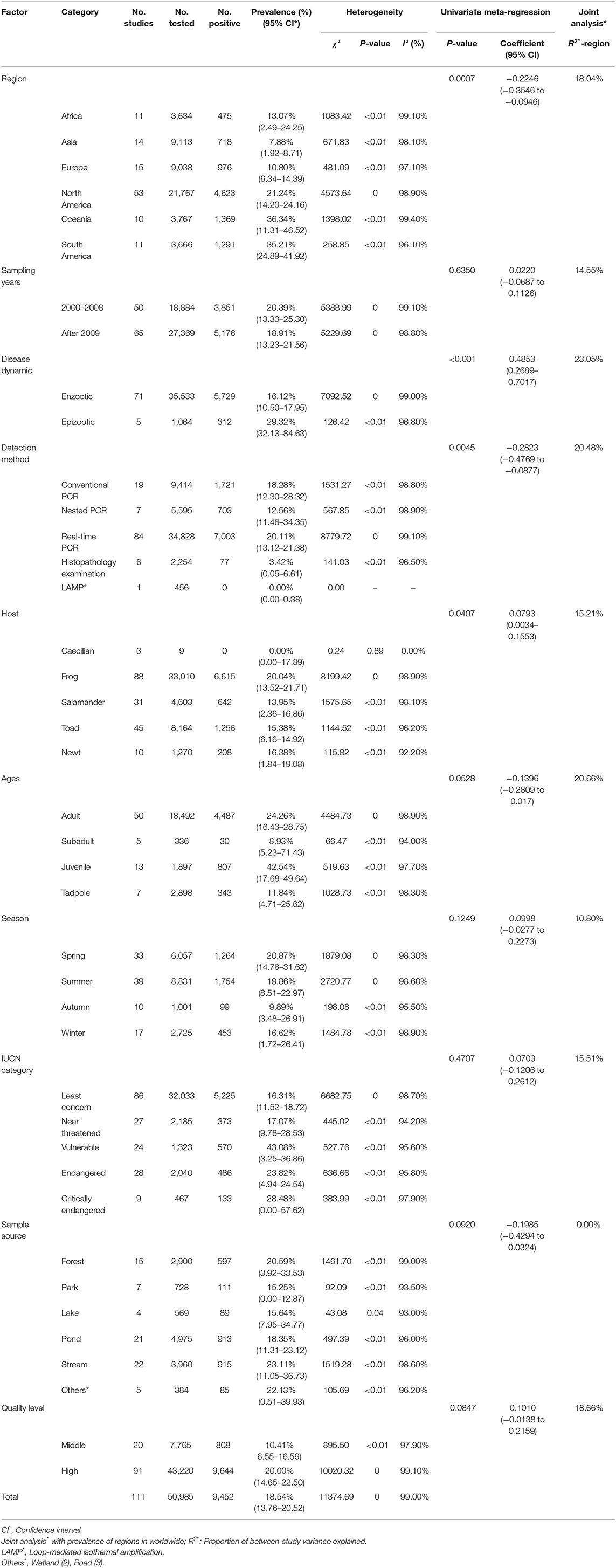
Table 1. Pooled worldwide prevalence of Batrachochytrium dendrobatidis by region.
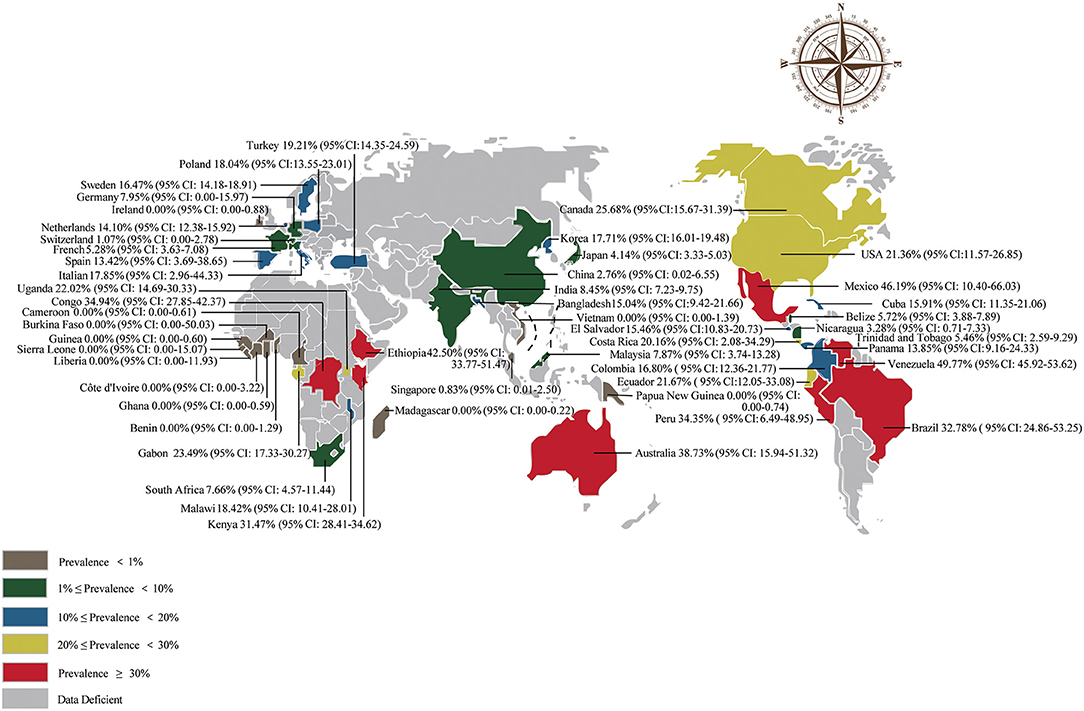
Figure 3. Map of the prevalence of Batrachochytrium dendrobatidis in amphibians worldwide.
We next estimated the possible potential risk factors for Bd prevalence, including geographical distribution, sampling year, detection methods, host, season, age, IUCN category and sample source (Table 1). The geographical distribution, disease dynamic, detection method and host were identified as main potential risk factors for Bd prevalence (P < 0.05). The pooled prevalence of Bd in the 2000–2008 group was 20.39% (95% CI: 13.33–25.30), which was higher than 18.91% (95% CI: 13.23–21.56) in the after 2009 group (Table 1). The epizootic disease had a significantly higher prevalence of 29.32% (95% CI: 32.13–84.63) relative to enzootic disease. Between real-time PCR, nested PCR, conventional PCR, histopathology examination and LAMP, the first method detected the highest prevalence at 20.11% (95% CI: 13.12–21.38). However, the frogs had the highest prevalence of 20.04% (95% CI: 13.52–21.71) relative to other groups. The prevalence of Bd in juvenile amphibians was higher at 42.54% (95% CI: 17.68–49.64) than the prevalence in other ages. Meanwhile, we found that the prevalence in spring was higher at 20.87% (95% CI: 14.78–31.62) than in other seasons. The prevalence in vulnerable amphibians was 43.08% (95% CI: 3.25–36.86) and was higher than in other categories. However, the prevalence from stream samples was higher at 23.11% (95% CI: 11.05–36.73) than in other sample sources.
Following this, we analyzed the geographical factors subgroup factors including latitude, longitude, elevation, monthly average rainfall and monthly average temperature (Table 2). The Bd prevalence in the south latitude range (20–40°; 32.38%, 95% CI: 16.83–40.35) and in the west longitude range (100–160°; 31.81%, 95% CI: 14.17–41.80) was significantly higher than other latitude and longitude ranges (P < 0.05). In the 1,700–5,000 m elevation, the prevalence was 33.15% (95% CI: 9.64–34.27), which was significantly higher than other elevation ranges (P < 0.05). The monthly average rainfall in the over 150 mm precipitation range had the highest prevalence at 28.58% (95% CI: 12.69–30.42). The prevalence in the monthly average temperature range of 17–25°C reached 26.48% (95% CI: 18.18–38.13), which was significantly higher than other temperature ranges (P < 0.05). The heterogeneity of each subgroup was explained by region (the covariate), ranging from 0 to 30.53% (R2-region) (Tables 1, 2).
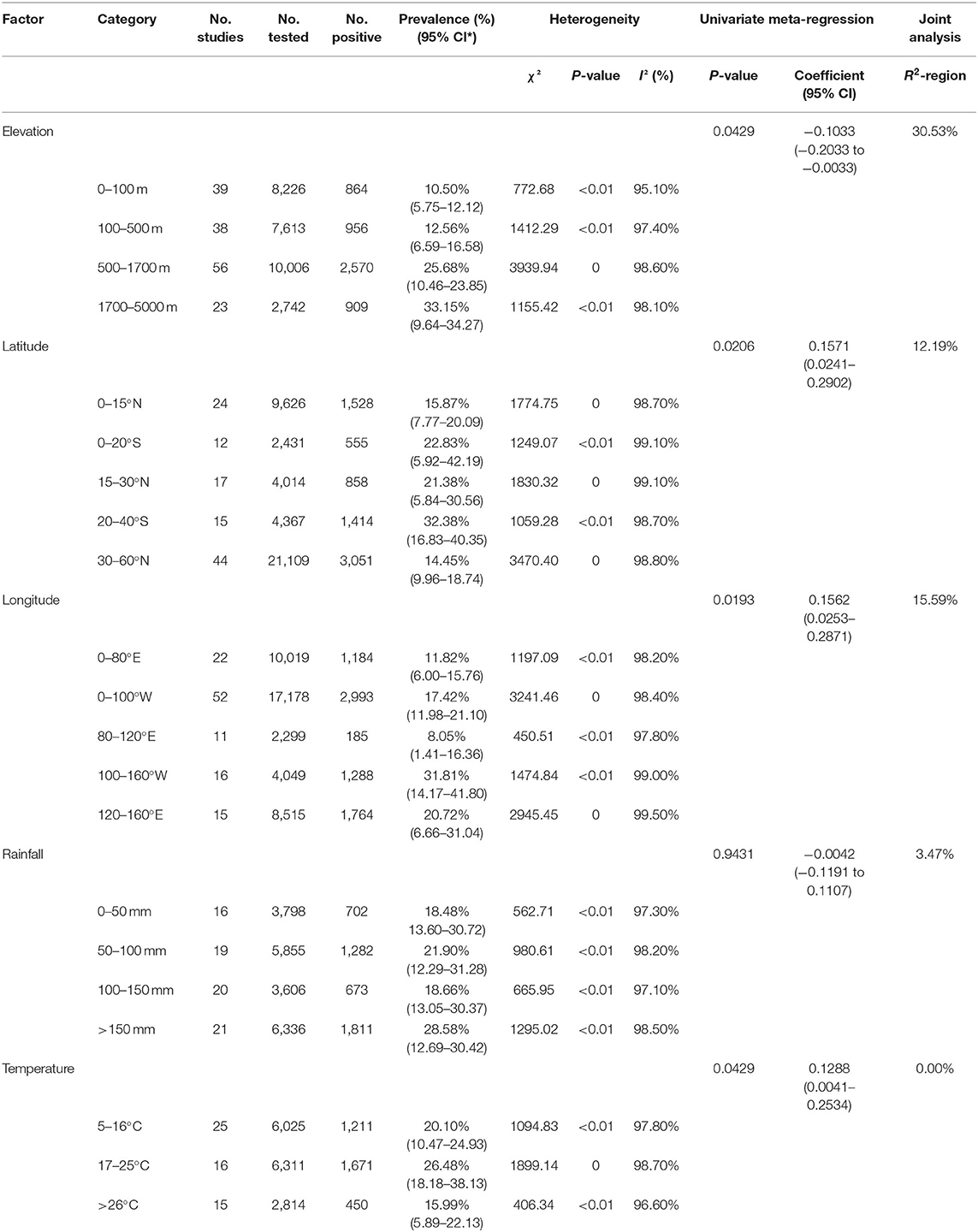
Table 2. Estimated pooled prevalence of Batrachochytrium dendrobatidis by geographical factor.
Chytridiomycosis is a worldwide infectious disease of amphibians, causing a decrease in biodiversity and a loss of economic benefits (42). Detailed knowledge of the epidemiological status of chytridiomycosis in amphibians provides a foundation for implementing efficient protective measures.
Therefore, we conducted a systematic review and meta-analysis of global Bd infection. The overall prevalence of Bd in amphibians worldwide from 2000 to 2021 was 18.54% (95% CI: 13.76–20.52; Supplementary Table 3).
In 2008, Office International des Epizooties (OIE) listed Batrachochytrium dendrobatidis as notifiable pathogen, because the international community recognized the impact of the global trade in amphibians on the spreading of chytridiomycosis (43). This legislation enabled countries connected to the World Trade Organization to specifically detect and restrict the trade of infected amphibians. Therefore, we chose 2008 as the entry point for our analysis of Bd infection in amphibians worldwide. We found that the Bd prevalence (18.91%) decreased after this policy was enacted (Table 1). Implementing similar policies might help to reduce Bd infection. Therefore, we suggest that more relevant legislation be implemented to protect amphibian species.
In our study, the Bd prevalence in South America and Oceania was basically the same and significantly higher than that in Asia (P < 0.05; Table 1). In view of our geographic subgroup analysis, we found that the highest Bd prevalence was in the latitude range 20–40°S and the longitude range 100–160°W (Table 2), which is also mainly concentrated in Oceania, Africa, South America and North America. Although Bd infection is mostly global, the distribution of fungus causing amphibian deaths has so far been limited to a few regions, mainly in Oceania, South America and North America (13, 44). Similarly, we found a high Bd prevalence in Australia, Brazil, Peru, Venezuela, Mexico, and the United States (21–49%; Supplementary Table 3). However, there were some areas where the prevalence of Bd was zero, mostly in Africa and Asia (Supplementary Table 3), which may be under-explored studies on the dynamics of amphibian chytridiomycosis (45). Or maybe frog populations in Africa have co-evolved with Bd to show resistance, even if the Bd was originated Africa (11, 46). When the elevation was between 1,700 and 5,000 m, the prevalence was significantly higher than that in the 0–100, 100–500, and 500–1,700 m groups (P < 0.05; Table 2). The joint analysis showed that regions explained 30.53% of heterogeneity in elevation subgroup (Table 2). These results may be due to the fact that different continents have different elevation characteristics. At high elevation areas, the prevalence of Bd increased and the survival rate of infected amphibians have been shown to be reduced (47, 48). In the entire Neotropical region, environmental conditions not only promote the abundance and diversity of amphibians, but also contribute to the growth and survival of Bd, especially in the cool and humid environments of many uplands (13). In central America, most amphibian declines occurred at elevations above 500 m (49). Since Bd is dependent on aquatic habitats (50) and intolerant to desiccation (51), higher levels of precipitation favor moist microhabitats and promote its growth and development (52, 53). The prevalence and infection intensity of chytridiomycosis are significantly correlated with temperature and rainfall in eastern Australia (54, 55). High environmental humidity may have promoted the growth of Bd, which is associated with high disease severity (56). In order to prevent the spread of Bd between various regions, we suggest that the domestic and international trade in amphibians should follow the veterinary supervision and quarantine guidelines issued by the IUCN and the OIE (43). According to the environmental conditions of specific locations and habitats, comprehensively evaluate the distribution, prevalence and infection intensity of Bd, and formulate more effective disease management strategies. In high-prevalence areas of Bd (more at risk compared to those in low-prevalence areas), more precautionary measures should be taken. These measures could include 1) closely stricter monitoring (increased intensity of supervisory personnel) the health of amphibians entering and exiting these areas and 2) continuously check the prevalence of Bd in amphibian habitats in these areas. If Bd is identified, fungicides could be used to control its spread (57).
For the seasonal subgroup, amphibians had the highest prevalence in spring (Table 1). The occurrence of chytridiomycosis has obvious seasonality (58). The fungal prevalence and infection intensity decreased from spring to summer within a population of frogs, which can clear fungal infections later in summer and at higher temperatures (59, 60). Bd is usually not randomly distributed in amphibian communities (47, 61), but rather is closely related to changes in microclimatic conditions. Therefore, we analyzed the most suitable conditions for Bd growth in combination with temperature and rainfall subgroups. Our results showed that amphibians were more susceptible to Bd infections at a temperature of 17–25°C (P < 0.05) and in rainfall of >150 mm (Table 2). On large geographic scales, temperatures and rainfall are the most fatal environmental factors affecting the occurrence of fungi, increasing the possibility of infection and transmission (10, 62). In view of the environmental scope of the above areas, we recommend multiple methods to control the spread of diseases such as Bd. Chemical methods are used in small epidemic areas, such as antifungal itraconazole, as well as environmental disinfectants (16, 63). Bio-augmentation methods have been proposed for more complex ecological environment, such as enhancing the amphibian cutaneous probiotic microorganisms (which can inhibit the growth of Bd) and vaccination to prevent fungal invasion (64).
In the 111 articles surveyed here, real-time PCR methods had a higher Bd prevalence (Table 1). Real-time PCR is more sensitive than traditional PCR and histopathology examination, with a higher and faster detection rate, making the results more objective rather than subjective (65). It can accurately detect Bd infection intensity in degraded samples or the low levels of organisms, increasing the detection of prevalence, and it is suitable for wildlife, laboratory animals, and the food trade as well as environmental samples (66). Therefore, most researchers prefer to use the real-time PCR method for the detection of Bd as early as possible. In addition, there are articles using nested PCR and Loop-mediated isothermal amplification (LAMP). Nested PCR and LAMP hhhimprove the sensitivity of traditional PCR, but are more prone to contamination (67, 68). Consequently, they have been rarely widely used. In fact, the use of appropriate detection methods is vital for helping relevant personnel obtain accurate testing data and better understand the epidemic status of a disease.
Among Bd infections in amphibians of different ages, the point estimate of juveniles was higher than the other three age ranges, but this was not significant (P = 0.0528; Table 1). The susceptibility to Bd was shown to vary at the different stages of life (61). The results have shown that the Bd prevalence in younger life stages was higher than in older life stages (larvae: 18.7%, juveniles: 40%, sub-adults: 5.5%; adults: 6.6%), and that juvenile stages had the highest prevalence of Bd (69). As frogs get older, Bd infection can be cleared (70). Fungi have mobile zoospores that have the ability to spread rapidly in water, which might promote tadpole infections (71), but tadpoles rarely died from these infections (51, 72). Thus, tadpoles might be the reservoirs for this disease and increase the activity of this Bd in the environment, leading to the infection of adult specimens (27, 73). Therefore, we recommend that relevant personnel pay attention to the detection Bd of tadpoles and juvenile amphibians, and control the fungal loads in environments where tadpoles live to reduce the prevalence.
Water sources can be one of the environmental reservoirs of Bd, because it can survive in water alone for several weeks (8, 71, 74). In a subgroup analysis of sample sources, the Bd prevalence of amphibians inhabiting streams was shown to be higher than in other places (Table 1). However, species living on roads had the lowest prevalence. Therefore, especially cooler and shaded lentic streams might be more conducive to Bd transmission among amphibians (75, 76). Thus, we analyzed the subgroups of different hosts, and frogs had the highest prevalence (P < 0.05; Table 1). There was a great difference in the degree of Bd infection between anurans (frogs and toads) and caudates (salamanders and newts) (72). According to statistical data, 36% of caudates are terrestrial, while only 1.7% of anurans are terrestrial (77). Therefore, species living longer in water were more susceptible to infection (78). In the light of the IUCN classification of amphibians, the prevalence of Bd in vulnerable species was the highest at 43.08% (Table 1). Many species were listed as threatened by the IUCN, because they were infected with Bd resulting in population declines (79). Therefore, we propose strengthening the protective measures for those habitats that are suitable for Bd growth, such as manipulating the habitat environment for temperature or humidity and for endangered species, captive breeding may be established to ensure populations.
In our research, there were 20 medium-quality articles and 91 high-quality articles (Table 1). In the medium quality articles, we found that some of them did not have random sampling, definite sampling time, sampling location or clear detection methods (Supplementary Table 4). Since these medium-quality articles did not provide detailed data, the potential risk factors of Bd infection cannot be accurately measured, leading to biased results. Thus, we recommend that researchers should find as many factors as possible influencing fungal infection, so as to more effectively prevent these diseases and lay scientific groundwork for future research work.
This study has many advantages, such as its rich sample size, comprehensive coverage, clear method of analyses and wide range of risk factor assessments, which provides many evidence-based substantive suggestions for combating Bd spread. However, this study has also some shortcomings. First, although corresponding search formula is applied to collect articles, all relevant studies could not be fully found. Second, there was no information about the susceptibility of a species to Bd infection, it could be a potentially important risk factor for the implementation of controls. Third, only 13 out of 111 studies included in the investigation reported explicitly using random sampling, but others could not be ruled out the possibility of random sampling, which could result in a slight publication bias in the funnel plots and Egger's test, even through no publication bias was detected in this study. Therefore, the future study design still need to be comprehensive, such as clearly disclosing their sampling details. However, we are convinced that our report can reflect the situation of chytridiomycosis spread worldwide.
Chytridiomycosis has a wide distribution range and affects various amphibian species at all ages. In the last decade, the prevalence of Bd has shown a decreasing trend, and temperature and elevation have significantly affected it. In addition, the use of real-time PCR methods could help researchers obtain a more accurate estimate of Bd prevalence in amphibians. Meanwhile, we thus should focus on highly sensitive species and those listed as threatened by the IUCN. Therefore, we suggest that governments should formulate a reasonable management based on different geographical conditions, habitat types and species status. This study helps further elucidate potential risk factors for Bd infection and also provides a theoretical basis for related personnel to prevent and control chytridiomycosis transmission.
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding authors.
ZL performed the data extraction and drafted the manuscript. ZL and QW analyzed the data. KS and JF developed the paper concept, designed, supervised, and revised the manuscript. All authors contributed to the article and approved the submitted version.
This work was supported by the National Natural Science Foundation of China (grant numbers 31961123001, 32171525, and 31770403).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We thanked Dr. Qinglong Gong for his advice on the design and writing of the article.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fvets.2021.791237/full#supplementary-material
1. Collins JP. Amphibian decline and extinction: what we know and what we need to learn. Dis Aquat Organ. (2010) 92:93–9. doi: 10.3354/dao02307
2. Stuart SN. Status and trends of amphibian declines and extinctions worldwide. Science. (2004) 306:1783–6. doi: 10.1126/science.1103538
3. Becker CG, Fonseca CR, Haddad CFB, Batista RF, Prado PI. Habitat split and the global decline of amphibians. Science. (2007) 318:1775–7. doi: 10.1126/science.1149374
4. Hartel T, Schweiger O, Öllerer K, Cogalniceanu D, Arntzen JW. Amphibian distribution in a traditionally managed rural landscape of eastern Europe: probing the effect of landscape composition. Biol Conserv. (2010) 143:1118–24. doi: 10.1016/j.biocon.2010.02.006
5. Pounds JA, Bustamante MR, Coloma LA, Consuegra JA, Fogden MPL, Foster PN, et al. Widespread amphibian extinctions from epidemic disease driven by global warming. Nature. (2006) 439:161–7. doi: 10.1038/nature04246
6. Kolby JE, Daszak P. The emerging amphibian fungal disease, chytridiomycosis: a key example of the global phenomenon of wildlife emerging infectious diseases. Microbiol Spectr. (2016) 4:4.3.11. doi: 10.1128/microbiolspec.EI10-0004-2015
7. Berger L, Speare R, Daszak P, Green DE, Cunningham AA, Goggin CL, et al. Chytridiomycosis causes amphibian mortality associated with population declines in the rain forests of Australia and Central America. Proc Natl Acad Sci USA. (1998) 95:9031–6. doi: 10.1073/pnas.95.15.9031
8. Kilpatrick AM, Briggs CJ, Daszak P. The ecology and impact of chytridiomycosis: an emerging disease of amphibians. Trends Ecol Evol. (2010) 25:109–18. doi: 10.1016/j.tree.2009.07.011
9. Voyles J, Young S, Berger L, Campbell C, Voyles WF, Dinudom A, et al. Pathogenesis of chytridiomycosis, a cause of catastrophic amphibian declines. Science. (2009) 326:582–5. doi: 10.1126/science.1176765
10. Lips KR. Overview of chytrid emergence and impacts on amphibians. Philos Trans R Soc B. (2016) 371:20150465. doi: 10.1098/rstb.2015.0465
11. Weldon C, du Preez LH, Hyatt AD, Muller R, Spears R. Origin of the amphibian chytrid fungus. Emerg Infect Dis. (2004) 10:2100–5. doi: 10.3201/eid1012.030804
12. Schloegel LM, Hero JM, Berger L, Speare R, McDonald K, Daszak P. The decline of the sharp-snouted day frog (Taudactylus acutirostris): the first documented case of extinction by infection in a free-ranging wildlife species? Ecohealth. (2006) 3:35–40. doi: 10.1007/s10393-005-0012-6
13. Lips KR, Brem F, Brenes R, Reeve JD, Alford RA, Voyles J, et al. Emerging infectious disease and the loss of biodiversity in a Neotropical amphibian community. Proc Natl Acad Sci USA. (2006) 103:3165–70. doi: 10.1073/pnas.0506889103
14. Marca EL, Lips KR, Lötters S, Puschendorf R, Ibáñez R, Rueda-Almonacid JV, et al. Catastrophic population declines and extinctions in Neotropical harlequin frogs (Bufonidae: Atelopus). Biotropica. (2005) 37:190–201. doi: 10.1111/j.1744-7429.2005.00026.x
15. Schloegel LM, Daszak P, Cunningham AA, Speare R, Hill B. Two amphibian diseases, chytridiomycosis and ranaviral disease, are now globally notifiable to the World Organization for Animal Health (OIE): an assessment. Dis Aquat Organ. (2010) 92:101–8. doi: 10.3354/dao02140
16. Bosch J, Sanchez-Tomé E, Fernández-Loras A, Oliver JA, Fisher MC, Garner TW. Successful elimination of a lethal wildlife infectious disease in nature. Biol Lett. (2015) 11:20150874. doi: 10.1098/rsbl.2015.0874
17. Brannelly LA, McCallum HI, Grogan LF, Briggs CJ, Ribas MP, Hollanders M, et al. Kilpatrick, AM. Mechanisms underlying host persistence following amphibian disease emergence determine appropriate management strategies. Ecol Lett. (2021) 24:130–48. doi: 10.1111/ele.13621
18. Valencia-Aguilar A, Ruano-Fajardo G, Lambertini C, da Silva Leite D, Toledo LF, Mott T. Chytrid fungus acts as a generalist pathogen infecting species-rich amphibian families in Brazilian rainforests. Dis Aquat Organ. (2015) 114:61–7. doi: 10.3354/dao02845
19. Becker CG, Rodriguez D, Lambertini C, Toledo LF, Haddad CFB. Historical dynamics of Batrachochytrium dendrobatidis in Amazonia. Ecography. (2016) 39:954–60. doi: 10.1111/ecog.02055
20. Alvarado-Rybak M, Lepe-Lopez M, Peñafiel-Ricaurte A, Valenzuela-Sánchez A, Valdivia C, Mardones FO, et al. Bioclimatic and anthropogenic variables shape the occurrence of Batrachochytrium dendrobatidis over a large latitudinal gradient. Sci Rep. (2021) 11:1–12. doi: 10.1038/s41598-021-96535-w
21. Longo AV, Zamudio KR. Temperature variation, bacterial diversity and fungal infection dynamics in the amphibian skin. Mol Ecol. (2017) 226:4787–97. doi: 10.1111/mec.14220
22. Gervasi SS, Hunt EG, Lowry M, Blaustein AR. Temporal patterns in immunity, infection load and disease susceptibility: understanding the drivers of host responses in the amphibian-chytrid fungus system. Funct Ecol. (2014) 28:569–78. doi: 10.1111/1365-2435.12194
23. Bataille A, Fong JJ, Cha M, Wogan GOU, Baek HJ, Lee H, et al. Genetic evidence for a high diversity and wide distribution of endemic strains of the pathogenic chytrid fungus Batrachochytrium dendrobatidis in wild Asian amphibians. Mol Ecol. (2013) 22:4196–209. doi: 10.1111/mec.12385
24. Farrer RA, Weinert LA, Bielby J, Garner TW, Balloux F, Clare F, et al. Multiple emergences of genetically diverse amphibian-infecting chytrids include a globalized hypervirulent recombinant lineage. Proc Natl Acad Sci USA. (2011) 108:18732–6. doi: 10.1073/pnas.1111915108
25. Becker CG, Greenspan SE, Tracy KE, Dash JA, Lambertini C, Jenkinson TS, et al. Variation in phenotype and virulence among enzootic and panzootic amphibian chytrid lineages. Fungal Ecol. (2017) 26:45e50. doi: 10.1016/j.funeco.2016.11.007
26. Ruggeri J, Longo AV, Gaiarsa MP, Alencar LR, Lambertini C, Leite DS. Carvalho-e-Silva SP, Zamudio KR, Toledo LF, Martins M. Seasonal variation in population abundance and chytrid infection in stream-dwelling frogs of the Brazilian Atlantic Forest. PLoS ONE. (2015) 10:e0130554. doi: 10.1371/journal.pone.0130554
27. Valencia-Aguilar A, Toledo LF, Vital MVC, Mott T. Seasonality, environmental factors, and host behavior linked to disease risk in stream-dwelling tadpoles. Herpetologica. (2016) 72:98–106. doi: 10.1655/HERPETOLOGICA-D-15-00013
28. Bolochio BE, Lescano JN, Cordier JM, Loyola R, Nori J. A functional perspective for global amphibian conservation. Biol Conserv. (2020) 245:108572. doi: 10.1016/j.biocon.2020.108572
29. Mohneke M, Onadeko AB, Rödel MO. Medicinal and dietary uses of amphibians in Burkina Faso. Afr J Herpetol. (2011) 60:78–83. doi: 10.1080/21564574.2011.564660
30. O'Rourke DP. Amphibians used in research and teaching. ILAR J. (2007) 48:183–7. doi: 10.1093/ilar.48.3.183
31. Moher D, Liberati A, Tetzlaff J, Altman DG, PRISMA Group. Preferred reporting items for systematic reviews and meta-analyses: the PRISMA statement. PLoS Med. (2009) 6:e1000097. doi: 10.1371/journal.pmed.1000097
32. Skerratt LF, Berger L, Hines HB, McDonald KR, Mendez D, Speare R. Survey protocol for detecting chytridiomycosis in all Australian frog populations. Dis Aquat Organ. (2008) 80:85–94. doi: 10.3354/dao01923
33. Guyatt GH, Oxman AD, Vist GE, Kunz R, Falck-Ytter Y, Alonso-Coello P, et al. GRADE Working Group. GRADE: an emerging consensus on rating quality of evidence and strength of recommendations. BMJ. (2008) 336:924–6. doi: 10.1136/bmj.39489.470347.AD
34. Wei XY, Gong QL, Zeng A, Wang W, Wang Q, Zhang XX. Seroprevalence and risk factors of Toxoplasma gondii infection in goats in China from 2010 to 2020: a systematic review and meta-analysis. Prev Vet Med. (2021) 186:105230. doi: 10.1016/j.prevetmed.2020.105230
35. R Core Team. R: A Language Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing (2020). Available online at: https://www.R-project.org/
36. Gong QL, Wang Q, Yang XY Li DL, Du R. Seroprevalence and risk factors of the bluetongue virus in cattle in China from 1988 to 2019: a comprehensive literature review and meta-analysis. Front Vet Sci. (2021) 7:550381. doi: 10.3389/fvets.2020.550381
37. Barendregt JJ, Doi SA, Lee YY, Norman RE, Vos T. Meta-analysis of prevalence. J Epidemiol Community Health. (2013) 67:974–8. doi: 10.1136/jech-2013-203104
38. Assefa A, Bihon A. Bovine cysticercosis in Ethiopia: a systematic review and meta-analysis of prevalence from abattoir-based surveys. Prev Vet Med. (2019) 169:104707. doi: 10.1016/j.prevetmed.2019.104707
39. Wang ZD, Wang SC, Liu HH, Ma HY Li ZY, Wei F, Zhu XQ, et al. Prevalence and burden of Toxoplasma gondii infection in HIV-infected people: a systematic review and meta-analysis. Lancet HIV. (2017) 4:e177–88. doi: 10.1016/S2352-3018(17)30005-X
40. Ding H, Gao YM, Deng Y, Lamberton PH, Lu DB. A systematic review and meta-analysis of the seroprevalence of Toxoplasma gondii in cats in mainland China. Parasite Vector. (2017) 10:27. doi: 10.1186/s13071-017-1970-6
41. Wang W, Gong QL, Zeng A, Li MH, Zhao Q, Ni HB. Prevalence of Cryptosporidium in pigs in China: a systematic review and meta-analysis. Transbound Emerg Dis. (2021) 68:1400–13. doi: 10.1111/tbed.13806
42. Kriger KM, Hero JM. Chytridiomycosis, amphibian extinctions, and lessons for the prevention of future panzootics. Ecohealth. (2009) 6:6–10. doi: 10.1007/s10393-009-0228-y
43. Fisher MC, Garner TW, Walker SF. Global emergence of Batrachochytrium dendrobatidis and amphibian chytridiomycosis in space, time, and host. Annu Rev Microbiol. (2009) 63:291–310. doi: 10.1146/annurev.micro.091208.073435
44. Murray KA, Skerratt LF, Speare R, McCallum H. Impact and dynamics of disease in species threatened by the amphibian chytrid fungus, Batrachochytrium dendrobatidis. Conserv Biol. (2009) 32:1242–52. doi: 10.1111/j.1523-1739.2009.01211.x
45. Bell RC, Gata Garcia AV, Stuart BL, Zamudio KR. High prevalence of the amphibian chytrid pathogen in Gabon. Ecohealth. (2011) 8:116–20. doi: 10.1007/s10393-010-0364-4
46. Morehouse EA, James TY, Ganley AR, Vilgalys R, Berger L, Murphy PJ, et al. Multilocus sequence typing suggests the chytrid pathogen of amphibians is a recently emerged clone. Mol Ecol. (2003) 12:395–403. doi: 10.1046/j.1365-294X.2003.01732.x
47. Brem FMR, Lips KR. Batrachochytrium dendrobatidis infection patterns among Panamanian amphibian species, habitats, and elevations during epizootic and enzootic stages. Dis Aquat Organ. (2008) 81:189–202. doi: 10.3354/dao01960
48. Kriger KM, Hero JM. Altitudinal distribution of chytrid (Batrachochytrium dendrobatidis) infection in subtropical Australian frogs. Austral Ecol. (2008) 33:1022–32. doi: 10.1111/j.1442-9993.2008.01872.x
49. Young BE, Lips KR, Reaser JK, Ibáñez R, Salas AW, Cedeño JR, et al. Population declines and priorities for amphibian conservation in Latin America. Conserv Biol. (2001) 15:1213–23. doi: 10.1046/j.1523-1739.2001.00218.x
50. Farthing HN, Jiang J, Henwood AJ, Fenton A, Garner TWJ, Daversa DR, et al. Microbial grazers may aid in controlling infections caused by the aquatic zoosporic fungus Batrachochytrium dendrobatidis. Front Microbiol. (2021) 11:592286. doi: 10.3389/fmicb.2020.592286
51. Johnson ML, Speare R. Possible modes of dissemination of the amphibian chytrid Batrachochytrium dendrobatidis in the environment. Dis Aquat Organ. (2005) 65:181–6. doi: 10.3354/dao065181
52. Longo AV, Burrowes PA, Joglar RL. Seasonality of Batrachochytrium dendrobatidis infection in direct-developing frogs suggests a mechanism for persistence. Dis Aquat Organ. (2010) 92:253–60. doi: 10.3354/dao02054
53. Whitfield SM, Kerby J, Gentry LR, Donnelly MA. Temporal variation in infection prevalence by the amphibian chytrid fungus in three species of frogs at La Selva, Costa Rica. Biotropica. (2012) 44:779–84. doi: 10.1111/j.1744-7429.2012.00872.x
54. Drew A, Allen EJ, Allen LJ. Analysis of climatic and geographic factors affecting the presence of chytridiomycosis in Australia. Dis Aquat Organ. (2006) 68:245–50. doi: 10.3354/dao068245
55. Kriger KM, Hero JM. Large scale seasonal variation in the prevalence and severity of chytridiomycosis. J Zool. (2007) 271:352–9. doi: 10.1111/j.1469-7998.2006.00220.x
56. Narváez-Narváez DA, Cabrera-Andrade A, Merino-Viteri A. Paz-Y-Miño C, Burgos G, Genoy-Puerto A. Infection dynamics of Batrachochytrium dendrobatidis in two frog species inhabiting Quito' Metropolitan Guangüiltagua park, Ecuador. J Wildl Dis. (2021) 57:749–60. doi: 10.7589/JWD-D-20-00110
57. Jiménez RR, Alvarado G, Ruepert C, Ballestero E, Sommer S. The fungicide chlorothalonil changes the amphibian skin microbiome: a potential factor disrupting a host disease-protective trait. Appl Microbiol. (2021) 1:26–37. doi: 10.3390/applmicrobiol1010004
58. Berger L, Speare R, Hines HB, Marantelli G, Hyatt AD, McDonald KR, et al. Effect of season and temperature on mortality in amphibians due to chytridiomycosis. Aust Vet J. (2004) 82:434–9. doi: 10.1111/j.1751-0813.2004.tb11137.x
59. Kinney VC, Heemeyer JL, Pessier AP, Lannoo MJ. Seasonal pattern of Batrachochytrium dendrobatidis infection and mortality in Lithobates areolatus: affirmation of Vredenburg's “10,000 zoospore rule.” PLoS ONE. (2011) 6:e16708. doi: 10.1371/journal.pone.0016708
60. Woodhams DC, Alford RA, Marantelli G. Emerging disease of amphibians cured by elevated body temperature. Dis Aquat Organ. (2003) 55:65–7. doi: 10.3354/dao055065
61. Rachowicz LJ, Vredenburg VT. Transmission of Batrachochytrium dendrobatidis within and between amphibian life stages. Dis Aquat Organ. (2004) 61:75–83. doi: 10.3354/dao061075
62. Olson DH, Aanensen DM, Ronnenberg KL, Powell CI, Walker SF, Bielby J, et al. Mapping the global emergence of Batrachochytrium dendrobatidis, the amphibian chytrid fungus. PLoS ONE. (2013) 8:e56802. doi: 10.1371/journal.pone.0056802
63. Garner TWJ, Garcia G, Carroll B, Fisher MC. Using itraconazole to clear Batrachochytrium dendrobatidis infection and subsequent depigmentation of Alytes muletensis tadpoles. Dis Aquat Organ. (2009) 83:257–60. doi: 10.3354/dao02008
64. Cassone A. Fungal vaccines: real progress from real challenges. Lancet Infect Dis. (2008) 8:114–24. doi: 10.1016/S1473-3099(08)70016-1
65. Hyatt AD, Boyle DG, Olsen V, Boyle DB, Berger L, Obendorf D, et al. Diagnostic assays and sampling protocols for the detection of Batrachochytrium dendrobatidis. Dis Aquat Organ. (2007) 73:175–92. doi: 10.3354/dao073175
66. Boyle DG, Boyle DB, Olsen V, Morgan JA, Hyatt AD. Rapid quantitative detection of chytridiomycosis (Batrachochytrium dendrobatidis) in amphibian samples using real-time Taqman PCR assay. Dis Aquat Organ. (2004) 60:141–8. doi: 10.3354/dao060141
67. Rigotto C, Sincero TC, Simões CM, Barardi CR. Detection of adenoviruses in shellfish by means of conventional-PCR, nested-PCR, and integrated cell culture PCR (ICC/PCR). Water Res. (2005) 39:297–304. doi: 10.1016/j.watres.2004.10.005
68. Rostami A, Karanis P, Fallahi S. Advances in serological, imaging techniques and molecular diagnosis of Toxoplasma gondii infection. Infection. (2018) 46:303–15. doi: 10.1007/s15010-017-1111-3
69. Spitzen-van der Sluijs A, Canessa S, Martel A, Pasmans F. Fragile coexistence of a global chytrid pathogen with amphibian populations is mediated by environment and demography. Philos Trans R Soc B. (2017) 284:20171444. doi: 10.1098/rspb.2017.1444
70. Briggs CJ, Vredenburg VT, Knapp RA, Rachowicz LJ. Investigating the population-level effects of chytridiomycosis: an emerging infectious disease of amphibians. Ecology. (2005) 86:3149–59. doi: 10.1890/04-1428
71. Johnson ML, Speare R Survival of Batrachochytrium dendrobatidis in water: quarantine and control implications. Emerg Infect Dis. (2003) 9:922–25. doi: 10.3201/eid0908.030145
72. Blaustein AR, Romansic JM, Schessele EA, Han BA, Pessier AP, Longcore JE. Interspecific variation in susceptibility of frog tadpoles to the pathogenic fungus Batrachochytrium dendrobatidis. Conserv Biol. (2005) 19:1460–8. doi: 10.1111/j.1523-1739.2005.00195.x
73. Salla RF, Gamero FU, Ribeiro LR, Rizzi GM, Medico SE, Rissoli RZ, et al. Cardiac adaptations of bullfrog tadpoles in response to chytrid infection. J Exp Zool Part A. (2015) 323:487–96. doi: 10.1002/jez.1945
74. Catenazzi A, Von May R, Vredenburg VT. High prevalence of infection in tadpoles increases vulnerability to fungal pathogen in high-Andean amphibians. Biol Conserv. (2013) 159:413–21. doi: 10.1016/j.biocon.2012.11.023
75. Chestnut T, Anderson C, Popa R, Blaustein AR, Voytek M, Olson DH, et al. Heterogeneous occupancy and density estimates of the pathogenic fungus Batrachochytrium dendrobatidis in waters of North America. PLoS ONE. (2014) 9:e106790. doi: 10.1371/journal.pone.0106790
76. Rohr JR, Raffel TR. Linking global climate and temperature variability to widespread amphibian declines putatively caused by disease. Proc Natl Acad Sci USA. (2010) 107:8269–74. doi: 10.1073/pnas.0912883107
77. Bancroft BA, Han BA, Searle CL, Biga LM, Olson DH, Kats LB, et al. Species-level correlates of susceptibility to the pathogenic amphibian fungus Batrachochytrium dendrobatidis in the United States. Biodivers Conserv. (2011) 20:1911–20. doi: 10.1007/s10531-011-0066-4
78. Greenspan SE, Calhoun AJ, Longcore JE, Levy MG. Transmission of Batrachochytrium dendrobatidis to wood frogs (Lithobates sylvaticus) via a bullfrog (L. catesbeianus) vector. J Wildlife Dis. (2012) 48:575–82. doi: 10.7589/0090-3558-48.3.575
Keywords: Batrachochytrium dendrobatidis, meta-analysis, amphibians, chytridiomycosis, prevalence
Citation: Li Z, Wang Q, Sun K and Feng J (2021) Prevalence of Batrachochytrium dendrobatidis in Amphibians From 2000 to 2021: A Global Systematic Review and Meta-Analysis. Front. Vet. Sci. 8:791237. doi: 10.3389/fvets.2021.791237
Received: 08 October 2021; Accepted: 26 November 2021;
Published: 17 December 2021.
Edited by:
Brian D. Perry, University of Oxford, United KingdomReviewed by:
Gabriel Arriagada, Universidad de O'Higgins, ChileCopyright © 2021 Li, Wang, Sun and Feng. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Keping Sun, c3Vua3AxMjlAbmVudS5lZHUuY24=; Jiang Feng, ZmVuZ2pAbmVudS5lZHUuY24=
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.