 Attila D. Sándor
Attila D. Sándor Andrei Daniel Mihalca
Andrei Daniel Mihalca Cristian Domşa1
Cristian Domşa1- 1Department of Parasitology and Parasitic Diseases, University of Agricultural Sciences and Veterinary Medicine, Cluj-Napoca, Romania
- 2Department of Parasitology and Zoology, University of Veterinary Medicine, Budapest, Hungary
The soft ticks (Ixodida: Argasidae) are ectoparasites of terrestrial vertebrates with a wide geographic distribution, occurring on all continents. These ticks are obligate blood-feeders, most of them show high degrees of host-specialization and several species in arid and tropical regions are important parasites of livestock and men. Species commonly occurring on domestic animals and man are generally well-known, with many studies focusing on their ecology, distribution or vectorial role. However, wildlife-specialist soft ticks are less studied. Nearly half of all soft tick species are bat specialists, with five species (Carios vespertilionis, Chiropterargas boueti, Chiropterargas confusus, Reticulinasus salahi, and Secretargas transgariepinus) occurring in the Western Palearctic. There is no comprehensive study on the distribution, hosts or pathogens in these soft ticks, although most species were shown to carry several viral, bacterial, or protozoan pathogens and also to occasionally infest humans. Based on a literature survey and 1,120 distinct georeferenced records, we present here the geographical range, host selection and vectorial potential for bat-specialist soft ticks occurring in the Western Palearctic (chiefly Europe, North Africa, and the Middle East). Carios vespertilionis shows the largest distribution range and was found on most host species, being ubiquitous wherever crevice-roosting bats occur. All the other species were located only in areas with Mediterranean climate, with Ch. boueti, Chiropteraragas confusus, and R. salahi are missing entirely from Europe. These three species have a host spectrum of bats roosting primarily in caves, while S. transgariepinus and Ca. vespertilionis is feeding primarily on crevice-roosting bat species. All but one of these soft tick species are known to feed on humans and may be vectors of important disease agents (Rickettsia spp., Borrelia spp., Bartonella spp., Ehrlichia spp., Babesia spp., several nairo-, and flaviviruses). As several crevice-roosting bat species show a continuous adaptation to human-altered areas, with certain species becoming common city-dwellers in the Western Palearctic, the study of bat specialist soft ticks is also important from an epidemiologic point of view.
Introduction
Ticks (Ixodoidea) are obligate blood-feeding arthropods, with a wide-spread occurrence and ~935 species known as parasites of terrestrial vertebrates (1, 2). The group has an ancient origin, with their first appearance suggested for the Cretaceous [65–146 mya, (3)], and widespread diversification and dispersal occurring during the Tertiary some 5 to 65 mya ago (4). Taxonomically, modern ticks are divided into three families (Argasidae, Ixodidae, and Nuttalliellidae) with the two most speciose being the hard ticks (Ixodidae) with 742 recognized species (2), followed by soft ticks (Argasidae) with 193 species, as listed in the last comprehensive checklist of this later group (1). The Nuttalliellidae consist of a single species (Nuttalliella namaqua), and is considered to be the most ancient among the three tick families, showing several intermediate characters specific for the other two (5).
Argasidae includes two subfamilies, Argasinae and Ornithodorinae, both with several genera, and subgenera, with differing numbers according to different authors (1, 6–10). They have a world-wide distribution, with most species being distributed in the tropics and dry regions of the globe (6). Argasid ticks show diverse adaptation to using their hosts. Most members of the family are characterized by a single, prolonged larval blood feeding and multiple, short blood feeding events of subsequent developmental stages on several host individuals, however other adaptations (e.g., no larval feeding or lack of blood-feeding in adults, etc.) were recorded in certain species (11). By doing so, these ticks are capable of taking up pathogens (viral, bacterial, or protozoan) and transfering them to other hosts, thus they have important vectorial role (6). Most of soft ticks inhabit holes and crevices and have access to hosts only occasionally, hence they developed extreme adaptations to prolonged fasting and short feeding bouts whenever hosts are available (12). Their vectorial capacity for several important zoonotic diseases is well-known, including human relapsing fever (its causative agent transmitted by Ornithodoros spp.), tick-borne relapsing fevers (caused by several Borrelia spp. transmitted mainly by Ornithodoros and Argas spp.) or African swine fever (vectored by Ornithodoros moubata, Ornithodoros porcinus, Ornithodoros erraticus, or Ornithodoros savignyi) causing severe economic losses (6, 13).
Soft ticks have a special relationship with bats (Mammalia: Chiroptera). Bats are widely distributed, show high species diversity (being the second largest order of mammals) and several adaptations, which make them ideal host candidates for tick parasitism (14). Their morphological adaptations for flight hinders the range of their behavioral responses to reduce tick burden (e.g., their highly specialized limbs are inadequate for proper grooming), most species are social, spending their resting periods in dense groups and they are highly attached to their specific roosting sites, of which most are either underground (caves) or crevices in rocks or trees—excellent hiding places for soft ticks (11). Thus, several soft tick species-groups evolved specific associations with bat hosts. For example, all the known 17 species of the Nearctic soft tick genus Antricola (and Parantricola) are exclusive parasites of bats (15), together with all species belonging to the subgenus Carios, Chiropterargas, Nothoaspis, and Reticulinasus, and several other species from the genera Alectorobius and Ornithodoros (Supplementary Material and references therein). While most of these soft tick species are tropical in their distribution, there are at least five species which regularly occur on bats in the Western Palearctic. These species are Carios vespertilionis, Chiropterargas boueti, Chiropteraragas confusus, Reticulinasus salahi, and Secretargas transgariepinus. All these parasitize bats mainly roosting either inside caves (Ch. boueti, Chiropteraragas confusus, and R. salahi) or crevices (Ca. vespertilionis and S. transgariepinus).
Our knowledge on the distribution and ecology of bat-specialist soft tick species is scanty, as most of the literature only lists occurrence records or describe specific case reports, without a systematic review on their range, status and importance. Here, we collated the published records on these five soft tick species in the Western Palearctic, looking for data on their geographical distribution, host-parasite relationships and vectorial importance and also raising awareness on future challenges posed by some of these species on human health. In the wake of recent climate change events and urbanization trends in bats' distribution, we also intended to look for the abiotic (climate linked) and biotic (host distribution linked) factors regulating the distribution of bat specialist soft ticks in the Western Palearctic.
Materials and Methods
Database Creation
Our methodology followed a three-step algorithm. First a database search was performed, using keywords as: “soft ticks,” “bats,” “Argasidae,” and “Western Palearctic,” “Argas boueti,” “Argas confusus,” “Argas transgariepinus,” “Argas vespertilionis,” and “Ornithodoros salahi” in the following databases: Web of Science, Zoological Record, Google Scholar, and Global Biodiversity Information Facility (www.gbif.org). Subsequently, copies of the original publications were obtained and the references cited in these works were traced. This process was repeated until no new references were found. In the third step we extracted each individual host-tick record from the references, noting the location, date, host and parasite species, development stage (for ticks) and pathogen (if) mentioned. These records were introduced into a database and individually georeferenced to create distribution maps.
Distribution Maps
For the maps, we overlaid the different hosts' range with the presence data for each tick species. Each host range was set with transparency, so the more ranges overlap, the more intense the range color is—a proxy for multiple host species presence. For host ranges of main bat host species we used the freely available shape files from the website of the International Union for Conservation of Nature (IUCN) Red List (16). IUCN ranges were used previously primarily for conservation biology of bats (17) or other mammals (18), but also for establishing the relationships between bats, insect ectoparasites and their vectored pathogens (19). In the following step, we intersected the ranges with the contour of the Western Palearctic. Western Palearctic contour was delimited following the borders previously published (20, 21).
Host-Parasite Relationships
Using the database we mapped each host-parasite relationship and delimited the primary/accidental hosts. For deciding primary/accidental hosts of any soft tick species we used an arbitrary rule. Any bat species which held more than 5.0% of any specific soft tick's record is considered a primary host of the respective tick species, while hosts with <5.0 % of all cumulative records of a particular tick are considered accidental hosts, following a system previously proposed for bat-bat fly associations (22, 23).
Results
The complete database contains altogether 1,151 entries (4,856 individual ticks), collected from 899 hosts (4,378 ticks), together with a total of 65 cases of free ticks (involving 313 individuals), while collection circumstances were unknown for 156 cases (n = 165 ticks, only tick species and geographic location known). Altogether 44 bat species were recorded to host soft ticks, with most records noted for Ca. vespertilionis (Table 1). For a number of 16 cases the records mention only generic Chiroptera, while seven cases were assigned either to Myotis spp., Pipistrellus spp., or Plecotus spp. For 19 cases (1.9% of all records) the host is known, but it is not a bat species: 16 cases refer to humans, while one case each refer to a bird (Picus viridis), to a dog (Canis familiaris), while one to a rodent (Allactaga euphratica). Host species are listed in Tables 2, 3. Carios vespertilionis had the most diverse host spectrum, with altogether 42 different host species (6 primary and 36 secondary hosts), Ch. boueti had the most primary hosts (14), while R. salahi had a single primary host holding 87.7% of all records. Most ticks were recorded on crevice-dwelling bat species (76.6%), although for three species (Ch. boueti, Chiropteraragas confusus, and R. salahi) most primary bat hosts are cave-dwelling ones (24).
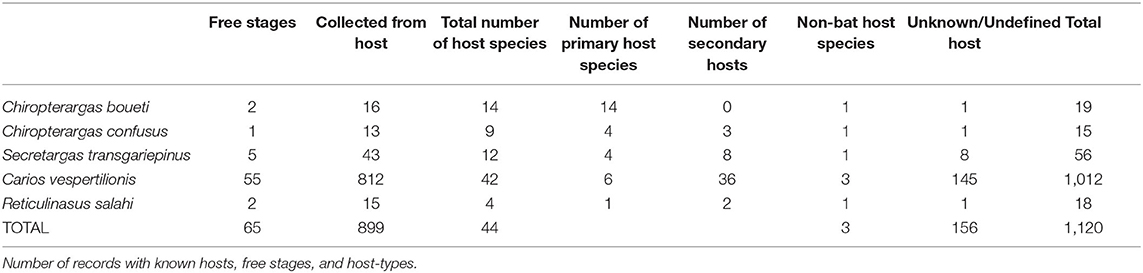
Table 1. Bat-specialist ticks recorded in the Western Palearctic.
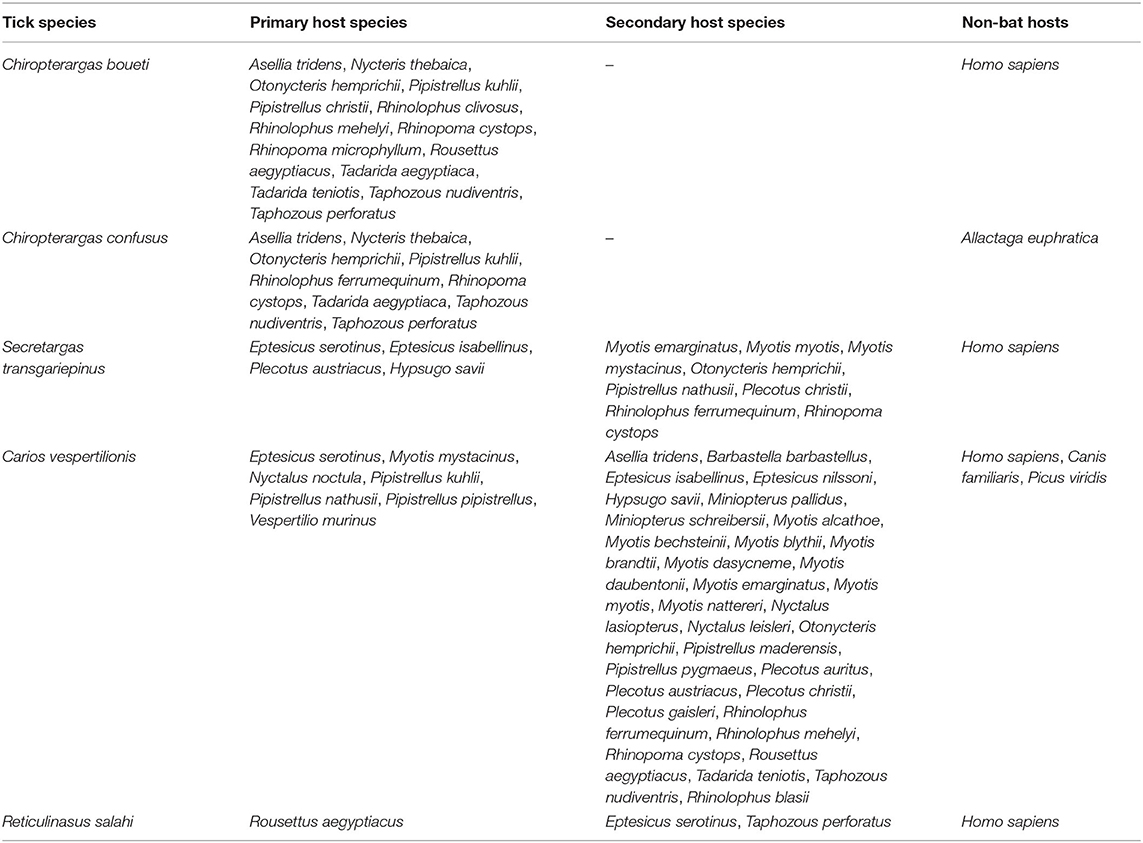
Table 2. Primary and secondary bat host species of soft ticks (Argasidae) in the Western Palearctic.
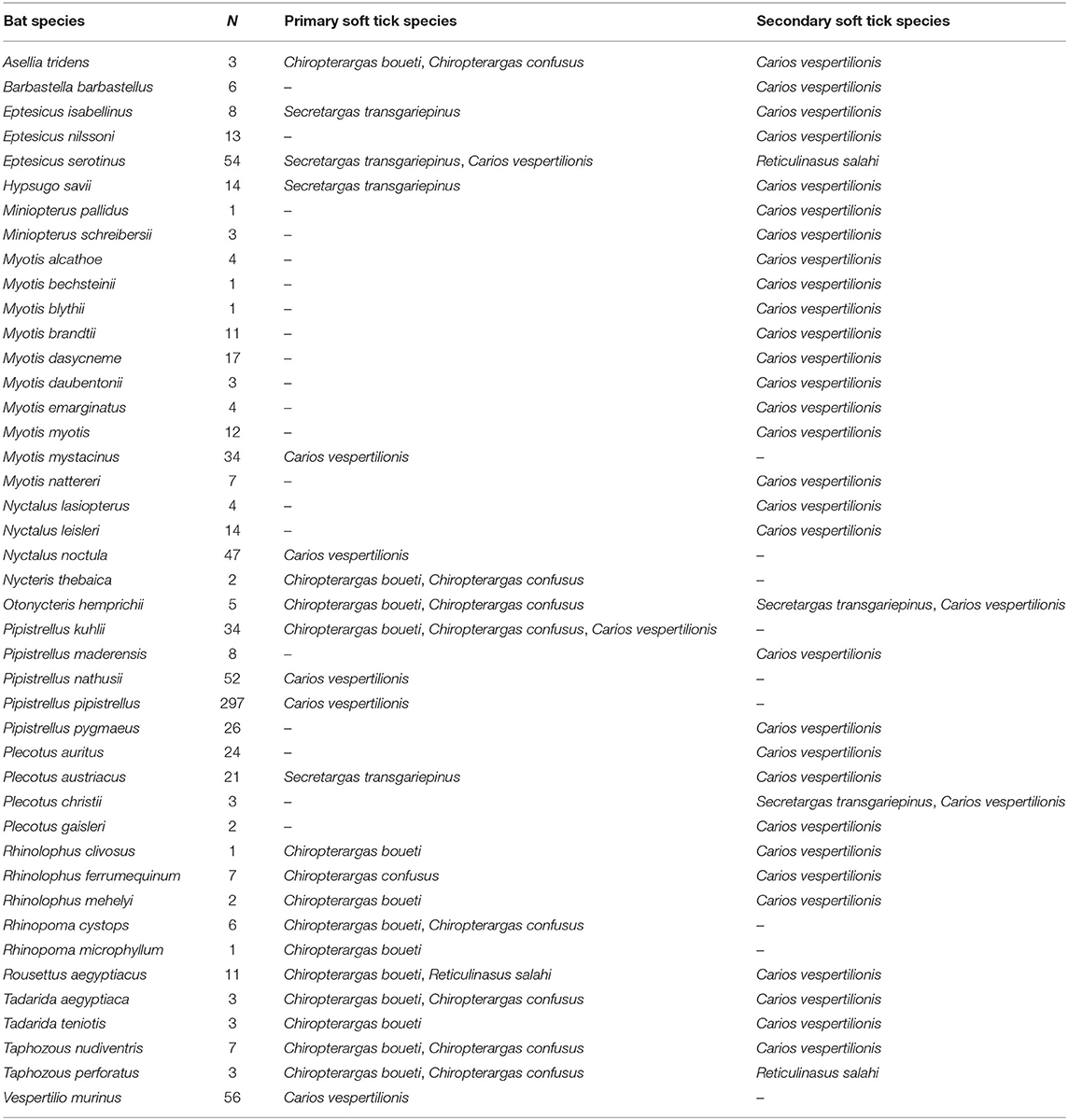
Table 3. List of bat species (Chiroptera) and their role as primary and secondary soft tick (Argasidae) hosts in the Western Palearctic (N, number of hosts with ticks).
Most tick records refer to subadult stages (only larvae being recorded on hosts, 93.13% of all ticks collected), with adults (males n = 25, females n = 67) and nymphs (n = 221) being collected from the environment or known bat roosts. Significantly more Ca. vespertilionis (mean intensity: 5.99 CI: 1.9–18.3) were collected from members of the genus Pipistrellus than from any other host species (x2 = 21.0216, p < 0.001).
Soft tick records showed a wide geographic distribution, covering most of the Western Palearctic, with significant differences between the extents of individual ranges. All five soft tick species show overlapping ranges in North Africa, most species (4/5) had a primarily Mediterranean range, with Ch. boueti, Chiropteraragas confusus, and R. salahi being exclusively found in North Africa and the Middle East (Figures 1, 2, 5). Carios vespertilionis and S. transgariepinus are distributed also in Europe (Figures 3, 4). Most records of soft ticks came from bats caught in (or in immediate vicinity of) man-made structures (buildings, ruins, and underground channels: 66%), with 13.6% being collected from caves. The rest were collected from bats caught in diverse habitats (roost unknown) while hosts were in active flight.
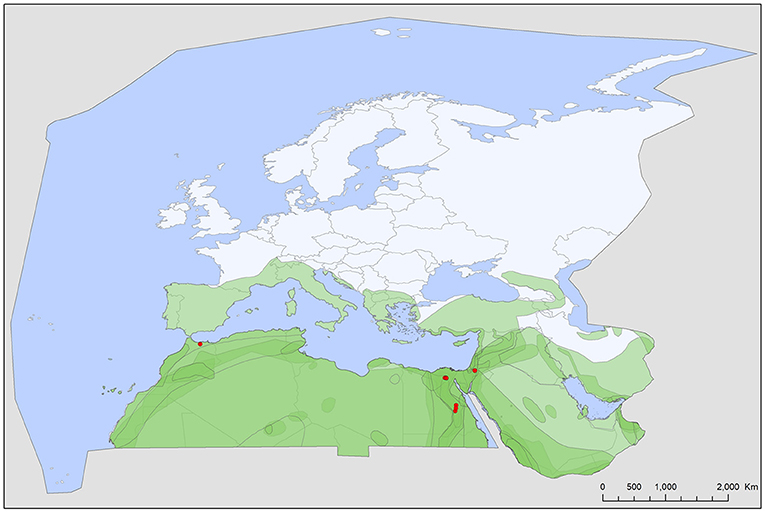
Figure 1. Geographic distribution of Chiropterargas boueti records in the Western Palearctic, overlayed to the geographic ranges for the 14 bat species studied as main hosts (Table 2) of this tick. Transparent layers were mapped on top of one another to highlight regions with dense range overlap. Some species have additional range overlap in Africa and Central and South Asia.
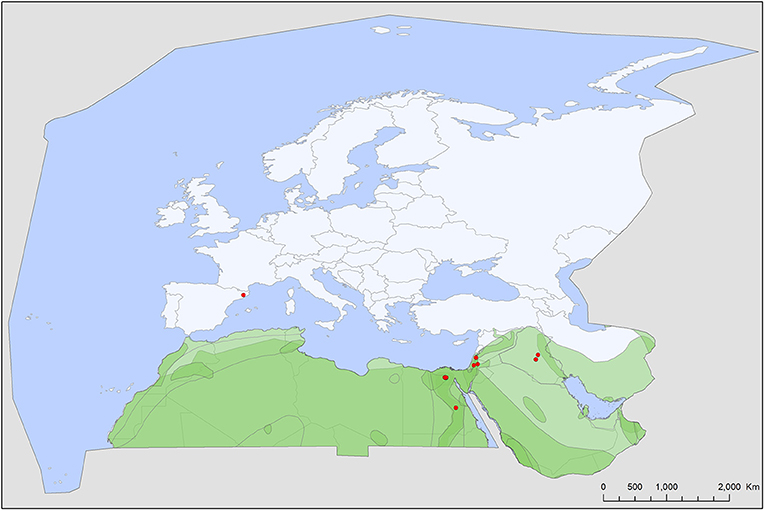
Figure 2. Geographic distribution of Chiropterargas confusus records in the Western Palearctic, overlayed to the geographic ranges for the nine bat species studied as main hosts (Table 2) of this tick. Transparent layers were mapped on top of one another to highlight regions with dense range overlap. Some species have additional range overlap in Africa and Central and South Asia.
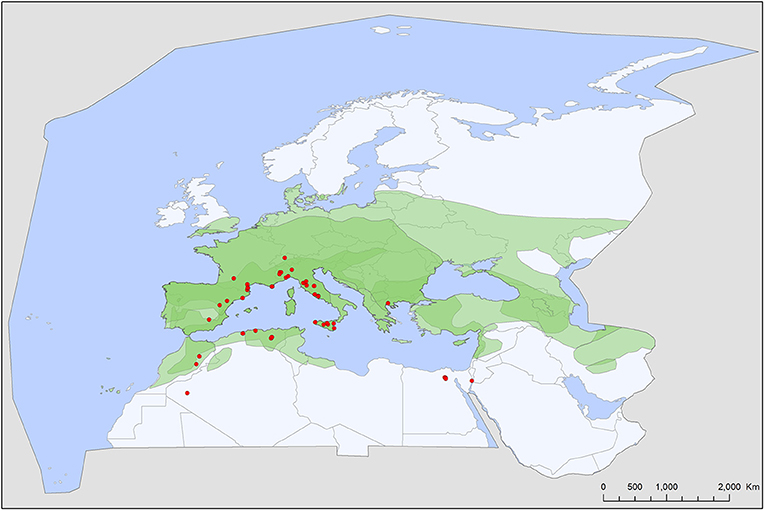
Figure 3. Geographic distribution of Secretargas transgariepinus records in the Western Palearctic, overlayed to the geographic ranges for the four bat species studied as main hosts (Table 2) of this tick. Transparent layers were mapped on top of one another to highlight regions with dense range overlap. Some species have additional range overlap in Africa and Central and South Asia.
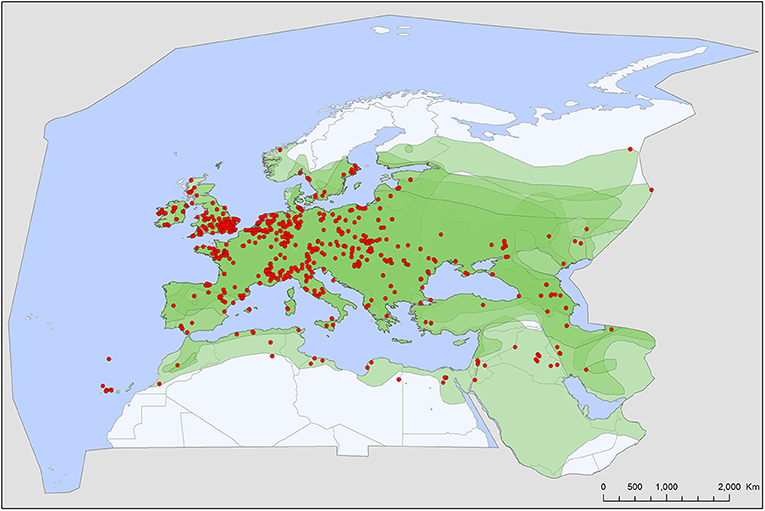
Figure 4. Geographic distribution of Carios vespertilionis records in the Western Palearctic, overlayed to the geographic ranges for the seven bat species studied as main hosts (Table 2) of this tick. Transparent layers were mapped on top of one another to highlight regions with dense range overlap. Some species have additional range overlap in Africa and Central and South Asia.
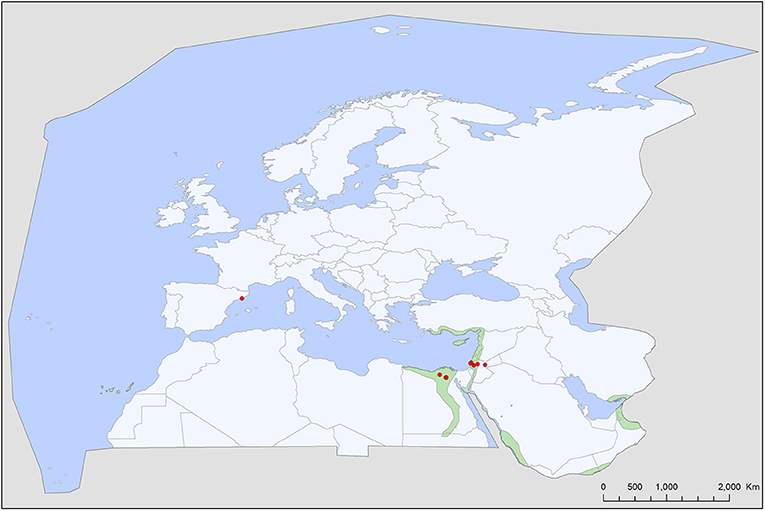
Figure 5. Geographic distribution of Reticulinasus salahi records in the Western Palearctic, overlayed to the geographic ranges for of Rousettus aegyptiacus, the sole primary host of this tick. R. aegyptiacus range extends into to the tropical and subtropical regions of Africa and Asia.
Several viral, bacterial, and piroplasmid pathogens were identified in two soft tick species of bats. The most common groups were bacteria (Bartonella spp., Borrelia spp., Coxiella burnetii, and Rickettsia spp.), but also five different viruses (belonging to flaviviruses and nairoviruses), as well two piroplasmids (Babesia spp.) were identified in soft ticks of bats (Table 4).

Table 4. List of bacterial, protozoan, and viral pathogens identified in bat specialist soft ticks.
Discussion
A total of five different soft tick species (Acari: Argasidae: Ca. vespertilionis, Ch. boueti, Ch. confusus, R. salahi, and S. transgariepinus) were recorded to be specialized to bats of the Western Palearctic. These ticks were found on 44 different bat species, showing diverse host-pattern (Figure 6). Most records came from a single tick species (Ca. vespertilionis, 88.7% of all records, Table 1), which not only has the highest number of host species, but also the widest distribution, covering the whole region of the Western Palearctic (Figure 4). Argasid ticks of bats primarily parasitize crevice dwelling host species, although there are three tick species (Ch. boueti, Chiropteraragas confusus, and R. salahi), for which most of the primary hosts are cave-dwelling bats. Soft tick occurrences showed a wide geographical distribution, covering most of the Western Palearctic. However, significant differences were found between the extent of individual ranges, with the range of three species being limited to North Africa and the Middle East. While overlapping areas are small, there is a region (northeastern part of Egypt and Israel) where all five species occur (Figures 1–5).
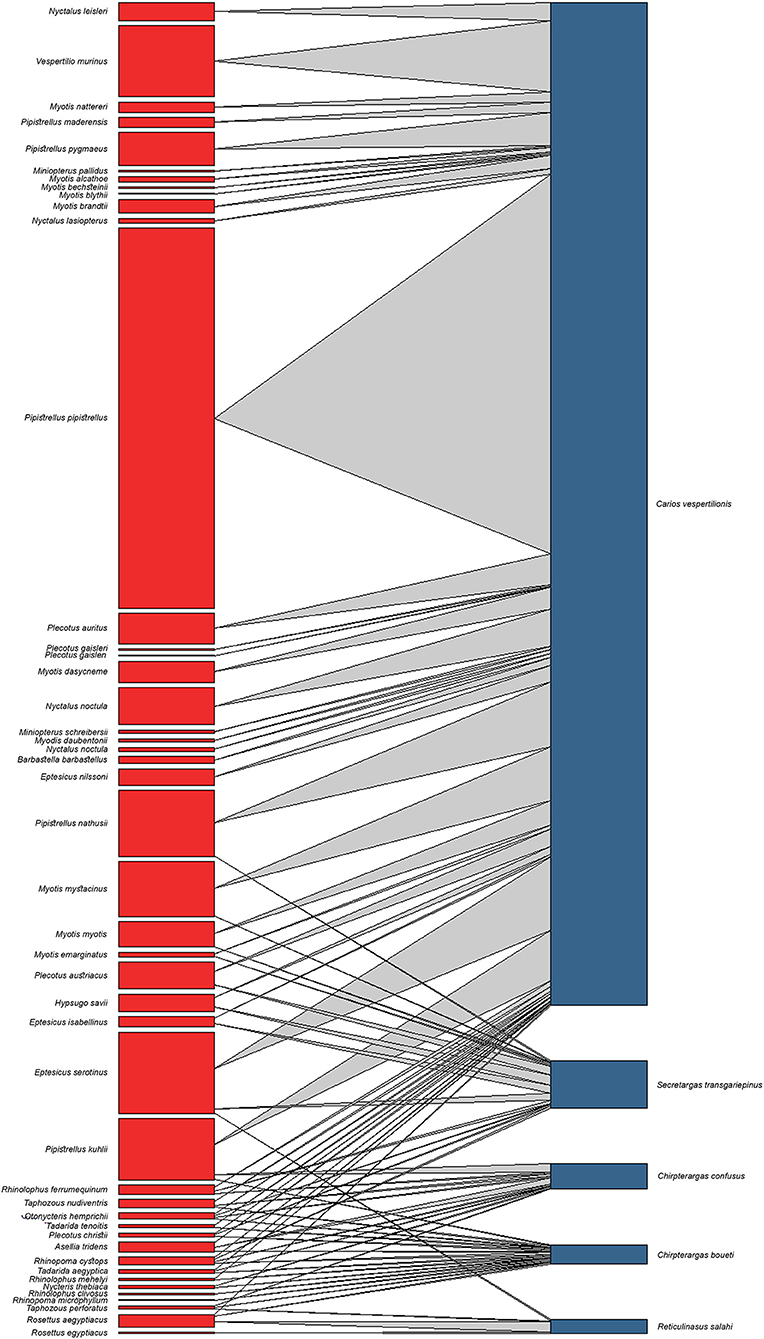
Figure 6. Quantitative interaction web based on bat specialist Argasidae ticks and their respective bat hosts. Links between nodes represent the sum of individual tick occurrences for a given bat species.
There is a considerable overlap between primary hosts among the different soft tick species. One bat species (Pipistrellus kuhlii) is the primary host for three different tick species, while further nine bat species regularly harbor two different argasid species (Table 2; Figure 6). Most tick species show a distribution that considerably overlaps with the range of their primary bat host (Figures 1, 2, 4, 5), with two notable exceptions. Secretargas transgariepinus shows a reduced range in comparison to its primary hosts' range, with several records in NE Africa, where primary hosts registered in the Western Palearctic do not occur (Figure 3). Records in this area came from bats exclusively distributed in Africa (Rhinopoma spp., Taphozous spp.), suggesting that on the African continent other primary hosts may occur. This species is well-known to regularly occur on bats performing large scale migrations like Pipistrellus spp. (24), hence several northern records may suggest accidental overshoots of argasid larvae collected from a bat in active migration (44). Another notable exception is the sole record of R. salahi in the Iberian Peninsula (Figure 5), far from the main range of its sole primary host, Rousettus aegyptius.
Specific Accounts
Chiropterargas boueti is a very poorly known species. Most information on this species was published in the original description (45), as well in its redescription (46). It has a wide distribution, primarily on the African continent, reaching Central and South Africa, with scattered records in Central Asia and the Middle East (47, 48). It is primarily a tick of cave dwelling tropical bats, with primary host species being Rhinopoma spp., with an extralimital occurrence in the Western Palearctic (Figure 1). Its ecology and vectorial capacity is unknown, while it is known to attack humans (46).
Chiropterargas confusus has a similar occurrence to the previous species, with which it shares most of its primary host species and also the occurrence records in the Western Palearctic (Figure 2). Its ecology and distribution are poorly known, with only a handful of records listed in Northern, Eastern and Southern Africa and Central Asia (46, 48, 49). In the Western Palearctic, this species has a narrow range, with records in NE Africa and the Middle East. There is no published information on its vectorial role. There is a putative record of its occurrence on a non-bat host (50), suggesting its suitability as a more generalist tick species.
Secretargas transgariepinus has a primarily tropical African distribution, with scattered records in North Africa and the Mediterranean region of Europe (51). It is primarily a parasite of crevice-dwelling bats, commonly occurring on Eptesicus spp., Hypsugo savii, and Plecotus spp. in the region (Tables 2, 3). The distribution of this argasid tick shows limited overlap with the range of its primary bat hosts in Europe, probably because its occurrence is limited by climatic factors (Figure 3). There is no clear seasonality in its records (Figure 7), and the apparent peak activity likely reflects an observation bias. The species is known for maternal care (52) and is a suspected vector (Table 4) for the Keterah virus (KTRO, nairoviruses) and spotted fever-causing bacteria of the genus Rickettsia (25–27). The species is regularly recorded on humans, with several cases known from Egypt and Italy (53).
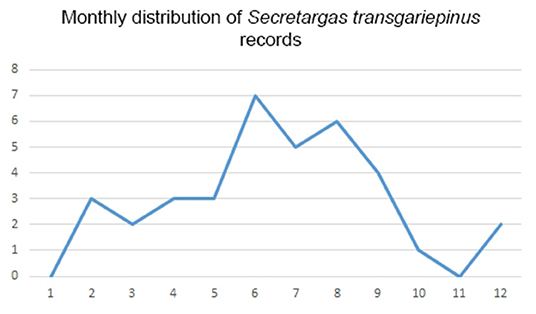
Figure 7. Monthly distribution of collection dates for Secretargas transgariepinus ticks.
Carios vespertilionis is the most common soft tick species of bats in the Western Palearctic (Table 1; Figure 4). It has the largest geographic distribution among bat ticks worldwide (54), with extensive morphological and genetic diversity along its wide range (36, 55). Its distribution mirrors the geographic range of the primary host species and it is the only soft tick species which may occur at the northernmost latitudes, wherever bats are present (Figure 4). It is also the species which has the highest number of records and known host species (Tables 1, 2). The species primarily occurs on crevice dwelling species (26 out of the 42 recorded host species, Table 2), with a particular affinity toward Pipistrellus spp., members of which usually host high number of individual ticks. These ticks may exert behavioral or even pathological impacts on their hosts (56), especially if they occur in high numbers (57, 58). While only larvae were recorded on hosts, roost sites (especially artificial ones) are important locations for adults, too (56, 59). This species was recorded in each month (Figure 8), and the seasonal distribution of records shows a summer peak. However, we suggest that this is mainly related to the timing of bat-research efforts in the field, rather than to a true activity peak of the ticks. Carios vespertilionis was recorded in multiple instances on humans (53, 60) and also on other vertebrates (Table 2) (61, 62). This species is known vector of several bacterial, protozoan and viral pathogens (Table 4 and references therein).
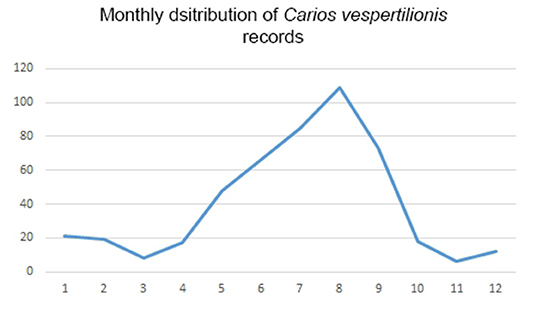
Figure 8. Monthly distribution of collection dates for Carios vespertilionis ticks.
Reticulinasus salahi is the host specialist tick of the Egyptian fruit bat, Rousettus aegyptiacus (63). It occurs in the Western Palearctic only where its primary host is present (north-east corner of Africa and the Middle East, but missing from Cyprus, Figure 5). It's single European record came from an accidental host (64). There is no information published on its vectorial capacity, although several cases are known when humans were infested by this argasid species (53, 63).
Apart of the species listed above, a few accidental records refer to several other Palearctic soft tick species that may also accidentally infest bats, as exemplified by two bird-specialists (Argas reflexus and Ornithodoros coniceps) and a rodent specialist (Ornithodoros tholozani) (65–68).
Two out of the five bat-specialist soft ticks recorded in the Western Palearctic have a wide range. These species (Ca. vespertilionis and S. transgariepinus) are parasites of crevice-dwelling species in the Western Palearctic and both have a wide palette of primary and accidental host species (Table 1; Figure 6). Their host species are small to middle sized insectivorous bats, which do not depend on the accessibility of large underground roost sites and regularly roost is small groups, actively seeking anthropogenic shelters (24). As these bat species (chiefly Pipistrellus spp., the group of small Myotis, Nyctalus noctula, Plecotus spp. and Eptesicus spp.) are feeding mainly on flying small moths and dipterans (24), they easily can find food and shelter even in the most urbanized areas of the region. Hence, it is not a surprise that these species show increase both in their range and populations. In addition, they are among the few bat species which became true urban dwellers (69). Especially large urban settings offer to these species not only hunting areas (70) and roost sites in the active period, but also suitable hibernating areas. During the last decades it has become an increasing trend for several such bat species to use large buildings (e.g., multistorey office buildings and block of flats) for autumn congregations or wintering sites in major cities (71). This tendency increased not only the number of these bats inside highly urbanized areas (72), but also the contacts with humans (73, 74). These bat species regularly harbor soft ticks (while their roosts offer habitat for adult ticks), and both Ca. vespertilionis and S. transgariepinus are known to be competent vectors for a series of viral, bacterial and protozoan pathogens (Table 4), some of which are zoonotic. While S. transgariepinus is currently a rare species in the Western Palearctic, whose range is seemingly limited by climatic factors, increasing temperatures in the near future may favor further range extension for this species, especially as its hosts will possibly have broader distribution. If these trends will continue in the near future, the increasing presence of bats and their soft ticks may pose a new epidemiologic challenge in highly urbanized areas.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author/s.
Author Contributions
AS, AM, and SH designed the study and acquired the budget. AS and ÁP screened the reference publications and built the database. CD analyzed the data and created the maps. AS wrote the manuscript. All authors performed critical revision and approved the final manuscript.
Funding
This research was supported by the by NKFIH-132794 (to SH and AS). ÁP was supported by the Collegium Talentum Programme of Hungary and by the NTP-NFTÖ-20 grant. While working on this project, AS was supported by the János Bolyai Research Scholarship of Hungarian Academy of Science and the ÚNKP 19-4-ÁTE-10 New National Excellence Program of the MIT.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank for help in gathering literature for Tamara Szentiványi and Mohamed L. Bendjeddou. The content of this manuscript has been published in part, as a chapter in the dissertation of ÁP.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fvets.2021.684737/full#supplementary-material
References
1. Guglielmone AA, Robbins RG, Apanaskevich DA, Petney TN, Estrada-Pena A, Horak IG, et al. The Argasidae, Ixodidae and Nuttalliellidae (Acari: Ixodida) of the world: a list of valid species names. Zootaxa. (2010) 2528:1–28. doi: 10.11646/zootaxa.2528.1.1
2. Guglielmone AA, Petney TN, Robbins RG. Ixodidae (Acari: Ixodoidea): descriptions and redescriptions of all known species from 1758 to december 31, 2019. Zootaxa. (2020) 4871:1–322. doi: 10.11646/zootaxa.4871.1.1
3. Peñalver E, Arillo A, Delclòs X, Peris D, Grimaldi DA, Anderson SR, et al. Ticks parasitised feathered dinosaurs as revealed by Cretaceous amber assemblages. Nat Commun. (2017) 8:1–13. doi: 10.1038/s41467-017-01550-z
4. de la Fuente J. The fossil record and the origin of ticks (Acari: Parasitiformes: Ixodida). Exp Appl Acarol. (2003) 29:331–44. doi: 10.1023/a:1025824702816
5. Mans BJ, De Klerk D, Pienaar R, Latif AA. Nuttalliella namaqua: a living fossil and closest relative to the ancestral tick lineage: implications for the evolution of blood-feeding in ticks. PLoS ONE. (2011) 6:e23675. doi: 10.1371/journal.pone.0023675
6. Hoogstraal H. Argasid and Nuttalliellid ticks as parasites and vectors. Adv Parasitol. (1985) 24:135–238. doi: 10.1016/S0065-308X(08)60563-1
7. Clifford CM, Kohls GM, Sonenshine DE. The systematics of the subfamily Ornithodorinae (Acarina: Argasidae). I. The genera and subgenera. Ann Entomol Soc Am. (1964) 57:429–37. doi: 10.1093/aesa/57.4.429
8. Camicas JL, Hervy JP, Adam F, Morel PC. The Ticks of the World Nomenclature, Described Stages, Hosts, Distribution (Acarida, Ixodida) (Including New Species Described Before 1/01/96). Paris: Éditions de l'Orstom (1998).
9. Mans BJ, Kelava S, Pienaar R, Featherston J, de Castro MH, Quetglas J, et al. Nuclear (18S-28S rRNA) and mitochondrial genome markers of Carios (Carios) vespertilionis (Argasidae) support Carios Latreille, 1796 as a lineage embedded in the Ornithodorinae: re-classification of the Carios sensu Klompen and Oliver (1993) clade into its respective subgenera. Ticks Tick Borne Dis. (2021) 12:101688. doi: 10.1016/j.ttbdis.2021.101688
10. Klompen JSH, Oliver JH. Systematic relationships in the soft ticks (Acari: Ixodida: Argasidae). Syst Entomol. (1993) 18:313–31. doi: 10.1111/j.1365-3113.1993.tb00669.x
11. Vial L. Biological and ecological characteristics of soft ticks (Ixodida: Argasidae) and their impact for predicting tick and associated disease distribution. Parasite. (2009) 16:191–202. doi: 10.1051/parasite/2009163191
12. Mans BJ, Neitz AWH. Adaptation of ticks to a blood-feeding environment: evolution from a functional perspective. Insect Biochem Mol Biol. (2004) 34:1–17. doi: 10.1016/j.ibmb.2003.09.002
13. Manzano-Romn R, Daz-Martn V, de la Fuente J, Prez-Snchez R. Soft ticks as pathogen vectors: distribution, surveillance control. In: Shah MM, editor. Parasitology. Rijeka: InTech. p. 125–62.
14. Klimpel S, Mehlhorn H. Bats (Chiroptera) as vectors of diseases parasites. In: Klimpel S, Mehlhorn K, editors. Parasitology Research Monographs. Berlin; Heidelberg: Springer-Verlag (2014). p. 25–61.
15. Barbier E, Bernard E, Dantas-Torres F. Ecology of Antricola ticks in a bat cave in north-eastern Brazil. Exp Appl Acarol. (2020) 82:255–64. doi: 10.1007/s10493-020-00544-9
16. IUCN. The IUCN Red List of Threatened Species. Version 2020-3. (2020) Available online at: https://www.iucnredlist.org/ (accessed February 19, 2021).
17. Frick WF, Kingston T, Flanders J. A review of the major threats and challenges to global bat conservation. Ann N Y Acad Sci. (2019) 1469:5–25. doi: 10.1111/nyas.14045
18. Sándor AD. Underground life is still safest: comments on ‘Danger underground and in the open - predation on blind mole rats (Rodentia, Spalacinae) revisited.' Mamm Rev. (2017) 47:230–5. doi: 10.1111/mam.12089
19. McKee CD, Krawczyk AI, Sándor AD, Görföl T, Földvári M, Földvári G, et al. Host phylogeny, geographic overlap, and roost sharing shape parasite communities in European bats. Front Ecol Evol. (2019) 7:69. doi: 10.3389/fevo.2019.00069
20. Masseti M, Bruner E. The primates of the western Palaearctic: a biogeographical, historical, and archaeozoological review. J Anthropol Sci. (2009) 87:33–91.
21. Ficetola GF, Falaschi M, Bonardi A, Padoa-Schioppa E, Sindaco R. Biogeographical structure and endemism pattern in reptiles of the Western Palearctic. Prog Phys Geogr. (2018) 42:220–36. doi: 10.1177/0309133318765084
22. Wenzel RL. The streblid batflies of Venezuela (Diptera: Streblidae). Brigham Young Univ Sci Bull Biol Ser. (1976) 20:1. doi: 10.5962/bhl.part.5666
23. Patterson BD, Dick CW, Dittmar K. Nested distributions of bat flies (Diptera: Streblidae) on Neotropical bats: artifact and specificity in host-parasite studies. Ecography (Cop). (2009) 32:481–7. doi: 10.1111/j.1600-0587.2008.05727.x
24. Dietz C, von Helversen O, Nill D. Bats of Britain, Europe and Northwest Africa. London: A&C Black (2009).
25. Hornok S, Szoke K, Meli ML, Sándor AD, Görföl T, Estók P, et al. Molecular detection of vector-borne bacteria in bat ticks (Acari: Ixodidae, Argasidae) from eight countries of the Old and New Worlds. Parasites and Vectors. (2019) 12:50. doi: 10.1186/s13071-019-3303-4
26. Reeves WK, Mans BJ, Durden LA, Miller MM, Gratton EM, Laverty TM. Rickettsia hoogstraalii and a Rickettsiella from the Bat Tick Argas transgariepinus, in Namibia. J Parasitol. (2020) 106:663–9. doi: 10.1645/20-46
27. Varma MGR, Converse JD. Keterah virus infections in four species of Argas ticks (Ixodoidea: Argasidae). J Med Entomol. (1976) 13:65–70. doi: 10.1093/jmedent/13.1.65
28. Zhmaeva ZM, Pchelkina AA, Belashova VS. Spontaneous infection of Argas vespertilionis with Rickettsia burnetii in the south of Kazakhstan. Med Parazitol (Mosk). (1966) 35:595–6.
29. Mediannikov O, Fenollar F, Socolovschi C, Diatta G, Bassene H, Molez J-F, et al. Coxiella burnetii in humans and ticks in rural Senegal. PLoS Negl Trop Dis. (2010) 4:e654. doi: 10.1371/journal.pntd.0000654
30. Lv J, De Marco MDMF, Goharriz H, Phipps LP, McElhinney LM, Hernández-Triana LM, et al. Detection of tick-borne bacteria and babesia with zoonotic potential in Argas (Carios) vespertilionis (Latreille, 1802) ticks from British bats. Sci Rep. (2018) 8:1–9. doi: 10.1038/s41598-018-20138-1
31. Socolovschi C, Kernif T, Raoult D, Parola P. Borrelia, Rickettsia, and Ehrlichia species in bat ticks, France, 2010. Emerg Infect Dis. (2012) 18:1966–75. doi: 10.3201/eid1812.111237
32. Zhao S, Yang M, Liu G, Hornok S, Zhao S, Sang C, et al. Rickettsiae in the common pipistrelle Pipistrellus pipistrellus (Chiroptera: Vespertilionidae) and the bat soft tick Argas vespertilionis (Ixodida: Argasidae). Parasit Vectors. (2020) 13:10. doi: 10.1186/s13071-020-3885-x
33. Wilhelmsson PE. Detektion Med Realtids—PCR av TBE—virus, Lyssavirus och Borrelia Species i Fladdermusfästingen Argas vespertilionis Och Fladdermusskinnbaggen Cimex pipistrelli. Linköping: Linköpings Universitet (2018).
34. Hubbard MJ, Baker AS, Cann KJ. Distribution of Borrelia burgdorferi sl spirochaete DNA in British ticks (Argasidae and Ixodidae) since the 19th century, assessed by PCR. Med Vet Entomol. (1998) 12:89–97. doi: 10.1046/j.1365-2915.1998.00088.x
35. Evans NJ, Bown K, Timofte D, Simpson VR, Birtles RJ. Fatal borreliosis in bat caused by relapsing fever spirochete, United Kingdom. Emerg Infect Dis. (2009) 15:1331–3. doi: 10.3201/eid1508.090475
36. Hornok S, Szoke K, Görföl T, Földvári G, Tu VT, Takács N, et al. Molecular investigations of the bat tick Argas vespertilionis (Ixodida: Argasidae) and Babesia vesperuginis (Apicomplexa: Piroplasmida) reflect “bat connection” between Central Europe and Central Asia. Exp Appl Acarol. (2017) 72:69–77. doi: 10.1007/s10493-017-0140-z
37. Gardner RA, Molyneaux DH, Stebbings RE. Studies on the prevalence of haematozoa of British bats. Mamm Rev. (1987) 17:75–80. doi: 10.1111/j.1365-2907.1987.tb00051.x
38. Hornok S, Corduneanu A, Kontschan J, Beko K, Szoke K, Gorfol T, et al. Analyses of separate and concatenated cox1 and 18S rRNA gene sequences indicate that the bat piroplasm Babesia vesperuginis is phylogenetically close to Cytauxzoon felis and the “prototheilerid” Babesia conradae. Acta Vet Hung. (2018) 66:107–15. doi: 10.1556/004.2018.010
39. Al'khovskii SV, L'vov DK, MIu S, Shchetinin AM, Deriabin PG, Samokhvalov EI, et al. The taxonomy of the Issyk-Kul virus (ISKV, Bunyaviridae, Nairovirus), the etiologic agent of the Issyk-Kul fever isolated from bats (Vespertilionidae) and ticks Argas (Carios) vespertilionis (Latreille, 1796). Vopr Virusol. (2013) 58:11–5.
40. Vargina SG, Kuchuk LA, Gershtein VI, Karas FR. Transmission of Issyk Kul virus by Argas vespertilionis ticks in experiment. Ecol Viruses. (1982) 123–7.
41. Lvov DK, Karas FR, Timofeev EM, Tsyrkin YM, Vargina SG, Veselovskaya OV, et al. “Issyk-Kul” virus, a new arbovirus isolated from bats Argas (Carios) vespertilionis (Latr., 1802) in the Kirghiz SSR. Arch Gesamte Virusforsch. (1973) 42:207–9. doi: 10.1007/BF01270841
42. Oba M, Omatsu T, Takano A, Fujita H, Sato K, Nakamoto A, et al. A novel Bunyavirus from the soft tick, Argas vespertilionis, in Japan. J Vet Med Sci. (2015) 78:443–5. doi: 10.1292/jvms.15-0536
43. Orlova MV, Kononova YV. Pathogens associated with specific bat ectoparasites (Chiroptera) (A review of literary data). Parazitologiia. (2018) 52:137–53.
44. Aeschlimann A, Büttiker W. Importations de Tiques en Suisse (Acarina Ixodoidea). Bull Soc Entomol Suisse. (1975) 48:69–75.
45. Roubaud E, Colas-Belcour J. Argas boueti, nouvelle espece d'ixodide parasite des chauve-souris au Soudan francais. Parasitology. (1933) 25:472–7. doi: 10.1017/S0031182000019685
46. Hoogstraal H. Bat ticks of the genus Argas (Ixodidea, Argasidae). The Subgenus Chiropterargas. Fieldiana Zool. (1955) 37:579–600. doi: 10.5962/bhl.title.2895
48. Hoogstraal H. Biological patterns in the Afghanistan tick fauna. In: Proceedings of the 3rd International Congress of Acarology. Dordrecht: Springer (1973). p. 511–4.
49. Hoogstraal H. African Ixodoidea. Volume I Ticks of the Sudan. United States. Naval Medical Research (1956). p. 1101. doi: 10.5962/bhl.title.6870
50. Mohammad MK. Species the soft tck genus Argas (Acarina, Ixodoidea) in Iraq. Bull Iraq Nat Hist Museum. (1999) 9:131–5.
51. Petney TN, Pfäffle MP, Estrada-Peña A. Argas transgariepinus White, 1846 (Figs. 10 and 11). In: Estrada-Peña A, Mihalca, AD, Petney, T, editors. Ticks of Europe and North Africa: A Guide to Species Identification. Cham: Springer (2017). p. 37–40.
52. Pienaar R, de Klerk DG, Putterill JF, Mans BJ. Notes on maternal behaviour in soft ticks: specifically observed in Argas (Argas) striatus Bedford, 1932 and Argas (Secretargas) transgariepinus White, 1846. Ticks Tick Borne Dis. (2018) 9:889–95. doi: 10.1016/j.ttbdis.2018.03.020
53. Péter Á, Barti L, Corduneanu A, Hornok S, Mihalca AD, Sándor AD. First record of Ixodes simplex found on a human host, with a review of cases of human infestation by bat tick species occurring in Europe. Ticks Tick Borne Dis. (2021) 12:101722. doi: 10.1016/j.ttbdis.2021.101722
54. Petney TN, Jaenson TGT, Pfäffle MP. Argas vespertilionis (Latreille, 1796) (Figs. 8 and 9). In: Estrada-Peña A, Mihalca A, Petney T, editors. Ticks of Europe and North Africa. Cham: Springer (2017). p. 33–36.
55. Hornok S, Szoke K, Tu VT, Kontschán J, Takács N, Sándor AD, et al. Mitochondrial gene heterogeneity of the bat soft tick Argas vespertilionis (Ixodida: Argasidae) in the Palaearctic. Parasit Vectors. (2017) 10:109. doi: 10.1186/s13071-017-2037-4
56. Del Cacho E, Estrada-Peña A, Sanchez A, Serra J. Histological response of Eptesicus serotinus (Mammalia: Chiroptera) to Argas vespertilionis (Acari: argasidae). J Wildl Dis. (1994) 30:340–5. doi: 10.7589/0090-3558-30.3.340
57. Walter G, Rackow W. Außergewöhnlich hoher Befall einer Nordfledermaus, Eptesicus nilssonii mit der Lederzecke, Argas vespertilionis (Argasidae). Nyctalus. (2007) 12:372–8.
58. Sándor AD, Corduneanu A, Péter Á, Mihalca AD, Barti L, Csosz I, et al. Bats and ticks: host selection and seasonality of bat-specialist ticks in eastern Europe. Parasit Vectors. (2019) 12:605. doi: 10.1186/s13071-019-3861-5
59. Walter G. Zum Ektoparasitenbefall der Fledermäuse und den potentiellen Auswirkungen. Myotis. (1996) 34:85–92.
60. Hoogstraal H. Bat ticks of the genus Argas (Ixodoidea, Argasidae), 3. The subgenus Carios, a redescription of A. (C.) vespertilionis (Latreille, 1802), and variation within an Egyptian population. Ann Entomol Soc Am. (1958) 51:19–26. doi: 10.1093/aesa/51.1.19
62. Jaenson TGT, Talleklint L, Lundqvist L, Olsen B, Chirico J, Mejlon H. Geographical distribution, host associations, and vector roles of ticks (Acari: Ixodidae, Argasidae) in Sweden. J Med Entomol. (1994) 31:240–56. doi: 10.1093/jmedent/31.2.240
63. Hoogstraal H. Ornithodoros salahi sp. nov. (Ixodoidea, Argasidae) from the Cairo Citadel, with notes on O. piriformis Warburton, 1918 and O. batuensis Hirst, 1929. J Parasitol. (1953) 39:256–63. doi: 10.2307/3273947
64. Estrada-Pena A, Sanches-Acedo C, Peribanez-Lopez MA. Nuevos datos relativos a la distribución de los ixodoidea en España (IV): primera cita de Ornithodoros (Reticulinasus) salahi Hoogstraal, 1953 (Acarina: Argasidae). Rev Iber Parasitol. (1989) 49:73.
65. Hoogstraal H, Clifford CM, Keirans JE. The Ornithodoros (Alectorobius) capensis group (Acarina: Ixodoidea: Argasidae) of the palearctic and oriental regions. O. (A.) coniceps identity, bird and mammal hosts, virus infections, and distribution in Europe, Africa, and Asia. J Parasitol. (1979) 65:395–407. doi: 10.2307/3280282
66. Theodor O, Costa M. New species and new records of Argasidae from Israel. Observations on the rudimentary scutum and the respiratory system of the larvae of the Argasidae. Parasitology. (1960) 50:365–86. doi: 10.1017/S0031182000025464
67. Rupp D, Zahn A, Ludwig P. Actual records of bat ectoparasites in Bavaria (Germany). Spixiana. (2004) 27:185–90.
68. Colas-Belcour J. Presence d'Ornithodoros coniceps en Tunisie. Arch Inst Pasteur Tunis. (1929) 18:265–7.
69. Jung K, Threlfall CG. Urbanisation and its effects on bats-a global meta-analysis. In: Voigt C, Kingston T, editors. Bats in the Anthropocene: Conservation of Bats in a Changing World. Cham: Springer (2016). p. 13–33.
70. Villarroya-Villalba L, Casanelles-Abella J, Moretti M, Pinho P, Samson R, Van Mensel A, et al. Response of bats and nocturnal insects to urban green areas in Europe. Basic Appl Ecol. (2021) 51:59–70. doi: 10.1016/j.baae.2021.01.006
71. Nusová G, Uhrin M, Kanuch P. Go to the city: urban invasions of four pipistrelle bat species in eastern Slovakia. Eur J Ecol. (2019) 5:23–6. doi: 10.2478/eje-2019-0004
72. Ancillotto L, Santini L, Ranc N, Maiorano L, Russo D. Extraordinary range expansion in a common bat: the potential roles of climate change and urbanisation. Sci Nat. (2016) 103:15. doi: 10.1007/s00114-016-1334-7
73. Kravchenko K, Vlaschenko A, Prylutska A, Rodenko O, Hukov V, Shuvaev V. Year-round monitoring of bat records in an urban area: Kharkiv (NE Ukraine), 2013, as a case study. Turkish J Zool. (2017) 41:530–48. doi: 10.3906/zoo-1602-51
Keywords: Chiroptera, host-specificity, Ixodoidea, soft ticks, zoonotic diseases
Citation: Sándor AD, Mihalca AD, Domşa C, Péter Á and Hornok S (2021) Argasid Ticks of Palearctic Bats: Distribution, Host Selection, and Zoonotic Importance. Front. Vet. Sci. 8:684737. doi: 10.3389/fvets.2021.684737
Received: 23 March 2021; Accepted: 14 May 2021;
Published: 22 June 2021.
Edited by:
Sebastián Muñoz-Leal, University of Concepcion, ChileReviewed by:
José M. Venzal, Centro Universitario Regional Litoral Norte Salto, Universidad de la Republica, UruguayBen J. Mans, Agricultural Research Council, South Africa
Copyright © 2021 Sándor, Mihalca, Domşa, Péter and Hornok. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Attila D. Sándor, YXR0aWxhLnNhbmRvckB1c2FtdmNsdWoucm8=