
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Vet. Sci. , 22 June 2021
Sec. Livestock Genomics
Volume 8 - 2021 | https://doi.org/10.3389/fvets.2021.665872
This article is part of the Research Topic Sheep and Goat Gene Exploration View all 19 articles
 Zhiying Wang1†
Zhiying Wang1† Bohan Zhou1†Tao Zhang1,2†Xiaochun Yan1Yongsheng Yu1Jinquan Li3,4,5Bujun Mei6
Bohan Zhou1†Tao Zhang1,2†Xiaochun Yan1Yongsheng Yu1Jinquan Li3,4,5Bujun Mei6 Zhixin Wang1Yanjun Zhang1Ruijun Wang1Qi Lv1Zhihong Liu2Yanhong Zhao2Chen Du7*
Zhixin Wang1Yanjun Zhang1Ruijun Wang1Qi Lv1Zhihong Liu2Yanhong Zhao2Chen Du7* Rui Su1*
Rui Su1*Objective: The purpose of this study was to discover the population structure and genetic diversity of Inner Mongolia White Cashmere goats (IMCGs) and demonstrate the effect of inbreeding on the live body weight (LBW), cashmere yield (CY), fiber length (FL), and fiber diameter (FD) of IMCGs.
Materials and Methods: All data were collected from pedigree information and production performance records of IMCGs from 1983 to 2019. The population structure and genetic diversity were analyzed by Endog 4.8 software. Inbreeding coefficients were obtained by the pedigree package in R. Then, a linear regression model was used to analyze how inbreeding influences economic traits in IMCGs. Four levels of inbreeding coefficients (Fi) were classified in this study, including Fi = 0, 0< Fi ≤ 6.25, 6.25< Fi ≤ 12.5 and Fi≥12.5. Variance analysis was performed to determine whether inbreeding levels had a significant effect on economic traits in IMCGs.
Results: The proportions of rams and dams in IMCGs for breeding were relatively small, with values of 0.8 and 20.5%, respectively. The proportion of inbred animals in the entire population was high, with values up to 68.6%; however, the average inbreeding coefficient and relatedness coefficient were 4.50 and 8.48%, respectively. To date, the population has experienced 12 generations. The average generation interval obtained in the present study was 4.11 ± 0.01 years. The ram-to-son pathway was lowest (3.97 years), and the ewe-to-daughter pathway was highest (4.24 years). It was discovered that the LBW, CY, and FL increased by 3.88 kg, 208.7 g, and 1.151 cm, respectively, with every 1% increase in the inbreeding coefficient, and the FD decreased by 0.819 μm with every 1% increase in the inbreeding coefficient. Additionally, multiple comparison analysis indicated that when the inbreeding coefficient was higher than 6.25%, the LBW showed an obvious decreasing trend. The threshold value of inbreeding depression in the CY is 12.5%. However, inbreeding depression has not been observed in the FL and FD.
Conclusion: Pedigree completeness needs to be further strengthened. The degree of inbreeding in this flock should be properly controlled when designing breeding programs.
As a typical small ruminant, cashmere goats are mainly distributed in dry and harsh climatic conditions in the tropics (1). Goat farming is practiced worldwide, with goat products having a favorable image (2, 3). The number of goats has increased globally, even in countries with high and intermediate incomes, despite major changes in agriculture due to industrial mergers, globalization, and technological advances in developed countries (4, 5). Most cashmere goats are dual-purpose breeds that produce cashmere, meat, and milk (6). China is the largest cashmere goat-producing country in the world. According to statistics, the total cashmere produced in China was ~15,437.76 tons in 2018, accounting for more than 2/3 of the world's total output. The export volume of cashmere reached 3,212 tons (http://www.fao.org/home/en/). The genetic resources of cashmere goats are very rich throughout the world and are mainly distributed in Asia, including Mongolia, Iran, Afghanistan, Kazakhstan, Kyrgyzstan, and Tajikistan. Inner Mongolia White Cashmere goats (IMCGs) and Liaoning Cashmere goats (LNCGs) are key breeds in China (7). The male parents of most goat breeds in China come from both varieties (8). IMCGs and LNCGs are not allowed to be exported and are listed in the directory of genetic resource protection. The number of goat breeds in China is ~69, including 18 cashmere goat breeds. There are 15 local varieties and 3 cultivated varieties (9).
Genetic diversity and population structure in animals are usually analyzed by using microsatellite and mitochondrial DNA variations (10, 11). In recent years, pedigree analysis has been an appropriate tool for evaluating genetic diversity and population structure in populations (12–16). Baena et al. (12) analyzed the genetic structure of the Mangalarga Marchador horse population in Brazil based on pedigree analysis and identified factors that may affect its genetic variability. Illa et al. (14) assessed the genetic diversity and population structure and appraised the efficiency of ongoing selective breeding programs in a closed nuclear herd of Nellore sheep through pedigree analysis. Vigeland (16) calculated relatedness coefficients by using pedigree information with inbred founders. Vatankhah et al. (15) described the population genetic structure and evaluated the state of conservation of genetic variability of Lori-Bakhtiari sheep in Iran. Goleman et al. (13) analyzed the degree of relatedness between individuals in the Polish hunting dog population and assessed the genetic variability of the population based on pedigree analysis. The analysis of genetic diversity and population structure will contribute to the conservation of animal genetic resources (14). IMCGs are an indigenous breed that is famous around the world due to its superior cashmere. In a previous study, the genetic diversity and population structure of IMCGs were analyzed by mitochondrial DNA and microsatellite polymorphism (17). This is the first study to analyze the population structure and genetic diversity of IMCGs based on pedigree information.
Many studies have reported that inbreeding in one flock may result in a decrease in production performance (14, 18). Yousefi et al. (19) quantified the effect of inbreeding on the average daily gain and the Kleiber ratio in native Mazandaran chickens. It was demonstrated that inbreeding had a significant effect on the average daily gain from hatching to 8 and 12 weeks of age (19). Paiva R demonstrated that MY305 was significantly affected by inbreeding (20). Todd et al. (18) analyzed the effects of inbreeding on covering success, gestation length, and foal sex ratio in Australian thoroughbred horses. Kiya et al. (21) reported that the inbreeding effect was significant for the longissimus muscle area and backfat thickness. In this study, the effect of the inbreeding coefficient on important economic traits in IMCGs was analyzed, which is helpful for properly designing mating schemes.
In this study, genealogical information of IMCGs for a period of 37 years from 1983 to 2019 was collected for genetic diversity and population structure analysis. A total of 53,381 individuals were recorded, including 25,032 male lambs and 28,349 female lambs. The pedigree was edited to check the inconsistencies in dam and sire registration, birth date and sex registration. All of the individuals were sorted by date of birth for the next analysis.
The data were provided by Inner Mongolia Yiwei White Cashmere Goat Co., Ltd, China. The goats are reared at Etuoke Banner, Ordos City, Inner Mongolia Autonomous Region, China with arid and semiarid areas. IMCGs graze year round with supplementary feeding in the winter. Artificial insemination was used in this flock. The mating ratio of male to female sheep was ~1:200–300. Mating usually occurred in early October of each year and lasted nearly 50 days. Lambs were born during the month of March. The feeding management and trait measurement methods have been described in detail in our previous studies (22). All birthing events were recorded, including the identification number, date of birth, birth status, sex, and birth weight. Pedigree information was comprehensive and clear with kids, sires, and dams. The traits evaluated in this study included the live body weight (LBW), cashmere yield (CY), fiber length (FL), and fiber diameter (FD). The basic statistics for each trait are shown in Table 1.
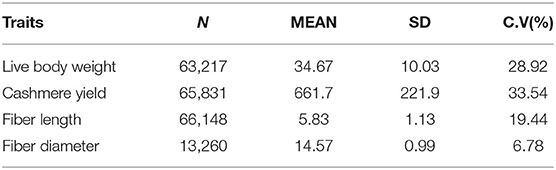
Table 1. The basic statistics of economic traits of IMCGs.
Endog 4.8 software was utilized to perform pedigree analysis and obtain corresponding parameters to illustrate the historical diversity and population structure (23). The depth and wholeness of the pedigree was determined by estimating the equivalent number of generations, and it was estimated by tracing back each ancestor in the pedigree history through numerous generations.
The founders are defined as individuals with one or both unknown parents. A 4-year time path was used to define the reference population because this time duration represents an approximate generation interval in goats (14). In the reference population, the effective number of founders and ancestors is useful to assess genetic diversity. The effective number of founders is characterized as the number of equally contributing founders that would be expected to produce the same genetic diversity as in the population under study (24). The formula is as follows:
where qk is the probability of gene origin for ancestor k. The effective number of ancestors (fa) reflects the minimum number of animals required to estimate the genetic diversity of the population under study, and it is a useful measure to determine the bottlenecks in the population that are the primary reason for genetic erosion in captive and domestic populations. It is estimated as:
where qj is the marginal contribution of ancestor j, which demonstrates the genetic contribution of an ancestor that is not explained by an earlier ancestor. In general, the effective number of ancestors should be smaller than the effective number of founders due to bottlenecks that reduce genetic variability.
The inbreeding coefficient (F) and the average relatedness (AR) coefficient were estimated by Meuwissen and Luo (24) and MaléCot (25), respectively. The AR coefficient of any animal is explained as the probability that an allele selected at random from the total population in the pedigree belongs to a particular animal; hence, it is equated as an account of the animal in the entire pedigree regardless of the pedigree information. F is defined as the probability that an individual has two identical alleles by descent. The change in breeding (ΔF) is estimated for each generation using the formulae suggested by Lacy (26).
where Fi is the individual inbreeding coefficient and t is the equivalent complete generation for this individual. The estimate of effective population size () was computed from by averaging ΔFi of n individuals included in a given reference subpopulation (27) as .
The genetic conservation index (GCI) for each of the individuals of the analyzed population was provided by Alderson (28). The index is computed from the genetic contributions of all of the identified founders as , where pi is the proportion of genes of founder i in the pedigree of an animal. The index is based on the assumption that the objective of a conservation program is to retain the full range of alleles possessed by the base population. In this respect, the ideal individual would receive equal contributions from all of the founder ancestors in the population, and consequently, the higher the GCI value is, the higher the values of an animal for conservation (28). The following parameters were estimated for each individual: (1) the number of fully outlined generations, detailed as the number of generations delineating the offspring of the furthest generation where the ancestors of second-generation individuals are known, and ancestors with unknown parents are considered founders (generation 0); (2) the maximum number of generations observed, determined as the number of generations separating the individual from its ultimate ancestor; and (3) equivalent complete generations are detailed as the sum over all known ancestors of the terms calculated as the aggregate of (1/2)n, where n is the number of generations separating the individual from each known ancestor. The average generation interval (GI) was pointed out as the average age of the birth of selected offspring. The estimate of GI for all of the pathways was estimated for the reference population, as this subpopulation is the most recent one that could accrue at least one generation on the farm.
Wright's (29) F-statistics are obtained as , and , where and are the mean coancestry and inbreeding coefficient for the entire metapopulation, respectively, and is the average coancestry for the subpopulation. D is the kinship distance for molecular coancestry (30, 31).
The parameter of founder genome equivalents (fg) can be defined as the number of founders that would be expected to produce the same genetic diversity as in the population under study if the founders were equally represented and no loss of alleles occurred. Parameter fg was obtained by the inverse of twice the average coancestry of the individuals included in a predefined reference population (30).
The basic statistics of each trait in this study are shown in Table 3. Except for the fiber diameter, the other three traits were collected from 1990 to 2019. The individual inbreeding coefficient was obtained by pedigree packages in R (32). A multiple linear regression model was used to analyze how the inbreeding coefficient influenced the four traits. In this study, the impact factors of each trait included the production year (1990–2019), flocks (1–12), age of individuals (1–7), age of dams (2–7) birth status (1, 2), and gender (1, 2). The inbreeding coefficient (Fi) is continuously viable; however, other factors are discrete variables. Therefore, these discrete variables need to be centered first. Then, regression analysis was carried out by using the lm function of R language (33). The regression model is as follows:
where yi is the vector of the observed value of the ithindividual. Yeari, Flocki, Agei, Dagei, Sexi, Bsi , and Fi are independent variables, while ei is the residual. γ00 is an intercept, which was also used as the overmean. γ01, γ02, γ03, γ04, γ05, γ06, and γ07 are the slope coefficients of Yeari, Flocki, Agei, Dagei, Sexi, Bsi , and Fi, which explained the incremental change in the dependent variable for each unit of change in the independent variables.
To assess how inbreeding influences the important economic traits of IMCGs, it is helpful to design a proper breeding scheme. Referring to corresponding studies (14), inbreeding coefficients (Fi) obtained by pedigree information were classified into four levels (Fi = 0; 0 < Fi ≤ 6.25; 6.25 < Fi ≤ 12.5; Fi ≥ 12.5). Variance analysis of inbreeding levels on each trait was performed by the aov function of R language.
The main demographic characteristics derived from genealogical data of IMCG flocks are summarized in Table 2. In this population, 0.8 and 20.5% are presented as sires and dams, respectively. Out of 53,381 animals, the number of founders (with one or more unknown parents) was 5,455, accounting for ~10.22%. Of the total animals, the ratio of individuals with no progeny was ~78.7%. The timeline trend of all animals and individuals with both known parents across years is presented in Figure 1. From 1983 to 1989, all animals in the pedigree were founders. The number of individuals with both unknown parents was relatively low from 1990 to 2012. After 1998, the number of individuals remained stable on this farm. Additionally, the pedigree completeness is shown in Figure 2. In total, of 53,381 animals, more than 90% had records with both parents. However, pedigree completion was poor when tracing back generations.

Table 2. Pedigree structure of Inner Mongolia White Cashmere goats.
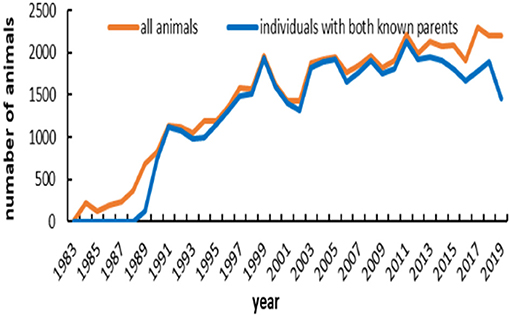
Figure 1. Number of animals and number of individuals with both known parents across years.
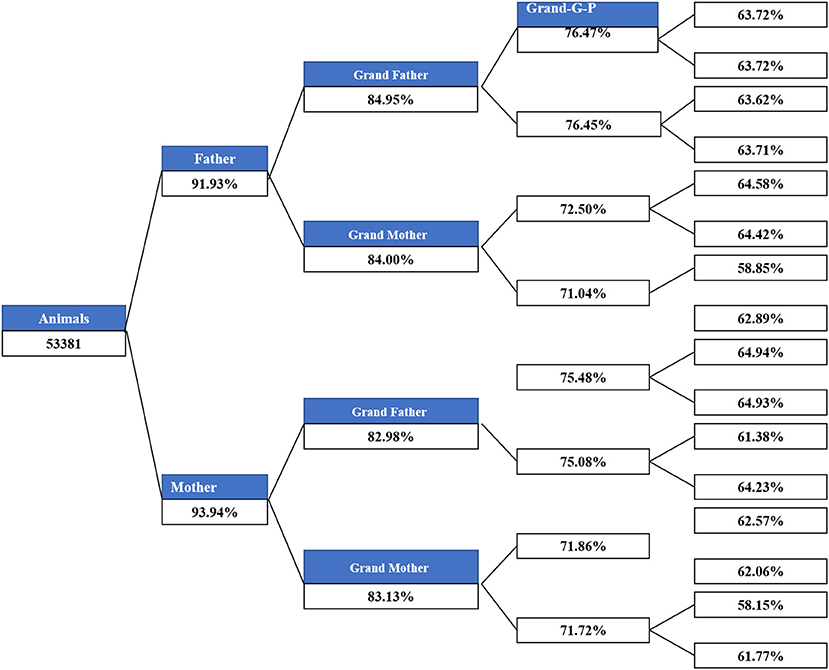
Figure 2. Pedigree completeness across generations.
The number of registered and inbred animals across birth years is shown in Figure 3. No inbred animal was found until 1993. The proportion of inbred animals in all registered individuals was more than 63.4%. An overall increasing trend did exist in the inbred animals for IMCGs from 1999 to 2005 and then remained stable. The number of inbred animals even reached 97.7% of the registered animals in 2005. The timeline trend of the average inbreeding coefficients is presented in Figure 4. Increasing trends of average inbreeding coefficients were observed in all registered animals. However, there was no regular trend for average inbreeding coefficients in the inbred animals. A decreasing trend for average inbreeding coefficients was observed from 1993 to 1998 in the inbred animals.
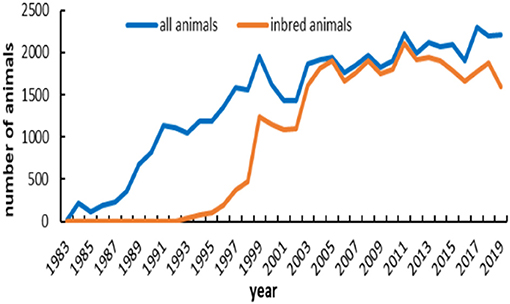
Figure 3. Registered and inbred individuals of IMCGs from 1983 to 2019.
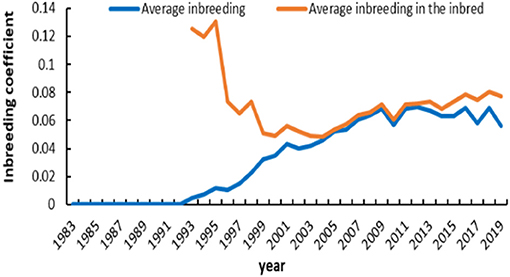
Figure 4. Average inbreeding coefficient in registered and inbred individuals in IMCGs across years.
The probabilities of gene origin parameters in the studied breed are presented in Table 3. Of the 8,592 animals in the reference population, there were 4,298 males and 4,303 females. The number of founder animals in the reference population was 1980. The number of ancestors contributing to the reference population of IMCGs was 1977, which accounted for 99.8% of founders. The effective numbers of founders and ancestors in the studied population were 27.87 and 23.42, respectively. The effective numbers of founders and ancestors in the reference population were 24 and 19, respectively. The parameters of mean maximum generations, complete generations and equivalent generations were 11, 7, and 8.55, and the increases in inbreeding in the corresponding generations were 0.76, 1.66, and 1.30%, respectively. Fifty percent of genetic diversity was explained by six influential ancestors in the reference cohort. The founder genome equivalent of the reference population was 11.78. F-statistics were used to assess genetic differentiation in subdivided populations, and the value was 5.7 × 10−5.
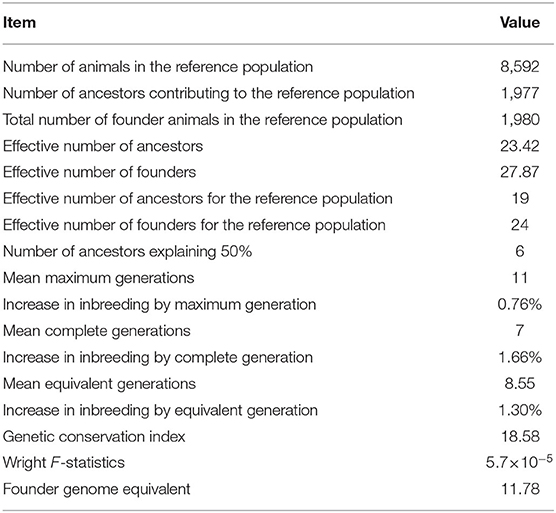
Table 3. Parameters of the probability of gene origin for the reference population of IMCGs.
The distribution of the inbreeding coefficient among IMCGs is presented in Table 4. A total of 36,602 animals were inbred. Approximately 55.7% of inbred animals had inbreeding lower than 6.25%. The inbreeding coefficient ranged from 0.98 to 35.93%. A total of 3.96% of inbred animals had an inbreeding coefficient > 25%. The average inbreeding value and mean average relatedness in the entire studied population were 4.50 and 8.48%, respectively. The average inbreeding in the inbred individuals was 6.57%. It is presumed that effective population size is considered the number of animals that breed in an ideal population and engender an equal amount of inbreeding in the population under study. The realized effective population size (Ner) was 15.16 ± 3.33.

Table 4. Inbreeding, average relatedness, and effective population size in IMCGs.
This population has gone through 12 generations by pedigree tracing (Table 5). With the increase in generations, the inbreeding coefficient and average relatedness coefficient show an overall increasing trend. More than 90% of the individuals in the population are inbred in the fifth generation. The mean relatedness coefficient ranged from 9.49 to 10.74% after the fifth generation, which was relatively high. In the first two generations, there were no inbred animals, and the average relatedness was also very low, with values of 0.05 and 1.79%. The estimation of generation intervals (in years) for the four pathways of the IMCGs is presented in Table 6. The average generation length obtained in the present study was 4.11 ± 0.01 years. The ram-to-son pathway was lowest (3.97 years), and the ewe-to-daughter pathway was highest (4.24 years). However, no significant differences in generation intervals were observed among the four pathways in IMCGs.
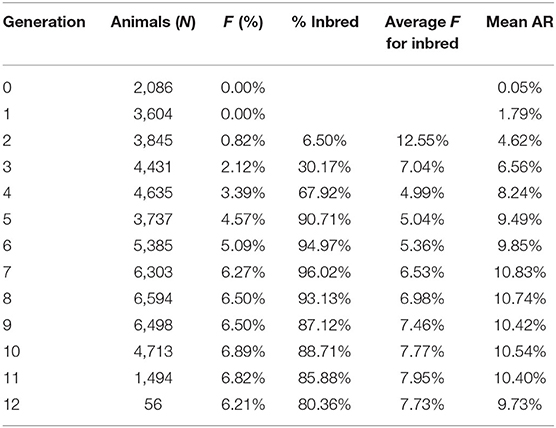
Table 5. Mean value of inbreeding (F) and percentage of endogamic animals of IMCGs using the maximum number of generations traced.
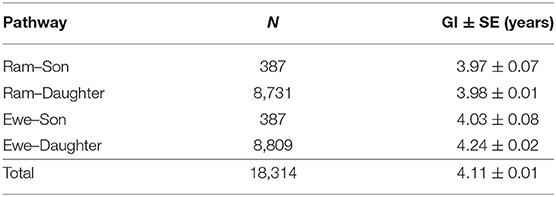
Table 6. Generation intervals (in years) for the four pathways of the IMCGs.
A linear regression model was established to assess the relationship between important economic traits and the inbreeding coefficients of IMCGs, and the results are shown in Table 7. The production year, herd, individual ages, age of dam, and sex had highly significant effects on all four traits (P < 0.01). The birth status had no significant effect on the FL and FD (P > 0.05), but the other three traits were significantly affected by the birth status (P < 0.01). Excluding the FL, the inbreeding coefficient had a significant effect on the other three traits. The results demonstrated that the LBW, CY, and FL increased by 3.88 kg, 208.7 g, and 1.151 cm, respectively, with every 1% increase in the inbreeding coefficient, and the FD decreased by 0.819 μm with every 1% increase in the inbreeding coefficient.
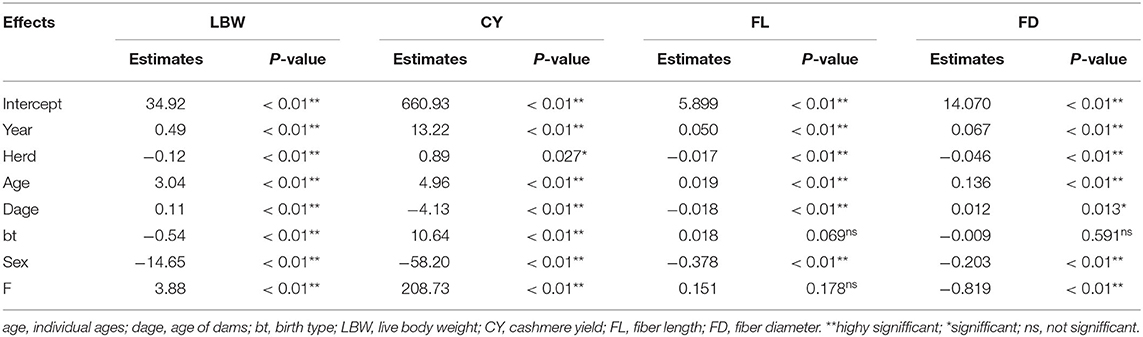
Table 7. Regression analysis of inbreeding on the economic traits of IMCGs.
Variance analysis was performed to determine the threshold value of the inbreeding value for each trait. The results are shown in Table 8. Inbreeding levels had a significant effect on all of the traits (P < 0.01). Additionally, the multiple comparison analysis indicated that when the inbreeding coefficient was more than 6.25%, the LBW had an obvious decreasing trend. The threshold value of inbreeding depression in the CY is 12.5%. However, inbreeding depression has been observed in the FL and FD.
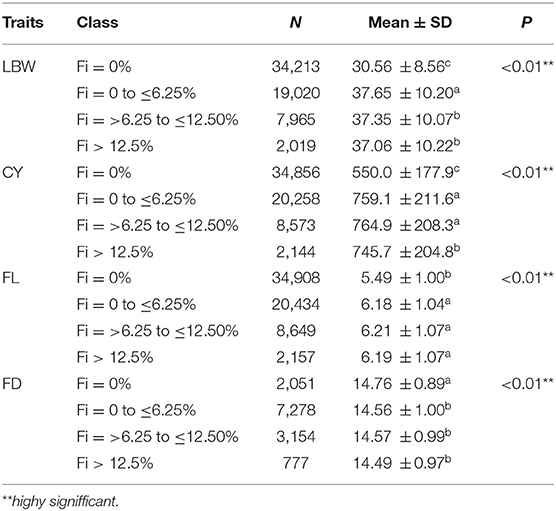
Table 8. Variance analysis of inbreeding levels on important economic traits of IMCGs.
In this population, 0.8 and 20.5% are presented as sires and dams, respectively. The fraction of males and females selected in IMCGs was relatively low. Individuals with no progeny accounted for 78.7% of all animals. All of these animals were sold as stud stocks to other flocks to improve performance or eliminated at the market for goat meat. Artificial insemination is used in this flock, so very few sires were selected. Out of 53,381 animals, the number of founders (with one or more unknown parents) was 5,455, accounting for ~10.22%. The results suggested a good depth in the pedigree in terms of completeness. Therefore, the accuracy of the estimated founders is reliable. The timeline trend of all animals with both known parents across the years indicated that the number of individuals with both unknown parents was low from 1990 to 2012; however, this value increased from 2013 to 2019. This may be related to incorrect pedigree information. After 1998, the population size remained stable. Our research team began to design a breeding plan for this flock in 1998, and thus, it had a reasonable population structure from then on.
This farm is representative of IMCGs. It has always undergone closed breeding and never introduced other breeds to cross. Therefore, the proportion of inbred animals among all registered individuals was more than 68.6%. A certain degree of inbreeding can improve production performance. Increasing trends of average inbreeding coefficients were observed in all registered animals. However, there was no regular trend for average inbreeding coefficients in the inbred animals. To maximize production performance, a reasonable mating plan is created every year. Most inbred animals showed inbreeding coefficients lower than 6.25%, and inbred animals with inbreeding coefficients > 25% accounted for ~4%. This may be related to our strict implementation of the proper mating scheme. A small fraction of high inbreeding individuals may be caused by natural mating or poor management. The average inbreeding value in the entire studied population was 4.50%, which is lower than that in the cashmere goat breed of the South Khorasan and Iranian Adani goat breeds reported by Joezy-Shekalgorabi et al. (34, 35). However, Illa et al. (14) reported that the average inbreeding coefficient in Nellore sheep was 3.32%, which was lower than that in the IMCG population. This difference may be explained by population structure, sample size, and management mode. The average relatedness in this population is 8.48%, which is higher than that in other ruminant breeds (36, 37). It is presumed that the effective population size is considered the number of animals that breed in an ideal population and engender an equal amount of inbreeding in the population under study. According to the FAO guidelines on preserving animal genetic resources, an effective population size of < 50 affects the fitness of the breed (38). The realized effective population size (Ner) noted in this population is 15.16 ± 3.33, which is far lower than that reported in Nellore sheep and Latxa dairy sheep (14, 39). This is caused by the differences among breeds, mating schemes, and breed plans. The F-statistic in this population is very low, which is far lower than that reported in other studies (37, 40). This indirectly reflects the genetic differentiation among the population. Thus, this result indicated that the genetic differentiation in the IMCG population was low.
Shortening the generation interval is one of the methods to increase genetic progress. It also may result in better economic returns. This outcome is the best choice for production enterprises. However, decreasing the ability of individuals to stay on the farm will intensify genetic variability losses, especially rams. The genetic contribution of those animals will be lower because of their short lifespan. The population in this study has gone through 12 generations by pedigree tracing. The average generation length obtained in the present study was 4.11 ± 0.01 years, which is longer than that estimated in Beetal goats. Tomar et al. (41) reported that the generation interval was 2.04 years for Beetal goats. Similar results were observed in other breeds of goats (34, 35, 42). The generation length is ~4 years for most goats. Compared to the sire–progeny pathways, a higher mean generation interval of the dam–progeny pathways was observed. This is probably attributable to the fact that breeding dams are usually kept for more years to produce offspring than sires. Similar results were obtained in Creole goats and Nellore sheep (14, 42). Animal conservation programs should balance the lowered generation intervals with decent annual genetic gains and breeding animals with sustained genetic variability on the farm.
The linear regression analysis demonstrated that the LBW, CY, and FL increased by 3.88 kg, 208.7 g, and 1.151 cm with every 1% increase in the inbreeding coefficient, and the FD decreased by 0.819 μm with every 1% increase in the inbreeding coefficient. It was illustrated that inbreeding had no negative impact on the economic traits under the current mating scheme. However, the variance of analysis of inbreeding levels indicated that when the inbreeding coefficient of most individuals was higher than 6.25 and 12.5%, the LBW and CY will produce inbreeding depression, respectively. Hence, the breeding plan should be updated appropriately. However, it is surprising to find that the increase in inbreeding resulted in a decrease in the FD, which is beneficial to the improvement of fiber quality. The results of this population are similar to those reported by Dai et al. (43). This study illustrated that inbreeding depression for fleece traits did not exist when the inbreeding coefficient reached 12.5%. Most studies have shown that inbreeding has no significant effect on fleece traits (36). Sousa et al. (44) reported that inbreeding had a significant effect on the body weight of Anglo Nubiana breed goats. Vostra-Vydrova et al. (37) indicated that inbreeding coefficients showed a significant negative influence on milk performance in the White Shorthair goat breed. This may be due to the differences in the studied traits, population structure, and data size.
Population structure across years in the Inner Mongolia Cashmere goat breed was documented in this study. A small number of unknown parents was found to be the reason for complete and detailed pedigree information. Although the proportion of inbred individuals in the entire population is high, the low average inbreeding coefficient and average relatedness obtained in the studied population indicated that the current mating scheme is relatively reasonable. Generally, animals with unknown parents are assumed to have no inbreeding. However, in reality, this will lead to underestimation of the inbreeding coefficient. An effective way to solve this problem is to ensure pedigree completeness. Alternatively, utilizing paternity testing methods helps to overcome the problem of pedigree incompleteness. Although inbreeding has not resulted in an obvious decrease in economic traits, it should be controlled properly when designing mating schemes.
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author/s.
TZ, XY, YY, and QL analyzed the data. RS, CD, and ZhiyW conceived of and coordinated the study. BM, RW, and JL helped in conceive of the study. ZhixW, YZhan, ZL, and YZhao help to collect data. ZhiyW and BZ wrote the manuscript. All authors read and approved the final manuscript.
TZ is only employed by the company Inner Mongolia Bigvet Co., Ltd.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors of the article would like to send their sincere gratitude to the Natural Science Foundation of the Inner Mongolia Autonomous Region (2019 MS03070), the Science-Technology Project of Inner Mongolia Region (2020SGG2693), the Inner Mongolia Agricultural University for second levels of outstanding doctorate (NDYB2016-05), the Natural Science for Youth Foundation (31702086), the Natural Science Foundation of China (No. 31760660), and the Youth Science and Technology Talent Support Program of Inner Mongolia Autonomous Region (No. NJYT- 17-A21). Additionally, the authors would like to thank the Inner Mongolia Yiwei White Cashmere Goat Co., Ltd. for providing the necessary data to complete this study.
1. Vazir NEA, Koshkooieh AE, Mehrjerdi AA, Mohammadabadi M, Babenko O, Bushtruk M, et al. Comparison of genetic diversity of leptin gene between wild goat and domestic goat breeds in iran. Malaysian Appl Biol. (2019) 48:85–93.
2. Gooki FG, Mohammadabadi M, Fozi MA, Soflaei M. Association of biometric traits with growth hormone gene diversity in raini cashmere goats. Walailak J Sci Technol. (2019) 16:499–508. doi: 10.48048/wjst.2019.3791
3. Khorshidi M, Mohammadabadi MR, Esmailizadeh AK, Barazandeh A, Babenko OI. Comparison of artificial neural network and regression models for prediction of body weight in Raini Cashmere goat. Iran J Appl Anim Sci. (2019) 9:453–61.
4. Gholamhoseini Gooki F, Mohammadabadi MR, Asadi Fozi M. Polymorphism of the growth hormone gene and its effect on production and reproduction traits in goat. Iran J Appl Anim Sci. (2018) 8:653–9.
5. Shamsalddini S, Mohammadabadi MR, Esmailizadeh AK. Polymorphism of the Prolactin gene and its effect on fiber traits in goat. Russ J Genet. (2016) 52:405–8. doi: 10.1134/S1022795416040098
6. Moghadaszadeh M, Mohammadabadi MR, Esmailizadeh AK. Association of Exon 2 of BMP15 gene with the litter size in the Raini Cashmere goat. Genet Millennium. (2015) 13:4062–7.
7. Jin M, Lu J, Fei X, Lu Z, Quan K, Liu Y, et al. Genetic signatures of selection for cashmere traits in chinese goats. Animals. (2020) 10:1905. doi: 10.3390/ani10101905
8. Qi Y, Luo J, Han X, Zhu Y, Chen C, Liu J, et al. Genetic diversity and relationships of 10 Chinese goat breeds in the Middle and Western China. Small Rumin Res. (2009) 82:88–93. doi: 10.1016/j.smallrumres.2009.01.015
9. National livestock and Poultry Genetic Resources Committee. Chinese Livestock and Poultry Genetic Resources. Special Livestock and Poultry, Sheep and Goats. China Agricultural Publishing House (2011).
10. Diwedi J, Singh AW, Ahlawat S, Sharma R, Arora R, Sharma H, et al. Comprehensive analysis of mitochondrial DNA based genetic diversity in Indian goats. Gene. (2020) 756:144–9. doi: 10.1016/j.gene.2020.144910
11. Phyu PP, Pichler R, Soe O, Aung PP, Than M, Shamsuddin M, et al. Genetic diversity, population structure and phylogeography of Myanmar goats. Small Rumin Res. (2017) 148:33–42. doi: 10.1016/j.smallrumres.2016.12.028
12. Baena MM, Gervásio IC, Rocha RDFB, Procópio AM, de Moura RS, et al. Population structure and genetic diversity of Mangalarga Marchador horses. Livestock Sci. (2020) 239:104–9. doi: 10.1016/j.livsci.2020.104109
13. Goleman M, Balicki I, Radko A, Jakubczak A, Fornal A. Genetic diversity of the Polish Hunting Dog population based on pedigree analyses and molecular studies. Livestock Sci. (2019) 229:114–7. doi: 10.1016/j.livsci.2019.09.017
14. Illa SK, Gollamoori G, Nath S. Evaluation of selection program by assessing the genetic diversity and inbreeding effects on Nellore sheep growth through pedigree analysis. Asian Aust J Anim Sci. (2019) 33:1369–77. doi: 10.5713/ajas.18.0553
15. Vatankhah M, Sigdel A, Abdollahi-Arpanahi R. Population structure of Lori-Bakhtiari sheep in Iran by pedigree analysis. Small Rumin Res. (2019) 174:148–55. doi: 10.1016/j.smallrumres.2019.02.019
16. Vigeland MD. Relatedness coefficients in pedigrees with inbred founders. J Math Biol. (2020) 81:185–207. doi: 10.1007/s00285-020-01505-x
17. Ma Y, Su R, Fan Y, Qiao X, Li X, Zhang L, et al. The complete mitochondrial DNA analyses of Inner Mongolia Cashmere goat type of Erlangshan species. Mitochondr DNA B. (2018) 3:44–5. doi: 10.1080/23802359.2017.1419084
18. Todd ET, Hamilton NA, Velie BD, Thomson PC. The effects of inbreeding on covering success, gestation length and foal sex ratio in Australian thoroughbred horses. BMC Genet. (2020) 21:41. doi: 10.1186/s12863-020-00847-1
19. Yousefi B, Gholizadeh M, Hafezian H. Quantifying the effect of inbreeding on average daily gain and Kleiber ratio in Mazandaran native chickens. Trop Anim Health. (2020) 52:1–7. doi: 10.1007/s11250-020-02347-x
20. Paiva R, Sousa J, Ferreira J, Cunha EE, Facó O. Population structure and effect of inbreeding on milk yield of Saanen goats in Brazilian production systems. Small Rumin Res. (2020) 192:106–14. doi: 10.1016/j.smallrumres.2020.106194
21. Kiya CK, Pedrosa VB, Muniz K, Gusmo AL, Pinto L. Population structure of a nucleus herd of Dorper sheep and inbreeding effects on growth, carcass, and reproductive traits. Small Rumin Res. (2019) 177:141–5. doi: 10.1016/j.smallrumres.2019.06.015
22. Wang Z, Wang R, Zhang W, Wang Z, Wang P, Liu H, et al. Estimation of genetic parameters for fleece traits in yearling Inner Mongolia Cashmere goats. Small Rumin Res. (2013) 109:15–21. doi: 10.1016/j.smallrumres.2012.07.016
23. Gutiérrez JP, Goyache F. A note on ENDOG: a computer program for analysing pedigree information. J Anim Breed. (2005) 122:357–60. doi: 10.1111/j.1439-0388.2005.00512.x
24. Meuwissen T, Luo Z. Computing inbreeding coefficients in large populations. Genet Select Evol. (1992) 24:1–9. doi: 10.1186/1297-9686-24-4-305
26. Lacy RC. Analysis of founder representation in pedigrees: founder equivalents and founder genome equivalents. Zool Biol. (1989) 8:111–23. doi: 10.1002/zoo.1430080203
27. Gonzalez-Recio O, Maturana ELD, Gutierrez JP. Inbreeding depression on female fertility and calving ease in Spanish dairy cattle. J Dairy Sci. (2007) 90:5744–52. doi: 10.3168/jds.2007-0203
28. Alderson GHL. A system to maximize the maintenance of genetic variability in small populations. Conserv Domest Livest. (1991) 2:18–9.
29. Wright S. Coefficients of Inbreeding and relationship. Am. Nat. (1922) 56:330–8. doi: 10.1086/279872
30. Caballero A, Toro MA. Interrelations between effective population size and other pedigree tools for the management of conserved populations. Genet Res. (2000) 75:331–43. doi: 10.1017/S0016672399004449
31. Caballero A, Toro MA. Analysis of genetic diversity for the management of conserved subdivided populations. Conserv Genet. (2002) 3:289–99. doi: 10.1023/A:1019956205473
32. Therneau TM, Sinnwell J. Pedigree functions. Acta Math Acad Sci Hungaricae. (2015) 37:481–96. doi: 10.1007/BF01895150
33. Marschner I. Fitting generalized linear models [R package glm2 version 1.2]. R J. (2017) 3:12–5. doi: 10.32614/RJ-2011-012
34. Joezy-Shekalgorabi S, Maghsoudi A, Taheri-Yeganeh A, Rajabi-Marand B. Pedigree analysis of Cashmere goat breed of South Khorasan. Ital J Anim Sci. (2016) 15:590–4. doi: 10.1080/1828051X.2016.1221748
35. Joezy-Shekalgorabi S, Maghsoudi A, Taheri-Yeganeh A, Rajabi-Marand B. Genetic variability of Iranian adani goat breed using pedigree analysis. J Anim Plant Sci. (2017) 27:1774–80.
36. Mokhtari MS, Damaneh M, Gutierrez JP. Genetic variability and population structure of Raeini Cashmere goats determined by pedigree analysis. J Livestock Sci Technol. (2017) 5:43–50.
37. Vostra-Vydrova H, Hofmanova B, Moravcikova N, Rychtarova J, Vostry L. Genetic diversity, admixture and the effect of inbreeding on milk performance in two autochthonous goat breeds. Livestock Sci. (2020) 240:104–63. doi: 10.1016/j.livsci.2020.104163
38. FAO IDAD UNEP. Secondary Guidelines for Development of National Farm Animal Genetic Resources Management Plans. Anim Record Medium Input Prod Environ. (1998) 35:1369–75.
39. Granado-Tajada I, Rodríguez-Ramilo ST, Legarra A, Ugarte E. Inbreeding, effective population size, and coancestry in the Latxa dairy sheep breed. J Dairy Sci. (2020) 103:5215–26. doi: 10.3168/jds.2019-17743
40. Berihulay H, Li Y, Liu X, Gebreselassie G, Islam R, Liu W, et al. Genetic diversity and population structure in multiple Chinese goat populations using a SNP panel. Anim Genet. (2019) 50:242–9. doi: 10.1111/age.12776
41. Tomar SS, Arun K, Singh RB. Population analysis of a flock of Beetal goats for demographic parameters. Indian J Anim Res. (2000) 34:133–5.
42. Ginja C, Gama LT, Martin-Burriel I, Lanari MR, Revidatti MA, Aranguren-M endez JA, et al. Genetic diversity and patterns of population structure in Creole goats from the Americas. Anim Genet. (2017) 48:315–29. doi: 10.1111/age.12529
43. Dai S, Wang C, Wang Z, Wang Z, Zhang Y, Na Q, et al. Inbreeding and its effects on fleece traits of Inner Mongolia cashmere goats. Small Rumin Res. (2015) 128:50–3. doi: 10.1016/j.smallrumres.2015.04.007
Keywords: inner Mongolia white cashmere goats, genetic diversity, regression analysis, population structure, inbreeding
Citation: Wang Z, Zhou B, Zhang T, Yan X, Yu Y, Li J, Mei B, Wang Z, Zhang Y, Wang R, Lv Q, Liu Z, Zhao Y, Du C and Su R (2021) Assessing Genetic Diversity and Estimating the Inbreeding Effect on Economic Traits of Inner Mongolia White Cashmere Goats Through Pedigree Analysis. Front. Vet. Sci. 8:665872. doi: 10.3389/fvets.2021.665872
Received: 09 February 2021; Accepted: 10 May 2021;
Published: 22 June 2021.
Edited by:
Fabyano Fonseca Silva, Universidade Federal de Viçosa, BrazilReviewed by:
Radovan Kasarda, Slovak University of Agriculture, SlovakiaCopyright © 2021 Wang, Zhou, Zhang, Yan, Yu, Li, Mei, Wang, Zhang, Wang, Lv, Liu, Zhao, Du and Su. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Chen Du, ZHVjaGVuMTk4NjA3MjMwMEAxMjYuY29t; Rui Su, c3VydWl5dUAxMjYuY29t
†These authors have contributed equally to this work and share first authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.