
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Vet. Sci., 30 April 2021
Sec. Parasitology
Volume 8 - 2021 | https://doi.org/10.3389/fvets.2021.650531
 Amany M. Ramez1†
Amany M. Ramez1† Ehab Kotb Elmahallawy2*†
Ehab Kotb Elmahallawy2*† Gehad E. Elshopakey3
Gehad E. Elshopakey3 Amira A. Saleh4Samar M. Moustafa5
Amira A. Saleh4Samar M. Moustafa5 Ashraf Al-Brakati6Walied Abdo7,8
Ashraf Al-Brakati6Walied Abdo7,8 Dina M. M. El-Shewehy1
Dina M. M. El-Shewehy1Schistosomiasis, a major parasitic illness, has high morbidity and negative financial effects in subtropical and tropical countries, including Egypt. The present study investigated the therapeutic effects of Spirulina platensis (SP) and matcha green tea (MGT) in Schistosoma mansoni-infected mice combined with tracing their possible antioxidant and anti-inflammatory impacts and their protective potency. A total of 60 Swiss albino mice were randomly allocated into six groups (n = 10): control group (CNT, received normal saline); SP–MGT group [received oral SP (3 g/kg bodyweight/day) plus MGT (3 g/kg bodyweight/day)]; S. mansoni group (infected with S. mansoni cercariae, 100 ± 10/mouse, using the tail immersion method); SP-infected group (infected with S. mansoni and received oral SP); MGT-infected group (received oral MGT after S. mansoni infection); and SP–MGT-infected group (received combined treatment of SP and MGT after S. mansoni infection). Treatment with SP and MGT started 4 weeks after S. mansoni infection and ended 10 weeks after. SP and MGT treatment (SP-infected and MGT-infected groups) and the combined treatment (SP–MGT-infected group) minimized the hepatic damage induced by S. mansoni; circulating alanine aminotransferase and aspartate transaminase decreased, and total protein, albumin, and globulin serum levels increased. The serum level of malondialdehyde significantly declined, and catalase, glutathione peroxidase, superoxide dismutase, and total antioxidant capacity increased in SP-infected, MGT-infected, and SP–MGT-infected groups compared with the infected group. Co-administration of SP and MGT reduced serum cytokine levels (tumor necrosis factor-alpha, interferon-gamma, and interleukin-13) and increased interleukin-10 levels after S. mansoni infection compared with the infected group. Moreover, treatment with SP and/or MGT decreased the number of granulomas in hepatic and splenic tissues compared with the infected group. Collectively, our results suggest that combined SP and MGT treatment is effective for S. mansoni infection. Liver and spleen tissue alterations were improved, the antioxidant systems were stimulated, and the inflammatory response was suppressed. Further research is recommended to investigate the mechanisms of the combined SP and MGT treatment effects to facilitate the development of novel therapies against this disease.
Schistosomiasis is a debilitating waterborne disease caused by helminthic parasites belonging to the genus Schistosoma (1). Despite vigorous control efforts, it is still the most widespread tropical disease. Schistosomiasis, which has high morbidity, affects ~210 million people in ~76 countries, in addition to malaria and tuberculosis (2). Importantly, in Sub-Saharan Africa, 200,000 cases or more of schistosomiasis are reported annually (3). One of the causative agents of schistosomiasis is Schistosoma mansoni, which migrate from the blood to the lungs and liver and finally reside as male and female worms in the mesenteric veins. During the chronic phase of the disease, intense granulomatous lesions develop in the intestine, liver, lungs, brain, spleen, and pelvic organs (4). Drug-resistant Schistosoma strains have emerged in endemic areas due to the continuous and irregular use of the same chemotherapy against the disease (5). Thus, the development of new, dynamic, and safe antischistosomal drugs, particularly from natural plant extracts (6), is imperative.
Of note, a natural antioxidant and herbal medications are being evaluated. Recently, increased attention has been paid to the use of herbal medications as remedies for various parasitic diseases (7–10). Spirulina platensis (SP) is a natural spiral-shaped, multicellular photosynthetic, blue-green microalga (Cyanobacterium). It is commonly used as a nutritional supplement for both humans and animals (11, 12). SP has a high nutritional value; it is rich in carbohydrates (15–25%), proteins (55–70%), sterols, and polyunsaturated fatty acids (18%) (13, 14). SP contains phycocyanin, mixed carotenoids, phytonutrients, essential amino acids, gamma-linolenic acid, linoleic acid, and palmitic acid, which can improve the defense system and induce potent scavenging activities to reactive oxygen species (15, 16). Moreover, the regular consumption of SP can improve the hematological profile owing to the presence of calcium, iron, copper, magnesium, and several vitamins (B6 and B12 and folic acid) essential for hemopoiesis (17). As a food additive, SP has antitumor, antibacterial, antiviral, anticancer, anti-HIV, and anti-inflammatory activities (18–20).
Green tea (Camellia sinensis) is consumed worldwide, especially in Japan and China (21). Polyphenolic catechins, including epicatechin gallate, epicatechin, epigallocatechin, and epigallocatechin gallate, are the most abundant biologically active green tea components. These components have a potentially wide range of therapeutic benefits, including anti-inflammatory, antioxidant, anticancer, antidiabetic, anticholesterol, anti-obesity, anti-mutation, anthelminthic, and antimicrobial effects (22–25). The protection of tea leaves from sunlight increases their amino acid content, particularly theanine, which has a lower catechin content (26). Matcha is a finely powdered green tea manufactured from tea leaves cultivated under shade for almost 3 weeks before being harvested (27). In recent years, the consumption of matcha has increased instead of green tea. However, research on the protective effects of matcha is scarce. In addition, high caffeine levels were detected in matcha; the young leaves and buds of camellia plants have more caffeine than mature leaves (28). Given the information mentioned earlier, matcha is considered the best-grade green tea; it is rich in theanine and caffeine but little catechin contents, unlike the popular green tea (29). To the best of our knowledge, there is little data regarding the antioxidant and anti-inflammatory properties of matcha and SP against S. mansoni infection. Therefore, the present study was conducted to evaluate the therapeutic effects of SP and matcha green tea (MGT) on S. mansoni infection. The antioxidant and anti-inflammatory impacts of SP and matcha and their ability to protect the hepatic and splenic tissues after S. mansoni infection were investigated.
Ethical approval and protocol approval for this study were obtained from the Animal Ethics Committee of the Faculty of Veterinary Medicine, Kafrelsheikh University, Egypt. The study protocol complied with all the relevant Egyptian legislation concerning the publication and research. The ethical approval number is KFS-2019/2.
The SP algae were kindly supplied by the Department of Botany, Faculty of Science, Mansoura University (Mansoura, Egypt). The MGT powder was provided by Aiya Co., Ltd. (Aichi, Japan). The matcha consisted mainly of polyphenol (10.0%), caffeine (3.0%), fiber (39.0%), protein (31.0%), calcium (0.42%), potassium (2.7%), iron (0.02%), and vitamins (C, 0.06%; A, 0.005%; and carotene, 0.029%).
Schistosoma Biological Supply Program at Theodor Bilharz Research Institute (Giza, Egypt) provided Biomphalaria alexandrina snails that shed S. mansoni cercariae. Briefly, the infected snails were kept for approximately 4 weeks in a test tube containing distilled water and then exposed to artificial light at 28°C ± 1 for 2 h to induce shedding of cercariae. The number of cercariae was then counted under a microscope, and each animal was infected by a 1-h exposure of the tail to a suspension containing 100 ± 10 cercariae (86).
Male Swiss albino mice (n = 60), weighing 20 ± 25 g, were purchased from the Schistosome Biological Supply Program unit at Theodor Bilharz Research Institute (Giza, Egypt). Before administering the different treatments, the mice were quarantined in plastic cages for 2 weeks and provided with water and control diet ad libitum. During the experimental period, all animals were strictly handled under standard environmental conditions (12-h light/dark cycles; relative humidity, 60%; room temperature, 22 ± 2°C).
As presented in Figure 1, the animals were randomly allocated into the following six experimental groups (n = 10 per group):
1. Control group (CNT): mice received oral physiological saline every day and kept without infection.
2. SP–MGT group: mice were treated with 3 g/kg bodyweight of SP (30) suspended in sterile physiological saline (100 ml) plus 3 g/kg bodyweight/day of MGT (31) suspended in distilled water using a gastrostomy tube.
3. S. mansoni group (infected): mice were infected with S. mansoni cercariae (100 ± 10/mouse) using the tail immersion method.
4. SP-infected group: mice were infected with S. mansoni and treated orally with an SP suspension at the same dose as mentioned earlier.
5. MGT-infected group: mice were infected with S. mansoni and treated orally with MGT suspension as described earlier.
6. SP–MGT-infected group: mice were treated with a combination of SP and MGT after S. mansoni infection in the same manner and same doses as described earlier.
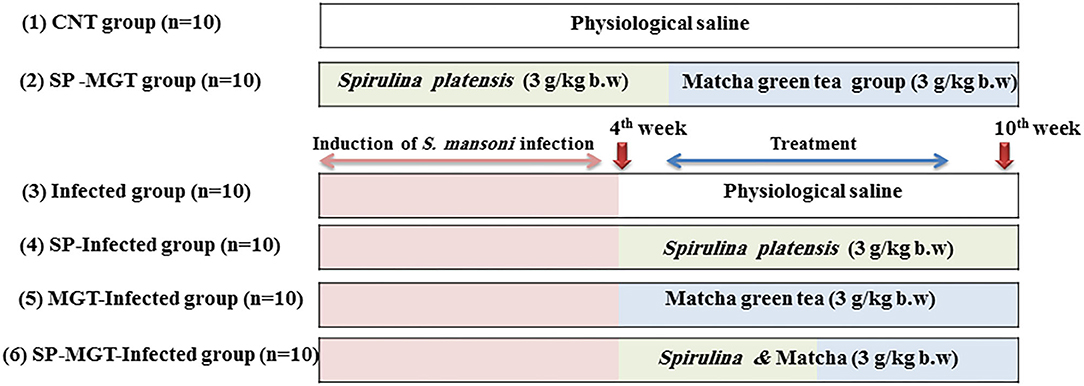
Figure 1. Time scheme, treatment protocol, and doses of used SP and MGT.
The treatment with SP and MGT started 4 weeks after S. mansoni infection and continued until the 10th week. Blood samples were collected from the retro-orbital venous sinus in plain test tubes 24 h after the last treatment. The samples were left to clot for approximately 15 min and then centrifuged at 3,000 rpm (4°C) to obtain the serum. The separated serum samples were kept at −80°C for further analysis. The mice were killed, and the liver and spleen tissues were collected and preserved in 10% neutral buffered formalin for histopathological assessment.
Serum alanine and aspartate aminotransferases (ALT and AST, respectively) activities (Randox, UK), as well as the total protein and albumin levels (Stanbio Laboratory, USA), were measured using the Photometer 5010V5+ (BM Co, Germany) following the standard protocols.
Malondialdehyde (MDA), total antioxidant capacity (TAC), catalase (CAT), superoxide dismutase (SOD), and glutathione peroxidase (GSH-Px) were measured in serum samples using commercial test kits obtained from Bio-Chain Inc., USA.
The serum levels of tumor necrosis factor-alpha (TNF-α), interferon-gamma (INF-γ), and interleukins (IL-10 and IL-13) were evaluated using commercial ready-made ELISA Kits (Quantikine Co., USA).
After storing the hepatic and splenic tissues in 10% formalin, the tissues were removed and immersed in serial ascending dilutions of ethanol. The tissue specimens were then dipped in paraffin to obtain the paraffin blocks. The blocks were cut into 5-μm-thick sections. Subsequently, the sections were stained with hematoxylin and eosin, as previously described by Bancroft and Layton (80).
Data were expressed as mean ± standard error of the mean (SEM). The differences among the biochemical parameters, antioxidant/oxidative stress, and inflammatory biomarkers were analyzed via one-way analysis of variance, followed by Duncan's comparisons tests. All statistical analyses were conducted using the SPSS software version 20 (version 20, USA). Differences were considered statistically significant at P < 0.05.
The effect of SP and/or MGT on liver activity was evaluated by measuring the serum levels of ALT, AST, total (T) protein, albumin, and globulins in S. mansoni-infected mice and shown in Table 1. S. mansoni infection (infected group) exhibited elevation of serum ALT and AST and reduction of the total protein, albumin, and globulin levels compared with the control group (P < 0.05). Contrarily, the concomitant administration of SP and/or MGT significantly reduced the hepatic damage as demonstrated in the SP-infected, MGT-infected, and SP–MGT-infected groups relative to the infected non-treated group (P < 0.05). Treatment with the combination of SP and MGT (SP–MGT-infected group) significantly restored serum total protein and globulin to concentrations similar to those of the uninfected controls (P < 0.05).

Table 1. Serum liver biomarkers in response to co-treatment with Spirulina platensis and/or matcha green tea in S. mansoni infected mice.
The antioxidant effects of SP and/or MGT against S. mansoni-mediated oxidative stress are presented in Figures 2, 3. Serum MDA was significantly elevated, and the CAT, SOD, and GSH-Px activities and TAC levels were significantly reduced in infected untreated mice compared with the control group. The treatment of S. mansoni-infected mice with SP and/or MGT significantly attenuated the changes in MDA, TAC, CAT, SOD, and GSH-Px activities (P < 0.05 vs. infected). In addition, SP–MGT-treated mice had higher activities of CAT, SOD, GSH-Px, and TAC compared with the control mice (P < 0.05).
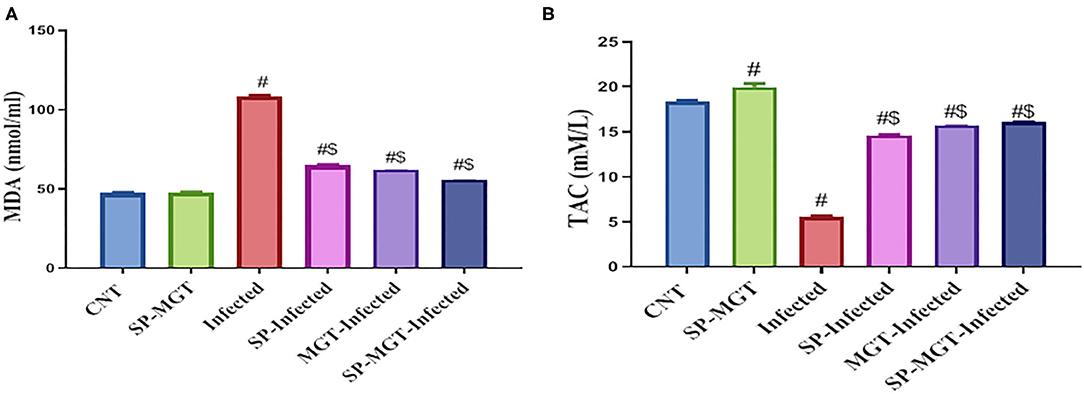
Figure 2. Malondialdehyde (MDA; A), and total antioxidant capacity (TAC; B) levels in serum of mice in response to co-treatment with Spirulina platensis and/or matcha green tea in S. mansoni infected mice. Data were displayed as mean ± SEM (n = 10/group). # or $ refers to the statistical significance at P < 0.05 against control and S. mansoni (infected) groups, respectively.
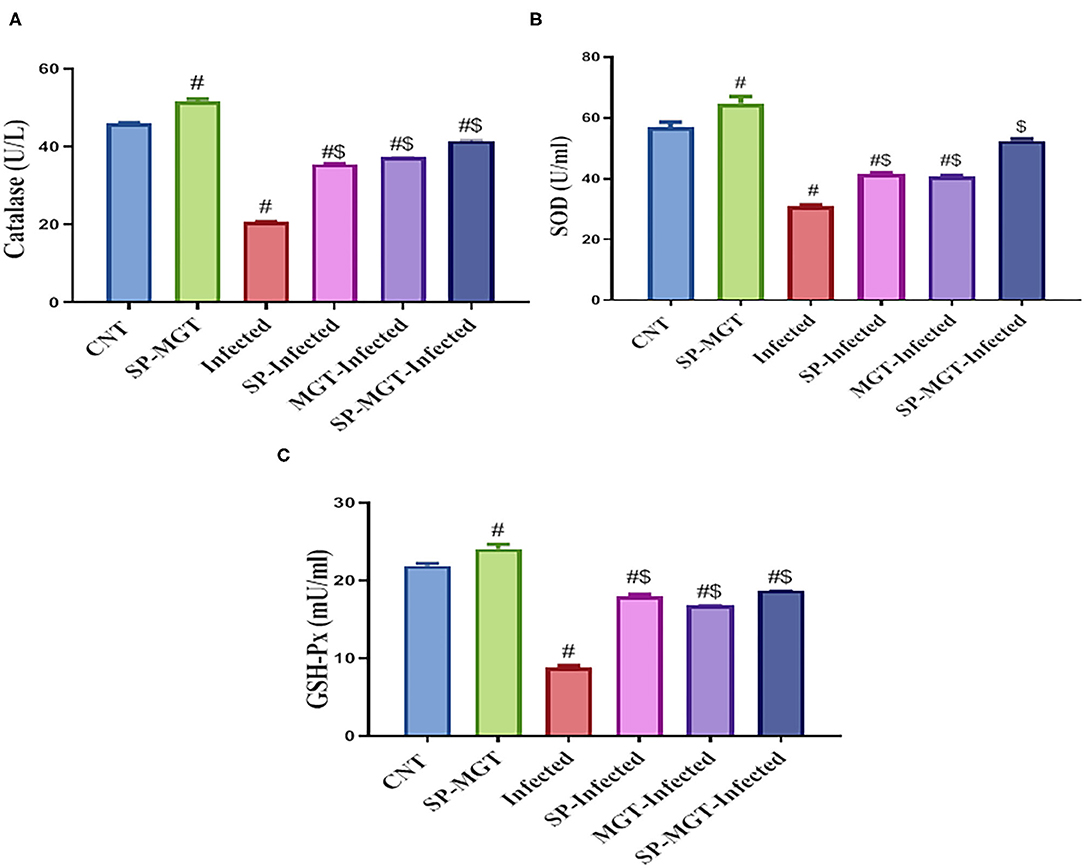
Figure 3. Catalase (CAT; A), superoxide dismutase (SOD; B), and glutathione peroxidase (GSH-Px; C) activities in serum of mice in response to co-treatment with Spirulina platensis and/or matcha green tea in S. mansoni infected mice. Data were displayed as mean ± SD (n = 10/group). # or $ refers to the statistical significance at P < 0.05 against control and S. mansoni (infected) groups, respectively.
The serum levels of TNF-α, IFN-γ, and IL-13 were significantly elevated, whereas that of IL-10 significantly reduced in S. mansoni-infected mice compared with the serum levels in the uninfected controls (Figure 4) (P < 0.05). Treatment with SP and/or MGT attenuated the changes in serum cytokines induced by S. mansoni infection; TNF-α, IFN-γ, and IL-13 were reduced, and IL-10 was significantly increased in SP- and/or MGT-treated infected mice compared with untreated infected mice (Figure 4) (P < 0.05).

Figure 4. Tumor necrosis factor-alpha (TNF-α; A), interferon-gamma (IFN-γ; B) levels, interleukin 10 (IL-10; C), and interleukin-13 (IL-13; D) in serum of mice in response to co-treatment with Spirulina platensis and/or matcha green tea in S. mansoni infected mice. Data were displayed as mean ± SEM (n = 10/group). # or $ refers to the statistical significance at P < 0.05 against control and S. mansoni (infected) groups, respectively.
The liver sections are presented in Figure 5. Control animals exhibited normal hepatocytes radiating in cords around the central vein. The hepatic tissues of animals treated with SP or MGT were normal. Schistosoma-infected animals exhibited numerous granulomatous lesions around the parasites and their eggs in the liver. The lesions were usually associated with marked inflammation and fibrosis, including infiltration of lymphocytes, macrophages, and large numbers of eosinophils. The number of granulomas decreased in infected animals treated with MGT compared with the number of lesions in infected untreated animals (Figure 6). Animals treated with SP exhibited smaller granulomas, mostly around the disintegrated parasite. Conversely, infected animals treated with both SP and MGT had fewer and smaller granulomas and mild periportal lymphocytic cell infiltration compared with untreated infected mice (P > 0.01). Interestingly, the combination group exhibited a significant decrease in these parameters in comparison with a single treatment of SP or MGT (P > 0.01).
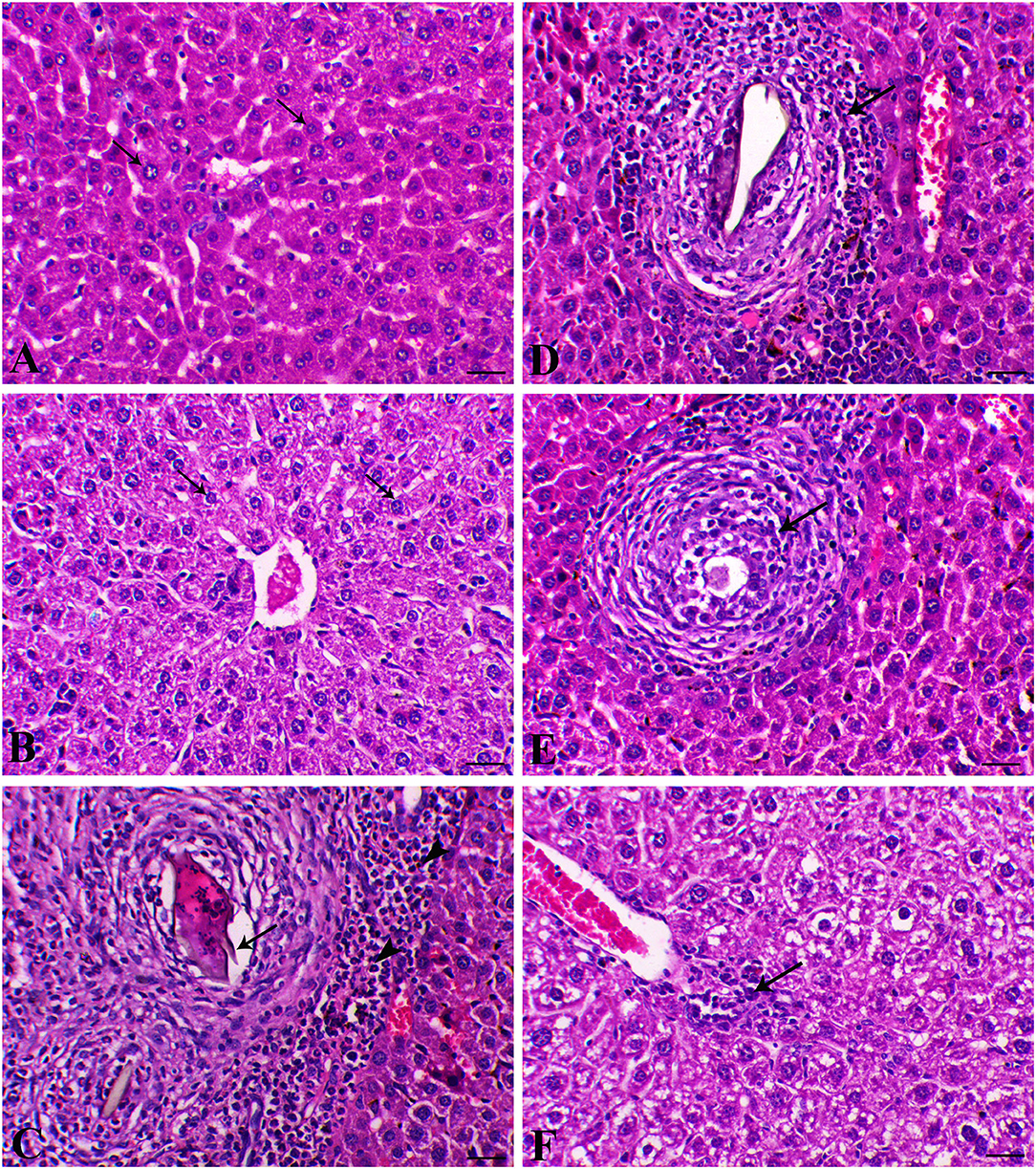
Figure 5. Hepatic histological sections of control group (A) (arrows indicate normal polygonal hepatic cell with central vesicular nucleus), SP–MGT (B) (arrows indicate normal hepatocytes), Infected group (C) (arrow indicates the schistosome egg, which revealing lateral spine, arrowheads indicates eosinophils), SP-infected group (D) (arrow indicates small-sized granuloma), MTG-infected group (E) (arrow indicates small-sized granuloma), and SP + MGT-infected group (F) (arrow indicates mild lymphocytic cells infiltration).
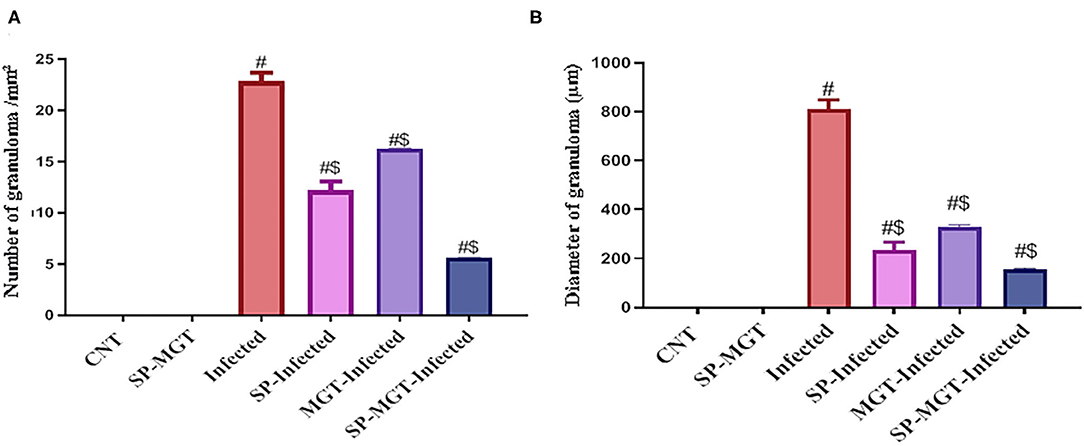
Figure 6. Quantitative scoring of number (A) and diameter (B) of granuloma in hepatic tissues. Data were displayed as mean ± SEM (n = 5/group). # or $ refers to the statistical significance at P < 0.05 against control and S. mansoni (infected) groups, respectively.
The splenic sections are presented in Figure 7. Control animals exhibited normal red and white pulps. Animals treated with only SP or MGT exhibited normal lymphoid cells around the central arteriole. Red pulp congestion associated with hemosiderin pigment deposition was observed in infected animals. In addition, a marked degree of lymphoid depletion accompanied by the appearance of reticular fibers was observed in the white pulp of infected mice. Schistosoma-infected mice treated with MGT exhibited marked decreases in lymphoid necrosis with lymphoid cell proliferation. Restoration of lymphoid follicles occurred in infected animals treated with SP, whereas in infected animals treated with both SP and MGT, the lymphoid follicles were normal.
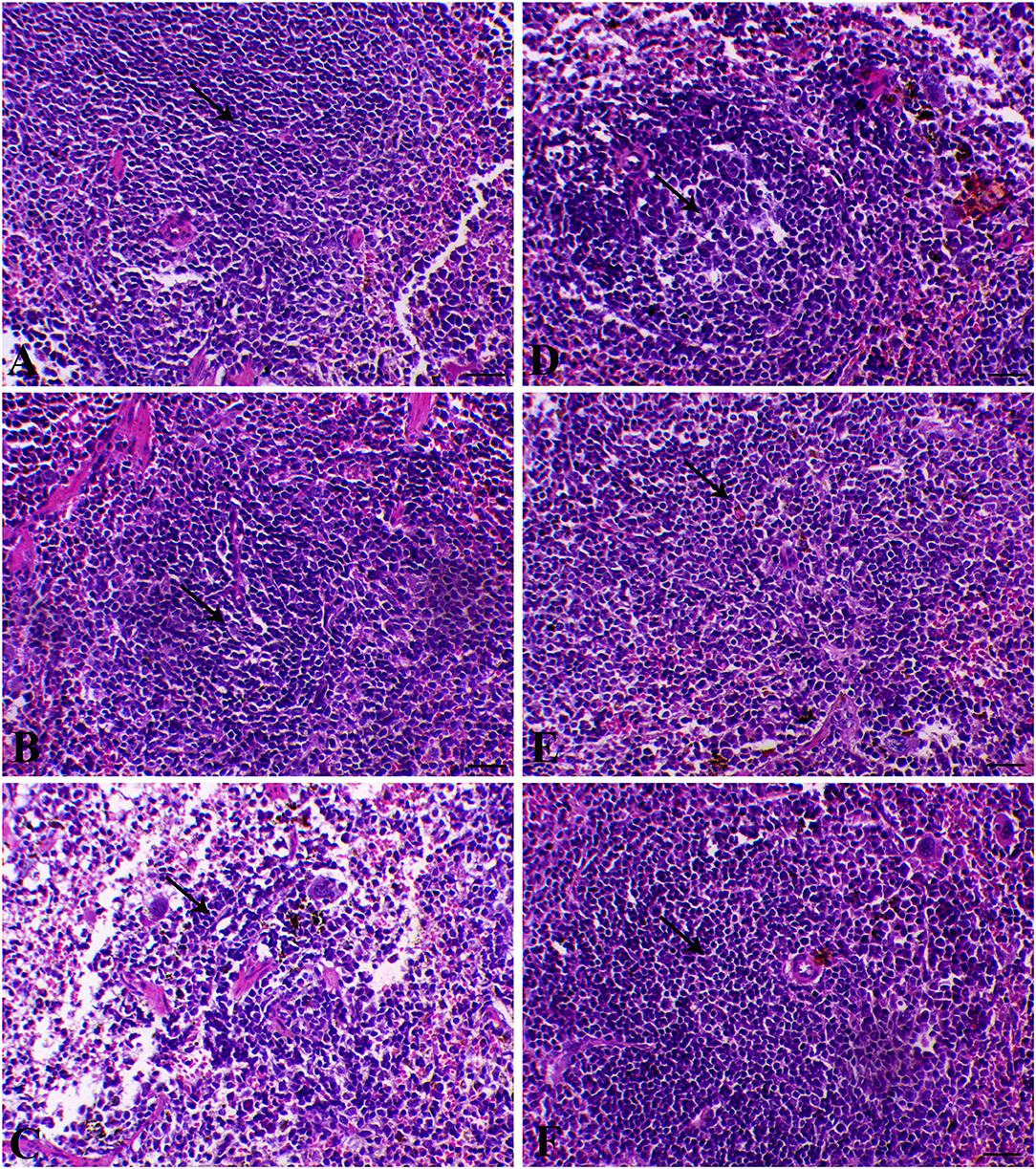
Figure 7. Splenic histological sections of control group (A) (arrow indicates normal lymphoid follicle), SP–MGT group (B) (arrow indicates normal lymphoid follicle), Infected group (C) (arrow indicates severe degree of lymphoid depletion), SP-infected group (D) (arrow indicates plenty of lymphoid cells), MTG-infected group (E) (arrow indicates an increase of lymphoid elements), and SP + MGT-infected group (F) (arrow indicates normal lymphoid pulp).
Schistosomiasis is one of the most widespread parasitic sicknesses. In addition to the high economic costs, schistosomiasis negatively impacts the well-being of the people in subtropical and tropical countries (32). In the last few decades, researchers have aimed to develop effective antischistosomal drugs with few adverse effects (33). However, this objective seems to be out of reach. Green tea is a widely consumed drink worldwide. It contains several active compounds, including polyphenols (tea catechins), methylxanthines, and essential oils (34). Previous reports have demonstrated the antiparasitic and biological activities of green tea (23, 35, 36). In this report, we demonstrated the antischistosomal activity of SP and MGT through their hepatosplenic antioxidant and anti-inflammatory protective effects.
S. mansoni infection causes hepatocellular injury and increases the circulating levels of hepatic enzymes, such as ALT and AST. In addition, it impairs protein synthesis (37). In our mouse model, the circulating levels of AST and ALT were significantly increased, whereas the total protein, albumin, and globulins were decreased after S. mansoni infection. Other investigators also illustrated hepatic damage mediated by S. mansoni infection (elevated ALT, AST, and ALP levels) in animal models (38–40). Liver damage also occurs, as indicated by the presence of inflammatory granulomas resulting from egg deposition as well as worms and their toxins (41). S. mansoni infection also leads to the reduction of the levels of total protein, albumin, and globulin (42–44). Moreover, hypoalbuminemia is associated with collagen deposition (40). Serum protein and albumin levels may also decrease with S. mansoni infection due to malabsorption after the extrusion of large egg numbers and intestinal mucosal damage or decreased synthesis due to hepatic cell injury (45). In our study, the serum biochemical data correlated with the histopathological results, and both confirmed liver damage.
Liver damage was resolved by the administration of SP and MGT, especially the combined treatment. Mohamed et al. (66) revealed that serum ALT, AST, and ALP activities were significantly reduced after treatment of S. mansoni-infected mice with blue-green algae, which is consistent with our results. The SP administration also significantly attenuated the elevated serum AST and ALT after doxorubicin treatment in rats; the presence of β-carotene in SP correlated with the decline in cell damage, particularly DNA damage (46). Al-Qahtani and Binobead (78) attributed the protective effects of SP against D-galactosamine-induced liver damage to the phenolic compounds in SP. SP can prevent hepatocyte deformations, inhibit hepatic enzymes, and prevent further cell damage. Numerous studies also revealed the protective potency of green tea toward various poisons and medicines, such as malathion (25), leflunomide (47), and micromycin (48). Our results were similar to the results of Dobrzynska et al. (81), Augustyniak et al. (79), El-Beshbishy et al. (60), and Miranda et al. (83). In these publications, green tea extracts lowered serum liver enzymes and protected liver cells from damage. In addition, co-administration of green tea ameliorated S. mansoni-induced liver damage in mice, as demonstrated by the significant decreases in ALT levels, the restoration of hepatocyte structure, and the decrease in perivascular collagen fibers (49).
A dynamic physiological connection exists between parasitic diseases and the antioxidant defense mechanisms of the infected host (50). Highly significant increases in hepatic lipid peroxides and glutathione depletion were found to occur in S. mansoni-infected mice (51). Interestingly, oxidative stress due to schistosomiasis increased the level of MDA (39) and reduced levels of SOD, CAT, and GSH-Px (41). Jatsa et al. (82) also demonstrated increased liver MDA concentration, with significant inhibition of glutathione concentration and CAT activity after schistosomiasis. The increased MDA levels might be due to the liberation of a huge amount of superoxide radicals by macrophages present in hepatic granulomas during S. mansoni infection. In addition, CAT is an endogenous antioxidant enzyme, and its depletion indicates an increase in the number of free radicals, inducing cellular damage (52). The cellular antioxidant system disturbances in schistosomiasis might arise in response to the stimulation of the host immune system, leading to the release of oxygen-derived free radicals as a premier non-specific defense response toward parasitic infection (53). Oxidative injury after S. mansoni infection was previously described, including increased MDA and NO levels and decreased GSH, CAT, and SOD activities (49, 54, 55). Our work confirmed the oxidative damage induced by schistosomiasis, which is indicated by significantly elevated serum levels of MDA and reduced CAT, SOD, and GSH-Px activities as well as TAC levels in S. mansoni-infected mice. The oxidative damage was reversed by SP and MGT supplementation.
The potential antioxidant activity of SP may be linked to its constituents, including chlorophyll, carotene, phenolic compounds, selenium, gamma-linolenic acids, and tocopherol. These constituents have the ability to protect against free radical-induced cellular transformation (56, 57). Another active component of SP is C-phycocyanin, which exhibited superoxide and hydroxyl radical scavenging activity and inhibition of lipid peroxidation (58). Moreover, pretreatment with SP reduces the toxic effects of cadmium and doxorubicin, as indicated by reduced MDA and NO levels and increased GSH and SOD levels in the liver tissue (46, 59). Among the major constituents of green tea are polyphenols, especially catechins, which prevent lipid peroxidation and have strong superoxide, hydrogen peroxide, and nitric acid scavenging activity (25, 60). Another active component of matcha is theanine, which can be converted to glutamine (29). Glutamate can alleviate mitochondrial damage, rejuvenate intermediate metabolites of the TCA cycle, and increase ATP production via the oxidative phosphorylation process to quench oxidative damage (61). Our findings are in agreement with a previous report demonstrating the role of green tea in quenching oxidative stress mediated by S. mansoni through the elevation of the TAC, SOD, and CAT activities in the liver and reduction of hepatic lipid peroxidation (49).
Of note, cytokines are critical in the pathogenesis of S. mansoni infection and are responsible for the extent of fibrosis and granuloma formation (62). The key mediator cytokine of liver fibrosis in S. mansoni infections is IL-13 (63). TNF-α participates in the maintenance of the granulomatous response (64). IFN-γ elicits free radical production, M1 macrophage expansion, and apoptosis (85). In addition, IL-10 plays an important role in the modulation of cytokine networks in schistosomiasis. The association between hepatic granuloma and fibrosis after S. japonicum infection and other mediators (IL-13 and tissue transglutaminases) has been reported (65). Silveira-Lemos et al. (84) also revealed that TNF-α and IL-10 modulate the development of granuloma in both humans and experimental schistosomiasis models. In line with these investigations, our study demonstrated that S. mansoni parasitic infection mediated the production of inflammatory fibrogenic factors via the elevation of serum TNF-α, IFN-γ, and IL-13 levels coupled with reduced IL-10 levels. Blue-green algae significantly reduced the serum levels of TNF-α in S. mansoni-infected mice (66). The anti-inflammatory effects of S. platensis were attributed to the inhibition of the NF-κB pathway and the subsequent suppression of pro-inflammatory cytokine production (67). Furthermore, phycocyanin, an active SP component, effectively limits the cyclooxygenase-2 inflammatory pathway and myeloperoxidase activity, scavenges free radicals, and inhibits lipid peroxidation (68). In this study, the inflammatory cytokines (TNF-α, IFN-γ, IL-10, and IL-13) returned to control serum levels after SP and/or MGT treatment of S. mansoni-infected mice, demonstrating the anti-inflammatory effects of SP and MGT. Green tea and green tea flower extracts downregulate the expression of inflammation markers, including IL-β1, IL-6, and TNFα, in human gingival epithelial keratinocytes and mouse liver treated with lipopolysaccharides (69, 70). Theanine, the main component of MGT, and its metabolites, such as glutamine and ethyleneimine, alleviate inflammation by reducing NF-κB activation and neutrophil accumulation in tissues and dampening the Ca2+ channels to block inflammatory mediator release (71).
Schistosomiasis causes the formation of hepatic granulomas and fibrosis, in addition to necrotic changes in the liver (33). In agreement with previous studies, our experiments revealed that chronic inflammation is among the pathological lineaments of S. mansoni, causing intense fibrosis in the infected tissues and organs (72). An increased number and diameter of granulomas and extensive fibrous tissue accumulation were detected in response to S. mansoni in several previous works (55, 73). In agreement with previous studies (74, 75), we detected parasitic egg granulomas with an increased spleen size after S. mansoni infection. SP has no harmful effects or organ toxicity; however, it remarkably reduced the incidence of liver tumors, indicating its prospective therapeutic effects on our model (56, 57). Surprisingly, green tea treatment of mice infected with S. mansoni caused an obvious refinement of most hepatocytes surrounding the blood vessels and reduced perivascular collagenous fibers, whereas some hepatocytes still exhibited cytoplasmic degeneration (49). The spleen and liver are connected via the portal vein system; thus, the spleen may contribute to the development of liver fibrosis through the infiltration of monocytes and cytokine production. As presented in our results, S. mansoni infection resulted in a series of histopathological changes in the spleen indicated by marked lymphoid necrosis within the white pulp associated with the infiltration of macrophages and histiocytes. Interestingly, treatment of mice with a combination of SP and MGT restored the normal histology of the spleen and lymphoid follicles. These changes are consistent with several previous reports demonstrating the antioxidant roles of SP and MGT in the restoration of lymphoid follicles and splenic pulps via altered inflammatory gene expression in the splenocytes (76, 77).
Taken together, the present findings emphasize that S. mansoni infection induces hepatosplenic injury in mice. Combined treatment with SP and MGT is a new favorable natural approach for minimizing the pathological alterations in the liver and spleen after S. mansoni infection, as indicated by restored antioxidant status, reduced inflammatory markers, and normal tissue architecture. Future investigations should consider the pharmacological potentials of SP and MGT, as well as other transduction pathways involved in their antischistosomal activity.
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author/s.
The animal study was reviewed and approved by the ethical approval and the protocol of this experiment were authorized by the Animal Ethical Committee of the Faculty of Veterinary Medicine, Kafrelsheikh University, Egypt, which complies with all relevant Egyptian legislations in publication and research. The ethical approval number is KFS-2019/2.
AR, EE, GE, and DE-S were involved in the conception of the research idea and participated in methodology design, supervision, and data analysis and interpretation. EE, GE, AS, SM, AA-B, and WA participated in methodology and data analysis. AR, EE, GE, WA, and DE-S drafted and prepared the manuscript for publication and revision. All authors contributed to the article and approved the submitted version.
This work was supported by the Taif University Researchers Supporting Program (project number: TURSP-2020/151), Taif University, Saudi Arabia. The funders had no role in data collection and analysis, decision to publish, or preparation of the manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors thank Taif University Researchers Supporting Program (project number: TURSP-2020/151), Taif University, Saudi Arabia, for their support.
SP, Spirulina platensis; MGT, matcha green tea; AST, aspartate transferase; ALT, alanine transferase; MDA, malondialdehyde; CAT, catalase; GSH-Px, glutathione peroxidase; SOD, superoxide dismutase; TAC, total antioxidant capacity; TNF-α, tumor necrosis factor; IFN-γ, interferon-gamma; IL-10, interleukin-6; IL-13, interleukin-13.
1. Tekwu EM, Bosompem KM, Anyan WK, Appiah-Opong R, Owusu KB-A, et al. In vitro assessment of anthelmintic activities of Rauwolfia vomitoria (Apocynaceae) stem bark and roots against parasitic stages of Schistosoma mansoni and cytotoxic study. J Parasitol Res. (2017) 2017:1–11. doi: 10.1155/2017/2583969
2. Steinmann P, Keiser J, Bos R, Tanner M, Utzinger J. Schistosomiasis and water resources development: systematic review, meta-analysis, and estimates of people at risk. Lancet Infect Dis. (2006) 6:411–25. doi: 10.1016/S.1473-3099(06)70521-7
3. Van der Werf MJ, de Vlas SJ, Brooker S, Looman CW, Nagelkerke NJ, Habbema JDF, et al. Quantification of clinical morbidity associated with schistosome infection in sub-Saharan Africa. Acta Trop. (2003) 86:125–39. doi: 10.1016/S0001-706X(03)00029-9
4. Elbaz T, Esmat G. Hepatic and intestinal schistosomiasis. J Adv Res. (2013). 4:445–52. doi: 10.1016/j.jare.2012.12.001
5. Lawn SD, Lucas SB, Chiodini PL. Case report: Schistosoma mansoni infection: failure of standard treatment with praziquantel in a returned traveller. Trans R Soc Trop Med Hyg. (2003) 97:100–1. doi: 10.1016/S0035-9203(03)90038-1
6. Hamed MA, Hetta MH. Efficacy of Citrus reticulata and Mirazid in treatment of Schistosoma mansoni. Mem Inst Oswaldo Cruz. (2005) 100:771–8. doi: 10.1590/S0074-02762005000700017
7. Kayser O, Kiderlen A, Croft S. Natural products as antiparasitic drugs. Parasitol Res. (2003) 90:S55–62. doi: 10.1007/s00436-002-0768-3
8. Elmahallawy EK, Jimenez-Aranda A, Martinez AS, Rodriguez-Granger J, Navarro-Alarcon M, Gutierrez-Fernandez J, et al. Activity of melatonin against Leishmania infantum promastigotes by mitochondrial dependent pathway. Chem Biol Interact. (2014) 220:84–93. doi: 10.1016/j.cbi.2014.06.016
9. Alkazzaz MA, Aziz ARA, Elmahallawy EK, Hassan AA. Hematological profile in schistosoma mansoni infected mice treated with commiphora molmol extract compared with praziquantel. PSM Biol Res. (2018) 3:77–84.
10. Elmahallawy EK, Elshopakey GE, Saleh AA, Agil A, El-Morsey A, El-Shewehy DMM, et al. S-Methylcysteine (SMC) Ameliorates Intestinal, Hepatic, and Splenic Damage Induced by Cryptosporidium parvum Infection Via Targeting Inflammatory Modulators and Oxidative Stress in Swiss Albino Mice. Biomedicines. (2020) 8:1–17. doi: 10.3390/biomedicines8100423
11. Chamorro G, Salazar M, Araujo KG, Dos Santos CP, Ceballos G, Castillo LF. Update on the pharmacology of Spirulina (Arthrospira), an unconventional food. Arch Latin Nutr. (2002) 52:232−40.
12. Mostafa SS, Gawish FA. Towards to control Biomphalaria alexandrina snails and the free living larval stages of Schistosoma mansoni using the microalga Spirulina platensis. Austr J Bas Appl Sci. (2009) 3:4112–9.
13. Ravi M, De SL, Azharuddin S, Paul SF. The beneficial effects of Spirulina focusing on its immunomodulatory and antioxidant properties. Nutr Dietary Suppl. (2010) 2:73–83. doi: 10.2147/NDS.S9838
14. Salama AF, Abdel-Hamid NM, El-Sheekh M, Tosson E, Gabr AM. Spirulina platensis microalgae protects against diethyl nitrosamine carcinogenic effect on female albino rats. Alex J Vet Sci. (2017) 53:167–79. doi: 10.5455/ajvs.263145
15. Simpore J, Kabore F, Zongo F, Dansou D, Bere A, Pignatelli S, et al. Nutrition rehabilitation of undernourished children utilizing Spiruline and Misola. Nutr J. (2006) 5:1–7. doi: 10.1186/1475-2891-5-3
16. Ambrozova JV, Misurcova L, Vicha R, Machu L, Samek D, Baron M, et al. Influence of extractive solvents on lipid and fatty acids content of edible freshwater algal and seaweed products, the green microalga Chlorella kessleri and the cyanobacterium Spirulina platensis. Molecules. (2014) 19:2344–60. doi: 10.3390/molecules19022344
17. Kaur K, Sachdeva R, Kochhar A. Effect of spirulina supplementation on the nutrient adequacy and health status of non-insulin-dependent diabetes mellitus (NIDDM) male subjects. Stud Ethno Med. (2009) 3:119–26. doi: 10.1080/09735070.2009.11886348
18. Francavilla M, Trotta P, Luque R. Phytosterols from Dunaliella tertiolecta and Dunaliella salina: a potentially novel industrial application. Bioresour Technol. (2010) 101:4144–50. doi: 10.1016/j.biortech.2009.12.139
19. Ibañez E, Cifuentes A. Benefits of using algae as natural sources of functional ingredients. J Sci Food Agric. (2013). 93:703–9. doi: 10.1002/jsfa.6023
20. Bashir S, Sharif MK, Javed MS, Amjad A, Khan AA, Shah, et al. Safety assessment of Spirulina platensis through sprague dawley rats modeling. Food Sci Technol. (2020) 40:376–81. doi: 10.1590/fst.41918
21. Emoto Y, Yoshizawa K, Kinoshita Y, Yuki M, Yuri T, Yoshikawa Y, et al. Green tea extract-induced acute hepatotoxicity in rats. J Toxicol Pathol. (2014) 27:163–74. doi: 10.1293/tox.2014-0007
22. Fujiki H, Suganuma M, Matsuyama S, Miyazaki K. Cancer prevention with green tea polyphenols for the general population, and for patients following cancer treatment. Curr Cancer Ther Rev. (2005) 1:109–14. doi: 10.2174/1573394052952474
23. Taha H, El-Khatib R, Khodair M. In vitro studies on the antiparasitic effects of some medicinal herbs on the growth behaviour of Fasciola gigantica and Eimeria stiedae. J Egypt Vet Med Assoc. (2006) 66:67–82.
24. Abdolahi M, Khordandi L, Ahrari K. The protective effect of green tea extract on acetaminophen induced nephro-toxicity in mice. J Arak Univ Med Sci. (2010) 13:90–6.
25. Raoofi R, Jahromi HK, Jahromi ZK, Abedi HA, Sameni H, Pourahmad M. Antioxidant effects of green-tea on biochemical and histopathological changes of liver in male rats poisoned by malathion insecticide. Int J Med Res Health Sci. (2016) 5:361–70.
26. Ashihara H. Occurrence, biosynthesis and metabolism of theanine (γ-glutamyl-L-ethylamide) in plants: a comprehensive review. Nat Prod Commun. (2015) 10:1934578X1501000525. doi: 10.1177/1934578X1501000525
27. Nakamura E, Tomita I, Matsuura T. Composition and functionality of “matcha” of different qualities. Jpn J Food Chem Saf. (2018) 25:7–14.
28. Ashihara H, Suzuki T. Distribution and biosynthesis of caffeine in plants. Front Biosci. (2004) 9:1864–76. doi: 10.2741/1367
29. Unno K, Furushima D, Hamamoto S, Iguchi K, Yamada H, Morita A, et al. Stress-reducing function of matcha green tea in animal experiments and clinical trials. Nutrients. (2018) 10:1468. doi: 10.3390/nu10101468
30. Hu J, Li Y, Pakpour S, Wang S, Pan Z, Liu J, et al. Dose effects of orally administered spirulina suspension on colonic microbiota in healthy mice. Front Cell Infect Microbiol. (2019) 9:243. doi: 10.3389/fcimb.2019.00243
31. Hamden K, Carrea U SE, Ayadi Marki F, Masmoudi H, El Feki A. Positive effects of green tea on hepatic dysfunction, lipid peroxidation and antioxidant defence depletion induced by cadmium. Biol Res. (2008) 41:331–9. doi: 10.4067/S0716-97602008000300009
32. WHO. Inter-Country Meeting on Strategies to Eliminate Schistosomiasis From 621 the Eastern Mediterranean region. 2007, Muscat (2018).
33. El-Banhawey MA, Ashry MA, EL-Ansary AK, Aly SA. Effect of Curcuma longa or parziquantel on Schistosoma mansoni infected mice liver—histological and histochemical study. Indian J Exp Biol. (2007) 45:877–89.
34. Paveto C, Güida MC, Esteva MI, Martino V, Coussio J, Flawi, á MM, et al. Anti-Trypanosoma cruzi activity of green tea (Camellia sinensis) catechins. Antimicrob Agents Chemother. (2004) 48:69–74. doi: 10.1128/AAC.48.1.69-74.2004
35. Molan A, Sivakumaran S, Spencer P, Meagher L. Green tea flavan-3-ols and oligomeric proanthocyanidins inhibit the motility of infective larvae of Teladorsagia circumcincta and Trichostrongylus colubriformis in vitro. Res Vet Sci. (2004) 77:239–43. doi: 10.1016/j.rvsc.2004.04.010
36. Murthy PK, Joseph SK. Plant products in the treatment and control of filariasis and other helminth infections and assay systems for antifilarial/anthelmintic activity. Planta Med. (2011) 77:647–61. doi: 10.1055/s-0030-1250452
37. Silva FL, Del-Rei RP, Fraga DB, Leony LM, Souza AM, Santos FL. Alterations in the lipid profiles and circulating liver enzymes in individuals infected by Schistosoma mansoni. Rev Soc Bras Med Trop. (2018) 51:795–801. doi: 10.1590/0037-8682-0113-2018
38. Mahmoud M, El-Abhar H, Saleh S. The effect of Nigella sativa oil against the liver damage induced by Schistosoma mansoni infection in mice. J Ethnopharmacol. (2002) 79:1–11. doi: 10.1016/S0378-8741(01)00310-5
39. Botros SS, Mahmoud MR, Moussa MM, Nosseir MM. Immunohistopathological and biochemical changes in Schistosoma mansoni-infected mice treated with artemether. J Infect. (2007) 55:470–7. doi: 10.1016/j.jinf.2007.07.022
40. El-Lakkany NM, Hammam OA, El-Maadawy WH, Badawy AA, Ain-Shoka AA, Ebeid FA. Anti-inflammatory/anti-fibrotic effects of the hepatoprotective silymarin and the schistosomicide praziquantel against Schistosoma mansoni-induced liver fibrosis. Parasit Vect. (2012) 5:9. doi: 10.1186/1756-3305-5-9
41. Dkhil MA. Role of berberine in ameliorating Schistosoma mansoni-induced hepatic injury in mice. Biol Res. (2014) 47:8. doi: 10.1186/0717-6287-47-8
42. El-Rigal N-S, Hetta MH. Effect of Citrus reticulata on serum protein fractions of mice after Schistosoma mansoni infection. JApSc. (2006) 6:1447–55. doi: 10.3923/jas.2006.1447.1455
43. Mohammed EH, Eltayeb M, Ibrahim H. Haematological and biochemical morbidity of Schistosoma haematobium in school children in Sudan. Sultan Qaboos Univ Med J. (2006) 6:59.
44. Barros L, Costa-Silva M, Biolchini C, Neves R, Machado-Silva J. Effect of praziquantel administration on hepatic stereology of mice infected with Schistosoma mansoni and fed a low-protein diet. Braz J Med Biol Res. (2009) 42:812–5. doi: 10.1590/S0100-879X2009000900007
45. Oliveira SA, Souza BSF, Guimarães-Ferreira CA, Barreto ES, Souza SC, Freitas LAR, et al. Therapy with bone marrow cells reduces liver alterations in mice chronically infected by Schistosoma mansoni. World J Gastroenterol. (2008) 14:5842. doi: 10.3748/wjg.14.5842
46. Hassanen MR, Mahfouz MK, Farid AS, Fadlullah AH. Biochemical effects of spirulina platensis against oxidative stress caused by doxorubicin. Benha Vet Med J. (2015) 28:147–54. doi: 10.21608/bvmj.2015.32493
47. Chen JH, Tipoe GL, Liong EC, So HS, Leung KM, Tom WM, et al. Green tea polyphenols prevent toxin-induced hepatotoxicity in mice by down-regulating inducible nitric oxide–derived prooxidants. Am J Clin Nutr. (2004) 80:742–51. doi: 10.1093/ajcn/80.3.742
48. Xu C, Shu WQ, Qiu ZQ, Chen JA, Zhao Q, Cao J. Protective effects of green tea polyphenols against subacute hepatotoxicity induced by microcystin-LR in mice. Environ Toxicol Pharmacol. (2007) 24:140–8. doi: 10.1016/j.etap.2007.04.004
49. Dajem SMB, Shati AA, Adly MA, Ahmed OM, Ibrahim EH, Mostafa OM. Green tea (Camellia sinesis) ameliorates female Schistosoma mansoni-induced changes in the liver of Balb/C mice. Saudi J Biol Sci. (2011) 18:361–8. doi: 10.1016/j.sjbs.2011.06.003
50. Connors VA, Deburon I, Granath W. Schistosoma mansoni: interleukin-1 increases phagocytosis and superoxide production by hemocytes and decreases output of cercariae in schistosome-susceptible Biomphalaria glabrata. Exp Parasitol. (1995) 80:139–48. doi: 10.1006/expr.1995.1016
51. Rizk M, Fayed T, Badawy M, El-Regal NS. Effect of different durations of Schistosoma mansoni infection on the levels of some antioxidants in mice. Med J Islam World Acad Sci. (2006) 16:25−34.
52. El-Rigal N, Metwally N, Mohamed A, Mohamed N, Rizk M. Protection against oxidative damage induced by Schistosoma mansoni using susceptible/resistant nucleoproteins from Biomphalaria alexandrina snails. Asian J Biol Sci. (2011) 4:445–56. doi: 10.3923/ajbs.2011.445.456
53. Fahmy SR, Rabia I, Mansour EM. The potential role of mefloquine against Schistosoma mansoni infection by prohibition of hepatic oxidative stress in mice. J Basic Appl Zool. (2014) 67:40–7. doi: 10.1016/j.jobaz.2014.09.002
54. Essam AM, Ashraf AE. Effect of curcumin on hematological, biochemical and antioxidants parameters in Schistosoma mansoni infected mice. Int J Sci. (2013) 2:1–14.
55. Kadry SM, Mohamed AM, Farrag EM, Fayed DB. Influence of some micronutrients and Citharexylum quadrangular extract against liver fibrosis in Schistosoma mansoni infected mice. Afr J Pharm Pharmacol. (2013). 7:2628–38. doi: 10.5897/AJPP12.620
56. Upasani C, Khera A, Balararnan R. Effect of lead with vitamin E. C, or Spirulina on malondialdehyde, conjugated dienes and hydroperoxides in rats. Indian J Exp Biol. (2001) 39:70–4.
57. Ismail MF, Ali DA, Fernando A, Abdraboh ME, Gaur RL, Ibrahim WM, et al. Chemoprevention of rat liver toxicity and carcinogenesis by Spirulina. Int J Biol Sci. (2009) 5:377. doi: 10.7150/ijbs.5.377
58. Pham TX, Park, Y.-K., Bae M, Lee J.-Y. The potential role of an endotoxin tolerance-like mechanism for the anti-inflammatory effect of Spirulina platensis organic extract in macrophages. J Med Food. (2017) 20:201–10. doi: 10.1089/jmf.2016.0119
59. Karadeniz A, Cemek M, Simsek N. The effects of Panax ginseng and Spirulina platensis on hepatotoxicity induced by cadmium in rats. Ecotoxicol Environ Saf. (2009) 72:231–5. doi: 10.1016/j.ecoenv.2008.02.021
60. El-Beshbishy HA, Tork OM, El-Bab MF, Autifi MA. Antioxidant and antiapoptotic effects of green tea polyphenols against azathioprine-induced liver injury in rats. Pathophysiology. (2011) 18:125–35. doi: 10.1016/j.pathophys.2010.08.002
61. Van Wyngene L, Vandewalle J, Libert C. Reprogramming of basic metabolic pathways in microbial sepsis: therapeutic targets at last? EMBO Mol Med. (2018) 10:e8712. doi: 10.15252/emmm.201708712
62. Aly IR, Hendawy MA, Ali E, Hassan E, Nosseir MM. Immunological and parasitological parameters after treatment with dexamethasone in murine Schistosoma mansoni. Mem Inst Oswaldo Cruz. (2010) 105:729–35. doi: 10.1590/S0074-02762010000600001
63. Chiaramonte MG, Cheever AW, Malley JD, Donaldson DD, Wynn TA. Studies of murine schistosomiasis reveal interleukin-13 blockade as a treatment for established and progressive liver fibrosis. Hepatology. (2001) 34:273–82. doi: 10.1053/jhep.2001.26376
64. de Jesus AR, Magalhaes A, Miranda DG, Miranda RG, Araújo MI, de Jesus AA, et al. Association of type 2 cytokines with hepatic fibrosis in human Schistosoma mansoni infection. Infect Immunity. (2004) 72:3391–7. doi: 10.1128/IAI.72.6.3391-3397.2004
65. Kim Y, Eom S, Kim K, Lee YS, Choe J, Hahn JH, et al. Transglutaminase II interacts with rac1, regulates production of reactive oxygen species, expression of snail, secretion of Th2 cytokines and mediates in vitro and in vivo allergic inflammation. Mol Immunol. (2010) 47:1010–22. doi: 10.1016/j.molimm.2009.11.017
66. Mohamed AH, Osman GY, Salem TA, Elmalawany AM. The hepatoprotective activity of blue green algae in Schistosoma mansoni infected mice. Exp Parasitol. (2014) 145:7–13. doi: 10.1016/j.exppara.2014.06.016
67. de Freitas Brito A, Silva AS, de Oliveira CVC, de Souza AA, Ferreira PB, de Souza ILL, et al. Spirulina platensis prevents oxidative stress and inflammation promoted by strength training in rats: dose-response relation study. Sci Rep. (2020) 10:1–8. doi: 10.1038/s41598-020-63272-5
68. Wu Q, Liu L, Miron A, Klímov, á B, Wan D, Kuča K. The antioxidant, immunomodulatory, and anti-inflammatory activities of Spirulina: an overview. Arch Toxicol. (2016) 90:1817–40. doi: 10.1007/s00204-016-1744-5
69. Chen BT, Li WX, He RR, Li YF, Tsoi B, Zhai YJ, et al. Anti-inflammatory effects of a polyphenols-rich extract from tea (Camellia sinensis) flowers in acute and chronic mice models. Oxid Med Cell Longevity. (2012) 2012:1–7. doi: 10.1155/2012/537923
70. Hagiu A, Attin T, Schmidlin PR, Ramenzoni LL. Dose-dependent green tea effect on decrease of inflammation in human oral gingival epithelial keratinocytes: in vitro study. Clin Oral Investig. (2020) 24:2375–83. doi: 10.1007/s00784-019-03096-4
71. Malkoç M, Patan H, Yaman SÖ, Türedi S, Kerimoglu G, Kural BV, et al. l-theanine alleviates liver and kidney dysfunction in septic rats induced by cecal ligation and puncture. Life Sci. (2020) 249:117502. doi: 10.1016/j.lfs.2020.117502
72. Meneghin A, Hogaboam CM. Infectious disease, the innate immune response, and fibrosis. J Clin Invest. (2007) 117:530–8. doi: 10.1172/JCI30595
73. El-Lakkany N, Seif el-Din SH, Badawy AA, Ebeid FA. Effect of artemether alone and in combination with grapefruit juice on hepatic drug-metabolizing enzymes and biochemical aspects in experimental Schistosoma mansoni. Int J Parasitol. (2004) 34:1405–12. doi: 10.1016/j.ijpara.2004.08.012
74. da Silva AM, Corrêa CL, Neves RH, Machado-Silva JR. A high-fat diet associated with acute schistosomiasis mansoni causes disorganization in splenic architecture in mice. Exp Parasitol. (2012) 132:193–9. doi: 10.1016/j.exppara.2012.06.015
75. Bauomy AA, Dkhil MA, Diab MS, Amer OS, Zrieq RM, Al-Quraishy S. Response of spleen and jejunum of mice infected with Schistosoma mansoni to Mulberry treatment. Pak J Zool. (2014) 46:753–61.
76. Pham TX, Lee Y, Bae M, Hu S, Kang H, Kim MB, et al. Spirulina supplementation in a mouse model of diet-induced liver fibrosis reduced the pro-inflammatory response of splenocytes. Br J Nutr. (2019) 121:748–55. doi: 10.1017/S0007114519000126
77. Jakubczyk K, Kochman J, Kwiatkowska A, Kaldunska J, Dec K, Kawczuga D, et al. Antioxidant properties and nutritional composition of matcha green tea. Foods. (2020) 9:1–10. doi: 10.3390/foods9040483
78. Al-Qahtani WH, Binobead MA. Anti-inflammatory, antioxidant and antihepatotoxic effects of Spirulina platensis against d-galactosamine induced hepatotoxicity in rats. Saudi J Biol Sci. (2019) 26:647–52. doi: 10.1016/j.sjbs.2018.01.003
79. Augustyniak A, Waszkiewicz E, Skrzydlewska E. Preventive action of green tea from changes in the liver antioxidant abilities of different aged rats intoxicated with ethanol. Nutrition. (2005) 21:925–32. doi: 10.1016/j.nut.2005.01.006
80. Bancroft J, Layton C. The Hematoxylins and Eosin. Bancroft's Theory and Practice of Histological Techniques. Elsevier (2013).
81. Dobrzynska I, Sniecinska A, Skrzydlewska E, Figaszewski Z. Green tea modulation of the biochemical and electric properties of rat liver cells that were affected by ethanol and aging. Cell Mol Biol Lett. (2004) 4:709–21.
82. Jatsa HB, Kenfack CM, Simo DN, Feussom NG, Nkondo ET, Tchuente, et al. Schistosomicidal, hepatoprotective and antioxidant activities of the methanolic fraction from Clerodendrum umbellatum Poir leaves aqueous extract in Schistosoma mansoni infection in mice. BMC Comp Altern Med. (2015) 15:248. doi: 10.1186/s12906-015-0788-z
83. Miranda DD, Arçari DP, Pedrazzoli J, Carvalho P d.O., Cerutti SM, et al. Protective effects of mate tea (Ilex paraguariensis) on H2O2-induced DNA damage and DNA repair in mice. Mutagenesis. (2008) 23:261–5. doi: 10.1093/mutage/gen011
84. Silveira-Lemos D, Teixeira-Carvalho A, Martins-Filho OA, Oliveira LFA, Costa-Silva MF, Matoso LF, et al. Eosinophil activation status, cytokines and liver fibrosis in Schistosoma mansoni infected patients. Acta Trop. (2008) 108:150–9. doi: 10.1016/j.actatropica.2008.04.006
85. Vartanian T, Li Y, Zhao M, Stefansson K. Interferon-gamma-induced oligodendrocyte cell death: implications for the pathogenesis of multiple sclerosis. Mol Med. (1995) 1:732. doi: 10.1007/BF03401888
Keywords: Spirulina platensis, matcha green tea, Schistosoma mansoni, oxidative stress, inflammation
Citation: Ramez AM, Elmahallawy EK, Elshopakey GE, Saleh AA, Moustafa SM, Al-Brakati A, Abdo W and El-Shewehy DMM (2021) Hepatosplenic Protective Actions of Spirulina platensis and Matcha Green Tea Against Schistosoma mansoni Infection in Mice via Antioxidative and Anti-inflammatory Mechanisms. Front. Vet. Sci. 8:650531. doi: 10.3389/fvets.2021.650531
Received: 07 January 2021; Accepted: 23 March 2021;
Published: 30 April 2021.
Edited by:
Jiyu Zhang, Lanzhou Institute of Husbandry and Pharmaceutical Sciences, Chinese Academy of Agricultural Sciences (CAAS), ChinaReviewed by:
Colin Matthew Fitzsimmons, University of Cambridge, United KingdomCopyright © 2021 Ramez, Elmahallawy, Elshopakey, Saleh, Moustafa, Al-Brakati, Abdo and El-Shewehy. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ehab Kotb Elmahallawy, ZWVoYWFAdW5pbGVvbi5lcw==
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.