 Ana M. Palomar1*
Ana M. Palomar1* Jesús Veiga2
Jesús Veiga2 Aránzazu Portillo1
Aránzazu Portillo1 Sonia Santibáñez1
Sonia Santibáñez1 Radovan Václav3
Radovan Václav3 Paula Santibáñez1
Paula Santibáñez1 José A. Oteo1
José A. Oteo1 Francisco Valera2
Francisco Valera2- 1Centre of Rickettsiosis and Arthropod-Borne Diseases, Hospital Universitario San Pedro-Center for Biomedical Research of La Rioja (CIBIR), Logroño, Spain
- 2Departamento de Ecología Funcional y Evolutiva, Estación Experimental de Zonas Áridas -Consejo Superior de Investigaciones Científicas (EEZA-CSIC), Ctra. de Sacramento s/n, La Cañada de San Urbano, Almería, Spain
- 3Institute of Zoology, Slovak Academy of Sciences, Bratislava, Slovakia
The knowledge of the distribution, richness and epidemiological importance of soft ticks of the genus Argas is incomplete. In Spain, five Argas species have been recorded, including three ornitophilic nidicolous ticks, but their associated microorganisms remain unknown. This study aimed to investigate ticks from bird nests and their microorganisms. Ticks were collected extensively from natural cavities and nest-boxes used by European rollers (Coracias garrulus) and little owls (Athene noctua) in Southeastern and Central Spain. Ticks were morphologically and genetically identified and corresponding DNA/RNA tick extracts were analyzed [individually (n = 150) or pooled (n = 43)] using specific PCR assays for bacteria (Anaplasmataceae, Bartonella, Borrelia, Coxiella/Rickettsiella, and Rickettsia spp.), viruses (Flaviviruses, Orthonairoviruses, and Phenuiviruses), and protozoa (Babesia/Theileria spp.). Six Argas genotypes were identified, of which only those of Argas reflexus (n = 8) were identified to the species level. Two other genotypes were closely related to each other and to Argas vulgaris (n = 83) and Argas polonicus (n = 33), respectively. These two species have not been previously reported from Western Europe. Two additional genotypes (n = 4) clustered with Argas persicus, previously reported in Spain. The remaining genotype (n = 22) showed low sequence identity with any Argas species, being most similar to the African Argas africolumbae. The microbiological screening revealed infection with a rickettsial strain belonging to Rickettsia fournieri and Candidatus Rickettsia vini group in 74.7% of ticks, mainly comprising ticks genetically related to A. vulgaris and A. polonicus. Other tick endosymbionts belonging to Coxiella, Francisella and Rickettsiella species were detected in ten, one and one tick pools, respectively. In addition, one Babesia genotype, closely related to avian Babesia species, was found in one tick pool. Lastly, Anaplasmataceae, Bartonella, Borrelia, and viruses were not detected. In conclusion, five novel Argas genotypes and their associated microorganisms with unproven pathogenicity are reported for Spain. The re-use of nests between and within years by different bird species appears to be ideal for the transmission of tick-borne microorganisms in cavity-nesting birds of semiarid areas. Further work should be performed to clarify the taxonomy and the potential role of soft Argas ticks and their microorganisms in the epidemiology of zoonoses.
Introduction
Soft ticks of the genus Argas Latreille, 1795 (Ixodida; Argasidae) are distributed worldwide and include around 60 species (1). Of them, only eight species have been described in the Western Palearctic region, specifically, Argas gilcolladoi, Argas persicus, Argas reflexus, Argas transgariepinus, Argas vespertilionis, Argas macrostigmatus, Argas vulgaris, and Argas polonicus (2–5). All but the latter three species have been reported in Spain (Southwestern Europe) as parasites of birds or bats (6). The majority of Argas spp. are nidicolous and birds are exclusive vertebrate hosts for several species, mainly those of Persicargas subgenera, while humans are accidental hosts (7, 8). The genus Argas includes species responsible for the transmission of pathogens of medical and veterinary interest. Apart from conditions caused directly by soft ticks, such as toxicosis and anaphylaxis (9, 10), these ticks carry microorganisms that could be agents of infectious diseases. Specifically, Argas species can vector bacterial pathogens such as Borrelia anserina and Aegyptianella spp. and viruses such as Issyk-kul virus (11, 12). Other microorganisms with unproved pathogenicity have been detected in Argas ticks: bacteria from genera Anaplasma, Bartonella, Borrelia, Coxiella, Ehrlichia, Francisella, Rickettsia, and Rickettsiella, viruses belonging to Flaviviridae, Orthomyxoviridae, Orthonairoviridae, Phenuiviridae, and Reoviridae families, and protozoans such as Babesia and Hemolivia spp. (7, 13–17).
The lack of information on the natural history and distribution of various Argas species, their incorrect or incomplete taxonomic description, and the fact that some species share morphological features but have not been molecularly examined, are responsible for the poor knowledge of Argas ticks in Spain. Moreover, their role in the epidemiology of tick-borne microorganisms has not been studied in this country. Here, we aim at describing soft ticks from natural and artificial nests occupied by different cavity-nesting birds in Spain and the prevalence of selected tick-borne microorganisms.
Materials and Methods
Study Area and Study System
The main study area (~50 km2) lies in the Desert of Tabernas (Almería province, SE Spain, 37.08°N, 2.35°W). The landscape mostly consists of open shrubland with olive and almond groves interspersed among numerous dry riverbeds—ramblas. The climate in this area is semiarid Mediterranean with a marked water deficit during long, hot summer months. The mean annual rainfall is ~ 230 mm, with high inter- and intra-annual variability (18). Tick samples also were collected in Segovia and Guadalajara provinces (both in the interior of the Iberian Peninsula), whose climate is Mediterranean with some continental characteristics.
In the main study area in Almería, natural cavities in sandstone cliffs, seminatural cavities in stone bridges and abandoned farmhouses and nest boxes provide nest sites for cavity-nesting birds, namely the European roller (Coracias garrulus, hereafter roller), the little owl (Athene noctua) and the rock/feral pigeon (Columba livia, hereafter pigeon). In this study, we sampled ticks in cavities occupied by rollers and little owls. The roller is a migratory bird that arrives at its breeding grounds in the study area during the second fortnight of April whereas the little owl is a resident bird. Both species rear a single brood per year (19). In contrast to these species, the pigeon is a resident bird that breeds at any time of the year in our study area and does not use nest boxes. Other species breeding in natural and seminatural cavities mainly include jackdaws (Corvus monedula), and common kestrels (Falco tinnunculus), whereas Scops owls (Otus scops), spotless starlings (Sturnus unicolor), and house sparrows (Passer domesticus) can breed in nest boxes.
Given nest-site limitation in the study area, both intra- and interspecific competition for suitable nesting holes occur and individual cavities can be re-used by different species both within and between years. This is frequently the case in Almería, so that many samples were collected from natural and seminatural cavities of rollers and little owls previously used by pigeons. The samples from Segovia were collected from a natural tree hole occupied by rollers but excavated by the Iberian green woodpecker (Picus sharpei), whereas the samples from Guadalajara were taken from rollers breeding in nest boxes.
Tick Collection and Preservation
In the framework of a long-term project of cavity-nesting birds in the Desert of Tabernas, cavities and nest boxes have been routinely inspected during each breeding season since 2005 and both nestlings and nest material periodically examined for ectoparasites. Ticks were collected from cavities occupied by breeding rollers and little owls during 2009, 2012, 2015, and 2018–2020. Additionally, four tick individuals were obtained from roller nests in Central Spain (Guadalajara and Segovia) in 2004 (Table 1). Ticks collected until 2018 were preserved in ethanol, while ticks obtained in 2019 and 2020 were kept fresh upon delivery to the Centre of Rickettsiosis and Arthropod-borne Diseases (CRETAV). Before frozen at −80°C until later analysis, fresh ticks were identified and a single leg of each specimen was dissected.
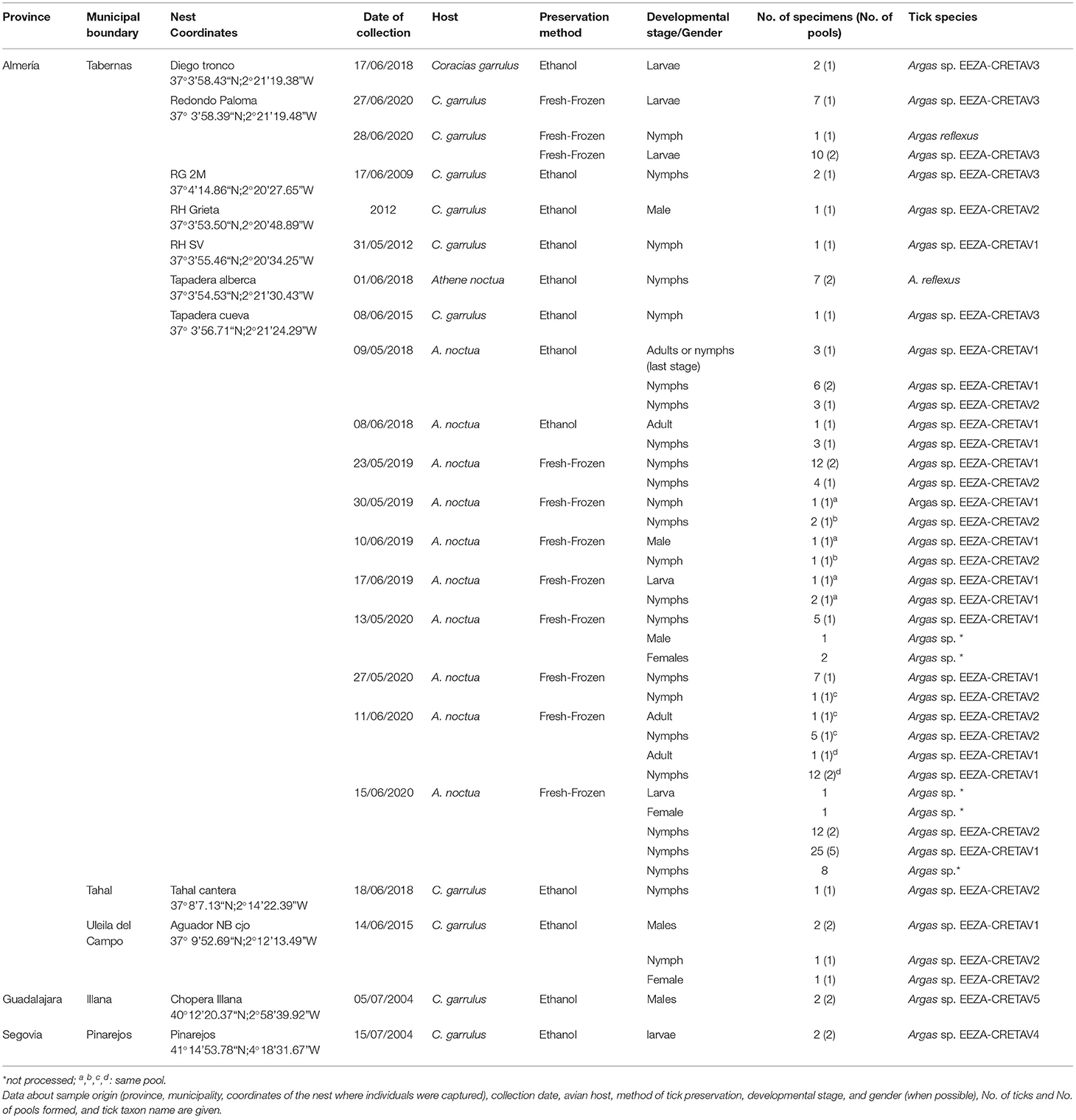
Table 1. Tick samples used in this study.
Tick Identification
The taxonomic identification of the ticks was carried out using morphological keys (20–23). Tick individuals were surface-sterilized and DNA was individually extracted from a single leg of each tick specimen using incubations with ammonium hydroxide (24). The obtained DNA templates were used for genetic characterization by the amplification and sequencing of the 16S rRNA fragment gene (25). Two other mitochondrial genes, 12S rRNA, and cytochrome oxidase subunit I (COI), were also used in analyses (Supplementary Table 1).
Microbial Screening
Ticks were pooled (from 1 to 7 specimens; whole larvae and body halves for the other life stages) according to tick species or genotype, origin and date of collection and, when possible, tick developmental stage. DNA extracts from pools of ticks preserved in ethanol were obtained using the Qiagen DNA DNeasy blood and tissue kit (Qiagen, Hilden, Germany), following the manufacturer's recommendations. Ticks of each pool that were preserved frozen were homogenized in 600 μL of culture medium with antibiotics [Dulbecco's Modified Eagle Medium (DMEM) with 100 units/mL penicillin and 100 μg/mL streptomycin, Sigma]. Four hundred μL of the homogenate were used for nucleic acid extraction (DNA and RNA) and the remaining 200 μL were preserved at −80°C for future analysis. The DNA and RNA were extracted using the DNeasy blood and tissue kit and RNeasy Mini kit (Qiagen, Hilden, Germany), respectively, following the manufacturer's recommendations.
The quality of nucleic acid extraction was checked using the 16S rRNA PCR assay (25). Positive samples were subjected to microbial screening using specific PCR assays for the analysis of (i) bacteria: Anaplasmataceae family, Bartonella spp., Borrelia spp., Coxiella spp., Rickettsiella spp., and spotted fever group (SFG) Rickettsia spp., (ii) viruses: Flaviviridae, Orthonairoviridae and Phenuiviridae families, and (iii) protozoans: Babesia and Theileria spp. Negative and positive controls (DNA or cDNA extracts of Anaplasma phagocytophilum, Bartonella henselae, Borrelia spielmanii, Borrelia miyamotoi, Coxiella-like of Rhipicephalus bursa, Rickettsia amblyommatis, Crimean-Congo haemorrhagic fever virus, Japanese encephalitis virus, Uukuniemi uukuvirus virus, and Babesia sp. from Rhipicephalus microplus) were included in all the PCR assays performed. Primers and PCR conditions are described in the Supplementary Table 1. The SFG Rickettsia was screened in tick legs for all ticks. In addition, pools formed by specimens that gave negative results were also tested for Rickettsia spp. Moreover, all the pools were screened for the presence of the remaining bacteria and protozoans. Lastly, the viral screening was performed on tick pools comprising specimens of fresh/frozen ticks (Table 1).
Prevalence of Infection
The prevalence of infection (PI) was estimated by:
PI = (No. of positive ticks/total No. of ticks analyzed) × 100%.
When microorganisms were amplified from pools of more than one tick, prevalence was calculated assuming that each positive pool contained only one positive tick. This estimate, known as minimum infectious rate (MIR), is calculated as follow:
MIR = (No. of positive pools/total No. of individual ticks analyzed) × 100%.
Analysis of Nucleotide Sequences
Nucleotide sequences were analyzed using the BLAST search (https://blast.ncbi.nlm.nih.gov/Blast.cgi), and the resulting sequences were submitted to GeneBank. The Clustal Omega online software (https://www.ebi.ac.uk/Tools/msa/clustalo/) was used for multiple sequence alignment. Phylogenetic analyses were conducted with MEGA X (http://www.megasoftware.net) using the maximum likelihood method including all sites. The nucleotide substitution model was selected according to the Akaike information criterion implemented in MEGAX. Confidence values for individual branches of the resulting trees were determined by bootstrap analysis with 500 replicates.
Results
Tick Identification
One hundred and sixty-three ticks, mainly nymphs, were collected from bird nests in Almería (n = 159), Guadalajara (n = 2), and Segovia (n = 2). Arthropods were obtained from nest material in cavities occupied by little owl (n = 129) and roller (n = 34) or, in few cases, from nestlings of these species (Table 1). All the specimens were morphologically identified as Argas spp. and 150 specimens were further studied molecularly. Examination of morphological characters enabled the identification of eight nymphs as A. reflexus, but the morphological identification of the remaining ticks could not be accurately performed with available keys. The A. reflexus identification was corroborated molecularly based on 16S rRNA fragment gene (Table 2). The molecular identification was not conclusive for the remaining 142 tick samples, which were grouped based on the 16S rRNA results into five different genotypes, designated as Argas spp. EEZA-CRETAV1–5 (Tables 1, 2). Based on 16S rRNA gene analyses, Argas sp. EEZA-CRETAV1 (n = 83) and Argas sp. EEZA-CRETAV2 (n = 33) were closest to A. vulgaris and A. polonicus, respectively (Table 2). In turn, the Argas sp. EEZA-CRETAV3 genotype (n = 22) shared the highest identity (<92.2%) with Argas africolumbae. Lastly, Argas sp. EEZA-CRETAV4 (n = 2) and Argas sp. EEZA-CRETAV5 genotypes (n = 2) shared highest identities with A. persicus (Table 2). The phylogeny inferred from 16S rRNA analysis corroborates the BLAST results (Figure 1). Phylogenetic analyses based on 12S rRNA and COI fragment genes could not be performed because of the lack of homologue sequences for the majority of Argas spp.
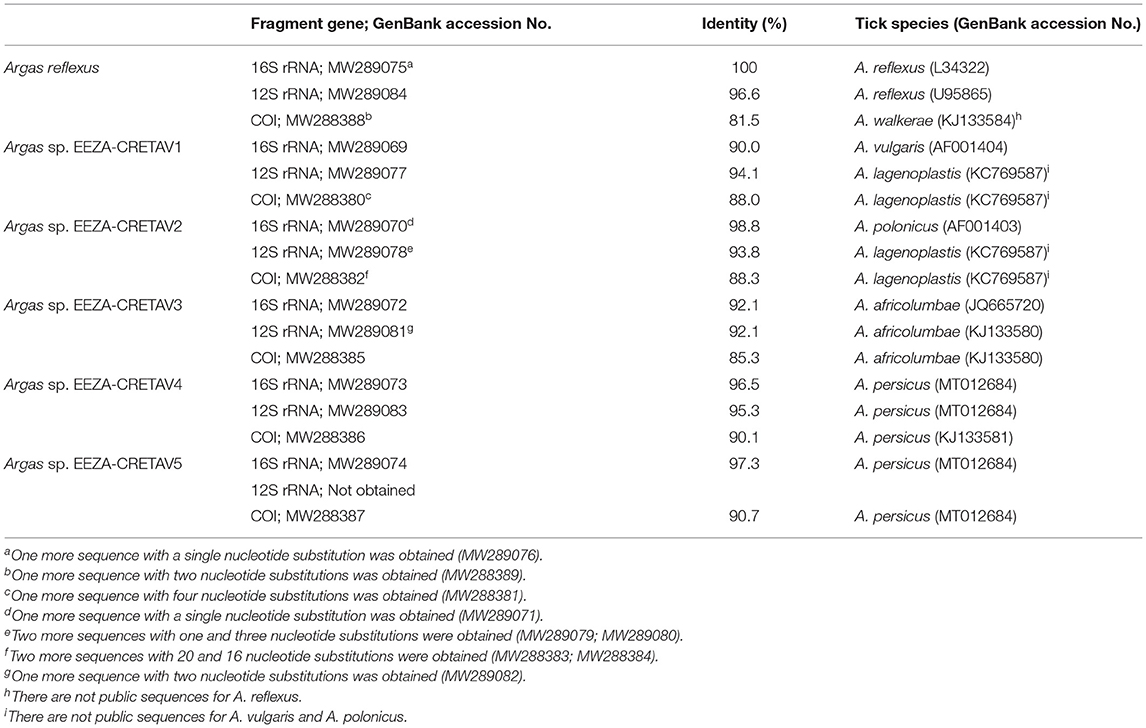
Table 2. Highest similarities of the Argas genotypes detected in this study reached with public sequences from GenBank.
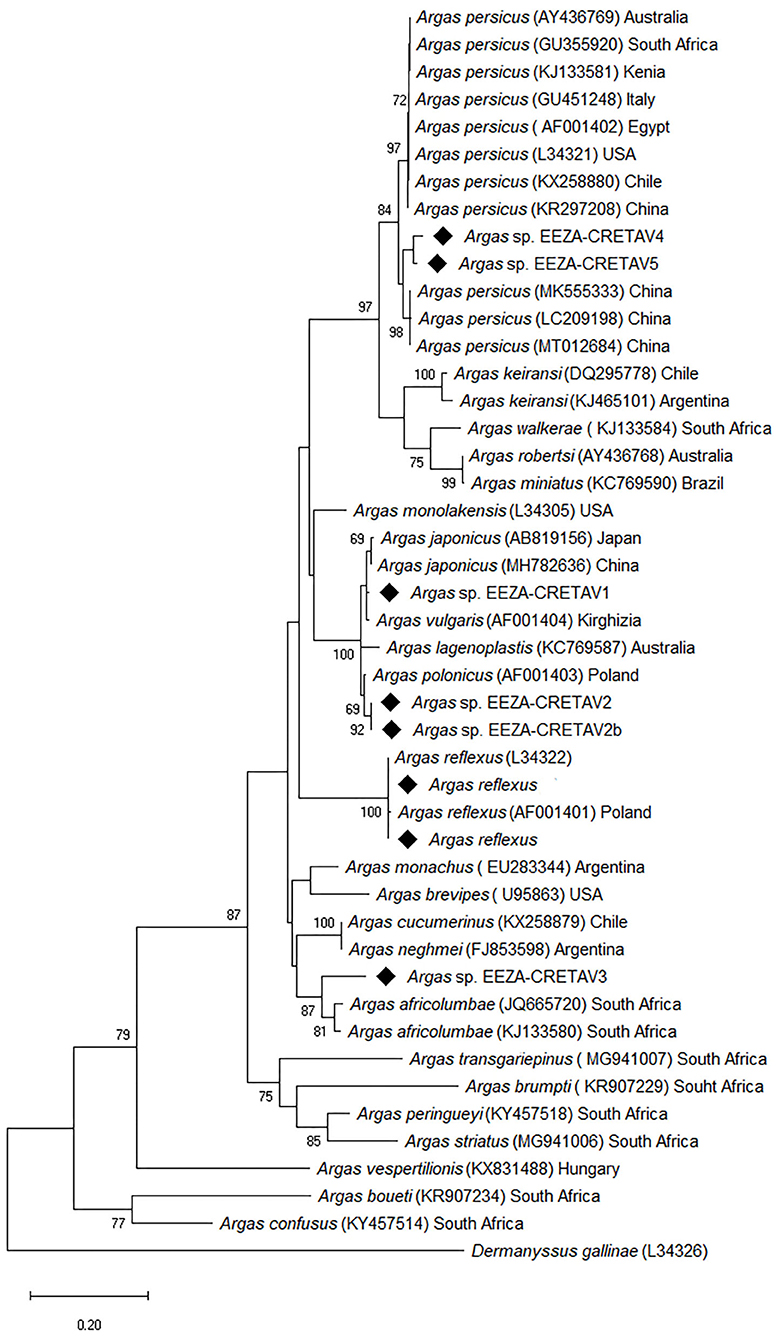
Figure 1. Phylogenetic tree based on 16S rRNA analysis showing the relationships between tick species and genotypes identified in this study and published validated Argas species. The evolutionary analysis was inferred using the maximum likelihood method and general time reversible + G model with Mega X. The analysis involved 46 nucleotide sequences and a total of 441 positions in the final dataset. The tree is drawn to scale, with branch lengths measured in the number of substitutions per site. Numbers (>65%) shown at the nodes correspond to bootstrapped percentages (for 500 repetitions). The GenBank accession number of sequences used in the analysis is shown in brackets after Argas taxon name and before sample origin. Sequences obtained in this study are marked with diamond. Dermanysus gallinae is used as outgroup.
Argas spp. EEZA-CRETAV1–3 specimens were collected in Almería, whereas those of Argas spp. EEZA-CRETAV4–5 were obtained in Segovia and Guadalajara. Also, some of the ticks of Argas spp. EEZA-CRETAV1–2 genotypes were collected from the same nests (Table 1).
Bacterial Screening
All the DNA extracts (individual samples and pools) gave positive results for the tick-16S rRNA PCR assay and, consequently, were screened for bacteria (Table 3). Amplicons for ompA gene were obtained from 112 DNA extracts from tick legs (PI = 74.7%). Specifically, Rickettsia was amplified from 81 and 31 samples belonging to Argas sp. EEZA-CRETAV1 (PI = 97.6%) and Argas sp. EEZA-CRETAV2 (PI = 93.9%) samples, respectively. DNA extracts corresponding to individual tick-leg samples from A. reflexus and Argas sp. EEZA-CRETAV3–5 specimens were negative, but body-halve extracts of these specimens were further analyzed using pooled samples. The nucleotide extracts from these pools were negative for all A. reflexus specimens, but yielded positive results for two out of six pools of Argas sp. EEZA-CRETAV3 specimens (MIR = 9.1%) (comprising a nymph collected from a roller nest in Almería in 2015 and larvae that were attached to a roller nestling in Almería in 2020), and all the pools of Argas sp. EEZA-CRETAV4–5 specimens (MIR = 100% for the two pools) (Table 3).
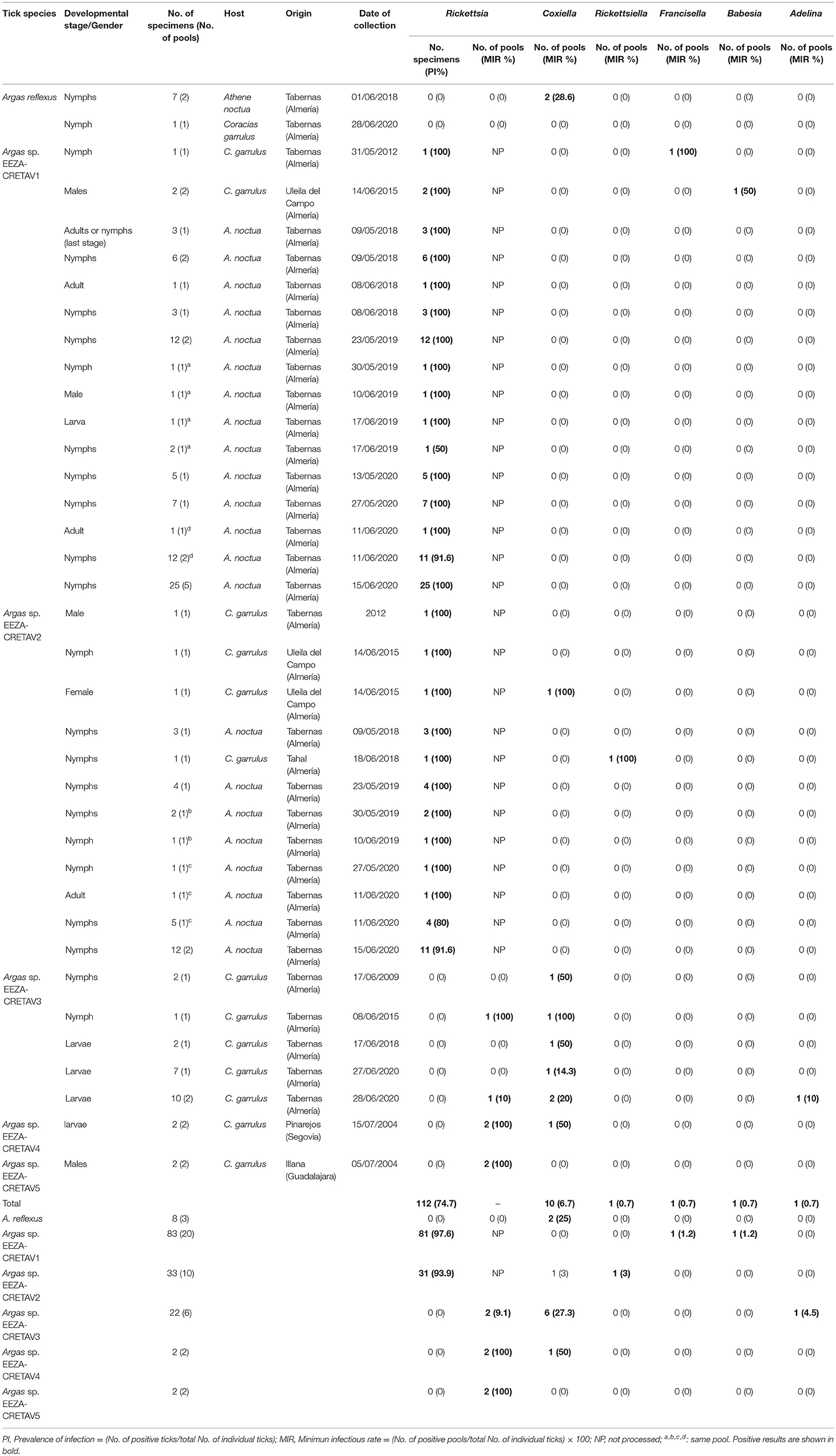
Table 3. Microorganisms detected in this study.
All the ompA gene sequences obtained were identical and showed the highest identity with Rickettsia fournieri (Table 4). Selected Rickettsia-positive samples were further genetically characterized by the amplification of five more rickettsial fragment genes (26). Nucleotide sequences for the respective genes were identical and showed highest identities with R. fournieri and Candidatus Rickettsia vini (Table 4). The phylogenetic tree based on the concatenated fragment genes of the Rickettsia strain detected, designated as Rickettsia sp. EEZA-CRETAV, corroborated the close relation with both R. fournieri and Ca. R. vini (Figure 2).

Table 4. Identities between fragment genes of Rickettsia sp. EEZA-CRETAV detected in the present study and published sequences from Rickettsia fournieri and Candidatus Rickettsia vini.

Figure 2. Phylogenetic tree showing the relationships between Rickettsia sp. EEZA-CRETAV and published Rickettsia spp. taxa. The evolutionary analysis was inferred using the maximum likelihood method and general time reversible + G model with Mega X, by concatenating fragments of six genes (sca4, 16s rRNA, ompB, ompA, 17-kDa, and gltA). The analysis involved 34 nucleotide sequences and a total of 4,120 positions in the final dataset. The tree is drawn to scale, with branch lengths measured in the number of substitutions per site. Numbers (>65%) shown at the nodes correspond to bootstrapped percentages (for 500 repetitions). The GenBank accession number of the sequences used in the analysis is shown in brackets after Rickettsia taxon name and the corresponding strain. Sequences obtained in this study are marked with diamond.
A total of 27 nucleotide sequences were obtained using the rpoB PCR assay selected for the Coxiella/Rickettsiella detection but highest identities with validated bacterial species reached <85% for 14 samples. The genetic analysis of the amplicon obtained from an Argas sp. EEZA-CRETAV1 nymph, collected in Almería in 2012, showed the highest identity with Francisella persicus (PI = 0.7% and PI = 1.2% for Argas sp. EEZA-CRETAV1) (Tables 3, 5). The novel Francisella strain molecularly described in this study was designated as Francisella sp. EEZA-CRETAV. Coxiella-like strains were successfully amplified from 10 pools (MIR = 6.7%). Two out of three A. reflexus pools (with ticks collected in a little owl nest in Almería in 2018) showed infection with a Coxiella strain previously detected in this tick species (MIR = 25%) (Table 5). A new strain of Coxiella spp., designated as Coxiella sp. EEZA-CRETAV1, was amplified in all the pools of Argas sp. EEZA-CRETAV3 specimens (n = 6, MIR = 27.3%). One more Coxiella strain, Coxiella sp. EEZA-CRETAV2, was detected in two pools of different tick genotypes, an Argas sp. EEZA-CRETAV2 (MIR = 3%) female collected in Almería in 2015 and an Argas sp. EEZA-CRETAV4 (MIR = 50%) larva collected in Segovia in 2004 (Table 3). Nucleotide sequences corresponding to rpoB an groEL genes of these two novel strains shared 95.3 and 97% identity, respectively, and reached highest identities with Coxiella strains detected in Ornithodoros ticks (Table 5). Moreover, pools integrated over a single specimen were also submitted for groEL analysis and a nymph belonging to Argas sp. EEZA-CRETAV2, collected in Almería in 2018, showed infection with Rickettsiella sp. (PI = 0.7% and PI for Argas sp. EEZA-CRETAV2 = 3%; Table 3). The corresponding amplicon, designated as Rickettsiella EEZA-CRETAV, showed highest identities with Rickettsiella species amplified from Ornithodoros normandi (Table 5).
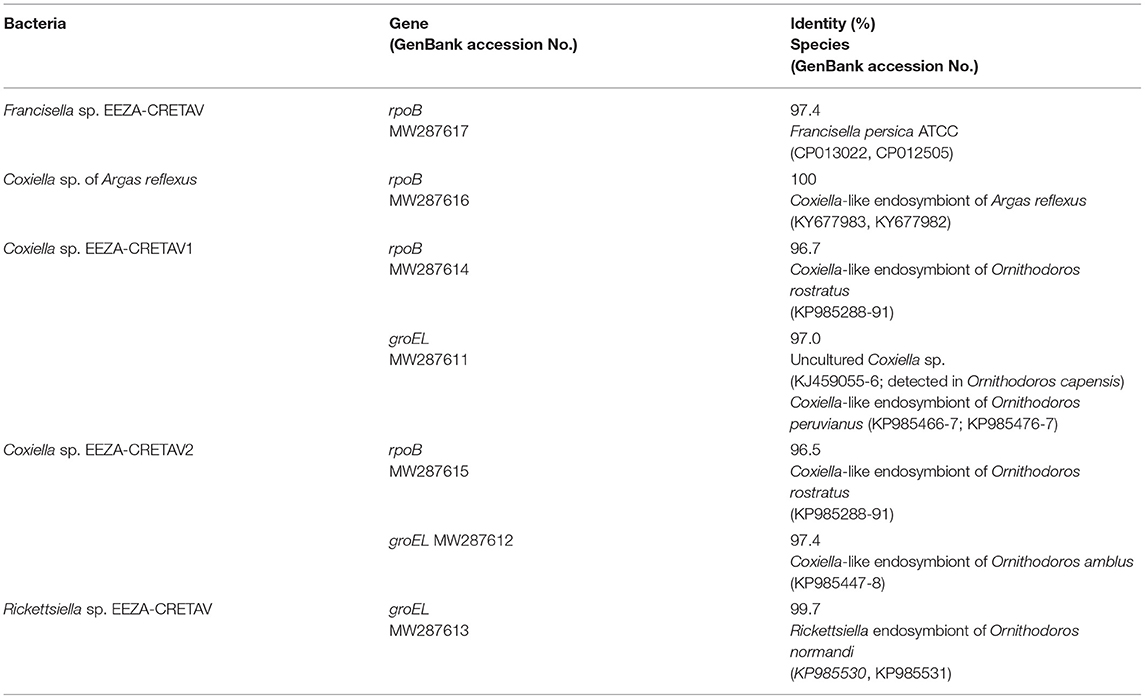
Table 5. Highest identities reached between fragment genes of Francisella, Coxiella, and Rickettsiella spp. detected in the present study and published sequences.
All the tick pools were examined by PCR assays for the presence of Anaplasmataceae, Bartonella, and Borrelia species and all gave negative results.
Viral Screening
Twenty-one pools originated from fresh/frozen ticks [n = 111 ticks: 1 tick (1 pool) of A. reflexus, 67 (12) of Argas sp. EEZA-CRETAV1, 26 (5) of Argas sp. EEZA-CRETAV2 and 17 (3) of Argas sp. EEZA-CRETAV3 specimens; Table 1] were analyzed for the presence of viruses belonging to families Flaviviridae, Orthonairoviridae, and Phenuiviridae. No sequences were amplified using the selected PCR assays.
Protozoan Screening
Tick DNA from 43 pools was analyzed using a PCR assay that amplifies 18S rRNA gene of Babesia and Theileria spp. (Table 3, Supplementary Table 1). Babesia sp. was detected from a male Argas sp. EEZA-CRETAV1 collected in Almería in 2015 (MIR = 0.7%, MIR = 1.2% for Argas sp. EEZA-CRETAV1) (Table 3). Three more ticks (one Argas sp. EEZA-CRETAV1 and two Argas sp. EEZA-CRETAV2 ticks) collected simultaneously from the same nest gave negative results. Based on the analysis of 18S rRNA gene, the closets Babesia sp. from the detected strain, designated as Babesia sp. EEZA-CRETAV, was Babesia ardeae (KY436057; 95.8% identity) (Figure 3). Two more genes, ITS1 and ITS2, were also examined, but currently there are no B. ardeae sequences available for these markers. The analysis of these genes showed highest (<82%) identities with Babesia poelea (accession no. DQ200887). A pool formed by 4 Argas sp. EEZA-CRETAV3 larvae, collected in Almería in 2020, showed presence of a coccidian parasite Adelina bambarooniae (AF494059) (MIR = 0.7%) (Table 3).

Figure 3. Maximum likelihood trees of Babesia species based on 18S rRNA analysis. The evolutionary analysis was inferred using Tamura-Nei model + G model with Mega X. The analysis involved 39 nucleotide sequences and a total of 483 positions in the final dataset. The tree is drawn to scale, with branch lengths measured in the number of substitutions per site. Numbers (>65%) shown at the nodes correspond to bootstrapped percentages (for 500 repetitions). The GenBank accession number of the sequences used in this analysis is shown in brackets after Babesia taxon name. The species found in this study is marked with diamond and the species detected in birds with asterisk. Taxon names are followed by GenBank accession numbers and collection location where available. Plasmodium falciparium is used as outgroup.
Sequences Submission to a Public Database
The nucleotide sequences of ticks and microorganisms detected in this study (n = 42) were deposited in the GenBank database under accession numbers showed in Table 6.

Table 6. GenBank accession numbers of the nucleotide sequences obtained in this study, differing from published sequences.
Discussion
Soft ticks are important vectors of microbial agents of animal and human diseases. Despite this, the knowledge of argasid tick species and their associated microorganisms is generally scarce. Herein, the occurrence of five novel Argas spp. genotypes, in addition to A. reflexus, collected in the Iberian Peninsula (Spain) in nests occupied by little owls and European rollers is reported. Moreover, we detected the presence of tick-borne microorganisms belonging to genera Rickettsia, Coxiella, Francisella, Rickettsiella, and Babesia in Argas ticks. In contrast, Anaplasmataceae, Bartonella, and Borrelia bacterium species and viruses belonging to the families Flaviviridae, Orthonairoviridae, and Phenuiviridae have not been detected.
Tick Identification
Of the six Argas genotypes detected in this study, only one could be identified to the species level, namely, A. reflexus. The morphological identification of ticks is challenging, even for experts (27), while the molecular approach appears to be an accurate tool for tick identification (25, 27). Nevertheless, increased effort for molecular characterization of more Argas species is needed for a reliable taxonomic inference based on molecular tools. In order to confirm if the genotypes identified in this study represent validated or potentially novel tick species, a further morphological analysis including unfed larva specimens, should be performed along with rigorous molecular analyses of Argas ticks from diverse geographical locations.
Argas reflexus specimens, known as the pigeon tick, have been collected in two bird nests in Almería (Southern Spain), one occupied by little owl (n = 7) and one by roller (n = 1). This species occurs in Spain, and it is a well-known ectoparasite of little owls, whereas information regarding roller infestations is scarce (6, 20, 28). Argas sp. EEZA-CRETAV4 and Argas sp. EEZA-CRETAV5, genotypes amplified in Central Spain, clustered molecularly with A. persicus from China (29), but in a different branch than other A. persicus specimens (Figure 1). Nevertheless, the broad genetic divergence of this group (29, 30), also revealed in the phylogeny inferred herein (Figure 1), suggests that cryptic species could occur in this taxon known as fowl tick. This worldwide distributed tick has been previously reported in Spain and is known to infest wild birds also in other countries (6, 28). In addition, ticks of three more Argas genotypes have been identified, all of them in Almería. The Argas sp. EEZA-CRETAV1–2 genotypes clustered together and appear to be closely related with A. vulgaris and A. polonicus, respectively (Figure 1). Neither of these two tick species have been previously reported from western Europe and their occurrence is only documented in a few eastern European countries (20, 21, 31). These genotypes have been found in the nest material of roller and little owls, occurring in the same nests in several cases (Table 1). In the study area, there is a high competition among cavity-nesting birds for suitable cavities and the same cavity can be successively used by little owls, pigeons and rollers (32). This fact could explain the infestation of both bird species by the same nidicolous tick taxa, i.e., A. reflexus and Argas sp. EEZA-CRETAV1–2. In contrast, Argas sp. EEZA-CRETAV3, closely related to A. africolumbae, has been found only in nests occupied by rollers. The roller is a long-distant migrant species (trans-Saharan migrant) and the Spanish populations overwinter in different African regions (33). It is worth mentioning that A. africolumbae is an ornithophilic tick that occurs in Africa: South Africa, Kenya, Tanzania and Burkina Faso (34, 35). It is well-known that birds can serve as dispersers of ticks and tick-borne microorganisms, even though this information pertains mainly to hard ticks (36, 37). Some studies suggest that the role of birds as dispersers of soft ticks is less important, due to the biology of these ticks (nidicolous behavior and shorter blood-feeding time), but the role of migratory birds as reservoirs or amplifiers of tick-borne microorganisms associated with soft ticks remains to be better investigated (38, 39).
Tick-Borne Microorganisms
The microbiological screening of ticks is important to identify the local risks of emergence of tick-borne diseases. Argas species, including A. reflexus and A. persicus, have been described as important pests and vectors of diseases in poultry and wild birds, being responsible for high economic losses (7, 11, 40). These tick species have also been recorded biting humans and causing anaphylaxis episodes (8, 10). Although humans are accidental hosts of Argas ticks and the ticks carry numerous microorganisms, human pathogens among them, the role of these ticks as vectors of human infectious agents has not been proven.
The most prevalent microorganisms amplified in this study belong to the Rickettsia genus (α-Proteobacteria; Rickettsiaceae). The high prevalence of Rickettsia spp. found in ticks of Argas sp. EEZA-CRETAV1–2 specimens suggests that the bacterium is a common endosymbiont of ticks of the two genotypes. The detection of the newly described Rickettsia strain in tick-body samples, but not in tick-leg samples of ticks of other genotypes (Argas sp. EEZA-CRETAV3–5), suggests that this Rickettsia species may not be a true intracellular endosymbiont and its presence in the former genotypes could be acquired through feeding on infected hosts or by cofeeding. The phylogenetic analysis of Rickettsia sp. EEZA-CRETAV reveals its close relation with R. fournieri and Ca. R. vini, both Rickettsia species associated with ornithophilic nidicolous ticks (41, 42). While R. fournieri has been described only once from A. lagenoplastis in Australia (41), Ca. R. vini has been detected in several European countries in Ixodes spp. (36, 42–45). The single study performed suggests that Ca. R. vini is not pathogenic (43). Nevertheless, the two Rickettsia taxa are closely related to other Rickettsia species that are recognized as human pathogens, specifically Rickettsia japonica and Rickettsia heilongjiangensis (46, 47). Thus, the epidemiology and pathological potential of Rickettsia strains such as Rickettsia sp. EEZA-CRETAV, in addition to R. fournieri and Ca. R. vini, should be further investigated. Likewise, the isolation of Rickettsia sp. EEZA-CRETAV is necessary to gain an insight into the epidemiological importance of this strain.
In addition to the Rickettsia taxon, this study has revealed for the first time different proteobacterial tick endosymbionts in Argas spp. from Spain, namely, Coxiella, Rickettsiella (Gamma-proteobacterium; Coxiellaceae) and Francisella (Gamma-proteobacterium; Francisellaceae) species. The detected species, commonly known as Rickettsiella-like, Coxiella-like, and Francisella-like, are intracellular obligatory endosymbionts important for tick survival. They play some role in B vitamins biosynthesis and their presence may interfere with the transmission (positively or negatively) of other microorganisms, including tick-borne pathogens (48). They are related to species responsible for important human diseases. For instance, Coxiella burnetii and Francisella tularensis cause Q fever and tularemia, respectively (49, 50). Coxiella-like species have been implicated in human and avian diseases (51, 52). Three different Coxiella strains have been successfully detected in this study. Of them, the Coxiella sp. from A. reflexus was homologous to the species previously detected in the same tick species, but the remaining two strains, designated as Coxiella sp. EEZA-CRETAV1–2, differ from the scarcely-known Coxiella-like strains of Argas species (14, 53–55). To date, Rickettsiella has been mainly reported in hard ticks (Ixodes spp. and Haemaphysalis spp.) and soft ticks belonging to the genus Ornithodoros (14). The presence of Rickettsiella in Argas ticks has been suggested for the bat tick A. transgariepinus from Namibia, but the available 16S rRNA sequences shared low identities with the published Rickettsiella sequences (17). In the present study, a Rickettsiella sp. similar to that of O. normandi from Tunisia (55) was detected in an Argas sp. EEZA-CRETAV2 specimen. Lastly, a strain closely related to F. persicus was found in an Argas sp. EEZA-CRETAV1 specimen. Francisella persicus, formerly Wolbachia persica, is an endosymbiont of Argas arboreus (previously referred to as A. persicus) (56). This bacterium has not been identified as an animal or human pathogen, but the analysis of its genome shows that this species conserves an important number of potentially functional virulence-associated genes, suggesting that it could be pathogenic to mammals (57).
Bacteria of Anaplasmataceae family, Bartonella, and Borrelia spp. have not been detected in the ticks analyzed. Argas spp. are recognized vectors of B. anserine, the agent of the avian spirochetosis, a worldwide distributed disease of veterinary importance that has not been reported from Spain (11, 40). The lack of virus detection in this study was unexpected, because diverse viruses are readily detected in Argas ticks (7). Scarce tick-borne viruses have been described from Spain and all but Meaban-like virus, a flavivirus found in Ornithodoros maritimus (38), are associated to ixodid ticks. Some of these viruses have a great relevance for human health, e.g., Crimean-Congo hemorrhagic fever virus, whose arrival in infected ticks has been suggested to take place through migratory birds (37, 58). This fact highlights the importance of studying viruses in soft ticks associated to birds in Spain, an important area in the migratory routes of many avian species, because what it is not sought, it is not found (59).
Two apicomplexan parasites have been found in this work, Babesia sp. and A. bambarooniae, though the latter species is not known as a tick-borne microorganism. In turn, the apicomplexan piroplasms Babesia (Aconoidasida; Babesiidae) are mainly vectored by ixodid ticks, though argasid ticks also were suggested as potential vectors (60). Babesia sp. EEZA-CRETAV has been amplified from an Argas sp. EEZA-CRETAV3 tick associated with rollers and the presence of the protozoan in the blood of rollers cannot be rejected. Sixteen Babesia species responsible for avian piroplasmosis, in addition to several strains that are not fully identified, are known (61). Of these, Babesia frugilegica, Babesia shortti and Babesia benneti have been reported from Spain (62–64). Babesia sp. EEZA-CRETAV is closely related to some of the scarcely genetically characterized Babesia species, mainly to B. ardeae (Figure 3). This species has been detected in Asia and its pathogenicity is unknown (61). This strain is also close to the human-pathogenic B. ducani that has been identified in North America, the United Kingdom and Australia (65). To our knowledge, Babesia spp. have not been identified in owls.
Conclusions
The sedentary lifestyle of soft ticks could imply a limited role of these ticks in the circulation of infectious agents (66). However, as indicated by this study, the high re-use of cavities within and between years by different bird species could importantly enhance the spread of microorganisms associated with soft nidicolous ticks, such as Argas ticks.
This study highlights the richness of nidicolous Argas ticks associated with cavity-nesting birds in a semi-desert area in Western Europe, and suggests that the diversity of this genus in Spain might be underestimated. Moreover, this work provides the first report of Rickettsia sp., Coxiella spp., Rickettsiella sp., Francisella sp. and Babesia sp., from soft ticks in Spain, and A. bambarooniae from Ixodida.
Further research should be carried out to confirm if the new genotypes of ticks and their microorganisms represent novel taxa and, additionally, to understand their role in the epidemiology of zoonoses using the One Health approach.
Data Availability Statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
Author Contributions
APa, RV, JO, and FV designed the initial study. JV and FV carried out the field work. APa performed the tick identification and tick processing. APa, APo, SS, and PS implemented the analysis of microorganisms. APa and FV wrote the first draft of the manuscript. All authors contributed to data interpretation and revisions.
Funding
RV was funded by the Research Grant Agency (VEGA) of the Ministry of Education, Science, Research and Sport of the Slovak Republic and Slovak Academy of Sciences (2/0023/20). FV received financial support from the projects CGL2014-55969 and PGC2018-097426-B-C22 (Spanish Ministry of Universities, Spanish State Research Agency, FEDER Program, European Union). JV was funded by a predoctoral grant (BES-2015-075951) of the Spanish Ministry of Science and Innovation.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We would like to thank Agustín Estrada-Peña (University of Zaragoza) for helping with the taxonomy of Argas spp. We thank L. Bolonio and I. E. Cardiel (CEBIME, Conservación y Estudio de la Biodiversidad en Medios Esteparios) for providing tick samples. Teresa Martínez, Maite Amat, Miguel Ángel Calero, and Stanislav Kolencik helped with fieldwork. Junta de Andalucía kindly provided permits to sample birds and their nests. We thank Dr. Didier Raoult (Unité de Recherche sur les Maladies Infectieuses et Tropicales Emergentes, France), Dr. J. Stephen Dumler (The Johns Hopkins Hospital, USA), Dr. Houssam Attoui [UMR Virologie, National Institute for Agriculture, Food, and Environment (INRAE), France], Dr. Luis M. Hernández-Triana (Wildlife Zoonoses and Vector-borne Diseases Research Group, Animal and Plant Health Agency, United Kingdom), Dr. Joaquim Ruiz (Universidad Cientifica del Sur, Peru), Dr. Volker Fingerle (German National Reference Centre for Borrelia), and Dr. Matthias Niedrig (Centre for Biosafety, Robert Koch Institute, Germany) for providing positive controls.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fvets.2021.637837/full#supplementary-material
References
1. Guglielmone AA, Robbins RG, Apanaskevich DA, Petney TN, Estrada-Peña A, Horak IG, et al. The argasidae, ixodidae and nuttalliellidae (Acari: Ixodida) of the world: a list of valid species names. Zootaxa. (2010) 2528:1–28. doi: 10.11646/zootaxa.2528.1.1
2. Dusbábek F. Argas (Argas) vulgaris Filippova, 1961, a new member of Czechoslovak tick fauna. Folia Parasitol. (1976) 23:281–3.
3. Dikaev BI. [Argasid tick fauna (Argasidae) of the Chechen-Ingush ASSR]. Parazitologiia. (1981) 15:76–8.
4. Estrada-Peña A, Lucientes J, Sánchez C. Argas (Persicargas) gilcolla n. sp. (Acarina: Argasidae): a parasite of the griffon vulture, Gyps fulvus, in Spain. J Parasitol. (1987) 73:824–8.
5. Estrada-Peña A, Pfäffle M, Baneth G, Kleinerman G, Petney TN. Ixodoidea of the western palaearctic: a review of available literature for identification of species. Ticks Tick Borne Dis. (2017) 8:512–25. doi: 10.1016/j.ttbdis.2017.02.013
6. Cordero del Campillo M, Castañón Ordóñez L, Reguera Feo A. [Índice-Catálogo de Zooparásitos Ibéricos]. Spain: Ediciones Universidad de León (1994).
7. Hoogstraal H. Argasid and nuttalliellid ticks as parasites and vectors. Adv Parasitol. (1985) 24:135–238. doi: 10.1016/S0065-308X(08)60563-1
8. Estrada-Peña A, Jongejan F. Ticks feeding on humans: a review of records on human-biting Ixodoidea with special reference to pathogen transmission. Exp Appl Acarol. (1999) 23:685–715. doi: 10.1023/a:1006241108739
9. Rosenstein M. Paralysis in chickens caused by larvae of the poultry tick, Argas persicus. Avian Dis. (1976) 20:407–9. doi: 10.2307/1589281
10. Boni E, Incorvaia C. Near-fatal anaphylaxis with Kounis syndrome caused by Argas reflexus bite: a case report. Clin Mol Allergy. (2020) 18:4. doi: 10.1186/s12948-020-00121-w
11. Gothe R, Buchheim C, Schrecke W. Argas (Persicargas) persicus and Argas (Argas) africolumbae as natural vectors of Borrelia anserina and Aegyptianella pullorum in upper Volta. Berliner Munchener Tierarztliche Wochenschrift. (1981) 94:280–5.
12. Vargina SG, Kuchuk LA, Gershtein VI, Karas FR. [Transmission of Issyk Kul virus by Argas vespertilionis ticks in experiment]. Sborn nauch Tr Inst Virus Im Ivanov Akad Med Nauk SSSR (1982). p. 123–7. doi: 10.1128/genomeA.00662-15
13. Karadjian G, Chavatte J, Landau I. Systematic revision of the adeleid haemogregarines, with creation of Bartazoon n. g., reassignment of Hepatozoon argantis Garnham, 1954 to Hemolivia, and molecular data on Hemolivia stellata. Parasite. (2015) 22:31. doi: 10.1051/parasite/2015031
14. Duron O, Binetruy F, Noël V, Cremaschi J, McCoy KD, Arnathau A, et al. Evolutionary changes in symbiont community structure in ticks. Mol Ecol. (2017) 26:2905–21. 10.1111/mec.14094. doi: 10.1111/mec.14094
15. Lafri I, El Hamzaoui B, Bitam I, Leulmi H, Lalout R, Mediannikov O, et al. Detection of relapsing fever Borrelia spp., Bartonella spp. and Anaplasmataceae bacteria in argasid ticks in Algeria. PLoS Negl Trop Dis. (2017) 11:e0006064. doi: 10.1371/journal.pntd.0006064
16. Lv J, Fernández de Marco MM, Goharriz H, Phipps LP, McElhinney LM, Hernández-Triana LM, et al. Detection of tick-borne bacteria and babesia with zoonotic potential in Argas (Carios) vespertilionis (Latreille, 1802) ticks from British bats. Sci Rep. (2018) 8:1865. doi: 10.1038/s41598-018-20138-1
17. Reeves WK, Mans BJ, Durden LA, Miller MM, Gratton EM, Laverty TM. Rickettsia hoogstraalii and a Rickettsiella from the Bat Tick Argas transgariepinus, in Namibia. J Parasitol. (2020) 106:663–9. doi: 10.1645/20-46
18. Lázaro R, Rodrigo FS, Gutiérrez L, Domingo F, Puigdefábregas J. Analysis of a thirty-year rainfall record (1967–1997) from semi-arid SE Spain: a plant ecological perspective. J Arid Environ. (2001) 48:373–95. doi: 10.1006/jare.2000.0755
19. Cramp S. The Complete Birds of the Western Palearctic on CDROM. Oxford: Oxford University Press (1998).
20. Filippova NA. [Argasid Ticks (Argasidae). Fauna of USSR, Arachnoidea 4(3)]. Moskva: ‘Nauka’ Publishing House (1966).
21. Siuda K, Hoogstraal H, Clifford CM, Wassef HY. Observations on the subgenus Argas (Ixodoidea: argasidae: Argas). 17. Argas (A.) polonicus sp. N. parasitizing domestic pigeons in Krakow, Poland. J Parasitol. (1979) 65:170–81.
23. Estrada-Peña A, Mihalca AD, Petney T. Ticks of Europe North Africa. A Guide to Species Identification. Switzerland: Springer International Publishing AG (2017). doi: 10.1007/978-3-319-63760-0
24. Portillo A, Santos AS, Santibáñez S, Pérez-Martínez L, Blanco JR, Ibarra V, et al. Detection of a non-pathogenic variant of Anaplasma phagocytophilum in Ixodes ricinus from La Rioja, Spain. Ann NY Acad Sci. (2005) 1063:333–6. doi: 10.1196/annals.1355.053
25. Black WC, Piesman J. Phylogeny of hard and soft tick taxa (Acari:Ixodida) based on mitochondrial 16S rDNA sequences. Proc Natl Acad Sci USA. (1994) 91:10034–8. doi: 10.1073/pnas.91.21.10034
26. Palomar AM, Portillo A, Santibáñez P, Santibáñez S, García-Álvarez L, Oteo JA. Genetic characterization of Candidatus Rickettsia vini, a new rickettsia amplified in ticks from La Rioja, Spain. Ticks Tick Borne Dis. (2012) 3:319–21. doi: 10.1016/j.ttbdis.2012.10.025
27. Estrada-Peña A, D'Amico G, Palomar AM, Dupraz M, Fonville M, Heylen D, et al. A comparative test of ixodid tick identification by a network of European researchers. Ticks Tick Borne Dis. (2017) 8:540–6. doi: 10.1016/j.ttbdis.2017.03.001
28. Doss MA, Farr MM, Roach KF, Anastos G. Special Publication No. 3, Ticks Tickborne Diseases, II. Hosts, Part 1. A-F. United States Government Printing Office. (1974). Available in: http://hdl.handle.net/1969.1/92068 (accessed January 24, 2021).
29. Feng J, Wu M, Wulan, Huang T, Zhang J, Renbatu N, et al. Identification of two genotypes of argas persicus and associated rickettsia-specific genes from different regions of inner mongolia. J Parasitol. (2019) 105:92–101. doi: 10.1645/18-27
30. Muñoz-Leal S, Venzal JM, Nava S, Reyes M, Martins TF, Leite RC, et al. The geographic distribution of Argas (Persicargas) miniatus and Argas (Persicargas) persicus (Acari: Argasidae) in America, with morphological and molecular diagnoses from Brazil, Chile and Cuba. Ticks Tick Borne Dis. (2018) 9:44–56. doi: 10.1016/j.ttbdis.2017.10.009
31. Dusbábek F. Identity of Argas (Argas) polonicus populations in Czechoslovakia and Poland. Folia Parasitol. (1985) 32:163–71.
32. Valera F, Václav R, Calero-Torralbo MA, Martínez T, Veiga J. Natural cavity restoration as an alternative to nest box supplementation. Restor Ecol. (2019) 27:220–7. doi: 10.1111/rec.12841
33. Rodríguez-Ruiz J, de la Puente J, Parejo D, Valera F, Calero-Torralbo MA, Reyes-González JM, et al. Disentangling migratory routes and wintering grounds of Iberian near-threatened European Rollers Coracias garrulus. PLoS ONE. (2014) 9:e115615. doi: 10.1371/journal.pone.0115615
34. Hoogstraal H, Kaiser MN, Walker JB, Ledger JA, Converse JD, Rice RGA. “Observations on the subgenus Argas (Ixodoidea: Argasidae: Argas) 10. A. (A.) africolumbae, n. 541 sp., a Pretoria virus-infected parasite of birds in southern and eastern Africa”. J Med Entomol. (1975) 12:194–210. doi: 10.1093/jmedent/12.2
35. Kraiss A, Gothe R. The life cycle of Argas (Argas) africolumbae under constant abiotic and biotic conditions. Vet Parasitol. (1982) 11:365–73. doi: 10.1016/0304-4017(82)90104-2
36. Palomar AM, Santibáñez P, Mazuelas D, Roncero L, Santibáñez S, Portillo A, et al. Role of birds in dispersal of etiologic agents of tick-borne zoonoses, Spain, 2009. Emerg Infect Dis. (2012) 18:1188–91. doi: 10.3201/eid1807.111777
37. Palomar AM, Portillo A, Santibáñez P, Mazuelas D, Arizaga J, Crespo A, et al. Crimean-Congo hemorrhagic fever virus in ticks from migratory birds, Morocco. Emerg Infect Dis. (2013) 19:260–3. doi: 10.3201/eid1902.121193
38. Arnal A, Gómez-Díaz E, Cerdà-Cuélla M, Lecollinet S, Pearce-Duvet J, Busquets N, et al. Circulation of a Meaban-like virus in yellow-legged gulls and seabird ticks in the western Mediterranean basin. PLoS ONE. (2014) 9:e89601. doi: 10.1371/journal.pone.0089601
39. Kada S, McCoy KD, Boulinier T. Impact of life stage-dependent dispersal on the colonization dynamics of host patches by ticks and tick-borne infectious agents Parasit Vectors. (2017) 10:375. doi: 10.1186/s13071-017-2261-y
40. Lisbôa RS, Teixeira RC, Rangel CP, Santos HA, Massard CL, Fonseca AH. Avian spirochetosis in chickens following experimental transmission of Borrelia anserina by Argas (Persicargas) miniatus. Avian Diseases. (2009) 53:166–8. doi: 10.1637/8377-061508-Reg.1
41. Diop A, Barker SC, Eberhard M, Barker D, Nguyen TT, Di Pinto F, et al. Rickettsia fournieri sp. nov, a novel spotted fever group rickettsia from Argas lagenoplastis ticks in Australia. Int J Syst Evol Microbiol. (2018) 68:3781–4. doi: 10.1099/ijsem.0.003057
42. Palomar AM, Portillo A, Crespo A, Santibáñez S, Mazuelas D, Oteo JA. Prevalence of ‘Candidatus rickettsia vini’ in Ixodes arboricola ticks in the north of Spain, 2011–2013. Parasit Vectors. (2015) 8:110. doi: 10.1186/s13071-015-0724-6
43. Nováková M, Costa FB, Krause F, Literak I, Labruna MB. Rickettsia vini n. sp. (Rickettsiaceae) infecting the tick Ixodes arboricola (Acari: Ixodidae). Parasit Vectors. (2016) 9:469. doi: 10.1186/s13071-016-1742-8
44. Nováková M, Heneberg P, Heylen D, Medvecký M, Muñoz-Leal S, Šmajs D, et al. Isolated populations of Ixodes lividus ticks in the Czech Republic and Belgium host genetically homogeneous Rickettsia vini. Ticks Tick Borne Dis. (2018) 9:479–84. doi: 10.1016/j.ttbdis.2017.12.018
45. Al-Khafaji AM, Bell-Sakyi L, Fracasso G, Luu L, Heylen D, Matthysen E, et al. Isolation of Candidatus Rickettsia vini from Belgian Ixodes arboricola ticks and propagation in tick cell lines. Ticks Tick Borne Dis. (2020) 11:101511. doi: 10.1016/j.ttbdis.2020.101511
46. Ando S, Kurosawa M, Sakata A, Fujita H, Sakai K, Sekine M, et al. Human Rickettsia heilongjiangensis infection, Japan. Emerg Infect Dis. (2010) 16:1306–8. doi: 10.3201/eid1608.100049
47. Uchida T. Rickettsia japonica, the etiologic agent of Oriental spotted fever. Microbiol. Immunol. (1993) 37:91–102. doi: 10.1111/j.1348-0421.1993.tb03185.x
48. Bonnet SI, Binetruy F, Hernández-Jarguín AM, Duron O. The Tick Microbiome: why non-pathogenic Microorganisms Matter in Tick Biology and Pathogen Transmission. Front Cell Infect Microbiol. (2017) 7:236. doi: 10.3389/fcimb.2017.00236
49. Ellis J, Oyston PC, Green M, Titball RW. Tularemia. Clin Microbiol Rev. (2002) 15:631–46. doi: 10.1128/CMR.15.4.631-646.2002
50. Oteo JA, Pérez-Cortés S, Santibáñez P, Gutiérrez E, Portillo A, Blanco JR, et al. Q fever endocarditis associated with a cardiovascular implantable electronic device. Clin Microbiol Infect. (2012) 18:E482–4. doi: 10.1111/j.1469-0691.2012.03992.x
51. Shivaprasad HL, Cadenas MB, Diab SS, Nordhausen R, Bradway D, Crespo R, et al. Coxiella-like infection in psittacines and a toucan. Avian Dis. (2008) 52:426–32. doi: 10.1637/8192-120707-Reg
52. Angelakis E, Mediannikov O, Jos SL, Berenger JM, Parola P, Raoult D. Candidatus Coxiella massiliensis infection. Emerg Infect Dis. (2016) 22:285–8. doi: 10.3201/eid2202.150106
53. Hosseini-Chegeni A, Kayedi MH. Molecular detection of Coxiella (Gammaproteobacteria: Coxiellaceae) in Argas persicus and Alveonasus canestrinii (Acari: Argasidae) from Iran. Microb Pathog. (2020) 139:103902. doi: 10.1016/j.micpath.2019.103902
54. Reeves WK. Molecular evidence for a novel Coxiella from Argas monolakensis (Acari: Argasidae) from Mono Lake, California, USA. Exp Appl Acarol. (2008) 44:57–60. doi: 10.1007/s10493-008-9128-z
55. Duron O, Noël V, McCoy KD, Bonazzi M, Sidi-Boumedine K, Morel O, et al. The recent evolution of a maternally-inherited endosymbiont of ticks led to the emergence of the Q fever pathogen, Coxiella burnetii. PLoS Pathog. (2015) 11:e1004892. doi: 10.1371/journal.ppat.1004892
56. Larson MA, Nalbantoglu U, Sayood K, Zentz EB, Cer RZ, Iwen PC, et al. Reclassification of Wolbachia persica as Francisella persica comb. nov. and emended description of the family Francisellaceae. Int J Syst Evol Microbiol. (2016) 66:1200–5. doi: 10.1099/ijsem.0.000855
57. Gerhart JG, Dutcher HA, Brenner AE, Moses AS, Grubhoffer L, Raghavan R. Multiple Acquisitions of Pathogen-Derived Francisella Endosymbionts in Soft Ticks. Genome Biol Evol. (2018) 10:607–15. doi: 10.1093/gbe/evy021
58. Estrada-Peña A, Palomar AM, Santibáñez P, Sánchez N, Habela MA, Portillo A, et al. Crimean-Congo hemorrhagic fever virus in ticks, Southwestern Europe, 2010. Emerg Infect Dis. (2012) 18:179–80. doi: 10.3201/eid1801.111040
59. Oteo JA, Palomar AM. Crimean-Congo haemorrhagic fever: “What is not sought is not found”. Med Clin. (2018) 150:266–7. doi: 10.1016/j.medcle.2018.01.015
60. Peirce MA, Parson NJ. Babesia ugwidiensis, a new species of avian piroplasm from Phalacrocoracidae in South Africa. Parasite. (2012) 19:375–9. doi: 10.1051/parasite/2012194375
61. Chavatte J, Okumura C, Landau I. Redescription of Babesia ardeae Toumanoff, 1940, a parasite of Ardeidae, including molecular characterization. Parasitol Res. (2017) 116:1089–97. doi: 10.1007/s00436-017-5394-1
62. Blanco G, Merino S, Tella JL, Fargallo JA, Gajón A. Hematozoa in two populations of the threatened red-billed chough in Spain. J Wildl Dis. (1997) 33:642–5. doi: 10.7589/0090-3558-33.3.642
63. Merino S. Babesia bennetti n. sp. from yellow-legged gull (Larus cachinnans, Aves, Laridae) on Benidorm Island, Mediterranean Sea. J Parasitol. (1998) 84:422–4. doi: 10.2307/3284504
64. Muñoz E, Molina R, Ferrer D. Babesia shortti infection in a common kestrel (Falco tinnunculus) in Catalonia (northeastern Spain). Avian Pathol. (1999) 28:207–9. doi: 10.1080/03079459994957
65. Young KM, Corrin T, Wilhelm B, Uhland C, Greig J, Mascarenhas M, et al. Zoonotic Babesia: a scoping review of the global evidence PLoS ONE. (2019) 14:e0226781. doi: 10.1371/journal.pone.0226781
Keywords: soft ticks, Argas spp., nidicolous, cavity-nesting birds, tick-borne bacteria, tick-borne viruses, tick-borne protozoa, Spain
Citation: Palomar AM, Veiga J, Portillo A, Santibáñez S, Václav R, Santibáñez P, Oteo JA and Valera F (2021) Novel Genotypes of Nidicolous Argas Ticks and Their Associated Microorganisms From Spain. Front. Vet. Sci. 8:637837. doi: 10.3389/fvets.2021.637837
Received: 04 December 2020; Accepted: 24 February 2021;
Published: 29 March 2021.
Edited by:
Sebastián Muñoz-Leal, University of Concepcion, ChileReviewed by:
José M. Venzal, Universidad de la Republica, UruguayQuan Liu, Foshan University, China
Copyright © 2021 Palomar, Veiga, Portillo, Santibáñez, Václav, Santibáñez, Oteo and Valera. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ana M. Palomar, YW1wYWxvbWFyQHJpb2phc2FsdWQuZXM=