
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Vet. Sci. , 07 December 2020
Sec. Animal Behavior and Welfare
Volume 7 - 2020 | https://doi.org/10.3389/fvets.2020.600949
This article is part of the Research Topic Ruminant Grazing Behavior: A Tool to Improve Product Quality and Ecosystem Services View all 13 articles
 Alessandra Nicolao1,2
Alessandra Nicolao1,2 Mauro Coppa3
Mauro Coppa3 Matthieu Bouchon4
Matthieu Bouchon4 Enrico Sturaro2
Enrico Sturaro2 Dominique Pomiès1
Dominique Pomiès1 Bruno Martin1
Bruno Martin1 Madeline Koczura1*†
Madeline Koczura1*†Rearing dairy calves with their mothers could teach them how to graze, optimizing grass use, and improving their welfare and performance. We tested the short-term effects of dam-calf contact experience on grazing and social behavior of weaned calves, monitored over seven days for their first post-weaning grazing experience. “Dam” (D) calves were reared and grazed with their mothers until weaning. “Mixed” calves (M) were separated from their mothers after 4 ± 0.5 weeks, they experienced dam-calf contact, but not grazing. “Standard” (S) calves had never experienced either dam-calf contact (separated at birth) or grazing. Each group grazed an equivalent pasture plot offering heterogeneous herbage. Scan sampling of calves' activities was performed every 5 min, 6 h per day, on Days 0, 1, 2, 3, and 7. Daily, the time when calves started grazing after introduction to pasture, and the number and duration of their grazing cycles were measured. Daily activities were differentiated into ingestion, rumination, and idling. The proportion of time that calves spent grouped with other individuals or isolated, and standing or lying were recorded. When grazing, their bites were characterized by botanical family group, height of the selected bite and vegetation status. Individual average daily gains from the 2-week periods before and after grazing were calculated, and were equivalent between groups (313 ± 71 g/d). On Day 0, D-calves started grazing immediately (1 ± 4.1 min), unlike M- and S-calves (39 ± 4.1 and 23 ± 4.1 min), and D-calves grazed patches of dry grass 21.7 times less than M-calves and 16.9 times less than S-calves. Dry herbage patch preference and grazing start time differences disappeared on Day 1. Calves spent the same time ingesting and idling, but M-calves spent on average 1.6 times less ruminating than D- or S-calves. The D-calves showed grazing behavior similar to that of adult cows, selecting grasses throughout pasture utilization, although legumes and forbs were present in the grazed layer. On the contrary, M- and S-calves did not express any specific preference. The S-calves spent more time isolated but had more positive reciprocal interactions than the calves in the other groups.
Maximizing production while reducing costs and labor are the main aims of modern dairy systems. This trend often results in an intensification of farming practices, which weakens societal acceptance of dairy production systems (1). Consumers are taking ever greater interest in how their food is produced, and are increasingly aware of environmental issues and animal welfare (2, 3). In dairy production, the most common welfare concerns are the separation of calves from their dams (4) and restricted access to pasture for animals in intensive systems (5). Pasture for dairy cattle offers several advantages for animal welfare and health, such as expression of natural behavior and possible reduction of lameness and claw disorders (6–8) or increased movement with positive effects on longevity (9). Grazing systems also reduce management and feeding costs for the farmer (10, 11). In commercial dairy farms, calves are usually separated from their dams close to birth, and rarely experience grazing during their early lives (12). In France, 60 % of dairy farms use seasonal batch calving during autumn and winter, in order to turn out animals to pasture in the following spring (13). Then, calves and heifers usually graze from spring to autumn, before their first year of age, but only 2% of dairy farms turn out calves to pasture before 6 months of age (13). This strategy allows the synchronization of the peak of herbage growth and the peak of lactation of dairy cows, with fresh herbage covering a large part of their nutritional requirements (14). At the same time, calves have also grown and matured sufficiently and are able to be moved to pasture.
Le Cozler et al. (13) reported that only 4% of farmers keep calves with their dam at later than 24 h, but this practice is increasingly used. Michaud et al. (15) investigated farms using a suckling practice in France (Massif Central, East and West of France), and found that 62 farms out of 102 kept calves with their dam or with a foster cow between 1 and 60 days of age. The presence of the dam in the early stages of a calf's life can have positive effects on its social interactions, feeding behavior, and growth (16–18). The dam is the primary social model and plays an important role in the acquisition of foraging behavior and feed selection (19, 20). Pullin et al. (20) found that lambs grazing with their dam spent more time foraging, were more active, developed long-term feed preferences and learned aversion to toxic feed more effectively than lambs grazing alone. Young animals learn by emulation of social models or by trial and error, although in most cases this last is less efficient (21). Calves usually are neophobic: they tend to choose feed and places they already know, so that individual learning in a new environment takes more time than learning by social models (22, 23). Lopes et al. (24) observed that heifers with early grazing experience, compared to inexperienced heifers, affected grazing behavior and milk production only in the first days on pasture, but showed that the animals would generally adapt to a new environment and a novel feed easily, especially during their first year of life. Dairy calves that have learned to graze with their dam might therefore more efficiently recognize herbage quality and select specific patches when turned out to pasture after weaning, compared to calves that never grazed before. However, it is unclear whether this advantage holds only in the first grazing day or is more persistent.
In the present study, the following hypotheses were tested, comparing three groups of calves with contrasting rearing experience on their first grazing days after weaning. We expected calves that had experienced dam-calf contact and grazing in their early life to show grazing and probably social behavior that was different from that of inexperienced calves, and more typical of adult dairy cows. The longer dam-calf contact lasted (a few weeks or until weaning), the greater would be the expected differences in calves' social behavior. The present study also evaluated the persistence of the expected differences in grazing or social behavior in the short term after weaning.
The experiment was performed in 2019 at the INRAE experimental farm of Marcenat (DOI: https://doi.org/10.15454/1.5572318050509348E12), located in the Massif Central (45°15′N, 2°55′E; 1150 m a.s.l.). All animal-related procedures were carried out in accordance with the guidelines for animal research of the French Ministry of Agriculture and all other applicable national and European regulations for experimentation with animals (https://www.recherche-animale.org/sites/default/files/charte_nationale_portant_sur_l_ethique_de_l_experimentation_animale_243579.pdf). The experiment started February 12. The early grazing period started July 22 and ended July 29. Three breed-balanced groups of eight dairy calves (Holstein and Montbéliarde) with different experience backgrounds were compared (Table 1): a group of “Standard” calves (S) that had been separated at birth from their dam and had never experienced grazing, a group of “Dam” calves (D) that had been reared and grazed with their dam until weaning, and a group of “Mixed” calves (M) that had been separated from their dam at 4 ± 0.5 weeks of age and had never experienced grazing. All calves were weaned at age of 10.9 ± 1.1 weeks. Before weaning, D-calves were housed separately from their dams at night and had free access to the dam cowshed during the day. Starting from May 5, when the calves were 4.6 ± 3.2 weeks old, the day cowshed access was replaced by free access to pasture with dams. The M-calves, until age 4.0 ± 0.5 weeks, were reared in the same way, except that they had no access to pasture. From this age until weaning, they were reared like S-calves, i.e., in separate housing and fed bulk milk with an automatic milk dispenser. D- and M-calves were reunited with their dams after morning milking at 9:00 a.m. and separated before evening milking at 3:30 p.m. At weaning, all calves were moved to a new pen, with one pen for each group to prevent mixing. In this pen, calves ingested 0.5 kg/d/calf of hay and 2.0 kg/d/calf of concentrate (Startivo, Centraliment, 15006 Aurillac). Hay was distributed in the evening with no refusal left in the morning. Concentrate was distributed half in the morning and half in the evening, until the end of the study. After the last weaning, all calves spent at least six days indoors all together to allow the latest weaned calves to adapt to the new conditions. At the beginning of the grazing period (week 15), D-, M- and S-calves were, respectively, 14.9 ± 3.2, 16.1 ± 2.8 and 15.3 ± 3.6 weeks old and weighed 131 ± 18.3 kg, 123 ± 17.4 kg, and 128 ± 23.5 kg respectively, on average. They had been weaned for 30 ± 22, 33 ± 20 and 33 ± 24 days, respectively. Calves were turned out to pasture on July 22 (Day 0), from 9:00 a.m. to 5:00 p.m.
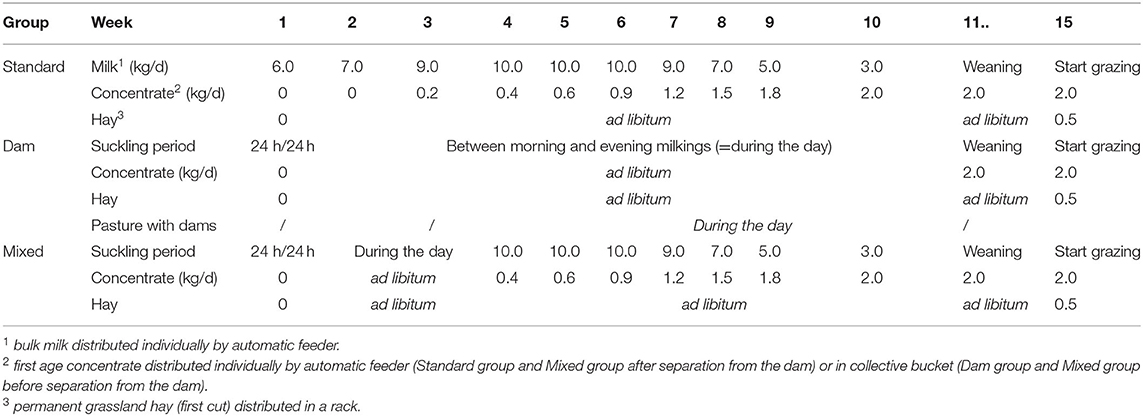
Table 1. Feeding plan (milk, concentrate, and hay) of the three groups of calves (Standard, Dam, Mixed) during the first 15 weeks of age.
Calves grazed a permanent pasture divided into three equivalent neighboring 0.15 ha plots. No close visual contact was allowed between the three groups of animals, by fencing the plots so that they were at least 15 m apart. To encourage grazing selection for all three groups of calves, the whole plot was strip-mown 28 days before the grazing period started. At Day 0, plots were thereby composed of alternate 3 m strips dominated by mature vegetation and vegetative regrowth. The botanical composition of the whole pasture was determined using the vertical point-quadrat method from Daget and Poissonet (25). The pasture was dominated by Lolium perenne (39.0%), Agrostis tenuis (15.0%), and Trifolium repens (13.5%). Rumex obtusifolius was also present (3.1%). At the beginning of the experiment, three 10 cm × 3 m grass samples were collected on each plot, perpendicularly to the mown and unmown strips, equally harvesting the same length from both. They were oven-dried at 60°C for 72 h and analyzed for proximate composition as described by Coppa et al. (26) (Table 2).
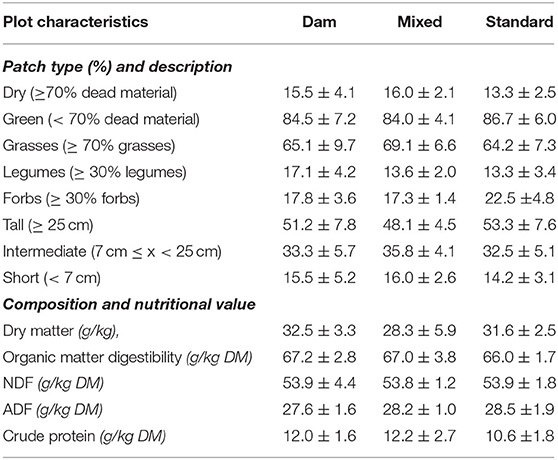
Table 2. Characteristics of vegetation offered on the experimental plots (mean ± standard deviation).
Calves were weighed once a week, and individual average daily gain (ADG) from the 2-week periods before and after the grazing period started was calculated.
Individual daily activities and behavior were observed by scan sampling at 5-min intervals (27, 28) on the day the calves encountered the pasture for the first time (Day 0), the next three days (Day 1, Day 2, Day 3) and one week later (Day 7), focusing on the first days as most of the differences were expected here (24). On each plot, four calves were randomly assigned to two observers for 6 h per day (9:00–12:00 a.m. and 2:00–5:00 p.m.). For observations, calves were always identified by the same numbers painted on their back. At the end of the afternoon, the calves went back indoors for the night where they were fed with hay and concentrate (Startivo, Centraliment, 15006 Aurillac). Observers, randomly assigned to a group of calves, changed experimental group between each morning and afternoon. Each day, the time taken by calves to start grazing was measured. A calf was considered to have started grazing if it was observed taking a bite in at least three out of four successive observations (29), following the flowchart in Figure 1. From the time the calf started grazing, the grazing cycle lasted until it showed more than three other successive activities (i.e., it stopped grazing for at least 15 min), according to Manzocchi et al. (30). The duration of a grazing cycle and the number of grazing cycles, as just described, were calculated following the flowchart in Figure 2. Daily activities were then differentiated into three groups: ingestion (grazing and drinking water), rumination, and idling. The latter comprised four subcategories: resting (observation, sleep, self-grooming), positive interactions (licking, sniffing, head play), negative interactions (head-butting, pushing, fighting) and ad hoc activities (walking, exploring, stereotypies, vocalizing) (Table 3). The daily proportion of ingestion, rumination and idling time was calculated as a percentage of the total daily observations. The daily proportion of resting time, socializing time and ad hoc activities was calculated as a percentage of the idling activities. Each time one of the activities was recorded, observers also indicated whether the calf was grouped with other calves or isolated, i.e., at least 3 m away from other calves, and whether it was standing or lying. The daily proportions of time spent grouped and standing were calculated over the total number of observations of the day. When calves were grazing, their bites were characterized by botanical group (grasses, legumes and forbs), the height of the selected bite (tall, intermediate, short vegetation) and the vegetation status (“dry” or “green”), according to Koczura et al. (31). Briefly, patches were characterized according to the visually estimated proportion of dry senescent herbage, of botanical family groups and of their height (26, 32). A patch was coded as “dry” if the dry senescent vegetation represented more than 70 % of the bite, as “green” if it was <70%; as dominated by “grasses” if the bite contained more than 70% of grasses, by legumes or forbs if they represented more than 30%; tall if herbage height was ≥ 25 cm, and small if it was ≤ 7 cm, as detailed in Table 2. Observers were able to get close to calves due to their adaptation to human presence achieved during the pre-weaning experiment. When calves ingested forbs, observers reported whether or not they selected Rumex thanks to a binary variable (1 = the calf tried to eat Rumex at least one time in the observation day). The daily proportion of vegetation type ingested by calves was calculated as a percentage of observations comprising the vegetation type compared to the total number of grazing observations of the day.
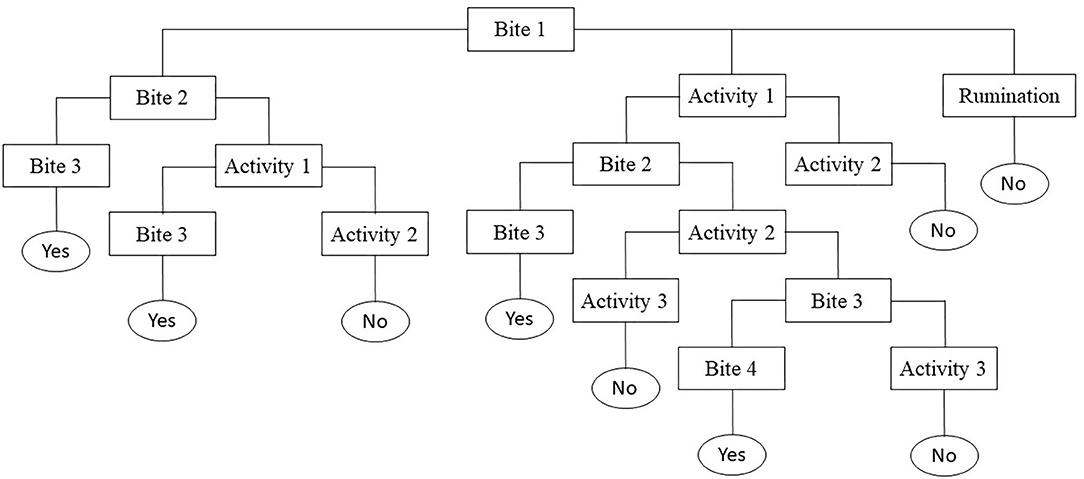
Figure 1. Flowchart of the conceptual scheme used during observations to tell whether calves started grazing at Bite 1 (Yes = at least three bites over four observations; No = flowchart restart to the next observation).
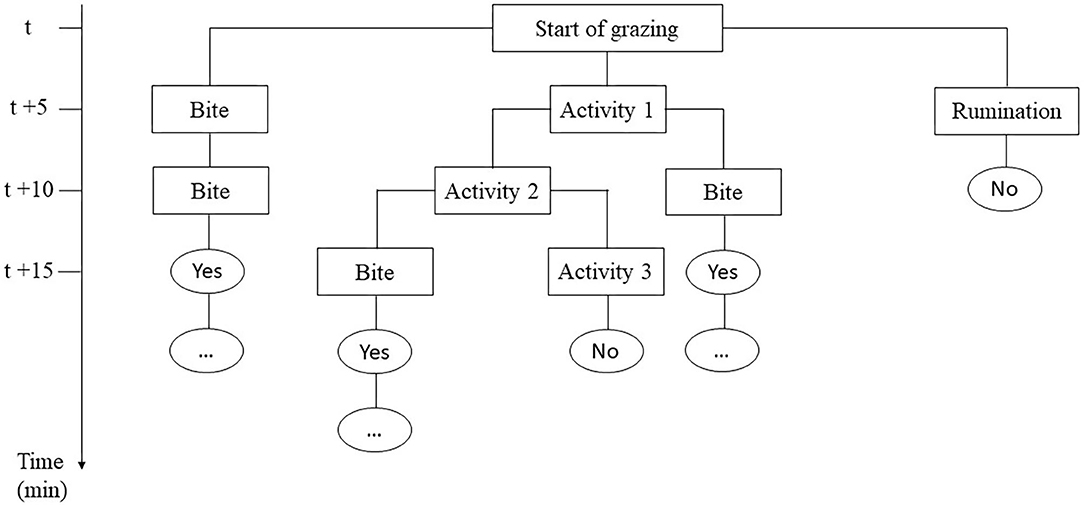
Figure 2. Flowchart of the conceptual scheme used during observations to tell whether a grazing cycle was established (Yes or No) and measure its duration.
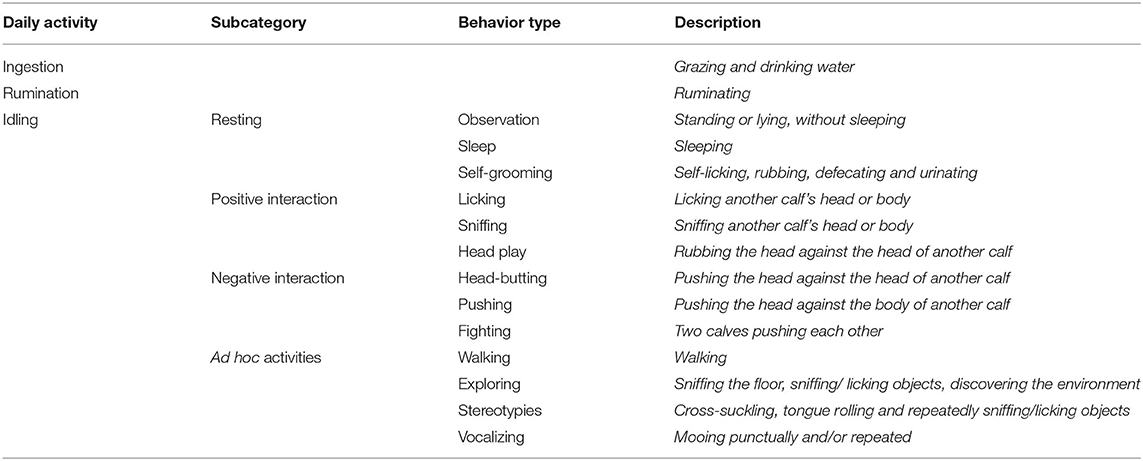
Table 3. Description of daily activities recorded by scan-sampling differentiated in four subcategories.
The weather was exceptionally hot on the afternoons of Day 2 and Day 3. The average maximum daily temperature during these afternoons was 31.2°C, whereas between 2000 and 2019, the average maximum temperature in July was 21.6°C (INRAE CLIMATIK 2.1.5, Marcenat weather station). Behavior observations at pasture were therefore made throughout the day on Day 0, Day 1, and Day 7, but only in the morning on Day 2 and Day 3. The daily ingestion, rumination and idling activities, together with the number and duration of grazing cycles, were accordingly calculated only for Day 0, Day 1, and Day 7, as the morning alone was not considered representative of the ingestion and rumination cycles of a whole day. On the other hand, the characterization of grazed bites and time needed to start grazing were calculated on mornings only for all days, the numbers of bites observed during the morning being considered sufficient and representative to express preference, as differences between morning and afternoon on those days were equivalent.
Daily activities and grazing cycles were analyzed with a repeated MIXED model on SAS 9.4 software (SAS Institute Inc., Cary, NC, USA). Group (Dam, Mixed or Standard), day (only 0, 1, or 7) and their interaction were included as fixed effects. The individual calf was considered as the subject of repetition, with day being the repeated factor. We used a compound symmetry covariance structure. Time to start grazing and herbage selection were analyzed with the same model, except that the day effect included all days. Average daily gain was analyzed with a similar repeated model, which included group, period (before or after pasture) and their interaction as fixed effects, calf as subject, and period as repeated factor. In this last model, the number of days since each calf had been weaned was used as a covariate. The effect of the age and BW of calves were tested as covariates as well, but were found to be non-significant, and so were not finally included in the model. For all data, normality of residuals was checked using the Shapiro-Wilk test. The frequency of times calves tried at least 1 time to include Rumex in their bites was compared between groups using a Chi2 test. Significance was set at p < 0.05.
During the 2 weeks before start of grazing, the ADG of D-, M- and S-calves did not differ significantly (p = 0.177), at 285, 355, and 480 g/d, respectively. During the following 2 weeks it increased by 313 g/d on average for all the groups.
Once turned out to pasture, the daily activities of the three groups of weaned dairy calves did not strongly differ (interaction groups × day non-significant). Overall, during Days 0, 1, and 7, calves in the three groups spent almost the same time ingesting (p = 0.081, on Day 1 M-calves tended to spend less time ingesting than D- and S-calves) and the same time idling (Table 4), but M-calves spent on average less time ruminating than D- and S-calves (1.54 times less). During idling activities, D-calves spent more time resting than M- or S-calves (1.16 and 1.14 times more, respectively), and M-calves spent more time in ad hoc activities than D-calves (1.24 times more). The S-calves had more positive social interactions than the calves in the other two groups. On Day 0 and Day 1, S-calves spent more time lying than D- or M-calves (Figure 3I). On Day 0 and Day 1 they spent more time isolated than calves in the other two groups (Figure 3II).
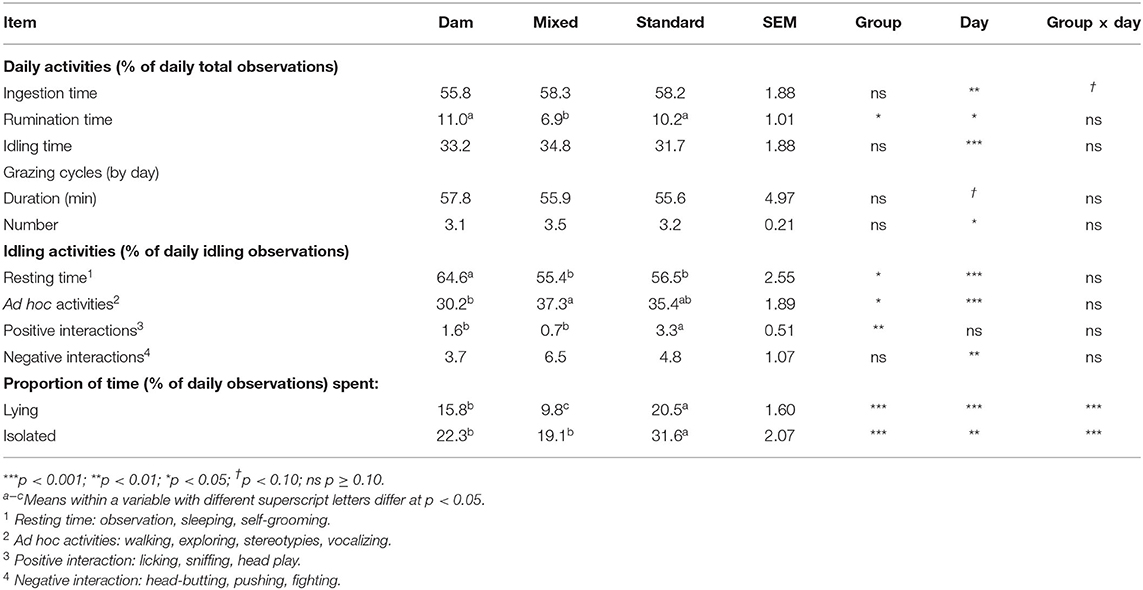
Table 4. Effect of early dam-calf contact and grazing experience on post-weaning daily activities and grazing cycles of dairy calves (Day 0, 1 and 7 after start of grazing).
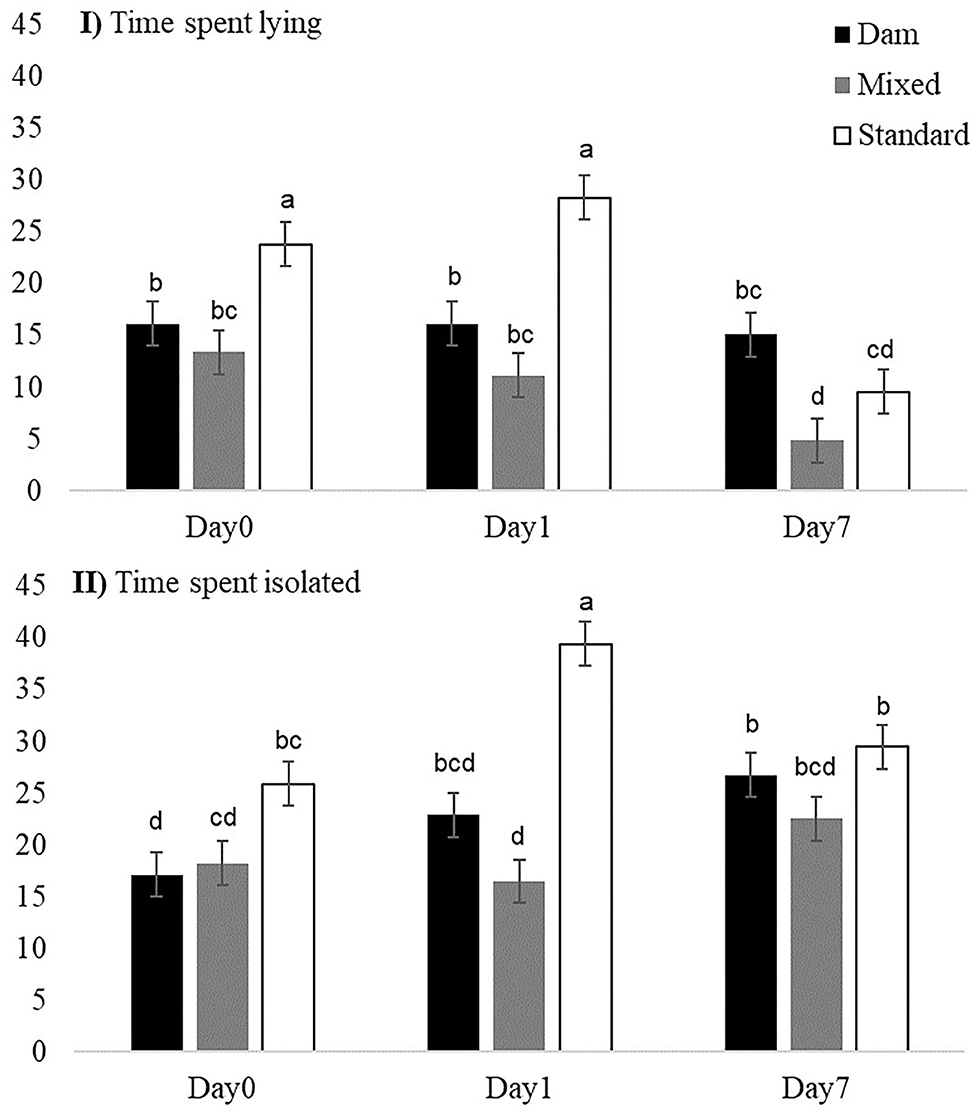
Figure 3. Effect of early dam-calf contact and grazing experience on (I) daily time spent lying (%) and (II) isolated (%) by calves on Day 0, Day 1, and Day 7 at pasture. Bars are standard errors. a−dMeans within a variable with different superscript letters differ at p < 0.05.
When moved to pasture, D-calves started grazing immediately (Table 5), whereas it took S-calves 23 ± 4.1 min to actively start to graze. The M-calves needed a further 20 min. On Day 0, the herbage selection was different between groups: D-calves grazed “dry” patches 21.71 times less than M-calves and 16.90 times less than S-calves. On Day 0, no differences between groups were observed for botanical composition and height, except for forbs: on that day, S-calves grazed 13.73 times more forbs than M-calves and 3.89 times more than D-calves. On Day 1, Day 3, and Day 7 all three groups of calves started grazing 5 ± 2.8 min after arriving on pasture, whereas M- and S-calves started grazing 15 ± 0.7 min after D-calves on Day 2. From Day 1, M- and S-calves reduced their proportion of “dry” patches to meet that of D-calves, with no longer any significant differences between groups. Overall, we found that the proportion of tall vegetation in the bites decreased from Day 1 to Day 7 and conversely that the proportion of short vegetation increased in the bites from Day 1 to Day 7. On Day 2 and Day 3, M-calves showed a higher proportion of intermediate vegetation than D- and S-calves (2.07 and 3.03 times more, on average). D-calves continuously maintained stable the proportion of grasses in their bites throughout the plot utilization, while M- and S-calves decreased their proportion over time (0.75 times less from Day 0 to Day 7, on average), increasing in parallel those of legumes and forbs (7.57 and 4.46 more times on average, respectively). On Day 0, none of D-calves grazed Rumex, on the contrary to M- and S-calves (4 and 6 calves, respectively) (Figure 4). This difference disappeared in the following days, already on Day 1.
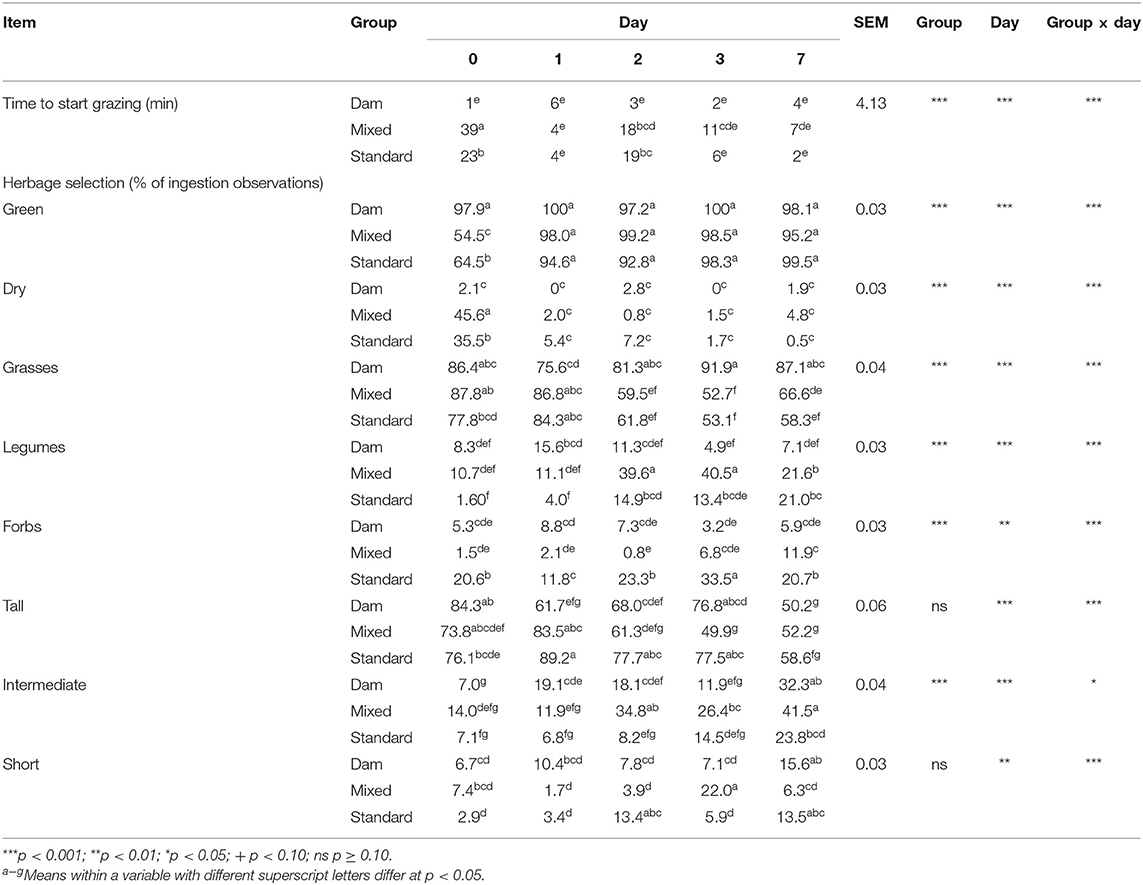
Table 5. Effect of early dam-calf contact and grazing experience on time to start grazing after introduction to pasture and characteristics of selected bites by dairy calves.
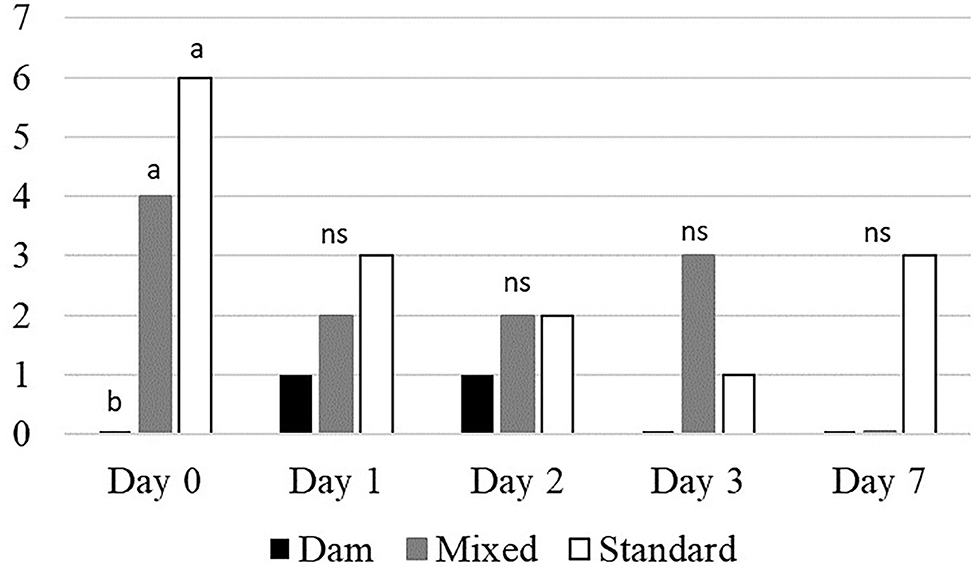
Figure 4. Overall number of calves that tasted Rumex each day (n = 8 calves × 3 groups). a−bMeans with different superscript differ at p < 0.05, Khi2 test; ns, not significant.
To our knowledge, only a few published studies have focused on dairy calf grazing behavior, and this is the first time that the effects of an early dam-calf contact have been investigated on calf grazing behavior, directly after weaning. As expected, the main differences in calf grazing behavior were mostly observed on the first day on pasture (Day 0): D-calves started grazing immediately when moved to pasture, whereas S- and M-calves started 23 and 43 min later, respectively. In several studies, it is reported that inexperienced heifers need a few hours (12) or a few days (24) to start grazing. This lag occurs even when animals are put on pasture with experienced heifers. In our study, the three groups were separated to prevent visual contact between experienced and non-experienced animals. Calves that had experienced pasture with dams in their early life then immediately remembered how to graze, unlike calves from the other groups. This is consistent with findings of Lopes et al. (24), who observed that heifers that had once experienced pasture instantly remembered how to graze the following year. The M- and S-calves took slightly longer to start to graze, probably because they had no social model or experienced individuals to emulate (33, 34). Also, inexperienced heifers spend more time exploring and tasting grass than ingesting it, compared to experienced heifers (33), which was numerically the case for our M- and S-calves here before they started grazing, even though exploring time was not long enough to statistically analyze it (data not shown). After starting grazing (on Day 0), M- and S-calves selected mainly “dry” patches, unlike D-calves, which directly grazed only “green” ones. This suggests that inexperienced calves could be neophobic (22): they were probably reluctant to try novel feed, and without a social model, were inclined to choose feed they already knew, or with similar characteristics to hay. Nevertheless, these differences were no longer seen in the following days, showing that calves can soon learn how to graze and cope with novelty.
Once they started grazing, all the calves followed the same pattern from Day 0 to Day 7: they first selected tall vegetation and then intermediate and short herbage as pasture utilization progressed. This is consistent with the selection of vegetation by stratum by experienced grazing cows under rotational grazing (35): once the upper layer is grazed, the height of the patch decreases, moving down to the lower layers (26). However, the botanical selection of the vegetation seemed different between groups: M- and S-calves ingested mainly grasses during the first days, as grasses are almost exclusive in the top layer, and then increasingly legumes and forbs [present in the intermediate and low layers, because of their smaller size; (34)]. On the contrary, D-calves constantly selected grasses until the seventh day of observation, whatever the height of the layer present on the plot. This suggests that inexperienced calves did not select vegetation according to its botanical composition, but rather ingested species according to their presence in the topmost layer as they utilized the plot. Calves that had experienced pasture with dams seem to have learnt to graze like adult cows, which are known to select grasses even on biodiverse pasture (26, 35). Furthermore, M- and S-calves tried to ingest Rumex, especially during the first day at pasture, while D-calves rarely approached it. Rumex is one of the main oxalate-producing plants: oxalate can cause poisoning in livestock if present in 10% or more of the dry weight of the plant (36). It is therefore important that cattle learn how to avoid it. This suggests again that calves that have grazed with their dams learnt to choose or avoid some plants (20), while inexperienced calves learnt by trial and error (21). Even though calves that did not experience grazing showed different grazing behavior than D-calves on the first day at pasture, their behavior evolved very quickly (less than a week) into behavior similar to adult cows. This implies that dam-calf contact close to birth has little impact on longer term grazing behavior.
Having experienced dam contact and/or pasture affected the time to start grazing and herbage selection behavior of dairy calves, but did not influence their daily ingestion time or the duration and number of their grazing cycles. All the groups of calves, regardless of their different previous experience, had the same grazing rhythm throughout the trial: this confirms that inexperienced animals exhibit similar grazing times to experienced animals, as found by Lopes et al. (24) and Hessle et al. (19). While idling, M-calves behaved differently from D-calves in ad hoc activities (i.e., walking, exploring, stereotypies and vocalizing). This was consistent with the finding of Arrazola et al. (33) highlighting that inexperienced calves spent more time walking and exploring compared to experienced calves, that spent more time inactive. Besides, M-calves spent less time lying than the calves in the other groups. Wilcox et al. (37) demonstrated that standing behavior could indicate a stress condition of the calves, especially in case of chronic stress. Even if we did not directly measure stress of the calves, it could not be excluded that repeating stress factors over time by splitting separation and weaning could have induced a stressful behavior for M-calves. We also found that M-calves spent less time ruminating than D-calves, while the latter spent more time resting. As rumination time is proportional to forage intake, this result suggests that although the ingestion time was similar between groups, M-calves may have ingested less forage than D-calves, as found by Arrazola et al. (33). However, the calves' daily forage intake was not monitored in the present study. A different digestibility of dry senescent and vegetative or tall and short patches (leaf to stem ratio) could also have affected rumination time, but the day by day differences among groups in patches characteristics are not consistent with the trend observed in rumination time. Furthermore, no differences in ADG between groups were observed before and after calves started grazing, even though in the literature inexperienced grazers were found to be nutritionally disadvantaged because of modest foraging behavior that could affect their live weight gains (19). This suggests that the calf daily forage intake was not different between groups. We cannot therefore confirm that the foraging skills of inexperienced calves were inferior, but we can assert that they were not typical of an adult cow.
To the best of our knowledge, this is the first study to investigate dam-calf contact effects after weaning. Valníčková et al. (38) did not find any effect of dam-calf contact on social interactions or play behavior during colostrum feeding. Le Neindre and Sourd (39) found that heifers reared with foster cows dominated more than heifers reared without cow contact. We thus expected that calves reared with their dams would be more sociable or have more dominant behaviors than artificially reared ones, but we found no differences in negative interactions (i.e., dominance behaviors, such as head-butting, pushing, or fighting) between groups at pasture. Nevertheless, we observed that S-calves had more positive interactions with their companions (particularly licking) than did calves in the other two groups. Pinheiro Machado et al. (40) found that licking behavior between grazing dairy cows was not a random choice but showed a companion's preference for socio-positive interactions. Furthermore, they observed that licking was more persistent in long-established social groups. This could suggest that D- and M-calves may have created bonds rather with dams than with other calves, compared to S-calves, but this point requires further investigation. Besides, the higher proportion of time spent isolated by S-calves, compared to D- and M-calves, could suggest that they exhibit less gregarious behavior. It is however difficult to interpret, because of a lack of literature on this topic.
Early life experience with dam and/or pasture influenced calves' foraging skills in the short term after weaning, especially on the first grazing day. Calves that had already experienced pasture with their dams immediately started to graze the day they were turned out to pasture in groups after being weaned. They instantly selected “green” patches of vegetation while grazing, unlike calves that had been housed indoors the whole time, which ingested predominantly senescent herbage on their first day. Daily ingestion time and duration and number of grazing cycles were not affected by previous experience. Nevertheless, botanical selection throughout pasture utilization and rejection of toxic plants (Rumex) showed that young calves could already exhibit post-weaning grazing behavior similar to that of adult cows when put on pasture early with their dam. This study provides evidence that separation of dairy calves from their dams close to birth has little impact on grazing behavior, as they grazed similarly to adult cows already in the short term (less than a week after being introduced to pasture). We spotted some differences in social behavior between the calves that experienced dam-calf contact and those that did not, but these differences are not easy to interpret and should be investigated in future studies. Further investigation is also needed to evaluate whether an early grazing experience with their dams could provide positive effects on behavior in the long term and performance in the future lactating careers of these calves.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethical review and approval was not required for the animal study because the experiment was performed at Marcenat, INRAE experimental farm (Certificate of Authorization to Experiment on Living Animals No. D 15-114-01). No ethical approval was required because defined severity level was 0.
AN, MK, MC, MB, DP, and BM contributed to the conception and design of the study. AN, MK, MC, DP, and BM participated in the collection of behavioral observations data. AN and MK did the data curation and treatment. MK performed the statistical analysis. AN and MK wrote the first draft of the manuscript. MK and MC supervised the experiment and the writing of the manuscript. All authors contributed to manuscript revision, and all read and approved the submitted version.
This study was part of the EU-project ProYoungStock funded by CORE Organic Cofund funding bodies, which are partners of the Horizon 2020 ERA-Net project CORE Organic Cofund. This research was financed by the French government IDEX-ISITE initiative 16-IDEX-0001 (CAP 20-25). AN was supported by a doctoral fellowship from the Fondazione Cariparo at University of Padova (Italy).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors thank the staff of the INRAE farm at Marcenat (UE Herbipôle) for animal care, Anna Mathieu for her active participation in the trial, Nadège Aigueperse for advice on experimental design, and Cyril Labonne, André Guittard and Vanessa Conte for their participation in the collection of behavioral observations data.
1. Cembalo L, Caracciolo F, Lombardi A, Del Giudice T, Grunert KG, Cicia G. Determinants of individual attitudes toward animal welfare-friendly food products. J Agricult Environ Ethics. (2016) 29:237–54. doi: 10.1007/s10806-015-9598-z
2. Brombin A, Pezzuolo A, Brščić M. Are we ready for the big change in the dairy production system? Res Vet Sci. (2019) 126:17–9. doi: 10.1016/j.rvsc.2019.08.006
3. Weary DM, Von Keyserlingk MAG. Public concerns about dairy-cow welfare: How should the industry respond? Anim Prod Sci. (2017) 57:1201–9. doi: 10.1071/AN16680
4. Busch G, Weary DM, Spiller A, Von Keyserlingk MAG. American and German attitudes towards cowcalf separation on dairy farms. PLoS ONE. (2017) 12:174013. doi: 10.1371/journal.pone.0174013
5. Schuppli CA, von Keyserlingk MAG, Weary DM. Access to pasture for dairy cows: responses from an online engagement. J Anim Sci. (2014) 92:5185–92. doi: 10.2527/jas.2014-7725
6. Charlton GL, Rutter SM. The behaviour of housed dairy cattle with and without pasture access: a review. Appl Anim Behav Sci. (2017) 192:2–9. doi: 10.1016/j.applanim.2017.05.015
7. Hernandez-Mendo O, Von Keyserlingk MAG, Veira DM, Weary DM. Effects of pasture on lameness in dairy cows. J Dairy Sci. (2007) 90:1209–14. doi: 10.3168/jds.S0022-0302(07)71608-9
8. Olmos G, Boyle L, Hanlon A, Patton J, Murphy JJ, Mee JF. Hoof disorders, locomotion ability and lying times of cubicle-housed compared to pasture-based dairy cows. Livestock Sci. (2009) 125:199–207. doi: 10.1016/j.livsci.2009.04.009
9. Ruhland K, Gränzer W, Groth W, Pirchner F. Blood levels of hormones and metabolites, erythrocytes and leukocytes and respiration and pulse rate of heifers after alpage. J Anim Breed Genet. (1999) 116:415–23. doi: 10.1046/j.1439-0388.1999.00203.x
10. Hanson JC, Johnson DM, Lichtenberg E, Minegishi K. Competitiveness of management-intensive grazing dairies in the mid-Atlantic region from 1995 to 2009. J Dairy Sci. (2013) 96:1894–904. doi: 10.3168/jds.2011-5234
11. White SL, Benson GA, Washburn SP, Green JT. Milk production and economic measures in confinement or pasture systems using seasonally calved Holstein and Jersey cows. J Dairy Sci. (2002) 85:95–104. doi: 10.3168/jds.S0022-0302(02)74057-5
12. Costa JHC, Costa WG, Weary DM, Machado Filho LCP, von Keyserlingk MAG. Dairy heifers benefit from the presence of an experienced companion when learning how to graze. J Dairy Sci. (2016) 99:562–8. doi: 10.3168/jds.2015-9387
13. Le Cozler Y, Recoursé O, Ganche E, Giraud D, Danel J, Bertin M, et al. A survey on dairy heifer farm management practices in a Western-European plainland, the French Pays de la Loire region. J Agricult Sci. (2012) 150:518–33. doi: 10.1017/S0021859612000032
14. Ramsbottom G, Horan B, Berry DP, Roche JR. Factors associated with the financial performance of spring-calving, pasture-based dairy farms. J Dairy Sci. (2015) 98:3526–40. doi: 10.3168/jds.2014-8516
15. Michaud A, Cliozier A, Bec H, Chassaing C, Disenhaus C, Drulhe T, et al. Déléguer l ' allaitement des veaux laitiers aux vaches? Résultats d'enquêtes auprès des éleveurs. Renc Rech Ruminants. (2018) 24:66–9.
16. Flower FC, Weary DM. Effects of early separation on the dairy cow and calf: 2. Separation at 1 day and 2 weeks after birth. Appl Anim Behav Sci. (2001) 70:275–84. doi: 10.1016/S0168-1591(00)00164-7
17. Krohn CC. Effects of different suckling systems on milk production, udder health, reproduction, calf growth and some behavioural aspects in high producing dairy cows - a review. Appl Anim Behav Sci. (2001) 72:271–80. doi: 10.1016/S0168-1591(01)00117-4
18. Wagner K, Seitner D, Barth K, Palme R, Futschik A, Waiblinger S. Effects of mother versus artificial rearing during the first 12 weeks of life on challenge responses of dairy cows. Appl Anim Behav Sci. (2015) 164:1–11. doi: 10.1016/j.applanim.2014.12.010
19. Hessle AK. Effects of social learning on foraging behaviour and live weight gain in first-season grazing calves. Appl Anim Behav Sci. (2009) 116:150–5. doi: 10.1016/j.applanim.2008.08.004
20. Pullin AN, Pairis-Garcia MD, Campbell BJ, Campler MR, Proudfoot KL, Fluharty FL. The effect of social dynamics and environment at time of early weaning on short- and long-term lamb behavior in a pasture and feedlot setting. Appl Anim Behav Sci. (2017) 197:32–9. doi: 10.1016/j.applanim.2017.09.003
21. Thorhallsdottir AG, Provenza FD, Balph DF. Ability of lambs to learn about novel foods while observing or participating with social models. Appl Anim Behav Sci. (1990) 25:25–33. doi: 10.1016/0168-1591(90)90066-M
22. Launchbaugh KL, Provenza FD, Werkmeister MJ. Overcoming food neophobia in domestic ruminants through addition of a familiar flavor and repeated exposure to novel foods. Appl Anim Behav Sci. (1997) 54:327–34. doi: 10.1016/S0168-1591(96)01194-X
23. Costa JHC, Daros RR, von Keyserlingk MAG, Weary DM. Complex social housing reduces food neophobia in dairy calves. J Dairy Sci. (2014) 97:7804–10. doi: 10.3168/jds.2014-8392
24. Lopes F, Coblentz W, Hoffman PC, Combs DK. Assessment of heifer grazing experience on short-term adaptation to pasture and performance as lactating cows. J Dairy Sci. (2013) 96:3138–52. doi: 10.3168/jds.2012-6125
25. Daget P, Poissonet J. Une méthode d'analyse phytologique des prairies. Ann Agronom. (1971) 22:5–41.
26. Coppa M, Farruggia A, Ravaglia P, Pomiès D, Borreani G, Le Morvan A, et al. Frequent moving of grazing dairy cows to new paddocks increases the variability of milk fatty acid composition. Animal. (2015) 9:604–13. doi: 10.1017/S1751731114003000
27. Mitlöhner FM, Morrow-Tesch JL, Wilson SC, Dailey JW, McGlone JJ. Behavioral sampling techniques for feedlot cattle. J Anim Sci. (2001) 79:1189–93. doi: 10.2527/2001.7951189x
28. Palacio S, Bergeron R, Lachance S, Vasseur E. The effects of providing portable shade at pasture on dairy cow behavior and physiology. J Dairy Sci. (2015) 98:6085–93. doi: 10.3168/jds.2014-8932
29. Farruggia A, Dumont B, D'Hour P, Egal D. How does protein supplementation affect the selectivity and performance of Charolais cows on extensively grazed pastures in late autumn? Grass For Sci. (2008) 63:314–23. doi: 10.1111/j.1365-2494.2008.00644.x
30. Manzocchi E, Koczura M, Coppa M, Turille G, Kreuzer M, Berard J. Grazing on upland pastures part-time instead of full-time affects the feeding behavior of dairy cows and has consequences on milk fatty acid profiles. Animalsl. (2019) 9:908; doi: 10.3390/ani9110908
31. Koczura M, Martin B, Bouchon M, Turille G, Berard J, Farruggia A, et al. Grazing behaviour of dairy cows on biodiverse mountain pastures is more influenced by slope than cow breed. Animal. (2019) 13:2594–602. doi: 10.1017/S175173111900079X
32. Dumont B, Rook AJ, Coran C, Röver KU. Effects of livestock breed and grazing intensity on biodiversity and production in grazing systems. 2. Diet selection. Grass For Sci. (2007) 62:159–171. doi: 10.1111/j.1365-2494.2007.00572.x
33. Arrazola A, Dicker K, Vasseur E, Bergeron R. The effect of early housing and companion experience on the grazing and ruminating behaviour of naïve heifers on pasture. Appl Anim Behav Sci. (2020) 226:104993. doi: 10.1016/j.applanim.2020.104993
34. Shingu Y, Ueda K, Tada S, Mitani T, Kondo S. Effects of the presence of grazing-experienced heifers on the development of foraging behavior at the feeding station scale for first-grazing season calves. Anim Sci J. (2017) 88:1120–127. doi: 10.1111/asj.12735
35. Coppa M, Farruggia A, Pradel P, Lombardi G, Martin B. An improved grazed class method to estimate species selection and dry matter intake by cows at pasture. Ital J Anim Sci. (2011) 10:e13. doi: 10.4081/ijas.2011.e13
36. James LF. Oxalate poisoning in livestock. In: Keeler RF, Van Kampen KR, James LF, editors. Effects of Poisonous Plants on Livestock. Amsterdam: Publusher Academic Press (1978). doi: 10.1016/b978-0-12-403250-7.50020-0
37. Wilcox CS, Schutz MM, Rostagno MR, Lay DC, Eicher SD. Repeated mixing and isolation: measuring chronic, intermittent stress in Holstein calves. J Dairy Sci. (2013) 96:7223–33. doi: 10.3168/jds.2013-6944
38. Valníčková B, Stěhulová I, Šárová R, Špinka M. The effect of age at separation from the dam and presence of social companions on play behavior and weight gain in dairy calves. J Dairy Sci. (2015) 98:5545–56. doi: 10.3168/jds.2014-9109
39. Le Neindre P, Sourd C. Influence of rearing conditions on subsequent social behaviour of Friesian and Salers heifers from birth to six months of age. Appl Anim Behav Sci. (1984) 12:43–52. doi: 10.1016/0168-1591(84)90095-9
Keywords: grazing behavior, dairy calves, grazing experience, dam-calf contact, post-weaning, social interactions, first grazing
Citation: Nicolao A, Coppa M, Bouchon M, Sturaro E, Pomiès D, Martin B and Koczura M (2020) Early-Life Dam-Calf Contact and Grazing Experience Influence Post-Weaning Behavior and Herbage Selection of Dairy Calves in the Short Term. Front. Vet. Sci. 7:600949. doi: 10.3389/fvets.2020.600949
Received: 31 August 2020; Accepted: 16 November 2020;
Published: 07 December 2020.
Edited by:
Emily Patterson-Kane, Independent Researcher, Rolling Meadows, IL, United StatesReviewed by:
Tony Waterhouse, Scotland's Rural College, United KingdomCopyright © 2020 Nicolao, Coppa, Bouchon, Sturaro, Pomiès, Martin and Koczura. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Madeline Koczura, bWFkZWxpbmUua29jenVyYUBpbnJhZS5mcg==
†Present address: Madeline Koczura, Université Clermont Auvergne, INRAE, VetAgro Sup, UMR0874 Ecosystème Prairial, Clermont-Ferrand, France
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.