 Hans-Peter Fuehrer1*
Hans-Peter Fuehrer1* Ana Margarida Alho2
Ana Margarida Alho2 Feodora Natalie Kayikci1Bita Shahi Barogh1Hugo Rosa3José Tomás3Hugo Rocha3Josef Harl4
Feodora Natalie Kayikci1Bita Shahi Barogh1Hugo Rosa3José Tomás3Hugo Rocha3Josef Harl4 Luís Madeira de Carvalho2*
Luís Madeira de Carvalho2*- 1Department of Pathobiology, Institute of Parasitology, University of Veterinary Medicine, Vienna, Austria
- 2CIISA - Centro de Investigação Interdisciplinar em Sanidade Animal, Faculdade de Medicina Veterinária, Universidade de Lisbon, Lisbon, Portugal
- 3Guarda Nacional Republicana, Lisbon, Portugal
- 4Department of Pathobiology, Institute of Pathology, University of Veterinary Medicine, Vienna, Austria
Vector-borne diseases of zoonotic and/or veterinary relevance have been increasingly reported in horses globally, although data regarding working and military horses is lacking. Portuguese military horses may constitute a risk group for these pathogens, as they frequently work outdoors in various regions of the country. This study included 101 apparently healthy horses belonging to the Portuguese National Republican Guard. Blood samples were analyzed to determine the presence and prevalence of piroplasms, Anaplasmataceae, Rickettsia spp., and filarioid helminths. Overall 32.7% of the horses gave positive results for Theileria equi. Two genotypes of T. equi were verified. No positive results were recorded for Anaplasma spp., Rickettsia spp., filarioid helminthes, and Babesia caballi. As equine piroplasmosis is a severe infectious tick-borne disease responsible for significant losses in equine production and with numerous impacts in the international movement of horses, adequate treatment, and preventive measures are needed to reduce exposure to vectors and future infections.
Introduction
Vector-borne diseases (VBDs) have been increasingly reported in horses worldwide (1). Several equine VBDs are of zoonotic relevance and horses potentially serve as sentinels for human infections. The distribution and spread of vector-borne pathogens are limited by the presence of competent arthropod vectors (e.g., ticks, fleas, and mosquitoes) capable of transmitting these pathogens. Military horses may constitute a risk group for VBDs, as they frequently work outdoors in different areas and thus are exposed to vectors present there.
Equine piroplasmosis is a tick-borne disease of equids such as horses, donkeys, mules, and zebras, and is caused by the protozoan parasites Theileria equi and Babesia caballi (2). Infected animals may carry B. caballi for several years and T. equi for a whole lifetime (2).
Theileria equi (Laveran, 1901) Mehlhorn, Schein 1998 is one of the most important pathogens of horses in many parts of the world, including Southern Europe. Clinical signs range from asymptomatic to acute, subacute, and chronic cases with fever, anemia, inappetence, and spleno- and hepatomegaly. In severe cases, the infection can lead to death. Transplacental transmission from mares to fetuses can lead to abortion (3), causing significant losses in the equine industry. Additionally, international horse trade facilitates the spread to non-endemic areas (4). Currently, more than 20 potential tick vectors of T. equi are known (5, 6). Species of the genera Dermacentor, Rhipicephalus, and Hyalomma are known as competent vectors and several Ixodes, Haemaphysalis, and Amblyomma species are discussed to transmit the parasite as well (2). Various tick species that may act as vectors for equine piroplasms are present in mainland Portugal, namely Rhipicephalus sanguineus, Rhipicephalus annulatus, Rhipicephalus bursa, Dermacentor marginatus, Hyalomma lusitanicum, and Hyalomma marginatum (7, 8). Several genotypes of T. equi have been reported in different populations of equids (2, 9). There is evidence that specific genotype assortment occurs within the tick vectors (6). In Portugal, previous studies show different prevalence rates for T. equi, which may be associated with the kind of horses/facility involved, the study area and the diagnostic tests used. In horse stud farms in the Ribatejo region, central Portugal, using Complement Fixation (CF), seroprevalence was 45.3%, while using blood smears, the infection was prevalent on 42,6% of the examined horses, with a potential transuterine transmission in 80% of positive mares (10, 11). Further South, in Alentejo region, using CF and IFAT the seroprevalence for Theileria equi was 85.1% and recently, using qPCR/nPCR, 56% of the horses examined from the Lisbon and Alentejo areas showed a prevalence of 56% (12, 13).
Members of the family Anaplasmataceae (e.g., Anaplasma, Neoehrlichia, and Ehrlichia) are gram-negative, intracellular bacteria infecting domestic and wild animals, but also humans. Anaplasma phagocytophilum has been documented in various animals including horses and is the causative agent of equine, canine, and human granulocytic anaplasmosis. Various strains are known and horses can harbor strains of zoonotic potential (14). In Europe, the main vector of this tick-borne pathogen is Ixodes ricinus, which is also present in mainland Portugal (7). Anaplasma phagocytophilum has previously been detected in I. ricinus on Madeira and in Ixodes ventalloi in mainland Portugal (Baixa de Palmela, Setubal district) (15, 16). Anaplasma phagocytophilum is regularly reported in serology-based studies on horses in Europe including Portugal [e.g., (8, 16, 17)].
Rickettsia species are obligate intracellular, gram-negative bacteria transmitted by various types of arthropods [e.g., ixodid ticks are the main vectors of spotted fever group rickettsiae (18)]. Several studies have documented Rickettsia spp. in horses.
Filarioid nematodes are also parasites of horses in Europe. Because of their asymptomatic to minor symptomatic effects, these parasites are neglected and understudied. Adults of Setaria equina are located in the peritoneal cavities of horses. Microfilariae of this worldwide distributed helminth can be found in the peripheral blood (19). Mosquitoes of the genera Culex and Aedes are the vectors of S. equina. Another filarioid species parasitizing medial layers or outside layers of tissues within the artery wall of horses is Onchocerca boehmi (syn. Elaeophora boehmi) (20, 21).
Little is known about the risk of military horses regarding VBDs. In fact, few studies have been conducted so far and no surveillance mechanisms are in place to assess geographical range and prevalence in the country. Considering the emergence of VBDs in Europe, as well as the lack of data on this topic, an epidemiological study was conducted, in order to identify the presence and prevalence of the most significant bacterial and parasitic VBDs of zoonotic and/or non-zoonotic relevance in Portuguese military horses using molecular analysis followed by sequence analysis of positive DNA products.
Materials and Methods
Overall 101 military horses (Puro Sangue Lusitano breed) belonging to the Portuguese National Republican Guard (GNR), stabled at guard facilities in metropolitan Lisbon were included in this study. Horses from various job sites were included (55 GNR Lisbon, 46 GNR Lisbon Braço de Prata Expo, 2 GNR Évora, 1 GNR Faro, and 1 GNR Tomar), and information on sex and age were determined. All horses were apparently healthy with no clinical signs compatible with VBDs. Horses were dewormed with moxidectin and praziquantel administered annually (with the exception of sportive horses, which were dewormed twice a year). Horses were not treated against tick infestation. Blood samples were collected at the jugular vein from apparently healthy horses in June (n = 55) and July (n = 46) in 2017.
A total of 50 μl of blood was spotted on Whatman® filter paper (VWR International GmBH, Vienna, Austria) and dried at room temperature. Afterward, each filter paper was sealed in an envelope separately. Filter papers were shipped to the University of Veterinary Medicine Vienna for molecular analysis. Sections of ~4 mm in diameter were cut out of the center of the blood spots with sterile blades. DNA was extracted with a modified chelex-based technique using InstaGene™ matrix (Bio-Rad Laboratories, Hercules, California) as established previously (22). Extracted DNA was stored at −20°C until further analysis. Samples were screened for the presence of DNA of various vector-borne pathogens of zoonotic and non-zoonotic relevance using specific broad-range PCR assays, under conditions reported previously [(23); Table 1] as follows: piroplasms (incl. Babesia and Theileria) within the 18S rRNA gene [BTH-1F/BTH-1R; (24)]; Anaplasmataceae (incl. Anaplasma, Ehrlichia, and Neoehrlichia) within the 16S gene [EHR16SD/EHR16SR; (26)]; Rickettsia spp. within the 23S/5S rRNA gene [ITS-F/ITS-R; (27)] and filiarioid helminths targeting a fragment of the mitochondrial cytochrome c oxidase subunit I gene [COI; H14FilaCOIFw/H14FilaCOIRv; (28)]. Additionally, all samples were further analyzed with a Babesia specific PCR analysis [Babfor/Babrev; (25)]. PCR products were analyzed by gel electrophoresis on 2% agarose gels stained with Midori Green Advance DNA stain (Nippon Genetics Europe, Düren, Germany). Positive reaction products were commercially purified and sequenced at LGC Genomics GmbH (Berlin, Germany).
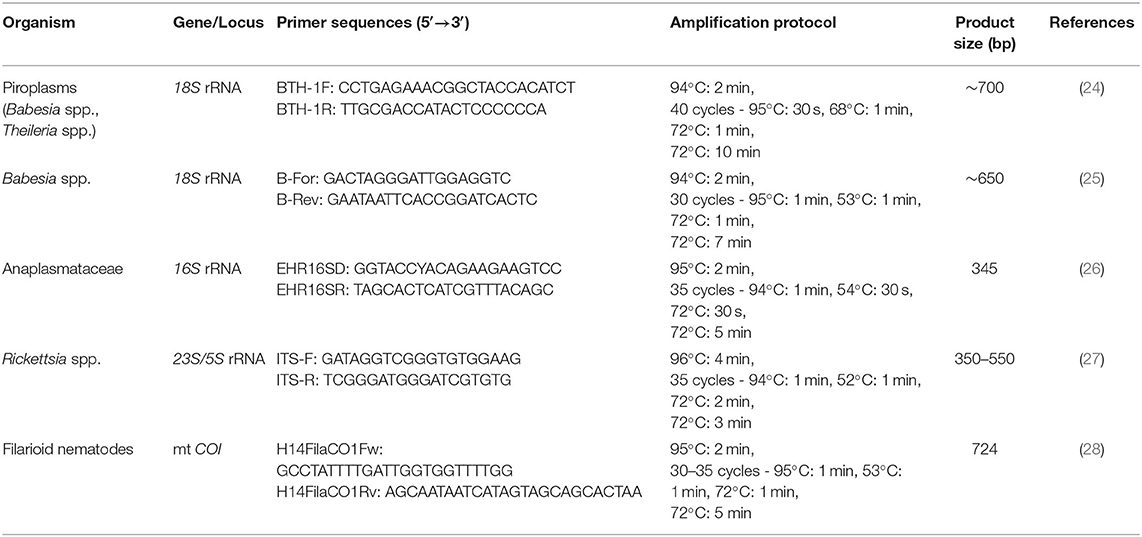
Table 1. Primer sequences and PCR protocols used for molecular analysis of vector-borne pathogens in military horses.
Phylogenetic Analysis of the T. equi 18S Sequences
To show the diversity of 18S lineages of T. equi, Bayesian inference (BI) and Maximum likelihood (ML) trees were calculated based on the sequences of the present study, and data was mined from NCBI GenBank. The GenBank sequences were retrieved by performing a BLAST search on a 530 bp 18S section of Theileria spp. To identify the lineages of T. equi, the sequences were aligned and sorted with MAFFT v.7 applying the default settings (29). The sequences were visually inspected using BioEdit (30) and all sequences were removed which did not cover the entire length, contained ambiguity characters, and/or obvious sequencing errors, resulting in a total of 220 T. equi sequences. The sequences of the present study and NCBI GenBank were combined and a sequence of Theileria ovis (AY533144) was added for outgroup comparison. All sequences were re-aligned with MAFFT v.7 applying the option “G-INS-i” and then collapsed to haplotypes using DAMBE (31). The alignment contained 49 unique T. equi lineages and 535 sites, of which all 15 gap positions were removed before the phylogenetic analysis. The best-fit substitution model for the 520 bp alignment was inferred using the model search implemented in the W-IQ-TREE web server [http://iqtree.cibiv.univie.ac.at/; (32)], resulting in the model HKY+G. A ML consensus tree was calculated from 1000 bootstrap replicates using W-IQ-TREE. A BI tree was calculated with MrBayes v.3.2.7 (33). The Bayesian analysis was run for 10 million generations (2 runs each with 4 chains, one of which was heated) and every thousandth tree was sampled. The first 25% of trees were discarded as burn-in and a 50% majority-rule consensus tree was calculated from the remaining 7,500 trees.
Results
In this study, 101 military horses were included of which 60 were males and 41 mares. The age ranged between 4 and 24 years (x = 12.4 years; xmed = 12 years). Overall 33 animals (32.7%; CI95: 24.3–42.3%) were infected with a vector-borne pathogen, namely Theileria equi. Seventeen males and 16 mares aged between 4 and 18 years (x = 10.5) gave positive signals at the molecular analysis for piroplasms. All of the positive horses came from Lisbon (23 at GNR Lisbon Braço de Prata Expo and 10 at GNR Lisbon). Two genotypes of T. equi were recorded of which 24 were genotype 1 and seven genotype 2. Both genotypes were present in GNR Lisbon Braço de Prata Expo and GNR Lisbon. The phylogenetic tree of the 18S sequences shows three main sequence groups or clades, one of which may be divided into two sub-groups (Figure 1). The T. equi lineages identified in the present study clustered in the first and second clade. Of the 33 T. equi positive samples, 24 featured a lineage (genotype 1, 100% match with HM229407), which clustered in the first clade. This lineage and similar ones were isolated from horses in China, Kazakhstan, Russia, Mongolia, Saudi Arabia, South Korea, Spain, Sudan, and Switzerland. Three slightly distinct lineages (KF597077, KF597078, and KF597081) in this clade were isolated from the waterbuck Kobus ellipsiprymnus in Kenya. Seven horses sampled in the present study featured a T. equi lineage, which clustered in the second clade (Figure 1). This lineage (genotype 2, 100% match with EU888906) and similar ones were isolated from horses in Brazil, Croatia, India, Jordan, Egypt, Israel, Saudi Arabia, South Africa, Spain, Turkey, and the USA. The phylogenetic tree (Figure 1) features a third clade, which can be subdivided into two subclades (3a and 3b). Subclade 3a contains lineages isolated from horses in Egypt, Israel, Palestine, Turkey, Nigeria, and Sudan. All records come from the Middle East and Northern and Eastern Africa. Two slightly deviating lineages (KF597074 and KF597076) were isolated from the waterbuck K. ellipsiprymnus in Kenya. Subclade 3b features lineages isolated from horses in Brazil, China, Cuba, Egypt, Iraq, Israel, South Africa, and the USA.
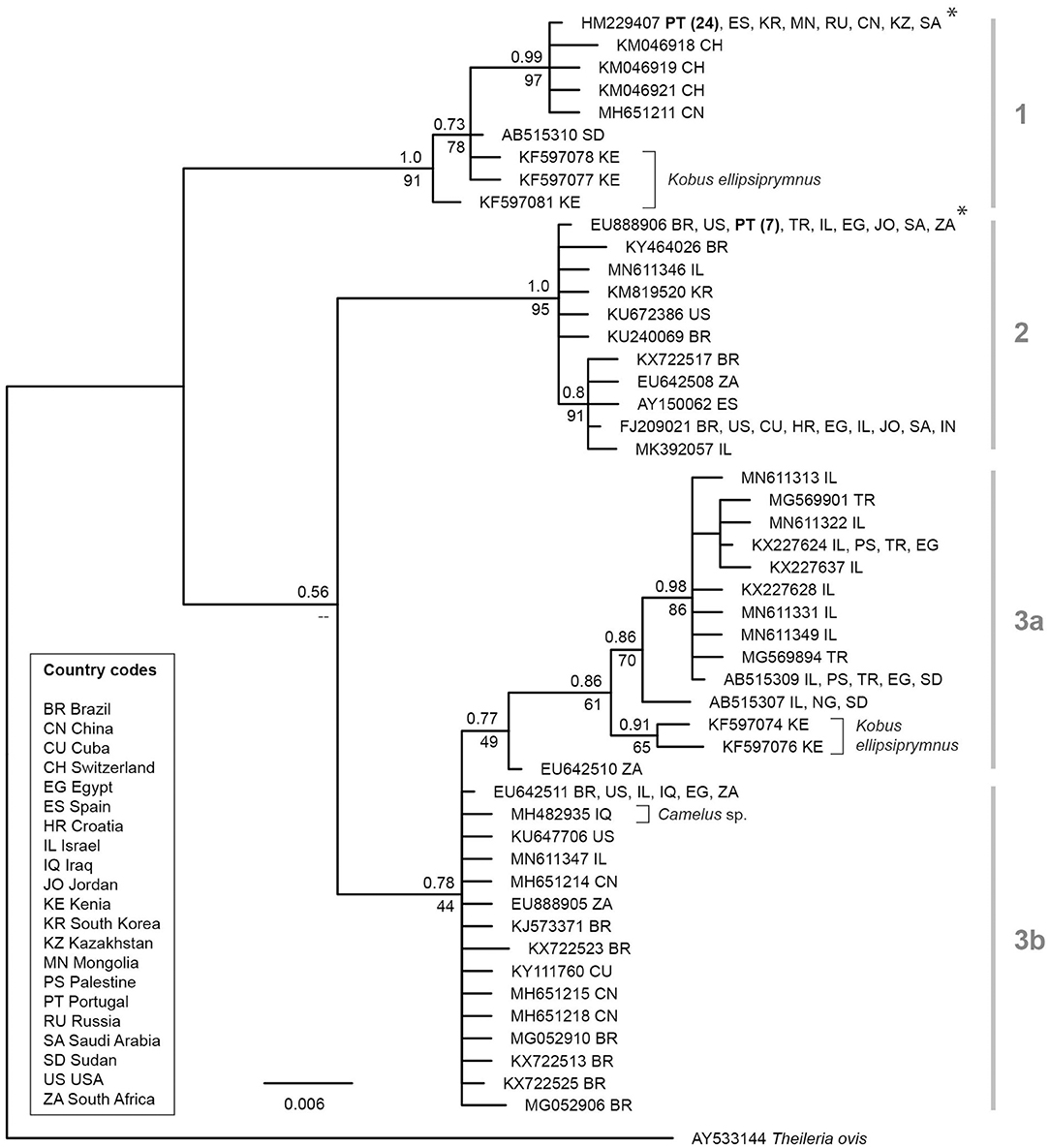
Figure 1. Bayesian inference tree of T. equi lineages based on 530 bp sections of the 18S. The BI posterior probabilities and ML bootstrap values are indicated at most nodes. The scale bar indicates the expected mean number of substitutions per site according to the model of sequence evolution applied. Asterisks mark the lineages isolated from horses in Portugal in the present study. As outgroup a sequence of Theileria ovis (AY533144) was included.
Analyses for the presence of DNA of zoonotic agents (e.g., Anaplasma spp. and Rickettsia spp.) but also other non-zoonotic pathogens (e.g., Babesia caballi and filarioid helminths like Setaria equina and Onchocerca boehmi) gave negative results.
Discussion
Prevalence of equine piroplasmosis vary in endemic regions in Europe for T. equi and B. caballi, depending on study design and diagnostic techniques used [IFAT, ELISA, PCR; (reviewed in 2)]. The prevalence of T. equi is generally higher than that for B. caballi. In healthy military horses included in this study, 32.7% (33/101) tested positive for the presence of DNA of T. equi. These data confirm a relatively high prevalence of T. equi in military horses in Portugal (mainly from the Lisbon area). In the Alentejo region (Southern Portugal) serological analyses with IFAT of 154 horses resulted in 85.1% positive for T. equi and 65.6% for B. caballi (12). Out of 73 “Puro Sangue Lusitano” horses from Vila Viçosa (Alentejo region, Southern Portugal) 53.4% were positive for T. equi using cELISA (34). Ribeiro et al. (8), combining serology and microscopy, reported T. equi in 19.1% and B. caballi in 11.7% of 162 horses in North of Portugal. Moreover, using PCR techniques, T. equi was detected in 56% of horses from central Southern Portugal that presented with clinical or subclinical signs (13). DNA of T. equi was found in two out of nine Rhipicephalus bursa male ticks collected from horses in the Comunidade Intermunicipal do-Ave (Northern Portugal), but because of the proof of DNA only, the vector capacity of this tick species remains unclear (35).
In the present study, horses between 4 and 18 years were infected with this parasite. It is known that the prevalence of equine piroplasmosis increases with age and that equids infected with T. equi remain life-long carriers of this parasite (2). Moreover, it is discussed that male and female horses have different susceptibilities to infection, but the results of various studies are contradictory. Host activity also increases the chance for horses to become infected (2). Grazing was documented to double the risk of becoming infected with B. caballi and T. equi (36). Outdoor or mixed indoor/outdoor type of housing was also reported as a risk factor for T. equi (8). Military horses included in this study are regularly outside, but are also stabled at military facilities. However, regular travel activity to various regions in the country increases the possibility of tick contact. Furthermore, the type of horse breed may also influence the risk of T. equi infections (37).
Interestingly we were not able to detect DNA of B. caballi in military horses from this study, although an additional Babesia-specific PCR was run. Previous studies have shown that horses seropositive for B. caballi (but also T. equi) are often not positive at PCR analysis (38). However, it is recommended to run cELISA, IFAT, and PCR to assure the identification of acute and chronic equine piroplasm infections (39). Recent research carried out on Lisbon horses, found a 7% prevalence for Babesia spp., meaning it may be a residual, but persistent agent, in this area (13).
Anaplasmataceae were not detected in the blood samples of healthy horses analyzed in this study. Using serology, A. phagocytophilum has previously been confirmed in nine of 302 horses from mainland Portugal (16) and 13% of 162 horses in Northern Portugal (8). However, it has been shown that seropositivity to A. phagocytophilum was significantly higher if compared to PCR results [e.g., (40)]. Therefore, we cannot exclude that horses in this study had previously been in contact with this pathogen. It is discussed that climate change might increase the threat of disease in European horse populations (41).
Although no horses were positive for Rickettsia spp. in this study, several Rickettsia species have been documented in horses in Europe. Spotted fever-group rickettsiae were reported in 15.03% of 479 horses in Italy using serology (17). Using IFAT, Rickettsia helvetica was documented in 36.5% of 63 horses in Sweden (42). In another study, R. helvetica and R. monacensis were confirmed in ticks collected from ponies in Poland using PCR techniques (43). On Corsica, the presence of Rickettsia slovaca and Rickettsia aeschlimannii in ticks collected on horses was confirmed (44).
DNA of microfilaria of filarioid helminths was not detected in the present study. However, although infections with these parasites are mainly asymptomatic and neglected, they have been reported in horses in several regions in Europe. In Hungary, microfilariae of S. equina were reported in 9.2% (18/195) of horses analyzed, and infection was associated with the presence of still waters nearby (45).
Although only one parasite species was documented, it can be concluded that T. equi is among the most important vector-borne transmitted agents infecting horses in Europe. As equine piroplasmosis is a severe infectious tick-borne disease responsible for significant losses in equine reproduction, and with numerous impacts on the international movement of horses, treatment and preventive measures are needed to reduce exposure to future infections.
Data Availability Statement
The 18S sequences of Theileria equi were deposited in NCBI GenBank under the accession numbers MT767139-MT767169.
Ethics Statement
This animal study was reviewed and approved by GNR–5373/17/CDF/GAB. Whenever possible, blood sampling was performed during normal sanitary surveys for the GNR horse population. Written informed consent was obtained from the owners for the participation of their animals in this study.
Author Contributions
H-PF, AA, and LM contributed to the conception and design of the study. AA, HRos, JT, and HRoc conducted field and laboratory work. FK and BS conducted the lab work. H-PF and JH analyzed the sequences and performed the phylogenetic analysis. H-PF, AA, JH, and LM wrote and reviewed the manuscript. All authors read and approved the final manuscript.
Funding
This project was funded by UID/CVT/00276/2020 (CIISA). H-PF was supported with a Short Term Scientific Mission (STSM)—COST Action TD1303.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer CG declared a past co-authorship with one of the author H-PF to the handling editor.
Acknowledgments
This study was performed within the framework of the COST action TD1303, EurNegVec.
References
1. Brown C, Torres A. Foreign Animal Diseases. 7th ed. Boca Raton, FL: Boca Publications (2008) p. 472.
2. Onyiche TE, Suganuma K, Igarashi I, Yokoyama N, Xuan X, Thekisoe O. A review on equine piroplasmosis: epidemiology, vector ecology, risk factors, host immunity, diagnosis and control. Int J Environ Res Public Health. (2019) 16:E1736. doi: 10.3390/ijerph16101736
3. Tirosh-Levy S, Gottlieb Y, Mimoun L, Mazuz ML, Steinman A. Transplacental transmission of Theileria equi is not a common cause of abortions and infection of foals in Israel. Animals. (2020) 10:E341. doi: 10.3390/ani10020341
4. Wise LN, Kappmeyer LS, Mealey RH, Knowles DP. Review of equine piroplasmosis. J Vet Intern Med. (2013) 27:1334–46. doi: 10.1111/jvim.12168
5. Scoles GA, Ueti MW. Vector ecology of equine piroplasmosis. Annu Rev Entomol. (2015) 60:561–80. doi: 10.1146/annurev-ento-010814-021110
6. Tirosh-Levy S, Steinman A, Einhorn A, Apanaskevich DA, Mumcuoglu KY, Gottlieb Y. Potential tick vectors for Theileria equi in Israel. Med Vet Entomol. (2020) 34:291–4. doi: 10.1111/mve.12435
8. Ribeiro AJ, Cardoso L, Maia JM, Coutinho T, Cotovio M. Prevalence of Theileria equi, Babesia caballi, and Anaplasma phagocytophilum in horses from the north of Portugal. Parasitol Res. (2013) 112:2611–17. doi: 10.1007/s00436-013-3429-9
9. Qablan MA, Oborník M, PetrŽelková KJ, Sloboda M, Shudiefat MF, Horín P, et al. Infections by Babesia caballi and Theileria equi in Jordanian equids: epidemiology and genetic diversity. Parasitology. (2013) 140:1096–103. doi: 10.1017/S0031182013000486
10. Carvalho-Varela M, Pereira da Fonseca IM, de Carvalho LMM, Sabino-Serra JM, Castelo-Branco A, Afonso-Roque MM, et al. Epidemiogical aspects of horse parasitic diseases in the Ribatejo region (Portugal). In: VIII International Congress of Parasitology (Izmir) (1994). p. 321.
11. Rego BMCD. Study of the natural infection by protozoa of the genus Babesia and Theileria in an equine stud-farm from Ribatejo (MSc dissertation). Integrated Master in Veterinary Medicine, Faculty of Veterinary Medicine, Technical University of Lisbon, Lisbon, Portugal (2008). p. 69.
12. Malta MJPV. Diagnosis horse babesiosis and theileriosis in alentejo (MSc dissertation) Master in Tropical Veterinary Medicine and Animal Production, Faculty of Veterinary Medicine, Technical University of Lisbon, Lisbon, Portugal (2001). p. 89.
13. Barros CJG. Equine piroplasmosis: molecular diagnosis and evaluation of hematological and inflammatory biomarkers changes in horses with clinical and subclinical disease (MSc Dissertation) Integrated Master in Veterinary Medicine, Faculty of Veterinary Medicine, University of Lisbon p. 71.
14. Matei IA, Estrada-Peña A, Cutler SJ, Vayssier-Taussat M, Varela-Castro L, Potkonjak A, et al. A review on the eco-epidemiology and clinical management of human granulocytic anaplasmosis and its agent in Europe. Parasit Vectors. (2019) 12:599. doi: 10.1186/s13071-019-3852-6
15. Santos AS, Santos-Silva MM, Almeida VC, Bacellar F, Dumler JS. Detection of Anaplasma phagocytophilum DNA in Ixodes ticks (Acari: Ixodidae) from Madeira Island and Setubal District, mainland Portugal. Emerg Infect Dis. (2004) 10:1643–8. doi: 10.3201/eid1009.040276
16. Santos AS, Bacellar F, Dumler JS. A 4-year study of Anaplasma phagocytophilum in Portugal. Clin Microbiol Infect. (2009) 15(Suppl. 2):46–7. doi: 10.1111/j.1469-0691.2008.02172.x
17. Ebani VV. Serological evidence of Anaplasma phagocytophilum and spotted fever group Rickettsia spp. exposure in horses from Central Italy. Pathogens. (2019) 8:E88. doi: 10.3390/pathogens8030088
18. Parola P, Paddock CD, Socolovschi C, Labruna MB, Mediannikov O, Kernif T, et al. Update on tick-borne rickettsioses around the world: a geographic approach. Clin Microbiol Rev. (2013) 26:657–702. doi: 10.1128/CMR.00032-13
19. Coleman SU, Klei TR, French DD. Prevalence of Setaria equina (Nematode: Onchocercidae) in southeastern Louisiana horses. J Parasitol. (1985) 71:512–3. doi: 10.2307/3281548
20. Supperer R. Filarosen der Pferde in Österreich. Wiener Tierärztliche Monatsschrift. (1953) 40:193–220.
21. Lia RP, Mutafchiev Y, Veneziano V, Giannelli A, Abramo F, Santoro M, et al. Filarial infection caused by Onchocerca boehmi (Supperer, 1953) in a horse from Italy. Parasitol Res. (2017) 116:191–8. doi: 10.1007/s00436-016-5277-x
22. Fuehrer HP, Starzengruber P, Swoboda P, Khan WA, Matt J, Ley B, et al. Indigenous Plasmodium ovale malaria in Bangladesh. Am J Trop Med Hyg. (2010) 83:75–8. doi: 10.4269/ajtmh.2010.09-0796
23. Cézanne R, Mrowietz N, Eigner B, Duscher GG, Glawischnig W, Fuehrer HP. Molecular analysis of Anaplasma phagocytophilum and Babesia divergens in red deer (Cervus elaphus) in Western Austria. Mol Cell Probes. (2017) 31:55–8. doi: 10.1016/j.mcp.2016.07.003
24. Zintl A, Finnerty EJ, Murphy TM, de Waal T, Gray JS. Babesias of red deer (Cervus elaphus) in Ireland. Vet Res. (2011) 42:7. doi: 10.1186/1297-9716-42-7
25. Blaschitz M, Narodoslavsky-Gföller M, Kanzler M, Stanek G, Walochnik J. Babesia species occurring in Austrian Ixodes ricinus ticks. Appl Environ Microbiol. (2008) 74:4841-6. doi: 10.1128/AEM.00035-08
26. Brown GK, Martin AR, Roberts TK, Aitken RJ. Detection of Ehrlichia platys in dogs in Australia. Aust Vet J. (2001) 79:554–8. doi: 10.1111/j.1751-0813.2001.tb10747.x
27. Vitorino L, Zé-Zé L, Sousa A, Bacellar F, Tenreiro R. rRNA intergenic spacer regions for phylogenetic analysis of Rickettsia species. Ann N Y Acad Sci. (2003) 990:726–33. doi: 10.1111/j.1749-6632.2003.tb07451.x
28. HodŽić A, Alić A, Fuehrer HP, Harl J, Wille-Piazzai W, Duscher GG. A molecular survey of vector-borne pathogens in red foxes (Vulpes vulpes) from Bosnia and Herzegovina. Parasit Vectors. (2015) 8:88. doi: 10.1186/s13071-015-0692-x
29. Katoh K, Standley DM. MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol Biol Evol. (2013) 30:772–80. doi: 10.1093/molbev/mst010
30. Hall TA. BioEdit: a user-friendly biological sequences alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp Ser. (1999) 41:95–8.
31. Xia X, Xie Z. DAMBE: software package for data analysis in molecular biology and evolution. J Heredity. (2001) 92:371–3. doi: 10.1093/jhered/92.4.371
32. Trifinopoulos J, Nguyen LT, von Haeseler A, Minh BQ. W-IQ-TREE: a fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. (2016) 44:W232–5. doi: 10.1093/nar/gkw256
33. Ronquist F, Teslenko M, Van Der Mark P, Ayres DL, Darling A, Höhna S, et al. MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Syst Biol. (2012) 61:539–42. doi: 10.1093/sysbio/sys029
34. Sousa S, Almeida A, Simão L, Anastácio S, Madeira de Carvalho LM. Theleria equi infection in Puro Sangue Lusitano horses from Vila Viçosa. In: Proceedings of the II Iberian Congress of Veterinary Epidemiology (Barcelona) (2010). p. 19.
35. Ferrolho J, Antunes S, Santos AS, Velez R, Padre L, Cabezas-Cruz A, et al. Detection and phylogenetic characterization of Theileria spp. and Anaplasma marginale in Rhipicephalus bursa in Portugal. Ticks Tick Borne Dis. (2016) 7:443–8. doi: 10.1016/j.ttbdis.2016.01.004
36. Moretti A, Mangili V, Salvatori R, Maresca C, Scoccia E, Torina A, et al. Prevalence and diagnosis of Babesia and Theileria infections in horses in Italy: a preliminary study. Vet J. (2010) 184:346–50. doi: 10.1016/j.tvjl.2009.03.021
37. Montes Cortés MG, Fernández-García JL, Habela Martínez-Estéllez MÁ. Seroprevalence of Theileria equi and Babesia caballi in horses in Spain. Parasite. (2017) 24:14. doi: 10.1051/parasite/2017015
38. Grandi G, Molinari G, Tittarelli M, Sassera D, Kramer LH. Prevalence of Theileria equi and Babesia caballi infection in horses from northern Italy. Vector Borne Zoonotic Dis. (2011) 11:955–956. doi: 10.1089/vbz.2010.0193
39. Mahmoud MS, El-Ezz NT, Abdel-Shafy S, Nassar SA, El Namaky AH, Khalil WK, et al. Assessment of Theileria equi and Babesia caballi infections in equine populations in Egypt by molecular, serological and hematological approaches. Parasit Vectors. (2016) 9:260. doi: 10.1186/s13071-016-1539-9
40. Passamonti F, Veronesi F, Cappelli K, Capomaccio S, Coppola G, Marenzoni ML, et al. Anaplasma phagocytophilum in horses and ticks: a preliminary survey of Central Italy. Comp Immunol Microbiol Infect Dis. (2010) 33:73–83. doi: 10.1016/j.cimid.2008.08.002
41. Dziegiel B, Adaszek L, Winiarczyk M, García-Bocanegra I, Carbonero A, Debiak P, et al. Comparative analysis of 16S RNA nucleotide sequences of Anaplasma phagocytophilum detected in the blood of horses from various parts of Europe. J Med Microbiol. (2013) 62:1891–1896. doi: 10.1099/jmm.0.058636-0
42. Elfving K, Malmsten J, Dalin AM, Nilsson K. Serologic and molecular prevalence of Rickettsia helvetica and Anaplasma phagocytophilum in wild cervids and domestic mammals in the central parts of Sweden. Vector Borne Zoonotic Dis. (2015) 15:529–34. doi: 10.1089/vbz.2015.1768
43. Skotarczak B, Wodecka B, Rymaszewska A, Adamska M. Molecular evidence for bacterial pathogens in Ixodes ricinus ticks infesting Shetland ponies. Exp Appl Acarol. (2016) 69:179–89. doi: 10.1007/s10493-016-0027-4
44. Grech-Angelini S, Stachurski F, Vayssier-Taussat M, Devillers E, Casabianca F, Lancelot R, et al. Tick-borne pathogens in ticks (Acari: Ixodidae) collected from various domestic and wild hosts in Corsica (France), a Mediterranean island environment. Transbound Emerg Dis. (2020) 67:745–57. doi: 10.1111/tbed.13393
Keywords: equine piroplasmosis, military horses, Theileria equi, vector-borne diseases, zoonosis, Portugal
Citation: Fuehrer H-P, Alho AM, Kayikci FN, Shahi Barogh B, Rosa H, Tomás J, Rocha H, Harl J and Madeira de Carvalho L (2020) Survey of Zoonotic and Non-zoonotic Vector-Borne Pathogens in Military Horses in Lisbon, Portugal. Front. Vet. Sci. 7:591943. doi: 10.3389/fvets.2020.591943
Received: 05 August 2020; Accepted: 04 September 2020;
Published: 15 October 2020.
Edited by:
Rodrigo Morchón García, University of Salamanca, SpainReviewed by:
Claudio Genchi, University of Milan, ItalyJacob Lorenzo-Morales, University of La Laguna, Spain
Copyright © 2020 Fuehrer, Alho, Kayikci, Shahi Barogh, Rosa, Tomás, Rocha, Harl and Madeira de Carvalho. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Hans-Peter Fuehrer, aGFucy1wZXRlci5mdWVocmVyQHZldG1lZHVuaS5hYy5hdA==; Luís Madeira de Carvalho, bWFkZWlyYWRlY2FydmFsaG9AZm12LnVsaXNib2EucHQ=