
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
BRIEF RESEARCH REPORT article
Front. Vet. Sci. , 19 November 2020
Sec. Veterinary Epidemiology and Economics
Volume 7 - 2020 | https://doi.org/10.3389/fvets.2020.588919
This article is part of the Research Topic Antimicrobial Resistance in Zoonotic Bacteria in Developing Countries: The Role of Food Animal Production in Public Health View all 11 articles
 Nadia Coppola1
Nadia Coppola1 Bibiana Freire2
Bibiana Freire2 Ana Umpiérrez3
Ana Umpiérrez3 Nicolás F. Cordeiro1Pablo Ávila1Gustavo Trenchi4Gustavo Castro2María Laura Casaux5
Nicolás F. Cordeiro1Pablo Ávila1Gustavo Trenchi4Gustavo Castro2María Laura Casaux5 Martín Fraga5
Martín Fraga5 Pablo Zunino3
Pablo Zunino3 Inés Bado1*
Inés Bado1* Rafael Vignoli1*
Rafael Vignoli1*The aim of this work was to detect Escherichia coli isolates displaying resistance to oxyimino-cephalosporins, quinolones, and colistin in feces from livestock in Uruguay. During 2016–2019, fecal samples from 132 broiler and layer chicken flocks, 100 calves, and 50 pigs, were studied in Uruguay. Samples were cultured on MacConkey Agar plates supplemented with ciprofloxacin, ceftriaxone, or colistin. E. coli isolates were identified by mass spectrometry and antibiotic susceptibility testing was performed by disk diffusion agar method and colistin agar test. Antibiotic resistance genes were detected by polymerase chain reaction and sequencing. The most frequently detected resistance gene was qnrB19, recovered from 87 animals. Regarding plasmid-mediated quinolone resistance genes, qnrS1 was the second in prevalence (23 animals) followed by qnrE1, found in 6 chickens and two calves. Regarding resistance to oxyimino-cephalosporins, 8 different β-lactamase genes were detected: blaCTX−M−8 and blaCMY−2 were found in 23 and 19 animals, respectively; next, blaCTX−M−2 and blaSHV−12 in 7 animals each, followed by blaCTX−M−14 in 5, blaCTX−M−15 and blaSHV2a in 2, and blaCTX−M−55 in a single animal. Finally, the mcr-1 gene was detected only in 8 pigs from a single farm, and in a chicken. Isolates carrying blaCMY−2 and blaSHV−12 were also found in these animals, including two isolates featuring the blaCMY−2/mcr-1 genotype. To the best of our knowledge, this is the first work in which the search for transferable resistance to highest priority critically important antibiotics for human health is carried out in chickens and pigs chains of production animals in Uruguay.
The interaction between humans and animals is quite diverse and may lead to cases of zoonosis and/or anthropozoonosis (1). Over the past decades the aforementioned interaction has constantly increased worldwide partly due to animal husbandry practices, the growth of the companion animal market, climate change, and ecosystem disruption. In this context, bacterial transmission may occur through food products (e.g., meat or eggs) or through direct contact, in particular in farmers, veterinarians, or abattoir workers (2).
As the human-animal connection escalates, so does the threat of pathogen spread (2, 3). With current rapid transport systems, a pathogen emerging today in any given country can easily be carried unnoticed in people, animals, plants, or food products, to distant parts of the world in <24 h (4).
On the other hand, anthropogenic changes to the ecosystem increase the number of shared habitats between humans and animals, thus exposing both to new pathogens. In this regard, several authors have described the occurrence in humans and several animal species of the pandemic strain, Escherichia coli 025:H4 ST131, carrying the extended-spectrum beta-lactamase CTX-M-15. This particular event indicates an interspecies transmission from humans to pets and livestock and has been particularly described across Europe (5).
Recently, the WHO, FAO, and OIE organizations have coined the term “One Health” which regards the environment and human and animal health as a single entity. In this context, antibiotic resistance is a major concern (6).
The highest consumption of antibiotics occurs in animal husbandry, reaching in several countries 80% of the annual total antimicrobial consumption, with a higher consumption estimated by the year 2030. In this sense, the food industry may be accountable for the spread and increase of antibiotic resistance mechanisms (6, 7).
The impact on humans of antibiotic-resistant bacteria of animal origin is reflected in the increase of enteropathogens such as Salmonella spp. resistant to oxyimino-cephalosporins and fluoroquinolones, responsible among others for severe pediatric infections. Moreover, resistance to azithromycin and ciprofloxacin has been noted in species such as Campylobacter jejuni, and ESBL-producing Escherichia coli (8). Particularly in the United States, enterobacteria are responsible for 140,000 healthcare-associated infections annually, with 26,000 infections attributable to ESBL-producing enterobacteria, representing 19% of hospital-acquired infections.
In this respect, foodborne infections due to these resistant enteropathogens are a risk to humans due to possible therapeutic failure (8). Evidence indicates that a reduction in antibiotic consumption in animal husbandry would lead to a decrease in bacterial antibiotic resistance levels (9).
Antibiotics, in animal husbandry, are mainly used to treat infections, furthermore, they are also used as a prophylactic, through group treatment by incorporating them in drug premixes at relatively high concentrations, and more worryingly as growth promoters (although this practice has already been banned in some countries) (8).
Particularly noteworthy is the emergence of transferable resistance to critical antibiotics of highest priority for human medicine such as oxyimino-cephalosporins, fluoroquinolones and polymyxins, due to: extended spectrum β-lactamases (ESBLs) and plasmidic cephalosporinases (pAmpC); plasmid-mediated quinolone resistance (PMQR) genes [e.g., qnr and aac(6′)-Ib-cr]; and mcr alleles, respectively (10).
Infections caused by multidrug resistant microorganisms (MRM) lead to longer hospital stays (6.4–12.7 days), increased morbimortality (6.5%) and elevated economic costs (18,588–29,069 US$ per patient) (11, 12). Additionally, the occurrence of MRM in production animals may result in economic losses in trade and agriculture commerce; an example of the latter was observed in Norway, where the presence of resistant E. coli in retail chicken meat resulted in a 20% decrease in sales (13).
In Uruguay, measures have been taken to restrict the use of antibiotics, both in human health and in animal husbandry. The use of antibiotics as growth promoters in the latter is forbidden, and cattle under treatment are not destined for human consumption (https://www.proa.hc.edu.uy/), (Decree N° 98/011). In addition, in March 2019 Decree 141/019 was established, prohibiting the import, export, manufacture, sale, use, possession and marketing of veterinary products containing the substance “colistin” in its composition, either alone or associated with other chemicals (either as raw material or finished product), or incorporated into animal feed.
Finally, in 2017 Umpierrez et al. reported for the first time the occurrence of antibiotic- resistance genes in E. coli isolated from cattle in Uruguay. In that study, the authors detected multidrug-resistant E. coli isolated from calves carrying blaCTX−M−14 and PMQR genes, among other resistance determinants (14).
The aim of this work was to detect E. coli isolates displaying resistance to oxyimino-cephalosporins, quinolones, and colistin in feces from livestock in Uruguay.
During 2016–2019, fecal samples from 282 animals were studied in Uruguay: 100 from calves, 132 from broiler and layer chicken flocks and 50 from pigs of different ages; samples were obtained from five, 13, and five establishments, respectively. Between 5 and 20 animals were sampled in each establishment (see Table 1).
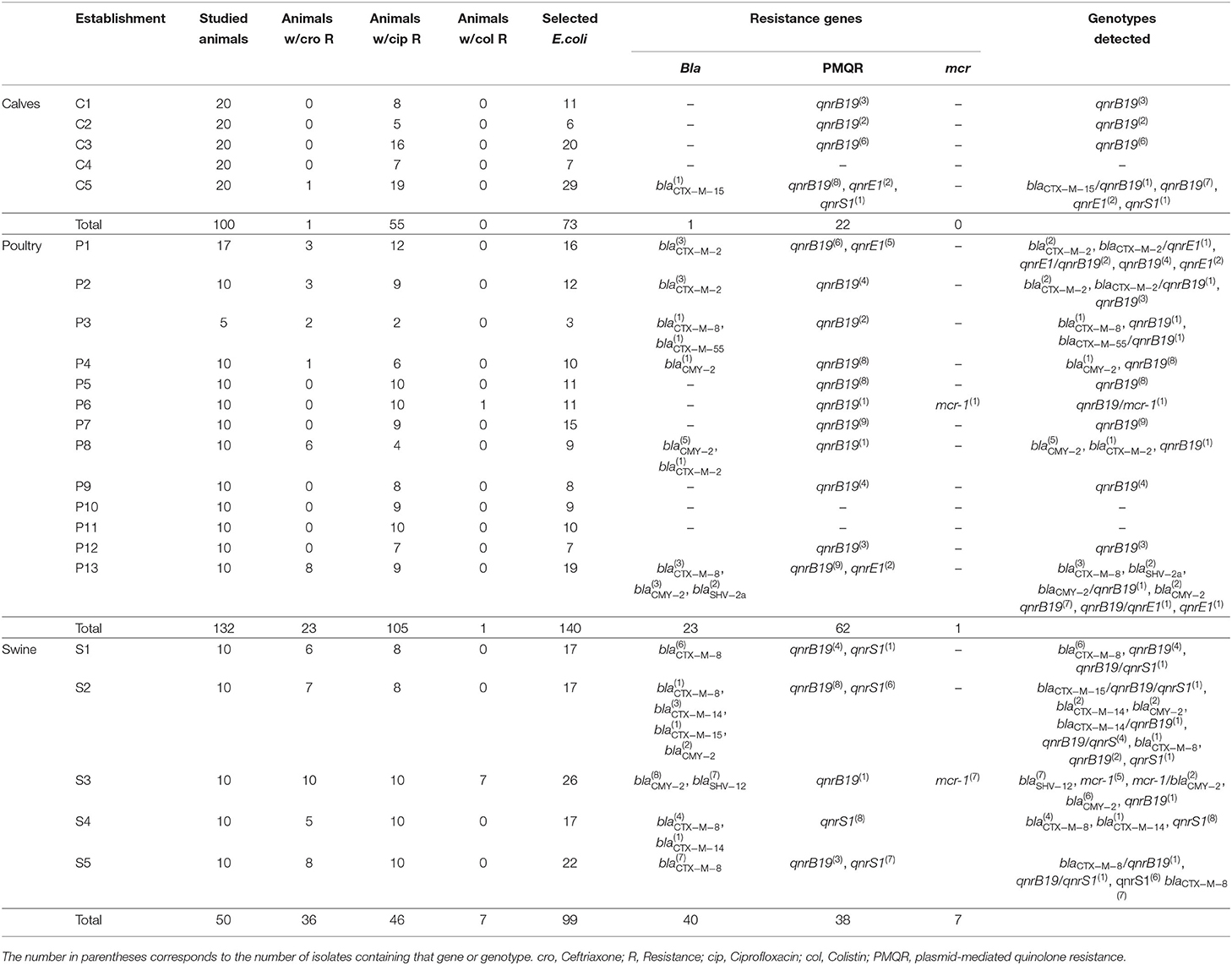
Table 1. Main results from the studied establishments.
Pig and bovine feces were collected wearing latex gloves directly from animals; conversely, chicken samples were taken directly from cloacae with sterile swabs. All samples were refrigerated at 4°C and sent, within 24 h, to Departamento de Bacteriología y Virología (Instituto de Higiene, Montevideo, Uruguay), or to Plataforma de Investigación en Salud Animal for processing (Colonia, Uruguay).
Samples were pre-enriched in Luria Bertani broth for 12 h at 37°C. Next, 10 μl of the broth were cultured on MacConkey Agar plates (Oxoid Ltd., Basingstoke, UK) supplemented with 0.125 mg/L ciprofloxacin (ION, Montevideo, Uruguay) or 1 mg/L ceftriaxone (Libra, Montevideo, Uruguay), or 3 mg/L colistin (Sigma-Aldrich St. Louis MO USA).
Putative E. coli colonies were identified by matrix assisted laser desorption ionization–time of flight (MALDI-TOF) mass spectrometry (Bruker, MA) in the facilities of Instituto de Higiene (Montevideo, Uruguay).
We then tested susceptibility to the following antibiotics: amoxicillin-clavulanic acid (AMC), ceftriaxone (CRO), ceftazidime (CAZ), ciprofloxacin (CIP), gentamicin (CN), amikacin (AK), and trimethoprim-sulfamethoxazole (SXT). Results were interpreted according to the Clinical Standard Laboratory Institute 2020 guidelines (15); isolates displaying intermediate resistance levels were considered as resistant.
Isolates showing resistance to 3rd generation cephalosporins underwent phenotypic screening for ESBLs and/or AmpC β-lactamases with the synergy double-disc method, using ESBL and AmpC inhibitors (AMC and boronic acid, respectively) plus 3rd generation cephalosporins according to Cordeiro et al. (16).
Minimum inhibitory concentration to colistin was determined for E. coli isolates recovered from MacConkey supplemented with the aforementioned antibiotic, by colistin agar test according to CLSI (15); E. coli ATCC 25922 was used as a quality control.
Resistance genes were sought by polymerase chain reaction (PCR) and confirmed by sequencing. We searched for the following resistance genes:
(a) oxyimino-cephalosporins
blaCTX−M−group−1, blaCTX−M−group−2, blaCTX−M−group−3, blaCTX−M−group−4, blaCTX−M−group−25, blaCTX−M−8, blaCTX−M−9, blaTEM, blaSHV, blaPER−2, blaAmpC (17);
(b) quinolones
aac (15)Ib-cr, qnrA, B, C, D, E, S, Vc, qepA (17);
(c) colistin
Detection of plasmid-mediated colistin resistance genes (mcr-1, mcr-2, and mcr-3) was performed by Real-Time PCR according to Li et al. (18) and mcr 4 was sought by PCR according to Carattoli et al. (19).
The detailed list of primers used can be found in Supplementary Table 1.
During the present work, 687 E. coli isolates were recovered: 334 from calves, 200 from poultry and 153 from pigs.
The 687 isolates were subjected to antibiotic susceptibility determination and detection of antibiotic resistance genes, as described in the materials and methods section. However, to avoid duplicate results, for each animal studied, only those isolates that presented phenotypic or genotypic differences are presented. Accordingly, we selected 73 bovine isolates, 141 poultry isolates and 99 pig isolates (Supplementary Table 2).
The most frequently detected resistance was to fluoroquinolones, being present in all of the analyzed farms. In 214/282 (75.9%) of the studied animals, we detected ciprofloxacin-resistant E. coli isolates (observed in 55 calves, 110 chickens, and 49 pigs) (Table 1). The farms displaying the highest levels of resistance corresponded to those of pig farming, yielding resistant isolates in 98% of the studied animals (49/50), followed by poultry farming with 83.3% (110/132), and lastly calf stables with 55% of the studied animals.
Conversely, resistance to oxyimino-cephalosporins was detected in 60/282 animals (21.4%), albeit quite heterogeneously: 72% of the sampled swine yielded resistant isolates (36/50) distributed along all the analyzed farms, followed by poultry farms where 17.4% of the sampled animals harbored resistant isolates (23/132), in 6/13 studied farms. Finally, a single calf harboring resistant isolates was detected among the studied establishments.
Resistance to colistin was the lowest of the three tested antibiotics, only 8 animals carried resistant isolates (2.8%); seven corresponded to pigs and the remaining case to a chicken.
The most frequently detected resistance gene was qnrB19, which was present in E. coli isolates recovered from 87 animals belonging to the 3 production lines. Regarding PMQR genes, qnrS1 was the second most frequent, being detected in 23 animals. Sparing a single case corresponding to a calf, the remaining 22 cases corresponded to isolates recovered from swine. The third gene in frequency was qnrE1, found in 8 isolates obtained from chickens and calves.
Regarding resistance to oxyimino-cephalosporins, 8 different β-lactamase genes were detected, the most frequent being blaCTX−M−8 and blaCMY−2 found in 22 and 19 animals, respectively; next, blaCTX−M−2 and blaSHV−12 were both detected in 7 animals, followed by blaCTX−M−14 in 4, blaCTX−M−15 and blaSHV2a in 2, and blaCTX−M−55 in a single animal (Table 1).
Finally, transferable colistin resistance genes were detected in 2 establishments, belonging to a pig and a poultry farm; in the former, mcr-1 was found in 8 animals, whereas in the latter in a single animal. In 3 pig-derived isolates, mcr-1 was found along with blaCMY−2, while in the poultry isolate mcr-1 was detected alongside qnrB19.
As previously mentioned, in the present work we selected 73 E. coli isolates obtained from 100 calves, 141 isolates obtained from 132 chickens, and 99 isolates obtained from 50 pigs, thus yielding genotype/studied animal ratios of 0.73 (73/100), 1.08 (140/132), and 1.77 (99/50) for calves, poultry, and swine, respectively. Additionally, 30% of the E. coli resistant genotypes recovered from calves were linked to transferable resistance genes (22/73); conversely, in poultry, and swine, the frequency of isolates harboring transferable resistance genes was 55% (78/140) and 76% (75/99), respectively (Table 1).
This is the first work tackling the presence of transferable resistance to antibiotics considered as highest priority critically important antibiotics for human health in three chains of production animals in Uruguay.
The most alarming situation was observed in swine husbandry, where the highest percentage of animals harboring resistant isolates, the highest number of bacterial genotypes per animal and the highest percentage of transferable resistance genes were observed. Taking into account this scenario it is possible to hypothesize that, from the three populations studied, the swine digestive tract is where the best condition for resistance genes horizontal transfer events occur.
Recently, Van Boeckel et al. have reported that the global consumption of antibiotics per kilogram of animal in cattle, poultry, and swine husbandry is 45, 148, and 172 mg, respectively (7). Our data show some degree of correlation with that study, in the sense that a greater use of antibiotics entails a higher detection of resistance mechanisms on account of the selective pressure imposed.
Nevertheless, resistance to colistin (at least in this study) is lower than values reported in other countries of our region and the rest of the world, such as Ecuador, Argentina, and Spain (20–22). During this work we detected the presence of 8/50 pigs (16%) carrying mcr-1-harboring E. coli, all of them corresponding to the same farm. In 5/7 animals, the genes mcr-1 and blaCMY−2 were found in separate E. coli isolates, yet in 3 animals those genes were found within the same isolate (Supplementary Table 2). Recently, we reported the detection of the first E. coli isolates of human origin carrying mcr-1 in Uruguay. One of these isolates harbored blaCMY−2 and mcr-1, albeit in different plasmids; in this sense, mcr-1 was encoded in an IncI2-type plasmid (23). More studies are needed to determine if there is a relationship between these isolates (i.e., mcr-1–blaCMY−2 bearing isolates of human and animal origin) or the genetic platforms that encode them. In addition, we also found an isolate obtained from poultry, carrying mcr-1. Presumably, the mandatory ban in Uruguay on veterinary usage of colistin, in any of its forms, will have beneficial effects by reducing the selection pressure on microorganisms carrying mcr-1. However, keeping in mind that mcr alleles are frequently associated with genes conferring resistance to other antibiotics widely used in veterinary medicine, such as quinolones and oxyimino cephalosporins, co-selection events are likely to occur. This has been demonstrated by the presence of the mcr gene along with ESBL, carbapenemases or plasmid-mediated quinolone resistance mechanisms such as qnrB or qnrS in different plasmids (IncI2, IncX4, y IncHI2) (24–27).
Regarding resistance to 3rd generation cephalosporins and quinolones in swine and poultry, the most frequently detected genes were blaCTX−M−8 and blaCMY−2, and qnrB19, respectively. A similar situation was found in human Salmonella enterica isolates in recent surveillance studies in our country (16). Since this microorganism is a primary pathogen and is associated with episodes of gastroenterocolitis, it could act as a doorway for resistance genes circulating in agricultural and veterinary environments. Notwithstanding, the intake of E. coli strains harboring the same or other resistance mechanisms could go unnoticed in the event of asymptomatic gut colonization.
In this concern, colonization with E.coli carrying resistance genes to antibiotics of critical use could be considered as a silent zoonosis, contributing to the resistance gene pool of microorganisms present in the gastrointestinal tract of animals and humans. It has been observed that gut colonization by E. coli strains with reduced susceptibility to fluoroquinolones can last from 2 weeks to 6 months, whereas the presence of β-lactamase-producing E. coli in outpatients can last up to 4 months in feces. Conversely, the persistence of ESBL-producing E. coli and K. pneumoniae in feces from recently discharged patients can last an average of 98 days (range 14–182 days) (28, 29).
Among the various ESBLs detected in poultry we also found blaCTX−M−55. This β-lactamase had never been reported in our country; yet in our neighboring country Brazil, it has been reported both in poultry and humans, usually associated to the glutathione transferase gene fosA3, responsible of conferring resistance to fosfomycin (30, 31). In order to assess this probable association in our poultry-derived isolates, we also performed PCR detection of fosA3, obtaining positive results (data not shown).
Although the main mechanism of resistance to oxyimino-cephalosporins in enterobacteria of human origin is blaCTX−M−15, this ESBL gene occasionally occurred in the isolates analyzed in the present work. In Uruguay blaCTX−M−2, blaCTX−M−8, blaSHV−12, and blaSHV−2 are among the most frequently detected β-lactamase genes, mainly in pediatric samples; more studies are needed to determine if there is any link with the microorganisms detected in this work or, perhaps, with the genetic structures that encode these genes (32–34).
In a previous report, we found E. coli isolates obtained from cattle showing some degree of resistance to fluoroquinolones, namely, 7.3% were carriers of PMQR (mainly qnrB2 and qnrS) (14). In the present work, the number of isolates carrying PMQR genes rose to 31.5% (23/73); furthermore, we also detected a change in the circulating alleles. In this regard, gene qnrB19 was the most frequently found, while the presence of qnrE1 and qnrS1 was detected in two and one isolate, respectively. The occurrence of qnrE1 has been recently reported in isolates of Salmonella enterica from cattle in Brazil (35), however it has not been described outside the Salmonella genus in neither cattle nor poultry; thus, in this work we confirm the circulation of such gene in E. coli both in cattle and poultry.
The qnrE1 gene was first reported in Argentina in 2017, in a human clinical isolate obtained in 2007 (36). Interestingly, in 2011 we reported the occurrence in Uruguay of a qnrB variant, termed qnrBKp737 (defined by a partial sequence of 606 bp obtained from a PCR amplification product) (37). The in silico translation of the partial nucleotide sequence displayed 26 amino acid differences with QnrB1; further comparison showed that qnrBKp737 was identical to qnrE1 (unpublished data). Apparently, this resistance mechanism scarcely reported so far, is also a long-standing problem in the context of “One Health” in our region.
One limitation of our work is the fact that we did not conduct a study of risk factors to determine which variables influence the selection of resistant microorganisms.
In light of our results, it will be necessary to carry out new studies encompassing a greater number of establishments with a design that allows us to analyze these aspects.
Beyond these limitations, the wide distribution of fluoroquinolone-resistant isolates in the animals analyzed is alarming and may reflect the widespread use of antimicrobials such as enrofloxacin. Since this class of antibiotics is on the list of highest priority critically important antibiotics for human health, their use in veterinary medicine should be drastically limited.
In conclusion, we have detected transferable resistance genes to the three antibiotics considered critical to human health, present in feces from farm animals in Uruguay. Several of such genes have also been reported previously in microorganisms of human origin in our country. Tackling the problem of antimicrobial resistance requires comprehensive approaches, including prudent use of antibiotics and surveillance under the “One Health” concept.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
NC, AU, and PÁ: carrying out of antibiotic sensitivity studies and detection of resistance genes by PCR and preparation of the manuscript. BF: pig samples collection, antibiotic sensitivity studies, and detection of resistance genes by PCR. NFC: real-time PCR for mcr-1, 2, and 3, interpretation of results, and preparation of the manuscript. GT: poultry samples collection, interpretation of results, and preparation of the manuscript. GC: pig samples collection, interpretation of results, and preparation of the manuscript. MC and MF: calves sample collection, interpretation of results, and preparation of the manuscript. PZ: experimental design, general coordination, interpretation of results, and elaboration of the manuscript. IB and RV: experimental design, general coordination, sequence analysis, interpretation of results, and elaboration of the manuscript. All authors contributed to the article and approved the submitted version.
This work was supported by grants from Agencia Nacional de Investigación e Innovación (ANII) and Espacio Interdisciplinario Universidad de la República to RV. This work was funded by the project PL-15 from Instituto Nacional de Investigación Agropecuaria (INIA). This work was also funded by grant to NC from Comisión Sectorial de Investigación Científica (CSIC).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fvets.2020.588919/full#supplementary-material
1. Hubalek Z. Emerging human infectious diseases: anthroponoses, zoonoses, and sapronoses. Emerg Infect Dis. (2003) 9:403–4. doi: 10.3201/eid0903.020208
2. Aarestrup FM. The livestock reservoir for antimicrobial resistance: a personal view on changing patterns of risks, effects of interventions and the way forward. Philos Trans R Soc Lond B Biol Sci. (2015) 370:20140085. doi: 10.1098/rstb.2014.0085
3. DeHart RL. Health issues of air travel. Annu Rev Public Health. (2003) 24:133–51. doi: 10.1146/annurev.publhealth.24.100901.140853
4. Wilson ME. The traveller and emerging infections: sentinel, courier, transmitter. J Appl Microbiol. (2003) 94(Suppl.):1S−11S. doi: 10.1046/j.1365-2672.94.s1.1.x
5. Ewers C, Grobbel M, Stamm I, Kopp PA, Diehl I, Semmler T, Fruth A, et al. Emergence of human pandemic O25:H4-ST131 CTX-M-15 extended-spectrum-beta-lacta-mase-producing Escherichia coli among companion animals. J Antimicrob Chemother. (2010) 65:651–60. doi: 10.1093/jac/dkq004
6. World Health Organization. Plan de Acción Mundial sobre la Resistencia a los Antimicrobianos. (2016) Available online at: http://www.who.int/antimicrobial-resistance/global-action-plan/es/
7. Van Boeckel TP, Brower C, Gilbert M, Grenfell BT, Levin SA, Robinson TP, et al. Global trends in antimicrobial use in food animals. Proc Natl Acad Sci USA. (2015) 112:5649–54. doi: 10.1073/pnas.1503141112
8. Lekshmi M, Ammini P, Kumar S, Varela MF. The food production environment and the development of antimicrobial resistance in human pathogens of animal origin. Microorganisms. (2017) 5:11. doi: 10.3390/microorganisms5010011
9. World Health Organization. WHO Guidelines on Use of Medically Important Antimicrobials in Food-Producing Animals: Web Annex A: Evidence Base. World Health Organization (2017).
10. World Health Organization. Critically Important Antimicrobials for Human Medicine, 5th rev. Geneva (2017).
12. Roberts RR, Hota B, Ahmad I, Douglas Scott R 2nd, Foster SD, Abbasi F, et al. Hospital and societal costs of antimicrobial-resistant infections in a Chicago Teaching Hospital: implications for antibiotic stewardship. Clin Infect Dis. (2009) 49:1175–84. doi: 10.1086/605630
13. Patrick DM, Chambers C, Purych D, Chong M, George D, Marra F. Value of anaggregate index in describing the impact of trends in antimicrobial resistance for Escherichia coli. Can J Infect Dis Med Microbiol. (2015) 26:33–8. doi: 10.1155/2015/274298
14. Umpiérrez A, Bado I, Oliver M, Acquistapace S, Etcheverría A, Padola LN, et al. Zoonotic potential and antibiotic resistance of Escherichia coli in neonatal calves in Uruguay. Microbes Environ. (2017) 32:275–82. doi: 10.1264/jsme2.ME17046
15. CLSI. Performance Standards for Antimicrobial Susceptibility Testing: 30th Informational Supplement. M100-S30. Wayne, PA: Clinical and Laboratory Standards Institute (2020).
16. Cordeiro NF, Nabón A, García-Fulgueiras V, Álvez M, Sirok A, Camou T, et al. Analysis of plasmid mediated quinolone and oxyimino-cephalosporin resistance mechanisms in Uruguayan Salmonella enterica isolates from 2011 to 2013. J Glob Antimicrob Resist. (2016) 6:165–71. doi: 10.1016/j.jgar.2016.06.002
17. Garcia-Fulgueiras V, Zapata Y, Papa-Ezdra R, Ávila P, Caiata L, Seija V, et al. First characterization of K. pneumoniae ST11 clinical isolates harboring blaKPC-3 in Latin America. Rev Argent Microbiol. 52:211–6. doi: 10.1016/j.ram.2019.10.003
18. Li J, Shi X, Yin W, Wang Y, Shen Z, Ding S, et al. A multiplex SYBR green real-time PCR assay for the detection of three colistin resistance genes from cultured bacteria, feces, and environment samples. Front Microbiol. (2017) 8:2078. doi: 10.3389/fmicb.2017.02078
19. Carattoli A, Villa L, Feudi C, Curcio L, Orsini S, Luppi A, et al. Novel plasmid-mediated colistin resistance mcr-4 gene in Salmonella and Escherichia coli, Italy 2013, Spain and Belgium, 2015 to 2016. Eurosurveillance. (2017) 22:30589. doi: 10.2807/1560-7917.ES.2017.22.31.30589
20. Yamamoto Y, Calvopina M, Izurieta R, Villacres I, Kawahara R, Sasaki M, et al. Colistin-resistant Escherichia coli with mcr genes in the livestock of rural small-scale farms in Ecuador. BMC Res Notes. (2019) 12:121. doi: 10.1186/s13104-019-4144-0
21. Faccone D, Moredo FA, Giacoboni GI, Albornoz E, Alarcón L, Nievas VF, et al. Multidrug-resistant Escherichia coli harbouring mcr-1 and blaCTX-M genes isolated from swine in Argentina. J Glob Antimicrob Resist. (2019) 18:160–2. doi: 10.1016/j.jgar.2019.03.011
22. García V, García-Meniño I, Mora A, Flament-Simon SC, Díaz-Jiménez D, Blanco JE, et al. Co-occurrence of mcr-1, mcr-4 and mcr-5 genes in multidrug-resistant ST10 enterotoxigenic and Shiga toxin-producing Escherichia coli in Spain (2006-2017). Int J Antimicrob Agents. (2018) 52:104–8. doi: 10.1016/j.ijantimicag.2018.03.022
23. Papa-Ezdra R, Grill Diaz F, Vieytes M, García-Fulgueiras V, Caiata L, Ávila P, et al. First three Escherichia coli isolates harbouring mcr-1 in Uruguay. J Glob Antimicrob Resist. (2020) 20:187–90. doi: 10.1016/j.jgar.2019.07.016
24. Wales AD, Davies RH. Co-selection of resistance to antibiotics, biocides and heavy metals, and its relevance to foodborne pathogens. Antibiotics. (2015) 4:567–604. doi: 10.3390/antibiotics4040567
25. Alba P, Leekitcharoenphon P, Franco A, Feltrin F, Ianzano A, Caprioli A, et al. Molecular epidemiology of mcr-encoded colistin resistance in enterobacteriaceae from food-producing animals in Italy revealed through the EU harmonized antimicrobial resistance monitoring. Front Microbiol. (2018) 9:1217. doi: 10.3389/fmicb.2018.01217
26. Haenni M, Poirel L, Kieffer N, Châtre P, Saras E, Métayer V, et al. Co-occurrence of extended spectrum? lactamase and MCR-1 encoding genes on plasmids. Lancet Infect Dis. (2016) 16:281–2. doi: 10.1016/S1473-3099(16)00007-4
27. Mulvey MR, Mataseje LF, Robertson J, Nash JH, Boerlin P, Toye B, et al. Dissemination of the mcr-1 colistin resistance gene. Lancet Infect Dis. (2016) 16:289–90. doi: 10.1016/S1473-3099(16)00067-0
28. Treisman GJ, Angelino AF. Duration of stool colonization in patients infected with extended- spectrum β-lactamase–producing Escherichia coli and Klebsiella pneumoniae. Reply to Rogers and Curry. Clin Infect Dis. (2008) 46:1321–2. doi: 10.1086/533474
29. Oplustil CP, Nunes R, Mendes C. Multicenter evaluation of resistance patterns of Klebsiella pneumoniae, Escherichia coli, Salmonella spp and Shigella spp isolated from clinical specimens in Brazil: RESISTNET surveillance program. Brazil J Infect Dis. (2001) 5:8–12. doi: 10.1590/S1413-86702001000100002
30. Cunha MPV, Lincopan N, Cerdeira L, Esposito F, Dropa M, Franco LS, et al. Coexistence of CTX-M-2, CTX-M-55, CMY-2, FosA3, and QnrB19 in extraintestinal pathogenic Escherichia coli from poultry in Brazil. Antimicrob Agents Chemother. (2017) 61:e02474–16. doi: 10.1128/AAC.02474-16
31. Fernandes MR, Sellera FP, Moura Q, Souza TA, Lincopan N. Draft genome sequence of a CTX-M-8, CTX-M-55 and FosA3 co-producing Escherichia coli ST117/B2 isolated from an asymptomatic carrier. J Glob Antimicrob Resist. (2018) 12:183–4. doi: 10.1016/j.jgar.2018.01.015
32. Garcia-Fulgueiras V, Araujo L, Bado I, Cordeiro NF, Mota MI, Laguna G, et al. Allodemic distribution of plasmids co harbouring CTX-M-15/Aac(6')Ib-cr/QnrB in K. pneumoniae is the main source of ESBL in Uruguay's Paediatric Hospital. J Glob Antimicrob Resist. (2017) 9:68–73. doi: 10.1016/j.jgar.2017.01.008
33. Vignoli R, García-Fulgueiras V, Cordeiro NF, Bado I, Seija V, Aguerrebere P, et al. Extended-spectrum β-lactamases, transferable quinolone-resistance and Virulotyping in extraintestinal E. coli in Uruguay. J infect Dev Ctries. (2016) 10:43–52. doi: 10.3855/jidc.6918
34. Bado I, Gutiérrez C, García-Fulgueiras V, Cordeiro NF, Araújo Pirez L, Seija V, et al. CTX-M-15 in combination with aac(6′)-Ib-cr is the most prevalent mechanism of resistance both in Escherichia coli and Klebsiella pneumoniae, including K. pneumoniae ST258, in an ICU in Uruguay. J Glob Antimicrob Resist. (2016) 6:5–9. doi: 10.1016/j.jgar.2016.02.001
35. Monte DF, Lincopan N, Cerdeira L, Fedorka-Cray PJ, Landgraf M. Early dissemination of qnrE1 in Salmonella enterica Serovar Typhimurium from livestock in South America. Antimicrob Agents Chemother. (2019) 63:e00571–19. doi: 10.1128/AAC.00571-19
36. Albornoz E, Tijet N, De Belder D, Gomez S, Martino F, Corso A, et al. qnrE1, a member of a new family of plasmid-located quinolone resistance genes originated from the chromosome of Enterobacter spp. Antimicrob Agent Chemother. (2017) 61:e02555–16. doi: 10.1128/AAC.02555-16
Keywords: swine, poultry, ESBL, E. coli, MCR-1, CTX-M-8, CMY-2, qnrB19
Citation: Coppola N, Freire B, Umpiérrez A, Cordeiro NF, Ávila P, Trenchi G, Castro G, Casaux ML, Fraga M, Zunino P, Bado I and Vignoli R (2020) Transferable Resistance to Highest Priority Critically Important Antibiotics for Human Health in Escherichia coli Strains Obtained From Livestock Feces in Uruguay. Front. Vet. Sci. 7:588919. doi: 10.3389/fvets.2020.588919
Received: 29 July 2020; Accepted: 19 October 2020;
Published: 19 November 2020.
Edited by:
Ioannis Magouras, City University of Hong Kong, Hong KongReviewed by:
Lucy Brunton, Royal Veterinary College (RVC), United KingdomCopyright © 2020 Coppola, Freire, Umpiérrez, Cordeiro, Ávila, Trenchi, Castro, Casaux, Fraga, Zunino, Bado and Vignoli. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Rafael Vignoli, cnZpZ25vbGlAaGlnaWVuZS5lZHUudXk=; Inés Bado, aWJhZG9AaGlnaWVuZS5lZHUudXk=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.