
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
BRIEF RESEARCH REPORT article
Front. Vet. Sci., 25 November 2020
Sec. Veterinary Epidemiology and Economics
Volume 7 - 2020 | https://doi.org/10.3389/fvets.2020.547843
This article is part of the Research TopicAntimicrobial Resistance in Zoonotic Bacteria in Developing Countries: The Role of Food Animal Production in Public HealthView all 11 articles
 David Ortega-Paredes1
David Ortega-Paredes1 Sofía de Janon1Fernando Villavicencio2Katherine Jaramillo Ruales2Kenny De La Torre3
Sofía de Janon1Fernando Villavicencio2Katherine Jaramillo Ruales2Kenny De La Torre3 José E. Villacís2,3
José E. Villacís2,3 Jaap A. Wagenaar4,5Jorge Matheu6Camila Bravo-Vallejo7Esteban Fernández-Moreira8
Jaap A. Wagenaar4,5Jorge Matheu6Camila Bravo-Vallejo7Esteban Fernández-Moreira8 Christian Vinueza-Burgos1*
Christian Vinueza-Burgos1*Antimicrobial resistance (AMR) is a major health threat for public and animal health in the twenty-first century. In Ecuador, antibiotics have been used by the poultry industry for decades resulting in the presence of multi-drug resistant (MDR) bacteria in the poultry meat production chain, with the consequent risk for public health. This study evaluated the prevalence of ESBL/AmpC and mcr genes in third-generation cephalosporin-resistant Escherichia coli (3GC-R E. coli) isolated from broiler farms (animal component), broiler carcasses (food component), and human enteritis (human component) in Quito-Ecuador. Samples were collected weekly from November 2017 to November 2018. For the animal, food, and human components, 133, 335, and 302 samples were analyzed, respectively. Profiles of antimicrobial resistance were analyzed by an automated microdilution system. Resistance genes were studied by PCR and Sanger sequencing. From all samples, 122 (91.7%), 258 (77%), and 146 (48.3%) samples were positive for 3GC-R E. coli in the animal, food, and human components, respectively. Most of the isolates (472/526, 89.7%) presented MDR phenotypes. The ESBL blaCTX-M-55, blaCTX-M-3, blaCTX-M-15, blaCTX-M-65, blaCTX-M-27, and blaCTX-M-14 were the most prevalent ESBL genes while blaCMY-2 was the only AmpC detected gene. The mcr-1 gene was found in 20 (16.4%), 26 (10.1%), and 3 (2.1%) of isolates from animal, food, and human components, respectively. The implication of poultry products in the prevalence of ESBL/AmpC and mcr genes in 3GC-R must be considered in the surveillance of antimicrobial resistance.
The World Health Organization recognizes antimicrobial resistance (AMR) as a major health threat in the 21st century (1). A global projection predicts that the increase of deaths linked to AMR will develop from 700,000 in 2016 to 10 million deaths per year in 2050 while 100 trillion USD could be lost by 2050 (2). In this scenario, the use of antibiotics in food animal production is one of the most important issues contributing to the AMR crisis. In fact, over 50% of antibiotic production is used by the meat industry and an increase of 50% in antibiotic usage for farming is estimated by 2030. Moreover, up to a 160% increase in antibiotic usage in food animals is expected in Latin American countries in absence of changes (3).
Poultry production is an important sector for the study of AMR because of the common usage of antibiotics in this industry. Additionally, it is expected that poultry will be the main animal production industry by 2025 (4). This issue is especially significant in developing countries where antimicrobials are not only used to treat infections but also prophylactically and as growth promoters (5). In Ecuador, poultry products are the most important source of animal protein with a per capita consumption of poultry meat of 30.4 kg/year (6).
In Ecuador, commonly used antibiotics in poultry production include quinolones, fosfomycin, and colistin, which are listed by WHO as critically important antimicrobials for human medicine, with quinolones and colistin even highest prioritized (7). This practice has promoted the dissemination of multi-drug resistant (MDR) bacteria, principally extended-spectrum β-lactamases and AmpC-producing (3GC-R) Escherichia coli, which is commonly studied as a sentinel organism to understand the epidemiology of AMR (8). In Ecuador, colistin was banned for use in food animals in 2019. However, mcr genes have been recently identified in E. coli isolated from animals, humans, and the environment (9–11).
A relationship between 3GC-R E. coli isolated from poultry products and humans has been suggested previously (12–16). In Ecuador, Vinueza-Burgos et al. described a high prevalence of 3GC-R E. coli and the presence of mcr-1 in poultry farms (17). However, there is no data about the prevalence of 3GC-R and colistin-resistant E. coli in broiler carcasses and humans in this location. Therefore, this study aimed to evaluate the state of 3GC-R and mcr genes in E. coli in broiler farms, chicken carcasses at retail level and human stool samples in Ecuador.
Samples for E. coli isolation were collected weekly from November 2017 to November 2018. Under local legislation, ethical approval was not required for collecting chicken caeca and carcasses during sampling. The participants for the human component were informed about the objective of the study; the participation was voluntary (all volunteers provided a written consent) and all personal information was anonymized. This project was approved by the committee of bioethics from the National Institute of Public Health “Leopoldo Izquieta Pérez” (Protocol ID: CEISH-INSPI-005). The sample distribution for each component considered local characteristics (location of farms in the zone of Quito, distribution of retail shops in the city, and location of healthcare centers).
The sample size for the animal component was calculated considering the number of industrial farms close to Quito, the number of batches that these farms produce in 1 year, and the prevalence of 3GC-R E. coli in poultry farms previously reported in this zone (17). For the food and human components, the minimum sample size was calculated considering an infinite population and at a 0.9 of confidence.
For the animal component,133 flocks from 69 farms close to Quito were sampled during the study period. For every sampled flock, 25 caeca from 25 chicken were randomly collected at the slaughterhouse and transported to the laboratory in an icebox within 2 h. At the laboratory, a sample pool of 25 g. was obtained and homogenized by hand as previously described (17).
For the food component, 335 carcasses were collected in three kinds of markets as follows: 125 samples from supermarkets, 126 samples from small shops, and 84 samples from open markets. Sampling places were distributed in both northern and southern areas of Quito. A sampling of chicken carcasses was performed alternately between the north and south of the city. Each sample consisted of one carcass collected in its original bag and transported to the laboratory in an icebox within 2 h. At the laboratory, 25 g of breast skin of every carcass were aseptically collected for bacteriological analysis.
For the human component, stool samples were collected in two health care centers located in the urban periphery of Quito. The inclusion criteria of the patients from whom samples were taken were: individuals with two or more episodes of diarrhea or vomiting in the last 24 h. Stool samples were transported to the laboratory in an icebox within 2 h and 25 g of feces were collected for bacteriological analysis.
All samples were homogenized with 225 ml of buffered peptone water (BPW; Difco, BD, Sparks, MD) and incubated at 37°C for 18–24 h. A loopful of each sample was streaked onto chromogenic Tryptone Bile X-Glucuronide (TBX) agar (BioRad, Hercules, California, USA) supplemented with cefotaxime (3 mg/l). Positive plates were considered when at least one typical colony could be selected (when available, two colonies were selected for further analysis) and confirmed to be E. coli using Triple Sugar Iron (TSI) agar (Difco, BD, Detroit, USA) and by PCR as described elsewhere (18). From the TSI medium, one loopful was suspended in 300 μl of sterile water and used to extract DNA by the boiling method. Another loopful was used to subculture the isolate in trypticase soy broth (TSB) (Difco, BD, Detroit, USA) and stored with glycerol (60%) at −80°C.
Resistance profiles to antibiotics commonly tested for Enterobacteraceae were obtained for all isolates using the Vitek® 2 system with AST-N271 cards (BioMérieux, Marcy-l'Étoile, France). The following antibiotics were tested: ampicillin, ampicillin + sulbactam, cephalothin, cefuroxime, ceftriaxone, cefotaxime, ceftazidime, cefepime, ertapenem, meropenem, amikacin, gentamicin, ciprofloxacin, norfloxacin, fosfomycin, nitrofurantoin, and trimethoprim + sulfamethoxazole. The MIC for colistin was tested in isolates positive for mcr genes by PCR using BD Phoenix™ M50 with NMIC/ID 94 panel (Becton Dickinson, Nueva Jersey, USA). Antimicrobial resistance phenotypes were obtained following the manufacturer's instructions. E. coli ATCC 25922 was used as a quality control strain. Results were evaluated using the clinical breakpoints recommended by CLSI (19). Isolates resistant to at least 3 antibiotic classes were considered as MDR. The mcr 1 to 5, blaCTX-M, blaTEM, blaSHV, and blaCMY genes were tested by PCR in all isolates as previously described (17, 20). Obtained amplicons were sequenced at Macrogen (Seoul-South Korea). Sequences were analyzed using Genious Prime software with the ResFinder database (21).
Significate differences of the prevalence of 3GC-R E. coli within and among each component were calculated using a χ2 test. This statistical test was also used to assess differences in antibiotics resistance rates and the presence of resistance genes variants as well as determining differences among farms, types of markets, and location of health care centers (p < 0.05). The 95% confidence intervals (CI95%) for the prevalence of 3GC-R E. coli was obtained by Binomial “exact” calculation. All tests were carried out in RStudio V.1.2.
A total of 526 samples were positive for 3GC-R E. coli. The highest prevalence was identified in the animal component (91.7%; CI95%: 90.8–92.7), followed by the food component (77%; CI95%: 76.3–77.8) and the human component (48.3%; CI95%: 47.9–48.8). χ2 test identified significant differences among components (p < 0.05). However, no significant differences were observed within farms, types of shops, and locations of health centers (p > 0.05).
All the isolates were resistant to ampicillin, cephalothin, cefuroxime, ceftriaxone, cefotaxime, and trimethoprim + sulfamethoxazole. High resistance rates (more than 80%), were registered for cefepime and ceftazidime in the three components. Resistance to ciprofloxacin and norfloxacin ranged from 70 to 80% in the animal and food components and were significantly higher than rates in the human component, where half of the isolates were resistant. AMR to the combination β-lactam + β-lactamase inhibitor (Ampicillin + Sulbactam) was around 55% in the three components.
Resistance to fosfomycin, nitrofurantoin, and gentamicin showed significant differences between the three components ranging from 30 to 50% in animal and food components and from 24 to 30% in the human component, while for cefepime the resistance rate was higher in humans (p < 0.05). Five isolates (two from the animal, one from food, and two from human components) were resistance to carbapenems (ertapenem and/or meropenem). None of the isolates were resistant to Amikacin (Table 1). Additionally, 89.7% of the isolates (472/526), presented MDR patterns with three up to seven groups of antibiotics. The most frequent pattern included resistance to β-lactams, fluoroquinolones, and folate pathway inhibitors (Table 2, Supplementary File 1). Distribution of MIC values for every tested antibiotic in each component shown in Supplementary File 4.
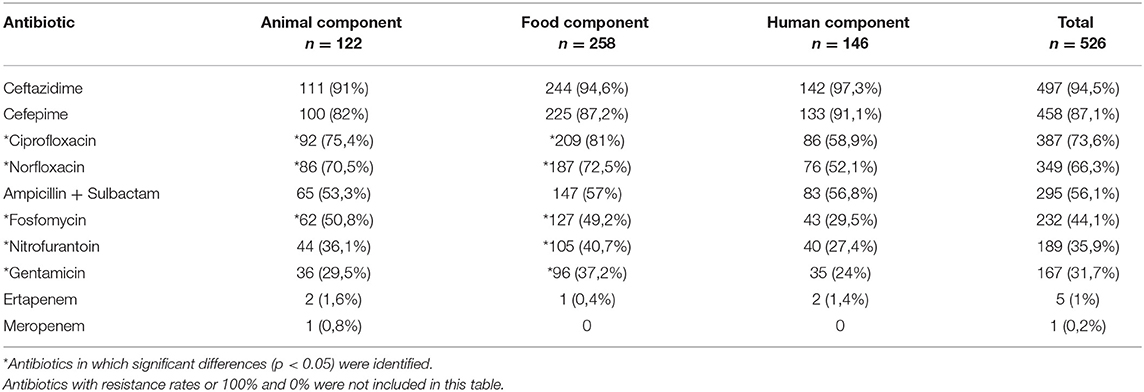
Table 1. Antibiotic resistance rates in each component.
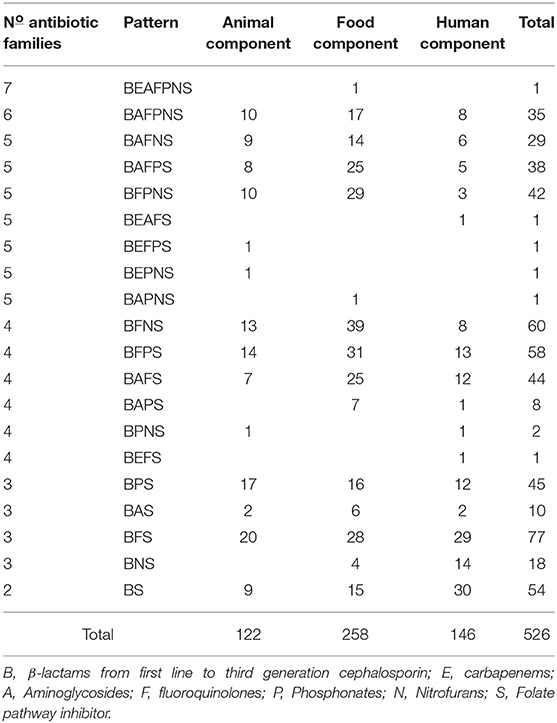
Table 2. Antimicrobial resistance patterns by antibiotic family.
Group 1 of blaCTX-M genes was the most prevalent family of ESBL genes, followed by group 9, group 2, and group 8. The allele blaCTX-M-55, belonging to group 1, was identified as the most prevalent variant in the three components, followed by blaCTX-M-3 in animal and food components and blaCTX-M-15 in the human component. Among blaCTX-M group 9, the most frequent allele in the three components was blaCTX-M-65 followed by blaCTX-M-27 (animal and food components) and blaCTX-M-14 (human component). The blaCTX-M-2 gene was present in the animal and food components, while the blaCTX-M-8 gene was detected in animal and human components. Moreover, ESBL variants of blaSHV and broad-spectrum β-lactamases blaTEM were frequently identified in the three components. Additionally, blaCTX-M-1, blaCTX-M-17, and blaCTX-M-2 were registered only in poultry isolates, blaCTX-M-8 was identified in food and human components, and blaCTX-M-15, blaCTX-M-14, and blaCTX-M-27 were identified only in the human component. Finally, only the AmpC gene blaCMY-2 was detected in the samples from the three components (Table 3, Supplementary File 2).
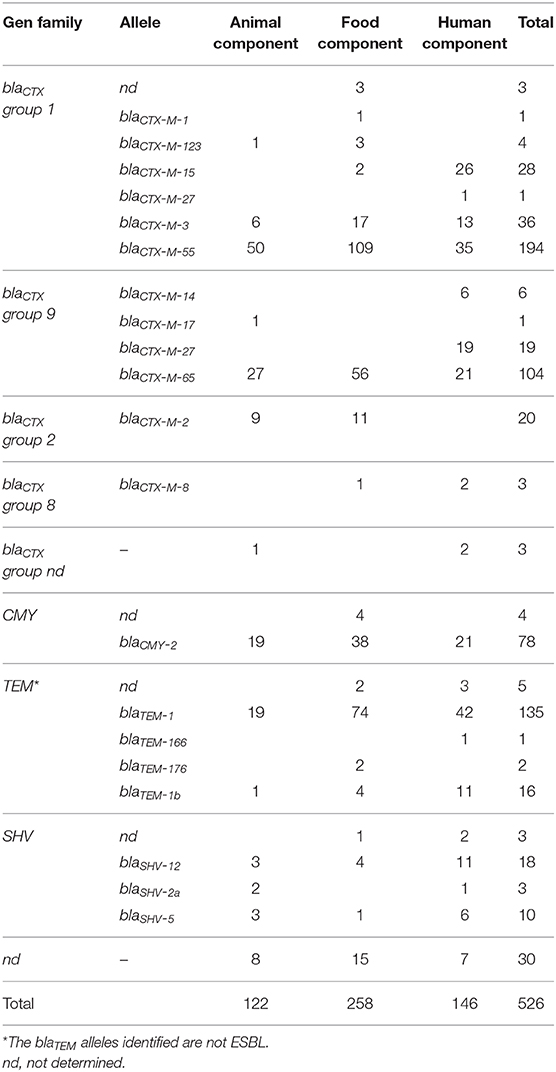
Table 3. Prevalence of ESBL/AmpC genes in the cefotaxime-resistant Escherichia coli isolates.
The mcr-1 gene was found in 16.4% (20/122), 10.1% (26/258), and 2.1% (3/146) of isolates from the animal, food, and human components, respectively. Colistin resistant isolates showed MIC values from 2 to 4 μg/ml. In these isolates, the allele blaCTX-M-55 was dominant in animal and food components, followed by blaCTX-M-65, blaCTX-M-2, and blaCTX-M-3. Additionally, blaCMY-2 and blaSHV-5 genes were detected in these components while the alleles blaCTX-M-15 and blaCTX-M-55 were the only ones detected mcr-1 positive isolates from humans (Supplementary File 3).
Studies of 3GC-R E. coli in poultry production remain scarce in the Andean region of South America (15, 20). Additionally, only a few of these studies have evaluated the prevalence of mcr genes in 3GC-R E. coli from poultry (17). To the best of our knowledge, this issue has not been evaluated with a multi-component approach in this region.
The high prevalence of 3GC-R E. coli in poultry showed in this study is in concordance with a previous report (17). These issues could be related to the rapid dissemination of 3GC-R mediated by horizontal transfer in broiler farms (22). On the other hand, the prevalence in the food component was significantly lower than in the animal component. This fact has been observed in Brazil, where 3GC-R E. coli was isolated from 54.2 and 29.2% of samples coming from animals and chicken carcasses, respectively (23). Besides, around 50% of the human stool samples were positive for 3GC-R E. coli. Prevalence of 3GC-R E. coli in healthy carriers has been reported in the Netherlands, Japan, India, Libya, and Sweden ranging from >5 to 19% (24). Moreover, E. coli ESBL has been registered in 18.8% of pediatric patients with diarrhea (25). Our results suggest that the poultry environment in Ecuador is a reservoir of 3GC-R E. coli.
Carbapenem resistance mediated by blaKPC-2, blaKPC-3, blaOXA-48, blaNDM-1, and blaVIM-2 has been reported in Enterobacteriaceae in South American countries in human isolates (26). In Ecuador, blaKPC-2 is the most prevalent carbapenemase gene detected in E. coli in hospitals (27, 28), and recently it was detected in an urban river in Citrobacter freundii (29). However, there are no reports of carbapenemases from poultry in our region (carbapenems are not used for poultry production). In our study, the isolates resistant to imipenem/meropenem from the three components were negative for the blaKPC gene by PCR testing (data not shown). Therefore, this resistance could be related to not tested carbapenemase genes or to a combination of ESBL enzymes and porin loss (30).
In this study, resistance to fluoroquinolones was significantly higher in isolates originated in poultry (up to 80%) than in isolates originated in the human component (55%). These data are concordant with the common use of quinolones in the poultry industry in Ecuador (31). Additionally, ciprofloxacin is commonly used as an empiric treatment of community-acquired E. coli infections in humans. These practices could explain the high resistance rates observed for fluoroquinolones which has also been reported in other countries of the region (32). Even though resistance to fluoroquinolones could be explained by chromosomal point mutations (33), quinolone resistance mediated by mobile resistance genes (qnr) has been frequently associated with ESBL production in Enterobacteriaceae (34) suggesting that the co-transference of these genes is a common event.
Additionally, all the isolates in this study were resistant to trimethoprim + sulfamethoxazole. This finding could be explained by the presence of class 1 integrons. These site-specific recombination systems typically have a sul gene in their 3′CS end, and frequently present aminoglycoside, quinolone, and β-lactam resistance genes in the variable region. These features promote the selection and evolution of these genetic platforms in mixed antibiotic pressure environments (35), which could explain our findings. However, a genetic analysis to test those elements is necessary to confirm this hypothesis.
The resistance rates to nitrofurantoin reported in this study are higher than the ones reported previously in Ecuador and Colombia (36). However, this antibiotic is not used in the poultry industry. The mechanism of resistance and the reason for the increase of resistance rates reported in this study remain unknown. Our results also showed that half of the isolates from poultry and about 30% of human isolates were resistant to fosfomycin. In South American countries resistance to fosfomycin in E. coli isolated from human infections has been reported ranging from 2 to 3% (37, 38). On the other hand, a study carried out in Brazil reported a lower prevalence of this resistance in poultry. There are no other reports of fosfomycin resistance in poultry in neighboring countries, but a close relation of Enterobacteria isolated from poultry between Peru and Ecuador has been described before, proposing the hypothesis of a common epidemiology of these bacteria in the Andes region (30, 31). Nitrofurantoin and fosfomycin are antibiotics prescribed for the treatment of infections caused by MDR and extremely resistant enterobacteria (39, 40). Our findings highlight the urgency of a better regulation of the usage of these antibiotics in Ecuador.
Finally, the aminoglycoside amikacin is not used in poultry production and is restricted to complicated infections in humans (41), so the susceptibility of all isolates to amikacin is not surprising.
In this study, blaCTX-M-55 was the most prevalent allele of the blaCTX-M group, followed by blaCTX-M-65 and blaCTX-M-2 in the poultry components. These results show a change in the prevalence of blaCTX-M genes compared with the report a previous report where blaCTX-M-65, blaCTX-M-55, and blaCTX-M-3 were the most prevalent alleles in poultry isolates (17). These outcomes differ with reports in other Latino-American countries as Colombia, where the most dominant allele was blaCMY-2 (42), and Brazil where blaCTX-M-8 and blaCTX-M-2 are the most important alleles in poultry (37, 38, 43, 44). Additionally, in the human component blaCTX-M-55, blaCTX-M-15, and blaCTX-M-65 were the most prevalent alleles. Based on clinical evidence, the change of the dominance of blaCTX-M-15 to blaCTX-M-55 in humans in Ecuador was already hypostatized in 2016 by Zurita et al. (45).
The presence of component-specific alleles of blaCTX-M genes also suggests the existence of specific reservoirs. Moreover, it has been hypothesized that the ecological characteristics were animal husbandry is carried out in the region (e.g., altitude, lack of seasons, etc.) could be related to specificities in the epidemiology of these genetic determinants (15, 29). Additionally, it should be noted that the methods used to screen the presence of 3GC-R E. coli could give biased information. For example, the use of selective media containing ceftazidime could contribute to recover isolates carrying ceftazidimases with low affinity to cefotaxime; principally alleles of blaCTX-M, blaTEM, blaSHV, blaPER, blaVEB, blaTLA, and blaGES/IBC (46). On the other hand, media supplemented with cefoxitin could be used for the proper isolation of AmpC β-lactamase-producing E. coli.
In Ecuador the mcr genes have been reported before in E. coli isolated from humans and animals (9–11, 15); but, to the best of our knowledge, this is the first time that genetic determinants for colistin resistance are studied in a multiple-component frame in our region. From all mcr genes tested in this study, only the mcr-1 gene was detected. This gene was reported in a previous study with a lower prevalence (17) which could indicate that the prevalence of mcr-1 in the poultry production has increased. On the other hand, the prevalence of mcr-1 in the human component remains low. This relation has been emphasized before, pointing out the animal production origin of this gene (47). Although the use of colistin as a growth promotor has been recently banned in Ecuador, the former intensive use of this antibiotic in the poultry production could be seen as the main trigger for the prevalence observed in the animal component. Our results show that poultry production is an important reservoir of 3GC-R E. coli that harbors the mcr-1 gene.
It is worth to mention that, as some of the tested antibiotics in this study are not used in poultry production (e.g., cephalosporins and nitrofurantoin), the resistance of E. coli to these drugs could be mediated by co-selection events (selection of mixed AR under the pressure of single agent) (48). Genomic elements as conjugating plasmids, insertion sequences, and integrons could play a main role in the dissemination and accumulation of AMR determinants in the poultry production environment. Therefore, co-selection and co-resistance processes should be considered when implementing strategies for AMR control. Besides, environmental factors as contamination of upstream rivers with antibiotics and resistant bacteria should be considered in this analysis (49).
Poultry production has been recognized as an important environment for the evolution of AMR worldwide (50, 51). Indeed, new configurations of resistance genes have been described from poultry production. Important examples are the colocation of mcr-1 and mcr-3 in plasmids (52), and the emergence of E. coli strains co-carrying ESBL and fosA3 genes (38). These findings suggest that the antibiotic pressure in poultry production promotes the active recombination and selection of MDR genotypes. Therefore, the presence of strains with new combinations of genetic determinants is a real possibility that should be further studied. In fact, the MDR patterns found in this study suggest the presence of multiple resistance mechanisms that deserve a deeper analysis at the genomic level.
The high prevalence of 3GC-R E. coli reported in this study is worrisome in terms of public health and highlights the need for health policies to prevent the increase of AR in the country.
Additionally, the high prevalence of 3GC- R E. coli registered in our study poses a risk of transmission to humans via the food chain. However, the implication of poultry products in the epidemiology of 3GC-R E. coli needs further research since other sources for the human acquisition of these bacteria should be considered.
To the best of our knowledge, this study shows for the first time, data on 3GC-R E. coli with a multi-component approach in Latin America. We stress the importance of MDR phenotypes and genetic determinants that are spreading rapidly worldwide.
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author/s.
DO-P: sampling, interpreted the results, and wrote the manuscript. SJ and KDLT: sampling, interpreted the results, and database management. FV and JV: designed the study, sampling, and performed the experiments in human isolates. JW and JM: designed the study. EF-M: designed the study, provided the critical input, and interpreted the results. CB-V and KR: performed the colistin phenotyping experiments. CV-B: designed the study, provided the critical input, interpreted the results, and wrote the manuscript. KR and KDLT: performed critical laboratory analysis and data analysis. All authors contributed to the article and approved the submitted version.
This project was funded by the WHO Advisory Group on Integrated Surveillance on AMR (AGISAR) grant AGISAR-JM/lg.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors want to thank José Luis Medina, Janina Mantilla, Vladimir Naranjo, and Andrea Márquez for their kind help in sample processing.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fvets.2020.547843/full#supplementary-material
Supplementary File 1. Antimicrobial resistance patterns by Antibiotic.
Supplementary File 2. Genotyping of the isolates.
Supplementary File 3. Phenotyping and genotyping of mcr-1 positive strains.
Supplementary File 4. Distribution of MIC values.
1. IACG. No time to wait: Securing the Future from Drug-Resistant Infections. WHO, editor (2019). Available online at: https://www.who.int/docs/default-source/documents/no-time-to-wait-securing-the-future-from-drug-resistant-infections-en.pdf?sfvrsn=5b424d7_6
2. O'Neill J. Tackling Drug-Resistant Infections Globally: Final Report and Recommendations. The Review on Antimicrobial Resistance. (2016). Available online at: https://amr-review.org
3. Van Boeckel TP, Brower C, Gilbert M, Grenfell BT, Levin SA, Robinson TP, et al. Global trends in antimicrobial use in food animals. Proc Natl Acad Sci USA. (2015) 112:5649–54. doi: 10.1073/pnas.1503141112
4. OECD/FAO. Agricultural Outlook 2016–2025. (2016). Available online at: https://doi.org/10.1787/agr_outlook-2016-10-en
5. Vinueza-Burgos C, Wautier M, Martiny D, Cisneros M, Van Damme I, De Zutter L. Prevalence, antimicrobial resistance and genetic diversity of Campylobacter coli and Campylobacter jejuni in Ecuadorian broilers at slaughter age. Poult Sci. (2017) 96:2366–74. doi: 10.3382/ps/pew487
6. Conave. Estadísticas Del Sect Avícola. (2019). Available online at: https://www.conave.org/informacion-sector-avicola-publico/
8. WHO. Global Priority List of Antibiotic-Resistant Batceria to Guide Research, Discovery, and Development of New Antibiotics. Geneva: WHO (2017). Available online at: https://www.who.int/news-room/detail/27-02-2017-who-publishes-list-of-bacteria-for-which-new-antibiotics-are-urgently-needed
9. Ortega-Paredes D, Barba P, Zurita J. Colistin-resistant Escherichia coli clinical isolate harbouring the mcr-1 gene in Ecuador. Epidemiol Infect. (2016) 144:2967–70. doi: 10.1017/S0950268816001369
10. Ortega-Paredes D, Haro M, Leoro-Garzón P, Barba P, Loaiza K, Mora F, et al. Multidrug-resistant Escherichia coli isolated from canine faeces in a public park in Quito, Ecuador. J Glob Antimicrob Resist. (2019) 18:263–8. doi: 10.1016/j.jgar.2019.04.002
11. Yamamoto Y, Calvopina M, Izurieta R, Villacres I, Kawahara R, Sasaki M, et al. Colistin-resistant Escherichia coli with mcr genes in the livestock of rural small-scale farms in Ecuador. BMC Res Notes. (2019) 12:1–5. doi: 10.1186/s13104-019-4144-0
12. Overdevest I, Willemsen I, Rijnsburger M, Eustace A, Xu L, Hawkey P, et al. Extended-spectrum β-lactamase genes of Escherichia coli in chicken meat and humans, The Netherlands. Emerg Infect Dis. (2011) 17:1216–22. doi: 10.3201/eid1707.110209
13. Kluytmans JAJW, Overdevest ITMA, Willemsen I, Kluytmans-van den Bergh MFQ, van der Zwaluw K, Heck M, et al. Extended-spectrum β-lactamase-producing Escherichia coli from retail chicken meat and humans: comparison of strains, plasmids, resistance genes, and virulence factors. Clin Infect Dis. (2013) 56:478–87. doi: 10.1093/cid/cis929
14. Moser KA, Zhang L, Spicknall I, Braykov NP, Levy K, Marrs CF, et al. The role of mobile genetic elements in the spread of antimicrobial-resistant Escherichia coli from chickens to humans in small-scale production poultry operations in rural Ecuador. Am J Epidemiol. (2018) 187:558–67. doi: 10.1093/aje/kwx286
15. Pacholewicz E, Liakopoulos A, Swart A, Gortemaker B, Dierikx C, Havelaar A, et al. Reduction of extended-spectrum-β-lactamase- and AmpC-β-lactamase-producing Escherichia coli through processing in two broiler chicken slaughterhouses. Int J Food Microbiol. (2015) 215:57–63. doi: 10.1016/j.ijfoodmicro.2015.08.010
16. Mughini-Gras L, Dorado-García A, van Duijkeren E, van den Bunt G, Dierikx CM, Bonten MJM, et al. Attributable sources of community-acquired carriage of Escherichia coli containing β-lactam antibiotic resistance genes: a population-based modelling study. Lancet Planet Heal. (2019) 3:e357–69. doi: 10.1016/S2542-5196(19)30130-5
17. Vinueza-Burgos C, Ortega-Paredes D, Narvaéz C, De Zutter L, Zurita J. Characterization of cefotaxime resistant Escherichia coli isolated from broiler farms in Ecuador. PLoS ONE. (2019) 14:e207567. doi: 10.1371/journal.pone.0207567
18. Bej AK, Dicesare JL, Haff L, Atlas RM. Detection of Escherichia coli and Shigella spp. in water by using the polymerase chain reaction and gene probes for UID. Appl Environ Microbiol. (1991) 57:1013–7.
19. CLSI. Antimicrobial Susceptibility Testing Standards. CLSI Supplement for Global Application. (2020). Available online at: http://em100.edaptivedocs.net/GetDoc.aspx?doc=CLSIM100~ED30:2020&scope$=$user
20. Rebelo AR, Bortolaia V, Kjeldgaard JS, Pedersen SK, Leekitcharoenphon P, Hansen IM, et al. Multiplex PCR for detection of plasmid-mediated colistin resistance determinants, mcr-1, mcr-2, mcr-3, mcr-4 and mcr-5 for surveillance purposes. Eurosurveillance. (2018) 23:1–11. doi: 10.2807/1560-7917.ES.2018.23.6.17-00672
21. Zankari E, Hasman H, Cosentino S, Vestergaard M, Rasmussen S, Lund O, et al. Identification of acquired antimicrobial resistance genes. J Antimicrob Chemother. (2012) 67:2640–4. doi: 10.1093/jac/dks261
22. Hoek AHAM Van, Veenman C, Florijn A, Huijbers PMC, Graat EAM, Greeff S De, et al. Longitudinal study of ESBL Escherichia coli carriage on an organic broiler farm. J Antimicrob Chemother. (2018) 73:3298–304. doi: 10.1093/jac/dky362
23. Apostolakos I, Mughini-Gras L, Fasolato L, Piccirillo A. Assessing the occurrence and transfer dynamics of ESBL/pAmpC-producing Escherichia coli across the broiler production pyramid. PLoS ONE. (2019) 14:e217174. doi: 10.1371/journal.pone.0217174
24. González D, Gallagher E, Zúñiga T, Leiva J, Vitas AI. Prevalence and characterization of β-lactamase-producing Enterobacteriaceae in healthy human carriers. Int Microbiol. (2019) 23:171–7. doi: 10.1007/s10123-019-00087-z
25. Bagus Wasito E, Shigemura K, Osawa K, Fardah A, Kanaida A, Raharjo D, et al. Antibiotic susceptibilities and genetic characteristics of extended-spectrum beta-lactamase-producing Escherichia coli isolates from stools of pediatric diarrhea patients in Surabaya, Indonesia. Jpn J Infect Dis. (2017) 70:378–82. doi: 10.7883/yoken.JJID.2016.234
26. Reyes JA, Melano R, Cárdenas PA, Trueba G. Mobile genetic elements associated with carbapenemase genes in South American enterobacterales. Brazilian J Infect Dis. (2020) 24:231–8. doi: 10.1016/j.bjid.2020.03.002
27. Zurita J, Alcocer I, Ortega-Paredes D, Barba P, Yauri F, Iñiguez D, et al. Carbapenem-hydrolysing β-lactamase KPC-2 in Klebsiella pneumoniae isolated in Ecuadorian hospitals. J Glob Antimicrob Resist. (2013) 1:229–30. doi: 10.1016/j.jgar.2013.06.001
28. Soria-Segarra C, Soria-Segarra C, Catagua-González A, Gutiérrez-Fernández J. Carbapenemase producing Enterobacteriaceae in intensive care units in Ecuador: results from a multicenter study. J Infect Public Health. (2019) 13:80–8. doi: 10.1016/j.jiph.2019.06.013
29. Ortega-Paredes D, Barba P, Mena-López S, Espinel N, Crespo V, Zurita J. High quantities of multidrug-resistant Escherichia coli are present in the Machángara urban river in Quito, Ecuador. J Water Health. (2020) 18:67–76. doi: 10.2166/wh.2019.195
30. Lartigue MF, Poirel L, Poyart C, Réglier-Poupet H, Nordmann P. Ertapenem resistance of Escherichia coli. Emerg Infect Dis. (2007) 13:315–7. doi: 10.3201/eid1302.060747
31. Vinueza C. Salmonella and Campylobacter in broilers at slaughter age: a possible source for carcasses contamination in Ecuador (Ph.D. thesis), Ghent University, Ghent, Belgium (2017).
32. Vieira DC, Lima WG, de Paiva MC. Plasmid-mediated quinolone resistance (PMQR) among Enterobacteriales in Latin America: a systematic review. Mol Biol Rep. (2020) 47:1471–83. doi: 10.1007/s11033-019-05220-9
33. Correia S, Poeta P, Hébraud M, Capelo JL, Igrejas G. Mechanisms of quinolone action and resistance: where do we stand? J Med Microbiol. (2017) 66:551–9. doi: 10.1099/jmm.0.000475
34. Farajzadehsheikh A, Veisi H, Shahin M, Getso M, Farahani A. Frequency of quinolone resistance genes among extended-spectrum?-lactamase (ESBL)-producing Escherichia coli strains isolated from urinary tract infections. Trop Med Health. (2019) 47:1–7. doi: 10.1186/s41182-019-0147-8
35. Ortega-Paredes D, Zurita J. Integrones, plataformas bacterianas de recombinación. Rev Ecuat Med Cienc Biol. (2013) XXXIV:167–85. doi: 10.26807/remcb.v34i1-2.242
36. Donado-godoy P, Byrne BA, León M, Castellanos R, Vanegas C, Coral A, et al. Prevalence, resistance patterns, and risk factors for antimicrobial resistance in bacteria from retail chicken meat in Colombia. J Food Prot. (2015) 78:751–9. doi: 10.4315/0362-028X.JFP-14-349
37. Bartoloni A, Sennati S, Di Maggio T, Mantella A, Riccobono E, Strohmeyer M, et al. Antimicrobial susceptibility and emerging resistance determinants (blaCTX-M, rmtB, fosA3) in clinical isolates from urinary tract infections in the Bolivian Chaco. Int J Infect Dis. (2016) 43:1–6. doi: 10.1016/j.ijid.2015.12.008
38. Cyoia PS, Koga VL, Nishio EK, Houle S, Dozois CM, De Brito KCT, et al. Distribution of ExPEC virulence factors, bla CTX-M, fosA3, and mcr-1 in escherichia coliisolated from commercialized chicken carcasses. Front Microbiol. (2019) 10:3254. doi: 10.3389/fmicb.2018.03254
39. Gardiner BJ, Stewardson AJ, Abbott IJ, Peleg AY. Nitrofurantoin and fosfomycin for resistant urinary tract infections: old drugs for emerging problems. Aust Prescr. (2019) 42:14–9. doi: 10.18773/austprescr.2019.002
40. Sheu CC, Chang YT, Lin SY, Chen YH, Hsueh PR. Infections caused by carbapenem-resistant Enterobacteriaceae: an update on therapeutic options. Front Microbiol. (2019) 10:80. doi: 10.3389/fmicb.2019.00080
41. Ramirez MS, Tolmasky ME. Amikacin: uses, resistance, and prospects for inhibition. Molecules. (2017) 22:2267. doi: 10.3390/molecules22122267
42. Castellanos LR, Donado-Godoy P, León M, Clavijo V, Arevalo A, Bernal JF, et al. High heterogeneity of Escherichia coli sequence types harbouring ESBL/AmpC genes on IncI1 plasmids in the Colombian poultry chain. PLoS ONE. (2017) 12:e170777. doi: 10.1371/journal.pone.0170777
43. Vinueza-Burgos C, Cevallos M, Ron-Garrido L, Bertrand S, De Zutter L. Prevalence and diversity of Salmonella serotypes in ecuadorian broilers at slaughter age. PLoS ONE. (2016) 11:e159567. doi: 10.1371/journal.pone.0159567
44. Salazar GA, Guerrero-López R, Lalaleo L, Avilés-Esquivel D, Vinueza-Burgos C, Calero-Cáceres W. Presence and diversity of Salmonella isolated from layer farms in central Ecuador: [Version 2; peer review: 2 approved]. F1000Research. (2019) 8:1–12. doi: 10.12688/f1000research.18233.2
45. Zurita J, Ortega-Paredes D, Barba P. First description of Shigella sonnei Harboring bla(CTX-M-55) outside Asia. J Microbiol Biotechnol. (2016) 26:2224–7. doi: 10.4014/jmb.1605.05069
46. Bonnet R. Growing group of extended-spectrum B-lactamases: the CTX-M enzymes. Antimicrob Agents Chemother. (2004) 48:1–14. doi: 10.1128/AAC.48.1.1
47. Gharaibeh MH, Shatnawi SQ. An overview of colistin resistance, mobilized colistin resistance genes dissemination, global responses, and the alternatives to colistin: a review. Vet World. (2019) 12:1735–46. doi: 10.14202/vetworld.2019.1735-1746
48. Cantón R, Ruiz-Garbajosa P. Co-resistance: an opportunity for the bacteria and resistance genes. Curr Opin Pharmacol. (2011) 11:477–85. doi: 10.1016/j.coph.2011.07.007
49. Ortega-Paredes D. High quantities of multidrug-resistant Escherichia coli are present in the Machángara urban river in Quito, Ecuador. J Water Health. (2020) 18:67–76. doi: 10.1348/000712610X493494
50. Lazarus B, Paterson DL, Mollinger JL RB. Do human extraintestinal Escherichia coli infections resistant to expanded-spectrum cephalosporins originate from food-producing animals? A systematic review. Clin Infect Dis. (2014) 1:439–52. doi: 10.1093/cid/ciu785
51. Saliu E-M, Vahjen W, Zentek J. Types and prevalence of extended–spectrum beta–lactamase producing Enterobacteriaceae in poultry. Anim Heal Res Rev. (2017) 18:46–57. doi: 10.1017/S1466252317000020
Keywords: AmpC beta-lactamases, broiler farms, broiler carcasses, E. coli, extended-spectrum beta-lactamase (ESBL), human, mcr-1
Citation: Ortega-Paredes D, de Janon S, Villavicencio F, Ruales KJ, De La Torre K, Villacís JE, Wagenaar JA, Matheu J, Bravo-Vallejo C, Fernández-Moreira E and Vinueza-Burgos C (2020) Broiler Farms and Carcasses Are an Important Reservoir of Multi-Drug Resistant Escherichia coli in Ecuador. Front. Vet. Sci. 7:547843. doi: 10.3389/fvets.2020.547843
Received: 31 March 2020; Accepted: 29 October 2020;
Published: 25 November 2020.
Edited by:
Josefina Leon-Felix, Consejo Nacional de Ciencia y Tecnología (CONACYT), MexicoReviewed by:
Andrea Isabel Moreno Switt, Pontificia Universidad Católica de Chile, ChileCopyright © 2020 Ortega-Paredes, de Janon, Villavicencio, Ruales, De La Torre, Villacís, Wagenaar, Matheu, Bravo-Vallejo, Fernández-Moreira and Vinueza-Burgos. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Christian Vinueza-Burgos, Y3ZpbnVlemFAdWNlLmVkdS5lYw==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.