
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Vet. Sci. , 10 December 2020
Sec. Parasitology
Volume 7 - 2020 | https://doi.org/10.3389/fvets.2020.00527
This article is part of the Research Topic Antiparasitic Activity and the Modes of Action of Natural Products and Traditional Medicines View all 10 articles
 Julia M. Souza1
Julia M. Souza1 Érica A. A. de Carvalho1
Érica A. A. de Carvalho1 Ana Carolina B. B. Candido1Rafael P. de Mendonça2Maria Fernanda da Silva2
Ana Carolina B. B. Candido1Rafael P. de Mendonça2Maria Fernanda da Silva2 Renato L. T. Parreira1Fernanda G. G. Dias2
Renato L. T. Parreira1Fernanda G. G. Dias2 Sérgio R. Ambrósio1Andrea T. Arantes3Ademar A. da Silva Filho4
Sérgio R. Ambrósio1Andrea T. Arantes3Ademar A. da Silva Filho4 Aline N. Nascimento5
Aline N. Nascimento5 Monique R. Costa5
Monique R. Costa5 Mirela I. Sairre5
Mirela I. Sairre5 Rodrigo C. S. Veneziani1*
Rodrigo C. S. Veneziani1* Lizandra G. Magalhães1,2*
Lizandra G. Magalhães1,2*The efficacy of Licochalcone A (LicoA) and its two analogs were reported against Leishmania (Leishmania) amazonensis and Leishmania (Leishmania) infantum in vitro, and in experimental model of L. (L.) infantum in vitro. Initially, LicoA and its analogs were screened against promastigote forms of L. (L.) amazonensis. LicoA was the most active compound, with IC50 values of 20.26 and 3.88 μM at 24 and 48 h, respectively. Against amastigote forms, the IC50 value of LicoA was 36.84 μM at 48 h. In the next step, the effectivity of LicoA was evaluated in vitro against promastigote and amastigote forms of L. (L.) infantum. Results demonstrated that LicoA exhibited leishmanicidal activity in vitro against promastigote forms with IC50 values of 41.10 and 12.47 μM at 24 and 48 h, respectively; against amastigote forms the IC50 value was 29.58 μM at 48 h. Assessment of cytotoxicity demonstrated that LicoA exhibited moderate mammalian cytotoxicity against peritoneal murine macrophages; the CC50 value was 123.21 μM at 48 h and showed about 30% of hemolytic activity at concentration of 400 μM. L. (L.) infantum-infected hamsters and treated with LicoA at 50 mg/kg for eight consecutive days was able to significantly reduce the parasite burden in both liver and spleen in 43.67 and 39.81%, respectively, when compared with negative control group. These findings suggest that chalcone-type flavonoids can be a promising class of natural products to be considered in the search of new, safe, and effective compounds capable to treat canine visceral leishmaniosis (CVL).
Leishmania parasites are the etiological agents that cause leishmaniasis, and the parasites are transmitted by the bite of infected female phlebotomine sandflies (1). Human disease is classified into three main clinical forms: cutaneous leishmaniasis (CL), mucocutaneous leishmaniasis (MCL), and visceral leishmaniasis (VL) (1). CL is the most common form of the disease, and the manifestations can depend of the parasite species and host immune responses (2). Leishmania (Viannia) braziliensis and Leishmania (Leishmania) amazonensis are the main species responsible by CL in Brazil, and also by diffuse cutaneous leishmaniasis (DCL) (2). On the other hand, VL exists in two forms, zoonotic and anthroponotic, which are caused by Leishmania (Leishmania) infantum and Leishmania (Leishmania) donovani, respectively (3). Among the different mammal species, dogs can be considered as the main reservoirs of L. (L.) infantum, and this parasite also is responsible by canine visceral leishmaniasis (CVL) (4). Due the extension of CVL in urban regions in South American countries, this parasite has the attention of public health and the scientific community (4, 5).
The chemotherapy of leishmaniasis on human depends on three drugs, pentavalent antimonials, such as meglumine antimoniate (MA), amphotericin B (AmpB), and miltefosine (6). These drugs have high cost, limited efficacy, long administration protocols, severe side-effects, and development of drug resistance upon repeated use. In relation on CLV, miltefosine is an alternative for treatment of animals in regions where this drug is not used in humans; however, studies shown that miltefosine reduces the parasitic load of dogs infected with L. (L.)infantum, but no parasitological cure (4, 7, 8).
Licochalcone A (LicoA, 1) (Figure 1) is a naturally occurring chalcone-type flavonoid isolated from roots of the Chinese licorice (Glycyrrhiza inflata) that can be considered an antiparasitic hit compound (9). It can chemically describe as an α,β-unsaturated bisphenylic ketone (chalcone skeleton) substituted with two phenolic hydroxyl groups, one methoxy moiety, and an isoprenoid side chain. Previous works described the significant in vitro and in vivo effect of LicoA against Leishmania (L.) major and L. (L.) donovani, which are, respectively, associated with the CL and VL forms (10, 11). Another chalcone-type flavonoid compound from licorice roots extracts is Echinatin (2) (Figure 1). It has been reported to have a broad range of bioactivities, such as hepatoprotective (12) anti-inflammatory, antioxidant (13, 14), antimicrobial (15), and cardioprotective (16).
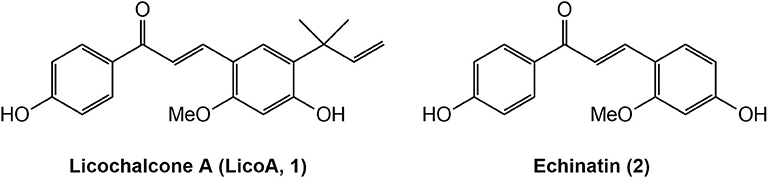
Figure 1. Structures of natural products Licochalcone A (LicoA 1) and Echinatin (2).
Considering that LicoA has demonstrated its potential against other forms of the parasite and that other analogs of LicoA could also present promising results, the aim of this study is to evaluate the in vivo leishmanicidal effect of LicoA and its two analogs against L. (L.) amazonensis and Leishmania (L.) infantum in vitro and in an experimental model of Leishmania (Leishmania) infantum in hamster. The analogs were synthesized and constitute structures similar to the natural products LicoA (1) and also Echinatin (2) (Figure 1). The analog LLA1 (1a) (Figure 2) is a derivate of LicoA that belongs to the class of chalcones containing the group O-prenil (17). In another study, the evaluation of a series of prenyloxy and geranyloxy chalcones against Leishmania (Leishmania) mexicana and Trypanosoma cruzi demonstrated that the position of the substituent has an influence on the activity and selectivity of these compounds (18). The antiparasitic activity of the analog LLA2 (2a) (Figure 2) has not been investigated yet.
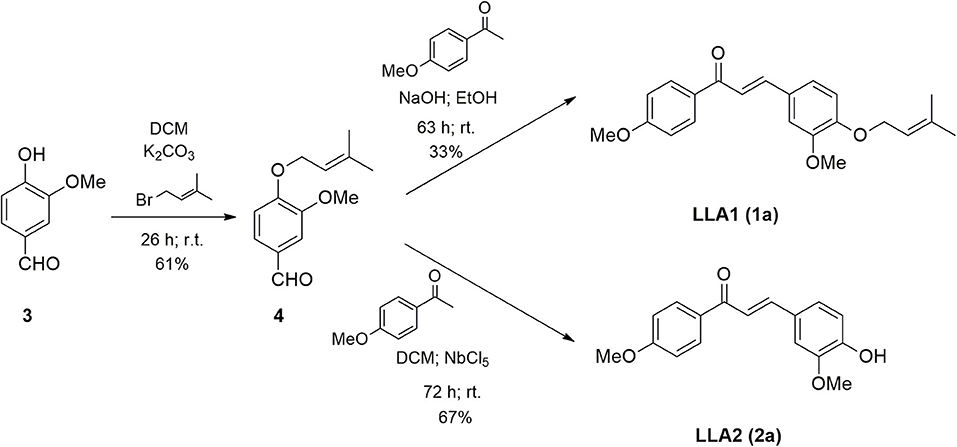
Figure 2. Scheme of synthesis of LLA1 (1a) and LLA2 (1a).
The reagents and solvents used in the synthesis of the analogs of LicoA (LLA1 1a and LLA2 2a) were obtained from commercial sources and used directly without further purification. NbCl5 was supplied by Companhia Brasileira de Metalurgia e Mineração (CBMM). Thin layer chromatography (TLC) was performed on Sigma-Aldrich silica gel matrix, pre-coated plates with fluorescent indicator 254 nm (Sigma-Aldrich, St Louis, USA). 1H and 13C NMR spectra were recorded on Varian equipment (500 MHz) from Federal University of ABC or Bruker equipment (DPX-300 and DPX-400) from University of São Paulo, Ribeirão Preto-SP, using the solvent deuterated chloroform (CDCl3) or dimethylsulfoxide (DMSO-d6). HRMS were obtained in instrument micrOTOFfrom University of São Paulo, Ribeirão Preto-SP. AmpB was purchased from Sigma-Aldrich, and MA (Glucantime®) was obtained from Sanofi-Aventis (Sanofi-Aventis, São Paulo, Brazil), which each 5 mL of aqueous solution contained 1.5 g of MA and represented 405 mg of pentavalent antimonial (Sanofi-Aventis).
The isolation and identification of LicoA was performed as previously described (19) and its chemical structure was identified by 1H- and 13C-NMR analysis data in comparison with the literature (20). Purity of LicoA (1) was estimated to be higher than 95% by both 13C NMR and HPLC analysis using different solvent systems.
The potassium carbonate was previously macerated with a mortar and pistil. Vanillin (3) (0.4526 g; 3 mmol), potassium carbonate (1.6970 g; 12 mmol), prenyl bromide (0.353 mL, 3 mmol), tetrabutyl ammonium chloride (0.0816 g; 0.3 mmol), and dichloromethane (DCM) were added to a round-bottom flask. The reaction was kept under stirring and at room temperature. The reaction was monitored by TLC until the practically total consumption of vanillin was verified (26 h). Then, the material was dissolved in dichloromethane and transferred to a separating funnel. The organic layer was separated and the aqueous layer extracted twice with dichloromethane. The combined organic extracts were washed with brine and the solution obtained was dried over anhydrous magnesium sulfate, filtered, and the solvent was evaporated under reduced pressure. The crude product obtained was purified over silica gel column chromatography using the mixture of 80% n-hexane in ethyl acetate as eluent to yield 0.4007 g of the pure product (61%) of 4. 1H-RMN (CDCl3, 300 MHz) δ: 1.75 (d, 3H, J = 1.2 Hz); 1.80 (d, 3H, J = 1.2 Hz); 4.58 (d, 2H. J = 6.7 Hz); 5.48 (tsept, 1H, J = 6.7 Hz; J = 1.2 Hz); 7.00 (d, J = 8.7 Hz); 7.82 (d, J = 8.7 Hz); 9.87 (s, 1H). 13C-RMN (CDCl3, 75 MHz) δ: 18.18 (CH3), 25.72 (CH3); 65.17 (CH2); 114.95 (CH); 118.87 (CH); 129.86 (C); 131.88 (CH); 138.89 (C); 163.98 (C), 190.64 (CH).
To a round-bottom flask, sodium hydroxide (0.0462 g; 0.9 mmol), ethanol (4 mL), and p-methoxyacetophenone (0.0754 g; 0.45 mmol) were added. The mixture was kept in an ice bath for 20 min. Then, the prenylated vanillin (4) obtained in the previous reaction (0.1001 g; 0.45 mmol) was added and the temperature was spontaneously raised to room temperature. The reaction was kept under stirring at room temperature until the total reagent consumption was observed by TLC (63 h). Ethanol was removed under reduced pressure. Then the material was dissolved in ethyl acetate and transferred to a separatory funnel. The aqueous phase was extracted twice with ethyl acetate. The combined organic phases were washed with brine. The organic extract was dried over anhydrous magnesium sulfate, filtered, and the solvent was evaporated under reduced pressure. The crude product obtained was purified over silica gel column chromatography using the mixture of 30% ethyl acetate in n-hexane as the eluent to yield 0.0498 g of the pure product (33%) of 1a as a yellow powder. 1H-RMN (CDCl3, 500 MHz) δ: 1.76 (d, 3H, J = 1.0 Hz); 1.80 (d, 3H, J = 1.0 Hz); 3.89 (s, 3H); 3.94 (s, 3H); 4.64 (d, 2H, J = 6.8 Hz); 5.52 (thept, 1H. J = 6.8 Hz; J = 1.0 Hz); 6.90 (d, 1H, J = 8.2 Hz); 6.99 (d, 2H, J = 8.8 Hz); 7.17 (d, 1H, J = 2.1 Hz); 7.21 (dd, 1H, J = 8.2 Hz; J = 2.1 Hz); 7.41 (d, 1H, J = 15.6 Hz); 7.76 (d, 1H, J = 15.6 Hz); 8.04 (d, 2H, J = 8.8 Hz). 13C-RMN (CDCl3, 125 MHz) δ: 18.28 (CH3); 25.84 (CH3); 55.47 (CH3); 55.97 (CH3); 65.77 (CH2); 110.27 (CH); 112.58 (CH); 113.77 (CH); 119.42 (CH); 119.66 (CH); 122.87 (CH); 127.91 (C); 130.71 (CH); 131.34 (C); 138.19 (C); 144.27 (CH); 149.55 (C); 150.62 (C); 163.25 (C); 188.81 (C).
To a round bottom flask fitted with a drying tube were added niobium pentachloride (0.8998 g; 1.35 mmol) and dichloromethane (10 mL). Then, p-methoxyacetophenone (0.1346 g, 0.9 mmol) and prenylated vanillin (0.2085 g, 0.9 mmol) were added. The reaction mixture was stirred at room temperature for 72 h. After the period, an extraction with dichloromethane was carried out, then the solvent was eliminated under reduced pressure and the crude residue was purified over silica gel column chromatography using the mixture of n-hexane and ethyl acetate (2:1) as eluent to yield 0.1814 g (67%) of 2a. 1H NMR (CDCl3, 500 MHz) δ: 3.87 (s, 3H); 3.93 (s, 3H); 6.95 (d, 1H, J = 8.3 Hz); 6.97 (d, 2H, J = 9.0 Hz); 7.12 (d, 1H, J = 1.9 Hz); 7.20 (dd, 1H, J = 1.9 Hz; J = 8.3 Hz); 7.39 (d, 1H, J = 15.4 Hz); 7.74 (d, 1H, J = 15.4 Hz); 8.03 (d, 2H, J = 9.0 Hz). 13C NMR (CDCl3, 125 MHz) δ: 55.46 (CH3); 55.97 (CH3); 110.11 (CH); 113.78 (CH); 114.92 (CH); 119.42 (CH); 123.16 (CH); 127.59 (C); 130.73 (CH); 131.27 (C); 144.46 (CH); 146.88 (C); 148.22 (C); 163.28 (C); 188.90 (C). HRMS m/z [M+H]+ calcd: 285.1121; found: 285.1124.
Male Balb/c mice (Mus musculus) were acquired from the animal houses of the University of São Paulo (Ribeirão Preto, São Paulo, Brazil) and male golden hamsters (Mesocricetus auratus) (110 g) were acquired from ANILAB–Laboratory Animal Creation and Trade Ltd. (Paulínia, São Paulo, Brazil). The experiments were conducted in accordance with the Brazilian legislation regulated by the National Council for the Control of Animal Experimentation (CONCEA) and approved by the University of Franca's Ethics Committee for Animal Care under protocol number 046/15 (Approval Date: November 09, 2015).
L. (L.) amazonensis (IFLA/BR/67/PH8.) was maintained as promastigote forms in Roswell Park Memorial Institute medium (RPMI 1640 medium) (Gibco, Grand Island, NY, USA), supplemented with antibiotics (penicillin 10.000 UI/mL and streptomycin 10 mg μg/mL) (Cultilab, Campinas, BR), and 10% bovine fetal serum (FBS) (Cultilab), at 25°C. L. (L.) infantum (MHOM/BR/1972/LD) was maintained as promastigote forms in M199 medium (Gibco), supplemented with 10% FBS (Cultilab), antibiotics (Cultilab), 5% human urine and 0.25% hemin (Sigma-Aldrich) at 25°C, and amastigote forms were obtained from the spleen of golden hamsters after 60 to 70 days of infection (21). Peritoneal murine macrophages were obtained by washing the peritoneal cavity of BALB/c mice with RPMI 1640 medium (Gibco) after 72 h of the application of 5 mL of the sodium thioglycolate (Sigma-Aldrich) at 3%, and then were cultivated in RPMI 1640 medium (Gibco) supplemented with antibiotics and 10% FBS (Gibco), at 37°C in a 5% CO2 humidified incubator.
Initially, a screening was performed against promastigote forms of L. (L.) amanzonensis. Briefly, promastigotes (1 × 106 parasites/well) were distributed in 96-well plates, and the compounds LicoA (1), LLA1 (1a), LLA2 (2a), and MA were added at concentrations 0.78–400.0 μM, and AmpB was added at concentrations 0.0025–1.56 μM. The plates were incubated at same conditions previously described during 48 h, and the leishmanicidal activity was determined by the inhibition of growth of the promastigote forms by counting the total number of live promastigote in Neubauer's chamber (Global Glass, São Paulo, BR) using an optical microscope (Nikon New York, USA). The compound (LicoA, 1) that exhibited activity against promastigote forms of L. (L.) amazonensis also was evaluated against promastigote forms of L. (L.) infantum as previously described and against amastigote forms of both Leishmania parasites. To evaluate the leishmanicidal activity against amastigote forms, peritoneal murine macrophages were seemed (2 × 105 cells per well) in 24-well plates on slide chambers and incubated at 37°C for 24 h. After the 24 h, the cells were infected with promastigote forms of L. (L.) amazonensis, previously acidified (22) or with amastigote forms of L. (L.) infantum at a ratio of 10:1 (parasites/macrophage) for 4 h, and subsequently the cells were incubated with the LicoA (1) (6.25–100 μM), AmpB (0.095–0.005 μM), or MA (12.5–400 μM) 48 h. The slides were stained with Giemsa (Synth, Diadema, BR), analyzed using an optical microscopy (Nikon), and the parasite load was defined by the number of infected macrophages X number of intracellular amastigotes/number of total macrophages (23). Parasites cultured in medium with 0.1% were used as negative control and parasites cultured in medium with MA or AmpB were used as positive controls.
Peritoneal murine macrophages were seemed (2 × 105 cells per well) in 96-well plates and LicoA (1), AmpB, and MA were added at concentrations 12.5–400 μM. The cells were incubated at 37°C in a 5% CO2 humidified incubator at 37°C during 48 h, and the cytotoxic activity was determined using MTT assay (23). Cells cultured in medium with 0.1% DMSO were used as negative control and cells cultured in medium with 25% DMSO were used as positive control.
Defibrinated sheep blood (Newprov, Pinhais, PR, BR) was diluted in 0.9% saline solution and a suspension at 3% of erythrocytes was transferred (100 μL) in 96-well plates. LicoA (1) and AmpB were added (12.5–400 μM), and the plates were incubated for 30 min at 37°C. The hemolytic activity was determined in the cell supernatant by optical density reading at 415 nm (Libra S12–Biochrom–Cambridge, RU) Distilled water was used as a positive control, and 0.9% saline solution with 0.1% DMSO was used as negative control.
The in vivo leishmanicidal activity of LicoA (1) was assessment in experimental model of Leishmania (L.) infantum and conducted as described by (21) with adaptations. Male golden hamsters were infected by intraperitoneal route with 1.0 × 108 L. (L.) infantum amastigotes. Four weeks after infection, the animals were randomly separated in 4 groups of 6 animals each. Two groups of treated animals received intraperitoneal doses of 20 and 50 mg/kg of body weight of LicoA (1), respectively, for eight consecutive days. Negative control group received the same number of injections of phosphate buffered saline (PBS) and animals from positive control group was treated with MA (Sanofi-Aventis) at dose 50 mg/kg of body weight during eight consecutive days by intraperitoneal route. Thirty five days after treatment the animals were sacrificed and the parasite burden was evaluated both in spleen and liver by limiting dilution method (24).
In order to evaluate the hepatotoxicity, groups with 6 male golden hamsters no-infected were treated intraperitoneally with LicoA (1) (20 mg/kg or 50 mg/kg of body weight per day) or MA (50 mg/kg of body weight per day) for 15 days. After this period, serum concentrations of alanine aminotransferase (ALT) and aspartate aminotransferase (AST) were determined using sets of commercial reagents (Labtest, Minas Gerais, Brazil). Negative control group received the same number of injections of PBS.
The analyses were performed using GraphPad Prism 7 program (GraphPad Software, San Diego, CA, USA). Both IC50 and CC50 values were calculated with the aid of sigmoid dose–response curves, and the selectivity index (SI) values were determined by CC50/IC50. In vitro experiments were performed in triplicate and repeat two times. In vivo, statistical analysis was performed using one-way ANOVA followed by Dunnett's multiple comparison test.
According to Figure 2, the chalcone containing O-prenyl (LLA) was synthesized in two steps. The vanillin prenylation reaction was carried out under conditions of phase transfer catalysis (PTC) that allows the use of aqueous potassium carbonate solution and a transfer catalyst (25). The most commonly used catalysts for this type of reaction are quaternary ammonium salts. Tetrabutyl ammonium chloride (TBAC) was used because it has large substituent groups (n-butyl) that favor the reaction. Despite the long reaction time, the procedure was simple, at room temperature and product 4 was obtained with a yield of 61%. The prenylated chalcone (LLA1) (1a) was prepared via the Claisen-Schmidt condensation between the prenylated vanillin and commercial p-methoxyacetophenone. The reaction was carried out under basic conditions and the yield of the product LLA1 (1a) was 33% after purification. The synthesis of LLA2 (2a) was carried out through the Claisen-Schmidt condensation by acid catalysis (Figure 2). The reaction between the prenylated vanillin and the p-methoxyacetophenone promoted the formation of chalcone, together with the hydrolysis of the prenyl group in the presence of Lewis acid (niobium pentachloride), providing the product 2a with 67% yield, after purification by chromatography on silica gel column. The use of niobium pentachloride as an acid catalyst is an alternative for the synthesis of chalcones through Claisen-Schmidt.
Initially, the leishmanicidal activity of LicoA (1) and analogs LLA1 (1a) and LLA2 (2a) was evaluated against promastigotes, and the active compound was subsequently evaluated against amastigote forms of L. (L.) amazonensis. LicoA (1) exhibited promising leishmanicidal activity in vitro with IC50 values of 20.26 and 3.88 μM at 24 and 48 h, respectively, against promastigote and IC50 value of 36.84 against amastigote forms of L (L.) amazonensis. LLA1 (1a) was the second most active against promastigote forms with IC50 values of 74.94 and 67.16 μM at 24 and 48 h, and LLA2 (2a) exhibited modest activity in vitro against promastigote forms of L. (L.) amazonensis (IC50 values > 300 μM). AmpB, used as positive control against L. (L.) amazonensis, exhibited IC50 values against promastigote forms of 0.27 and 0.065 μM at 24 and 48 h, respectively; and against amastigote forms the IC50 value was 0.022 at 48 h. As LicoA (1) was the most active, in the next step, the effectivity of LicoA (1) was evaluated in vitro against promastigote and amastigote forms of L. (L.) infantum. LicoA (1) exhibited leishmanicidal activity in vitro against promastigote forms with IC50 values of 41.10 and 12.47 μM at 24 and 48 h, respectively; against amastigote forms the IC50 value was 29.58 μM after 48 h. MA, used as positive control exhibited IC50 values > 400 μM in both promastigote and amastigote forms (Table 1).
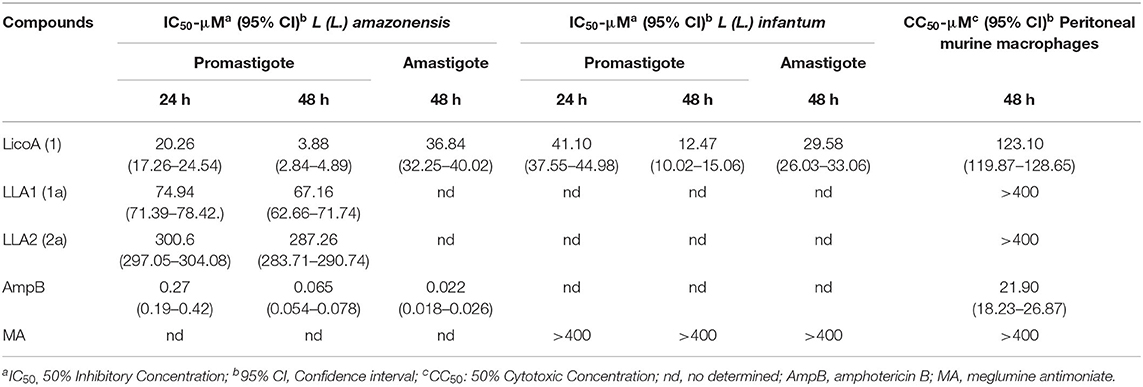
Table 1. Leishmanicidal activity and mammalian cytotoxicity of LicoA and its analogs.
LicoA (1) at concentrations evaluated exhibited moderate mammalian cytotoxicity against peritoneal murine macrophages, the CC50 values was 123.21 μM at 48 h. AmpB, exhibited mammalian cytotoxicity with CC50 values of 21.90 μM. On the other hand, MA showed no cytotoxicity at concentrations evaluated (Table 1). In the relationship between activity against intracellular amastigotes, and cytotoxicity in peritoneal murine macrophages, it was possible to determine that selectivity index (SI) of the LicoA (1) was 3.85 to L (L.) amazonensis and 4.16 to L. (L) infantum (data not shown). The hemolytic activity also was evaluated, and LicoA (1) showed about 30% of hemolytic activity at highest concentration evaluated (400 μM). On the other hand, AmpB showed 60% of hemolysis at concentration 100 μM and at higher concentrations (≥ 200 μM) was observed 100% of hemolysis.
L. (L.) infantum-infected hamsters were treated during eight consecutive days at doses of 20 and 50 mg/kg of body weight per day of LicoA (1) 4 weeks after infection, and 35 days after treatment the animals were sacrificed. LicoA (1) at 20 and 50 mg/kg of body weight per day was able to significantly reduce the parasite burden in liver in 15.27 and 43.67%, respectively, and in spleen in 20.69 and 39.81%, respectively, when compared with negative control group. Positive control group that received 50 mg/kg of body weight per day of MA during eight consecutive days also showed significant reduction of parasite burden in 55.87% (liver) and of 63.57% (spleen) (Figures 3A,B). Besides, the average body weight animals showed no significant variation among negative, LicoA (1), and MA groups. However, after the treatment, 100% survival rate was registered in LicoA and MA groups, but in the negative group, the mortality of 33% of animals was observed (data not shown). Also, the animals no-infected and treated intraperitoneally at doses 20 and 50 mg/kg of body weight per day of LicoA (1) showed no significant alterations in the serum levels of aminotransaminases when compared with negative control group, but in the MA group, a significant increase in the serum levels of aminotransaminases was observed (Figures 4A,B).
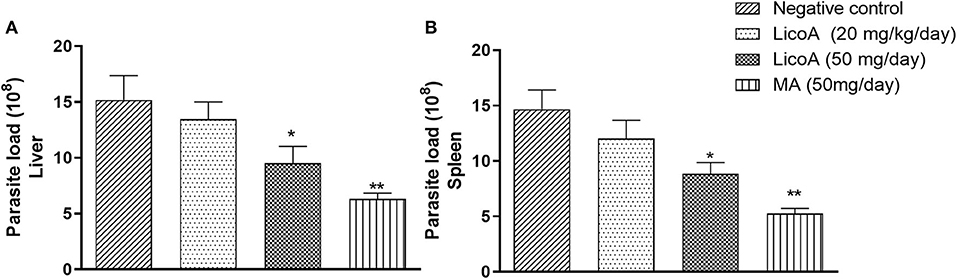
Figure 3. Liver (A) and spleen (B) parasite load of L. (L.) infantum-infected hamsters treated intraperitoneal with 20 and 50 mg/kg of body weight of LicoA (1) during eight consecutive days 4 weeks after infection. Thirty-five days after treatment the animals were sacrificed and the parasite burden was evaluated both in spleen and liver by limiting dilution method. Negative control group received the same number of injections of phosphate buffered saline (PBS) and animals from positive control group was treated with meglumine antimoniate (MA) (Glucantime®-Sanofi-Aventis) at dose 50 mg/kg of body. An asterisk indicates statistically significant differences as compared to the negative control group (PBS) (*p < 0.05, **p < 0.01).
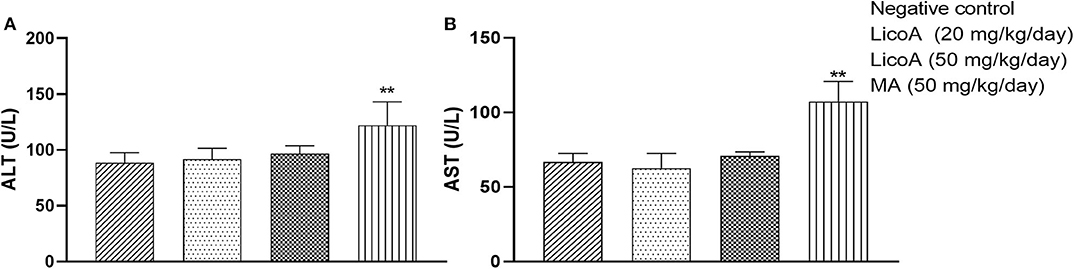
Figure 4. Serum concentrations of aminotransaminases ALT (alanine aminotransferase) (A) and AST (aspartate aminotransferase) (B) in no-infected hamsters and treated intraperitoneal with 20 and 50 mg/kg of body weight per day of LicoA (1) or treated with meglumine antimoniate (MA) (Glucantime®-Sanofi-Aventis) at dose 50 mg/kg of body weight per day during 15 days. Negative control group received the same number of injections of phosphate buffered saline (PBS) and animals from positive control group. An asterisk indicates statistically significant difference as compared to the negative control group (PBS) (**p < 0.01).
Secondary metabolites from plants have been demonstrated promising leishmanicidal activity in vitro and in vivo, including compounds from chalcone class (8). Then, our decision to investigate LicoA (1) and its two analogs [LLA1 (1a), LL2 (2a)] was triggered by our previous study which demonstrated that LicoA (1) possess leishmanicidal activity against L. (L.) major and L. (L.) donovani in vitro and in vivo, which are parasites that cause CL and VL, respectively (10, 11). Besides, a search of the literature reports that LicoA (1) is of interest due to be inexpensive. Thus, this information encouraged us to evaluate the effect of LicoA and its two analogs against L. (L.) amazonensis and L. (L.) infantum, which are parasites involved in the DCL and CVL, respectively.
In this study, the synthesis of LLA1 and LLA2 was performed. As described, the prenylated chalcone LLA1 (1a) was prepared via the Claisen-Schmidt condensation between the prenylated vanillin and commercial p-methoxyacetophenone (Figure 2). The yield of this reaction was low; however, the difficulty in preparing prenylated chalcones was also found in another study that reports 16% yield for condensation of prenylated benzaldehydes (26). The analog LLA2 (2a) was obtained when the Claisen-Schmidt condensation between the prenylated vanillin and the p-methoxyacetophenone was performed using Lewis acid niobium pentachloride in dichloromethane. Niobium pentachloride promoted the formation of chalcone and hydrolysis of the prenyl group. Niobium pentachloride (NbCl5) has a wide variety of applications in organic synthesis, for example, in Diels-Alder reactions, multicomponent reactions, and polymerization reactions. In addition, niobium compounds are more readily available in Brazil due to the presence of the largest niobium reserves in the world (27). The obtained result shows that niobium pentachloride can be an efficient alternative for the synthesis of chalcones.
Among the compounds evaluated, LicoA (1) was the most promising compound against Leishmania parasites under the in vitro experimental conditions used. In general, compounds with IC50 values <10 μM in vitro in infective protozoan forms are considered as a hit and lead candidates for further in vivo evaluations (28). Thus, this study demonstrated that LicoA (1) possess moderate activity against amastigote forms of L (L.) amazonensis and L. (L.) infantum. On the other hand, previous studies demonstrated that LicoA (1) inhibits the in vitro growth of L. (L.) major and L. (L.) donovani promastigotes with IC50 value of ~0 μM for both parasites in 72 h, and a reduction in the number of amastigotes was observed at 2.5 μM after 72 h of incubation with LicoA (1) (10, 11). It should be noted that promastigote forms were more susceptible to LicoA (1) than amastigotes forms, and this effect can be attributed by distinct metabolism effects, as previous described in the literature (23).
By comparing the leishmanicidal activity observed to LicoA (1) with the positive control, the results demonstrated that promastigote and amastigote forms of L (L.) amazonensis were susceptible to AmpB. In contrast, promastigote and amastigote forms of L. (L.) infantum were not susceptible to the positive control, MA at conditions evaluated in this study. This results reinforces the hypothesis that pentavalent antimonials, such as MA act as prodrugs that require biological reduction to the trivalent to exert anti-leishmanial activity (29, 30). Although MA did not show leishmanicidal activity in vitro, this compound was choice to positive control in vivo assay, and then was evaluated under in vitro conditions. Additionally, we choice MA rather than sodium stibogluconate (SSG) because previous study has demonstrated that MA can be more effective than SSG in the treatment of leishmaniosis (31). As the experiments were performed in different periods, MA was not used as positive control against L. (L.) amazonensis in vitroI; however, pentavalent antimonials are considered first line agents in the therapy against leishmaniasis (6).
Despite of the moderate cytotoxic effect of LicoA (1) observed in our study, previous study has demonstrated that LicoA (1) shows low-toxicity in HFF cell in vitro (32). Also, study has demonstrated that LicoA (1) induces hemolysis to trigger cell shrinkage and phospholipid scrambling of the cell membrane at concentrations higher than 30 μM after 24 h (33).
Although the results in vitro have been better against L. (L.) amazonensis, the in vivo assay was performed against Leishmania (L.) infantum because this parasite is responsible by CVL, and also dogs are the main urban domestic reservoir of VL (3, 4). Several advances in the treatment of CVL have been made in the last years, but the pharmacotherapeutic options are still not satisfactory (5–7, 34). MA and allopurinol (alone or in combination) are drugs for CVL treatment and are able to achieve clinical cure (35). Other leishmanicidal drugs such as miltefosine in combination with allopurinol have also demonstrated leishmanicidal efficacy in infected dogs (36). Unfortunately, drug treatment cannot impede disease transmission since treated animals remain carriers of the parasite (5, 7, 34–36). Moreover, other aspects like toxicity and the rise of Leishmania strains that are resistant to the current use drugs highlights the importance of the development of new, safe and effective compounds capable to treat CVL and actually to eliminate the infection (5, 21, 34–36). Then, the in vivo activity of LicoA (1) was evaluated in experimental model of Leishmania (L.) infantum in hamster. The doses of LicoA (1) and the scheme of treatment were based on previous reports and also on dose of MA (11, 21). The results demonstrated a reduction of parasite burden in both liver and spleen to hamsters infected with L. (L.) infantum after treatment with LicoA (1). The positive control group, MA, showed better results than compared with LicoA (1), but both compounds significantly reduced the parasite burden. Previous study has shown that BALB/c mice infected with L. (L.) major and treated with LicoA (1) at doses 2.5 and 5 mg/kg of body weight by intraperitoneal administration 7 days after infection during 39 days completely prevented lesion development (11). Also, it was observed that the treatment of hamsters infected with L. (L.) donovani with intraperitoneal administration of LicoA (1) at a dose of 20 mg/kg of body weight per day for six consecutive days resulted in a >96% reduction of parasite load in the liver and the spleen compared with untreated control (11). However, our results demonstrated that at a dose of 20 mg/kg of body weight per day resulted in a reduction of parasite load of the 15.25 and 20.69% in liver and spleen, respectively. The intrinsic and/or acquired susceptibility of the parasite species can have contribute to differences of LicoA (1) susceptibility among the Leishmania species in vitro and in vivo when compared with the effect of LicoA (1) against L. (L.) donovani and L. (L.) major (10, 11). Previous study has demonstrated that intrinsic variation in miltefosine susceptibility of Leishmania clinical isolates was observed for visceral leishmaniasis in Nepal and cutaneous leishmaniasis in Peru (36). Another study demonstrated intraspecies differences in natural susceptibility to AmpB of clinical isolates of Leishmania subgenus Viannia (37). In this same study, it was demonstrated that isolates and strains maintained in the laboratory were less sensitive to AmpB when compared to clinical isolated (37).
No evidence of hepatotoxicity was found since serum concentrations of aminotransaminases showed no changed between LicoA-treated and non-treated groups. Previous study also demonstrated that LicoA (1) at doses higher than 1,000 mg/kg, administered orally once a day for 2 weeks did not cause any signs of toxicity. Also, intraperitoneal administration of LicoA (1) at doses of 100 mg/kg in rats and 150 mg/kg in hamsters did not show any toxicity (11). In addition, study has been shown that LicoA (1) has a protective effect against Acetaminophen overdose-induced hepatotoxicity (38). These findings suggest the safety of LicoA (1) in animal model independently of the route of administration. Notwithstanding positive control group (animal treated with MA) has showed better results than compared with LicoA (1) at dose 50 mg/kg of body weight per day, pentavalent antimonials cause serious side effects (4, 7, 8). Also, the MA administration can not be administered by the oral route due irritation of intestinal mucosa (39), but LicoA (1) could be evaluated by the oral route in the future research in L. (L.) infantum-infected hamsters.
About the action mechanism, studies suggest that the effect of LicoA may be due to the altering of both ultrastructure and function of the mitochondria of Leishmania parasites (40). Moreover, studies involving in silico approaches also reinforced the antileishmanial potential of LicoA by denoting its capability to inhibit the NADPH-dependent fumarate reductase, which is the key enzyme in the fumarate respiration. This anaerobic process is essential for the growth and survival of the parasite in their host, but is absent in the normal cells of the mammalian hosts (41, 42). In addition, studies of the modeling and docking suggest that LicoA is a potent inhibitor of cysteine proteases A, which are the essential virulence factors in leishmania parasitic (43–45). Also, study has been shown that chalcones act against malarial papain-like cysteine proteases, which is an important mechanism of the hemoglobin degradation (46).
Given in consideration that CVL manifests through splenomegaly possibly associated with hepatomegaly, and keeping in mind the lack of hepatoxicity of LicoA (1) and its in vitro effect against L (L.) amazonensis and L (L.) infantum, and in vivo effect in hamster-infected with L (L.) infantum, further studies to address the effects LicoA (1) and other chalcones on pathological changes associate to leishmaniosis are crucial. In addition, the results suggest that chalcone-type flavonoids are a promising class of natural products to be considered in the search of new compounds capable to treat CVL, and the association of such compounds with nanostructured carriers may lead to formulations that can increase the efficacy, selectivity, and safety of the CVL treatment.
The raw data supporting the conclusions of this article will be available by request from the corresponding author.
The experiments were conducted in accordance with the Brazilian legislation regulated by the National Council for the Control of Animal Experimentation (CONCEA) and approved by the University of Franca's Ethics Committee for Animal Care under protocol number 046/15 (Approval Date: November, 09, 2015).
JS, EC, AC, MF, RM, and FD carried out the biological experiments and maintenance of Leishmania parasites. RP, AN, and MC contributed to the isolation, synthesis, and characterization of the compounds. SA, AS, and MS supervised the chemical part of the study. AA, LM, and RV supervised the work and provided the facilities for biological and chemical activities. All authors contributed to the article and approved the submitted version.
This study was supported by the National Council for Scientific and Technological Development, Brazil–CNPq (Fellowship: 303946/2018-0), Coordination for the Improvement of Higher Education Personnel, Brazil-CAPES (Finance code 001 and Fellowships: 88881.133783/2016-01 and 88882.36577/2019-01). The authors thank grants 2013/11164-4; 2016/18045-9; 2016/24456-1 from São Paulo Research Foundation, Brazil-FAPESP for the financial support.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors are also thankful to Dr. André F. Tempone from Instituto Adolfo Lutz (São Paulo, Brazil) for providing the Leishmania (L.) infantum strain used in this work. The authors would also like to thank CBMM for supplying niobium pentachloride samples and the Centagro–(Centro Tecnológico Agropecuário Ltda) for technical support.
1. World Health Organization. What is Leishmaniais. (2020). https://www.who.int/leishmaniasis/disease/en/ (accessed May 11, 2020).
2. Christensen SM, Belew AT, El-Sayed NM, Tafuri WL, Silveira FT, Mosser DM. Host and parasite responses in human diffuse cutaneous leishmaniasis caused by L. amazonensis. PLoS Negl Trop Dis. (2019) 13:e0007152. doi: 10.1371/journal.pntd.0007152
3. Jain K, Jain NK. Vaccines for visceral leishmaniasis: a review. J Immunol Methods. (2015) 422:1–12. doi: 10.1016/j.jim.2015.03.017
4. Werneck GL. Visceral leishmaniasis in Brazil: rationale and concerns related to reservoir control. Rev Saúde Pública. (2014) 48:851–55. doi: 10.1590/S0034-8910.2014048005615
5. Belo VS, Struchiner CJ, Werneck GL, Barbosa DS, de Oliveira RB, Neto RGT, et al. A systematic review and meta-analysis of the factors associated with Leishmania infantum infection in dogs in Brazil. Vet Parasitol. (2013) 195:1–13. doi: 10.1016/j.vetpar.2013.03.010
6. Akbari M, Oryan A, Hatam G Application of nanotechnology in treatment of leishmaniasis: a review. Acta Trop. (2017) 172: 86–90. doi: 10.1016/j.actatropica.2017.04.029
7. Dos Santos Nogueira F, Avino VC, Galvis-Ovallos F, Pereira-Chioccola VL, Moreira MAB, Romariz APPL, et al. Use of miltefosine to treat canine visceral leishmaniasis caused by Leishmania infantum in Brazil. Parasit Vectors. (2019) 12:79. doi: 10.1186/s13071-019-3323-0
8. Tajuddeen N, Isah MB, Suleiman MA, van Heerden FR, Ibrahim MA. The chemotherapeutic potential of chalcones against leishmaniases: a review. Int J Antimicrob Agents. (2018) 51:311–18. doi: 10.1016/j.ijantimicag.2017.06.010
9. Borsari C, Santarem N, Torrado J, Olías AI, Corral MJ, Baptista C, et al. Methoxylated 2'-hydroxychalcones as antiparasitic hit compounds. Eur J Med Chem. (2017) 126:1129–35. doi: 10.1016/j.ejmech.2016.12.017
10. Chen M, Christensen SB, Blom J, Lemmich E, Nadelmann L, Fich K, et al. Licochalcone A, a novel antiparasitic agent with potent activity against human pathogenic protozoan species of Leishmania. Antimicrob. Agents Chemother. (1993) 37:2550–56. doi: 10.1128/AAC.37.12.2550
11. Chen M, Christensen SB, Theander TG, Kharazmi A. Antileishmanial activity of licochalcone A in mice infected with Leishmania major and in hamsters infected with Leishmania donovani. Antimicrob Agents Chemother. (1994) 38:1339–44. doi: 10.1128/AAC.38.6.1339
12. Lin Y, Kuang Y, Li K, Whang S, Ji S, Chen K, et al. Nrf2 activators from Glycyrrhiza inflata and their hepatoprotective activities against CCl4-induced liver injury in mice. Bioorg Med Chem Lett. (2017) 25:5522–30. doi: 10.1016/j.bmc.2017.08.018
13. Fu Y, Chen J, Li YJ, Zheng YF, Li P. Antioxidant and anti-inflammatory activities of six flavonoids separated from licorice. Food Chem. (2013) 141:1063–71. doi: 10.1016/j.foodchem.2013.03.089
14. Liang M, Li X, Ouyang X, Xie H, Chen D. Antioxidant mechanisms of echinatin and licochalcone A. Molecules. (2019) 24:3–16. doi: 10.3390/molecules24010003
15. Haraguchi H, Tanimoto K, Tamura Y, Mizutani K, Kinoshita T. Mode of antibacterial action of retrochalcones from Glycyrrhiza inflata. Phytochemistry. (1998) 48:125–9. doi: 10.1016/S0031-9422(97)01105-9
16. Tian X, Liu C, Jiang HL, Zhang Y, Han J, Liu J, et al. Cardioprotection provided by Echinatin against ischemia/reperfusion in isolated rat hearts. BMC Cardiovascular Disorders. (2016) 16:119–27. doi: 10.1186/s12872-016-0294-3
17. Passalacqua TG, Dutra LA, de Almeida L, Arenas-Velásquez AM, Esteves-Torres FA, Yamasaki PR, et al. Synthesis and evaluation of novel prenylated chalcone derivatives as anti-leishmanial and anti-trypanosomal compounds. Bioorg Med Chem Lett. (2015) 25:3342–5. doi: 10.1016/j.bmcl.2015.05.072
18. Espinoza-Hicks JC, Chacón-Vargas KF, Hernandez-Rivera JL, Nogueda-Torres B, Tamariz J, Sanchez-Torres LE, et al. Novel prenyloxy chalcones as potential leishmanicidal and trypanocidal agents: design, synthesis and evaluation. Eur J Med Chem. (2019) 167:402–13. doi: 10.1016/j.ejmech.2019.02.028
19. Fontes L BA, Dos Santos Dias D, de Carvalho LSA, Mesquita HL, da Silva RL, Dias AT, et al. Immunomodulatory effects of licochalcone A on experimental autoimmune encephalomyelitis. J Pharm Pharmacol. (2014) 66:886–94. doi: 10.1111/jphp.12212
20. Wang QE, Lee FSC, Wang X. Isolation and purification of inflacoumarin A and licochalcone A from licorice by high-speed counter-current chromatography J Chromatogr A. (2004) 1048:51–7. doi: 10.1016/S0021-9673(04)01086-6
21. Salerno Pimentel IA, Paladi Cde S, Katz S, de Souza Júdice WA, Cunha RL, Barbiéri CL. In vitro and in vivo activity of an organic tellurium compound on Leishmania (Leishmania) chagasi. PLoS ONE. (2012) 7:e48780. doi: 10.1371/journal.pone.0048780
22. Leirião P, Rodrigues CD, Albuquerque SS, Mota MM. Survival of protozoan intracellular parasites in host cells. EMBO Rep. (2004) 5:1142–7. doi: 10.1038/sj.embor.7400299
23. Amaral M, de Sousa FS, Silva TAC, Junior AJG, Taniwaki NN, Johns DM, et al. A semi-synthetic neolignan derivative from dihydrodieugenol B selectively affects the bioenergetic system of Leishmania infantum and inhibits cell division. Sci Rep. (2019) 9:6114. doi: 10.1038/s41598-019-42273-z
24. Lima HC, Bleyenberg JA, Titus RG. A simple method for quantifying Leishmania in tissues of infected animals. Parasitol Today. (1997) 13:80–2. doi: 10.1016/S0169-4758(96)40010-2
25. Marzorati L, Barazzone GC, Bueno-Filho MA, Wladislaw B, Di Vitta C. New methodology for the preparation of N-tosyl aziridine-2-carboxylates. Tetrahedron Lett. (2007) 48:6509–13. doi: 10.1016/j.tetlet.2007.07.054
26. Kromann H, Larsen M, Boesen T, Schonning K, Nielsen SF. Synthesis of prenylated benzaldehydes and their use in the synthesis of analogues of licochalcone A. Eur J Med Chem. (2004) 39:993–1000. doi: 10.1016/j.ejmech.2004.07.004
27. Arpini BH, Bartolomeu AA, Andrade CKZ, Silva-Filho LC, Lacerda V. Recent advances in using niobium compounds as catalysts in organic chemistry. Current Organic Synthesis. (2015) 12:570–83. doi: 10.2174/157017941205150821125817
28. Katsuno K, Burrows JN, Duncan K, Hooft Van Huijsduijnen R, Kaneko T, Kita K, et al. Hit and lead criteria in drug discovery for infectious diseases of the developing world. Nat Rev Drug Discov. (2015) 14:751–8. doi: 10.1038/nrd4683
29. Miguel DC, Zauli-Nascimento RC, Yokoyama-Yasunaka JK, Katz S, Barbiéri CL, Uliana SR. Tamoxifen as a potential antileishmanial agent: efficacy in the treatment of Leishmania braziliensis and Leishmania chagasi infections. J Antimicrob Chemother. (2009) 63:365–68. doi: 10.1093/jac/dkn509
30. Borborema SET, Osso JA, Andrade HF Jr, Nascimento Nd. Biodistribution of meglumine antimoniate in healthy and Leishmania (Leishmania) infantum chagasi-infected BALB/c mice. Mem Inst Oswaldo Cruz. (2013) 108:623–30. doi: 10.1590/0074-0276108052013014
31. Yesilova Y, Surucu HA, Ardic N, Mksoy M, Yesilova A, Oghumu S, et al. Meglumine Antimoniate is more effective than sodium stibogluconate in the treatment of cutaneous leishmaniasis. J Dermatolog Treat. (2016) 27:83–87. doi: 10.3109/09546634.2015.1054778
32. Si H, Xu C, Zhang J, Zhang X, Li B, Zhou X, et al. Licochalcone A: an effective and low-toxicity compound against Toxoplasma gondii in vitro and in vivo. Int J Parasitol Drugs Drug Resist. (2018) 8:238–45. doi: 10.1016/j.ijpddr.2018.02.006
33. Egler J, Lang F. Licochalcone A induced suicidal death of human erythrocytes. Cell Physiol Biochem. (2015) 37:2060–70. doi: 10.1159/000438565
34. Travi BL, Cordeiro-da-Silva A, Dantas-Torres F, Miró G. Canine visceral leishmaniasis: diagnosis and management of the reservoir living among us. PLoS Negl Tro. Dis. (2018) 12:e0006082. doi: 10.1371/journal.pntd.0006082
35. Denerolle P, Bourdoiseau G. Combination allopurinol and antimony treatment versus antimony alone and allopurinol alone in the treatment of canine leishmaniasis (96 cases). J Vet Intern Med. (1999) 13:413–5 doi: 10.1111/j.1939-1676.1999.tb01455.x
36. Miró G, Oliva G, Cruz I, Cañavate C, Mortarino M, Vischer C, et al. Multicentric, controlled clinical study to evaluate effectiveness and safety of miltefosine and allopurinol for canine leishmaniosis. Vet Dermatol. (2009) 20:397–04. doi: 10.1111/j.1365-3164.2009.00824.x
37. Yardley V, Croft SL, De Doncker S, Dujardin JC, Koirala S, Rijal S, et al. The sensitivity of clinical isolates of Leishmania from Peru and Nepal to miltefosine. Am J Trop Med Hyg. (2005) 73:272–5. doi: 10.4269/ajtmh.2005.73.272
38. Franco-Muñoz C, Manjarrés-Estremor M, Ovalle-Bracho C. Intraspecies differences in natural susceptibility to amphotericine B of clinical isolates of Leishmania subgenus Viannia. PLoS ONE. (2018) 13:e0196247. doi: 10.1371/journal.pone.0196247
39. Lv H, Xiao Q, Zhou J, Feng H, Liu G, Ci X. Licochalcone a upregulates Nrf2 antioxidant pathway and thereby alleviates acetaminophen-induced hepatotoxicity. Front Pharmacol. (2018) 9:147. doi: 10.3389/fphar.2018.00147
40. Rodrigues RF, Charret KS, da Silva EF, Echevarria A, Amaral FV, Leon LL, et al. Antileishmanial Activity of 1,3,4-thiadiazolium-2-aminide in mice infected with Leishmania amazonensis. Antimicrob Agents Chemother. (2009) 53:839–42. doi: 10.1128/AAC.00062-08
41. Zhai L, Blom J, Chen M, Christensen SB, Kharazmi A. The antileishmanial agent licochalcone A interferes with the function of parasite mitochondria. Antimicrob Agents Chemother. (1995) 39:2742–8. doi: 10.1128/AAC.39.12.2742
42. Chen M, Zhai L, Christensen SB, Theander TG, Kharazmi A. 2001. Inhibition of fumarate reductase in Leishmania major L. donovani by chalcones. Antimicrob Agents Chemother. (2001) 45:2023–29. doi: 10.1128/AAC.45.7.2023-2029.2001
43. Merlino A, Vieites M, Gambino D, Laura Coitiño E. Homology modeling of T. cruzi and L. major NADH-dependent fumarate reductases: Ligand docking, molecular dynamics validation, and insights on their binding modes. J Mol Graph Model. (2014) 48:47–59. doi: 10.1016/j.jmgm.2013.12.001
44. Rana S, Mahato JP, Kumar M, Sarsaiya S. Modeling and docking of cysteine protease-A (CPA) of Leishmania donovani. J Appl Pharm Sci. (2017) 7:179–84. doi: 10.7324/JAPS.2017.70924
45. Rana S, Sivaperumal R. Virtual screening and molecular docking of Anti-Antileishmanial for selected pharmacophore for visceral Leishmaniasis. J Drug Deliv Ther. (2018) 8:230–5. doi: 10.22270/jddt.v8i6-s.2120
Keywords: cutaneous leishmaniasis, visceral leishmaniasis, canine visceral leishmaniasis, licochalcone A, leishmanicidal activity
Citation: Souza JM, de Carvalho ÉAA, Candido ACBB, de Mendonça RP, Fernanda da Silva M, Parreira RLT, Dias FGG, Ambrósio SR, Arantes AT, da Silva Filho AA, Nascimento AN, Costa MR, Sairre MI, Veneziani RCS and Magalhães LG (2020) Licochalcone a Exhibits Leishmanicidal Activity in vitro and in Experimental Model of Leishmania (Leishmania) Infantum. Front. Vet. Sci. 7:527. doi: 10.3389/fvets.2020.00527
Received: 16 May 2020; Accepted: 08 July 2020;
Published: 10 December 2020.
Edited by:
Jiyu Zhang, Chinese Academy of Agricultural Sciences, ChinaReviewed by:
Elmo Eduardo Almeida Amaral, Instituto Oswaldo Cruz, BrazilCopyright © 2020 Souza, de Carvalho, Candido, de Mendonça, Fernanda da Silva, Parreira, Dias, Ambrósio, Arantes, da Silva Filho, Nascimento, Costa, Sairre, Veneziani and Magalhães. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lizandra G. Magalhães, bGl6YW5kcmEubWFnYWxoYWVzQHVuaWZyYW4uZWR1LmJy; Rodrigo C. S. Veneziani, cm9kcmlnby52ZW5lemlhbmlAdW5pZnJhbi5lZHUuYnI=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.