 Mikayla F. A. Baxter1
Mikayla F. A. Baxter1 Juan D. Latorre1
Juan D. Latorre1 Sami Dridi1
Sami Dridi1 Ruben Merino-Guzman2
Ruben Merino-Guzman2 Xochitl Hernandez-Velasco2
Xochitl Hernandez-Velasco2 Billy M. Hargis1
Billy M. Hargis1 Guillermo Tellez-Isaias1*
Guillermo Tellez-Isaias1*- 1Department of Poultry Science, University of Arkansas, Fayetteville, AR, United States
- 2Departamento de Medicina y Zootecnia de Aves, Facultad de Medicina Veterinaria y Zootecnia, Universidad Nacional Autónoma de México, Mexico City, Mexico
Intestinal health is essential for feed efficiency and growth in animal agriculture and is dependent on barrier function, inflammation and dysbiosis. Our laboratory has published a nutritional model to induce gut inflammation using rye as a source of energy in poultry. More recently, we have used this model as an assessment of a nutritional rehabilitation model for better understanding of childhood undernutrition. The objective of this brief research report was to use a well-establish malabsorption model in broiler chickens using corn and rye as an energy source to identify several intestinal health biomarkers in the serum. To screen for inflammatory biomarkers, seven commercially available tests were used including Griess, superoxide dismutase, thiobarbituric acid reactive substances, Total antioxidant capacity, extracellular-signal-regulated kinase, Citrulline, and Interferon-ɤ; total IgA from cloacal swab was also measured. In the present study, chickens fed rye had a significant (P < 0.05) reduction in body weight and body weight gain at 10 day when compared with chickens that received the corn diet. In the second phase of the experiment, chickens that remain with the corn diet had significant differences in body weight and body weight gain. No significant differences were observed for any of the four antioxidant biomarkers evaluated in the sera (P > 0.05). However, significant differences were observed in serum citrulline and IFN-ɤ, as well as in cloacal IgA, in broiler chickens fed with rye, suggesting their potential use as biomarkers to study intestinal inflammation.
Introduction
Intestinal health is essential for feed efficiency and growth in animal agriculture and is dependent on barrier function, inflammation, and dysbiosis (1, 2). Determining reliable, high throughout biomarkers to measure intestinal inflammation, and barrier function in poultry is continuously being investigated. In a recent study, Chen et al. (3) developed an inflammatory gut barrier failure model in chickens using high NSP (Non-starch polysaccharides) diet, and coccidiosis. Other biomarkers have also been evaluated to assess enterocyte health. Inflammation associated with oxidative stress can induce physiological changes in gene expression suggesting that inflammation-induced oxidative stress plays a crucial role in intestinal function (2, 3). Mitochondrial respiration is essential in maintaining TEER (Trans epithelial electrical resistance), suggesting oxidation plays an essential role in tight junction stability in Caco-2 cells (4). Decreases in mitochondrial ATP production resulted in a decrease in permeability and increase in the gene expression of occludin and claudin-1 but a decrease in the gene expression claudin-2 and claudin-7 providing a direct link between intestinal permeability, mitochondrial function, and cellular energy status (4). Oxidative stress is often measured by looking at metabolites produced during oxidation or enzyme activity. Superoxide dismutase (SOD) is an antioxidant enzyme that neutralizes the harmful by-products of metabolism (5). Thiobarbituric acid reactive substances (TBARS) are metabolites produced during peroxidation; total antioxidant capacity detects the antioxidant potential in a sample, and the Griess assay uses nitrite and nitrate breakdown to measure nitric oxide concentration within the cell (6). Our laboratory has published a nutritional model to induce gut inflammation using rye as a source of energy in poultry (3, 7–13). More recently, we have used this model as an assessment of a nutritional rehabilitation model for better understanding of childhood undernutrition (14). Citrulline is produced exclusively by the enterocytes of the small bowel, is the nitrogen end product of glutamine metabolism and can be converted to arginine (15). In pre-weaned piglets, plasma citrulline levels correlated with the intestinal absorption markers mannitol suggesting that citrulline can be used as a marker for intestinal function (16). The extracellular signal-regulated kinase (ERK) is a primary signaling pathway in the regulation of intestinal epithelial proliferation, survival, and wound healing (17). Therefore, it is plausible that ERK activity in the serum could reflect the intestinal damage incurred during stress.
On the other hand, secretory IgA (SIgA) is an essential part of the adaptive humoral immune system and the primary immunoglobulin that neutralizes pathogens on external mucosal surfaces while resisting proteases (18–20). LPS-induced TNF-α (Tumor necrosis factor-Alpha) is a transcription factor, which induces the expression of TNF-α, a pro-inflammatory cytokine (21). Interferon-gamma (IFN-ɤ), is another proinflammatory cytokine of the innate immune system with immunomodulatory and immuno-stimulatory properties (22–25). Hence, it is likely that both, innate and adaptive immune response, may provide a viable biomarker to assess intestinal health. The objective of this brief research report was to use a malabsorption model in broiler chickens to evaluate several intestinal health biomarkers in the serum and total IgA from gut mucosa.
Materials and Methods
Animals and Experimental Design
All animals and animal procedures were approved by the Institutional Animal Care and Use Committee at the University of Arkansas in Fayetteville. One hundred and sixty day old mixed sex broiler chicks from Cobb-Vantress, Silom Springs, AR, USA (n = 40 chickens/group) were used in this experiment. On the day of hatch, chicks were neck tagged and randomly allocated to one of four dietary treatments on group floor pens in temperature control rooms which were set using standard management practices. All diets were antibiotic-free and formulated to meet or exceed the current broiler nutritional requirements according to the National Research Council (1994; Table 1). The dietary treatments were (1) a control corn-based diet that chick consumed throughout the trial (corn-corn); (2) an early phase malnutrition diet where chicks were fed a rye-based diet during the first phase of the experiment and then in the second phase were fed a corn-based diet (rye-corn); (3) a control rye-based diet that chick consumed throughout the trial (rye-rye); (4) a late phase malnutrition diet where chicks were fed a corn-based diet during the first phase of the experiment than in the second phase were fed a rye-based diet (corn-rye). The experiment was divided into two phases, the first phase was from the day of hatch to day 10 and the second phase was from day 10 to day 20. At the end of the first phase, before the diets were switched, half the chickens were euthanized (n = 20 chicks/ diet). At the end of the second phase, the remaining chicks (n = 20 chicks/diets were euthanized to measure intestinal permeability after the diets were switched (corn-corn, rye-corn, rye-rye, corn-rye).
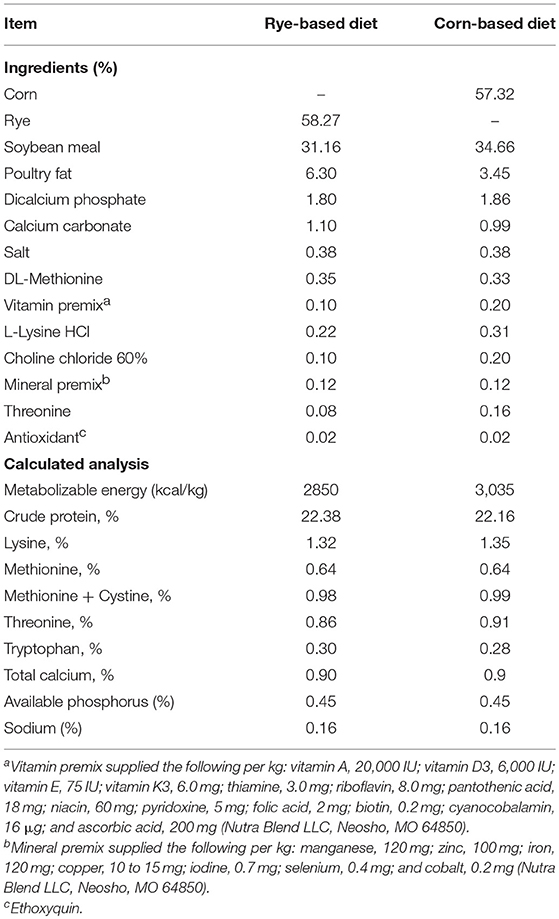
Table 1. Composition and nutrient content of the experimental diets (%).
Assay Kits
To screen for inflammatory biomarkers, eight commercially available kits were purchased, and assays were conducted following the detailed description of the protocols for each assay, respectively. Oxidative stress kits were generic, and all kits were specific for chicken. The TBARS Assay kit, the SOD assay kit, and the antioxidant Assay kit were purchased from Cayman Chemical Company (Ann Arbor, Michigan, USA). The Griess reagent kit was purchased from Invitrogen™ Molecular Probes (Waltham, MA, USA) to determine nitrate concentration as an indicator of nitric oxide. The ERK and Citrulline sandwich ELISA kits were purchased from MyBioSource (San Diego, California, USA). IFN-ɤELISA kit was purchased from Invitrogen Corporation (Frederick, Maryland, USA). Total IgA concentrations were measured in the cloaca as previously described by Merino-Guzmán et al. (19).
Statistical Analysis
All data were subjected to analysis of variance as a completely randomized design using the General Linear Models procedure of SAS (26). Data are expressed as mean ± standard error. Significant differences among the means were determined by using Duncan's multiple range test at P < 0.05.
Results and Discussion
Inflammation is the endpoint of stress, regardless of its origin or nature (biological, environmental, nutritional, physical, chemical, or psychological). Stress and inflammation are innate responses involving hormones, immune cells, and molecular mediators, which are essential mechanisms for the survival and the healing process in all forms of life (20). During chronic inflammation, the increased production of reactive oxygen species induces peroxidation of lipids in cell membranes as well as mitochondria membranes (27). The long-term damage of this vital organelle has a profound impact on all cells of the individual. It is well accepted that in animals, the interactions between diet ingredients, gut microbiome, nervous system, immune system, and endocrine system play critical roles in metabolic and gastrointestinal disorders (28–30).
Evaluation of a nutritional rehabilitation model on body weight and body weight gain in broiler chickens fed rye or corn at varying time points are summarized in Table 2. In the present study, feed intake was not recorded. However, chickens fed rye had a significant (P < 0.05) reduction in body weight and body weight gain at 10 day when compared with chickens that received the corn diet. In the second phase of the experiment, chickens that remain with the corn diet had significant differences in body weight and body weight gain (Table 2). The reduction if body weight and body weight gain observed in the present study, are in agreement with previous studies conducted in our laboratory using a rye diet in modern broilers (11). In that study, a Bacillus direct fed microbial (DFM) was included in the same rye diet as a treatment group. Interestingly, although differences were observed in feed intake between control rye diet and DFM treated group, chickens that received the Bacillus spore based probiotic in the feed had a significant reduction in feed conversion ratio, digesta viscosity and liver bacterial translocation when compared with control non treated chickens (11). Similar results have been observed using other high non-starch polysaccharide diets (12, 13).
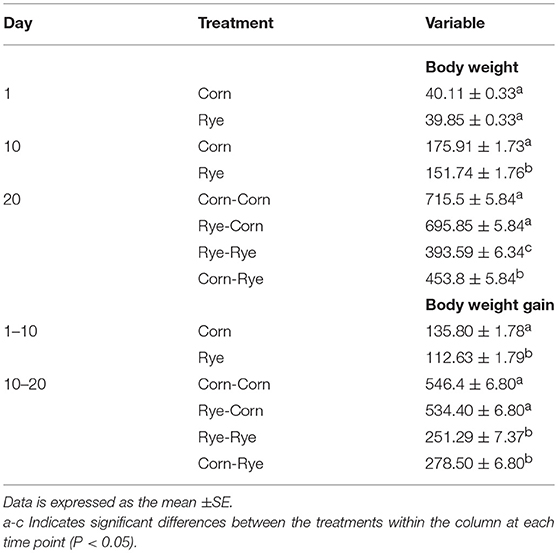
Table 2. Evaluation of a nutritional rehabilitation model on body weight and body weight gain in broiler chickens fed rye or corn at varying time points.
The oxidative stress has been related not only to the increased production of free radicals but also to changes to the scavenging capacity of antioxidant systems. To adapt to oxidative stress, the antioxidant systems in the body contain antioxidant enzymes such as SOD and Glutathione peroxidase which are employed to protect the body from oxidative stress (31). Dietary supplementation of several nutraceuticals have been shown to markedly ameliorate oxidative damage induced by heat stress in broiler chickens (32, 33). Furthermore, serum biochemical biomarkers such as glucose, proteins, calcium, phosphorus, and alkaline phosphatase concentrations have been reported to evaluate seasonal variations between male and female broiler chickens (34). Similar biomarkers have also been used to evaluate the antioxidant and immune modulatory properties of dietary supplements (35–37).
At 10 days of age, only three biomarkers were evaluated (SOD, IgA, and Citrulline). No statistical differences (P > 0.05) were observed in any of these biomarkers between corn or rye treated chickens (data not shown). Similarly, at day 20, no differences were detected for any of the four antioxidant biomarkers in the sera (Table 3). These metabolites may be processed in the liver preventing them from being detected in the sera. Previous research used commercially available kits to detect the oxidative stress differences in the plasma between thermoneutral and acute heat stress-treated chickens. Acute heat stress significantly increased plasma TBARS but had no significant effect on SOD and ferric/reducing antioxidant power (38). Other studies have also shown that systemic increases in oxidative stress markers are likely dependent on the type of stress (39, 40).
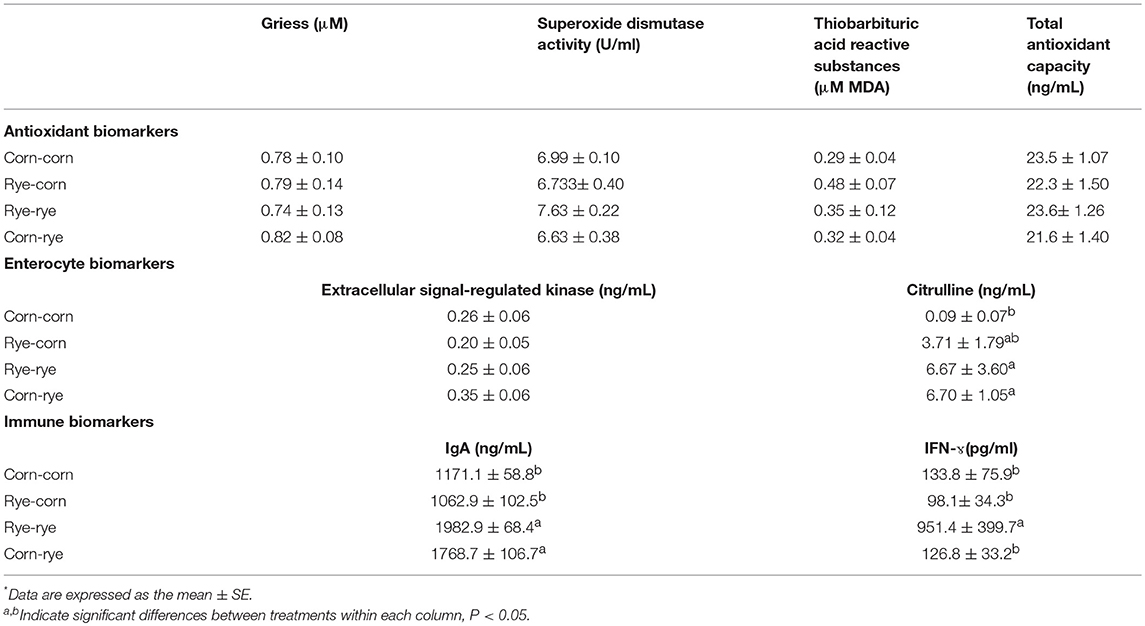
Table 3. Evaluating biomarkers in the sera of broiler chickens fed rye and corn at 20 days of age*.
While no significant differences were observed on ERK between treated groups, a significant concentration in serum citrulline was observed in chickens fed rye-rye or chickens fed corn-rye (Table 3).
On the other hand, circulating citrulline is mainly produced by enterocytes of the small bowel. For this reason, plasma or serum citrulline concentration has been proposed as a biomarker of remnant small bowel mass and function (41). Hence, in humans decreased level of plasma citrulline correlated with the reduced enterocyte mass independently of nutritional and inflammatory status (42, 43). Intestinal citrulline production originates mainly from the proximal small bowel, and probably from the middle and upper parts of intestinal villi. The primary precursor is glutamine, and derived amino acids, either circulating or exogenous (44). Glutamine, arginine, and possibly other amino acids such as proline or ornithine, also contribute to the intestinal production of citrulline (45).
Interestingly, using the same nutritional rehabilitation model from the present study, Baxter et al. (14), demonstrated that duodenum of modern broilers fed corn in the second phase of the experiment had statistically lower villi height (VH) and shorter crypt depth than rye-fed chicks. Previous research has found that the chicks consuming a high fiber diet had a higher epithelial turnover and an increase in VH, to try to compensate for the poor digestibility (46–48). Therefore, the increasing concentration of serum citrulline observed in chickens that received the rye diets correlates with our previous findings that rye increases duodenal surface area. Hence, our results confirm that serum citrulline is correlated to small bowel length as has been demonstrated in mammals (43–45).
The intestinal mucosa releases anti-microbial proteins and IgA. IgA is an antibody isotype specialized in protecting the intestinal mucosa, as well as inhibiting inflammatory processes, neutralizing bacterial toxins, and enhancing nonspecific defense mechanisms (e.g., lactoperoxidase and lactoferrin) (19, 49). Secretory IgA provides the first line of defense by preventing pathogen entry into the mucosa (50–52). Chicks fed rye in the second phase of the experiment had significantly higher levels of cloacal IgA than those fed a corn-based diet (Table 3). In human medicine, fecal secretory IgA is measured as an indicator of intestinal infection, celiac disease, and food allergies (44, 53, 54). In chickens, Salmonella enterica infection and treatment with a probiotic Enterococci faecium resulted in an increase SIgA in an intestinal wash compared to the control (43).
Furthermore, a rye-based diet has been associated with pathogenic bacteria, specifically Clostridium perfringens (3, 9–11). Therefore, the higher abundance of cloacal IgA in the chicks fed rye in the second phase of the experiment is likely due to the dysbacteriosis, intestinal inflammation, and gut permeability associated with rye diets in poultry (9, 10). The rye-rye treatment group also had a significantly higher amount of serum IFN-ɤcompared to any of the other dietary treatment groups (Table 3). This finding suggests that a rye-based diet increased the amount of systemic pro-inflammatory cytokine IFN-ɤ. Pro-inflammatory cytokines like IFN-ɤhave also been reported to increase the expression of the pore-forming tight junction protein like claudin-2 and decrease expression of the pore-sealing claudin tight junctions like claudin-1, 3, 4, 5, and 8 (46, 55). Therefore, it is evident that broilers consuming a rye-based diet for 20 days increased the systemic level of IFN-ɤwhich likely contributed to an increase in intestinal permeability by modifying tight junction distribution within the intestinal tract as has been previously demonstrated (14). In summary, cloacal IgA, serum citrulline, and IFN-ɤmay be used as potential biomarkers to study intestinal inflammation in chickens.
Author Contributions
MB, GT-I, and SD conceived and planned the study. JL and GT-I supervised all research. GT-I and BH analyzed and interpreted data. RM-G, BH, SD, and XH-V revised the first version of the manuscript. XH-V and GT-I were responsible for the final editing of the manuscript. All the authors approved the final version of the manuscript.
Funding
This research was supported by the Arkansas Bioscience Institute under the project: development of an avian model for evaluation early enteric microbial colonization on the gastrointestinal tract and immune function.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. Gilani S, Howarth GS, Kitessa SM, Tran CD, Forder REA, Hughes RJ. New biomarkers for increased intestinal permeability induced by dextran sodium sulphate and fasting in chickens. J Anim Physiol Anim Nutr. (2017) 101:e237–45. doi: 10.1111/jpn.12596
2. Ducatelle R, Goossens E, Meyer F, Eeckhaut V, Antonissen G, Haesebrouck F, et al. Biomarkers for monitoring intestinal health in poultry: present status and future perspectives. Vet Res. (2018) 49:43. doi: 10.1186/s13567-018-0538-6
3. Chen J, Tellez G, Richards JD, Escobar J. Identification of potential biomarkers for gut barrier failure in broiler chickens. Front Vet Sci. (2015) 2:14. doi: 10.3389/fvets.2015.00014
4. Janssen-Duijghuijsen LM, Grefte S, de Boer VC, Zeper L, van Dartel DA, van der Stelt I, et al. Mitochondrial ATP depletion disrupts Caco-2 monolayer integrity and internalizes claudin 7. Front Physiol. (2017) 8:794. doi: 10.3389/fphys.2017.00794
5. Xu JX, Cao CY, Sun YC, Wang LL, Li N, Xu SW, et al. Effects on liver hydrogen peroxide metabolism induced by dietary selenium deficiency or excess in chickens. Biol Trace Elem Res. (2014) 159:174–82. doi: 10.1007/s12011-014-0002-z
6. Ding AH, Nathan CF, Stuehr DJ. Release of reactive nitrogen intermediates and reactive oxygen intermediates from mouse peritoneal macrophages. Comparison of activating cytokines and evidence for independent production. J Immunol. (1988) 141:2407–12.
7. MacAuliffe T, McGinnis J. Effect of antibiotic supplements to diets containing rye on chick growth. Poult Sci. (1971) 50:1130–4.
8. Campbell GL, Campbell LD, Classen HL. Utilisation of rye by chickens: effect of microbial status, diet gamma irradiation and sodium taurocholate supplementation. Br Poult Sci. (1983) 24:191–203.
9. Tellez G, Latorre JD, Kuttappan VA, Kogut MH, Wolfenden A, Hernandez-Velasco X, et al. Utilization of rye as energy source affects bacterial translocation, intestinal viscosity, microbiota composition, and bone mineralization in broiler chickens. Front Genet. (2014) 5:339. doi: 10.3389/fgene.2014.00339
10. Tellez G, Latorre JD, Kuttappan VA, Hargis BM, Hernandez-Velasco X. Rye affects bacterial translocation, intestinal viscosity, microbiota composition and bone mineralization in turkey poults. PLoS ONE. (2015) 10:e0122390. doi: 10.1371/journal.pone.0122390
11. Latorre J, Hernandez-Velasco X, Bielke L, Vicente J, Wolfenden R, Menconi A, et al. Evaluation of a Bacillus direct-fed microbial candidate on digesta viscosity, bacterial translocation, microbiota composition and bone mineralization in broiler chickens fed on a rye-based diet. Br Poult Sci. (2015) 56:723–32. doi: 10.1080/00071668.2015.1101053
12. Latorre JD, Hernandez-Velasco X, Kuttappan VA, Wolfenden RE, Vicente JL, Wolfenden AD, et al. Selection of Bacillus spp. for cellulase and xylanase production as direct-fed microbials to reduce digesta viscosity and Clostridium perfringens proliferation using an in vitro digestive model in different poultry diets. Front Vet Sci. (2015) 2:25. doi: 10.3389/fvets.2015.00025
13. Latorre JD, Hernandez-Velasco X, Vicente JL, Wolfenden R, Hargis BM, Tellez G. Effects of the inclusion of a Bacillus direct-fed microbial on performance parameters, bone quality, recovered gut microflora, and intestinal morphology in broilers consuming a grower diet containing corn distillers dried grains with solubles. Poult Sci. (2017) 96:2728–35. doi: 10.3382/ps/pex082
14. Bederska-Lojewska D, Swiatkiewicz S, Arczewska-Włosek A, Schwarz T. Rye non-starch polysaccharides: their impact on poultry intestinal physiology, nutrients digestibility and performance indices-a review. Ann Anim Sci. (2017) 17:351–69. doi: 10.1515/aoas-2016-0090
15. Van Krimpen M, Torki M, Schokker D. Effects of rye inclusion in grower diets on immune competence-related parameters and performance in broilers. Poult Sci. (2017) 96:3324–37. doi: 10.3382/ps/pex152
16. Baxter MF, Latorre JD, Koltes DA, Dridi S, Greene ES, Bickler SW, et al. Assessment of a nutritional rehabilitation model in two modern broilers and their jungle fowl ancestor: a model for better understanding childhood undernutrition. Front Nutr. (2018) 5:18. doi: 10.3389/fnut.2018.00018
17. Windmueller HG, Spaeth AE. Source and fate of circulating citrulline. Am J Physiol. (1981) 241:E473–80.
18. Berkeveld M, Langendijk P, Verheijden JHM, Taverne MAM, van Nes A, van Haard P, et al. Citrulline and intestinal fatty acid-binding protein: longitudinal markers of postweaning small intestinal function in pigs? J Anim Sci. (2008) 86:3440–9. doi: 10.2527/jas.2008-1167
19. Iizuka M, Konno S. Wound healing of intestinal epithelial cells. World J Gastroenterol. (2011) 17:2161–71. doi: 10.3748/wjg.v17.i17.2161
20. Hermans D, Pasmans F, Heyndrickx M, Van Immerseel F, Martel A, Van Deun K, et al. A tolerogenic mucosal immune response leads to persistent Campylobacter jejuni colonization in the chicken gut. Crit Rev Microbiol. (2012) 38:17–29. doi: 10.3109/1040841X.2011.615298
21. Merino-Guzmán R, Latorre JD, Delgado R, Hernandez-Velasco X, Wolfenden AD, et al. Comparison of total immunoglobulin A levels in different samples in Leghorn and broiler chickens. Asian Pac J Trop Biomed. (2017) 7:116–20. doi: 10.1016/j.apjtb.2016.11.021
22. Staley M, Conners MG, Hall K, Miller LJ. Linking stress and immunity: immunoglobulin A as a non-invasive physiological biomarker in animal welfare studies. Horm Behav. (2018) 102:55–68. doi: 10.1016/j.yhbeh.2018.04.011
23. Hong YH, Lillehoj HS, Lee SH, Dalloul RA, Lillehoj EP. Analysis of chicken cytokine and chemokine gene expression following Eimeria acervulina and Eimeria tenella infections. Vet Immunol Immunopathol. (2006) 114:209–23. doi: 10.1016/j.vetimm.2006.07.007
24. Utech M, Ivanov AI, Samarin SN, Bruewer M, Turner JR, Mrsny RJ, et al. Mechanism of IFN-gamma-induced endocytosis of tight junction proteins: myosin II-dependent vacuolarization of the apical plasma membrane. Mol Biol Cell. (2005) 16:5040–52. doi: 10.1091/mbc.e05-03-0193
25. Wang F, Graham WV, Wang Y, Witkowski ED, Schwarz BT, Turner JR. Interferon-gamma and tumor necrosis factor-alpha synergize to induce intestinal epithelial barrier dysfunction by up-regulating myosin light chain kinase expression. Am J Pathol. (2005) 166:409–19.
26. Nava P, Koch S, Laukoetter MG, Lee WY, Kolegraff K, Capaldo CT, et al. Interferon-gamma regulates intestinal epithelial homeostasis through converging beta-catenin signaling pathways. Immunity. (2010) 32:392–402. doi: 10.1016/j.immuni.2010.03.001
27. Cavalcanti YVN, Brelaz MCA, Neves JK, Ferraz JC, Pereira VR. Role of TNF-alpha, IFN-gamma, and IL-10 in the development of pulmonary tuberculosis. Pulm Med. (2012) 2012:745483. doi: 10.1155/2012/745483
29. Büyükgüzel E, Büyükgüzel K, Snela M, Erdem M, Radtke K, Ziemnicki K. Effect of boric acid on antioxidant enzyme activity, lipid peroxidation, and ultrastructure of midgut and fat body of Galleria mellonella. Cell Biol Toxicol. (2013) 29:117–29. doi: 10.1007/s10565-013-9240-7
30. Mayer EA, Knight R, Mazmanian SK, Cryan JF, Tillisch K. Gut microbes and the brain: paradigm shift in neuroscience. J Neurosci. (2014) 34:15490–6. doi: 10.1523/JNEUROSCI.3299-14.2014
31. Kelly JR, Kennedy PJ, Cryan JF, Dinan TG, Clarke G, Hyland NP. Breaking down the barriers: the gut microbiome, intestinal permeability and stress-related psychiatric disorders. Front Cell Neurosci. (2015) 9:392. doi: 10.3389/fncel.2015.00392
32. Tellez G. Prokaryotes versus eukaryotes: who is hosting whom? Vet Vet Sci. (2014) 1:3. doi: 10.3389/fvets.2014.00003
33. Huang C, Jiao H, Song Z, Zhao J, Wang X, Lin H. Heat stress impairs mitochondria functions and induces oxidative injury in broiler chickens. J Anim Sci. (2015) 93:2144–53. doi: 10.2527/jas.2014-8739
34. Akhavan-Salamat H, Ghasemi HA. Alleviation of chronic heat stress in broilers by dietary supplementation of betaine and turmeric rhizome powder: dynamics of performance, leukocyte profile, humoral immunity, and antioxidant status. Trop Anim Health Prod. (2016) 48:181–8. doi: 10.1007/s11250-015-0941-1
35. Kamel NN, Ahmed AM, Mehaisen GM, Mashaly MM, Abass OA. Depression of leukocyte protein synthesis, immune function and growth performance induced by high environmental temperature in broiler chickens. Int J Biometeorol. (2017) 61:1637–45. doi: 10.1007/s00484-017-1342-0
36. Devi TG, Samanta AK, Ghosh TK, Samanta R, Debbarma N. Seasonal variations in serum biochemical constituents of broiler chickens. Indian J Poult Sci. (2016) 51:180–3. doi: 10.5958/0974-8180.2016.00025.8
37. Abou-Elkhair R, Ahmed H, Selim S. Effects of black pepper (Piper nigrum), turmeric powder (Curcuma longa) and coriander seeds (Coriandrum sativum) and their combinations as feed additives on growth performance, carcass traits, some blood parameters and humoral immune response of broiler chickens. Asian-Austr J Anim Sci. (2014) 27:847–54. doi: 10.5713/ajas.2013.13644
38. Flees J, Rajaei-Sharifabadi H, Greene E, Beer L, Hargis BM, Ellestad L, et al. Effect of Morinda citrifolia (Noni)-enriched diet on hepatic heat shock protein and lipid metabolism-related genes in heat stressed broiler chickens. Front Physiol. (2017) 8:919. doi: 10.3389/fphys.2017.00919
39. Tokusoglu O, Simsek A, Parvizi M, Eymen D. Turmeric curcuminoid polyphenolics as antioxidant and anticarcinogenic agents. Nat Sci Discov. (2015) 1:56–61. doi: 10.20563/nsd.50644
40. Lin H, Decuypere E, Buyse J. Acute heat stress induces oxidative stress in broiler chickens. Comp Biochem Physiol A Mol Integr Physiol. (2006) 144:11–7. doi: 10.1016/j.cbpa.2006.01.032
41. Lin H, Decuypere E, Buyse J. Oxidative stress induced by corticosterone administration in broiler chickens (Gallus gallus domesticus): 1.Chronic exposure. Comp Biochem Physiol B Biochem Mol Biol. (2004) 139:737–44. doi: 10.1016/j.cbpc.2004.09.013
42. Altan Ö, Pabuçcuoglu A, Altan A, Konyalioglu S, Bayraktar H. Effect of heat stress on oxidative stress, lipid peroxidation and some stress parameters in broilers. Br Poult Sci. (2003) 44:545–50. doi: 10.1080/00071660310001618334
43. Crenn P, Messing B, Cynober L. Citrulline as a biomarker of intestinal failure due to enterocyte mass reduction. Clin Nutr. (2008) 27:328–39. doi: 10.1016/j.clnu.2008.02.005
44. Crenn P, Coudray-Lucas C, Thuillier F, Cynober L, Messing B. Postabsorptive plasma citrulline concentration is a marker of absorptive enterocyte mass and intestinal failure in humans. Gastroenterology. (2008) 119:1496–505. doi: 10.1053/gast.2000.20227
45. Korpe P S, Petri W A. Environmental enteropathy: critical implications of a poorly understood condition. Trends Mol Med. (2012) 18:328–36. doi: 10.1016/j.molmed.2012.04.007
46. Moinard C, Cynober L. Citrulline: a new player in the control of nitrogen homeostasis. J Nutr. (2007) 137:1621S−5S. doi: 10.1093/jn/137.6.1621S
47. Bischoff SC, Barbara G, Buurman W, Ockhuizen T, Schulzke JD, Serino M, et al. Intestinal permeability-a new target for disease prevention and therapy. BMC Gastroenterol. (2014) 14:189. doi: 10.1186/s12876-014-0189-7
48. Jin L, Reynolds LP, Redmer DA, Caton JS, Crenshaw JD. Effects of dietary fiber on intestinal growth, cell proliferation, and morphology in growing pigs. J Anim Sci. (1994) 72:2270–8.
49. Yu B, Chiou PWS. Effects of crude fiber level in the diet on the intestinal morphology of growing rabbits. Lab Anim. (1996) 30:143–8.
50. Zulkifli I, Rahayu HI, Alimon A, Vidyadaran M, Babjee S. Gut micoflora and intestinal morphology of commercial broiler chickens and Red Jungle Fowl fed diets containing palm kernel meal. Arch Geflügelk. (2009) 73:49–55.
51. Siddiqui I, Majid H, Abid S. Update on clinical and research application of fecal biomarkers for gastrointestinal diseases. World J Gastrointest Pharmacol Ther. (2017) 8:39–46. doi: 10.4292/wjgpt.v8.i1.39
52. Karaffová V, Bobíková K, Husáková E, Levkut M, Herich R, Revajová V, et al. Interaction of TGF-ß4 and IL-17 with IgA secretion in the intestine of chickens fed with E. faecium AL41 and challenged with S. enteritidis. Res Vet Sci. (2015) 100:75–9. doi: 10.1016/j.rvsc.2015.04.005
53. Gutzeit C, Magri G, Cerutti A. Intestinal IgA production and its role in host-microbe interaction. Immunol Rev. (2014) 260:76–85. doi: 10.1111/imr.12189
54. Neutra MR, Kozlowski PA. Mucosal vaccines: the promise and the challenge. Nat Rev Immunol. (2006) 6:148–58. doi: 10.1038/nri1777
Keywords: biomarkers, chickens, citrulline, IgA, intestinal integrity
Citation: Baxter MFA, Latorre JD, Dridi S, Merino-Guzman R, Hernandez-Velasco X, Hargis BM and Tellez-Isaias G (2019) Identification of Serum Biomarkers for Intestinal Integrity in a Broiler Chicken Malabsorption Model. Front. Vet. Sci. 6:144. doi: 10.3389/fvets.2019.00144
Received: 01 November 2018; Accepted: 25 April 2019;
Published: 09 May 2019.
Edited by:
Amlan Kumar Patra, West Bengal University of Animal and Fishery Sciences, IndiaReviewed by:
Alireza Seidavi, Islamic Azad University, IranVincenzo Tufarelli, University of Bari Aldo Moro, Italy
Vito Laudadio, University of Bari Aldo Moro, Italy
Copyright © 2019 Baxter, Latorre, Dridi, Merino-Guzman, Hernandez-Velasco, Hargis and Tellez-Isaias. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Guillermo Tellez-Isaias, Z3RlbGxlekB1YXJrLmVkdQ==