 Mathias Busch1
Mathias Busch1 Hugo Brouwer1
Hugo Brouwer1 Germaine Aalderink1
Germaine Aalderink1 Gerrit Bredeck2
Gerrit Bredeck2 Angela A. M. Kämpfer2
Angela A. M. Kämpfer2 Roel P. F. Schins2
Roel P. F. Schins2 Hans Bouwmeester1*
Hans Bouwmeester1*- 1Division of Toxicology, Wageningen University and Research, Wageningen, Netherlands
- 2IUF—Leibniz-Research Institute for Environmental Medicine, Duesseldorf, Germany
Plastic particles in the nanometer range–called nanoplastics–are environmental contaminants with growing public health concern. As plastic particles are present in water, soil, air and food, human exposure via intestine and lung is unavoidable, but possible health effects are still to be elucidated. To better understand the Mode of Action of plastic particles, it is key to use experimental models that best reflect human physiology. Novel assessment methods like advanced cell models and several alternative approaches are currently used and developed in the scientific community. So far, the use of cancer cell line-based models is the standard approach regarding in vitro nanotoxicology. However, among the many advantages of the use of cancer cell lines, there are also disadvantages that might favor other approaches. In this review, we compare cell line-based models with stem cell-based in vitro models of the human intestine and lung. In the context of nanoplastics research, we highlight the advantages that come with the use of stem cells. Further, the specific challenges of testing nanoplastics in vitro are discussed. Although the use of stem cell-based models can be demanding, we conclude that, depending on the research question, stem cells in combination with advanced exposure strategies might be a more suitable approach than cancer cell lines when it comes to toxicological investigation of nanoplastics.
1 Introduction
Plastic polymers have become indispensable in our everyday life due to their favorable physicochemical properties and relatively low production costs. Annual plastics production has risen to 368 million tons in 2019 (https://plasticseurope.org) and 71% of the subsequent plastic waste ends up in aquatic or terrestrial environments (Geyer et al., 2017; de Souza Machado et al., 2018). This plastic waste is continually exposed to physical stress through abrasion or UV light and will inevitably fragment into microplastics (<5 mm) and eventually nanoplastics (<100 nm) (EFSA, 2016). In addition to environment-derived micro- and nanoplastic particles, microplastics have also intentionally been included in consumer products and can subsequently fragment to the nanoscale size (Enfrin et al., 2020). Other direct sources of micro- and nanoplastics include the release of fibers from synthetic textiles (Cai et al., 2021) or abrasion of car tires (Järlskog et al., 2022).
Nanoplastics cannot easily be measured directly due to a lack of suitable analytical techniques (Schwaferts et al., 2019), but are assumed to be present in household dust (Zhang et al., 2020), particulate matter in outside air (Yao et al., 2022) and foodstuffs including salt (Kosuth et al., 2018), bivalves (Van Cauwenberghe and Janssen, 2014; Davidson and Dudas, 2016), fish (Kershaw and Rochman, 2015), honey (Liebezeit and Liebezeit, 2015) and water (Kosuth et al., 2018; Mason et al., 2018). Humans are therefore exposed to nanoplastics by ingestion and inhalation and subsequent health outcomes are poorly understood. Studies on human exposure to microplastic particles show that annual intake may exceed hundred thousands of particles (Cox et al., 2019; Mohamed Nor et al., 2021), while data on nanoplastic exposure is not available yet. Societal concerns on the potential health effects of micro- and nanoplastic exposure have been increasing, underpinning the need for risk assessment (Coffin et al., 2022; Felipe-Rodriguez et al., 2022).
Nanoplastics are not a homogeneous contaminant but represent a mixture of sheets, fragments and fibers of various sizes comprised mainly of polyethylene (PE), polypropylene (PP), polyethylene terephtalate (PET) and polyvinyl chloride (PVC) (https://plasticseurope.org, Karbalaei et al., 2018). This heterogeneity complicates risk assessment as past research has mostly focused on polystyrene (PS) nanospheres, which do not accurately represent the multitude of nanoplastics to which humans are exposed. Only few studies have considered alternative polymer types (Magrì et al., 2018; Busch et al., 2021b; Magrì et al., 2021; Sun et al., 2021) and even fewer studies have focused on non-spherical (Busch et al., 2022a) or weathered particles (Gopinath et al., 2021; Roursgaard et al., 2022). Despite the lack of data on prototypical environmental nanoplastics, considerable effort has been made to implement read-across methodologies based on the particles’ intrinsic properties, Mode of Action (MoA) or associated (bio)molecules, but these algorithms are not yet applicable to nanoplastics risk assessment (Walkey et al., 2014; Helma et al., 2017; Toropova and Toropov, 2022).
There is limited information available on nanoplastics toxicity in vivo and much of our current knowledge stems from rodent studies using microplastics. These studies need to be interpreted with caution, as some of these studies have been scrutinized because of issues related to particle characterization, (overload) exposure, detection of particles in tissues (mass balance) and methodological issues related to histopathology (Coffin et al., 2022; Gouin et al., 2022). Rodent studies using high concentrations of PS nanoplastics report bioavailability after ingestion or inhalation and particle translocation to lymph nodes, liver, spleen, and kidneys (Jani et al., 1989; Walczak et al., 2015a). More data on plastic particle toxicity is available in the context of occupational exposure to polymers (Mason, 1975; Heldaas et al., 1984; Hsiao et al., 2004). However, this information holds limited applicability to environmental nanoplastics exposure, as these studies were conducted to simulate high lung exposure concentrations in plastic industry workers (Hesterberg et al., 1992; Porter et al., 1999; Warheit et al., 2003), which is hardly comparable to the current levels of airborne plastic particles in a non-occupational environment.
Besides the direct toxicity of nanoplastics, the potential toxicity of associated chemicals and components of the eco-corona should also be considered. Nanoplastics frequently contain additives, e.g., phthalates, which may comprise up to 50% of the total mass of plastics (Andrady and Rajapakse, 2016; Hartmann et al., 2019) and often contain known toxicants like dioxins, polycyclic aromatic hydrocarbons, halogenated flame retardants and heavy metals (Bouwmeester et al., 2015; Hahladakis et al., 2018; Zimmermann et al., 2019), which could be released from the polymers after uptake (Peters et al., 2022). Furthermore, the surface of particles spontaneously adsorbs surrounding chemicals and biomolecules, leading to an eco-corona or bio-corona, respectively (Casals et al., 2010; Pulido-Reyes et al., 2017).
The plethora of possible polymers, shapes, sizes and exposure scenarios make exhaustive testing of nanoplastics a daunting task that is ethically, financially and temporally unfeasible using animal models. The acceptance of data generated in in vitro models for regulatory purposes is being spurred by the ban on animal testing in the cosmetics industry (Pistollato et al., 2021) and nanotoxicity testing will also have to rely on data generated in vitro (EFSA, 2021). Initiated by “toxicology of the 21st century” (Hartung, 2009), and to better explain MoAs, models that emulate human physiology best, should be put forward. This is boosting the development of in vitro models ultimately replacing animal based testing (Tannenbaum and Bennett, 2015). Currently, the main models used for in vitro oral or inhalation nanotoxicity testing are based on immortalized cell lines derived from cancer tissues, including Caco-2 for the intestine and A549 for the lung (Jones and Grainger, 2009; Wang M. et al., 2018; Fröhlich, 2018; EFSA, 2021). In addition to these conventional models, novel models including stem cell-based approaches have recently started being used in nanotoxicity testing. These novel methods offer advantages over cancer cell lines by having an unaltered genotype, allowing the presence of more physiologically relevant cell types and may help estimate interindividual variation. In this review, we discuss novel stem cell-based in vitro models of the primarily exposed organs, intestine and lung, as a new approach methodology for nanoplastics toxicity testing and how the resulting data can be used to guide future risk assessment.
2 Selection of in vitro models to study the toxicity of nanoplastics
Based on the knowledge on microplastics and engineered nanomaterials, nanoplastics mainly enter the human body through ingestion or inhalation. Therefore, these barriers are initially most at risk and studying potential toxic effects in vitro requires physiologically relevant models of the intestine and lung, respectively. While cell line-based models have been extensively used in toxicological research, these models lack essential features, as discussed below.
Establishing advanced barrier models requires an actively dividing and tissue-specific cell source, for example fully differentiated primary cells or somatic stem cells that can be directly isolated from the human body, often obtained from biopsies. However, the replicating ability of differentiated primary cells is limited, as differentiation generally occurs after exiting the cell cycle (Ma et al., 2019). Recently, commercially available constructs based on primary human tissue of the intestine or lung, have become available and applied for toxicological studies (Jackson et al., 2018; Janssen et al., 2021). These constructs are generated from human donors and are directly provided to the user for one-time experimental application. Although these primary models are highly relevant in terms of physiology, the user is dependent on donor availability and unobstructed delivery, as these models cannot be permanently established in a laboratory independently from the commercial provider.
In contrast to already differentiated primary tissue and cells, somatic stem cells are undifferentiated cells that divide to replenish dying cells under physiological conditions (Zakrzewski et al., 2019). These somatic stem cells, also called adult stem cells (ASCs), remain dormant until surrounding factors activate them to divide and terminally differentiate into functional cells (Urbán and Cheung, 2021). ASCs are multipotent, meaning that they can develop into more than one cell type, but they cannot produce all cell types of the body, as is the case for pluripotent cell types (Lyssiotis et al., 2011; Lairson et al., 2013). Despite improvements in long-term culture expansion of ASCs, their lifespan appears limited and inversely correlated to the donors age and highly affected by the extraction site (Bruder et al., 1997; Majors et al., 1997). Overall, the limited availability of biopsy material in combination with the finite dividing capacity of both primary cells and ASCs make them less suited to be used in toxicological studies. However, their use has greatly advanced the field of patient-specific disease modeling, i.e. cancer research and drug screening (Drost and Clevers, 2018; Driehuis et al., 2020).
Embryonic stem cells (ESCs) and induced pluripotent stem cells (iPSC), on the other hand, have gained increasing interest as a source of stem cells due to their ability to differentiate into all cell types of the organism and their high proliferative capacity (Shi et al., 2017). Human ESCs are pluripotent stem cells generally derived from the inner cell mass of blastocysts (Thomson et al., 1998). The required number of embryos used to generate the resulting ESC lines have only been reported in a small number of cases (Mehta, 2014), hinting at a low success rate, which in combination with the destructive process on the blastocyst have led to an extensive ethical debate on the use of human material (Wert and Mummery, 2003; Lo and Parham, 2009). Conversely, iPSCs do not rely on the use of embryos or invasive biopsies, forming a more ethical source of pluripotent cells. iPSCs are somatic cells (e.g. fibroblasts) reprogrammed into a pluripotent state through the forced overexpression of the four transcription factors OCT4, SOX2, KLF4 and MYC (Takahashi et al., 2007). From their pluripotent state, iPSCs have been successfully differentiated to obtain tissue-specific morphology and gene expression for various organs, including the intestine (Takahashi et al., 2018) and lung (Miller et al., 2019). Embryonic-like cells derived from iPSC differentiation can be used to study a wide variety of organs and exposure scenarios, due to their proliferative capacity and the wide spectrum of available differentiation protocols (Ahmed et al., 2021). The differentiation of iPSCs into mature models of human organs usually requires multiple sequential steps in which cells are exposed to different growth factors mimicking the in vivo cell environment during mammalian embryonic development (Spence et al., 2011; Abud et al., 2017). As a result, the differentiation of iPSCs leads to models with a better representation of different tissue-specific cell types encountered in vivo (Grouls et al., 2022), in contrast to cancer lines which only represent one dominant cell type.
Particle toxicity is often evaluated based on the range of reversible and irreversible cellular damage in addition to the cells’ ability to overcome these effects, as well as signaling cascades involved in processes like proliferation and inflammation. As cancer cell lines underwent genetic changes to ensure continuous proliferation in a tumor microenvironment despite challenging metabolic conditions (Hartwell and Kastan, 1994; Sonugur and Akbulut, 2019), this could potentially lead to a skewed view on the toxicity-induced effects of nanoplastics on tissue. By contrast, stem cell-derived tissue-like cells maintain physiological proliferative behavior to provide a more representative view for toxicity-induced injury compared to cancer cell lines, while simultaneously providing multiple cell types in the same in vitro model. Thus, we view iPSC as a promising approach for quantitative nanoplastics toxicity testing with high physiological relevance in the primarily exposed organs intestine and lung.
2.1 Intestinal models
The intestine is a major part of the human digestive system, responsible for the peristalsis of chyme and the uptake of nutrients from ingested food. Accordingly, the intestinal epithelium is constantly exposed to contaminants taken up with food and water, making it an important target organ for toxicities and diseases. Therefore, research in the fields of medicine, pharmacology and toxicology has been focused on the development of suitable models to study effects on the intestine outside of animals.
To date, several studies report intestinal effects of ingested micro- and nanoplastics in a variety of in vitro models based on cancer cell lines. Among the available cell lines, the colorectal cancer cell line Caco-2 is by far the most utilized one (Cortés et al., 2020; Domenech et al., 2021; Stock et al., 2021), as it spontaneously differentiates into an enterocyte-like phenotype (Pinto et al., 1983), representing the most abundant cell type in the human intestine. However, several attempts have been made to increase the complexity of Caco-2 based in vitro models by implementing additional cell lines when investigating plastic particles (see Table 1). For example, an advanced intestinal model included HT29-MTX-E12 as goblet cells and PMA-differentiated THP-1 cells as macrophages to expand the model by the presence of mucus and an immunocompetent cell type (Busch et al., 2021a). Furthermore, a Caco-2/HT29 model supplemented with Raji-B cells to induce the differentiation into microfold cells (M-cells), a cell type specialized in transport across the epithelial barrier (Mabbott et al., 2013), was used to investigate the effects of PS nanoplastics (Domenech et al., 2020). Establishing even higher levels of complexity, a quadruple culture model was used to study plastic particle effects by complementing a Caco-2/HT29-MTX co-culture with macrophages and dendritic cells derived from human blood monocytes (Lehner et al., 2020). However, the co-culturing of different cancer cell lines and/or primary cells requires extensive fine-tuning of optimal culture conditions, e.g. in terms of medium composition, whereas different cell populations in cultures derived from stem cells originate from the same single cell source.
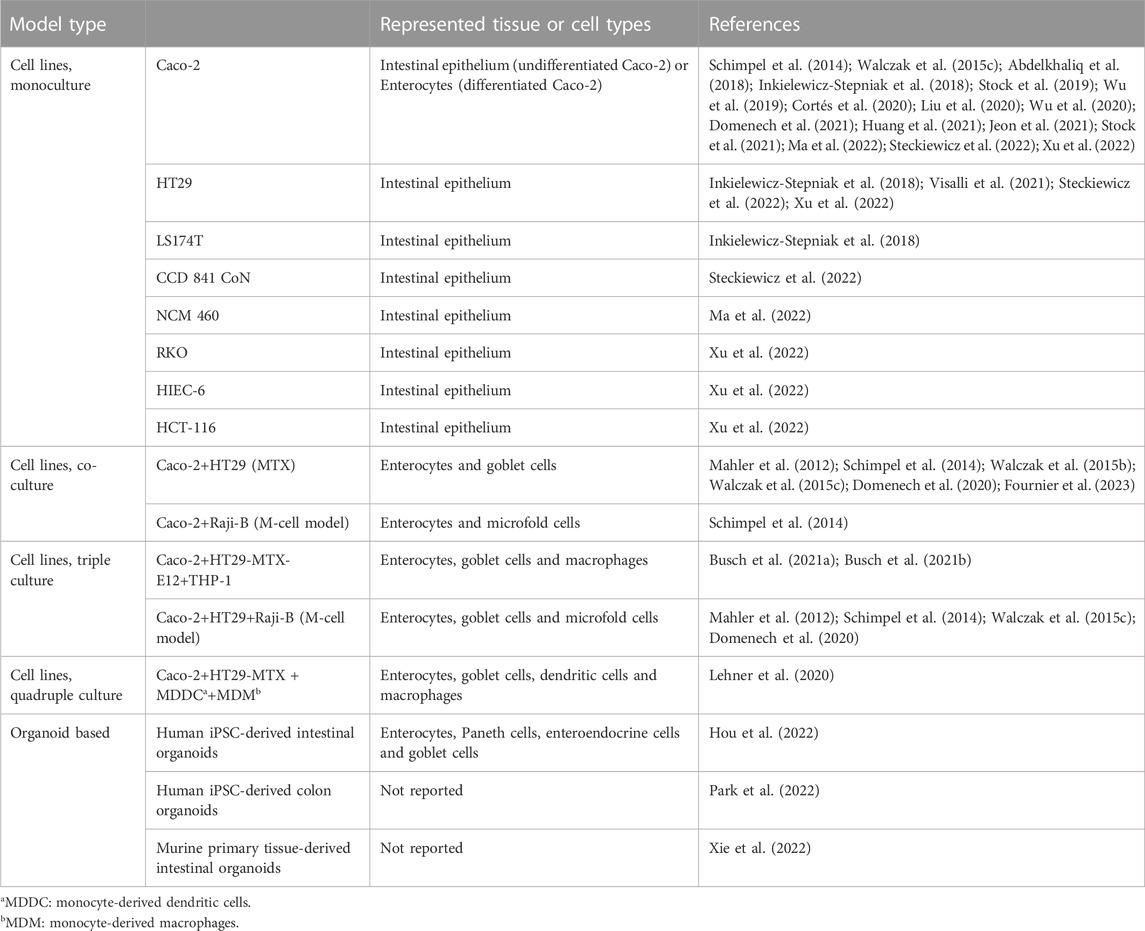
TABLE 1. Human intestinal in vitro models used for plastic particle toxicity assessment.
A breakthrough toward human cell-based models with advanced physiological relevance was seen through the establishment of crypt-based intestinal organoids, enabling the long-term culture of primary intestinal cells in absence of mesenchymal tissue (Sato et al., 2009; Sato et al., 2011). Since then, research interest in stem cells has increased dramatically, resulting in the generation of numerous culture and differentiation protocols (Cao et al., 2011; Spence et al., 2011; Forster et al., 2014; Miura and Suzuki, 2017; Hou et al., 2018; Pleguezuelos-Manzano et al., 2020; Stewart et al., 2020), establishing two principal approaches for the generation of self-organized intestinal spheroids: 1) isolation of intestinal crypts and 2) use of ESCs or iPSCs.
Uniquely, these spheroids represent all major epithelial cell lineages within the organoid (Sato et al., 2011), and allow to skew them towards specific cell types and rare lineages like tuft cells (Gerbe et al., 2016; Treveil et al., 2020; Inaba et al., 2021). Even the most sophisticated non-stem cell models still fail to replicate the lineage complexity to this extent (Lozoya-Agullo et al., 2017; Lehner et al., 2020; Kämpfer et al., 2022). For the purpose of hazard testing of nanoplastics, both overall effects on the tissue as well as effects in specific individual cell types may be relevant. Using organoids, cell type-specific uptake efficiencies were demonstrated for nanoplastics (Hou et al., 2022). The authors generated intestinal organoids from an iPSC line and reported the presence of enterocytes, Paneth cells, goblet cells and endocrine cells using immunofluorescence. Uptake of 50 nm PS nanoplastics was found to be cell type-dependent, which may have been overlooked or drastically underestimated in standard culture systems, as the authors report uptake primarily in goblet, Paneth and endocrine cells. Furthermore, some cell types may be more sensitive towards stressors or noxae (Busch et al., 2022b), and might, therefore, represent important targets for exposure studies.
Also their (patho)physiological identity renders intestinal stem cell models a highly versatile tool. The cultures can replicate the complexity of various regions of the intestinal tract–from duodenum to colon–as well as healthy or diseased tissue depending on the biopsy origin or growth/differentiation protocol (Sato et al., 2011; Tsai et al., 2017; Bandi et al., 2020). In contrast, standard cell models mostly rely on cancer-derived cell lines in mono- or co-culture that often require extensive differentiation times and further manipulation to obtain rather mature cell types (Hilgers et al., 1990; Lesuffleur et al., 1990) and disease-like phenotypes (Susewind et al., 2016; Kämpfer et al., 2017). Inherently diseased, these cell lines may be of limited physiological relevance for specific research questions due to biochemical and genetic differences compared to healthy intestinal tissue, e.g. the absence or limited presence of phosphoprotein p53 (Thant et al., 2008) and cytochrome (CYP) P450 isoforms (Ohta et al., 2020). While typically considered in the context of drug metabolization (Fritz et al., 2019), CYP enzymes also play a role in the effects of xenobiotics, e.g., organic pollutants (Takeshita et al., 2011; Abass et al., 2012), which may be critical contaminants to consider in plastic particle hazard assessment (Coffin et al., 2019; Brachner et al., 2020).
It is important to also realize potential differences between stem cell-based intestinal organoids and the in vivo situation, for example the limited presence of mucus, although refined protocols are being developed (Navabi et al., 2013; Sontheimer-Phelps et al., 2020). Whereas human organoids express physiologically relevant types of mucins–especially mucin 2 (Sato et al., 2011)—the overall luminal coverage and thickness of the mucus remains low (VanDussen et al., 2015). In the context of particle hazard assessment, mucus is a crucial determinant for the dose of nanoparticles that can interact with the epithelium (Bandi et al., 2020).
However, stem cell-derived intestinal cultures remain scarcely used for nanoplastics hazard assessment (Lu et al., 2017; Belair et al., 2020) with only three studies on plastic particles to date (Hou et al., 2022; Park et al., 2022; Xie et al., 2022), of which two studies failed to report the differentiation procedure and/or sufficiently characterize the organoids. Even with sufficient characterization, treatment of organoids with particles is a challenge, as the organoids’ morphology–the inward-facing apical side creating a lumen–and their maintenance in semi-solid media (e.g., Matrigel) impede particle exposures. Exposures would typically occur from the apical side, which is not accessible without manipulation, while diffusion of particles is obstructed in matrices like Matrigel. Various adaptions might resolve these challenges, including luminal microinjection or application of flow-through after puncturing the organoid (Leslie Jhansi et al., 2015; Sidar et al., 2019), reversed cell polarization (i.e. “apical-out” organoids) (Co et al., 2019) or breaking the organoids’ 3D structure down to a 2D structure. Especially the latter is applied frequently, either using the whole organoid (Kasendra et al., 2018; Madden et al., 2018; Roodsant et al., 2020; Yamada and Kanda, 2021; Breau et al., 2022) or selected cell types (Workman et al., 2018; Yoshida et al., 2020). While the structural change from 3D to 2D did not substantially modify the expression of intestinal markers or functional genes (Takahashi et al., 2021), these organoid-derived cell layers can develop a high transepithelial electrical resistance (TEER>1,500 Ω•cm2) (Wang Y. et al., 2018), which might indicate restricted passive transport across the cell layer and may limit the transport of nanoplastics. In contrast to manipulating mature organoids back into 2D layers, other protocols are designed to differentiate iPSCs directly into intestinal epithelial monolayers, without the detour of spheroid generation (Iwao et al., 2014; Kabeya et al., 2020), resulting in more realistic TEER values of ∼250 Ω•cm2 (Kabeya et al., 2018). Besides 2D culturing, adaptions creating accessible luminal space or using perfusion in tubular intestinal structures are becoming increasingly sophisticated (Naumovska et al., 2020; Nikolaev et al., 2020; Wilson et al., 2021) and offer promising alternative test systems.
2.2 Lung models
For inhaled particles, the lung epithelium forms an initial barrier and target site of interaction. Accordingly, in vitro testing of particulates for lung toxicity has long used cancer or immortalized epithelial cell lines (Hiemstra et al., 2018). While being less robust and reproducible, particles have also been tested on primary epithelial cells or tissue explant cultures from lungs of rodent or human origin (Crocker et al., 1973; Craighead et al., 1980; Mohr and Emura, 1985; Driscoll et al., 1995). The introduction of stem cell-based in vitro models and further advancement of associated tissue engineering methodologies in pulmonary research has contributed to an improved understanding of mechanisms of lung development, damage repair and tissue regeneration processes (Gotoh et al., 2014; Basil et al., 2020). However, the use of stem cell-based models for lung toxicity testing is still in its infancy compared to, for example, developmental neurotoxicity testing (Fritsche et al., 2021; Masui et al., 2022).
In sharp contrast, the specific field of particle toxicology has witnessed crucial progress through the development of improved in vitro assays that use lung epithelial cell models in air liquid interface (ALI) culture conditions (Adler et al., 1990; Blank et al., 2006; Lacroix et al., 2018). Combined with aerosol generating devices, this allows for a more realistic interaction between the test particles and the surfactant-containing apical side of the epithelial cells, in contrast to models in which cells are exposed to particle suspensions under submerged conditions (Ding et al., 2020). Another innovative in vitro approach involves the growing of lung epithelial cells on mechanically stretchable membranes to enable testing of particles under breathing movement mimicking conditions (Schmitz et al., 2019).
To date, the pulmonary toxicity of micro- or nano-sized plastics has been tested in various in vitro models and aforementioned specific experimental approaches. Most investigators have either used the A549 human alveolar type II-like epithelial cell line or the BEAS-2B human bronchial epithelial cell line under conventional submerged testing conditions (see Table 2). The toxicity of PS nanoplastics has been investigated in suspension on BEAS-2B cells as well as HPAEpiC human pulmonary alveolar epithelial cells, comprised of type I and type II cells (Yang et al., 2021). A microfluidic-chip test model with BEAS-2B cells was employed by Gupta et al. (2021) to study uptake of silica particles and PS. Furthermore, A549 cells were exposed on a membrane under cyclic stretch conditions to PS nanoparticles in order to mimic the effect of breathing movements (Roshanzadeh et al., 2020). Airway and alveolar epithelial toxicity has also been explored in ALI exposure settings. For example, the uptake and inflammatory properties of various PS particles were investigated in A549 cells, as well as in the Calu-3 human upper airway cell line under ALI exposure conditions (Meindl et al., 2021). To account for the potential contribution of alveolar macrophages, the authors also included a co-culture model of A549 and THP-1 cells (Meindl et al., 2021). To the best of our knowledge, to date only one study investigating plastics has been performed using a lung organoid-based testing approach. The toxicity of polyester fibers sampled from cloth dryers was investigated in human airway organoids that were generated from tissue resident ASCs obtained from three donors (Winkler et al., 2022). Using immunofluorescence and qRT-PCR-based markers, basal cells, ciliated cells, goblet cells, club cells, and even type I cells were identified. Interestingly, the authors exposed the organoid spheres after fragmentation to allow exposure to the fibers from the outside as well as the more relevant “inner cavities” of the organoids. While the outcomes of this study may be of limited relevance in view of the dimensions of the tested materials (average length: 700 ± 400 µm and average width 10 ± 5 µm), the work provides strong proof of principle for the use of lung organoids for hazard assessment of airborne nanoplastics.
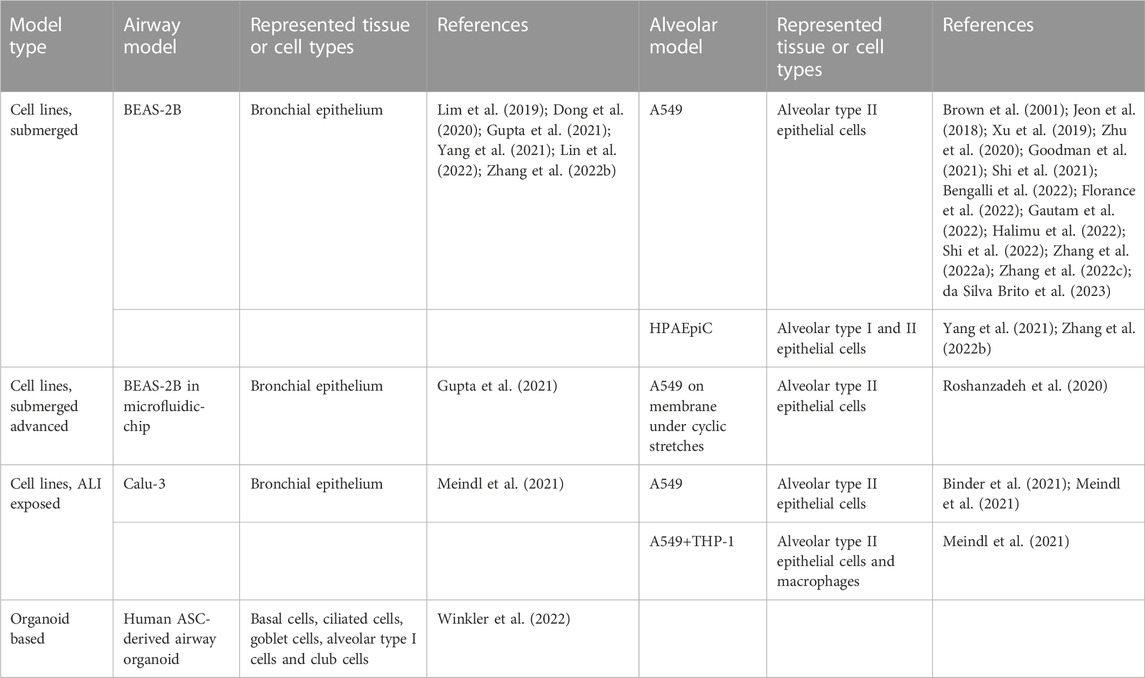
TABLE 2. Human lung in vitro models used for plastic particle toxicity assessment.
Similar to the intestine, a further introduction and application of stem cell-based models for pulmonary toxicity testing of nanoplastics seems promising. Herein, however, the principal advantage of exposing such models under ALI-conditions is to be emphasized in terms of dose and kinetics. Construction of complex organoid (e.g., scaffold) models, in such a way they that can be coupled to ALI exposure systems in a realistic, physiologically relevant manner, is a major challenge. If achieved, however, it will greatly contribute to improved safety evaluation of airborne nanoplastics.
3 Challenges and considerations when testing plastic particles in vitro
When studying the MoA of nanoplastics in an in vitro model, certain points should be carefully considered when designing the experiment, choosing relevant endpoints, or interpreting the data in regard to plastics hazard assessment:
Low density of polymers. Industrially produced plastics cover a whole spectrum of different densities, spanning from less than 1 g/cm³ (PP, PE), to around 1 g/cm³ (PS), to heavier polymers (PVC, polyamide (PA), PET). The density of cell culture medium is approximately 1 g/cm³, meaning that nanoplastics will either float or sediment during in vitro experiments, depending on their polymer composition. Polymers with a density lower than 1 g/cm³ will become buoyant when applied in a traditional cell culture system and will not establish contact with cells growing on the bottom of a well plate. So far, several approaches have been used to tackle this issue: one common approach is the inversion of the cell model, e.g. by seeding cells on cover slips that are subsequently inverted and exposed to buoyant particles from below (Stock et al., 2020), or by sealing a well plate with silicone gaskets and then inverting the entire plate (Watson et al., 2016). Furthermore, it has been reported that inverted in vitro models can be established by seeding cells on the basolateral side of transwell inserts (Stock et al., 2021), which also enables the application of more complex co-culture models (Busch et al., 2021b). Another approach to expose cells to buoyant plastic particles could be the use of ALI systems (Upadhyay and Palmberg, 2018). The absence of medium on the apical side of cells allows exposure to particles via particle-containing aerosols that sediment on the cell layers, regardless of the particles’ density. Although this approach has been used in an advanced in vitro model of the intestine (Lehner et al., 2020), exposure via aerosols is obviously more relevant for in vitro models representing the lung. A very specific approach to solve the buoyancy problem was introduced by Green et al. (1998). The authors embedded buoyant PE particles in the surface layer of agarose and seeded primary macrophages on top. However, due to the immobility of particles, this approach is only useful for phagocytizing cell models and not applicable for models of the intestinal or lung epithelium. A completely different approach to test buoyant polymers in vitro is the modification of the material to become denser. For example, mineral talc is a commonly used filler for PP to increase the materials’ performance (Ammar et al., 2017), which also increases the density of PP.
Differences in applied dose and effective dose. Apart from buoyant polymers, the low density of non-buoyant plastic particles needs to be considered as well. Polymers like PS are only marginally denser than cell culture medium and therefore exhibit very slow sedimentation rates. The amount of particles interacting with the cells (effective or delivered dose) is often small compared to the total amount of particles added to the in vitro system (applied dose). This may be further aggravated by particle agglomeration in the cell culture medium, which further decreases the density of the agglomerate (Hinderliter et al., 2010). Therefore, considerations of sedimentation kinetics for in vitro nanotoxicology research is important to avoid misinterpretation of concentration-dependent cellular response and uptake data (Teeguarden et al., 2007). In silico dosimetry models have been developed to predict the delivered dose based on the physicochemical properties of the particles and the setup of the in vitro exposure scenario. Currently, three main tools are used to estimate the delivered dose during in vitro experiments; the In vitro Sedimentation, Diffusion and Dosimetry model (ISDD), the In vitro Sedimentation, Diffusion, Dissolution and Dosimetry model (ISD3) and the Distorted Grid model (DG) (Hinderliter et al., 2010; DeLoid et al., 2015; Thomas et al., 2018). In most cases, the ISDD model is sufficient for predicting nanoplastics dosimetry. However, ISD3 and DG are better suited for modelling buoyant particles such as PE or PP, as well as polydisperse or degradable materials (Watson et al., 2016). As an example for a severe discrepancy between the applied dose and the effective dose that interacts with cells, ISDD sedimentation modelling of 50 nm PS nanoparticles showed only 17% deposition after 24 h at relevant in vitro conditions (Busch et al., 2021a). Contrastingly, heavier, metal-based engineered nanomaterials like silver nanoparticles of comparable size can sediment completely within 4 h under certain conditions (Kämpfer et al., 2021). Cell culture models grown under dynamic flow conditions to improve differentiation (Sargent et al., 2010) or implemented in microfluidic chips (Zhang et al., 2017; Kulthong et al., 2021) might improve the distribution of nanoplastics, increasing the correspondence between the applied and delivered dose. However, in silico dosimetry models are not yet equipped to predict nanoparticle sedimentation under dynamic flow conditions.
Adherence of macromolecules. Particles suspended in biological media like cell culture medium will acquire a biomolecular corona, i.e. proteins, lipids and carbohydrates, that adhere to the particles’ surface (Docter et al., 2015). As the adherence of these macromolecules will change the surface properties of the particles and therefore their interaction with biological systems (Monopoli et al., 2012; Cao et al., 2022), the possible role of the corona has to be considered during in vitro testing (Allen et al., 2006). For example, Abdelkhaliq et al. (2018) analyzed the protein corona on different PS nanoplastics via LC-MS/MS and reported differences based on the surface modification of the particles, which in turn impacted the uptake rates of the nanoplastics.
A physiologically relevant exposure should include the history of the particles, for example a simulated gastrointestinal digestion, or incubation in lung fluids. During in vitro experiments, nanoplastics are commonly applied in cell culture medium containing fetal calf serum (FCS), leading to adherence of serum proteins to the nanoplastic surface (Abdelkhaliq et al., 2018). Contrastingly, the corona of ingested nanoplastics will more likely consist of macromolecules from the food matrix and digestive fluids, while inhaled nanoplastics will exhibit a corona that may be acquired from exogenous environmental matrices (e.g. microbial components, semivolatile organic compounds) and the obvious endogenous molecules present in lung surfactant (Griese, 1999; Ichinose et al., 2005).
Interference with test assays and readouts. Besides changing and defining the surface identity, the adsorption of macromolecules, in particular target proteins of interest, to the surface of plastic particles, might distort the outcome of applied in vitro assays. Binding of extracellular lactate dehydrogenase (LDH), a common marker of membrane damage, might reduce the enzymes activity during the LDH assay, as it was reported for silver nanoparticles (Oh et al., 2014; Liang et al., 2015). Moreover, interference of amine-modified PS with the interleukin (IL)-8 ELISA was observed in the form of exaggerated IL-8 concentrations (Busch et al., 2022a). Therefore, assessing potential interferences of the tested materials with the used assays is necessary prior to toxicological investigations (Stone et al., 2009; Wilhelmi et al., 2012; Ong et al., 2014).
Use of fluorescently labeled nanoplastics. Often, it is important to know to what degree particles are internalized by target cells. In contrast to metal-based nanoparticles, where techniques like ICP-MS can be used to measure the cellular uptake of unlabeled particles (Mitrano et al., 2012), no equivalent techniques are currently available for polymeric particles. Instead, fluorescently labeled nanoplastics are commonly used to monitor the potency of cellular uptake, which can be quantified using confocal microscopy or flow cytometry. The use of fluorescent labels, while seemingly simple, require great care to avoid artifacts during the measurements. For instance, it is reported that fluorophores may leach out of the plastic particles, causing higher fluorescent signals in cells, or fluorescence in cell compartments not commonly available to nanoplastics (Tenuta et al., 2011; Catarino et al., 2019). Additionally, fluorophores might be pH-sensitive, leading to alterations in signal strength in subcellular compartments like lysosomes (Simonsen and Kromann, 2021), complicating interpretation of data. Another issue of using fluorescent particles is the inability to distinguish between internalization and surface adhesion of particles. Flow cytometry is commonly used to assess particle internalization, as it offers higher throughput than confocal microscopy and is more easily quantifiable. However, flow cytometry is unable to distinguish between fluorescence originating from within cells or from cell surfaces, necessitating further experiments to confirm actual internalization. This can be achieved by using dyes that are specifically quenched at the cell surface (Nuutila and Lilius, 2005; Dumont et al., 2017), by validating and correcting flow cytometry with confocal microscopy data (Gottstein et al., 2013), or by using a modified setup such as imaging flow cytometry (Phanse et al., 2012; Smirnov et al., 2015). Issues surrounding the use of fluorescence to determine nanoparticle internalization have received increased scientific interest and have been discussed in great detail in a recent review (FitzGerald and Johnston, 2021).
Size limitations depending on the exposed organ. Unlike engineered nanomaterials, environmental plastic particles do not have a clear size cutoff, covering a whole size spectrum from sub-nano size to several millimeters. In the case of ingested particles, size limitations only play a minor role, as even centimeter-sized plastics can be swallowed and are able to pass the human gastrointestinal tract. Therefore, the intestinal epithelium can, in theory, be exposed to particles of enormous size, justifying the testing of larger particles. For example, Lehner et al. (2020) applied microplastics up to 500 µm in an intestinal in vitro model. In contrast, the size of inhaled particles is strictly limited in the context of lung anatomy and physiology. Particles with an aerodynamic diameter of less than 10 µm are able to deposit in the tracheobronchial region, while only particles less than approximately 2.5 µm are small enough to reach the alveoli (Oberdorster et al., 2005; Xing et al., 2016). Therefore, when testing plastic particles in an in vitro model of the human respiratory system, the size of the test material has to be chosen accordingly.
Relevance of available model particles. When looking at available scientific literature regarding toxicological investigations of plastic particles, the majority of studies is carried out using spherical PS particles of a defined size. However, there is a strong discrepancy between available data and environmental occurrence: PS makes up only 6.1% of globally produced polymers (www.plasticseurope.org) and shapes of micro- and nanoplastics have been described to be primarily fibers, fragments and sheets (Hongprasith et al., 2020). Furthermore, plastic particles in the environment cover a whole size spectrum that might exhibit synergistic effects during co-exposure of differently sized particles (Liang et al., 2021). While PS spheres are widely commercially available, particles of other polymers or shapes are very limited. Currently, research groups mostly rely on producing their own model plastics via methods like laser ablation, cryocutting or milling (Magrì et al., 2018; Lionetto et al., 2021; Busch et al., 2022a), which result in much more relevant polymer types, shapes and size distributions, but might limit the comparability between different studies. The urgent need for standardized, widely available reference particles of relevant polymers, sizes and shapes has already been repeatedly expressed in recent reviews (Gouin et al., 2019; Brachner et al., 2020; Halappanavar and Mallach, 2021).
Chemicals in nanoplastic samples. Commercial nanoplastic suspensions are often used as model particles for the hazard assessment of plastic particles in toxicological studies. These suspensions might contain additives like preservatives, antimicrobials or surfactants, which may cause artifacts in toxicity tests. For example, the preservative sodium azide that was present in a commercial PS suspension was found to be the cause of acute toxicity towards Daphnia magna, instead of the PS particles themselves (Pikuda et al., 2019). Furthermore, nanoplastics produced from bulk plastic and/or plastic products might contain additives and unpolymerized monomers, like bisphenol A, heavy metals or styrene (Ajaj et al., 2021; Catrouillet et al., 2021; Gulizia et al., 2023). These chemicals might be released from the nanoplastics during incubation in the in vitro culture medium or in the cells that have taken up the plastic particles (Peters et al., 2022). To prevent incorrect interpretation of nanoplastics in vitro data and to distinguish between actual particle effects and effects stemming from associated chemicals, it is recommended to include a filtrate control in experiments (Petersen et al., 2022).
4 Conclusion and future outlook
Currently developed (advanced) in vitro models, due to their increased physiological relevance for human hazard assessment and suitability for in-depth MoA studies, are well positioned for contributing to solve the complex issues surrounding nanotoxicity. Different in vitro models are suited to tackle different challenges and depending on the research question, stem cell-based models might be a suitable approach for the investigation of nanoplastics uptake and effects in the primarily exposed organs intestine and lung, as they offer specific advantages over traditional immortalized cell lines.
Common knowledge derived from in vitro models includes the prediction of absorption rates, the derivation of MoAs and the estimation of potency by relating in vitro endpoints to in vivo endpoints (Bernauer et al., 2005). The derivation of MoAs highly depends on the physiological relevance of the model and partly on the cell types present. Conventional cell models benefit from a high relative robustness compared to stem cell models and are comparatively easy to culture, which may lead to a higher interlaboratory comparability and more accurate estimation of the benchmark dose (Black et al., 2022) when comparing different nanoparticles. This is especially true when comparing results found in different stem cell-based models, where a lack of consistency could imaginably complicate risk assessment. However, in the case of cellular processes altered due to the disease status of cancer cell lines, including genotoxicity, proliferation and cytokine production/response (Kauffman et al., 2013), iPSCs may be more relevant to study the real world situation. Similarly, when considering omics-based MoA screening approaches, the multitude of mutations commonly seen in cancer cell lines (Hartung and Daston, 2009) can alter RNA or protein expression and can mask the true effect of many small molecule compounds (Ben-David et al., 2018) and potentially also nanoplastics. Especially when using approaches such as systems toxicology (Hartung et al., 2012), small alterations in e.g. transcriptomics might help identifying important early signaling events at low exposure concentrations that might only lead to detectable toxicity at prolonged or repeated exposure. Furthermore, while the lack of robustness of stem cell-based models can be seen as a demerit, these models do allow for the estimation of relative sensitivity of subpopulations of cells or individuals with a particular genetic background. Such information can be especially relevant for risk assessment as this reduces the need for potentially inaccurate scaling factors. While in vivo studies are commonly used to derive the lower confidence limit of the benchmark dose (BMDL) and organ concentrations, special care must be taken when interpreting these results. The saying “mice are not men” (Warren et al., 2015) especially holds true regarding nanoparticles where differences in physical barriers including the amount of Peyers patches and M-cells, as well as the structure of the mucus layer in intestine and lung can cause further discrepancies between the human and animal situation (Kararli, 1995; Fagerholm et al., 1996; Ermund et al., 2013).
While the methodologies to scale in vitro nanoparticle toxicokinetics to in vivo kinetics are in the explorative phase, they are the only conceivable, consistent source of human toxicokinetic data. When considering in vitro models for the possibility of extrapolation from in vitro to in vivo (i.e. quantitative in vitro to in vivo extrapolations or QIVIVE), stem cell-based models seem to be favored as these consist of non-diseased tissue made up of the relevant cell types found in vivo. In view of replacing animal experiments, efforts have already been made to apply physiologically based kinetic (PBK) models to nanomaterials in order to extrapolate in vitro uptake data to the in vivo situation (Brouwer et al., 2023). Although the acceptance of PBK model data in a regulatory context is still limited (Lamon et al., 2019), using input data from physiologically relevant stem cell-based in vitro models might facilitate the development in this direction.
The identification of key events leading to adverse health outcomes and subsequent implementation in an adverse-outcome-pathway (AOP) frame is currently explored for nanomaterials. These AOPs, and associated key events, might aid in selecting relevant in vitro assays. It is expected that commonly used in vitro assays can be used, however some emphasis to typical “nano”-related endpoints are foreseen, such as cellular uptake, membrane damage, ROS generation or release of pro-inflammatory cytokines (Jagiello and Ciura, 2022). However, the molecular initiating events (MIE) of nanoparticles leading to toxicity are often unspecific, in direct contrast to the MIE of a chemical that binds to a receptor or target molecule (Halappanavar et al., 2020). Therefore, translation of in vitro effects to the in vivo situation in the case of nanomaterials including nanoplastics still requires extensive development, but will greatly benefit from in vitro models that closely mimic the in vivo situation, such as the stem cell-based models discussed in this review.
Despite the unique features that qualify stem cell-based models as a promising approach for future nanotoxicity studies, there are some limitations that can make the use of these models challenging. The cultivation and differentiation of stem cells into a mature model usually covers time spans of two weeks or more for a single experiment (Kabeya et al., 2018; Naumovska et al., 2020), and is quite expensive in terms of consumption of medium and the multitude of growth factors required for differentiation. The handling of stem cells requires highly trained operators, since wrong handling can easily lead to death or spontaneous differentiation of stem cells, e.g., by suboptimal cell confluence or inappropriate passaging (Kogut et al., 2014; Castro-Viñuelas et al., 2021; Yamamoto et al., 2022), which drastically impairs the robustness of the model itself. This issue of robustness is further supported by the lack of standardized protocols for stem cell-based models in the current literature.
Another current limitation of both cell line and stem cell-based models is the lack of immunocompetent cells, as immune cells like macrophages or neutrophils are derived from hematopoietic stem cells in the bone marrow, instead of tissue-resident stem cells, and transported via the blood (Epelman et al., 2014; Leiding, 2017). Organ models based on stem cells exclusively consist of the cell types that stem from the respective tissue-resident stem cells. Yet, pro-inflammatory effects appear to be one of the main mechanisms of nanoplastics toxicity, as they have been described in several in vivo studies (Choi et al., 2021; Sun et al., 2021; He et al., 2022; Tang et al., 2022). Similarly, nanoplastics have been shown to interact with components of the nervous system (Jung et al., 2020), which stem cell-based models of intestine and lung are currently lacking. However, the generation of immune and neural cells from iPSCs has been reported (McKinney, 2017; Trump et al., 2019; Gutbier et al., 2020) and the co-culture of mature stem cell-based models with additional cells is possible, e.g. intestinal organoids with an enteric nervous system (Noel et al., 2017; Takahashi et al., 2017; Workman et al., 2017; de Rus Jacquet, 2019; Choi et al., 2022). However this has, to the best of our knowledge, not been applied in the field of particle toxicology yet.
The limitations of stem cell-based models, such as the reduced robustness and the lack of standardized protocols might eventually be overcome, as the field develops rapidly and new insights are gained frequently. Similarly, the implementation of other relevant cell types into these models will be the next step towards highly versatile alternatives to animal testing, as it will be possible to investigate immune- or nervous system-related effects of nanoplastics in a physiologically relevant in vitro environment.
Lastly, recreating dynamic conditions as present in the intestine and lung by creating in vitro models in microfluidic devices (i.e. organ-on-a-chip) might also help to tackle some specific particle-related challenges such as the buoyancy issue. Initial work using a microfluidic-chip model has been performed by exposing human bronchial BEAS-2B cells to PS (Gupta et al., 2021). By generating scaffold-like (Shim et al., 2017) or tubular structures (Naumovska et al., 2020) inside of microfluidic devices that are exposed to a dynamic flow of particle suspension, the sedimentation behavior of buoyant particles will no longer be an obstacle when testing polymers like PP or PE, allowing more accurate dosimetry estimates in intestinal or non-ALI lung models. The adaption of protocols and settings of the microfluidic devices for the purpose of particle testing will require extensive optimization, such as flow rates, exposure time and exact dosimetry calculations. However, the successful integration of such advanced models into the portfolio of validated methods of in vitro nanotoxicology will greatly enhance the field.
Author contributions
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
Funding
This study was supported by the Netherlands Organization for Health Research and Development (ZonMw), within the MOMENTUM (Microplastics and Human Health Consortium) project (grant no. 458001101) and has received funding from the Plasticheal project under European Union’s Horizon 2020 research and innovation programme (grant no. 965196). This publication is also part of the LymphChip project (number NWA-ORC 2019 1292.19.019) of the NWA research program ‘Research on Routes by Consortia (ORC)’, which is funded by the Netherlands Organization for Scientific Research (NWO). The work was also supported by the Leibniz Research Alliance Advanced Materials Safety.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abass, K., Lämsä, V., Reponen, P., Küblbeck, J., Honkakoski, P., Mattila, S., et al. (2012). Characterization of human cytochrome P450 induction by pesticides. Toxicology 294 (1), 17–26. doi:10.1016/j.tox.2012.01.010
Abdelkhaliq, A., van der Zande, M., Punt, A., Helsdingen, R., Boeren, S., Vervoort, J. J. M., et al. (2018). Impact of nanoparticle surface functionalization on the protein corona and cellular adhesion, uptake and transport. J. Nanobiotechnology 16 (1), 70. doi:10.1186/s12951-018-0394-6
Abud, E. M., Ramirez, R. N., Martinez, E. S., Healy, L. M., Nguyen, C. H. H., Newman, S. A., et al. (2017). iPSC-derived human microglia-like cells to study neurological diseases. Neuron 94 (2), 278–293. doi:10.1016/j.neuron.2017.03.042
Adler, K. B., Cheng, P.-W., and Kim, K. C. (1990). Characterization of Guinea pig tracheal epithelial cells maintained in biphasic organotypic culture: Cellular composition and biochemical analysis of released glycoconjugates. Am. J. Respir. Cell Mol. Biol. 2 (2), 145–154. doi:10.1165/ajrcmb/2.2.145
Ahmed, U., Ahmed, R., Masoud, M. S., Tariq, M., Ashfaq, U. A., Augustine, R., et al. (2021). Stem cells based in vitro models: Trends and prospects in biomaterials cytotoxicity studies. Biomed. Mater 16 (4), 042003. doi:10.1088/1748-605X/abe6d8
Ajaj, A., J'Bari, S., Ononogbo, A., Buonocore, F., Bear, J. C., Mayes, A. G., et al. (2021). An insight into the growing concerns of styrene monomer and poly(styrene) fragment migration into food and drink simulants from poly(styrene) packaging. Foods 10 (5), 1136. doi:10.3390/foods10051136
Allen, L. T., Tosetto, M., Miller, I. S., O’Connor, D. P., Penney, S. C., Lynch, I., et al. (2006). Surface-induced changes in protein adsorption and implications for cellular phenotypic responses to surface interaction. Biomaterials 27 (16), 3096–3108. doi:10.1016/j.biomaterials.2006.01.019
Ammar, O., Bouaziz, Y., Haddar, N., and Mnif, N. (2017). Talc as reinforcing filler in polypropylene compounds: Effect on morphology and mechanical properties. Polym. Sci. 3 (2). doi:10.4172/2471-9935.100023
Andrady, A. L., and Rajapakse, N. (2016). “Additives and chemicals in plastics,” in Hazardous chemicals associated with plastics in the marine environment. The handbook of environmental chemistry. Editors H. Takada, and H. Karapanagioti (Cham): Springer).
Bandi, S. P., Kumbhar, Y. S., and Venuganti, V. V. K. (2020). Effect of particle size and surface charge of nanoparticles in penetration through intestinal mucus barrier. J. Nanoparticle Res. 22 (3), 62. doi:10.1007/s11051-020-04785-y
Basil, M. C., Katzen, J., Engler, A. E., Guo, M., Herriges, M. J., Kathiriya, J. J., et al. (2020). The cellular and physiological basis for lung repair and regeneration: Past, present, and future. Cell Stem Cell 26 (4), 482–502. doi:10.1016/j.stem.2020.03.009
Belair, D. G., Visconti, R. J., Hong, M., Marella, M., Peters, M. F., Scott, C. W., et al. (2020). Human ileal organoid model recapitulates clinical incidence of diarrhea associated with small molecule drugs. Toxicol Vitro 68, 104928. doi:10.1016/j.tiv.2020.104928
Ben-David, U., Siranosian, B., Ha, G., Tang, H., Oren, Y., Hinohara, K., et al. (2018). Genetic and transcriptional evolution alters cancer cell line drug response. Nature 560 (7718), 325–330. doi:10.1038/s41586-018-0409-3
Bengalli, R., Zerboni, A., Bonfanti, P., Saibene, M., Mehn, D., Cella, C., et al. (2022). Characterization of microparticles derived from waste plastics and their bio-interaction with human lung A549 cells. J. Appl. Toxicol. 42, 2030–2044. doi:10.1002/jat.4372
Bernauer, U., Oberemm, A., Madle, S., and Gundert-Remy, U. (2005). The use of in vitro data in risk assessment. Basic Clin. Pharmacol. Toxicol. 96 (3), 176–181. doi:10.1111/j.1742-7843.2005.pto960306.x
Binder, S., Cao, X., Bauer, S., Rastak, N., Kuhn, E., Dragan, G. C., et al. (2021). In vitro genotoxicity of dibutyl phthalate on A549 lung cells at air–liquid interface in exposure concentrations relevant at workplaces. Environ. Mol. Mutagen 62 (9), 490–501. doi:10.1002/em.22464
Black, M. B., Stern, A., Efremenko, A., Mallick, P., Moreau, M., Hartman, J. K., et al. (2022). Biological system considerations for application of toxicogenomics in next-generation risk assessment and predictive toxicology. Toxicol Vitro 80, 105311. doi:10.1016/j.tiv.2022.105311
Blank, F., Rothen-Rutishauser, B. M., Schurch, S., and Gehr, P. (2006). An optimized in vitro model of the respiratory tract wall to study particle cell interactions. J. Aerosol Med. 19 (3), 392–405. doi:10.1089/jam.2006.19.392
Bouwmeester, H., Hollman, P. C., and Peters, R. J. (2015). Potential health impact of environmentally released micro- and nanoplastics in the human food production chain: Experiences from nanotoxicology. Environ. Sci. Technol. 49 (15), 8932–8947. doi:10.1021/acs.est.5b01090
Brachner, A., Fragouli, D., Duarte, I. F., Farias, P. M. A., Dembski, S., Ghosh, M., et al. (2020). Assessment of human health risks posed by nano-and microplastics is currently not feasible. Int. J. Environ. Res. Public Health 17 (23), 8832. doi:10.3390/ijerph17238832
Breau, K. A., Ok, M. T., Gomez-Martinez, I., Burclaff, J., Kohn, N. P., and Magness, S. T. (2022). Efficient transgenesis and homology-directed gene targeting in monolayers of primary human small intestinal and colonic epithelial stem cells. Stem Cell Rep. 17 (6), 1493–1506. doi:10.1016/j.stemcr.2022.04.005
Brouwer, H., Van Oijen, F. L. N., and Bouwmeester, H. (2023). “Chapter 40 - potential human health effects following exposure to nano- and microplastics, lessons learned from nanomaterials,” in Present knowledge in food safety. Editors M. E. Knowles, L. E. Anelich, A. R. Boobis, and B. Popping (Academic Press), 590–605. doi:10.1016/B978-0-12-819470-6.00014-7
Brown, D. M., Wilson, M. R., MacNee, W., Stone, V., and Donaldson, K. (2001). Size-dependent proinflammatory effects of ultrafine polystyrene particles: A role for surface area and oxidative stress in the enhanced activity of ultrafines. Toxicol. Appl. Pharmacol. 175 (3), 191–199. doi:10.1006/taap.2001.9240
Bruder, S. P., Jaiswal, N., and Haynesworth, S. E. (1997). Growth kinetics, self-renewal, and the osteogenic potential of purified human mesenchymal stem cells during extensive subcultivation and following cryopreservation. J. Cell Biochem. 64 (2), 278–294. doi:10.1002/(sici)1097-4644(199702)64:2<278::aid-jcb11>3.0.co;2-f
Busch, M., Bredeck, G., Kampfer, A. A. M., and Schins, R. P. F. (2021a). Investigations of acute effects of polystyrene and polyvinyl chloride micro- and nanoplastics in an advanced in vitro triple culture model of the healthy and inflamed intestine. Environ. Res. 193, 110536. doi:10.1016/j.envres.2020.110536
Busch, M., Bredeck, G., Waag, F., Rahimi, K., Ramachandran, H., Bessel, T., et al. (2022a). Assessing the NLRP3 inflammasome activating potential of a large panel of micro- and nanoplastics in THP-1 cells. Biomolecules 12 (8), 1095. doi:10.3390/biom12081095
Busch, M., Kämpfer, A. A. M., and Schins, R. P. F. (2021b). An inverted in vitro triple culture model of the healthy and inflamed intestine: Adverse effects of polyethylene particles. Chemosphere 284, 131345. doi:10.1016/j.chemosphere.2021.131345
Busch, M., Ramachandran, H., Wahle, T., Rossi, A., and Schins, R. P. F. (2022b). Investigating the role of the NLRP3 inflammasome pathway in acute intestinal inflammation: Use of THP-1 knockout cell lines in an advanced triple culture model. Front. Immunol. 13, 898039. doi:10.3389/fimmu.2022.898039
Cai, Y., Mitrano, D. M., Hufenus, R., and Nowack, B. (2021). Formation of fiber fragments during abrasion of polyester textiles. Environ. Sci. Technol. 55 (12), 8001–8009. doi:10.1021/acs.est.1c00650
Cao, J., Yang, Q., Jiang, J., Dalu, T., Kadushkin, A., Singh, J., et al. (2022). Coronas of micro/nano plastics: A key determinant in their risk assessments. Part Fibre Toxicol. 19 (1), 55. doi:10.1186/s12989-022-00492-9
Cao, L., Gibson, J. D., Miyamoto, S., Sail, V., Verma, R., Rosenberg, D. W., et al. (2011). Intestinal lineage commitment of embryonic stem cells. Differentiation 81 (1), 1–10. doi:10.1016/j.diff.2010.09.182
Casals, E., Pfaller, T., Duschl, A., Oostingh, G. J., and Puntes, V. (2010). Time evolution of the nanoparticle protein corona. ACS Nano 4 (7), 3623–3632. doi:10.1021/nn901372t
Castro-Viñuelas, R., Sanjurjo-Rodríguez, C., Piñeiro-Ramil, M., Rodríguez-Fernández, S., López-Baltar, I., Fuentes-Boquete, I., et al. (2021). Tips and tricks for successfully culturing and adapting human induced pluripotent stem cells. Mol. Ther. Methods Clin. Dev. 23, 569–581. doi:10.1016/j.omtm.2021.10.013
Catarino, A. I., Frutos, A., and Henry, T. B. (2019). Use of fluorescent-labelled nanoplastics (NPs) to demonstrate NP absorption is inconclusive without adequate controls. Sci. Total Environ. 670, 915–920. doi:10.1016/j.scitotenv.2019.03.194
Catrouillet, C., Davranche, M., Khatib, I., Fauny, C., Wahl, A., and Gigault, J. (2021). Metals in microplastics: Determining which are additive, adsorbed, and bioavailable. Environ. Sci. Process Impacts 23 (4), 553–558. doi:10.1039/d1em00017a
Choi, S., Choi, J., Cheon, S., Song, J., Kim, S.-Y., Kim, J.-e., et al. (2022). Pulmonary fibrosis model using micro-CT analyzable human PSC–derived alveolar organoids containing alveolar macrophage-like cells. Cell Biol. Toxicol. 38 (4), 557–575. doi:10.1007/s10565-022-09698-1
Choi, Y. J., Park, J. W., Lim, Y., Seo, S., and Hwang, D. Y. (2021). In vivo impact assessment of orally administered polystyrene nanoplastics: Biodistribution, toxicity, and inflammatory response in mice. Nanotoxicology 15 (9), 1180–1198. doi:10.1080/17435390.2021.1996650
Co, J. Y., Margalef-Català, M., Li, X., Mah, A. T., Kuo, C. J., Monack, D. M., et al. (2019). Controlling epithelial polarity: A human enteroid model for host-pathogen interactions. Cell Rep. 26 (9), 2509–2520. doi:10.1016/j.celrep.2019.01.108
Coffin, S., Bouwmeester, H., Brander, S., Damdimopoulou, P., Gouin, T., Hermabessiere, L., et al. (2022). Development and application of a health-based framework for informing regulatory action in relation to exposure of microplastic particles in California drinking water. Microplast nanoplast 2 (1), 12. doi:10.1186/s43591-022-00030-6
Coffin, S., Lee, I., Gan, J., and Schlenk, D. (2019). Simulated digestion of polystyrene foam enhances desorption of diethylhexyl phthalate (DEHP) and in vitro estrogenic activity in a size-dependent manner. Environ. Pollut. 246, 452–462. doi:10.1016/j.envpol.2018.12.011
Cortés, C., Domenech, J., Salazar, M., Pastor, S., Marcos, R., and Hernández, A. (2020). Nanoplastics as a potential environmental health factor: Effects of polystyrene nanoparticles on human intestinal epithelial caco-2 cells. Environ. Sci. Nano 7 (1), 272–285. doi:10.1039/c9en00523d
Cox, K. D., Covernton, G. A., Davies, H. L., Dower, J. F., Juanes, F., and Dudas, S. E. (2019). Human consumption of microplastics. Environ. Sci. Technol. 53 (12), 7068–7074. doi:10.1021/acs.est.9b01517
Craighead, J. E., Mossman, B. T., and Bradley, B. J. (1980). Comparative studies on the cytotoxicity of amphibole and serpentine asbestos. Environ. Health Perspect. 34, 37–46. doi:10.1289/ehp.803437
Crocker, T. T., O'Donnell, T. V., and Nunes, L. L. (1973). Toxicity of benzo( )pyrene and air pollution composite for adult human bronchial mucosa in organ culture. Cancer Res. 33 (1), 88–93.
da Silva Brito, W. A., Singer, D., Miebach, L., Saadati, F., Wende, K., Schmidt, A., et al. (2023). Comprehensive in vitro polymer type, concentration, and size correlation analysis to microplastic toxicity and inflammation. Sci. Total Environ. 854, 158731. doi:10.1016/j.scitotenv.2022.158731
Davidson, K., and Dudas, S. E. (2016). Microplastic ingestion by wild and cultured manila clams (venerupis philippinarum) from baynes sound, British columbia. Arch. Environ. Contam. Toxicol. 71 (2), 147–156. doi:10.1007/s00244-016-0286-4
de Rus Jacquet, A. (2019). Preparation and Co-culture of iPSC-derived dopaminergic neurons and astrocytes. Curr. Protoc. Cell Biol. 85 (1), e98. doi:10.1002/cpcb.98
de Souza Machado, A. A., Kloas, W., Zarfl, C., Hempel, S., and Rillig, M. C. (2018). Microplastics as an emerging threat to terrestrial ecosystems. Glob. Chang. Biol. 24 (4), 1405–1416. doi:10.1111/gcb.14020
DeLoid, G. M., Cohen, J. M., Pyrgiotakis, G., Pirela, S. V., Pal, A., Liu, J., et al. (2015). Advanced computational modeling for in vitro nanomaterial dosimetry. Part Fibre Toxicol. 12, 32. doi:10.1186/s12989-015-0109-1
Ding, Y., Weindl, P., Lenz, A.-G., Mayer, P., Krebs, T., and Schmid, O. (2020). Quartz crystal microbalances (QCM) are suitable for real-time dosimetry in nanotoxicological studies using VITROCELL®Cloud cell exposure systems. Part Fibre Toxicol. 17 (1), 44. doi:10.1186/s12989-020-00376-w
Docter, D., Strieth, S., Westmeier, D., Hayden, O., Gao, M., Knauer, S. K., et al. (2015). No king without a crown – impact of the nanomaterial-protein corona on nanobiomedicine. Nanomedicine 10 (3), 503–519. doi:10.2217/nnm.14.184
Domenech, J., Cortés, C., Vela, L., Marcos, R., and Hernández, A. (2021). Polystyrene nanoplastics as carriers of metals. Interactions of polystyrene nanoparticles with silver nanoparticles and silver nitrate, and their effects on human intestinal caco-2 cells. Biomolecules 11 (6), 859. doi:10.3390/biom11060859
Domenech, J., Hernandez, A., Rubio, L., Marcos, R., and Cortes, C. (2020). Interactions of polystyrene nanoplastics with in vitro models of the human intestinal barrier. Arch. Toxicol. 94 (9), 2997–3012. doi:10.1007/s00204-020-02805-3
Dong, C.-D., Chen, C.-W., Chen, Y.-C., Chen, H.-H., Lee, J.-S., and Lin, C.-H. (2020). Polystyrene microplastic particles: In vitro pulmonary toxicity assessment. J. Hazard Mater 385, 121575. doi:10.1016/j.jhazmat.2019.121575
Driehuis, E., Kretzschmar, K., and Clevers, H. (2020). Establishment of patient-derived cancer organoids for drug-screening applications. Nat. Protoc. 15 (10), 3380–3409. doi:10.1038/s41596-020-0379-4
Driscoll, K. E., Carter, J. M., Iype, P. T., Kumari, H. L., Crosby, L. L., Aardema, M. J., et al. (1995). Establishment of immortalized alveolar type II epithelial cell lines from adult rats. Vitro Cell Dev. Biol. Anim. 31 (7), 516–527. doi:10.1007/BF02634029
Drost, J., and Clevers, H. (2018). Organoids in cancer research. Nat. Rev. Cancer 18 (7), 407–418. doi:10.1038/s41568-018-0007-6
Dumont, C., Czuba, E., Chen, M., Villadangos, J. A., Johnston, A. P., and Mintern, J. D. (2017). DNA-based probes for flow cytometry analysis of endocytosis and recycling. Traffic 18 (4), 242–249. doi:10.1111/tra.12466
EFSA (2016). Presence of microplastics and nanoplastics in food, with particular focus on seafood. EFSA J. 14 (6), 4501. doi:10.2903/j.efsa.2016.4501
EFSA More, S., Bampidis, V., Benford, D., Bragard, C., Halldorsson, T., Hernández-Jerez, A., et al. (2021). Guidance on risk assessment of nanomaterials to be applied in the food and feed chain: Human and animal health. EFSA J. 19 (8), e06768. doi:10.2903/j.efsa.2021.6768
Enfrin, M., Lee, J., Gibert, Y., Basheer, F., Kong, L., and Dumée, L. F. (2020). Release of hazardous nanoplastic contaminants due to microplastics fragmentation under shear stress forces. J. Hazard Mater 384, 121393. doi:10.1016/j.jhazmat.2019.121393
Epelman, S., Lavine, K. J., and Randolph, G. J. (2014). Origin and functions of tissue macrophages. Immunity 41 (1), 21–35. doi:10.1016/j.immuni.2014.06.013
Ermund, A., Schütte, A., Johansson, M. E. V., Gustafsson, J. K., and Hansson, G. C. (2013). Studies of mucus in mouse stomach, small intestine, and colon. I. Gastrointestinal mucus layers have different properties depending on location as well as over the Peyer's patches. Am. J. Physiol. Gastrointest. Liver Physiol. 305 (5), 341–347. doi:10.1152/ajpgi.00046.2013
Fagerholm, U., Johansson, M., and Lennernäs, H. (1996). Comparison between permeability coefficients in rat and human jejunum. Pharm. Res. 13 (9), 1336–1342. doi:10.1023/A:1016065715308
Felipe-Rodriguez, M., Bohm, G., and Doran, R. (2022). What does the public think about microplastics? Insights from an empirical analysis of mental models elicited through free associations. Front. Psychol. 13, 920454. doi:10.3389/fpsyg.2022.920454
FitzGerald, L. I., and Johnston, A. P. R. (2021). It’s what’s on the inside that counts: Techniques for investigating the uptake and recycling of nanoparticles and proteins in cells. J. Colloid Interface Sci. 587, 64–78. doi:10.1016/j.jcis.2020.11.076
Florance, I., Chandrasekaran, N., Gopinath, P. M., and Mukherjee, A. (2022). Exposure to polystyrene nanoplastics impairs lipid metabolism in human and murine macrophages in vitro. Ecotoxicol. Environ. Saf. 238, 113612. doi:10.1016/j.ecoenv.2022.113612
Forster, R., Chiba, K., Schaeffer, L., Regalado, S. G., Lai, C. S., Gao, Q., et al. (2014). Human intestinal tissue with adult stem cell properties derived from pluripotent stem cells. Stem Cell Rep. 2 (6), 838–852. doi:10.1016/j.stemcr.2014.05.001
Fournier, E., Leveque, M., Ruiz, P., Ratel, J., Durif, C., Chalancon, S., et al. (2023). Microplastics: What happens in the human digestive tract? First evidences in adults using in vitro gut models. J. Hazard Mater 442, 130010. doi:10.1016/j.jhazmat.2022.130010
Fritsche, E., Haarmann-Stemmann, T., Kapr, J., Galanjuk, S., Hartmann, J., Mertens, P. R., et al. (2021). Stem cells for next level toxicity testing in the 21st century. Small 17 (15), 2006252. doi:10.1002/smll.202006252
Fritz, A., Busch, D., Lapczuk, J., Ostrowski, M., Drozdzik, M., and Oswald, S. (2019). Expression of clinically relevant drug-metabolizing enzymes along the human intestine and their correlation to drug transporters and nuclear receptors: An intra-subject analysis. Basic Clin. Pharmacol. Toxicol. 124 (3), 245–255. doi:10.1111/bcpt.13137
Fröhlich, E. (2018). Comparison of conventional and advanced in vitro models in the toxicity testing of nanoparticles. Artif. Cells Nanomed Biotechnol. 46, 1091–1107. doi:10.1080/21691401.2018.1479709
Gautam, R., Jo, J., Acharya, M., Maharjan, A., Lee, D., KC, P. B., et al. (2022). Evaluation of potential toxicity of polyethylene microplastics on human derived cell lines. Sci. Total Environ. 838, 156089. doi:10.1016/j.scitotenv.2022.156089
Gerbe, F., Sidot, E., Smyth, D. J., Ohmoto, M., Matsumoto, I., Dardalhon, V., et al. (2016). Intestinal epithelial tuft cells initiate type 2 mucosal immunity to helminth parasites. Nature 529 (7585), 226–230. doi:10.1038/nature16527
Geyer, R., Jambeck, J. R., and Law, K. L. (2017). Production, use, and fate of all plastics ever made. Sci. Adv. 3 (7), e1700782. doi:10.1126/sciadv.1700782
Goodman, K. E., Hare, J. T., Khamis, Z. I., Hua, T., and Sang, Q. A. (2021). Exposure of human lung cells to polystyrene microplastics significantly retards cell proliferation and triggers morphological changes. Chem. Res. Toxicol. 34 (4), 1069–1081. doi:10.1021/acs.chemrestox.0c00486
Gopinath, P. M., Twayana, K. S., Ravanan, P., John, T., Mukherjee, A., Jenkins, D. F., et al. (2021). Prospects on the nano-plastic particles internalization and induction of cellular response in human keratinocytes. Part Fibre Toxicol. 18 (1), 35. doi:10.1186/s12989-021-00428-9
Gotoh, S., Ito, I., Nagasaki, T., Yamamoto, Y., Konishi, S., Korogi, Y., et al. (2014). Generation of alveolar epithelial spheroids via isolated progenitor cells from human pluripotent stem cells. Stem Cell Rep. 3 (3), 394–403. doi:10.1016/j.stemcr.2014.07.005
Gottstein, C., Wu, G., Wong, B. J., and Zasadzinski, J. A. (2013). Precise quantification of nanoparticle internalization. ACS Nano 7 (6), 4933–4945. doi:10.1021/nn400243d
Gouin, T., Becker, R. A., Collot, A. G., Davis, J. W., Howard, B., Inawaka, K., et al. (2019). Toward the development and application of an environmental risk assessment framework for microplastic. Environ. Toxicol. Chem. 38 (10), 2087–2100. doi:10.1002/etc.4529
Gouin, T., Ellis-Hutchings, R., Thornton Hampton, L. M., Lemieux, C. L., and Wright, S. L. (2022). Screening and prioritization of nano- and microplastic particle toxicity studies for evaluating human health risks - development and application of a toxicity study assessment tool. Microplast nanoplast 2 (1), 2. doi:10.1186/s43591-021-00023-x
Green, T. R., Fisher, J., Stone, M., Wroblewski, B. M., and Ingham, E. (1998). Polyethylene particles of a 'critical size' are necessary for the induction of cytokines by macrophages in vitro. Biomaterials 19, 2297–2302. doi:10.1016/S0142-9612(98)00140-9
Griese, M. (1999). Pulmonary surfactant in health and human lung diseases: State of the art. Eur. Respir. J. 13 (6), 1455–1476. doi:10.1183/09031936.99.13614779
Grouls, M., Janssen, A. W. F., Duivenvoorde, L. P. M., Hooiveld, G. J. E. J., Bouwmeester, H., and van der Zande, M. (2022). Differential gene expression in iPSC-derived human intestinal epithelial cell layers following exposure to two concentrations of butyrate, propionate and acetate. Sci. Rep. 12 (1), 13988. doi:10.1038/s41598-022-17296-8
Gulizia, A. M., Patel, K., Philippa, B., Motti, C. A., van Herwerden, L., and Vamvounis, G. (2023). Understanding plasticiser leaching from polystyrene microplastics. Sci. Total Environ. 857, 159099. doi:10.1016/j.scitotenv.2022.159099
Gupta, G., Vallabani, S., Bordes, R., Bhattacharya, K., and Fadeel, B. (2021). Development of microfluidic, serum-free bronchial epithelial cells-on-a-chip to facilitate a more realistic in vitro testing of nanoplastics. Front. Toxicol. 3, 735331. doi:10.3389/ftox.2021.735331
Gutbier, S., Wanke, F., Dahm, N., Rümmelin, A., Zimmermann, S., Christensen, K., et al. (2020). Large-scale production of human iPSC-derived macrophages for drug screening. Int. J. Mol. Sci. 21 (13), 4808. doi:10.3390/ijms21134808
Hahladakis, J. N., Velis, C. A., Weber, R., Iacovidou, E., and Purnell, P. (2018). An overview of chemical additives present in plastics: Migration, release, fate and environmental impact during their use, disposal and recycling. J. Hazard Mater 344, 179–199. doi:10.1016/j.jhazmat.2017.10.014
Halappanavar, S., and Mallach, G. (2021). Adverse outcome pathways and in vitro toxicology strategies for microplastics hazard testing. Curr. Opin. Toxicol. 28, 52–61. doi:10.1016/j.cotox.2021.09.002
Halappanavar, S., van den Brule, S., Nymark, P., Gate, L., Seidel, C., Valentino, S., et al. (2020). Adverse outcome pathways as a tool for the design of testing strategies to support the safety assessment of emerging advanced materials at the nanoscale. Part Fibre Toxicol. 17 (1), 16. doi:10.1186/s12989-020-00344-4
Halimu, G., Zhang, Q., Liu, L., Zhang, Z., Wang, X., Gu, W., et al. (2022). Toxic effects of nanoplastics with different sizes and surface charges on epithelial-to-mesenchymal transition in A549 cells and the potential toxicological mechanism. J. Hazard Mater 430, 128485. doi:10.1016/j.jhazmat.2022.128485
Hartmann, N. B., Huffer, T., Thompson, R. C., Hassellov, M., Verschoor, A., Daugaard, A. E., et al. (2019). Are we speaking the same language? Recommendations for a definition and categorization framework for plastic debris. Environ. Sci. Technol. 53 (3), 1039–1047. doi:10.1021/acs.est.8b05297
Hartung, T., and Daston, G. (2009). Are in vitro tests suitable for regulatory use? Toxicol. Sci. 111 (2), 233–237. doi:10.1093/toxsci/kfp149
Hartung, T. (2009). Toxicology for the twenty-first century. Nature 460 (7252), 208–212. doi:10.1038/460208a
Hartung, T., van Vliet, E., Jaworska, J., Bonilla, L., Skinner, N., and Thomas, R. (2012). Systems toxicology. ALTEX 29 (2), 119–128. doi:10.14573/altex.2012.2.119
Hartwell, L. H., and Kastan, M. B. (1994). Cell cycle control and cancer. Science 266 (5192), 1821–1828. doi:10.1126/science.7997877
He, Y., Li, Z., Xu, T., Luo, D., Chi, Q., Zhang, Y., et al. (2022). Polystyrene nanoplastics deteriorate LPS-modulated duodenal permeability and inflammation in mice via ROS drived-NF-κB/NLRP3 pathway. Chemosphere 307, 135662. doi:10.1016/j.chemosphere.2022.135662
Heldaas, S. S., Langård, S. L., and Andersen, A. (1984). Incidence of cancer among vinyl chloride and polyvinyl chloride workers. Br. J. Ind. Med. 41 (1), 25–30. doi:10.1136/oem.41.1.25
Helma, C., Rautenberg, M., and Gebele, D. (2017). Nano-Lazar: Read across predictions for nanoparticle toxicities with calculated and measured properties. Front. Pharmacol. 8, 377. doi:10.3389/fphar.2017.00377
Hesterberg, T. W., McConnell, E. E., Miiller, W. C., Hamilton, R., and Bunn, W. B. (1992). Pulmonary toxicity of inhaled polypropylene fibers in rats. Fundam. Appl. Toxicol. 19 (3), 358–366. doi:10.1016/0272-0590(92)90174-G
Hiemstra, P. S., Grootaers, G., van der Does, A. M., Krul, C. A. M., and Kooter, I. M. (2018). Human lung epithelial cell cultures for analysis of inhaled toxicants: Lessons learned and future directions. Toxicol Vitro 47, 137–146. doi:10.1016/j.tiv.2017.11.005
Hilgers, A. R., Conradi, R. A., and Burton, P. S. (1990). Caco-2 cell monolayers as a model for drug transport across the intestinal mucosa. Pharm. Res. 7 (9), 902–910. doi:10.1023/A:1015937605100
Hinderliter, P. M., Minard, K. R., Orr, G., Chrisler, W. B., Thrall, B. D., Pounds, J. G., et al. (2010). Isdd: A computational model of particle sedimentation, diffusion and target cell dosimetry for in vitro toxicity studies. Part Fibre Toxicol. 7 (1), 36. doi:10.1186/1743-8977-7-36
Hongprasith, N., Kittimethawong, C., Lertluksanaporn, R., Eamchotchawalit, T., Kittipongvises, S., and Lohwacharin, J. (2020). IR microspectroscopic identification of microplastics in municipal wastewater treatment plants. Environ. Sci. Pollut. Res. Int. 27 (15), 18557–18564. doi:10.1007/s11356-020-08265-7
Hou, X., Chang, D. F., Trecartin, A., Barthel, E. R., Schlieve, C. R., Frey, M. R., et al. (2018). Short-term and long-term human or mouse organoid units generate tissue-engineered small intestine without added signalling molecules. Exp. Physiol. 103 (12), 1633–1644. doi:10.1113/EP086990
Hou, Z., Meng, R., Chen, G., Lai, T., Qing, R., Hao, S., et al. (2022). Distinct accumulation of nanoplastics in human intestinal organoids. Sci. Total Environ. 838, 155811. doi:10.1016/j.scitotenv.2022.155811
Hsiao, T.-J., Wang, J.-D., Yang, P.-M., Yang, P.-C., and Cheng, T.-J. (2004). Liver fibrosis in asymptomatic polyvinyl chloride workers. J. Occup. Environ. Med. 46 (9), 962–966. doi:10.1097/01.jom.0000137722.66767.38
Huang, W., Yin, H., Yang, Y., Jin, L., Lu, G., and Dang, Z. (2021). Influence of the co-exposure of microplastics and tetrabromobisphenol A on human gut: Simulation in vitro with human cell Caco-2 and gut microbiota. Sci. Total Environ. 778, 146264. doi:10.1016/j.scitotenv.2021.146264
Ichinose, T., Nishikawa, M., Takano, H., Sera, N., Sadakane, K., Mori, I., et al. (2005). Pulmonary toxicity induced by intratracheal instillation of Asian yellow dust (Kosa) in mice. Environ. Toxicol. Pharmacol. 20 (1), 48–56. doi:10.1016/j.etap.2004.10.009
Inaba, A., Arinaga, A., Tanaka, K., Endo, T., Hayatsu, N., Okazaki, Y., et al. (2021). Interleukin-4 promotes tuft cell differentiation and acetylcholine production in intestinal organoids of non-human primate. Int. J. Mol. Sci. 22 (15), 7921. doi:10.3390/ijms22157921
Inkielewicz-Stepniak, I., Tajber, L., Behan, G., Zhang, H., Radomski, M. W., Medina, C., et al. (2018). The role of mucin in the toxicological impact of polystyrene nanoparticles. Mater. (Basel) 11 (5), 724. doi:10.3390/ma11050724
Iwao, T., Toyota, M., Miyagawa, Y., Okita, H., Kiyokawa, N., Akutsu, H., et al. (2014). Differentiation of human induced pluripotent stem cells into functional enterocyte-like cells using a simple method. Drug Metab. Pharmacokinet. 29 (1), 44–51. doi:10.2133/dmpk.dmpk-13-rg-005
Jackson, G. R., Maione, A. G., Klausner, M., and Hayden, P. J. (2018). Prevalidation of an acute inhalation toxicity test using the EpiAirway in vitro human airway model. Appl Vitro Toxicol 4 (2), 149–158. doi:10.1089/aivt.2018.0004
Jagiello, K., and Ciura, K. (2022). In vitro to in vivo extrapolation to support the development of the next generation risk assessment (NGRA) strategy for nanomaterials. Nanoscale 14 (18), 6735–6742. doi:10.1039/d2nr00664b
Jani, P., Halbert, G. W., Langridge, J., and Florence, A. T. (1989). The uptake and translocation of latex nanospheres and microspheres after oral administration to rats. J. Pharm. Pharmacol. 41 (12), 809–812. doi:10.1111/j.2042-7158.1989.tb06377.x
Janssen, A. W. F., Duivenvoorde, L. P. M., Rijkers, D., Nijssen, R., Peijnenburg, A. A. C. M., van der Zande, M., et al. (2021). Cytochrome P450 expression, induction and activity in human induced pluripotent stem cell-derived intestinal organoids and comparison with primary human intestinal epithelial cells and Caco-2 cells. Arch. Toxicol. 95 (3), 907–922. doi:10.1007/s00204-020-02953-6
Järlskog, I., Jaramillo-Vogel, D., Rausch, J., Gustafsson, M., Strömvall, A.-M., and Andersson-Sköld, Y. (2022). Concentrations of tire wear microplastics and other traffic-derived non-exhaust particles in the road environment. Environ. Int. 170, 107618. doi:10.1016/j.envint.2022.107618
Jeon, S., Clavadetscher, J., Lee, D. K., Chankeshwara, S. V., Bradley, M., and Cho, W. S. (2018). Surface charge-dependent cellular uptake of polystyrene nanoparticles. Nanomater. (Basel) 8 (12), 1028. doi:10.3390/nano8121028
Jeon, S., Lee, D.-K., Jeong, J., Yang, S. I., Kim, J.-S., Kim, J., et al. (2021). The reactive oxygen species as pathogenic factors of fragmented microplastics to macrophages. Environ. Pollut. 281, 117006. doi:10.1016/j.envpol.2021.117006
Jones, C. F., and Grainger, D. W. (2009). In vitro assessments of nanomaterial toxicity. Adv. Drug Deliv. Rev. 61 (6), 438–456. doi:10.1016/j.addr.2009.03.005
Jung, B. K., Han, S. W., Park, S. H., Bae, J. S., Choi, J., and Ryu, K. Y. (2020). Neurotoxic potential of polystyrene nanoplastics in primary cells originating from mouse brain. Neurotoxicology 81, 189–196. doi:10.1016/j.neuro.2020.10.008
Kabeya, T., Mima, S., Imakura, Y., Miyashita, T., Ogura, I., Yamada, T., et al. (2020). Pharmacokinetic functions of human induced pluripotent stem cell-derived small intestinal epithelial cells. Drug Metab. Pharmacokinet. 35 (4), 374–382. doi:10.1016/j.dmpk.2020.04.334
Kabeya, T., Qiu, S., Hibino, M., Nagasaki, M., Kodama, N., Iwao, T., et al. (2018). Cyclic AMP signaling promotes the differentiation of human induced pluripotent stem cells into intestinal epithelial cells. Drug Metab. Dispos. 46 (10), 1411–1419. doi:10.1124/dmd.118.082123
Kämpfer, A. A. M., Busch, M., Buttner, V., Bredeck, G., Stahlmecke, B., Hellack, B., et al. (2021). Model complexity as determining factor for in vitro nanosafety studies: Effects of silver and titanium dioxide nanomaterials in intestinal models. Small 17 (15), e2004223. doi:10.1002/smll.202004223
Kämpfer, A. A. M., Shah, U.-K., Chu, S. L., Busch, M., Büttner, V., He, R., et al. (2022). “Interlaboratory comparison of an intestinal triple culture to confirm transferability and reproducibility,” in Vitromodels. doi:10.1007/s44164-022-00025-w
Kämpfer, A. A. M., Urban, P., Gioria, S., Kanase, N., Stone, V., and Kinsner-Ovaskainen, A. (2017). Development of an in vitro co-culture model to mimic the human intestine in healthy and diseased state. Toxicol Vitro 45 (1), 31–43. doi:10.1016/j.tiv.2017.08.011
Kararli, T. T. (1995). Comparison of the gastrointestinal anatomy, physiology, and biochemistry of humans and commonly used laboratory animals. Biopharm. Drug Dispos. 16 (5), 351–380. doi:10.1002/bdd.2510160502
Karbalaei, S., Hanachi, P., Walker, T. R., and Cole, M. (2018). Occurrence, sources, human health impacts and mitigation of microplastic pollution. Environ. Sci. Pollut. Res. Int. 25 (36), 36046–36063. doi:10.1007/s11356-018-3508-7
Kasendra, M., Tovaglieri, A., Sontheimer-Phelps, A., Jalili-Firoozinezhad, S., Bein, A., Chalkiadaki, A., et al. (2018). Development of a primary human Small Intestine-on-a-Chip using biopsy-derived organoids. Sci. Rep. 8 (1), 2871. doi:10.1038/s41598-018-21201-7
Kauffman, A., Gyurdieva, A., Mabus, J., Ferguson, C., Yan, Z., and Hornby, P. (2013). Alternative functional in vitro models of human intestinal epithelia. Front. Pharmacol. 4, 79. doi:10.3389/fphar.2013.00079
Kershaw, P. J., and Rochman, C. M. (2015). Sources, fate and effects of microplastics in the marine environment: Part 2 of a global assessment. Reports and studies. Group of Experts on the Scientific Aspects of Marine Environmental Protection (GESAMP) eng no. 93. Available at: IMO/FAO/Unesco-IOC/WMO/IAEA/UN/UNEP Joint.
Kogut, I., Roop, D. R., and Bilousova, G. (2014). “Differentiation of human induced pluripotent stem cells into a keratinocyte lineage,” in Epidermal cells: Methods and protocols. Editor K. Turksen (New York, NY: Springer New York), 1–12. doi:10.1007/7651_2013_64
Kosuth, M., Mason, S. A., and Wattenberg, E. V. (2018). Anthropogenic contamination of tap water, beer, and sea salt. PLoS One 13 (4), e0194970. doi:10.1371/journal.pone.0194970
Kulthong, K., Hooiveld, G. J. E. J., Duivenvoorde, L., Miro Estruch, I., Marin, V., van der Zande, M., et al. (2021). Transcriptome comparisons of in vitro intestinal epithelia grown under static and microfluidic gut-on-chip conditions with in vivo human epithelia. Sci. Rep. 11 (1), 3234. doi:10.1038/s41598-021-82853-6
Lacroix, G., Koch, W., Ritter, D., Gutleb, A. C., Larsen, S. T., Loret, T., et al. (2018). Air–liquid interface in vitro models for respiratory toxicology research: Consensus workshop and recommendations. Appl. Vitro Toxicol. 4 (2), 91–106. doi:10.1089/aivt.2017.0034
Lairson, L. L., Lyssiotis, C. A., Zhu, S., and Schultz, P. G. (2013). Small molecule–based approaches to adult stem cell therapies. Ann. Rev. Pharmacol. Toxicol. 53 (1), 107–125. doi:10.1146/annurev-pharmtox-011112-140300
Lamon, L., Asturiol, D., Vilchez, A., Cabellos, J., Damásio, J., Janer, G., et al. (2019). Physiologically based mathematical models of nanomaterials for regulatory toxicology: A review. Comput. Toxicol. 9, 133–142. doi:10.1016/j.comtox.2018.10.002
Lehner, R., Wohlleben, W., Septiadi, D., Landsiedel, R., Petri-Fink, A., and Rothen-Rutishauser, B. (2020). A novel 3D intestine barrier model to study the immune response upon exposure to microplastics. Arch. Toxicol. 94 (7), 2463–2479. doi:10.1007/s00204-020-02750-1
Leiding, J. W. (2017). Neutrophil evolution and their diseases in humans. Front. Immunol. 8, 1009. doi:10.3389/fimmu.2017.01009
Leslie Jhansi, L., Huang, S., Opp Judith, S., Nagy Melinda, S., Kobayashi, M., Young Vincent, B., et al. (2015). Persistence and toxin production by Clostridium difficile within human intestinal organoids result in disruption of epithelial paracellular barrier function. Infect. Immun. 83 (1), 138–145. doi:10.1128/IAI.02561-14
Lesuffleur, T., Barbat, A., Dussaulx, E., Zweibaum, A., and ZweibAum, A. (1990). Growth adaptation to methotrexate of HT-29 human colon carcinoma cells is associated with their ability to differentiate into columnar absorptive and mucus-secreting cells. Cancer Res. 50 (19), 6334–6343.
Liang, B., Zhong, Y., Huang, Y., Lin, X., Liu, J., Lin, L., et al. (2021). Underestimated health risks: Polystyrene micro- and nanoplastics jointly induce intestinal barrier dysfunction by ROS-mediated epithelial cell apoptosis. Part Fibre Toxicol. 18 (1), 20. doi:10.1186/s12989-021-00414-1
Liang, L., Cui, M., Zhang, M., Zheng, P., Deng, Z., Gao, S., et al. (2015). Nanoparticles' interference in the evaluation of in vitro toxicity of silver nanoparticles. RSC Adv. 5 (82), 67327–67334. doi:10.1039/c5ra05863e
Liebezeit, G., and Liebezeit, E. (2015). Origin of synthetic particles in honeys. Pol. J. Food Nutr. Sci. 65 (2), 143–147. doi:10.1515/pjfns-2015-0025
Lim, S. L., Ng, C. T., Zou, L., Lu, Y., Chen, J., Bay, B. H., et al. (2019). Targeted metabolomics reveals differential biological effects of nanoplastics and nanoZnO in human lung cells. Nanotoxicology 13 (8), 1117–1132. doi:10.1080/17435390.2019.1640913
Lin, S., Zhang, H., Wang, C., Su, X.-L., Song, Y., Wu, P., et al. (2022). Metabolomics reveal nanoplastic-induced mitochondrial damage in human liver and lung cells. Environ. Sci. Technol. 56 (17), 12483–12493. doi:10.1021/acs.est.2c03980
Lionetto, F., Corcione, C. E., Rizzo, A., and Maffezzoli, A. (2021). Production and characterization of polyethylene terephthalate nanoparticles. Polym. (Basel) 13 (21), 3745. doi:10.3390/polym13213745
Liu, S., Wu, X., Gu, W., Yu, J., and Wu, B. (2020). Influence of the digestive process on intestinal toxicity of polystyrene microplastics as determined by in vitro Caco-2 models. Chemosphere 256, 127204. doi:10.1016/j.chemosphere.2020.127204
Lo, B., and Parham, L. (2009). Ethical issues in stem cell research. Endocr. Rev. 30 (3), 204–213. doi:10.1210/er.2008-0031
Lozoya-Agullo, I., Araujo, F., Gonzalez-Alvarez, I., Merino-Sanjuan, M., Gonzalez-Alvarez, M., Bermejo, M., et al. (2017). Usefulness of caco-2/HT29-MTX and caco-2/HT29-MTX/raji B coculture models to predict intestinal and colonic permeability compared to caco-2 monoculture. Mol. Pharm. 14 (4), 1264–1270. doi:10.1021/acs.molpharmaceut.6b01165
Lu, W., Rettenmeier, E., Paszek, M., Yueh, M.-F., Tukey, R. H., Trottier, J., et al. (2017). Crypt organoid culture as an in vitro model in drug metabolism and cytotoxicity studies. Drug Metab. Dispos. 45 (7), 748–754. doi:10.1124/dmd.117.075945
Lyssiotis, C. A., Lairson, L. L., Boitano, A. E., Wurdak, H., Zhu, S., and Schultz, P. G. (2011). Chemical control of stem cell fate and developmental potential. Angew. Chem. Int. Ed. 50 (1), 200–242. doi:10.1002/anie.201004284
Ma, Y., Koh, J. Y. C., Lim, H. K., Shi, P., and Tay, C. Y. (2022). Elucidating the size-dependency of in vitro digested polystyrene microplastics on human intestinal cells health and function. Macromol. Chem. Phys. 223 (13), 2100454. doi:10.1002/macp.202100454
Ma, Y., McKay, D. J., and Buttitta, L. (2019). Changes in chromatin accessibility ensure robust cell cycle exit in terminally differentiated cells. PLoS Biol. 17 (9), e3000378. doi:10.1371/journal.pbio.3000378
Mabbott, N. A., Donaldson, D. S., Ohno, H., Williams, I. R., and Mahajan, A. (2013). Microfold (M) cells: Important immunosurveillance posts in the intestinal epithelium. Mucosal Immunol. 6 (4), 666–677. doi:10.1038/mi.2013.30
Madden, L. R., Nguyen, T. V., Garcia-Mojica, S., Shah, V., Le, A. V., Peier, A., et al. (2018). Bioprinted 3D primary human intestinal tissues model aspects of native physiology and ADME/tox functions. iScience 2, 156–167. doi:10.1016/j.isci.2018.03.015
Magrì, D., Sanchez-Moreno, P., Caputo, G., Gatto, F., Veronesi, M., Bardi, G., et al. (2018). Laser ablation as a versatile tool to mimic polyethylene terephthalate nanoplastic pollutants: Characterization and toxicology assessment. ACS Nano 12 (8), 7690–7700. doi:10.1021/acsnano.8b01331
Magrì, D., Veronesi, M., Sánchez-Moreno, P., Tolardo, V., Bandiera, T., Pompa, P. P., et al. (2021). PET nanoplastics interactions with water contaminants and their impact on human cells. Environ. Pollut. 271, 116262. doi:10.1016/j.envpol.2020.116262
Mahler, G. J., Esch, M. B., Tako, E., Southard, T. L., Archer, S. D., Glahn, R. P., et al. (2012). Oral exposure to polystyrene nanoparticles affects iron absorption. Nat. Nanotechnol. 7 (4), 264–271. doi:10.1038/nnano.2012.3
Majors, A. K., Boehm, C. A., Nitto, H., Midura, R. J., and Muschler, G. F. (1997). Characterization of human bone marrow stromal cells with respect to osteoblastic differentiation. J. Orthop. Res. 15 (4), 546–557. doi:10.1002/jor.1100150410
Mason, S. A., Welch, V. G., and Neratko, J. (2018). Synthetic polymer contamination in bottled water. Front. Chem. 6, 407. doi:10.3389/fchem.2018.00407
Mason, T. J. (1975). Cancer mortality in U.S. counties with plastics and related industries. Environ. Health Perspect. 11, 79–84. doi:10.1289/ehp.751179
Masui, A., Hirai, T., and Gotoh, S. (2022). Perspectives of future lung toxicology studies using human pluripotent stem cells. Arch. Toxicol. 96 (2), 389–402. doi:10.1007/s00204-021-03188-9
McKinney, C. E. (2017). Using induced pluripotent stem cells derived neurons to model brain diseases. Neural Regen. Res. 12 (7), 1062–1067. doi:10.4103/1673-5374.211180
Mehta, R. H. (2014). Sourcing human embryos for embryonic stem cell lines: Problems & perspectives. Indian J. Med. Res. 140, 106–111.
Meindl, C., Öhlinger, K., Zrim, V., Steinkogler, T., and Fröhlich, E. (2021). Screening for effects of inhaled nanoparticles in cell culture models for prolonged exposure. Nanomaterials 11 (3), 606. doi:10.3390/nano11030606
Miller, A. J., Dye, B. R., Ferrer-Torres, D., Hill, D. R., Overeem, A. W., Shea, L. D., et al. (2019). Generation of lung organoids from human pluripotent stem cells in vitro. Nat. Protoc. 14 (2), 518–540. doi:10.1038/s41596-018-0104-8
Mitrano, D. M., Lesher, E. K., Bednar, A., Monserud, J., Higgins, C. P., and Ranville, J. F. (2012). Detecting nanoparticulate silver using single-particle inductively coupled plasma–mass spectrometry. Environ. Toxicol. Chem. 31 (1), 115–121. doi:10.1002/etc.719
Miura, S., and Suzuki, A. (2017). Generation of mouse and human organoid-forming intestinal progenitor cells by direct lineage reprogramming. Cell Stem Cell 21 (4), 456–471. doi:10.1016/j.stem.2017.08.020
Mohamed Nor, N. H., Kooi, M., Diepens, N. J., and Koelmans, A. A. (2021). Lifetime accumulation of microplastic in children and adults. Environ. Sci. Technol. 55 (8), 5084–5096. doi:10.1021/acs.est.0c07384
Mohr, U., and Emura, M. (1985). The use of cell and organ culture from the respiratory tract for testing chemicals. Food Chem. Toxicol. 23 (2), 233–238. doi:10.1016/0278-6915(85)90022-5
Monopoli, M. P., Åberg, C., Salvati, A., and Dawson, K. A. (2012). Biomolecular coronas provide the biological identity of nanosized materials. Nat. Nanotechnol. 7 (12), 779–786. doi:10.1038/nnano.2012.207
Naumovska, E., Aalderink, G., Wong Valencia, C., Kosim, K., Nicolas, A., Brown, S., et al. (2020). Direct on-chip differentiation of intestinal tubules from induced pluripotent stem cells. Int. J. Mol. Sci. 21 (14), 4964. doi:10.3390/ijms21144964
Navabi, N., McGuckin, M. A., and Lindén, S. K. (2013). Gastrointestinal cell lines form polarized epithelia with an adherent mucus layer when cultured in semi-wet interfaces with mechanical stimulation. PLoS One 8 (7), e68761. doi:10.1371/journal.pone.0068761
Nikolaev, M., Mitrofanova, O., Broguiere, N., Geraldo, S., Dutta, D., Tabata, Y., et al. (2020). Homeostatic mini-intestines through scaffold-guided organoid morphogenesis. Nature 585 (7826), 574–578. doi:10.1038/s41586-020-2724-8
Noel, G., Baetz, N. W., Staab, J. F., Donowitz, M., Kovbasnjuk, O., Pasetti, M. F., et al. (2017). A primary human macrophage-enteroid co-culture model to investigate mucosal gut physiology and host-pathogen interactions. Sci. Rep. 7 (1), 45270. doi:10.1038/srep45270
Nuutila, J., and Lilius, E. M. (2005). Flow cytometric quantitative determination of ingestion by phagocytes needs the distinguishing of overlapping populations of binding and ingesting cells. Cytom. A 65 (2), 93–102. doi:10.1002/cyto.a.20139
Oberdorster, G., Oberdorster, E., and Oberdorster, J. (2005). Nanotoxicology: An emerging discipline evolving from studies of ultrafine particles. Environ. Health Perspect. 113 (7), 823–839. doi:10.1289/ehp.7339
Oh, S. J., Kim, H., Liu, Y., Han, H. K., Kwon, K., Chang, K. H., et al. (2014). Incompatibility of silver nanoparticles with lactate dehydrogenase leakage assay for cellular viability test is attributed to protein binding and reactive oxygen species generation. Toxicol. Lett. 225 (3), 422–432. doi:10.1016/j.toxlet.2014.01.015
Ohta, Y., Kazuki, K., Abe, S., Oshimura, M., Kobayashi, K., and Kazuki, Y. (2020). Development of Caco-2 cells expressing four CYPs via a mammalian artificial chromosome. BMC Biotechnol. 20 (1), 44. doi:10.1186/s12896-020-00637-8
Ong, K. J., MacCormack, T. J., Clark, R. J., Ede, J. D., Ortega, V. A., Felix, L. C., et al. (2014). Widespread nanoparticle-assay interference: Implications for nanotoxicity testing. PLoS One 9 (3), e90650. doi:10.1371/journal.pone.0090650
Park, S. B., Jung, W. H., Choi, K. J., Koh, B., and Kim, K. Y. (2022). A comparative systematic analysis of the influence of microplastics on colon cells, mouse and colon organoids. Tissue Eng. Regen. Med. 2022. doi:10.1007/s13770-022-00496-8
Peters, R., de Jong, N., de Haan, L., Wright, S., and Bouwmeester, H. (2022). Release and intestinal translocation of chemicals associated with microplastics in an in vitro human gastrointestinal digestion model. Microplast nanoplast 2 (1), 3. doi:10.1186/s43591-021-00022-y
Petersen, E. J., Barrios, A. C., Henry, T. B., Johnson, M. E., Koelmans, A. A., Montoro Bustos, A. R., et al. (2022). Potential artifacts and control experiments in toxicity tests of nanoplastic and microplastic particles. Environ. Sci. Technol. 56 (22), 15192–15206. doi:10.1021/acs.est.2c04929
Phanse, Y., Ramer-Tait, A. E., Friend, S. L., Carrillo-Conde, B., Lueth, P., Oster, C. J., et al. (2012). Analyzing cellular internalization of nanoparticles and bacteria by multi-spectral imaging flow cytometry. J. Vis. Exp. 64, e3884. doi:10.3791/3884
Pikuda, O., Xu, E. G., Berk, D., and Tufenkji, N. (2019). Toxicity assessments of micro- and nanoplastics can Be confounded by preservatives in commercial formulations. Environ. Sci. Technol. 6 (1), 21–25. doi:10.1021/acs.estlett.8b00614
Pinto, M., Robine-Leon, S. A., Appay, M. D., Kedinger, M., Triadou, N., Dussaulx, E., et al. (1983). Enterocyte-like differentiation and polarization of the human colon carcinoma cell line Caco-2 in culture. Biol. Cell 47, 323–330.
Pistollato, F., Madia, F., Corvi, R., Munn, S., Grignard, E., Paini, A., et al. (2021). Current EU regulatory requirements for the assessment of chemicals and cosmetic products: Challenges and opportunities for introducing new approach methodologies. Arch. Toxicol. 95 (6), 1867–1897. doi:10.1007/s00204-021-03034-y
Pleguezuelos-Manzano, C., Puschhof, J., van den Brink, S., Geurts, V., Beumer, J., and Clevers, H. (2020). Establishment and culture of human intestinal organoids derived from adult stem cells. Curr. Protoc. Immunol. 130 (1), e106. doi:10.1002/cpim.106
Porter, D. W., Castranova, V., Robinson, V. A., Hubbs, A. F., Mercer, R. R., Scabilloni, J., et al. (1999). Acute inflammatory reaction in rats after intratracheal instillation of material collected from a nylon flocking plant. J. Toxicol. Environ. Health A 57 (1), 25–45. doi:10.1080/009841099157845
Pulido-Reyes, G., Leganes, F., Fernández-Piñas, F., and Rosal, R. (2017). Bio-nano interface and environment: A critical review. Environ. Toxicol. Chem. 36 (12), 3181–3193. doi:10.1002/etc.3924
Roodsant, T., Navis, M., Aknouch, I., Renes, I. B., van Elburg, R. M., Pajkrt, D., et al. (2020). A human 2D primary organoid-derived epithelial monolayer model to study host-pathogen interaction in the small intestine. Front. Cell Infect. Microbiol. 10, 272. doi:10.3389/fcimb.2020.00272
Roshanzadeh, A., Park, S., Ganjbakhsh, S. E., Park, J., Lee, D.-H., Lee, S., et al. (2020). Surface charge-dependent cytotoxicity of plastic nanoparticles in alveolar cells under cyclic stretches. Nano Lett. 20 (10), 7168–7176. doi:10.1021/acs.nanolett.0c02463
Roursgaard, M., Hezareh Rothmann, M., Schulte, J., Karadimou, I., Marinelli, E., and Møller, P. (2022). Genotoxicity of particles from grinded plastic items in caco-2 and HepG2 cells. Front. Public Health 10, 906430. doi:10.3389/fpubh.2022.906430
Sargent, C. Y., Berguig, G. Y., Kinney, M. A., Hiatt, L. A., Carpenedo, R. L., Berson, R. E., et al. (2010). Hydrodynamic modulation of embryonic stem cell differentiation by rotary orbital suspension culture. Biotechnol. Bioeng. 105 (3), 611–626. doi:10.1002/bit.22578
Sato, T., Stange, D. E., Ferrante, M., Vries, R. G. J., van Es, J. H., van den Brink, S., et al. (2011). Long-term expansion of epithelial organoids from human colon, adenoma, adenocarcinoma, and barrett's epithelium. Gastroenterology 141 (5), 1762–1772. doi:10.1053/j.gastro.2011.07.050
Sato, T., Vries, R. G., Snippert, H. J., van de Wetering, M., Barker, N., Stange, D. E., et al. (2009). Single Lgr5 stem cells build crypt-villus structures in vitro without a mesenchymal niche. Nature 459 (7244), 262–265. doi:10.1038/nature07935
Schimpel, C., Teubl, B., Absenger, M., Meindl, C., Fröhlich, E., Leitinger, G., et al. (2014). Development of an advanced intestinal in vitro triple culture permeability model to study transport of nanoparticles. Mol. Pharm. 11 (3), 808–818. doi:10.1021/mp400507g
Schmitz, C., Welck, J., Tavernaro, I., Grinberg, M., Rahnenführer, J., Kiemer, A. K., et al. (2019). Mechanical strain mimicking breathing amplifies alterations in gene expression induced by SiO2 NPs in lung epithelial cells. Nanotoxicology 13 (9), 1227–1243. doi:10.1080/17435390.2019.1650971
Schwaferts, C., Niessner, R., Elsner, M., and Ivleva, N. P. (2019). Methods for the analysis of submicrometer- and nanoplastic particles in the environment. Trends Anal. Chem. 112, 52–65. doi:10.1016/j.trac.2018.12.014
Shi, Q., Tang, J., Wang, L., Liu, R., and Giesy, J. P. (2021). Combined cytotoxicity of polystyrene nanoplastics and phthalate esters on human lung epithelial A549 cells and its mechanism. Ecotoxicol. Environ. Saf. 213, 112041. doi:10.1016/j.ecoenv.2021.112041
Shi, X., Wang, X., Huang, R., Tang, C., Hu, C., Ning, P., et al. (2022). Cytotoxicity and genotoxicity of polystyrene micro- and nanoplastics with different size and surface modification in A549 cells. Int. J. Nanomedicine 17, 4509–4523. doi:10.2147/IJN.S381776
Shi, Y., Inoue, H., Wu, J. C., and Yamanaka, S. (2017). Induced pluripotent stem cell technology: A decade of progress. Nat. Rev. Drug Discov. 16 (2), 115–130. doi:10.1038/nrd.2016.245
Shim, K.-Y., Lee, D., Han, J., Nguyen, N.-T., Park, S., and Sung, J. H. (2017). Microfluidic gut-on-a-chip with three-dimensional villi structure. Biomed. Microdevices 19 (2), 37. doi:10.1007/s10544-017-0179-y
Sidar, B., Jenkins, B. R., Huang, S., Spence, J. R., Walk, S. T., and Wilking, J. N. (2019). Long-term flow through human intestinal organoids with the gut organoid flow chip (GOFlowChip). Lab. Chip 19 (20), 3552–3562. doi:10.1039/C9LC00653B
Simonsen, J. B., and Kromann, E. B. (2021). Pitfalls and opportunities in quantitative fluorescence-based nanomedicine studies - a commentary. J. Control Release 335, 660–667. doi:10.1016/j.jconrel.2021.05.041
Smirnov, A., Solga, M. D., Lannigan, J., and Criss, A. K. (2015). An improved method for differentiating cell-bound from internalized particles by imaging flow cytometry. J. Immunol. Methods 423, 60–69. doi:10.1016/j.jim.2015.04.028
Sontheimer-Phelps, A., Chou, D. B., Tovaglieri, A., Ferrante, T. C., Duckworth, T., Fadel, C., et al. (2020). Human colon-on-a-chip enables continuous in vitro analysis of colon mucus layer accumulation and physiology. Cell Mol. Gastroenterol. Hepatol. 9 (3), 507–526. doi:10.1016/j.jcmgh.2019.11.008
Sonugur, F. G., and Akbulut, H. (2019). The role of tumor microenvironment in genomic instability of malignant tumors. Front. Genet. 10, 1063. doi:10.3389/fgene.2019.01063
Spence, J. R., Mayhew, C. N., Rankin, S. A., Kuhar, M. F., Vallance, J. E., Tolle, K., et al. (2011). Directed differentiation of human pluripotent stem cells into intestinal tissue in vitro. Nature 470 (7332), 105–109. doi:10.1038/nature09691
Steckiewicz, K. P., Adamska, A., Narajczyk, M., Megiel, E., and Inkielewicz – Stepniak, I. (2022). Fluoride enhances polystyrene nanoparticles cytotoxicity in colonocytes in vitro model. Chem. Biol. Interact. 367, 110169. doi:10.1016/j.cbi.2022.110169
Stewart, C. J., Estes, M. K., and Ramani, S. (2020). “Establishing human intestinal enteroid/organoid lines from preterm infant and adult tissue,” in Innate lymphoid cells: Methods and protocols. Editor S. Amarnath (New York, NY: Springer US), 185–198. doi:10.1007/978-1-0716-0338-3_16
Stock, V., Bohmert, L., Donmez, M. H., Lampen, A., and Sieg, H. (2020). An inverse cell culture model for floating plastic particles. Anal. Biochem. 591, 113545. doi:10.1016/j.ab.2019.113545
Stock, V., Bohmert, L., Lisicki, E., Block, R., Cara-Carmona, J., Pack, L. K., et al. (2019). Uptake and effects of orally ingested polystyrene microplastic particles in vitro and in vivo. Arch. Toxicol. 93 (7), 1817–1833. doi:10.1007/s00204-019-02478-7
Stock, V., Laurisch, C., Franke, J., Donmez, M. H., Voss, L., Bohmert, L., et al. (2021). Uptake and cellular effects of PE, PP, PET and PVC microplastic particles. Toxicol Vitro 70, 105021. doi:10.1016/j.tiv.2020.105021
Stone, V., Johnston, H., and Schins, R. P. (2009). Development of in vitro systems for nanotoxicology: Methodological considerations. Crit. Rev. Toxicol. 39 (7), 613–626. doi:10.1080/10408440903120975
Sun, M., Ding, R., Ma, Y., Sun, Q., Ren, X., Sun, Z., et al. (2021). Cardiovascular toxicity assessment of polyethylene nanoplastics on developing zebrafish embryos. Chemosphere 282, 131124. doi:10.1016/j.chemosphere.2021.131124
Susewind, J., de Souza Carvalho-Wodarz, C., Repnik, U., Collnot, E. M., Schneider-Daum, N., Griffiths, G. W., et al. (2016). A 3D co-culture of three human cell lines to model the inflamed intestinal mucosa for safety testing of nanomaterials. Nanotoxicology 10 (1), 53–62. doi:10.3109/17435390.2015.1008065
Takahashi, K., Tanabe, K., Ohnuki, M., Narita, M., Ichisaka, T., Tomoda, K., et al. (2007). Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 131 (5), 861–872. doi:10.1016/j.cell.2007.11.019
Takahashi, Y., Inoue, Y., Kuze, K., Sato, S., Shimizu, M., Kiyono, H., et al. (2021). Comparison of gene expression and activation of transcription factors in organoid-derived monolayer intestinal epithelial cells and organoids. Biosci. Biotechnol. Biochem. 85 (10), 2137–2144. doi:10.1093/bbb/zbab136
Takahashi, Y., Sato, S., Kurashima, Y., Lai, C.-Y., Otsu, M., Hayashi, M., et al. (2017). Reciprocal inflammatory signaling between intestinal epithelial cells and adipocytes in the absence of immune cells. EBioMedicine 23, 34–45. doi:10.1016/j.ebiom.2017.07.027
Takahashi, Y., Sato, S., Kurashima, Y., Yamamoto, T., Kurokawa, S., Yuki, Y., et al. (2018). A refined culture system for human induced pluripotent stem cell-derived intestinal epithelial organoids. Stem Cell Rep. 10 (1), 314–328. doi:10.1016/j.stemcr.2017.11.004
Takeshita, A., Igarashi-Migitaka, J., Nishiyama, K., Takahashi, H., Takeuchi, Y., and Koibuchi, N. (2011). Acetyl tributyl citrate, the most widely used phthalate substitute plasticizer, induces cytochrome P450 3A through steroid and xenobiotic receptor. Toxicol. Sci. 123 (2), 460–470. doi:10.1093/toxsci/kfr178
Tang, X., Fan, X., Xu, T., He, Y., Chi, Q., Li, Z., et al. (2022). Polystyrene nanoplastics exacerbated lipopolysaccharide-induced necroptosis and inflammation via the ROS/MAPK pathway in mice spleen. Environ. Toxicol. 37 (10), 2552–2565. doi:10.1002/tox.23618
Tannenbaum, J., and Bennett, B. T. (2015). Russell and burch's 3Rs then and now: The need for clarity in definition and purpose. J. Am. Assoc. Lab. Anim. Sci. 54 (2), 120–132.
Teeguarden, J. G., Hinderliter, P. M., Orr, G., Thrall, B. D., and Pounds, J. G. (2007). Particokinetics in vitro: Dosimetry considerations for in vitro nanoparticle toxicity assessments. Toxicol. Sci. 95 (2), 300–312. doi:10.1093/toxsci/kfl165
Tenuta, T., Monopoli, M. P., Kim, J., Salvati, A., Dawson, K. A., Sandin, P., et al. (2011). Elution of labile fluorescent dye from nanoparticles during biological use. PLoS One 6 (10), e25556. doi:10.1371/journal.pone.0025556
Thant, A. A., Wu, Y., Lee, J., Mishra, D. K., Garcia, H., Koeffler, H. P., et al. (2008). Role of caspases in 5-FU and selenium-induced growth inhibition of colorectal cancer cells. Anticancer Res. 28 (6A), 3579–3592.
Thomas, D. G., Smith, J. N., Thrall, B. D., Baer, D. R., Jolley, H., Munusamy, P., et al. (2018). ISD3: A particokinetic model for predicting the combined effects of particle sedimentation, diffusion and dissolution on cellular dosimetry for in vitro systems. Part Fibre Toxicol. 15 (1), 6. doi:10.1186/s12989-018-0243-7
Thomson, J. A., Itskovitz-Eldor, J., Shapiro, S. S., Waknitz, M. A., Swiergiel, J. J., Marshall, V. S., et al. (1998). Embryonic stem cell lines derived from human blastocysts. Science 282 (5391), 1145–1147. doi:10.1126/science.282.5391.1145
Toropova, A. P., and Toropov, A. A. (2022). Nanomaterials: Quasi-SMILES as a flexible basis for regulation and environmental risk assessment. Sci. Total Environ. 823, 153747. doi:10.1016/j.scitotenv.2022.153747
Treveil, A., Sudhakar, P., Matthews, Z. J., Wrzesiński, T., Jones, E. J., Brooks, J., et al. (2020). Regulatory network analysis of Paneth cell and goblet cell enriched gut organoids using transcriptomics approaches. Mol. Omics 16 (1), 39–58. doi:10.1039/C9MO00130A
Trump, L. R., Nayak, R. C., Singh, A. K., Emberesh, S., Wellendorf, A. M., Lutzko, C. M., et al. (2019). Neutrophils derived from genetically modified human induced pluripotent stem cells circulate and phagocytose bacteria in vivo. Stem Cells Transl. Med. 8 (6), 557–567. doi:10.1002/sctm.18-0255
Tsai, Y.-H., Nattiv, R., Dedhia, P. H., Nagy, M. S., Chin, A. M., Thomson, M., et al. (2017). In vitro patterning of pluripotent stem cell-derived intestine recapitulates in vivo human development. Development 144 (6), 1045–1055. doi:10.1242/dev.138453
Upadhyay, S., and Palmberg, L. (2018). Air-liquid interface: Relevant in vitro models for investigating air pollutant-induced pulmonary toxicity. Toxicol. Sci. 164 (1), 21–30. doi:10.1093/toxsci/kfy053
Urbán, N., and Cheung, T. H. (2021). Stem cell quiescence: The challenging path to activation. Development 148 (3), dev165084. doi:10.1242/dev.165084
Van Cauwenberghe, L., and Janssen, C. R. (2014). Microplastics in bivalves cultured for human consumption. Environ. Pollut. 193, 65–70. doi:10.1016/j.envpol.2014.06.010
VanDussen, K. L., Marinshaw, J. M., Shaikh, N., Miyoshi, H., Moon, C., Tarr, P. I., et al. (2015). Development of an enhanced human gastrointestinal epithelial culture system to facilitate patient-based assays. Gut 64 (6), 911–920. doi:10.1136/gutjnl-2013-306651
Visalli, G., Facciolà, A., Pruiti Ciarello, M., De Marco, G., Maisano, M., and Di Pietro, A. (2021). Acute and sub-chronic effects of microplastics (3 and 10 µm) on the human intestinal cells HT-29. Int. J. Environ. Res. Public Health 18 (11), 5833. doi:10.3390/ijerph18115833
Walczak, A. P., Hendriksen, P. J. M., Woutersen, R. A., van der Zande, M., Undas, A. K., Helsdingen, R., et al. (2015a). Bioavailability and biodistribution of differently charged polystyrene nanoparticles upon oral exposure in rats. J. Nanoparticle Res. 17 (5), 231. doi:10.1007/s11051-015-3029-y
Walczak, A. P., Kramer, E., Hendriksen, P. J., Helsdingen, R., van der Zande, M., Rietjens, I. M., et al. (2015b). In vitro gastrointestinal digestion increases the translocation of polystyrene nanoparticles in an in vitro intestinal co-culture model. Nanotoxicology 9 (7), 886–894. doi:10.3109/17435390.2014.988664
Walczak, A. P., Kramer, E., Hendriksen, P. J., Tromp, P., Helsper, J. P., van der Zande, M., et al. (2015c). Translocation of differently sized and charged polystyrene nanoparticles in in vitro intestinal cell models of increasing complexity. Nanotoxicology 9 (4), 453–461. doi:10.3109/17435390.2014.944599
Walkey, C. D., Olsen, J. B., Song, F., Liu, R., Guo, H., Olsen, D. W. H., et al. (2014). Protein corona fingerprinting predicts the cellular interaction of gold and silver nanoparticles. ACS Nano 8 (3), 2439–2455. doi:10.1021/nn406018q
Wang, M., Lai, X., Shao, L., and Li, L. (2018a). Evaluation of immunoresponses and cytotoxicity from skin exposure to metallic nanoparticles. Int. J. Nanomedicine 13, 4445–4459. doi:10.2147/IJN.S170745
Wang, Y., Kim, R., Hwang, S.-H. J., Dutton, J., Sims, C. E., and Allbritton, N. L. (2018b). Analysis of interleukin 8 secretion by a stem-cell-derived human-intestinal-epithelial-monolayer platform. Anal. Chem. 90 (19), 11523–11530. doi:10.1021/acs.analchem.8b02835
Warheit, D. B., Webb, T. R., Reed, K. L., Hansen, J. F., and Kennedy, G. L. (2003). Four-week inhalation toxicity study in rats with nylon respirable fibers: Rapid lung clearance. Toxicology 192 (2), 189–210. doi:10.1016/S0300-483X(03)00304-4
Warren, H. S., Tompkins, R. G., Moldawer, L. L., Seok, J., Xu, W., Mindrinos, M. N., et al. (2015). Mice are not men. PNAS 112 (4), 345. doi:10.1073/pnas.1414857111
Watson, C. Y., DeLoid, G. M., Pal, A., and Demokritou, P. (2016). Buoyant nanoparticles: Implications for nano-biointeractions in cellular studies. Small 12 (23), 3172–3180. doi:10.1002/smll.201600314
Wert, G. d., and Mummery, C. (2003). Human embryonic stem cells: Research, ethics and policy. Hum. Reprod. 18 (4), 672–682. doi:10.1093/humrep/deg143
Wilhelmi, V., Fischer, U., van Berlo, D., Schulze-Osthoff, K., Schins, R. P. F., and Albrecht, C. (2012). Evaluation of apoptosis induced by nanoparticles and fine particles in RAW 264.7 macrophages: Facts and artefacts. Toxicol. Vitro 26 (2), 323–334. doi:10.1016/j.tiv.2011.12.006
Wilson, R. L., Hewes, S. A., Rajan, A., Lin, S.-C., Bomidi, C., Iida, T., et al. (2021). A millifluidic perfusion cassette for studying the pathogenesis of enteric infections using ex-vivo organoids. Ann. Biomed. Eng. 49 (4), 1233–1244. doi:10.1007/s10439-020-02705-8
Winkler, A. S., Cherubini, A., Rusconi, F., Santo, N., Madaschi, L., Pistoni, C., et al. (2022). Human airway organoids and microplastic fibers: A new exposure model for emerging contaminants. Environ. Int. 163, 107200. doi:10.1016/j.envint.2022.107200
Workman, M. J., Gleeson, J. P., Troisi, E. J., Estrada, H. Q., Kerns, S. J., Hinojosa, C. D., et al. (2018). Enhanced utilization of induced pluripotent stem cell–derived human intestinal organoids using microengineered chips. Cell Mol. Gastroenterol. Hepatol. 5 (4), 669–677. doi:10.1016/j.jcmgh.2017.12.008
Workman, M. J., Mahe, M. M., Trisno, S., Poling, H. M., Watson, C. L., Sundaram, N., et al. (2017). Engineered human pluripotent-stem-cell-derived intestinal tissues with a functional enteric nervous system. Nat. Med. 23 (1), 49–59. doi:10.1038/nm.4233
Wu, B., Wu, X., Liu, S., Wang, Z., and Chen, L. (2019). Size-dependent effects of polystyrene microplastics on cytotoxicity and efflux pump inhibition in human Caco-2cells. Chemosphere 221, 333–341. doi:10.1016/j.chemosphere.2019.01.056
Wu, S., Wu, M., Tian, D., Qiu, L., and Li, T. (2020). Effects of polystyrene microbeads on cytotoxicity and transcriptomic profiles in human Caco-2 cells. Environ. Toxicol. 35 (4), 495–506. doi:10.1002/tox.22885
Xie, S., Zhang, R., Li, Z., Liu, C., Chen, Y., and Yu, Q. (2022). Microplastics perturb colonic epithelial homeostasis associated with intestinal overproliferation, exacerbating the severity of colitis. Environ. Res. 217, 114861. doi:10.1016/j.envres.2022.114861
Xing, Y.-F., Xu, Y.-H., Shi, M.-H., and Lian, Y.-X. (2016). The impact of PM2.5 on the human respiratory system. J. Thorac. Dis. 8 (1), 69–74. doi:10.3978/j.issn.2072-1439.2016.01.19
Xu, M., Halimu, G., Zhang, Q., Song, Y., Fu, X., Li, Y., et al. (2019). Internalization and toxicity: A preliminary study of effects of nanoplastic particles on human lung epithelial cell. Sci. Total Environ. 694, 133794. doi:10.1016/j.scitotenv.2019.133794
Xu, X., Feng, Y., Han, C., Yao, Z., Liu, Y., Luo, C., et al. (2022). Autophagic response of intestinal epithelial cells exposed to polystyrene nanoplastics. Environ. Toxicol. 38, 205–215. doi:10.1002/tox.23678
Yamada, S., and Kanda, Y. (2021). “Evaluation of barrier functions in human iPSC-derived intestinal epithelium,” in Permeability barrier: Methods and protocols. Editor K. Turksen (New York, NY: Springer US), 27–35. doi:10.1007/7651_2021_346
Yamamoto, T., Arita, M., Kuroda, H., Suzuki, T., and Kawamata, S. (2022). Improving the differentiation potential of pluripotent stem cells by optimizing culture conditions. Sci. Rep. 12 (1), 14147. doi:10.1038/s41598-022-18400-8
Yang, S., Cheng, Y., Chen, Z., Liu, T., Yin, L., Pu, Y., et al. (2021). In vitro evaluation of nanoplastics using human lung epithelial cells, microarray analysis and co-culture model. Ecotoxicol. Environ. Saf. 226, 112837. doi:10.1016/j.ecoenv.2021.112837
Yao, Y., Glamoclija, M., Murphy, A., and Gao, Y. (2022). Characterization of microplastics in indoor and ambient air in northern New Jersey. Environ. Res. 207, 112142. doi:10.1016/j.envres.2021.112142
Yoshida, S., Miwa, H., Kawachi, T., Kume, S., and Takahashi, K. (2020). Generation of intestinal organoids derived from human pluripotent stem cells for drug testing. Sci. Rep. 10 (1), 5989. doi:10.1038/s41598-020-63151-z
Zakrzewski, W., Dobrzyński, M., Szymonowicz, M., and Rybak, Z. (2019). Stem cells: Past, present, and future. Stem Cell Res. Ther. 10 (1), 68. doi:10.1186/s13287-019-1165-5
Zhang, H., Zhang, S., Duan, Z., and Wang, L. (2022a). Pulmonary toxicology assessment of polyethylene terephthalate nanoplastic particles in vitro. Environ. Int. 162, 107177. doi:10.1016/j.envint.2022.107177
Zhang, J., Wang, L., and Kannan, K. (2020). Microplastics in house dust from 12 countries and associated human exposure. Environ. Int. 134, 105314. doi:10.1016/j.envint.2019.105314
Zhang, J., Wei, X., Zeng, R., Xu, F., and Li, X. (2017). Stem cell culture and differentiation in microfluidic devices toward organ-on-a-chip. Future Sci. OA 3 (2), FSO187. doi:10.4155/fsoa-2016-0091
Zhang, T., Yang, S., Ge, Y., Wan, X., Zhu, Y., Li, J., et al. (2022b). Polystyrene nanoplastics induce lung injury via activating oxidative stress: Molecular insights from bioinformatics analysis. Nanomaterials 12 (19), 3507. doi:10.3390/nano12193507
Zhang, Y. X., Wang, M., Yang, L., Pan, K., and Miao, A. J. (2022c). Bioaccumulation of differently-sized polystyrene nanoplastics by human lung and intestine cells. J. Hazard Mater 439, 129585. doi:10.1016/j.jhazmat.2022.129585
Zhu, K., Jia, H., Sun, Y., Dai, Y., Zhang, C., Guo, X., et al. (2020). Enhanced cytotoxicity of photoaged phenol-formaldehyde resins microplastics: Combined effects of environmentally persistent free radicals, reactive oxygen species, and conjugated carbonyls. Environ. Int. 145, 106137. doi:10.1016/j.envint.2020.106137
Keywords: iPSC, ASC, ESC, nanotoxicology, intestine, microplastics, particle research
Citation: Busch M, Brouwer H, Aalderink G, Bredeck G, Kämpfer AAM, Schins RPF and Bouwmeester H (2023) Investigating nanoplastics toxicity using advanced stem cell-based intestinal and lung in vitro models. Front.Toxicol 5:1112212. doi: 10.3389/ftox.2023.1112212
Received: 30 November 2022; Accepted: 17 January 2023;
Published: 27 January 2023.
Edited by:
Xiang Wang, University of California, Los Angeles, United StatesReviewed by:
Aditya Reddy Kolli, Philip Morris International, SwitzerlandElijah Petersen, National Institute of Standards and Technology (NIST), United States
Copyright © 2023 Busch, Brouwer, Aalderink, Bredeck, Kämpfer, Schins and Bouwmeester. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Hans Bouwmeester, aGFucy5ib3V3bWVlc3RlckB3dXIubmw=