 Madeline J. Stanley1,2*
Madeline J. Stanley1,2* Lisa Peters2,3
Lisa Peters2,3 Aidan Guttormson1,2
Aidan Guttormson1,2 Julien Tremblay4Jessica Wasserscheid4Lauren Timlick2
Julien Tremblay4Jessica Wasserscheid4Lauren Timlick2 Charles W. Greer4
Charles W. Greer4 José Luis Rodríguez Gil2,5Thor Halldorson3
José Luis Rodríguez Gil2,5Thor Halldorson3 Sonya Havens2Richard Grosshans1,6
Sonya Havens2Richard Grosshans1,6 Elliott Taylor7Gregg Tomy3
Elliott Taylor7Gregg Tomy3 David B. Levin1*
David B. Levin1* Vince P. Palace2,5*
Vince P. Palace2,5*- 1Department of Biosystems Engineering, University of Manitoba, Winnipeg, MB, Canada
- 2International Institute for Sustainable Development Experimental Lakes Area, Winnipeg, MB, Canada
- 3Center for Oil and Gas Research Laboratory, Department of Chemistry, University of Manitoba, Winnipeg, MB, Canada
- 4Genomics and Microbiomes Group, Energy, Mining and Environment Research Centre, National Research Council Canada, Montreal, QC, Canada
- 5Department of Environment and Geography, University of Manitoba, Winnipeg, MB, Canada
- 6International Institute for Sustainable Development, Winnipeg, MB, Canada
- 7Polaris Applied Sciences, Bainbridge Island, WA, United States
Conventional oil spill recovery practices can damage sensitive habitats, like freshwater shorelines, and leave residual oil in the environment, causing chronic exposure to interacting biota. Non-invasive remediation efforts are required to enhance recovery of residual oil after primary recovery without further environmental damage. Engineered Floating Wetlands (EFWs), also known as Floating Treatment Wetlands, have been used around the world for phytoremediation and biodegradation of aquatic contaminants. EFWs have the potential to increase crude oil biodegradation by enhancing surface area for microbial colonization and interaction with contaminants in the water column. To assess changes in the prokaryotic and eukaryotic microbial communities associated with EFWs in conjunction with primary recovery of shoreline washing, a contained oil spill of diluted bitumen was conducted in a shoreline enclosure in an experimental lake at the International Institute for Sustainable Development Experimental Lakes Area, northwestern Ontario, Canada, in 2019. Total polycyclic aromatic compound (PAC) concentrations in the water column of the experimental enclosure peaked 20 days post spill, declining to near background conditions by day 66, and were dominated by 2-, 3-, and 4-ring alkylated PACs. Although total sediment PACs were highly variable and were influenced by pyrogenic sources (e.g., retene from wood combustion/forest fires) in all sites, concentrations in the experimental enclosure indicated influence from diluted bitumen. The EFW prokaryotic community was diverse and evenly distributed, while the eukaryotic community had lower richness and evenness, with a few dominant organisms. To our knowledge, this was the first in-lake experiment studying EFWs for oil spill remediation under natural environmental conditions, responding to scientific and industrial research needs. While we were unable to confirm whether community shifts were a result of diluted bitumen or seasonal changes, EFWs supported natural microbial diversity, with presence of amplicon sequence variants capable of degrading PACs without the need for bacterial inoculation. Further research should assess EFW microbial changes with other oil products commonly transported in Canada. As well, assessing target PAC degradation rates, EFW surface area requirements, and microbial activity will continue to advance collective knowledge in this field on the potential of EFWs as a secondary remediation strategy.
1 Introduction
Canada has the third largest crude oil reserve in the world (Dupuis and Ucan-Marin, 2015; Natural Resources Canada, 2020) and is one of the top producers and exporters of oil (Natural Resources Canada, 2020). In the last 30 years, bitumen production in Canada has increased five-fold, from producing 135 thousand barrels per day (8% of crude oil production) in 1990 to 1.8 million barrels per day (∼40% of crude oil production) in 2019 (Canada Energy Regulator, 2021a)1. Bitumen is a highly viscous product, which must be diluted for some transport mechanisms (e.g., pipelines). Diluted bitumen, hereafter referred to as ‘dilbit’, is often a mixture of 70% bitumen and 30% diluent, such as condensate or naphtha (Lee et al., 2015; Canada Energy Regulator, 2021c). The export of dilbit increased from 17% to 63% of total Canadian crude oil exports from 1990 to 2019 (Canada Energy Regulator, 2021b)1. The increase in production and transport of bitumen or oil products puts nearby ecosystems at risk for accidental spill events, and thus spill responders and industry should have reliable and effective remediation methods in place for such incidents (Chang et al., 2014; Murray et al., 2018).
Conventional response methods, such as mechanical recovery, can be costly and invasive (Yavari et al., 2015). For example, mechanical recovery in shoreline environments can cause significant damage through excavation and/or removal of vegetation (Hoff, 1995; Pezeshki et al., 2000; Zhu et al., 2001; Lee et al., 2015) and chemical methods, such as dispersant use, can increase oil exposure to interacting organisms and dispersants have not been approved for use in freshwater in Canada (Lee et al., 2015). Biological remediation may be a less-invasive and cost-effective alternative with potential to increase degradation of spilled oil (Rehman et al., 2018). Biodegradation of oil, commonly referred to as monitored natural recovery, occurs when microorganisms metabolize petroleum carbon compounds for energy (Magar et al., 2009). This breakdown can be enhanced by nutrients (biostimulation) or bacterial inoculants (bioaugmentation), and can also be naturally stimulated by plants and their root exudates (Lee et al., 2015). In an oil spill scenario, enhanced metabolism of petroleum may be performed by increasing plant surface area through deploying engineered floating wetlands (EFWs). EFWs, also referred to as floating treatment wetlands (FTWs), are platforms of emergent vegetation with roots growing into the water column. This increases surface area interaction with contaminants for phytoremediation and microbial biofilm development for biodegradation (Tanner and Headley, 2011; Shahid et al., 2018). There has been growing interest and success in the use of EFWs to enhance microbial biodegradation of oil compounds, summarized in Stanley et al. (2022). This synthesis summarized findings from existing literature, concluding that the combination of plants and bacterial inoculants on EFWs had the greatest degradation potential, but plants alone were consistently more successful than bacteria alone. Authors further highlighted a need to study this remediation strategy in field-based trials and under variable environmental conditions, such as cold freshwater lakes. As well, a need to understand the potential of the native microbial community on EFW roots for in situ oil spill remediation, without bacterial inoculation (Stanley et al., 2022).
Research at the International Institute for Sustainable Development Experimental Lakes Area (IISD-ELA) in northwestern Ontario, Canada, explored the use of EFWs for secondary remediation of oil spills in freshwater shorelines of a boreal lake, as part of the FLOating Wetland Treatments to Enhance Remediation (FLOWTER) project. Canada’s boreal zone occupies 28% of land (552 million ha), of which 71 million ha is occupied by lakes, ponds, and rivers (Brandt et al., 2013). Conducting research at the IISD-ELA allows researchers to understand how oil spills may impact and be treated in similar environments. Studies conducted at the site also benefit from years of baseline data that is often missing to assess recovery following an oil spill incident (Lee et al., 2015).
The objective of this study was to monitor changes to the microbial community on EFW roots upon exposure to a controlled dilbit spill following primary recovery, and to monitor changes to polycyclic aromatic compound (PAC) chemistry in the water and sediment of the oiled and unoiled reference sites. We hypothesized that the community would shift in the presence of oil to microorganisms known to consume and degrade hydrocarbon compounds. To the best of our knowledge, this was the first freshwater, in-lake experiment to study controlled oil spills and EFWs.
2 Materials and methods
2.1 Research design and site description
The contained oil spill was conducted in a wetland shoreline enclosure on Lake 260, a boreal experimental lake at the IISD-ELA, in 2019 (volume = 1,975,969 m3, surface area = 332,460 m2, max depth = 15.64 m; IISD-ELA, 2022). Enclosures, previously described in Ankley et al. (2021), encompassed <20,000 L of shoreline aquatic habitat and were constructed with a floating collar (5 m wide x 10 m long) attached to an impermeable polypropylene curtain, sealed to the lake sediment and 5 m onto the shore with a double layer of sandbags (Curry Industries Ltd., Winnipeg, MB, Canada) (Supplementary Figure 1). The study included an experimental enclosure (oiled; EE), a reference enclosure (unoiled; RE), and a general surrounding lake reference site (unoiled; LR), noting there was no site replication. Due to limited appropriate wetland shoreline area and maximum depth (≤2 m), the EE was shortened to approximately 6 m long into the aqueous environment in efforts to maintain similar enclosure volumes.
2.1.1 Oil application and recovery
Cold Lake Winter Blend dilbit, sourced from pipeline stocks provided by the Canadian Association of Petroleum Producers (CAPP), was weathered on June 19–20, 2019 for 36h by exposing 9 kg of oil to sunlight and air movement in a 1.1 m diameter stainless steel pan over ∼220 L of lake water, as described in Palace et al. (2021). Weathering was performed to simulate oil that would typically reach the shoreline environment after a spill event. During the weathering process, oil undergoes evaporative loss of lighter compounds (e.g., BTEX), altering its chemical and physical characteristics, including an increase in viscosity and density that make recovery of an aquatic spill more difficult (Lee et al., 2015). After weathering, the oil was collected from the surface of the water in the weathering pan using slotted stainless steel spoons and 1,444 g was applied in the EE at the near shore (within 50 cm of the bank) environment on 21 June 2019. Shorelines were treated 4 days after oil addition (25 June 2019) to allow floating oil to strand naturally on the shoreline and to simulate conservative response times in remote locations. The EE received primary recovery of shoreline washing for 12 minutes through pumping enclosure water (1,600 L) over the shoreline with a low pressure manifold and collecting oil on the water surface with polypropylene sorbent media (Spill Ninja, MEP Brothers, Winnipeg, MB, Canada). Cleanup treatment followed response industry standards for wetland (vegetated) shorelines. Primary recovery removed 49.4 g of dilbit, leaving 1,394 g (97%) of residual oil on the shoreline and in the enclosure. The selected shoreline treatment method was non-invasive and did not include any attempts to scrub oil from surfaces or activities that would alter the shoreline and native vegetation. Residual oil was then treated in the EE on 26 June 2019, with secondary remediation of an EFW located in the middle to far shore environment of the enclosure. The RE also received shoreline washing on 27 June 2019, followed by EFW addition. In order to conduct such research, contingency measures were implemented to minimize further impact to the lake ecosystem beyond the experimental regions (Supplementary Material, Section 1.1).
2.1.2 Engineered floating wetlands
EFWs were BioHaven Floating Islands sourced and produced by Martin Ecosystems (LA, United States), under license by Floating Islands International (MT, United States). The platform was made of recycled polyethylene terephthalate plastic and coated in polyurea for protection, with holes to transplant emergent vegetation (Floating Islands International, 2024). EFWs (1.14 m2; 21 holes of 7.62 cm diameter) were originally established in 2017 with local emergent wetland plants, Gold Label coconut mulch, and soil, and re-transplanted in June 2019 to replace dead plant material with coconut mulch and Typha sp. (cattail) and three Carex spp. (sedge); C. atherodes/utriculata, C. lasiocarpa, and C. pseudocyperus/hystericina (identified by Chris Penner, personal communication with Dr. Richard Grosshans, IISD, n.d.). Noting, Carex are particularly difficult to identify due to the high number of species, classified by minor changes in plant features (Naczi, 1992, as cited in Starr et al., 2009). EFWs plants were measured for heights and number of live plants, however, due to insect activity, measurements are not discussed herein.
2.2 Polycyclic aromatic compound chemistry
2.2.1 Water chemistry
Water was collected 3 days pre-oil addition and on days 1 through 6, 8, 12, 20, 38, 66, 87, 209, 263, 356, and 411 post-oil addition for PAC chemistry (where day 0 was the date of oil addition). Total PACs (n = 44) included 16 United States Environmental Protection Agency (2014) priority parent PACs and 28 alkylated PACs (which includes parent and alkylated heterocyclic PACs) (Supplementary Table 1). Water was collected in 1 L amber bottles using a peristaltic pump from the far shore environment in the enclosures (∼0.5 m from enclosure end and collecting approximately halfway through the water column) and transported in a cooler to the Centre for Oil and Gas Research and Development (COGRAD) laboratory at the University of Manitoba (Winnipeg, MB, Canada). Within 24 h of collection, 250 mL of each sample was vacuum filtered (1.2 μm, Whatman GF/C filter) and liquid-liquid extraction was completed following methods previously described in Dearnley (2022). Briefly, filtrates were transferred into separatory funnels which received 20 μL of 5 ng/μL recovery internal standard (suite of d8-naphthalene, d8-acenaphthylene, d10-acenaphthene, d10-fluorene, d10-phenanthrene, d10-pyrene, d12-benz(a)anthracene, d12-chrysene, d12-benzo(b)fluoranthene, d12-benzo(k)fluoranthene, d12-benzo(a)pyrene, d12-indeno(1,2,3-c,d)pyrene, d14-dibenzo(a,h)anthracene, and d14-benzo(g,h,i)perylene) and 10 g of sodium chloride (NaCl). Samples were extracted twice with 50 mL dichloromethane (DCM) by gently swirling liquids for 1 min. DCM was collected into round bottom flasks (total 100 mL DCM) and was reduced to approximately 2 mL using rotary evaporation. Sodium sulphate (Na2SO4) was added to remove any residual water from the extraction process (Dearnley, 2022) and samples were transferred to 12 mL round bottom borosilicate glass tubes with a Teflon lined screw caps. The remaining sodium sulphate was rinsed three times with Optima ™ grade hexanes (ThermoFisher Scientific), which was then transferred into the glass test tubes. Solvent exchange from DCM to Optima ™ hexanes was conducted using a nitrogen gas evaporator (N-EVAP™111, Organomation Associates Inc., MA, USA and OA-SYS Heating System), until 1 mL of sample remained. Each sample was spiked with 20 μL of an instrument performance internal standard (5 ng/μL d10-anthracene) and was transferred into an amber GC vial with a Teflon cap. PACs were detected and quantified on an Agilent 7890 gas chromatograph coupled to a 7000C triple quadrupole mass spectrometer (GC-MS/MS) following methods and GC-MS/MS conditions outlined in Idowu et al. (2018).
During each sampling period, a field blank was collected by opening an amber bottle containing 200 mL Optima ™ water (ThermoFisher Scientific) for the duration of sample collection. PAC extraction of field blanks followed the same protocols described above for experimental samples. Final concentrations were calculated using internal standard recoveries and blank correction. Of all samples analyzed, GC-MS/MS runs for two samples (EE and RE) on day 209 failed and are not included.
Water was also regularly monitored for basic quality parameters and was sampled bi-weekly for nutrient chemistry, discussed in Supplementary Material, Sections 1.2 and 1.3.
2.2.2 Sediment chemistry
Sediment cores were collected with a sediment corer (4.6 cm diameter x 30 cm long clear polycarbonate tube) in three random locations (n = 3) from each site 3 days pre-oil addition and on days 38, 66, and 91 post-oil addition. Single sediment samples were collected on days 263 and 642. Sediment samples were collected in triplicate for PAC chemistry, unlike the single water samples, to capture spatial heterogeneity of PAC concentrations resulting from variation in oil submergence and/or sediment characteristics. The top 5 cm of each sediment core was transferred into a 125 mL amber jar and refrigerated overnight to remove overlying water once particles had settled and then frozen at −20°C. Sediment PACs were extracted at the COGRAD laboratory using accelerated solvent extraction methods described in Dearnley (2022) and quantified on GC-MS/MS as described above (Idowu et al., 2018).
2.3 Root microbial community
Three days before oil addition, single root samples were collected from a random location on each EFW. On days 38 and 91, root samples were collected from three random locations on each EFW (n = 3). Collected roots were transferred into a Whirl-pak® bag and placed onto dry ice until transported to a −80°C freezer before shipping to the National Research Council, Montreal, Quebec, Canada, for extraction and sequencing to assess microbial diversity.
2.3.1 Extraction, amplification and sequencing of gDNA
Root samples were extracted using the PowerSoil Max DNA extraction kit (cat#12988–10, QIAgen). Between 5 and 10 g of roots were weighed and transferred into a PowerMax Bead Tube containing 15 mL of PowerBead Solution. The extraction was done following manufacturer’s guidelines. The nucleotide acid elution was done with 3 mL of DNAse-free Ambion water. The extracted genomic DNA (gDNA) was quantified using the Quant-iT™ PicoGreen™ dsDNA Assay Kit (cat#. P7589, ThermoFisher Scientific).
Taxonomic 16S (prokaryotic) and 18S (eukaryotic) ribosomal ribonucleic acid (rRNA) gene amplifications were done on all the gDNA samples. Bacterial communities, including eubacteria and archaea, were targeted using the primer set 515F-Y (5′- GTGYCAGCMGCCGCGGTAA-3′)/926R (5′-CCGYCAATTYMTTTRAGTTT-3′). The 18S rRNA gene region of the eukaryotic ribosomal DNA was amplified using the primer set 565F (5′- CCAGCASCYGCGGTAATTCC-3′)/948R (5′- ACTTTCGTTCTTGATYRA-3′). Primers contained the required Illumina adaptors at the 5′end of the primer sequences (5′-TCGTCGGCAGCGTCAGATGTGTATAAGAGACAG-3′ for the forward primer and 5′-GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAG-3′ for the reverse primer).
The V4-V5 region of the bacterial 16S rRNA gene and the V4 region of the eukaryotic rRNA gene were amplified in 25 μL volumes using 12.5 μL of KAPA HiFi HotStart ReadyMixPCR Kit (Roche), 10 μM of each primer, Ambion nuclease-free water, 20 μg of BSA and 1 μL of gDNA. Thermal cycling conditions were as follows: initial denaturation at 95°C for 3 min; 30 cycles at 95°C for 30 s, 55°C (for 16S primers) or 50°C (for 18S primers) for 30 s, 72°C for 30 s; and a final elongation at 72 °C for 7 min. PCRs products were visualized on SyBrSafe-stained 1% agarose gels. PCR amplicons were purified using 0.8X volume of magnetic beads solution (Agencourt AMPure XP, Beckman Coulter Life Science) according to the manufacturer’s protocol. Unique codes were added to each sample by amplifying 5 μL of the purified PCR product with 12.5 μL of KAPA HiFi HotStart ReadyMixPCR Kit, 150 nM of each Nextera XT Index Primer (Illumina Inc., San Diego, CA, USA) and Ambion nuclease-free water for a total volume of 25 μL. Thermal cycling conditions were as follows: 3 min at 95°C, eight cycles of 30 s at 95°C, 30 s at 55°C, 30 s at 72°C, and a final elongation step of 5 min at 72°C. Indexed amplicons were purified with the magnetic beads as previously described and quantified using the picogreen fluorescence method and combined in an equimolar ratio.
Paired-end sequencing (2 × 250 bp) of the 16S and 18S rRNA gene amplicon pools was carried out on an Illumina MiSeq sequencer at the National Research Council (Montreal, QC, Canada) using the MiSeq Reagent kit V2, 500 cycles (cat# MS-102–2003, Illumina).
Sequencing data was analyzed using AmpliconTagger (Tremblay and Yergeau, 2019). Briefly, raw reads were scanned for sequencing adapters and PhiX spike-in sequences. Remaining paired-end reads were processed to remove primer sequences (pTrimmer v1.3.3; Zhang et al., 2019) and discard reads having an average quality Phred score lower than 20. The remaining sequences were processed for generating Amplicon Sequence Variants (ASVs) (DADA2 v1.12.1; Callahan et al., 2016). Since the quality filtering step was performed in a separate upstream step, we used more lenient parameters for the DADA2 workflow which is summarized as follows: filterAndTrim (maxEE = 8, truncQ = 0, maxN = 0, minQ = 0). Errors were learned using the learnErrors (nbases = 1e8) function for both forward and reverse filtered reads. Reads were then merged using the mergePairs (minOverlap = 10, maxMismatch = 0) function. Chimeras were removed with DADA2’s internal remove BimeraDeNovo (method = ‘consensus’) method followed by UCHIME reference (Rognes et al., 2016). ASVs were assigned a taxonomic lineage with the RDP classifier (Wang et al., 2007) using the complete SILVA release 138 database (Quast et al., 2012) limited to 16S and 18S entries. Taxonomic lineages were combined with the ASV abundance matrix obtained above to generate a raw ASV table, from which the prokaryotic or eukaryotic ASV tables were generated. A total of 6,365 and 3,263 ASVs were obtained for 16S and 18S data types respectively.
2.4 Data analyses
No statistical comparisons were made as there was no enclosure replication. Results can be used as a case example to assess whether EFW microbial communities change upon oil exposure, and to inform future replicated research to assess potential of plants and EFWs for secondary oil spill recovery. Trends were analyzed using R (4.2.2) in R Studio (2023.09.1 + 494; Posit Team, 2023; R Core Team, 2022) primarily with tidyverse (v. 2.0.0, Wickham et al., 2019). Trends in PAC concentration were prepared for dissolved-phase PACs and trends in mean PAC concentration with standard deviation (error bars) of site replicates were prepared for sediment PACs to visualize replicate variability within each site.
Rarefied ASVs, based on the lowest yield, were analyzed for richness (alpha and chao1) and diversity (Shannon and Simpson), as provided from the National Research Council of Canada, for Bray-Curtis beta diversity with a principal coordinate analysis (PCoA), and taxonomic profiles using R Studio (2023.09.1 + 494; Posit Team, 2023; R Core Team, 2022), visualized with ggplot2 (v. 3.4.1, Wickham, 2016). Additional R packages used for data analyses are included in Supplementary Table 2.
Metazoans and class Embryophyta (land plants) had high relative abundance but were removed to analyze the eukaryotic microbial community. Eukaryotic ASVs were inconsistently identified to genus, and often subphyla or clades were inserted into the classification. Edits (such as the removal of subphyla or classes confirmed with the SILVA database (R138.1) (SILVA, 2023; Quast et al., 2012; Yilmaz et al., 2014; Glöckner et al., 2017), World Register of Marine Species (WoRMS Editorial Board, 2023), and National Center for Biotechnology Information (Schoch et al., 2020) databases, and scientific literature) were made to correct or remove misplacement of taxonomic classifications (e.g., Phragmoplastophyta was misclassified as a phylum), however missing classifications once corrected (e.g., class/family) were not added. Taxonomic profiles present the lowest level identified.
3 Results and discussion
3.1 Polycyclic aromatic compound chemistry
3.1.1 Water chemistry
Total dissolved-phase PAC concentrations before oil addition were 290.21 ng/L, 27.38 ng/L, and 57.90 ng/L in the EE, RE, and LR, respectively (Figure 1). After oil addition to the EE, concentrations on day 1 were 1,591.23 ng/L, 95.84 ng/L, and 63.33 ng/L, respectively. The maximum concentration in the EE occurred on day 20, reaching 12,440.83 ng/L, within the chronic (>96 h) toxicity range for total polycyclic aromatic hydrocarbons (PAH) causing mortality (LC50) for freshwater biota (Lee et al., 2015). Total PACs on day 20 were 1,667.17 ng/L and 795.11 ng/L in the RE and LR, respectively. This peak was estimated to result from a precipitation event on day 19, a record rainfall of 145.6 mm in 24 h (IISD-ELA meteorological data), causing remobilization and reintroduction of oil from the shoreline and/or sediments. This was supported by the declining trend in total dissolved-phased PAC concentrations on days 8 and 12 after oil addition. Concentrations in the EE then declined to 2,336.03 ng/L on day 38, and later to 359.91 ng/L, nearing background conditions on day 66. Total PACs were relatively stable for the remainder of the experiment, where concentrations were last recorded on day 411 at 467.97 ng/L, 202.56 ng/L, and 119.51 ng/L in the in EE, RE, and LR, respectively. Figure 1 presents the first 87 days of exposure. The remaining data are presented in Supplementary Figure 2.
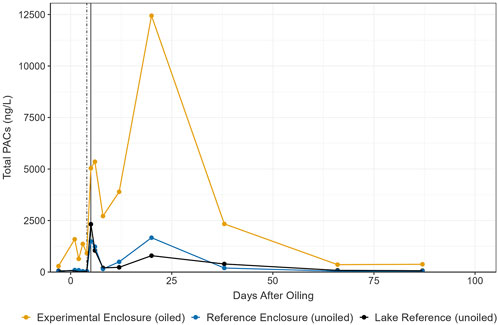
Figure 1. Total PACs (n = 44) in the water from 3 days pre-oil addition to day 87 post-oil addition. Vertical lines represent primary (dotted) and secondary (solid) remediation events.
Total PACs in the EE were primarily 2-, 3- and 4-ring alkylated PACs, with fewer ≥5-ring compounds (Supplementary Figures 3, 4), and with dominance (mean >100 ng/L) of alkylated naphthalenes (C2, C3, and C4), dibenzothiophenes (C1 and C2), C2-pyrene, and C1-fluorene. Following oil addition, alkylated PACs had a mean (range; n = 16) of 90.65% (66.79%–99.28%), 70.56% (16.89%–98.49%), and 60.06% (4.28%–99.87%) of total PACs in the EE, RE and LR, respectively. Alkylated PACs are more persistent compared to parent compounds, have greater chronic toxicity to biota (Zhu et al., 2001; Lee et al., 2015), and increase with continued weathering and evaporative processes (Lee et al., 2015). Total PACs were higher than previous oil spill research at the IISD-ELA (Palace et al., 2021; Rodriguez-Gil et al., 2021), which may be a result of the oil product, PACs measured, analytical methods (Rodriguez-Gil et al., 2021), remediation efforts, site volume, and environmental conditions. For example, during this study we experienced an extreme precipitation event which may have caused redistribution and mixing of oil into the water column from the oil impacted shoreline, whereas in previous studies oil had sunk to the lake bottom (Stoyanovich et al., 2021) with potential accumulation in sediment through deposition, partitioning, and/or by direct contact (Stoyanovich et al., 2022). This illustrates the limitations of comparing to prior studies and past spill events.
3.1.2 Sediment chemistry
Significant variability was evident among total sediment PAC concentrations within replicates and there were high concentrations observed in unoiled sites, making it difficult to discern differences between sites over the exposure period. This was in part due to significant contributions from retene, a three-ring PAC, associated with combustion of wood (i.e., forest fires) or diagenic processes (Ramdahl, 1983; Gabos et al., 2001). Forest fires are a regular and essential disturbance for boreal forest rejuvenation (Brandt et al., 2013), and the study region is often impacted by nearby fires resulting in ash deposition and smoke, and also has signs of a historic fire in the watershed (1979; personal communication with Paul Fafard, IISD-ELA, February 2023). In all sampling periods, percent retene of total PACs ranged from 0.00% to 92.29%, 22.03%to96.80%, and 28.04%to97.68% in the EE, RE, and LR, respectively. The differences in ranges between sites suggest the EE may have other compounds contributing to total PACs in some replicates and/or sample rounds, potentially those of petrogenic origin. To assess changes in sediment PACs between sites, we reanalyzed PAC data excluding retene from total PAC concentrations (n = 43) (Figure 2) (trends of the complete dataset, including retene is included in Supplementary Figure 5).
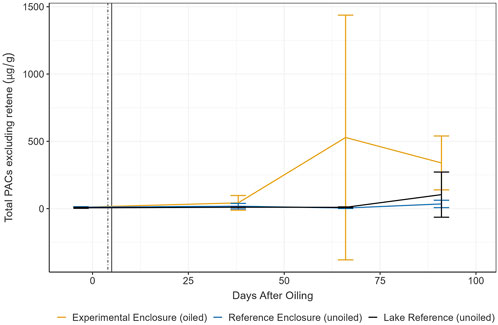
Figure 2. Mean (±SD) total sediment PACs (excluding retene; n = 43) from 3 days pre-oil addition to day 91 post-oil addition. Error bars represent standard deviation of triplicate samples at each site. Vertical lines represent primary (dotted) and secondary (solid) remediation events. A sediment sample from the EE on day 91 was lost.
Mean total PACs (±SD; triplicate samples at each site) excluding retene (n = 43) during pre-oil conditions were 10.46 ± 1.26 μg/g, 10.86 ± 3.94 μg/g, and 5.78 ± 3.76 μg/g in the EE, RE, and LR, respectively, with large contributions from C1-pyrene (14.50%–44.05%). Concentrations increased with high variability on day 38 in the EE to 42.64 ± 54.65 μg/g, with large contributions from C3-dibenzothiophene (4.54%–20.59%), C2-dibenzothiophene (5.31%–19.13%), and C1-pyrene (5.68%–15.50%), however the largest change occurred on day 66 at 528.39 ± 909.17 μg/g, mostly of C4-phenanthrene (1.72%–87.47%) and C1-pyrene (6.54%–23.58%) in some samples. The variability among replicates indicates potential influence of sample location, similarly observed in Stoyanovich et al. (2022). Any oil globules contained in the EE sediment samples would have contributed to increased PACs and replicate variability. Sediment PACs in the RE were 18.17 ± 20.60 μg/g and 4.23 ± 3.44 μg/g on day 38 and 66, respectively, and were 10.36 ± 1.50 μg/g and 9.76 ± 3.37 μg/g in the LR, respectively, mostly C1-pyrene. Concentrations decreased in the EE on day 91, at 339.42 ± 200.17 μg/g (n = 2), almost fully contributed by C1-pyrene (98.89%–99.00%), with little contribution from alkylated dibenzothiophenes or phenanthrene, suggesting potential degradation, loss, or influence of sample location. Concentrations in the RE and LR were 34.77 ± 27.62 μg/g and 103.93 ± 167.73 μg/g, respectively. By day 263, concentrations were similar amongst all sites at 30.31 μg/g, 31.30 μg/g, and 36.61 μg/g, respectively (88.65%–93.81% C1-pyrene) (Supplementary Figure 6). On day 642, sediment PACs remained at 31.80 μg/g in the EE, while the RE and LR had 19.16 μg/g and 1.25 μg/g, respectively. It is clear there is a large contribution of C1-pyrene in the sediment, with and without oil exposure, and it is possible C1-pyrene and other sediment PACs, may have resulted from forest fires (Vergnoux et al., 2011), however the increase on day 38 and 66 in the EE, is likely contributed from dilbit.
Despite background PACs skewing interpretation of total PAC trends between oiled and unoiled sites, increases in mean trends of parent and alkylated PACs in the EE indicate potential influence from dilbit (Supplementary Figure 5). However, there was large variability captured within site replicates, which is likely a result of spatial heterogeneity due to sediment characteristics and/or oil submergence and PAC sorption. To assess whether PACs arose from petrogenic or pyrogenic sources, sediment PAC ratios are discussed in Supplementary Material, Section 1.4.
3.2 Root microbial community
3.2.1 Diversity
Root microbial 16S (prokaryotes) and 18S (eukaryotes) rRNA gene Bray-Curtis beta diversity of rarefied ASVs was analyzed using a PCoA to identify differences between EFWs over the experimental period. The PCoA for prokaryotic and eukaryotic beta diversity had 40.13% and 41.28% of explained variation on the first two axes, respectively (Figure 3). The communities of all three EFWs on day −3 were clustered together on both PCoAs, indicating similarity before EFW deployment to their respective sites. The greatest differences in the prokaryotic community were between the LR and enclosures on days 38 and 91, which were clustered together, suggesting there may be an enclosure effect. Differences over the exposure period were observed for the enclosure sites, with greater differences between sites on day 91, indicating a potential seasonal or condition effect. There were no changes in the eukaryotic beta diversity in the LR, while there were changes in the enclosures over the exposure period, with greatest differences between the enclosure sites on day 91, displaying a potential site condition and/or seasonal effect.
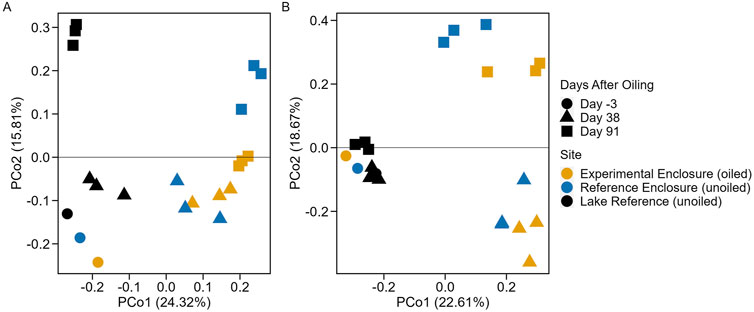
Figure 3. Principal Coordinate Analysis of 16S (A) and 18S (B) Bray-Curtis beta diversity in EFW root microbial biofilm.
Prokaryotes had greater richness, and diversity and evenness than eukaryotes, suggesting there may be no dominating prokaryotic organisms, while certain eukaryotic organisms may be dominant or selected for. While highly variable, the enclosures appear to have greater mean richness than the LR on days 38 and 91, but diversity indices are variable, further suggesting a potential enclosure effect (Supplementary Table 3). The enclosure design may limit wind and wave aeration, nutrient cycling, and other lake processes (see Supplementary Material, Section 1.2) that may influence microbial diversity and abundance. Similar enclosure effects on the aquatic prokaryotic community were identified in the Freshwater Oil spill Remediation Study which took place at the IISD-ELA studying the use of enhanced Monitored Natural Recovery and shoreline washing agent as secondary remediation methods following dilbit spills (Kharey et al., 2024).
3.2.2 Taxonomic profiles
3.2.2.1 Prokaryotes
Taxonomic profiles of rarefied ASVs revealed that among all sites and periods, prokaryotes were dominated by phyla Proteobacteria (41.48%–56.68% relative abundance [RA]), Bacteroidota (10.30%–20.94%), Planctomycetota (5.40%–10.93%), Acidobacteriota (3.74%–8.85%), and others, of a total of 45 bacteria and archaea phyla identified (Supplementary Figure 7). The Proteobacteria were mostly made of classes Alphaproteobacteria (20.34%–29.47% of class taxonomic profiles) and Gammaproteobacteria (18.89%–31.42% of class taxonomic profiles), which are two major groups involved in PAC degradation (Ghosal et al., 2016).
Taxonomic profiles of the top 20 prokaryotic ASVs (genera) represent approximately 40% of community RA, of 704 identified ASVs (Figure 4), indicating high community richness. For that reason, the next 21–40 most relatively abundant organisms are presented (Figure 5). The highest relative abundance was 7.71% for Bryobacter, an aerobic chemoorganotroph found in acidic wetland environments (e.g., sphagnum) (Kulichevskaya et al., 2010; Dedysh, 2019), a common moss in the study lake. Due to high richness and diversity, this section will discuss those with potential roles in oil spill remediation. However acknowledging that other microorganisms may support hydrocarbon degraders, through nitrogen or oxygen production (Radice et al., 2023) for example, and a diverse consortia is important for co-metabolism of complex, or a mixture of, PACs (Zhu et al., 2001; Cao et al., 2009; Lee et al., 2015; Ghosal et al., 2016).
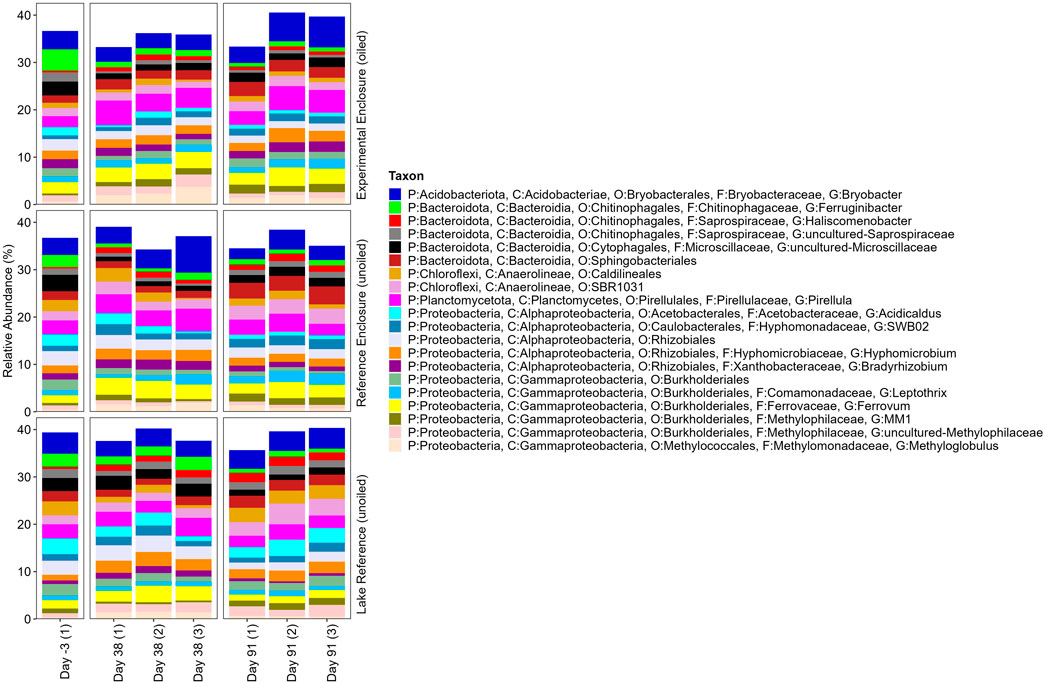
Figure 4. Taxonomic profile of the top twenty most relatively abundant 16S ASVs in EFW root biofilm replicates from the EE, RE, and LR 3 days pre-oil addition, and on days 38 and 91 post-oil addition. Taxon legend includes full taxonomic details to the lowest level identified as available.
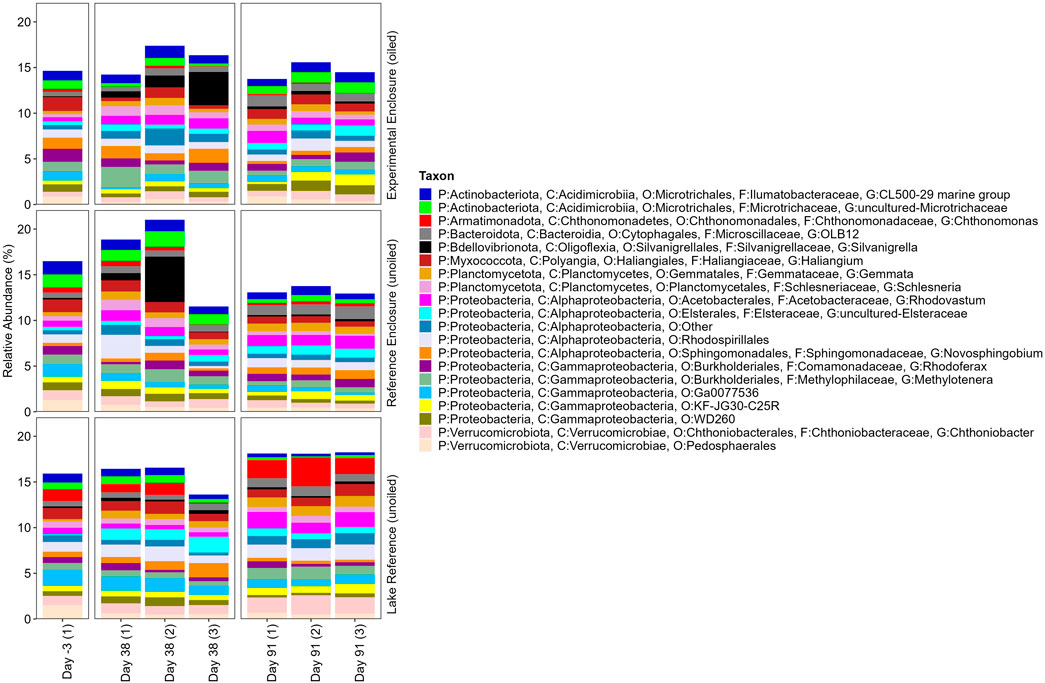
Figure 5. Taxonomic profile of the top twenty-one to forty most relatively abundant 16S ASVs in EFW root biofilm replicates from the EE, RE, and LR 3 days pre-oil addition, and on days 38 and 91 post-oil addition. Taxon legend includes full taxonomic details to the lowest level identified as available.
Three genera belong to order Rhizobiales, microorganisms in symbiosis with plants, known for their ability to assist in nitrogen fixation (Newton et al., 2011; Garrido-Oter et al., 2018; Fahde et al., 2023) and plant growth promotion, and can enhance phosphate solubility and accessibility (Fahde et al., 2023). Some have been reported to express various monooxygenase genes for degrading hydrocarbons and the desulfurization of hydrocarbons (e.g., dibenzothiophene) (Abbasian et al., 2016), and various species are known hydrocarbon degraders (Prince et al., 2018). Bradyrhizobium, previously found in co-occurrence with hydrocarbon degraders and involved in degradation (Yang et al., 2016), had variable abundance over the exposure period and between sites, peaking in the EE on day 91 (1.55%–2.20%), and on day 38 in the RE (1.84%–2.11%) and LR (1.24%–1.46%). Similarly, Hyphomicrobium, found in a consortia capable of degrading aromatic hydrocarbons (H. facile, strain Y3; Ozaki et al., 2006), peaked on day 91 in the EE (1.70%–2.99%) and on day 38 in the RE (1.96%–2.36%) and LR (2.38%–2.99%). Bradyrhizobium and Hyphomicrobium belong to families Xanthobacteraceae and Hyphomicrobiaceae, known to include hydrocarbon degraders and were detected in salt marsh sediments following the Deepwater Horizon oil spill (Beazley et al., 2012).
Pirellula are aerobic, heterotrophic bacteria, known for their role in biogeochemical processes (Glöckner et al., 2003). Some strains have P450 monooxygenases and epoxide hydrolase for detoxifying pollutants (Glöckner et al., 2003), and while not specific to oil, the expression of these enzymes may have important roles in aromatic ring cleavage. Pirellula increased in all sites by day 38 (2.47%–5.23%), highest in the EE.
Two ASVs belong to Chloroflexi, anaerobic bacteria that can degrade petroleum hydrocarbons (Cébron et al., 2022), that have been found in high abundance under oil contamination (Aburto-Medina et al., 2012; Brown et al., 2013; Abed et al., 2014; Peng et al., 2015; Cébron et al., 2022) and salt marsh sediments following the Deepwater Horizon spill, including class Anaerolineae (Engel et al., 2017). ASVs of the order Caldilineales had variable abundance among all sites, with the highest detection on day 38 in the EE (0.43%–1.34%) and RE (0.29%–2.88%), and on day 91 in the LR (2.75%–3.03%). Presence of order SBR1031 increased over the exposure period in all three sites, ranging from 1.64% to 2.16%, 2.91% to 3.18%, and 2.85% to 4.35% in the EE, RE, and LR, respectively.
Seven genera from order Burkholderiales were in the top forty most abundant prokaryotes. Burkholderiales are commonly found in the environment, and some have significant potential for PAC biodegradation (Pérez-Pantoja et al., 2012; Prince et al., 2018). For example, enrichment of Methylotenera, a methylotrophic bacterium, has been detected in oil and petroleum contaminated water and is suggested to be a hydrocarbon degrader (Thompson et al., 2017; Idomeh et al., 2021). Methylotenera’s abundance was highest in the EE by day 38 (1.06%–2.23%). Other methylotrophic Burkholderiales included MM1 and an uncultured genus from family Methylophilaceae. Ferrovum increased over the exposure period in the EE to a range of 2.56%–3.95% by day 91, while RAs were variable in the reference sites, highest by day 38. Leptothrix, a noted phenanthrene degrader (Bodour et al., 2003) and Rhodoferax, belong to family Comamonadaceae, which can express aromatic oxygenase-encoding genes (Pérez-Pantoja et al., 2012), and include hydrocarbon degrading species (Beazley et al., 2012). Leptothrix slightly increased in the enclosures over the exposure period, ranging from 1.13% to 2.01% and 1.49% to 2.32% by day 91 in the EE and RE, respectively. RA was lower in the LR, highest on day 91 (0.84%–1.11%). Rhodoferax was variable over time and between sites, highest on day −3 in the EE at 1.42% RA, however days 38 and 91 appeared relatively similar.
Xiao et al. (2013) studied microbial communities in production wells in the Chaoyanggou oil reservoir and found dominance of family Comamonadaceae in oil production wells, and family Hyphomonadaceae, among others, in formation water from production wells. Hyphomonadaceae has previously been correlated with the biodegradation of chemically dispersed Troll and Grane oils at cool temperatures (Ribicic et al., 2018). We observed SWB02 (family Hyphomonadaceae) increase in all sites from day −3 to 38, ranging from 0.80% to 1.58%, 1.25% to 2.28%, and 1.12% to 2.12% in the EE, RE, and LR, respectively.
Another methanotroph, Methyloglobulus known to degrade methane (Deutzmann et al., 2014), a potential by-product of oil biodegradation (Lee et al., 2015) and/or a potentially major hydrocarbon during spills (King et al., 2015), peaked in all sites by day 38. RA increased from 0.66% on day −3 to 1.99%–3.73% by day 38 in the EE, declining to 1.40%–1.97% by day 91. While RA ranged from 1.22% to 1.92% and 1.38% to 1.55% in the RE and LR by day 38, respectively.
Another well-known PAC degrader is Novosphingobium, a metabolically versatile sphingomonad often found in the rhizosphere (Sohn et al., 2004; Kertesz and Kawasaki, 2010; Lyu et al., 2014; Ghosal et al., 2016; Yang et al., 2016; Chettri and Singh, 2019; Rojo, 2021), capable of degrading a suite of PACs (Sohn et al., 2004). Novosphingobium was detected in all samples, peaking on day 38, ranging from 0.78% to 1.53%, 0.26% to 0.85%, and 0.66% to 1.56% in the EE, RE, and LR, respectively.
An ASV from order Sphingobacteriales increased over time in the enclosures, ranging from 2.27% to 3.00% and 3.15% to 3.80% in the EE and RE by day 91, respectively, while being more variable in the LR. Sphingobacteriales are known to include hydrocarbon degrading species (Beazley et al., 2012; Prince et al., 2018), identified in high abundance under oil contamination (Y. Wang et al., 2021), and some have been identified as late responders to oil contamination (Koo et al., 2015; Franchi et al., 2022). Family Flexibacteraceae from order Sphingobacteriales increased in a salt marsh following the Deepwater Horizon spill, as well as families from order Rhodospirillales (Beazley et al., 2012), which include genera capable of degrading alkanes, aromatics, and PACs (Prince et al., 2018, and citations within). An ASV from order Rhodospirillales peaked on day 91 in the EE (0.68%–1.34%) and LR (1.36%–1.66%), and on day 38 in the RE (0.26%–2.58%).
Other potential hydrocarbon degraders from phylum Bacteroidota include Haliscomenobacter (Aburto-Medina et al., 2012), and an uncultured Saprospiraceae (Taylor et al., 2021), which was a dominant organism after the wells were shut during the Deepwater Horizon spill (Dubinsky et al., 2013), possibly consuming complex carbon sources (McIlroy and Nielsen, 2014). Haliscomenobacter increased over time in the reference sites, reaching 1.10%–1.56% and 1.67%–1.99% in the RE and LR by day 91, respectively, and the EE peaked on day 38 (0.85%–1.20%). However, the uncultured Saprospiraceae declined in the EE (<1.00% on days 38 and 91), was variable in the RE (0.46%–1.30%), and remained stable in the LR over time (1.00%–1.89%).
3.2.2.2 Eukaryotes
Eukaryotic phyla were more variable in dominance between sites and over the experimental period than the prokaryotic community, with dominance by Ciliophora (16.49%–63.87%), Diatomea (3.75%–36.57%), Charophyta (0.00%–29.38%), Cercozoa (1.96%–18.76%), Dinoflagellata (0.91%–9.87%), Chytridiomycota (1.85%–9.11%), unidentified phyla (2.48%–12.25%), and others, out of 35 phyla identified (Supplementary Figure 8). Ciliophora were dominant on day −3 and on day 91, but slightly decreased in mean RA on day 38 in all sites. While on day 38, other phyla increased, such as Diatomea in the LR, and Charophyta, Dinoflagellata, Cercozoa, and others in the enclosures.
The top twenty eukaryotic ASVs represented approximately 40%–75% of the community and displayed an uneven distribution with some dominant ASVs (Figure 6). The RA explained by the top twenty declined in the enclosure sites on day 38, suggesting others may be selected for upon application to the enclosures, as observed in the top 21–40 eukaryotic ASVs (Figure 7). The following section will focus on organisms with potential roles in oil spill remediation.
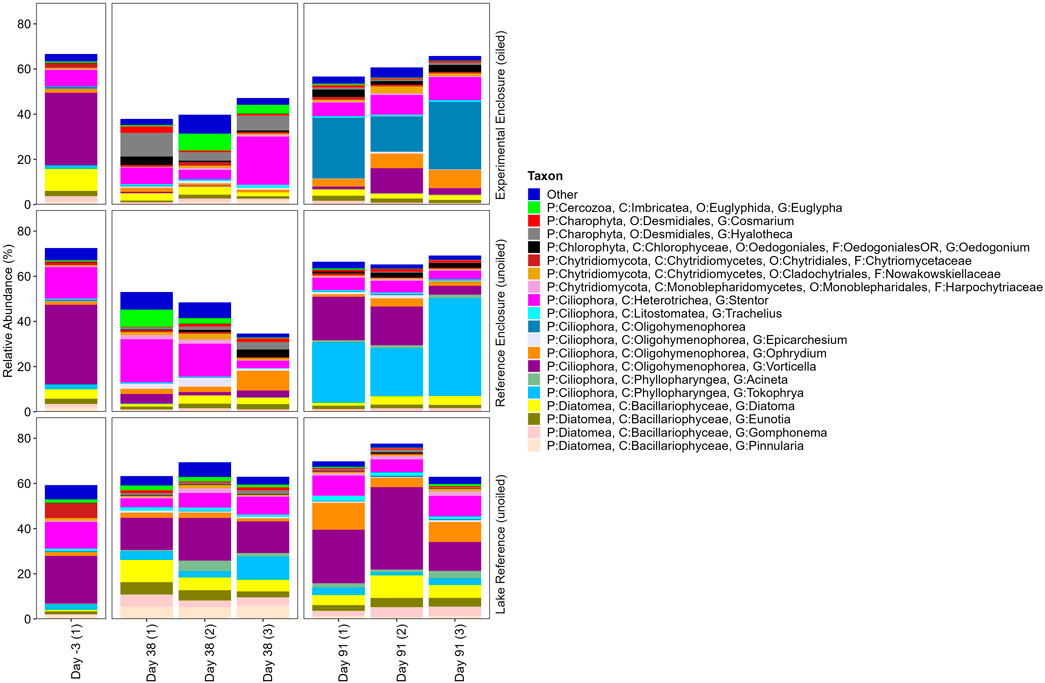
Figure 6. Taxonomic profile of the top twenty most relatively abundant 18S ASVs in EFW root biofilm replicates from the EE, RE, and LR 3 days pre-oil addition, and on days 38 and 91 post-oil addition. Taxon legend includes full taxonomic details to the lowest level identified as available.
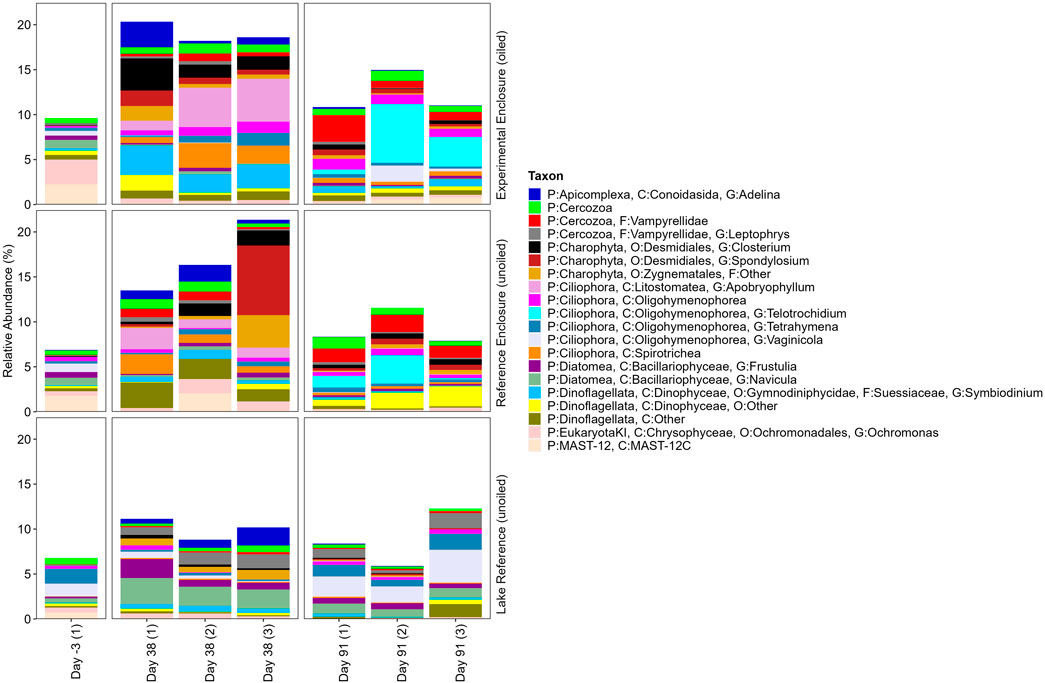
Figure 7. Taxonomic profile of the top twenty-one to forty most relatively abundant 18S ASVs in EFW root biofilm replicates from the EE, RE, and LR 3 days pre-oil addition, and on days 38 and 91 post-oil addition. Taxon legend includes full taxonomic details to the lowest level identified as available.
Some fungi are known hydrocarbon degraders due to their range of substrate specificity and enzyme production (e.g., peroxidases, laccases, monooxygenases) (Haritash and Kaushik, 2009; Kadri et al., 2017). Three unidentified ASVs belonging to phylum Chytridiomycota were identified in the top 20 eukaryotic ASVs. Chytridiomycota, often referred to as zoosporic fungi, are mostly found in aquatic ecosystems (Jobard et al., 2010; Prince, 2018; Naranjo-Ortiz and Gabaldón, 2019), and some are potential aromatic hydrocarbon degraders (Prince, 2018). For example, Phlyctochytrium reinboldtae were able to produce some metabolites of naphthalene, though mostly in small trace amounts (Cerniglia et al., 1978). Family Chytriomycetaceae were variable in RA among sites and time periods, but highest in the LR on day −3 (6.66%). Family Nowakowskiellaceae was also variable over the exposure period in reference locations, however increased in the EE, from 0.75% to 0.92%–3.18% on day −3 and 91, respectively. Family Harpochytriaceae had low RA in the EE (<1.00%), peaking on day 38 in the RE (0.21%–1.80%), while was variable for LR samples, ranging from 0.38% to 2.08% and 0.96% to 2.02% by days 38 and 91, respectively.
Algae are essential organisms involved in ecosystem productivity and function, and some are capable of metabolizing PACs (Ghosal et al., 2016; Prince, 2018). The top 40 ASVs had algal representatives from phylas Chlorophyta, Diatomea, Charophyta, and Dinoflagellata. Oedogonium, a filamentous green algae, had the highest RA by day 38 in the enclosures, increasing from 0.13% to 0.61%–3.68% in the EE, and from 0.12% to 0.22%–3.36% in the RE, while the LR had <0.50% RA on days 38 and 91. Oedogonium has been observed to tolerate vegetable oil spills and has affinity for low oxygen and nutrient rich conditions (Selala et al., 2014), while others found they were unable to adapt to crude oil presence (Oberholster et al., 2014). It has also been targeted for wastewater bioremediation and biofuel production (Adesalu et al., 2016 and citations within). Another green algae ASV from order Zygnematales increased by day 38 in all sites, highest in the RE. RAs ranged from 0.41% to 1.64%, 0.13% to 3.61%, and 0.61% to 1.05% in the EE, RE, and LR, respectively. Ochromonas, a chrysophyte algae, was highest on day −3 in the EE (2.76%) and by day 38 in the RE (0.33%–1.57%), while had low RA (<1.00%) in all other sample periods and sites.
There was a clear shift in diatom and desmid abundance between the enclosures and LR, which may result from differing site conditions. Desmid algae increased by day 38, often higher in the EE, while diatoms were often higher in the LR, similar to findings in Kamalanathan et al. (2021). Bruno et al. (1982) found periphyton communities from South Carolina streams dominated by desmid algae had greater uptake of benzo[a]pyrene than those dominated by diatoms, noting desmid cell structure and features (e.g., sheath) may be important for cellular sorption. Desmid Hyalotheca increased from 0.06% to 3.77%–10.51% in the EE by day 38, while RAs ranged from 0.95% to 3.41% and 0.75% to 1.80% in the RE and LR, respectively. Similarly, Cosmarium and Closterium were highest in the EE by day 38. Closterium sp. has previously been studied for degradation of Bonny light crude oil under different light regimes. It was observed that degradation was greater without light, when oil was the only source of carbon (Uzoh et al., 2015). Spondylosium increased by day 38 only in the enclosures from 0.00% on day −3 to 0.55%–1.73% and 0.00%–7.75% in the EE and RE, respectively. Diatoma had the largest RA of all diatoms (0.76%–10.01%), highest in the LR by day 91, while RA generally declined in the enclosures over the exposure period. Other diatoms peaked in the LR on day 38, including Eunotia (2.62%–5.50%), Gomphonema (2.92%–5.31%), Pinnularia (5.23%–5.86%), Frustulia (0.75%–2.10%), and Navicula (2.13%–2.90%), while RA in the enclosures were variable or generally declined over the exposure period. While RA was higher in the LR, some diatoms have been associated with oil or hydrocarbons. For example, Cerniglia et al. (1982) confirmed Navicula sp. could oxidize naphthalene to 1-naphthol at cool temperatures, while others observed temporal shifts to diatom assemblages (Eunotia, Gomphonema, and Pinnularia) after a vegetable and diesel oil spill in the Negro River, Brazil, based on diatom ecological guild (motile, erect, planktonic, and colonial) (De Faria et al., 2019). The variability of diatoms in our study may be the response of ecological guilds to environmental conditions or disturbance over the experimental period.
Dinoflagellates are common planktonic organisms found in aquatic environments and include both autotrophic and heterotrophic organisms. Some (e.g., Symbiodinium pilosum) are endosymbionts (Levy et al., 2007), and some have been found to tolerate or flourish during oil spill events or experiments (Kamalanathan et al., 2021). Symbiodinium and two unidentified dinoflagellates were identified in the top 40 ASVs. Symbiodinium had low RA (<1.00%) excluding samples from the EE on day 38 (2.02%–3.26%). One of the unidentified dinoflagellates had low RA (<1.00%) except day 38 in the EE (0.26%–1.74%) and day 91 in the RE (0.70%–2.23%). Similarly, the other unidentified dinoflagellate had low RA (<1.00%) except on day 38 in the RE (1.37%–2.83%) and on day 91 in the LR (0.04%–1.45%). Previous research found that dinoflagellates can be enhanced by the presence of oil degrading bacteria and growth promoting capabilities (Park et al., 2020), and others found that some marine species can ingest oil droplets under varying conditions (with or without food/chemical dispersants) (Almeda et al., 2014). Research by Almeda et al. (2018) found that oil and chemical dispersants disrupt the grazing pressure by heterotrophic dinoflagellates and ciliates, which allow for more tolerant, bloom forming dinoflagellates to flourish.
Similarly, some ciliates are natural predators to microorganisms and are ubiquitous in freshwater environments (Lara et al., 2007; Esteban et al., 2015; Robinson et al., 2022), however have also been studied for hydrocarbon degradation potential and tolerance to toxicity. Kachieng’a and Momba (2018, 2017) measured successful co-metabolism of petroleum hydrocarbons with a ciliate consortia, however Prince (2018) questioned whether it was biodegradation by the protozoans or by synergistic bacteria. Others have observed that ciliate grazing can result in bioaccumulation of oil (Rogerson and Berger, 1981) and can enhance bacterial biodegradation (Tso and Taghon, 2006), and their movement may result in oil dispersion in the water column, which could facilitate biodegradation (Gilbert et al., 2014). In fact, Holubar et al. (2000) found that presence of protozoan ciliates enhanced chemical oxygen demand degradation in petroleum contaminated sewage sludge treatment. Some researchers observed correlation between ciliate richness and high molecular weight PAHs (Moss et al., 2015), while others have found sensitivity to crude oil and dispersants (Almeda et al., 2018), indicating variable species tolerance to environmental conditions and pollutants (Jousset et al., 2010; Lara and Acosta-Mercado, 2012; Robinson et al., 2022).
There were fourteen ciliate genera in the top 40 ASVs that varied in dominance between sites and over time. Vorticella dominated the community on day −3 (21.15%–35.23%), however declined by day 38 in the enclosures (0.00%–4.23%) while remaining dominant in the LR (14.03%–18.93%), later increasing in all three sites (1.22%–36.54%). There were other dominant ciliates that had variable RA over the exposure period or between sites and replicates, such as Stentor (3.27%–21.37%) and Ophrydium (1.04%–11.88%), while some were site specific. Tokophrya was highest in the RE by day 91 (21.30%–43.27%), while it was much lower in the EE (0.00%) and LR (1.49%–3.21%). An unidentified ASV from class Oligohymenophorea increased drastically by day 91 in the EE, from 1.11% on day −3 to 15.74%–29.97%, while the RE and LR were <1.00%. Palace et al. (2021) previously identified unique operational taxonomic units of Oligohymenophorea in water of oil treated sites compared to unoiled sites in a shoreline enclosure study at the IISD-ELA. While research by Lara et al. (2007) found greater presence of Oligohymenophorea in non-polluted soil compared to PAH polluted soil. Telotrochidium increased from 0.00% on day −3 to 0.51%–6.54% and 0.15%–3.16% RA by day 91 in the EE and RE, respectively, while none were detected in the LR. Apobryophyllum increased from 0.00% to 1.08%–4.77% and 0.96%–2.38% in the EE and RE by day 38, respectively, and an unknown ASV from class Spirotrichea increased from 0.00% to 0.64%–2.73% and 0.70%–2.17%, respectively. Aprobryophullum belongs to class Litostomatea, which was previously identified as a dominant representative in an oil sand tailing reclamation site, and has been hypothesized to include species that have a high tolerance to hydrocarbon contamination (Richardson et al., 2020). Vagnicola had lower ciliate RA but increased in the LR by day 91. RA ranged from 0.03% to 1.80%, 0.00% to 0.06%, and 1.79% to 3.66% in the EE, RE, and LR by day 91, respectively. While shifts in our study cannot be attributed to oil, the dominance of ciliates may play a role in biodegradation (directly or by enhancing bacterial biodegradation), and/or other important biogeochemical processes.
Cercozoa, another group of bacterial grazers (Esteban et al., 2015), were in the top 40 ASVs, which similarly have had variable responses to oil. Dalby et al. (2008) identified Cercozoa as the dominant organism in chronically polluted seawater, but not in oligotrophic or crude oil contaminated microcosms. While Cobanli et al. (2022) found that Novel Clade 2 of Cercozoa increased between day 6 and 10 following dilbit additions to seawater microorganisms during the spring, but not the summer. ASV from family Vampyrellidae increased in the enclosures by day 91, from 0.78% to 2.96% and 1.34% to 1.92% in the EE and RE, respectively, and was <1.00% in the LR. Leptophrys was highest in the LR by days 38 (0.86%–1.55%) and 91 (0.32%–1.67%), while it comprised <1.00% in the enclosures. Euglypha increased in all sites by day 38, ranging from 0.58% to 7.44%, 0.36% to 7.67%, and 1.17% to 2.08%, in the EE, RE, and LR, respectively, while remaining samples were <1.00%, excluding day −3 in the LR (1.51%).
4 Conclusion
A contained oil spill was conducted in a wetland shoreline enclosure on Lake 260 at the IISD-ELA in order to better understand how microbial communities on EFW roots change following exposure to dilbit and to assess the potential of EFWs as a secondary remediation method following primary recovery of shoreline washing. There were greater shifts in diversity of eukaryotes on EFW roots than prokaryotes, but it was difficult to confirm if microbial community changes were attributed to oil exposure, enclosure effects, or seasonal change. However, organisms capable of degrading hydrocarbons or PACs, and those known to support degradation, were identified in this study, and some of these increased in the EE after oil addition.
While microbial results were inconclusive, PACs in the aqueous environment returned to near background conditions after ∼60 days, and fluctuations were linked to environmental conditions, further supporting the need to test efficiency of remediation efforts in the natural environment. It is possible the PACs were adsorbed to the sediment, underwent biodegradation, or further weathering processes. Sediment PACs were highly influenced by pyrogenic sources, however PAC concentrations increased in the EE after oil addition until day 91, later declining by day 263, at which point total PACs were similar among oiled and reference sites.
While the lack of enclosure replication limits our capacity to assess statistical significance of microbial and chemical changes among sites, this research responded to scientific and industrial research needs and confirmed that EFWs can support a naturally diverse microbial community, including those with oil degradation potential. This may be important for co-metabolism and/or other supporting processes. To our knowledge, this was the first, in-lake experiment to test EFWs for oil spill response. This research advances our knowledge in this field; however, we suggest that future field research be replicated to statistically conclude whether presence of oil causes a shift to hydrocarbon degrader dominance. We also believe there is a need to assess different oil products, rates of PAC degradation/removal, and EFW surface area requirements. Further, we suggest that researchers analyze microbial activity and/or gene expression, using methods such as metatranscriptomics or proteomics, to determine whether there are functional changes to the microbial community when exposed to oil, which will improve the collective knowledge on EFW potential to naturally remediate oil spills.
EFWs are currently deployed around the world as a cost-effective bioremediation strategy for various contaminants and aquatic environments, and their use for crude oil and hydrocarbon remediation is a relatively new and growing field. The current literature shows great potential for this method as a non-invasive strategy for oil spills, and continued research will provide guidance to spill responders on how to optimize deployments to enhance oil spill remediation.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: https://www.ncbi.nlm.nih.gov/, BioProject PRJNA1175668. The remaining datasets presented in this article are not readily available in online repositories as this work is part of a collaborative project, and some data presented may be used to interpret results in other partnering articles not yet published. Requests to access these datasets should be directed to the IISD Experimental Lakes Area Data Request page, available at: https://www.iisd.org/ela/researchers/data-requests/.
Author contributions
MS: Conceptualization, Data curation, Formal Analysis, Investigation, Methodology, Visualization, Writing–original draft. LP: Data curation, Investigation, Methodology, Writing–review and editing. AG: Investigation, Writing–review and editing. JT: Data curation, Formal Analysis, Investigation, Writing–review and editing. JW: Data curation, Investigation, Writing–review and editing. LT: Investigation, Writing–review and editing. CG: Conceptualization, Data curation, Funding acquisition, Investigation, Methodology, Resources, Writing–review and editing. JR: Writing–review and editing. TH: Data curation, Writing–review and editing. SH: Data curation, Investigation, Resources, Writing–review and editing. RG: Conceptualization, Funding acquisition, Investigation, Writing–review and editing. ET: Investigation, Methodology, Writing–review and editing. GT: Funding acquisition, Resources, Writing–review and editing. DL: Funding acquisition, Supervision, Writing–review and editing. VP: Conceptualization, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Writing–review and editing.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This research was funded by Genome Canada [GAPP R13-6336]; Mitacs Accelerate [Grant Number IT15202]; NSERC Collaborative Research and Development grant awarded to Gregg Tomy [NSERC File CRDPJ 532225-2018], and contributions from the International Institute for Sustainable Development Experimental Lakes Area. Funding was provided by Frontiers of Synthetic Biology to publish this article as open access. The authors declare that the FLOWTER study received Industrial partner support from the Canadian Association of Petroleum Producers, the Canadian Energy Pipeline Association [defunct], and the Myera Group, and in -kind contribution National Energy Board (now Canada Energy Regulator), TransCanada Pipelines, TransMountain Pipelines, and Enbridge. These funders were not involved in the collection, analysis, interpretation of data, the writing of this article, or the decision to submit it for publication.
Acknowledgments
Authors would like to acknowledge that this research was conducted at the International Institute for Sustainable Development Experimental Lakes Area field station, situated on the traditional land of the Anishinaabe Nation in Treaty 3 Territory and the homeland of the Métis Nation, and in Treaty 1 Territory, the ancestral lands of the Anishinaabe (Ojibwe), Ininiw (Cree), Anisininew (Ojibwe Cree), Dene, and Dakota Nations, and the homeland of the Red River Métis Nation. We thank the Digital Research Alliance of Canada for access to the Graham high performance compute cluster for the processing and analysis of DNA sequence data. We would like to acknowledge and thank all collaborators and students who supported project set up, and all researchers at the IISD Experimental Lakes Area for support and access to unpublished meteorological data. This article is part of a series of manuscripts from a doctoral dissertation (Stanley, 2024a), available online on 30 September 2025 at http://hdl.handle.net/1993/38302.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fsybi.2025.1517337/full#supplementary-material
Abbreviations
ASV, Amplicon sequence variant; Dilbit, Diluted bitumen; EE, Experimental enclosure; EFW, Engineered Floating Wetland; FLOWTER, FLOating Wetland Treatments to Enhance Remediation; IISD-ELA, International Institute for Sustainable Development Experimental Lakes Area; LR, Lake reference; PAC, Polycyclic aromatic compound; PAH, Polycyclic aromatic hydrocarbon; PCoA, Principal coordinate analysis; RE, Reference enclosure; RA, Relative abundance; rRNA, Ribosomal ribonucleic acid; SD, Standard deviation.
Footnotes
1Data presented from the Canadian Crude Oil Exports: A 30 Year Review (Canada Energy Regulator, 2021a; Canada Energy Regulator, 2021b) contains information licensed under the Open Government Licence- Canada. https://open.canada.ca/en/open-government-licence-canada
References
Abbasian, F., Palanisami, T., Megharaj, M., Naidu, R., Lockington, R., and Ramadass, K. (2016). Microbial diversity and hydrocarbon degrading gene capacity of a crude oil field soil as determined by metagenomics analysis. Biotechnol. Prog. 32, 638–648. doi:10.1002/btpr.2249
Abed, R. M. M., Al-Kharusi, S., Prigent, S., and Headley, T. (2014). Diversity, distribution and hydrocarbon biodegradation capabilities of microbial communities in oil-contaminated cyanobacterial mats from a constructed wetland. PLoS One 9, 1145700–e114622. doi:10.1371/journal.pone.0114570
Aburto-Medina, A., Adetutu, E. M., Aleer, S., Weber, J., Patil, S. S., Sheppard, P. J., et al. (2012). Comparison of indigenous and exogenous microbial populations during slurry phase biodegradation of long-term hydrocarbon-contaminated soil. Biodegradation 23, 813–822. doi:10.1007/s10532-012-9563-8
Adesalu, T. A., Julius, M. L., and Temenu, T. O. (2016). Molecular characterization, lipid analysis and GC-MS determination of bioactive compounds identified in a West African strain of the green alga Oedogonium (Chlorophyta). J. Pharmacogn. Phytochem. 5, 01–06.
Almeda, R., Connelly, T. L., and Buskey, E. J. (2014). Novel insight into the role of heterotrophic dinoflagellates in the fate of crude oil in the sea. Sci. Rep. 4, 7560–7569. doi:10.1038/srep07560
Almeda, R., Cosgrove, S., and Buskey, E. J. (2018). Oil spills and dispersants can cause the initiation of potentially harmful dinoflagellate blooms (“Red Tides”). Environ. Sci. Technol. 52, 5718–5724. doi:10.1021/acs.est.8b00335
Ankley, P. J., Xie, Y., Black, T. A., DeBofsky, A., Perry, M., Paterson, M. J., et al. (2021). Using zooplankton metabarcoding to assess the efficacy of different techniques to clean-up an oil-spill in a boreal lake. Aquat. Toxicol. 236, 105847. doi:10.1016/j.aquatox.2021.105847
Beazley, M. J., Martinez, R. J., Rajan, S., Powell, J., Piceno, Y. M., Tom, L. M., et al. (2012). Microbial community analysis of a coastal salt marsh affected by the Deepwater Horizon oil spill. PLoS One 7, e41305. doi:10.1371/journal.pone.0041305
Bodour, A. A., Wang, J. M., Brusseau, M. L., and Maier, R. M. (2003). Temporal change in culturable phenanthrene degraders in response to long-term exposure to phenanthrene in a soil column system. Environ. Microbiol. 5, 888–895. doi:10.1046/j.1462-2920.2003.00481.x
Brandt, J. P., Flannigan, M. D., Maynard, D. G., Thompson, I. D., and Volney, W. J. A. (2013). An introduction to Canada’s boreal zone: ecosystem processes, health, sustainability, and environmental issues. Environ. Rev. 21, 207–226. doi:10.1139/er-2013-0040
Brown, D., Wilson, S., Caffrey, S. M., Bordenave, S., Gieg, L. M., Ramos-Padron, E., et al. (2013). Microbial community and potential functional gene diversity involved in anaerobic hydrocarbon degradation and methanogenesis in an oil sands tailings pond. Genome 56, 612–618. doi:10.1139/gen-2013-0083
Bruno, M. G., Fannin, T. E., and Leversee, G. J. (1982). The disposition of benzo(a)pyrene in the periphyton communities of two South Carolina streams: uptake and biotransformation. Can. J. Bot. 60, 2084–2091. doi:10.1139/b82-255
Callahan, B. J., McMurdie, P. J., Rosen, M. J., Han, A. W., Johnson, A. J. A., and Holmes, S. P. (2016). DADA2: high-resolution sample inference from Illumina amplicon data. Nat. Methods 13, 581–583. doi:10.1038/nmeth.3869
Canada Energy Regulator (2021a) Annual Canadian crude oil production by type, Data from Canadian crude oil exports: a 30 year review. doi:10.35002/zd9v-jj06
Canada Energy Regulator (2021b) Annual crude oil exports volume by type, Data from Canadian crude oil exports: a 30 year review. https://open.canada.ca/en/open-government-licence-canada.
Canada Energy Regulator (2021c). Canadian crude oil exports: a 30 Year review. Available at: https://www.cer-rec.gc.ca/en/data-analysis/energy-commodities/crude-oil-petroleum-products/report/canadian-crude-oil-exports-30-year-review/index.html (Accessed February 23, 2022).
Cao, B., Nagarajan, K., and Loh, K. C. (2009). Biodegradation of aromatic compounds: current status and opportunities for biomolecular approaches. Appl. Microbiol. Biotechnol. 85, 207–228. doi:10.1007/s00253-009-2192-4
Cébron, A., Borreca, A., Beguiristain, T., Biache, C., and Faure, P. (2022). Taxonomic and functional trait-based approaches suggest that aerobic and anaerobic soil microorganisms allow the natural attenuation of oil from natural seeps. Sci. Rep. 12, 7245–7316. doi:10.1038/s41598-022-10850-4
Cerniglia, C. E., Gibson, D. T., and Van Baalen, C. (1982). Naphthalene metabolism by diatoms isolated from the Kachemak Bay region of Alaska. J. Gen. Microbiol. 128, 987–990. doi:10.1099/00221287-128-5-987
Cerniglia, C. E., Hebert, R. L., Szaniszlo, P. J., and Gibson, D. T. (1978). Fungal transformation of naphthalene. Arch. Microbiol. 117, 135–143. doi:10.1007/BF00402301
Chang, S. E., Stone, J., Demes, K., and Piscitelli, M. (2014). Consequences of oil spills: a review and framework for informing planning. Ecol. Soc. 19, 26. doi:10.5751/ES-06406-190226
Chettri, B., and Singh, A. K. (2019). Kinetics of hydrocarbon degradation by a newly isolated heavy metal tolerant bacterium Novosphingobium panipatense P5:ABC. Bioresour. Technol. 294, 122190. doi:10.1016/j.biortech.2019.122190
Cobanli, S. E., Wohlgeschaffen, G., Ryther, C., Macdonald, J., Gladwell, A., Watts, T., et al. (2022). Microbial community response to simulated diluted bitumen spills in coastal seawater and implications for oil spill response. FEMS Microbiol. Ecol. 98, fiac033–13. doi:10.1093/femsec/fiac033
Dalby, A. P., Kormas, K. A., Christaki, U., and Karayanni, H. (2008). Cosmopolitan heterotrophic microeukaryotes are active bacterial grazers in experimental oil-polluted systems. Environ. Microbiol. 10, 47–56. doi:10.1111/j.1462-2920.2007.01428.x
Dearnley, J. (2022). Assessing exposure of small-bodied fish to experimental crude oil spills in contained shoreline environments using biliary polycyclic aromatic compounds metabolites. Winnipeg, MB: University of Manitoba. [Master's Thesis].
Dedysh, S. N. (2019). “Bryobacter,” in Bergey’s manual of systematics of archaea and bacteria (John Wiley and Sons, Inc.), 1–5. in association with Bergey’s Manual Trust). doi:10.1002/9781118960608.gbm01667
De Faria, D. M., Costin, J. C., Tremarin, P. I., and Ludwig, T. A. V. (2019). Temporal changes in biological traits of diatom communities in response to an oil spill in a subtropical river. An. Acad. Bras. Cienc. 91, 201708633–e20170917. doi:10.1590/0001-3765201920170863
Deutzmann, J. S., Hoppert, M., and Schink, B. (2014). Characterization and phylogeny of a novel methanotroph, Methyloglobulus morosus gen. nov., spec. nov. Syst. Appl. Microbiol. 37, 165–169. doi:10.1016/j.syapm.2014.02.001
Dubinsky, E. A., Conrad, M. E., Chakraborty, R., Bill, M., Borglin, S. E., Hollibaugh, J. T., et al. (2013). Succession of hydrocarbon-degrading bacteria in the aftermath of the deepwater horizon oil spill in the Gulf of Mexico. Environ. Sci. Technol. 47, 10860–10867. doi:10.1021/es401676y
Dupuis, A., and Ucan-Marin, F. (2015) A literature review on the aquatic toxicology of petroleum oil: an overview of oil properties and effects to aquatic biota. Ottawa, ON: DFO Canadian Science Advisory Secretariat Research Document. Available at: https://publications.gc.ca/collections/collection_2015/mpo-dfo/Fs70-5-2015-007-eng.pdf.
Engel, A. S., Liu, C., Paterson, A. T., Anderson, L. C., Eugene Turner, R., and Overton, E. B. (2017). Salt marsh bacterial communities before and after the Deepwater Horizon oil spill. Appl. Environ. Microbiol. 83, 007844-17–e822. doi:10.1128/AEM.00784-17
Esteban, G. F., Finlay, B. J., and Warran, A. (2015). “Free-living protozoa,” in Thorp and covinch’s freshwater invertebrates: ecology and general Biology. Editors J. H. Thorp,, and D. C. Rogers (Elsevier Inc.), 113–132.
Fahde, S., Boughribil, S., Sijilmassi, B., and Amri, A. (2023). Rhizobia: a promising source of plant growth-promoting molecules and their non-legume interactions: examining applications and mechanisms. Agriculture 13, 1279. doi:10.3390/agriculture13071279
Floating Islands International (2024). Products. Available at: https://www.floatingislandinternational.com/products.html#biohavens (Accessed October 11, 2022).
Franchi, E., Cardaci, A., Pietrini, I., Fusini, D., Conte, A., De Folly D’Auris, A., et al. (2022). Nature-based solutions for restoring an agricultural area contaminated by an oil spill. Plants 11, 2250. doi:10.3390/plants11172250
Gabos, S., Ikonomou, M. G., Schopflocher, D., Fowler, B. R., White, J., Prepas, E., et al. (2001). Characteristics of PAHs, PCDD/Fs and PCBs in sediment following forest fires in northern Alberta. Chemosphere 43, 709–719. doi:10.1016/S0045-6535(00)00424-0
Garrido-Oter, R., Nakano, R. T., Dombrowski, N., Ma, K. W., McHardy, A. C., Schulze-Lefert, P., et al. (2018). Modular traits of the Rhizobiales root microbiota and their evolutionary relationship with symbiotic Rhizobia. Cell Host Microbe 24, 155–167.e5. doi:10.1016/j.chom.2018.06.006
Ghosal, D., Ghosh, S., Dutta, T. K., and Ahn, Y. (2016). Current state of knowledge in microbial degradation of polycyclic aromatic hydrocarbons (PAHs): a review. Front. Microbiol. 7, 1369. doi:10.3389/fmicb.2016.01369
Gilbert, D., Jakobsen, H. H., Winding, A., and Mayer, P. (2014). Co-transport of polycyclic aromatic hydrocarbons by motile microorganisms leads to enhanced mass transfer under diffusive conditions. Environ. Sci. Technol. 48, 4368–4375. doi:10.1021/es404793u
Glöckner, F. O., Kube, M., Bauer, M., Teeling, H., Lombardot, T., Ludwig, W., et al. (2003). Complete genome sequence of the marine planctomycete Pirellula sp. strain 1. Proc. Natl. Acad. Sci. U. S. A. 100, 8298–8303. doi:10.1073/pnas.1431443100
Glöckner, F. O., Yilmaz, P., Quast, C., Gerken, J., Beccati, A., Ciuprina, A., et al. (2017). 25 years of serving the community with ribosomal RNA gene reference databases and tools. J. Biotechnol. 261, 169–176. doi:10.1016/j.jbiotec.2017.06.1198
Haritash, A. K., and Kaushik, C. P. (2009). Biodegradation aspects of polycyclic aromatic hydrocarbons (PAHs): a review. J. Hazard. Mater. 169, 1–15. doi:10.1016/j.jhazmat.2009.03.137
Hoff, R. Z. (1995). Responding to oil spills in coastal marshes: the fine line between help and hindrance. HAZMAT Report 96-2. Seattle, WA: Hazardous Materials Response and Assessments Division, NOAA. Available at: https://response.restoration.noaa.gov/sites/default/files/Coastal_Marshes_508.pdf.
Holubar, P., Grudke, T., Moser, A., Strenn, B., and Braun, R. (2000). Effects of bacterivorous ciliated protozoans on degradation efficiency of a petrochemical activated sludge process. Water Res. 34, 2051–2060. doi:10.1016/S0043-1354(99)00378-4
Idomeh, J. E., Shittu, O. B., Oyedepo, J. A., Bada, B. S., Balogun, S. A., Idomeh, F. A., et al. (2021). Petroleum hydrocarbon impacted aquatic ecosystem reveals Methylotenera as the dominant genera in the Niger Delta Region of Nigeria. Geomicrobiol. J. 38, 879–894. doi:10.1080/01490451.2021.1973614
Idowu, I., Francisco, O., Thomas, P. J., Johnson, W., Marvin, C., Stetefeld, J., et al. (2018). Validation of a simultaneous method for determining polycyclic aromatic compounds and alkylated isomers in biota. Rapid Commun. Mass Spectrom. 32, 277–287. doi:10.1002/rcm.8035
IISD Experimental Lakes Area (IISD-ELA) (2022). IISD experimental lakes area: bathymetry data package. doi:10.6073/pasta/e9898c78448cbbe544057e689d3c06df
Jobard, M., Rasconi, S., and Sime-Ngando, T. (2010). Diversity and functions of microscopic fungi: a missing component in pelagic food webs. Aquat. Sci. 72, 255–268. doi:10.1007/s00027-010-0133-z
Jousset, A., Lara, E., Nikolausz, M., Harms, H., and Chatzinotas, A. (2010). Application of the denaturing gradient gel electrophoresis (DGGE) technique as an efficient diagnostic tool for ciliate communities in soil. Sci. Total Environ. 408, 1221–1225. doi:10.1016/j.scitotenv.2009.09.056
Kachieng’a, L., and Momba, M. N. B. (2017). Biodegradation of petroleum hydrocarbon functional groups in contaminated water using selected protozoan isolates. J. Environ. Chem. Eng. 5, 1885–1891. doi:10.1016/j.jece.2017.03.022
Kachieng’a, L., and Momba, M. N. B. (2018). The synergistic effect of a consortium of protozoan isolates (Paramecium sp., Vorticella sp., Epistylis sp. and Opercularia sp.) on the biodegradation of petroleum hydrocarbons in wastewater. J. Environ. Chem. Eng. 6, 4820–4827. doi:10.1016/j.jece.2018.07.005
Kadri, T., Rouissi, T., Kaur Brar, S., Cledon, M., Sarma, S., and Verma, M. (2017). Biodegradation of polycyclic aromatic hydrocarbons (PAHs) by fungal enzymes: a review. J. Environ. Sci. 51, 52–74. doi:10.1016/j.jes.2016.08.023
Kamalanathan, M., Schwehr, K. A., Labonté, J. M., Taylor, C., Bergen, C., Patterson, N., et al. (2021). The interplay of phototrophic and heterotrophic microbes under oil exposure: a microcosm study. Front. Microbiol. 12, 675328. doi:10.3389/fmicb.2021.675328
Kertesz, M. A., and Kawasaki, A. (2010). “Hydrocarbon-degrading sphingomonads: sphingomonas, spingobium, Novosphingobium, and sphingopyxis,” in Handbook of hydrocarbon and lipid microbiology. Editor K. N. Timmis (Berlin, Heidelberg: Springer), 1693–1705. doi:10.1007/978-3-540-77587-4_119
Kharey, G. S., Palace, V., Whyte, L., and Greer, C. W. (2024). Native freshwater lake microbial community response to an in situ experimental dilbit spill. FEMS Microbiol. Ecol. 100, fiae055. doi:10.1093/femsec/fiae055
King, G. M., Kostka, J. E., Hazen, T. C., and Sobecky, P. A. (2015). Microbial responses to the Deepwater Horizon Oil spill: from coastal wetlands to the deep sea. Ann. Rev. Mar. Sci. 7, 377–401. doi:10.1146/annurev-marine-010814-015543
Koo, H., Mojib, N., Huang, J. P., Donahoe, R. J., and Bej, A. K. (2015). Bacterial community shift in the coastal Gulf of Mexico salt-marsh sediment microcosm in vitro following exposure to the Mississippi Canyon Block 252 oil (MC252). 3 Biotech. 5, 379–392. doi:10.1007/s13205-014-0233-x
Kulichevskaya, I. S., Suzina, N. E., Liesack, W., and Dedysh, S. N. (2010). Bryobacter aggregatus gen. nov., sp. nov., a peat-inhabiting, aerobic chemo-organotroph from subdivision 3 of the acidobacteria. Int. J. Syst. Evol. Microbiol. 60, 301–306. doi:10.1099/ijs.0.013250-0
Lara, E., and Acosta-Mercado, D. (2012). A molecular perspective on ciliates as soil bioindicators. Eur. J. Soil Biol. 49, 107–111. doi:10.1016/j.ejsobi.2011.11.001
Lara, E., Berney, C., Harms, H., and Chatzinotas, A. (2007). Cultivation-independent analysis reveals a shift in ciliate 18S rRNA gene diversity in a polycyclic aromatic hydrocarbon-polluted soil. FEMS Microbiol. Ecol. 62, 365–373. doi:10.1111/j.1574-6941.2007.00387.x
Lee, K., Boufadel, M., Chen, B., Foght, J., Hodson, P., Swanson, S., et al. (2015). Expert panel report on the behaviour and environmental impacts of crude oil released into aqueous environments. Ottawa, ON: Royal Society of Canada.
Levy, M. G., Litaker, R. W., Goldstein, R. J., Dykstra, M. J., Vandersea, M. W., and Noga, E. J. (2007). Piscinoodinium, a fish-ectoparasitic dinoflagellate, is a member of the class Dinophyceae, subclass Gymnodiniphycidae: convergent evolution with Amyloodinium. J. Parasitol. 93, 1006–1015. doi:10.1645/GE-3585.1
Lyu, Y., Zheng, W., Zheng, T., and Tian, Y. (2014). Biodegradation of polycyclic aromatic hydrocarbons by Novosphingobium pentaromativorans US6-1. PLoS One 9, e101438. doi:10.1371/journal.pone.0101438
Magar, V. S., Chadwick, D. B., Bridges, T. S., Fuchsman, P. C., Conder, J. M., Dekker, T. J., et al. (2009). Technical guide: monitored natural recovery at contaminated sediment sites. Environ. Int. Corp. Available at: https://apps.dtic.mil/sti/pdfs/ADA512822.pdf.
McIlroy, S. J., and Nielsen, P. H. (2014). “The gamily Saprospiraceae,” in The Prokaryotes: other major lineages of bacteria and the archaea. Editors E. Rosenberg, E. F. De Long, S. Lory, E. Stackebrandt, and F. Thompson (Berlin, Heidelberg: Springer-Verlag Berlin Heidelberg), 863–889. doi:10.1007/978-3-642-38954-2
Moss, J. A., McCurry, C., Tominack, S., Romero, I. C., Hollander, D., Jeffrey, W. H., et al. (2015). Ciliated protists from the nepheloid layer and water column of sites affected by the Deepwater Horizon oil spill in the Northeastern Gulf of Mexico. Deep. Res. Part I Oceanogr. Res. Pap. 106, 85–96. doi:10.1016/j.dsr.2015.10.001
Murray, M. W., Allan, J. D., and Child, M. (2018). Potential ecological impacts of crude oil transport in the Great Lakes Basin. Gt. Lakes. Sci. Advis. Board, Sci. Prior. Comm. Available at: https://www.ijc.org/sites/default/files/2019-01/Potential%20Ecological%20Impacts%20of%20Crude%20Oil%20Transport%20in%20the%20Great%20Lakes%20Basin%20-%20Oct%202018.pdf.
Naczi, R. F. C. (1992). Systematics of Carex section griseae (cyperaceae). Ann Arbor (MI): University of Michigan. [PhD Thesis].
Naranjo-Ortiz, M. A., and Gabaldón, T. (2019). Fungal evolution: diversity, taxonomy and phylogeny of the Fungi. Biol. Rev. 94, 2101–2137. doi:10.1111/brv.12550
Natural Resources Canada (2020). Crude oil industry overview. Available at: https://natural-resources.canada.ca/our-natural-resources/energy-sources-distribution/fossil-fuels/crude-oil/crude-oil-industry-overview/18078 (Accessed April 26, 2023).
Newton, R. J., Jones, S. E., Eiler, A., McMahon, K. D., and Bertilsson, S. (2011). A guide to the natural history of freshwater lake bacteria. Microbiol. Mol. Biol. Rev. 75, 14–49. doi:10.1128/mmbr.00028-10
Oberholster, P., Cheng, P., Truter, C., Hill, L., and Botha, A. M. (2014). “Species specific responses of common freshwater wetland phytoplankton to crude oil by application of bioassays,” in Oil spills: environmental issues, prevention and ecological impacts. Editor A. Clifton (New York: Nova Science Publishers, Inc), 87–119.
Ozaki, S., Kishimoto, N., and Fujita, T. (2006). Isolation and phylogenetic characterization of microbial consortia able to degrade aromatic hydrocarbons at high rates. Microbes Environ. 21, 44–52. doi:10.1264/jsme2.21.44
Palace, V. P., Peters, L., Berard-Brown, N., Smyth, P., Shanoff, T., Wiseman, G., et al. (2021). “Assessing monitored natural recovery (MNR) for remediating crude oil spills in freshwater environments: the Freshwater Oil Spill Remediation Study (FOReSt) at the Experimental Lakes Area, Canada,” in 2021 international oil spill conference, 1–20.
Park, B. S., Erdner, D. L., Bacosa, H. P., Liu, Z., and Buskey, E. J. (2020). Potential effects of bacterial communities on the formation of blooms of the harmful dinoflagellate Prorocentrum after the 2014 Texas City “Y” oil spill (USA). Harmful Algae 95, 101802. doi:10.1016/j.hal.2020.101802
Peng, M., Zi, X., and Wang, Q. (2015). Bacterial community diversity of oil-contaminated soils assessed by high throughput sequencing of 16s rRNA genes. Int. J. Environ. Res. Public Health 12, 12002–12015. doi:10.3390/ijerph121012002
Pérez-Pantoja, D., Donoso, R., Agulló, L., Córdova, M., Seeger, M., Pieper, D. H., et al. (2012). Genomic analysis of the potential for aromatic compounds biodegradation in Burkholderiales. Environ. Microbiol. 14, 1091–1117. doi:10.1111/j.1462-2920.2011.02613.x
Pezeshki, S. R., Hester, M. W., Lin, Q., and Nyman, J. A. (2000). The effects of oil spill and clean-up on dominant US Gulf coast marsh macrophytes: a review. Environ. Pollut. 108, 129–139. doi:10.1016/S0269-7491(99)00244-4
Posit Team (2023). RStudio: integrated development environment for R. Available at: http://www.posit.co/.
Prince, R. C. (2018). “Eukaryotic hydrocarbon degraders,” in Taxonomy, genomics and ecophysiology of hydrocarbon-degrading microbes, Handbook of hydrocarbon and lipid microbiology. Editor T. J. McGenity (Springer), 1–20. doi:10.1007/978-3-540-77587-4_150
Prince, R. C., Amande, T. V., and McGenity, T. J. (2018). “Prokaryotic hydrocarbon degraders,” in Taxonomy, genomics and ecophysiology of hydrocarbon degrading microbes, handbook of hydrocarbon and lipid Mmcrobiology. Editor T. J. McGenity (Switzerland: Springer), 1–41. doi:10.1007/978-3-319-60053-6_15-1
Quast, C., Pruesse, E., Yilmaz, P., Gerken, J., Schweer, T., Yarza, P., et al. (2012). The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Res. 41, 590–596. doi:10.1093/nar/gks1219
Radice, R. P., Fabrizio, V.De, Donadoni, A., Scopa, A., and Martelli, G. (2023). Crude oil bioremediation: from bacteria to microalgae. Processes 11, 442. doi:10.3390/pr11020442
Ramdahl, T. (1983). Retene-a molecular marker of wood combustion in ambient air. Nature 306, 580–582. doi:10.1038/306580a0
R Core Team (2022). R: a language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing.
Rehman, K., Imran, A., Amin, I., and Afzal, M. (2018). Inoculation with bacteria in floating treatment wetlands positively modulates the phytoremediation of oil field wastewater. J. Hazard. Mater. 349, 242–251. doi:10.1016/j.jhazmat.2018.02.013
Ribicic, D., McFarlin, K. M., Netzer, R., Brakstad, O. G., Winkler, A., Throne-Holst, M., et al. (2018). Oil type and temperature dependent biodegradation dynamics - combining chemical and microbial community data through multivariate analysis. BMC Microbiol. 18, 83–15. doi:10.1186/s12866-018-1221-9
Richardson, E., Bass, D., Smirnova, A., Paoli, L., Dunfield, P., and Dacks, J. B. (2020). Phylogenetic estimation of community composition and novel eukaryotic lineages in Base Mine Lake: an oil sands tailings reclamation site in Northern Alberta. J. Eukaryot. Microbiol. 67, 86–99. doi:10.1111/jeu.12757
Robinson, C. V., Porter, T. M., McGee, K. M., McCusker, M., Wright, M. T. G., and Hajibabaei, M. (2022). Multi-marker DNA metabarcoding detects suites of environmental gradients from an urban harbour. Sci. Rep. 12, 10556–10612. doi:10.1038/s41598-022-13262-6
Rodriguez-Gil, J. L., Stoyanovich, S., Hanson, M. L., Hollebone, B., Orihel, D. M., Palace, V., et al. (2021). Simulating diluted bitumen spills in boreal lake limnocorrals - Part 1: experimental design and responses of hydrocarbons, metals, and water quality parameters. Sci. Total Environ. 790, 148537. doi:10.1016/J.SCITOTENV.2021.148537
Rogerson, A., and Berger, J. (1981). Effect of crude oil and petroleum-degrading micro-organisms on the growth of freshwater and soil protozoa. J. Gen. Microbiol. 124, 53–59. doi:10.1099/00221287-124-1-53
Rognes, T., Flouri, T., Nichols, B., Quince, C., and Mahé, F. (2016). VSEARCH: a versatile open source tool for metagenomics. PeerJ 4, e2584. doi:10.7717/peerj.2584
Rojo, F. (2021). A new global regulator that facilitates the co-metabolization of polyaromatic hydrocarbons and other nutrients in Novosphingobium. Environ. Microbiol. 23, 2875–2877. doi:10.1111/1462-2920.15527
Schoch, C., Ciufo, S., Domrachev, M., Hotton, C. L., Kannan, S., Khovanskaya, R., et al. (2020). NCBI Taxonomy: a comprehensive update on curation, resources and tools. Database 2020, baaa062. doi:10.1093/database/baaa062
Selala, C., Botha, A. M., De Klerk, L. P., De Klerk, A. R., Myburgh, J. G., and Oberholster, P. J. (2014). Using phytoplankton diversity to determine wetland resilience, one year after a vegetable oil spill. Water. Air. Soil Pollut. 225, 2051–2111. doi:10.1007/s11270-014-2051-5
SILVA, (2023). High quality ribosomal RNA databases. Available at: https://www.arb-silva.de/(Accessed August 8, 2023)
Shahid, M. J., Arslan, M., Ali, S., Siddique, M., and Afzal, M. (2018). Floating wetlands: a sustainable tool for wastewater treatment. Clean. - Soil, Air, Water 46. doi:10.1002/clen.201800120
Sohn, J. H., Kwon, K. K., Kang, J. H., Jung, H. B., and Kim, S. J. (2004). Novosphingobium pentaromativorans sp. nov., a high-molecular-mass polycyclic aromatic hydrocarbon-degrading bacterium isolated from estuarine sediment. Int. J. Syst. Evol. Microbiol. 54, 1483–1487. doi:10.1099/ijs.0.02945-0
Stanley, M. (2024a). Engineered floating wetlands as a secondary oil spill remediation strategy for freshwater shorelines. [PhD Thesis]. Winnipeg, MB: University of Manitoba.
Stanley, M. (2024b). Floating wetland treatments to enhance remediation (FLOWTER) study- diluted bitumen. Bethesda, Maryland: National Center for Biotechnology Information. BioProject PRJNA1175668 [Release Date February 14, 2025].
Stanley, M., Palace, V., Grosshans, R., and Levin, D. B. (2022). Floating treatment wetlands for the bioremediation of oil spills: a review. J. Environ. Manage. 317, 115416. doi:10.1016/j.jenvman.2022.115416
Starr, J. R., Naczi, R. F. C., and Chouinard, B. N. (2009). Plant DNA barcodes and species resolution in sedges (Carex, Cyperaceae). Mol. Ecol. Resour. 9, 151–163. doi:10.1111/j.1755-0998.2009.02640.x
Stoyanovich, S., Rodríguez-Gil, J. R., Hanson, M. L., Hollebone, B. P., Orihel, D. M., Palace, V. P., et al. (2021). Simulating diluted bitumen spills in boreal lake limnocorrals - part 2: factors affecting the physical characteristics and submergence of diluted bitumen. Sci. Total Environ. 790, 148580. doi:10.1016/j.scitotenv.2021.148580
Stoyanovich, S., Yang, Z., Hanson, M., Hollebone, B. P., Orihel, D. M., Palace, V., et al. (2022). Fate of polycyclic aromatic compounds from diluted bitumen spilled into freshwater limnocorrals. Sci. Total Environ. 819, 151993. doi:10.1016/j.scitotenv.2021.151993
Tanner, C. C., and Headley, T. R. (2011). Components of floating emergent macrophyte treatment wetlands influencing removal of stormwater pollutants. Ecol. Eng. 37, 474–486. doi:10.1016/j.ecoleng.2010.12.012
Taylor, N. M., Toth, C. R. A., Collins, V., Mussone, P., and Gieg, L. M. (2021). The effect of an adsorbent matrix on recovery of microorganisms from hydrocarbon-contaminated groundwater. Microorganisms 9, 90–19. doi:10.3390/microorganisms9010090
Thompson, H., Angelova, A., Bowler, B., Jones, M., and Gutierrez, T. (2017). Enhanced crude oil biodegradative potential of natural phytoplankton-associated hydrocarbonoclastic bacteria. Environ. Microbiol. 19, 2843–2861. doi:10.1111/1462-2920.13811
Tremblay, J., and Yergeau, E. (2019). Systematic processing of ribosomal RNA gene amplicon sequencing data. Gigascience 8, giz146–14. doi:10.1093/gigascience/giz146
Tso, S. F., and Taghon, G. L. (2006). Protozoan grazing increases mineralization of naphthalene in marine sediment. Microb. Ecol. 51, 460–469. doi:10.1007/s00248-006-9058-4
United States Environmental Protection Agency (2014). Priority pollutant list. 2. Available at: https://www.epa.gov/sites/default/files/2015-09/documents/priority-pollutant-list-epa.pdf (Accessed April 15, 2022).
Uzoh, C. V., Ifeanyi, V. O., Okwuwe, C. I., Oranusi, S. U., Braide, W., Iheukwumere, I. H., et al. (2015). Effect of light on the biodegradation of crude oil by the algae Closterium species. J. Nat. Sci. Res. 5, 112–118.
Vergnoux, A., Malleret, L., Asia, L., Doumenq, P., and Theraulaz, F. (2011). Impact of forest fires on PAH level and distribution in soils. Environ. Res. 111, 193–198. doi:10.1016/j.envres.2010.01.008
Wang, Q., Garrity, G. M., Tiedje, J. M., and Cole, J. R. (2007). Naïve Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 73, 5261–5267. doi:10.1128/AEM.00062-07
Wang, Y., Wang, J., Leng, F., and Chen, J. (2021). Effects of oil pollution on indigenous bacterial diversity and community structure of soil in Fushun, Liaoning Province, China. Geomicrobiol. J. 38, 115–126. doi:10.1080/01490451.2020.1817196
Wickham, H., Averick, M., Bryan, J., Chang, W., McGowan, L., François, R., et al. (2019). Welcome to the tidyverse. J. Open Source Softw. 4, 1686. doi:10.21105/joss.01686
Xiao, M., Zhang, Z. Z., Wang, J. X., Zhang, G. Q., Luo, Y. J., Song, Z. Z., et al. (2013). Bacterial community diversity in a low-permeability oil reservoir and its potential for enhancing oil recovery. Bioresour. Technol. 147, 110–116. doi:10.1016/j.biortech.2013.08.031
Yang, S., Wen, X., Shi, Y., Liebner, S., Jin, H., and Perfumo, A. (2016). Hydrocarbon degraders establish at the costs of microbial richness, abundance and keystone taxa after crude oil contamination in permafrost environments. Sci. Rep. 6, 37473. doi:10.1038/srep37473
Yavari, S., Malakahmad, A., and Sapari, N. B. (2015). A review on phytoremediation of crude oil spills. Water. Air. Soil Pollut. 226, 279–318. doi:10.1007/s11270-015-2550-z
Yilmaz, P., Parfrey, L. W., Yarza, P., Gerken, J., Pruesse, E., Quast, C., et al. (2014). The SILVA and “all-species living tree project (LTP)” taxonomic frameworks. Nucleic Acids Res. 42, 643–648. doi:10.1093/nar/gkt1209
Zhang, X., Shao, Y., Tian, J., Liao, Y., Li, P., Zhang, Y., et al. (2019). PTrimmer: an efficient tool to trim primers of multiplex deep sequencing data. BMC Bioinforma. 20, 236–6. doi:10.1186/s12859-019-2854-x
Zhu, X., Venosa, A. D., Suidan, M. T., and Lee, K. (2001). Guidelines for the bioremediation of marine shorelines and freshwater wetlands. Cincinnati, OH: U.S. Environmental Protection Agency. Available at: https://www.epa.gov/sites/default/files/2013-07/documents/guidelines_for_the_bioremediation_of_marine_shorelines_and_freshwater_wetlands.pdf.
Keywords: engineered floating wetland, diluted bitumen, oil spill, polycyclic aromatic compounds, microbial diversity, freshwater, bioremediation, floating treatment wetland
Citation: Stanley MJ, Peters L, Guttormson A, Tremblay J, Wasserscheid J, Timlick L, Greer CW, Rodríguez Gil JL, Halldorson T, Havens S, Grosshans R, Taylor E, Tomy G, Levin DB and Palace VP (2025) Assessing changes to the root biofilm microbial community on an engineered floating wetland upon exposure to a controlled diluted bitumen spill. Front. Synth. Biol. 3:1517337. doi: 10.3389/fsybi.2025.1517337
Received: 25 October 2024; Accepted: 20 January 2025;
Published: 24 February 2025.
Edited by:
Bryan J. Cassone, Brandon University, CanadaReviewed by:
Fiaz Ahmad, Northwestern Polytechnical University, ChinaTamer Abdelaziz, University of the Basque Country, Spain
Kexue Han, Tsinghua University, China
Copyright © 2025 Stanley, Peters, Guttormson, Tremblay, Wasserscheid, Timlick, Greer, Rodríguez Gil, Halldorson, Havens, Grosshans, Taylor, Tomy, Levin and Palace. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Madeline J. Stanley, bXN0YW5sZXlAaWlzZC1lbGEub3Jn; David B. Levin, ZGF2aWQubGV2aW5AdW1hbml0b2JhLmNh; Vince P. Palace, dnBhbGFjZUBpaXNkLWVsYS5vcmc=