 C. Dudreuilh1,2*
C. Dudreuilh1,2* S. Basu1,2
S. Basu1,2 O. Shaw3
O. Shaw3 H. Burton1,2
H. Burton1,2 N. Mamode1,2
N. Mamode1,2 F. Harris4
F. Harris4 T. Tree4P. Nedyalko5M. Terranova-Barberio5
T. Tree4P. Nedyalko5M. Terranova-Barberio5 G. Lombardi4
G. Lombardi4 C. Scottà4A. Dorling1,2
C. Scottà4A. Dorling1,2
- 1Department of Inflammation Biology, School of Immunology and Microbial Sciences, King’s College London, London, United Kingdom
- 2Centre for Nephrology, Urology and Transplantation, School of Immunology and Microbial Sciences, King’s College London, London, United Kingdom
- 3Synnovis Clinical Transplantation Laboratory, Guy’s Hospital, London, United Kingdom
- 4Peter Gorer Department of Immunobiology, King’s College London, London, United Kingdom
- 5NIHR Guy’s and St Thomas’ Biomedical Research Centre at Guy’s and St Thomas NHS Foundation Trust, St Thomas’ Hospital, London, United Kingdom
Introduction: Highly sensitised (HS) patients represent up to 30% of patients on the kidney transplant waiting list. When they are transplanted, they have a high risk of acute/chronic rejection and long-term allograft loss. Regulatory T cells (Tregs) (CD4+CD25hiCD127lo) are T cells involved in the suppression of immune alloresponses. A particular subset, called T follicular regulatory T cells (Tfr, CXCR5+Bcl-6+), is involved in regulating interactions between T effectors and B cells within the germinal centre and can be found in peripheral blood. Therefore, we wanted to identify specific subsets of Tregs in the peripheral blood of HS individuals.
Methods: We recruited prospectively healthy volunteers (HV) (n = 9), non-sensitised patients on haemodialysis (HD) (n = 9) and HS individuals, all of whom were on haemodialysis (n = 15).
Results: We compared the Treg phenotypes of HV, HD and HS. HS patients had more CD161+ Tregs (p = 0.02) and more CD45RA−CCR7− T effectors (Teffs) (p = 0.04, memory Teffs able to home to the germinal centre) compared to HVs. HS patients had more Bcl-6+ Tregs (p < 0.05), fewer Th1-like Tregs, more Th2-like Tregs (p < 0.001) and more CD161+ (p < 0.05) Tregs compared to HD patients. This population has been described to be highly suppressive. HD had a deficiency in a Th17-like CD161+ effector Treg cluster (cluster iii., CCR6+CCR4+CXCR3− CD39+CD15s+ICOS−CCR7−CD161+) (p < 0.05).
Discussion: This is the first study presenting a deep Treg phenotype in HS patients. We confirmed that HS patients had more of a Th17-like CD161+ effector Treg from population III (CD4+CD25hiCD127loCD45RA−) compared to non-sensitised patients on HD. The clinical relevance of this highly suppressive Tregs population remains to be determined in the context of transplantation.
Introduction
Highly sensitised (HS) patients represent up to 30% of patients on the kidney transplant waiting list. The new UK allocation scheme should be beneficial in prioritising HS patients for a transplant, but partly due to the COVID-19 pandemic and the lack of high-risk transplants being performed, it has not been possible to clearly identify its full impact to date. However, if we extrapolate from what happened in the US with a similar allocation scheme (1), the new UK allocation system may not benefit the most highly sensitised individuals (calculated reaction frequency (crF) 95%–100%), leading to their accumulation on the transplant waiting list.
Finding matched transplants for HS patients is challenging; even with a living donor they usually have to undergo pre-transplant optimisation, including desensitisation, delisting or participating in the UK sharing scheme. When transplanted, even with a well-matched donor, they have a higher risk of both acute and chronic rejection and long-term allograft loss (2). Regulatory T cells (Tregs, CD4+CD25hiCD127lo) are T cells involved in the suppression of immune alloresponses. A particular subset, called T follicular regulatory T cells (Tfr, CXCR5+Bcl-6+), is involved in regulating T effectors and B cell interactions within the germinal centre and can be found in the peripheral blood (3). Other Treg subsets T-bet+ CD45RO− and T-bet− CD45RO+ Tregs have been associated with the presence of donor-specific antibodies (DSA) and DSA+ antibody-mediated rejection (ABMR), respectively (4–6). The aim of the present study was to assess if we could identify specific subsets of Tregs in highly sensitised individuals compared to healthy volunteers (HVs) and individuals on haemodialysis.
Materials and methods
Patients and samples
This study used samples from three cohorts of individuals. Two cohorts were patient recruits from studies initiated and conducted at King’s College London, whereas the third used leucocyte cones from NHS Blood and Transplant (NHSBT). Informed consent was obtained from all participants after the nature and possible consequences of the studies were explained. The first group included sensitised patients (cRF > 50%) enrolled into the “Antibody Incompatible Transplantation - a prospective study” (AIT–IRAS project ID 204733; reference no. 16/WM/0370, West Midlands, Coventry & Warwickshire ethics committee, Research Ethics Committee 22/02/2019). All patients recruited by the first author (CD) between 2019 and 2021 (n = 14) were included. Samples were taken at enrolment and at 3-monthly periods thereafter (maximum number 4). Participants to this study provided written informed consent before inclusion. The second group of patients included haemodialysis controls included from the “Sensitisation develop in Kidney Patients (SIKP) study” (IRA project ID 276643; reference no. 21/SC/0156). Ethical approval was obtained from the South Central–Hampshire B Research Ethics Committee. Patients enrolled in the trial were identified as dialysis patients who had not been sensitised (cRF = 0) and were enrolled by the first author (CD). HV controls were recruited using leucocyte “cones” obtained through a leukoreduction system from anonymised healthy donor peripheral blood obtained from the National Blood Service (NHSBT, Tooting, London, UK) with informed consent and ethical approval (Institutional Review Board of Guy’s Hospital; reference no. 09/H0707/86/).
Isolation of peripheral blood mononuclear cells
All the samples were obtained from local haemodialysis units for SIKP and AIT and from NHSBT for leucocyte cones. Samples were processed using a local SOP by a single operator (CD). The peripheral blood mononuclear cells (PBMCs) were isolated using Lymphoprep separation (Lymphoprep; Stem Cell, Canada) and used fresh [resuspended in AIM V® media (Life Technologies, UK)] with 10% human AB serum (Sigma-Aldrich)–AIMV/AB.
Phenotyping panel
PBMCs were incubated with Near IR LIVE/DEAD (Invitrogen, USA) for 15 min in a cell incubator, at 37°C, containing 5% CO2. After washing, cells were stained with titrated amounts of fluorochrome-conjugated monoclonal-Ab. The Tregs panel consisted of: ICOS (C398.4A), CXCR5 (J252D4, CD25 (BC96), CCR7 (G043H7), CD3 (UCTH1), CCR6 (G034E3), CD69 (FN50), PD-1 (EG12.2H7), Helios (22F6), Ki67 (Ki67) and CD45RO (UCHL-1), all from BioLegend (San Diego, CA, USA); CXCR3 (1C6/CXCR3), CD15s (CSLEX1), CCR4 (1G10), CD4 (SK3), CD45RA (Hi100), CD39 (TU66), Bcl-6 (K112–91), GATA-3 (L50-B23) and CTLA4 (BNI3), all from BD Biosciences (Franklin Lakes, NJ, USA); CD127 (EBioRDR5), FOXP3 (PCH101) and T-bet (4B10), all from ThermoFisher (Waltham, MA, USA); and CD161 (191B8, Miltenyi, Bergisch Gladbach, Germany). The list of all the antibodies, clones, colours and quantities are available in Supplementary Materials Tables S1 and S2. Cells were stained in FACS buffer [as per Stroukov et al. (7)] for 15 min in a cell incubator, at 37°C, containing 5% CO2. After incubation, cells were washed, fixed [1% paraformaldehyde with 99% phosphate-buffered saline (PBS)], then washed with PBS 5% FCS and stored at 4°C before acquisition within 24 h on a BD LSRFortessa flow cytometer (KCL BRC Flow Cytometry Laboratory), using Diva software (BD, Ashland, OR, USA). Samples were then imported in FlowJo (BD) and Cytobank (Beckman Coulter) for unsupervised analyses; CITRUS was run using equal numbers of Tregs (CD4+CD25hiCD127lo) or Teffs (CD4+ non-Tregs), or Tregs events per sample and the following clustering channels: CXCR5, ICOS, CCR7, CXCR3, CD161, CCR6, CD15s, CCR4, CD69, PD-1, CD45RA and CD39 for extracellular markers; and FOXP3, Helios, GATA3, Bcl-6, T-bet, Ki67, CTLA4 and CD45RO for intracellular markers. Samples with a cell viability <60% were not used in the analysis.
Statistical analysis
The statistical analyses were performed using GraphPad Prism. P-values < 0.05 were considered statistically significant. Wilcoxon matched-pairs signed-rank tests were used for non-parametric paired data; Mann–Whitney tests were used for non-parametric unpaired data.
Where more than two groups were analysed together, the non-parametric one-way ANOVA test or Kruskal–Wallis test was employed. The p-values generated through these calculations were recorded. The CITRUS analysis was carried out using an equal sampling method, with a minimum cluster size of 5, five cross-validation folds, one false discovery rate and 315 events per file for Figure 5 and 714 events for Figure 7.
Results
We recruited 15 HS individuals on haemodialysis, nine non-sensitised patients on haemodialysis (HD) and compared them to nine HV leukocyte cones samples.
Patients’ clinical characteristics
The patients’ clinical characteristics are presented in Table 1. In the HD cohort, seven of nine (78%) patients were male and their mean age was 52 ± 9 years. Of them, two (22%) had a background of previous immunotherapy. All were non-sensitised with a cRF of 0 and a mean C-reactive protein (CRP) at the time of blood draw of 9 ± 13.5 mg/L. The mean time on haemodialysis was 2.4 ± 1.4 years. The clinical characteristics of the HS patients are presented in Table 2. The majority were male (9/15, 60%), with a mean age of 49 ± 16 years. Most were from a Black ethnicity (9/15, 60%) and had been on haemodialysis for 5.8 ± 3.3 years. Their mean cRF was 89% and CRP was 4.4 ± 3.7 mg/L. Ten of them had been primarily sensitised through a previous transplant, three through pregnancy and two after transfusion. The transfusion history of eight previously transplanted or pregnant patients was unknown. A comparison of the two groups is presented in Table 3. Patients in the HS group had been on haemodialysis for longer (p = 0.005) and more had a background of previous transplantation (p = 0.002).
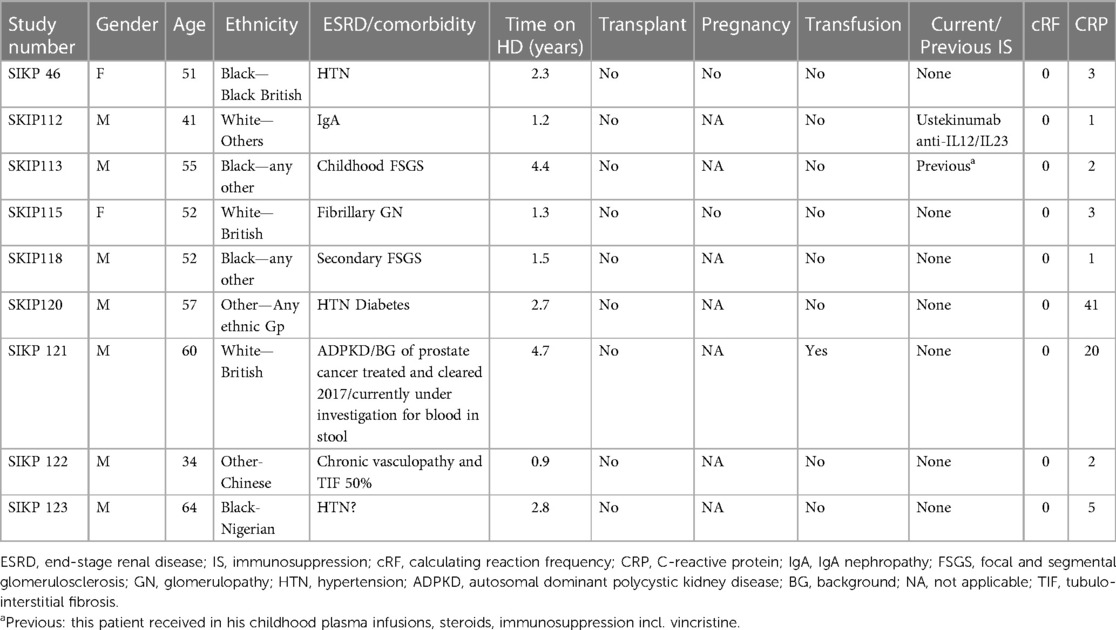
Table 1. HD clinical characteristics.
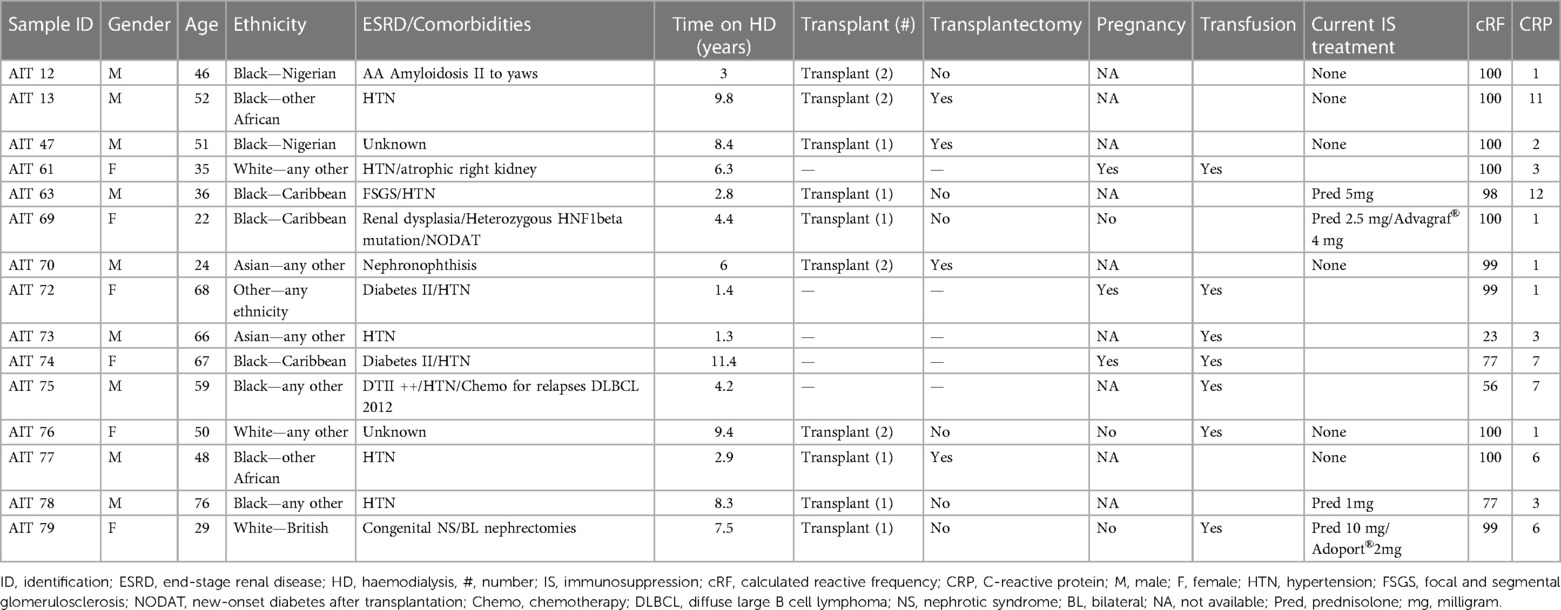
Table 2. HS clinical characteristics.
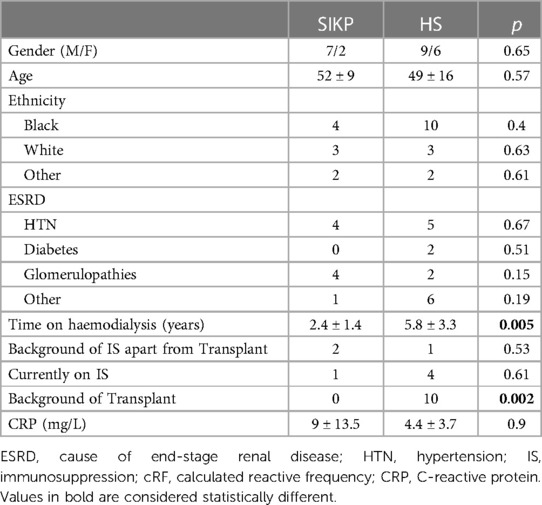
Table 3. Clinical characteristics of SIKP and HS patients.
Human blood T follicular regulatory cells are CXCR5+CCR7+CD45ra+ICOS-PD-1-
The Treg phenotype was studied in nine samples derived from the leucocyte cones of healthy volunteers (the gating strategy is described in Figure 1). The phenotype of blood Tfr (circulating T follicular regulatory cells, called bTfr) has mainly been described in mouse models (3); therefore, we started by identifying the markers expressed by the Tfr population in the blood of healthy individuals. Tfr were defined as Tregs that are CXCR5+ and the markers expressed by them are shown in Figure 2A. The majority of bTfr (defined as ICOS and PD-1-negative CXCR5+ Tregs) looked similar to Tfr and were CCR7+ (able to home to secondary lymphoid organs) and CD45RA+ (naïve) (Figure 2B), supporting the hypothesis that bTfr come from a “truncated” germinal centre response (3). bTfr could be human Tfr that have gone into the T cell zone within the lymph node, have been activated by the dendritic cell, but probably not enough to go through the germinal centre reaction and therefore recirculate into the blood (Supplementary Figure S1).

Figure 1. Gating strategy to identify Th-like subpopulations of Tregs.
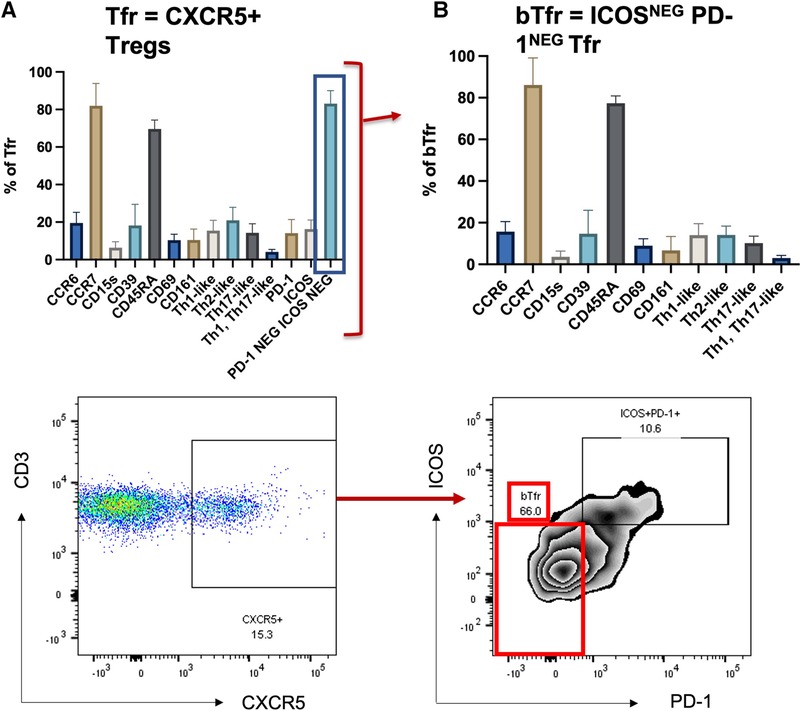
Figure 2. Tfr and bTfr extracellular phenotyping. Description of expression of different markers in the Tfr population (CXCR5+ Tregs) (left panel, A) and on the PD-1neg and ICOSneg (bTfr) subpopulation (right panel, B). The bTf is obtained by selecting PD-1 negative and ICOS negative cells out of the Tfr population.
Highly sensitised patients have more CD161+ Tregs compared to HV and unsensitised HD
We compared HS Treg phenotyping to unsensitised HD patients and healthy volunteers. The analysis was extended to patients who had been sensitised through transfusion and they had a different Treg signature (see Supplementary Material and Figures S2–S4). There was no difference in the proportion of Tregs between the three groups (Supplementary Figure S5). In a biased flow cytometry analysis, unsensitised HD patients had a reduction in Th1-like Tregs (p = 0.0026) and Teffs (p = 0.0085) and an increase in Th2-like Tregs (p = 0.0076) and Teffs (p = 0.005), due to a decrease in CXCR3 expression (p = 0.0030 and p = 0.0079, respectively) (Figure 3A for Tregs and Figure 4A for Teffs). They trended towards a decreased expression of CCR6 (p = 0.0637) and CD15s (p = 0.0644) in Tregs and displayed a trend towards a decrease in the Tfh population (CXCR5+ Teffs), even if non-significant (p = 0.0710).
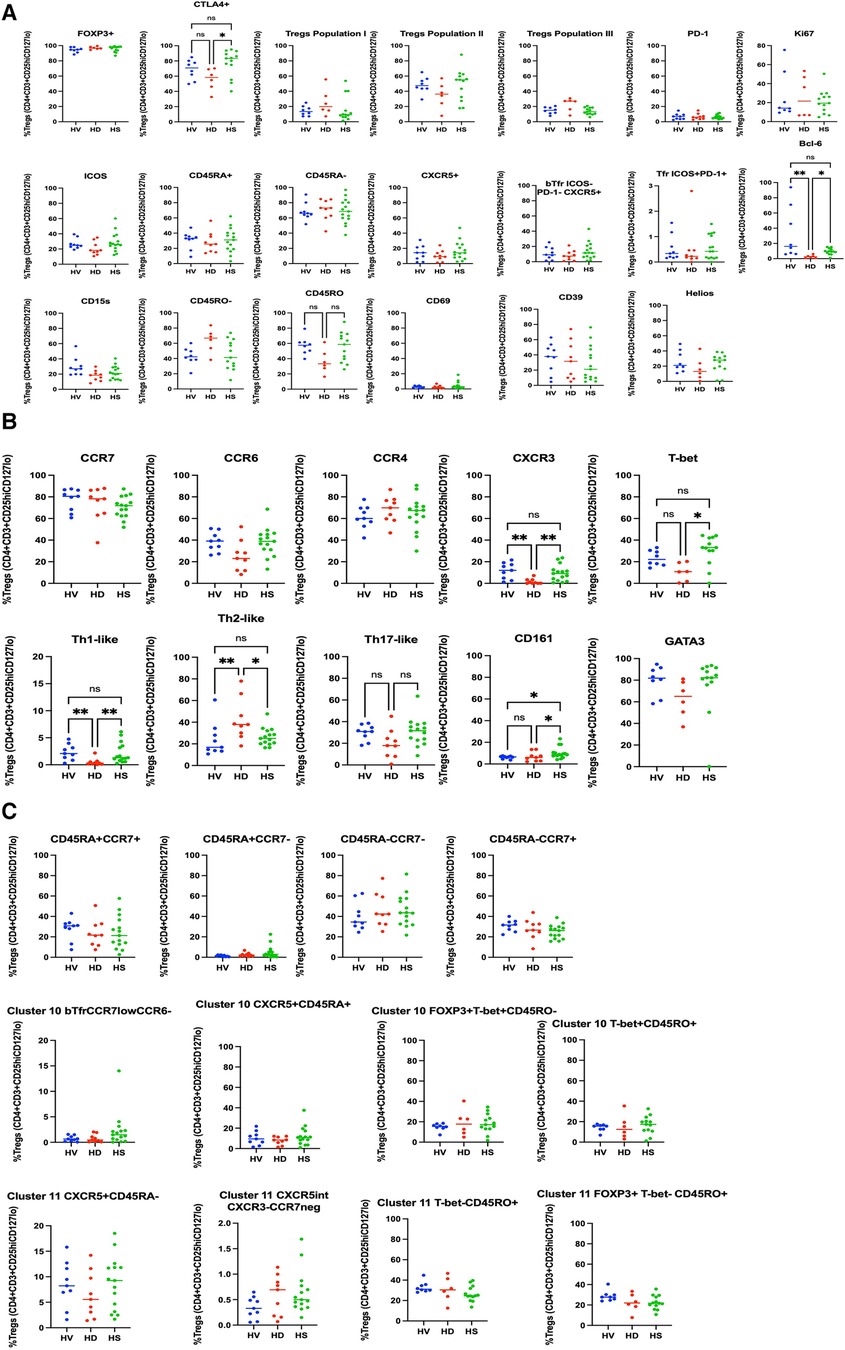
Figure 3. Phenotyping analysis of Tregs in HV, non-sensitised HD, and HS patients. In (A) some common Tregs markers, and the population I, II and III are presented. bTfr were defined as CXCR5+PD-1−ICOS-Tregs. In (B) the markers associated with the Th-like populations are presented. There was no difference in the CD45RA/CCR7 groups (C), but HS patients had a trend towards increase in the CD45RA+CCR7− naïve Tregs population (p = 0.0817).
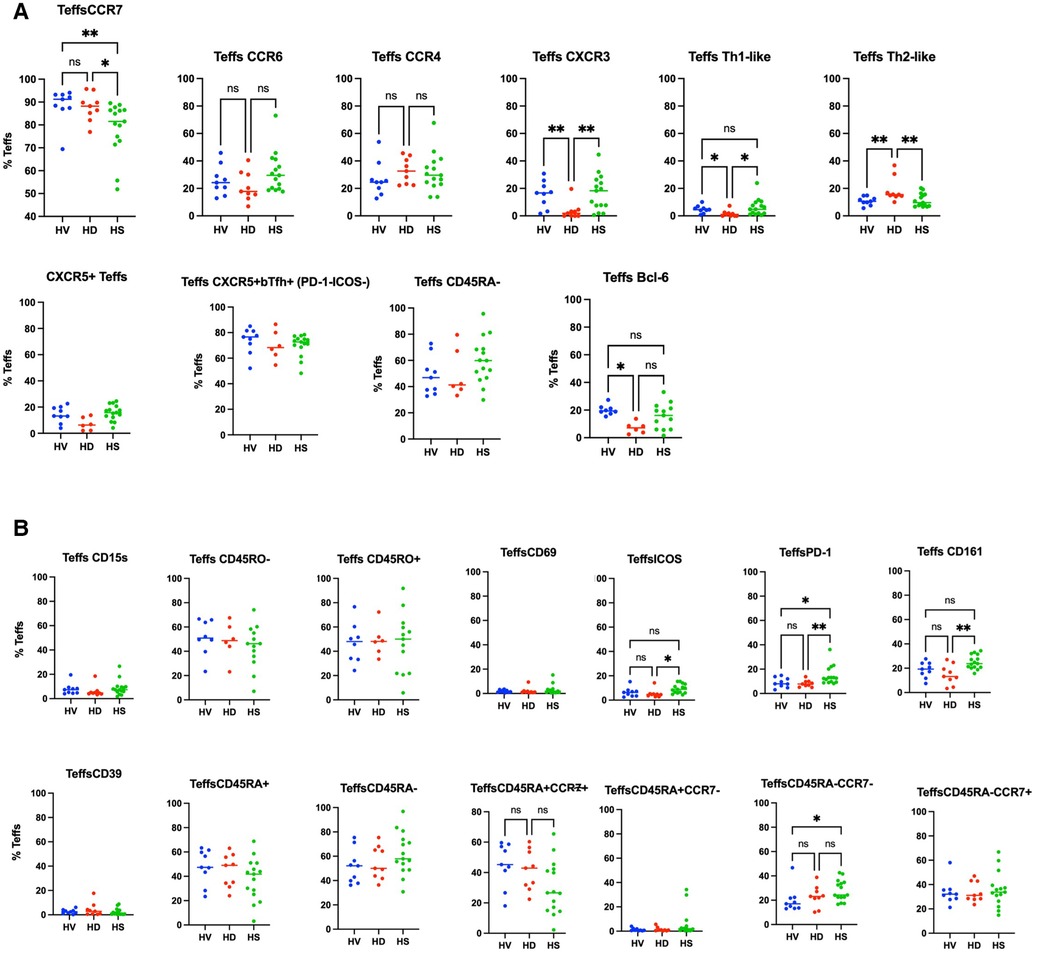
Figure 4. Phenotyping analysis of Teffs in HV, non-sensitised HD, and HS patients. In (A) some common Teffs markers are presented. In (B) the markers associated with the Th-like populations and subpopulations based on the expression of CD45RA and CCR7 are presented. There was no difference in the CD45RA/CCR7 groups (C), but HS patients had a trend towards increase in the CD45RA+CCR7− naïve Tregs population (p = 0.0817).
HS patients had a higher proportion of CD161+ Tregs compared to HV and non-sensitised HD patients (Figure 3A) (p = 0.02). They had a smaller proportion of CCR7+ Tregs compared to HV and HD, and a smaller proportion of Tregs expressing ICOS (p = 0.0138) and PD-1 (p = 0.0112) (Figure 3B). They had a tendency towards an increase in the CD45RA+CCR7−-naïve Tregs population (p = 0.0817). They had a decrease of CCR7+ Teffs (p = 0.009), a lower proportion of CD45RA−CCR7− Teffs compared to HV (p = 0.0409), balanced by a trend to an increase of the CD45RA+CCR7+ Teffs population (p = 0.0553). There was no difference in the expression of the other surface markers, the numbers of bTfr, of ICOS+PD-1+Tfr and the T-bet+CXCR5+CD45RO−ICOS−PD-1− Treg cluster- and T-bet−CXCR5intCD45RO+ICOS−PD-1int cluster, as also described by Louis et al. (4).
HS patients have more of a Th17-like CD161+ Treg cluster (CCR6+CCR4+CXCR3− CD39+CD15s+ICOS−CCR7−CD161+)
Using a CITRUS analysis (available on Cytobank), we compared Tregs from HD, HV and HS patients. HV and HS patients had the same proportion of CD69− clusters (Figure 5) (i) and a Th17-like CD161+ Treg cluster (iii.) compared to unsensitised HD. Unsensitised HD had higher proportion of three different CD69− Tregs clusters (i.). Both unsensitised HD and HS patients had a defect in an ICOS+Th17-like effector Tregs cluster (ii. 9752, ICOS+ CCR7− CCR6+CCR4+CXCR3−CD15s++CD39+) compared to HV (Figure 5). HS patients had a trend towards more of a Th17-like CD161+ Treg cluster (cluster iii., CCR6+CCR4+CXCR3− CD39+CD15s+ICOS−CCR7-CD161+). Interestingly, by using a biased analysis, we were able to identify that HS patients had more of cluster iii compared to unsensitised HD patients (Figure 6). This cluster of Tregs was particularly interesting, as it has been identified as a Th17-like CD161+ Tregs cluster in patients with chronic arthritis (13).
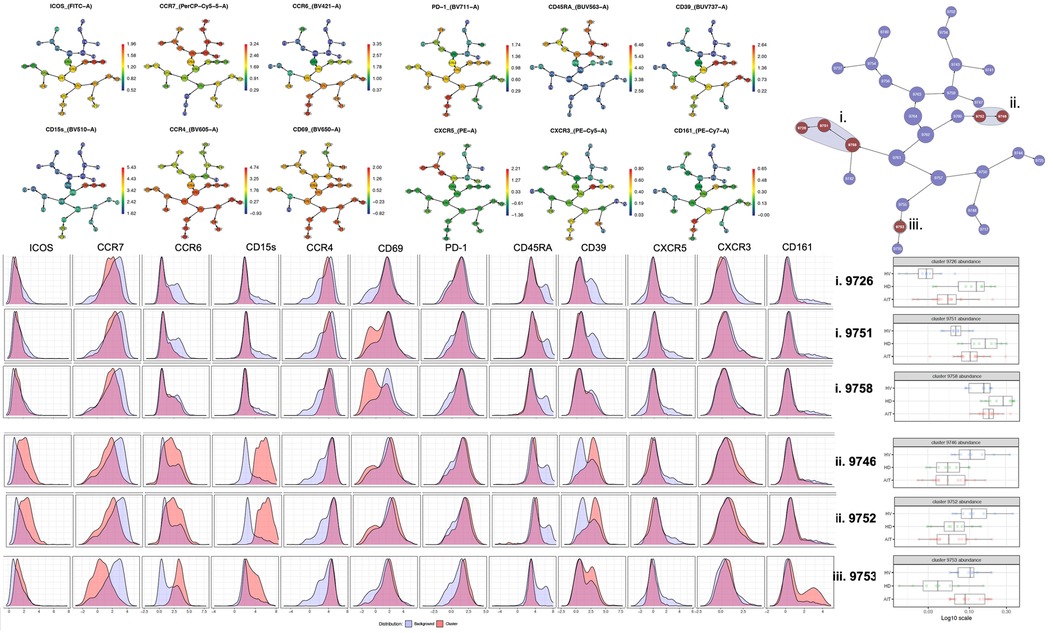
Figure 5. CITRUS analysis of Tregs from extracellular panel comparing HV, unsensitised HD and AIT patients.
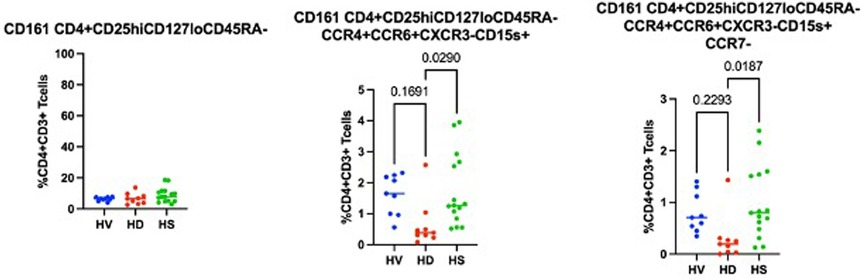
Figure 6. Biased analysis of cluster iii.
Non-sensitised HD patients had more of a T-bet+CD45RO− Tregs population and HS patients had a lower proportion of two T-bet- CD45RO+ Tregs clusters compared to HD patients
In a high-dimensional analysis using CITRUS as a clustering algorithm and focusing on intracellular markers, we found that unsensitised patients on dialysis had a trend towards more of a FOXP3lo T-bet+CD45RO− population (cluster 21621 and 21632) compared to HV and to HS patients (Figure 7). HS patients had a lower proportion of two T-bet− CD45RO+ Treg clusters (21660 and 21667) compared to non-sensitised HD patients (p = 0.01 and p = 0.03, respectively). These two groups of clusters were of particular interest as they look very close to the ones identified by Louis et al. (4), which were associated with the presence of DSA only (T-bet+ CD45RO− Tregs) and DSA/ABMR (T-bet− CD45RO+ Tregs).
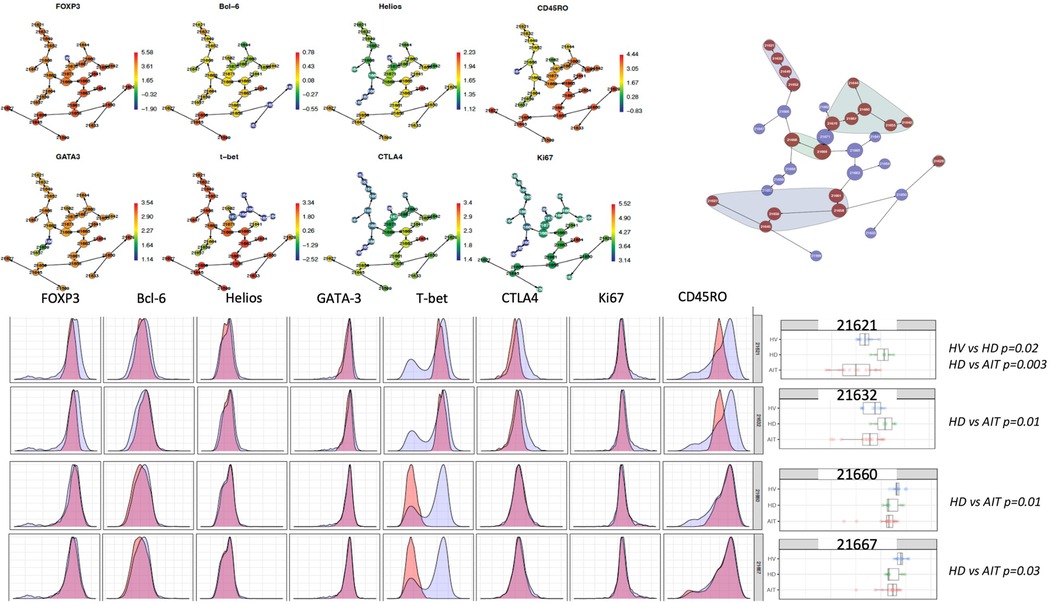
Figure 7. CITRUS analysis of intracellular panel.
Discussion
This is the first study presenting a deep Treg phenotype in HS patients. We demonstrated that they had more of a Th17-like CD161+ effector Tregs from population III (CD4+CD25hiCD127loCD45RA−) (cluster iii) compared to unsensitised HD patients.
We described for the first time the presence of Tfr in the blood (called bTfr) in non-sensitised HD and AIT patients and this study is one of the first presenting the presence of CXCR5+ Tregs circulating Tregs in HV (4). The identification of bTfr has come mainly from mouse models (8–11) and there are only a few studies that looked at bTfr specifically in HV (3). We confirmed that bTfr are ICOS−PD-1−, and express a high level of CCR7 and a high level of CD45RA. This supports the hypothesis of the “truncated germinal centre response,” where bTfr could be naïve Tfr that may have interacted with the DC within the T cell zone, but without being activated and would circulate back in the blood [see (3) and Figure 2].
The different groups of patients described have some common characteristics. The HS and unsensitised HD groups are not different for most of the controllable characteristics: age, sex, ethnicity, inflammatory status and cause of end-stage renal disease (ESRD). There was a tendency towards the male sex in the unsensitised HD group (7/9) as female patients are more likely to be sensitised through pregnancies on the top of the risk of being sensitised through blood transfusions. They were different in the cRF, but this was one of the inclusion/exclusion criteria; therefore, some differences were to be expected. In addition, there was a difference in the length of time on dialysis between the two groups. Patients in the SIKP group were on dialysis for a shorter period (2.4 ± 1.4 years) compared to patients in the HS group (5.8 ± 3.3 years) (p = 0.005). This introduces a bias in the way we can interpret the phenotyping data; however, there was no way to mitigate this bias. Patients with low cRF are much easier to transplant and therefore do not stay on dialysis; therefore, it was more difficult to find patients on the waiting list who had been there as long as the HS patients.
The type of sensitisation and outcomes after transplantation have not been described extensively. One review by Scornik et al. (12) discussed the impact of different routes of sensitisation and the risk of immunisation. They clearly stratified the immunogenicity from low to high immunogenicity as follow: Recent transfusion < Pregnancy alone < Multiple transfusion = Pregnancy + transfusion = Transplantation < Transplantation + Transfusion. In our study, patients who had been sensitised through transfusion had a higher number of ICOS+ Tregs (p = 0.049), CCR4+ Tregs (p = 0.0452), a lower proportion of CCR7+ Tregs (p = 0.0413), and a trend towards more population II Tregs (p = 0.0574) and Th17-like Tregs (p = 0.0631). This means that patients who had been sensitised through blood transfusion only (AIT 73 and AIT 75) had more Th17-like effector Tregs able to home towards the lymphoid organs. This may relate to the fact that patients who have been immunised by blood transfusion have less immunogenicity. These results need to be validated in a bigger cohort.
In the unbiased phenotyping analysis, our initial hypothesis was that non-sensitised patients on HD would have a different phenotype compared to HV, and that HS patients would have another signature phenotype compared to both HV and non-sensitised HD patients. However, first, we demonstrated that non-sensitised HD had fewer Bcl6+ Tregs and Teffs, and fewer Th1-like Tregs and Teffs, compared to healthy volunteers. We did not find any difference in the numbers of Tregs as a proportion of CD4+ cells. This is different from the previous literature describing either increases in the number of patients on HD (13, 14), or a decrease in the number (15), but is consistent with the work performed by Afzali et al. (16). Afzali et al. compared the Tregs phenotype in non-sensitised HD patients and healthy volunteers. They identified that Tregs from patients on HD had fewer Tregs from population II, that age did not impact the number of Tregs in this population, and there were no differences in expression of CD39, FOXP3, HLADR and CD27 by Tregs in HD and HV.
This study is, to the best of our knowledge, the first to phenotype Tregs in HS patients. When recruiting patients, we recruited them on a minimum immunosuppression to reduce as much as possible the impact of treatment on the T cell populations present. Interestingly, in the biased analysis, HS patients had more CD161 Tregs, fewer CCR7− Teffs, more CD45RA-CCR7− Teffs (memory Teffs capable to home to the germinal centre) and more PD-1 Teffs. Afzali et al. (16) demonstrated that Tregs from patients on haemodialysis had a lower proliferation rate and a decreased suppressive ability, which recovered after culture using IL2 and Rapamycin. While the functional capacities of Tregs were not studied in this manuscript, based on this previous published work, expanding Tregs using Rapamycin could be promising, particularly if it could be confirmed that the expansion process is reducing the number of cells producing IL-17, as demonstrated by Afzali et al. More interestingly, an unbiased analysis using CITRUS (Figure 7) confirmed that HS patients had more of a CD161+ effector Tregs from population III (CD4+CD25hiCD127loCD45RA−) (cluster iii) compared to unsensitised HD. This is particularly relevant as our group identified this specific cluster present in the inflamed joints of patients with inflammatory arthritis (17) and it could be a subset of Tregs involved in inflammation associated with sensitisation. Moreover, our group demonstrated that this specific subset of Tregs was highly suppressive (18) and produce cytokines favouring wound healing. HS patients had a lower proportion of two T-bet- CD45RO+ Tregs clusters compared to non-sensitised HD patients (p = 0.01 and p = 0.03, respectively). Recently, one study compared the Tregs phenotype between HV, patients with ESRD but not on HD and transplant patients (KTx group) (13). A major difference between these groups was that patients in the transplant group had functioning transplants and were on immunosuppression. In that study, they demonstrated that KTx patients showed increased frequencies of naïve and effector memory natural Tregs (nTregs), whereas central memory nTregs were reduced. Patients with ESRD had nTregs (CD4+CD25hiFOXP3+) that expressed higher levels of CD127 but lacked CD154 expression upon activation. However, we found no differences in this population in our analyses, demonstrating the complexity of the different groups.
This is the first study presenting a deep Treg phenotype in highly sensitised patients. We confirmed that HS patients had more of a Th17-like CD161+ effector Tregs from population III (CD4+CD25hiCD127loCD45RA−) compared to unsensitised HD patients. This is of particular interest for future areas of research, including strategies to restore a less pro-inflammatory phenotype of Tregs.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The studies involving humans were approved by AIT: reference no. 16/WM/0370, West Midlands, Coventry and Warwickshire Ethics Committee, Research Ethics Committee 22/02/2019; and SIKP: reference no. 21/SC/0156. Ethical approval was obtained from the South Central—Hampshire B Research Ethics Committee. The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study.
Author contributions
AD is the last author of this manuscript. He developed the initial idea and background behind this publication, obtained funding, supervised CD and edited the manuscript. CD wrote the paper, recruited patients, and performed the assays and analysis. SB helped recruit and process the samples. OS helped identify patients of interest to recruit. HB helped teach laboratory techniques, recruit patients and with the data analysis. MT-B and PN helped with the high-dimensional analysis. FH and TT helped design and troubleshoot the Tregs panel. NM helped recruit patients and was the principal investigator of the AIT study. GL developed the initial idea with AD and has supervised CD through her PhD. CS is the second supervisor of CD and has helped design the phenotyping data and suppression assays. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by the Medical Research Council (award number MR/S000852/1).
Acknowledgments
The authors acknowledge that the research is supported by the National Institute for Health Research (NIHR) Biomedical Research Centre based at Guy’s and St Thomas’ NHS Foundation Trust and King’s College London. The views expressed are those of the authors and not necessarily those of the NHS, the NIHR or the Department of Health.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors CD and GL declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/frtra.2023.1165320/full#supplementary-material
References
1. Schinstock CA, Smith BH, Montgomery RA, Jordan SC, Bentall AJ, Mai M, et al. Managing highly sensitized renal transplant candidates in the era of kidney paired donation and the new kidney allocation system: is there still a role for desensitization? Clin Transplant. (2019) 33(12):e13751. doi: 10.1111/ctr.13751
2. Manook M, Koeser L, Ahmed Z, Robb M, Johnson R, Shaw O, et al. Post-listing survival for highly sensitised patients on the UK kidney transplant waiting list: a matched cohort analysis. Lancet. (2017) 389(10070):727–34. doi: 10.1016/S0140-6736(16)31595-1
3. Dudreuilh C, Basu S, Scottà C, Dorling A, Lombardi G. Potential application of T-follicular regulatory cell therapy in transplantation. Front Immunol. (2021) 11:612848. doi: 10.3389/fimmu.2020.612848
4. Louis K, Fadakar P, Macedo C, Yamada M, Lucas M, Gu X, et al. Concomitant loss of regulatory T and B cells is a distinguishing immune feature of antibody-mediated rejection in kidney transplantation. Kidney Int. (2022) 101(5):1003–16. doi: 10.1016/j.kint.2021.12.027
5. Segundo DS, Millán O, Muñoz-Cacho P, Boix F, Paz-Artal E, Talayero P, et al. High proportion of pretransplantation activated regulatory T cells (CD4+CD25highCD62l+CD45RO+) predicts acute rejection in kidney transplantation: results of a multicenter study. Transplantation. (2014) 98(11):1213–8. doi: 10.1097/TP.0000000000000202
6. Boix F, Trujillo C, Muro M. Cell-mediated immunity (CMI) as the instrument to assess the response against the allograft: present and future. Curr Protein Pept Sci. (2018) 19(11):1092–106. doi: 10.2174/1389203719666180730164542
7. Stroukow Q, Mastronicola D, Albany CJ, Catak Z, Lombardi G, Scotta C. OMIP-090: a 20-parameter flow cytometry panel for rapid analysis of cell diversity and homing capacity in human conventional and regulatory T cells. Cytometry A. (2023) 103(5):362–7. doi: 10.1002/cyto.a.24720
8. Linterman MA, Pierson W, Lee SK, Kallies A, Kawamoto S, Rayner TF, et al. Foxp3+follicular regulatory T cells control T follicular helper cells and the germinal center response. Nat Med. (2011) 17(8):975–82. doi: 10.1038/nm.2425
9. Chung Y, Tanaka S, Chu F, Nurieva RI, Martinez GJ, Rawal S, et al. Follicular regulatory T cells expressing Foxp3 and Bcl-6 suppress germinal center reactions. Nat Med. (2011) 17(8):983–8. doi: 10.1038/nm.2426
10. Aloulou M, Carr EJ, Gador M, Bignon A, Liblau RS, Fazilleau N, et al. Follicular regulatory T cells can be specific for the immunizing antigen and derive from naive T cells. Nat Commun. (2016) 7(1):10579. doi: 10.1038/ncomms10579
11. Maceiras AR, Almeida SCP, Mariotti-Ferrandiz E, Chaara W, Jebbawi F, Six A, et al. T follicular helper and T follicular regulatory cells have different TCR specificity. Nat Commun. (2017) 8:15067. doi: 10.1038/ncomms15067
12. Scornik JC, Meier-Kriesche HU. Blood transfusions in organ transplant patients: mechanisms of sensitization and implications for prevention. Am J Transplant. (2011) 11(9):1785–91. doi: 10.1111/j.1600-6143.2011.03705.x
13. Landwehr-Kenzel S, Zobel A, Hoffmann H, Landwehr N, Schmueck-Henneresse M, Schachtner T, et al. Ex vivo expanded natural regulatory T cells from patients with end-stage renal disease or kidney transplantation are useful for autologous cell therapy. Kidney Int. (2018) 93(6):1452–64. doi: 10.1016/j.kint.2018.01.021
14. Carmona-Escamilla MA, Queipo G, García-Mosqueda LA, García-Covarrubias L, Fonseca-Sánchez MA, Villanueva-Ortega E, et al. Peripheral blood regulatory T cells are diminished in kidney transplant patients with chronic allograft nephropathy. Transplant Proc. (2018) 50(2):444–8. doi: 10.1016/j.transproceed.2018.01.001
15. Hendrikx TK, van Gurp EAFJ, Mol WM, Schoordijk W, Sewgobind VDKD, IJzermans JNM, et al. End-stage renal failure and regulatory activities of CD4+CD25bright+FoxP3+ T-cells. Nephrol Dial Transplant. (2009) 24(6):1969–78. doi: 10.1093/ndt/gfp005
16. Afzali B, Edozie FC, Fazekasova H, Scottà C, Mitchell PJ, Canavan JB, et al. Comparison of regulatory T cells in hemodialysis patients and healthy controls: implications for cell therapy in transplantation. CJASN. (2013) 8(8):1396–405. doi: 10.2215/CJN.12931212
17. Afzali B, Mitchell PJ, Edozie FC, Povoleri GA, Dowson SE, Demandt L, et al. CD161 expression characterizes a subpopulation of human regulatory T cells that produces IL-17 in a STAT3-dependent manner. Eur J Immunol. (2013) 43(8):2043–54. doi: 10.1002/eji.201243296
Keywords: highly sensitised, regulatory T cells, haemodialysis, T follicular regulatory cells, sensitisation
Citation: Dudreuilh C, Basu S, Shaw O, Burton H, Mamode N, Harris F, Tree T, Nedyalko P, Terranova-Barberio M, Lombardi G, Scottà C and Dorling A (2023) Highly sensitised individuals present a distinct Treg signature compared to unsensitised individuals on haemodialysis. Front. Transplant. 2:1165320. doi: 10.3389/frtra.2023.1165320
Received: 13 February 2023; Accepted: 20 September 2023;
Published: 30 October 2023.
Edited by:
Fadi Issa, University of Oxford, United KingdomReviewed by:
Manuel Muro, Virgen de la Arrixaca University Hospital, SpainByoung Chol Oh, Johns Hopkins University, United States
Thiago J. Borges, Massachusetts General Hospital and Harvard Medical School, United States
© 2023 Dudreuilh, Basu, Shaw, Burton, Mamode, Harris, Tree, Nedyalko, Terranova-Barberio, Lombardi, Scottà and Dorling. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: C. Dudreuilh Y2Fyb2xpbmUuZHVkcmV1aWxoQGtjbC5hYy51aw==