 Vera W. Pfeiffer
Vera W. Pfeiffer Jun Zhu3
Jun Zhu3 Karen Poh
Karen Poh- 1Nelson Institute for Environmental Studies, University of Wisconsin–Madison, Madison, WI, United States
- 2Department of Statistics, University of Wisconsin–Madison, Madison, WI, United States
- 3Animal Disease Research Unit, United States Department of Agriculture-Agricultural Research Service (USDA-ARS), Pullman, WA, United States
- 4Department of Planning and Landscape Architecture, University of Wisconsin-Madison, Madison, WI, United States
- 5Silvernail Studio for Geodesign, LLC, Black Earth, WI, United States
Introduction: Urbanization diminishes the extent of uncompacted, exposed soil and the coverage of wild plant species, yet still supports diverse pollinator communities when their habitat is maintained within the built environment. Floral abundance and richness are known to influence bee foraging behavior, and these factors vary across the landscape, especially across heterogeneous urban extents. This study assesses how floral resources are distributed across the city of Madison, WI and how these factors influence the distribution of foraging bumble bees.
Methods: We conducted a systematic walking transect survey of bumble bees across Madison, WI. The resulting point location data associated with more than 5,000 non-lethally surveyed bumble bees were analyzed with regard to floral resource explanatory variables as well as underlying land use zoning on more than 700 transects. We used Moran's I correlograms to investigate spatial autocorrelation in floral resource variables and bumble bee counts, then we fitted a generalized linear model predicting transect bee counts based on floral cover, density, species richness and wild plant species richness on the distribution of foraging bees. We employed a geographically-weighted regression model to explore non-stationarity in the effects of floral resource explanatory variables across the study extent.
Results: We found significant positive influence of flower cover, species richness, and weakly significant positive influence of wildflower species richness on foraging bee counts within the model as well as a significant positive influence of the land use zoning categorical variable. The effects of floral resource predictors on foraging bumble bees varies based on landscape context across the city.
Discussion: The results of this study show that landscapes with high cover as well as floral diversity maximize bumble bee foraging, and the positive effect of wildflower species richness stands out where floral cover and overall richness are also present. Given that urban landscapes are not homogeneous and that floral resources are not consistently distributed across the cityscape, valuable pollinator habitat should be protected, and supplemented where gaps persist.
Introduction
Pollination is an essential ecosystem service that enables the production of fruits and seeds and maintains the diversity of most plant populations in almost all global ecosystems (Klein et al., 2007; Ollerton et al., 2011; Garibaldi et al., 2013). However, global declines of many pollinators have triggered international interest in conservation science to address risk factors and isolate the most influential variables that promote bee community resilience (Goulson et al., 2008; Tuell et al., 2008; Garbuzov and Ratnieks, 2014). Floral abundance and floral species richness are known factors that influence bumble bee colony growth and foraging activity (Jha and Kremen, 2013; Crone and Williams, 2016). These resources have been assumed to decrease in urban ecosystems compared to forb-rich, semi-natural land cover (e.g., prairie), yet recent work has revealed that urban ecosystems likely provide intermediate amounts of floral resources that surpass other types of natural land cover (e.g., forb-poor grassland) (Goddard et al., 2010; Threlfall et al., 2015; Lynch et al., 2021). Comprehensive investigation of the distribution of floral resources and foraging bumble bees across an urban extent may illuminate which aspects of floral resource distribution shape bumble bee foraging patterns most across an urban ecosystem.
Foraging for food is an energy intensive process that must be continuously accomplished to sustain bumble bee colonies through the growing season (Dornhaus and Chittka, 2005). Bumble bees do not store much surplus food within the colonies, so constant, efficient foraging on nutritionally appropriate pollen sources is a necessity (Vaudo et al., 2018). Bumble bees are central place foragers, known to fly long distances to visit resource rich floral assemblages (Redhead et al., 2016) and landscape scale floral resources have been shown to influence bumble bee foraging distances in studies based in multiple landscapes (Jha and Kremen, 2013; Pope and Jha, 2018). Bumble bee activities and resource use are difficult to assess at broad landscape scales and are known to differ depending on the composition of the landscape (Hemberger and Gratton, 2018; Pfeiffer et al., 2019; Rollings and Goulson, 2019). Spiesman et al. (2016) found no influence of landscape scale floral resources on bumble bee colony growth, when local dominance of resource-rich flowers was high.
Landscape-scale studies have been conducted to confirm the importance of floral cover and native plant species richness to sustain bee communities, and further consideration of the distribution of these factors may help to illuminate the degree of their influence in heterogeneous urban ecosystems (Williams et al., 2012; Requier et al., 2020; O'Connell et al., 2021). Many studies focus on components of urban landscapes in isolation, such as city parks or residential gardens, rather than the comprehensive urban landscape (Bhattacharya et al., 2003; McFrederick and LeBuhn, 2006; Osborne et al., 2008). Developing an understanding of the distribution of important foraging resources and their use by foraging bees across complex spatial extents is important to improve the capacity of landscapes to sustain robust pollinator populations (Pywell et al., 2006; Brosi et al., 2008; Goulson et al., 2010). Comparing the influence of various explanatory factors on the distribution of bumble bee foragers can help to tune conservation guidelines to the context of a particular ecotype (Galpern et al., 2012).
This study investigates the spatial distribution of floral resources and foraging bumble bees around the city of Madison, and tests the predictive capacity of four aspects of floral resource distribution to estimate the distribution of foraging bumble bees. We hypothesize that (1) floral resources and bumble bee abundance are not uniformly distributed across the urban extent, and (2) floral cover, density, species richness, and wildflower species richness increase bumble bee foraging activity. Additionally, (3) we expect that the importance of the various aspects of floral cover fluctuate across the city based on relevance to local limiting factors. Specifically, we expect that floral community composition will be most important where there is sufficient floral cover or density. We explore patterns of non-stationarity in the effect size of floral resource explanatory variables across the study extent using geographically weighted regression. The results of this study identify what aspects of floral resource distribution confer the strongest influence on observed frequency of bumble bee foragers, informing conservation practitioners based on local context.
Materials and methods
Study area
We conducted a systematic transect survey across a 125 km2 extent covering the city of Madison, WI, USA to map the distribution of floral resources and foraging bumble bees in July and August of 2013 and 2014. The survey extent included the city of Madison, located on an isthmus bounded by a northern and southern lake, as well as the exurban agricultural land immediately adjacent to the eastern and western edges of the city. Several high density urbanized patches of mixed commercial/residential zones were located within both eastern and western residential zones, as well as the center of the grid (downtown Madison). Agricultural land was primarily located on the eastern and western edges of the city, across an approximately 6 × 3 km of surveyed terrain on each edge. The majority of the surveyed terrain was comprised by residential land use, characterized by a moderate range of impervious surface (~20–30% based on 1 m resolution classification), and grass lawn, with scattered patches of woodland and semi-natural grassland or restored prairie scattered throughout the extent.
Transects
An ~400 m transect was surveyed across each publicly accessible 400 × 400 m cell of the 125 km2 grid, by walking a maximally straight line from one side of the grid cell to another between 9 am and 5 pm, in July and August of 2013 and 2014, the time of year when bumblebees are most prolific. Due to extremely-restricted accessibility of the urban landscape based on the layout of residential property, most transects were located along one side of a street, and half the transect area was comprised by impervious surface. Frequently, large parking lots also boosted the amount of impervious surface covered by the transect path. After the maximally straight, approximately 400 m, publicly-accessible path from one side of the cell to another side of the cell was selected, an observer walked the path across the cell as a 10 m wide transect, at which time the observer recorded a list of flowering plants in anthesis, and recorded GPS points marking the occurrences of bumble bees. The mean and median transect lengths was 389 m and 409 m, respectively, while the minimum and maximum transects were 210 m and 664 m. When bumble bees were observed within 5 m of the observer, GPS coordinates were collected and their species was recorded along with the plant morphospecies they were visiting in most cases (Williams et al., 2014). Most native plants were identified to species, however many ornamental plants and rare plants were identified by morphospecies alone (i.e., >90 percent of floral ids were at species level).
Explanatory variables
The floral resource explanatory variables estimated for each transect included percent cover, density, flowering plant species richness, and prairie and lawn species richness. During the transect walk, the observer estimated the total percentage area of the transect that could support vegetation (non-impervious), as well as the average density of flowers within that area of non-impervious cover. After completion of the data collection, all flower morphospecies were classified as lawn, prairie, or garden species based on the ecotype where they were most often observed. The lawn species included “weedy” morphospecies that commonly persisted in areas with mowed lawn, including white clover (Trifolium repens), red clover (Trifolium pretense), birds-foot trefoil (Lotus corniculatus), crown vetch (Secuigera varia), dandelion (Taraxacum officinale), heal-all (Prunella vulgaris), sweet clover (Melilotus spp.), plantains (Plantago sp.), and chickweed (Cerastium spp.). Prairie species included primarily native, perennial species commonly found in restored prairie ecotypes (as well as gardens), but whether spontaneous or cultivated, require reduced mowing to produce flowers such as Joe-pye weed (Eutrochium purpureum), Boneset (Eupatorium perfoliatum), Cone flower (Echinacea purpurea), milkweeds (Asclepias spp.), rosinweed (Silphium integrifolium), cup plant (Silphium perfoliatum), vervain (Verbena spp.), Queen Anne's lace (Daucus carota) and others. Most other species were classified as garden species found primarily in managed gardens and dominated by cultivars.
Mapped transects were compared with city parcel spatial data layers, and each transect was assigned a categorical land use zoning type, loosely based on city zoning of the parcels where the transect was surveyed. Commercial use was prioritized, so that mixed residential and commercial zones were coded as commercial. Residential zones were the most common land use category. Transects that passed through city parks were coded as city parks. Agricultural zoning qualifications in the city parcel data were much less accurately linked to observed land use, so agricultural land use zoning was determined based on observation during the transect survey.
Statistical analysis
To test our expectation that floral resource descriptors and bumble bee counts were not uniformly distributed (hypothesis 1), and spatial autocorrelated at local scales – meaning nearest neighbor transects or nearest few transects, or nearest 50 transects were more similar than expected based on the distribution of values across the whole dataset – we investigated the scale of spatial autocorrelation in each of these variables using correlograms. Correlograms estimate Moran's I correlation metrics between classes of points separated by increasing spatial lags to illustrate the scales at which either positive or negative spatial autocorrelation is observed and where it dissipates. Signals at the most distant lags should be ignored because they are based on very few comparisons, because they are calculated only based on the most distant points.
The number of bumble bees foraging on each transect was modeled using a generalized linear regression model to estimate the effect of floral resource explanatory variables and land use zoning on bumble bee frequency to test hypotheses 2. The variance in count data was greater than the mean, indicating a pattern of over-dispersion that breaks the assumptions associated with the Poisson distribution. Both negative binomial and quasi-poisson model fitting were applied, resulting in similar patterns of variable strength and significance. The robust quasi-poisson fitting was used for the Poisson model to accommodate overdispersion commonly observed in count datasets. This fitting adjusts the standard error with an appropriate scalar, although the coefficient estimates remain the same as the standard Poisson model. Interactions between land use type and each floral resource variable were included to test hypothesis 3. Before fitting the model, the variance inflation factor of the explanatory variables was calculated to ensure that multicollinearity would not overestimate the variance explained by the model. All VIF scores fell below 2, well below commonly used cut-off values of 5 or 10.
The Moran's I test of residuals based on spatially weighted 10 nearest neighbor points was used to check for spatial structure in the residuals of the model, and the alternative hypothesis that existing spatial structure in the dataset was not captured in the model. However, the test statistic ranging between −1 and 1, was calculated to be 0.062 (p < 0.001), very close to zero indicating only a very small proportion of the variance in bee frequency was explained by spatial structure with a ten nearest point (k = 10) neighborhood covering an approximately 1.5 km radius neighborhood. Similarly small spatial structure was observed using a 25 or 100 point neighborhood, with Moran's I test statistics estimated as 0.016 p = 0.037 or 0.013 p = 0.001. Robust standard error was calculated separately for each coefficient estimate in the glm model using the “sandwich” package in R (R Core Team, 2013; Zeileis et al., 2018). To account for the spatially autocorrelated variance, the robust standard error for heteroscedasticity was used (ie. vcovHC) with White's estimator for large sample sizes (i.e., “HC0”). This method is tailored to take into account leverage points.
In the final component of the study, we employed a local quasi-Poisson model fitted by the geographically-weighted quasi-Poisson regression method to investigate non-stationarity in floral resource coefficient estimates across the study extent (Kalogirou, 2018). This exploratory statistical method is a type of local regression whereby generalized linear regression models are fit across the study extent based on a moving window, and regression coefficients are calculated for each data point. This analysis was accomplished using the “lctools” R package. Local regression coefficients were calculated based on an adaptive geographic window including the 50 closest points and each included data point is weighted by geographic distance.
Results
Overall, 784 transect were surveyed, including 408 residential transects, 222 commercial transects, 77 agricultural transects, and 77 transects through city parks. Across the transects 5,574 foraging bumble bees were documented, primarily comprised by Bombus impatiens, B. bimaculatus, B. vagans, B. griseocollis, B. rufocinctus, B. fervidus, B. auricomus, also, B. perplexus, B. sandersoni, B. ternarius, B. terricola, and B. affinis. Floral cover, density, species richness, as well as the number of prairie and lawn species were documented, and while the range of each of these variables was similar across each of the land use zoning types, the distribution of values for several of these explanatory variables differed between land cover zoning categories (i.e., agricultural, commercial, residential, and park) (Figure 1). Transects in agricultural zoned areas and city parks were characterized by a more uniform distribution of flower cover compared to commercial and residential transects which were more frequently characterized by lower flower cover and higher impervious surface than agricultural and park transects. Commercial and residential transects included more low and high values of species richness, while agricultural and city park transects included more high values of lawn and prairie species compared to commercial and residential transects.
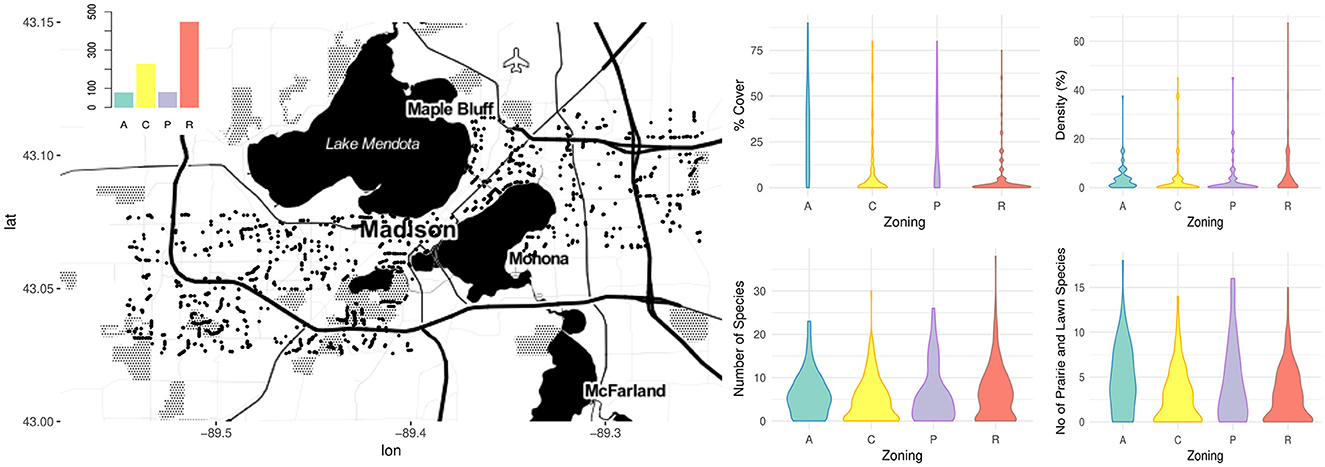
Figure 1. Map of the study extent and point locations of bumble bees observed along the transects. The upper left bar chart shows the number of transects sampled of each land use category (A – Agricultural, C – Commercial, P – Park, R – Residential), while the fiddle plots on the right show the distribution of flower resource explanatory variables by land use categorical variable zoning levels. Basemap data copyrighted OpenStreetMap contributors and available from https://www.openstreetmap.org.
Floral resources were more spatially autocorrelated than bees
Positive spatial autocorrelation was observed in bee count data up to about 5 km, and in floral resource variables up to about 2 or 3 km (Figure 2). Maximum Moran's I estimates of bee count data at very short distances reached 0.1 (Figure 2A), while floral resource variables reached a higher maximum of just over 0.15 (Figures 2B–D), which indicate that only a small proportion of the variation could be attributed to local spatial autocorrelation.
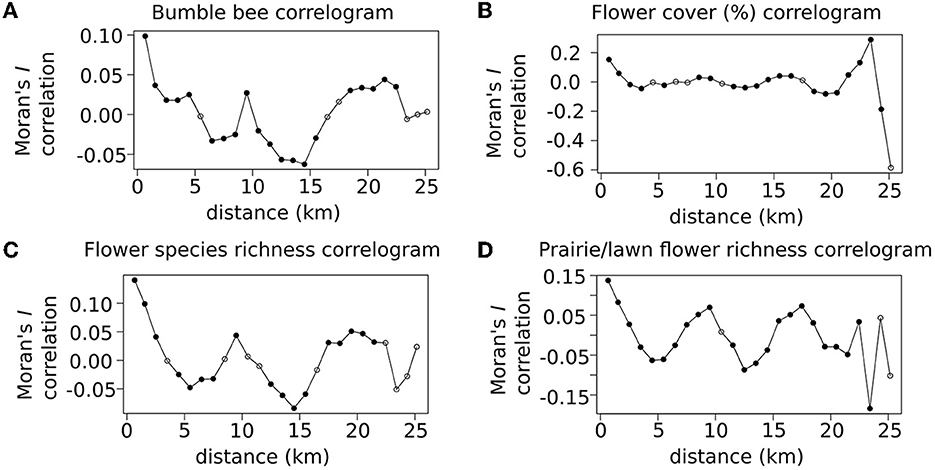
Figure 2. Correlograms depict the spatial structure of variance across scales within the study extent for (A) bee counts, as well as each floral resource variable, (B) floral cover, (C) floral species richness, and (D) Prairie/lawn flower species richness. Correlograms plot the Moran's I correlation statistic. Dark points in the correlogram indicate a statistically significant Moran's I value indicating positive or negative spatial autocorrelation between transects separated by the spatial lag value on the x-axis compared to the rest of the transects in the dataset.
The most bee-attractive flower species were prairie and lawn species
Fourteen flower species each supported more than 1% of the bumble bees observed in the survey data (accounting for over 55 observations). These “most visited species” accounted for ~70% of all bumble bee observations documented in our survey. These top visited species included, goldenrod (Solidago spp.) (12.4% of observations), white clover (Trifolium repens) (12.2%), thistle (Cirsium spp.) (11.2%), bee balm (Monarda spp.) (10.4%), garden mints (Mentha spp.) (5.6%), spirea (Spirea spp.) (5.5%), purple coneflower (Echinacea purpurea) (4.0%), and silphiums (Silphium spp.) (4%). Russian sage (Perovskia atriplicifolia), Birds-foot trefoil (Lotus corniculatus), knapweed (Centaurea spp.), crown vetch (Secuigera varia), joe pye weed (Eutrochium purpureum), and red clover (Trifolium pretense), each supported between 1 and 3% of the bee visits. Most of these species were classified as either lawn or prairie species rather than garden species. While these species were commonly distributed across the city, there were other common species (Table 1) that were rarely visited, including hosta (Hosta spp.), bell flower (Campanula latifolia), and lillies (Lilium spp.). The most visited flowers classified as garden types included, mint, Russian sage, and spirea.
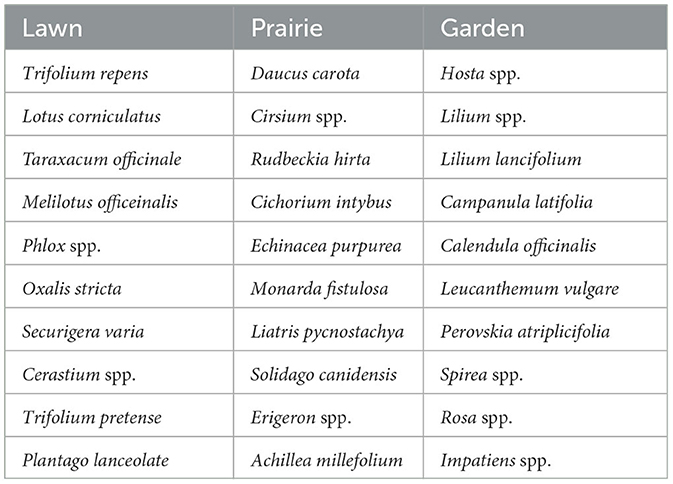
Table 1. Most common species of each flower ecotype group ordered by observation frequency.
Floral cover and diversity positively influenced the number of foraging bumble bees
The quasi-Poisson generalized linear regression model fit for the transect count data evidenced significant influence of flower cover as well as species richness, and a weakly significant influence of prairie and lawn flower species richness on the bee count data across the transects (Table 2). Additionally, the categorical factor of land use zoning, showed significant differences in the number of foraging bees where city parks supported 4 times more bumble bee foragers than agricultural transects on average, and residential and commercial transects supported about twice as many bees as agricultural transects (Figure 3C). The effect size of floral resource explanatory variables contrasted in terms of their unit increase influence on bee count in Figure 3. An additional global GLM fitted using a negative binomial distribution is presented in Supplementary Table 1, and shows similar patterns of predictive power for the explanatory variables. The negative binomial distribution model weighs small values more, while the quasi-poisson weighs the large values more.
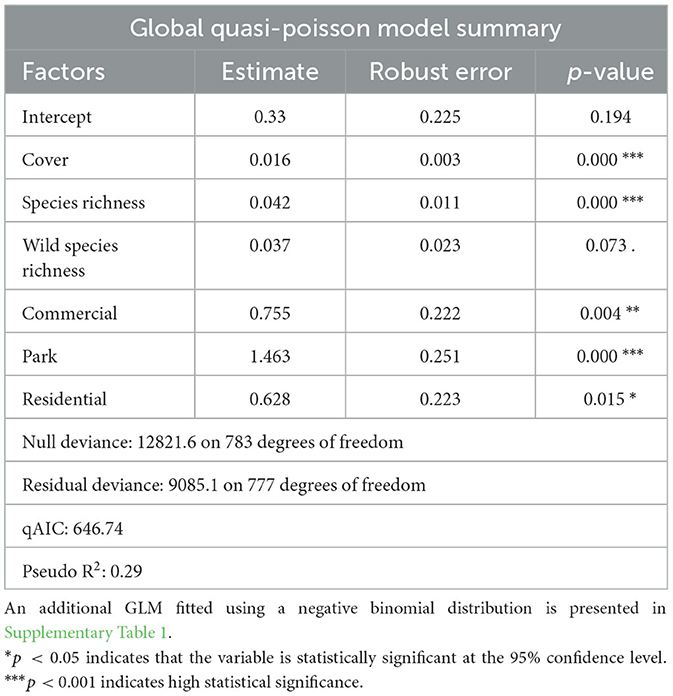
Table 2. Global model summary information for the quasi-Poisson fit regression model used to test the influence of floral resource explanatory variables and land use zoning on foraging bumble bee transect count data.
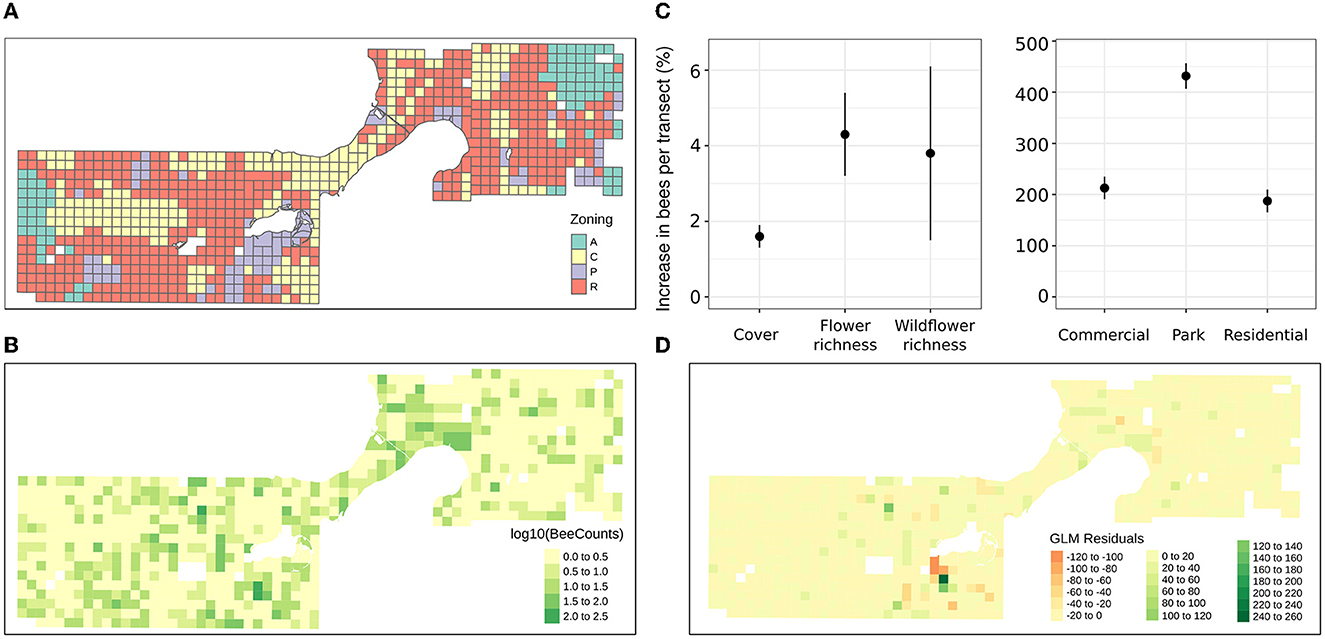
Figure 3. Transect grid polygon layer symbolized by (A) land use zoning type (B) log base 10 of the number of foraging bumble bees counted on each transect (2 = 100 bees) (C) estimated effects of unstandardized explanatory variables (expected increase in bees per variable unit, i.e., percent or additional species), and (D) the residuals of the global GLM, based on the fitted model and observed bumble bee count data.
Interactions between land use type and each floral resource variable were tested, yet interactions were only weakly significant in the full model, and when non-significant resource variable and zone class interaction subsets were eliminated, the weak significance dissipated, and more variance was explained by the primary floral resource factors.
Local models illustrated variance in effect sizes
A geographically-weighted generalized linear regression analysis was used to explore fluctuation of floral resource explanatory variable effect sizes across the study extent (Figures 4, 5). Local regressions were fit for each transect point, based on the data associated with the fifty closest transects, weighted by geographical distance so that data from the closest transects were more impactful in the regression model. The coefficients from each local regression model are plotted on each transect point to indicate the strength (Figure 4) and significance (Figure 5) of the explanatory variables in the local regression. Floral cover was the most consistently strong positive influence on bumble bee foraging counts across the study extent, consistent with the global quasipoisson generalized linear model. Density was the only non-significant floral resource explanatory variable. The slight positive influence of density was also widespread and highest in the agricultural and commercial areas.
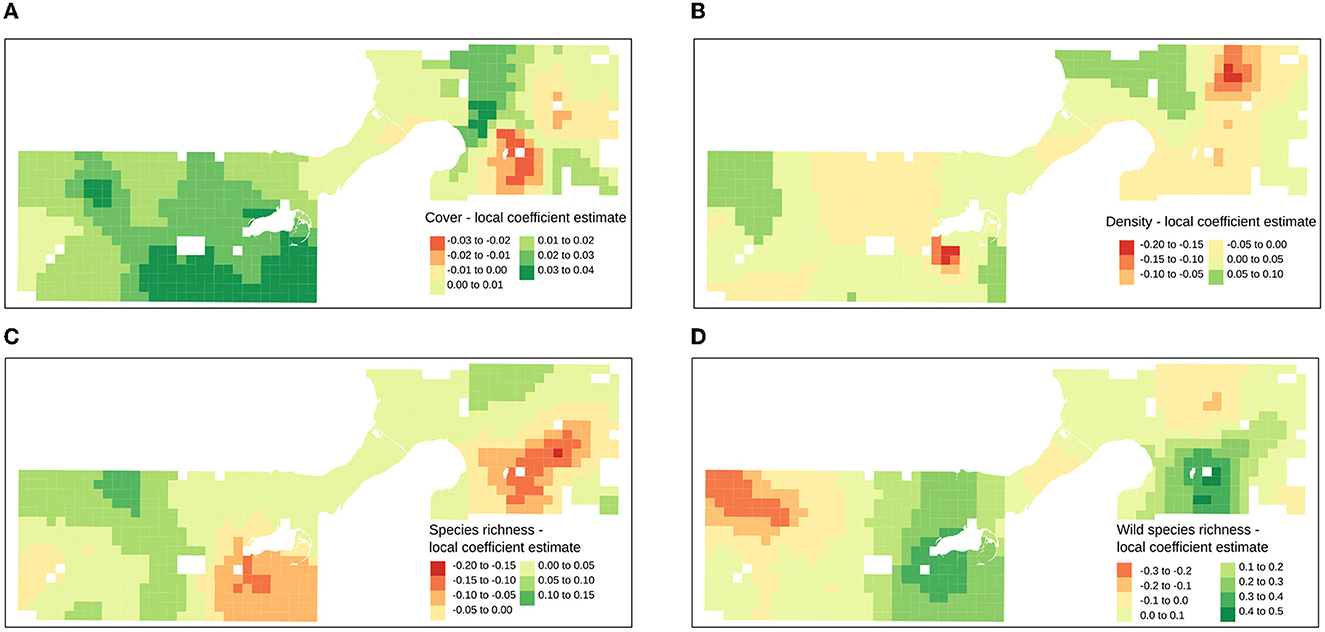
Figure 4. Raw explanatory variables and coefficient estimate results of geographically weighted regression mapped across study extent for (A) % flower cover (B) % flower density (C) flower species richness (D) wildflower species richness. Coefficient estimates can be interpreted as, −0.3 = 26%, −0.2 = 18%, −0.1 = 10% less bumble bees, 0.1 =11%, 0.2 = 22%, 0.3 = 35%, 0.4 = 49 % more bumble bees based on a marginal unit increase in the variable.
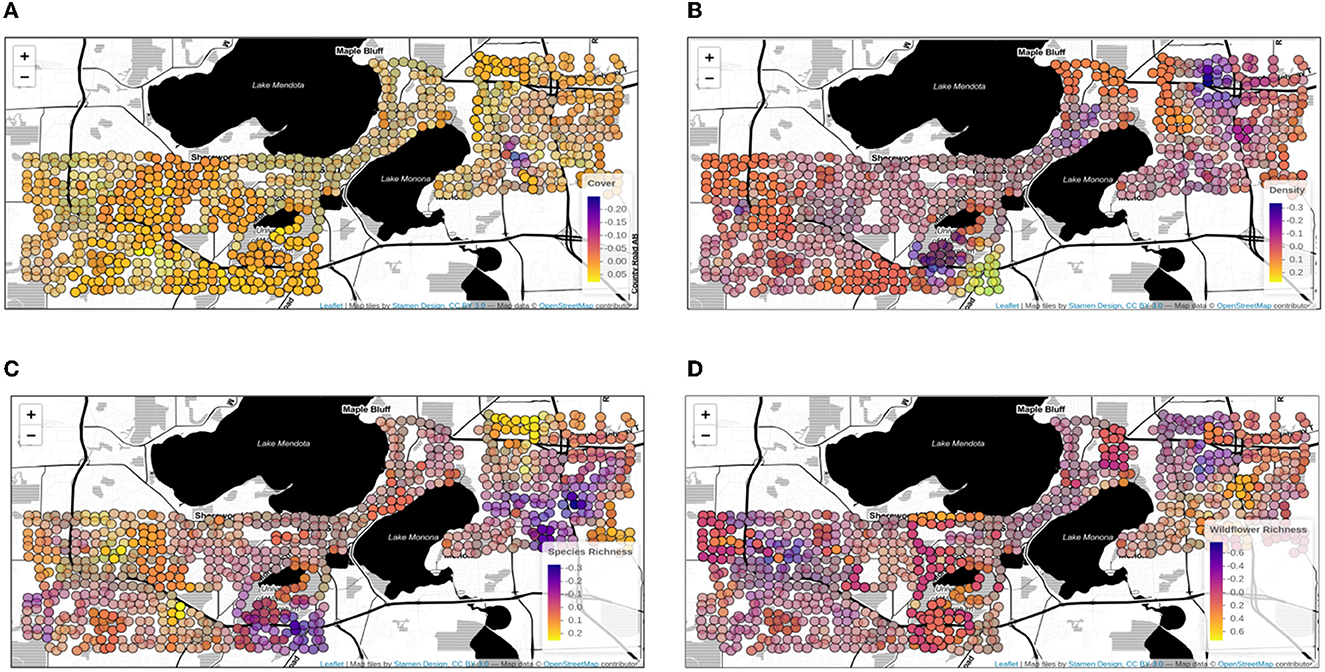
Figure 5. The geographically weighted regression coefficient estimates (same as Figure 4) mapped across study extent for (A) % flower cover (B) % flower density (C) flower species richness (D) wildflower species richness. Transparent dots indicate lack of statistical significance of variable effects within the local regression models, while opaque dots represent statistically significant coefficient estimates in the local regression models. Basemap data copyrighted OpenStreetMap contributors and available from https://www.openstreetmap.org.
While overall floral morphospecies richness and the richness of prairie and lawn flower morphospecies was positively correlated, this overall and non-managed subset of flower richness was distributed differently across land use zones. Commercial and residential areas sustained higher floral richness than parks and agricultural areas, however, parks and agricultural transects sustained rather higher richness of the lawn and prairie species subset. Across the study extent, the positive influence of overall species richness was more widespread and disappeared only in places where the positive influence of prairie and lawn species richness was particularly strong. These areas where prairie and lawn species richness stood out as the stronger effect in the glm regression model included areas with large, species-rich restored prairies (West – UW Madison Arboretum; East – Heritage Prairie, Elvehjem Park) surrounded by residential areas.
Discussion
The results of the study illustrate the spatial distribution of foraging bumble bees with regard to Madison's floral resources and the scales of spatial autocorrelation present in the bumble bee counts and floral resource variables. Furthermore, the global GLM of bumble bee abundance based on floral resources evidences the positive influence of floral cover and species richness, while the local regression results portray some variability in the influence of these floral resource variables across the urban extent.
Flower species and foraging bumble bees
Past studies have offered insight into bumble bee preferences for nutritious flower species by foraging bumble bees based on protein-lipid composition (Vaudo et al., 2016, 2018). Gardeners, pollinator enthusiasts, and ecologists have identified bumble bee-attractive flowers and encouraged the use of native plants in pollinator gardens for many years (Tuell et al., 2008; Williams et al., 2015). However, the benefit of lists or simplistic dichotomies has also been questioned, as pollinators tend to be generalists, and other nutritional or contextual factors may influence their foraging behavior (Garbuzov and Ratnieks, 2014; Rollings and Goulson, 2019). An investigation of urban plant-pollinator visitation networks between 24 pollinator morpho-types and 106 plant taxa revealed that elimination of all but four highly attractive plant taxa could maintain all observed pollinators (Lowenstein et al., 2018). Many flower species are likely useful to pollinators, and other factors like consistent availability of resources through the season might be more important factors to consider (Garbuzov and Ratnieks, 2014). Yet, dramatic differences in visitation across flower species (i.e., 14 species accounted for 70% of visits) underscore the variation in preference of foraging bumble bees (Lowenstein et al., 2018; Mach and Potter, 2018; Rollings and Goulson, 2019). To optimize pollinator habitat within urban gardens, selection of bee-attractive species that provide ample pollen and nectar resources is an important consideration.
Flower distribution and foraging bumble bees
Local significant positive spatial autocorrelation was observed for both floral resource variables and bumble bee observations, however autocorrelation levels were higher for floral resource variables and tapered off at shorter spatial lags (2 or 3 km) compared to bumble bee counts (5 km, Figure 2). The low positive Moran's I statistic values for the floral resource variables (I ~ 0.15–0.2), indicate that floral resources within a couple km are more similar to each other than the rest of the dataset, however not dramatically so. At a spatial lag of 4 or 5 km, almost all the floral resource variables exhibit negative spatial autocorrelation, indicating that variable values are more different from each other than the rest of the dataset. This appears to be driven by the dispersion of urban centers through the urban fabric, spacing heavily urbanized spaces between residential neighborhoods with lower coverage of impervious surface.
In recent years, an uptick in urban ecology studies has shed more light on the distribution of floral resources across cities (Ossola et al., 2019; Locke et al., 2021). A study of neighborhoods in Chicago revealed differences in the richness and composition of both spontaneous and cultivated plants between neighborhoods, with some differences explained by socioeconomic factors (Minor et al., 2023). Neighborhoods with lower racial or ethnic diversity had lower numbers of plants, while neighborhoods with intermediate numbers of Hispanic and white residents had the highest species richness, and a higher frequency of weedy species was reported in lower income neighborhoods (Lowenstein and Minor, 2016). Authors conclude that these patterns suggest evidence of disparities in plant-related ecosystem services.
Positive local autocorrelation of bumble bee counts (I ~ 0.1) was slightly less pronounced than autocorrelation of floral resource variables, and also extended for about twice as far. While this positive spatial autocorrelation is likely driven by differences in nesting or foraging resources between neighborhoods, the longer range of local autocorrelation is likely due to the mobility of the central place foraging bumble bees (Darvill et al., 2010). Differences in pollinator communities between urban spaces have been recorded in another recent urban pollinator study, wherein parks sustained higher richness and abundance of flower visiting insects than residential neighborhood blocks (Matteson et al., 2013), likely due to differences in habitat provisioned by these different land uses.
Global models and variance in the influence of floral resource variables on the number of foraging bumble bees
In the global GLM, we estimated average effect sizes of floral cover, density, overall floral richness, and wildflower richness for prediction of foraging bumble bees. This approach was used to test our hypotheses that our explanatory variables provided predictive capacity across our study extent. Consistent with other studies, we observed that these explanatory floral variables help predict bumble bee distribution (e.g., Matteson et al., 2013; Spiesman et al., 2016). However, we also expected that the influence of particular variables might differ based on local context. For instance, where floral resources are abundant and specious, the highest quality floral resource patches likely attract the most bumble bees. Investigating the residuals of the model could help to identify locations that surpass our expectations of foraging quality, and locations which fail to meet our predictions. In our map of global model residuals (Figure 3), we can see some high and low predictions of bumble bee foraging, especially several very attractive transects at the UW Madison Arboretum. We can hypothesize about what might cause this additional variation in bumble bee foraging, and set up new studies to test these hypotheses, but we also can explore potential variabilities in the strength of our predictor variables using geographically-weighted regression. This comprehensive investigation of fluctuation in the effect sizes of floral resource explanatory variables across the study extent contextualizes some differing results observed in studies that focus only on a particular ecotype within cities or exurban ecotypes.
In the global model, we tested for evidence of interactions between land use zoning class and floral resource variables, but finally none were statistically significant (α = 0.05). Several interactions were weakly significant, including, the interaction between city parks and native flower species richness, boosting the number of foraging bumble bees when they occurred together.
Floral cover and density
Loss of habitat including nesting and foraging resources are likely the most important factors contributing to loss of pollinators around the world (Goulson et al., 2008; Williams and Osborne, 2009; Winfree et al., 2009; Potts et al., 2010). Yet, it is unclear in what contexts various aspects of floral abundance and distribution may be most critical for local bumble bees (Crone and Williams, 2016; Spiesman et al., 2016). The geographically weighted regression results in this study highlight the pervasive positive influence of flower cover, and the slight positive influence of flower density – which is most apparent in agricultural zones and commercial zones. Floral resources are a primary limiting factor for pollinator populations, but local circumstances of distribution and phenology should be also be considered in depth. Studies located in resource pulse landscapes have observed beneficial results of late blooming mass-flowering crops, but not early blooming mass-flowering crops, suggesting increased floral resources as colonies are reaching maximum size has a much different effect than a bump in floral food resources while the colony is still growing. Bumble bees store very little extra food resources in the nest, so continuous access to floral resources during the season is crucial (Dornhaus and Chittka, 2005). Recent studies have documented variation in foraging distance in response to floral abundance in the surrounding landscape evidencing flexibility that is essential for bumble bees to respond to variability in the spatial and temporal distribution of resources (Jha and Kremen, 2013; Vasseur et al., 2013).
Several recent studies have isolated the efficiency of foragers in resource collection as the primary driver of colony growth (Williams et al., 2012; Spiesman et al., 2016; Requier et al., 2020). One study found no influence of landscape scale floral resources when local resources were high, and in this case found high flower species dominance as the primary driver of colony growth (Spiesman et al., 2016). The association between transect scale floral cover and foraging bumble bees seems quite clear, and persistently positive across the study extent. It's possible that the spatial scale at which density was estimated, and the heterogeneity within a transect, obscured the usefulness of this metric in our study.
Floral species richness, floral type, and foraging bumble bees
Floral richness positively influenced the count of foraging bumble bees more than the richness of prairie and lawn species alone. While many common lawn and prairie species were highly attractive to foraging bumble bees and supported the majority of foraging visits, there were highly bumble bee-attractive garden plants as well, including mints, Russian sage, and spirea. As micronutrients may vary greatly between plant species, the need for diverse floral resources may vary based on the nutritional content of the available assemblage.
In two parts of the city, a strong, positive influence of native plant species diversity overcame the influence of overall species richness in the geographically weighted regression results. This phenomenon appeared to result in compensatory negative coefficients in the overall floral species richness variable, where the prairie and lawn species subset surpassed it as a strong influential explanatory variable. These areas represented localities where species rich prairies were nested in residential communities. While it seems that, generally, floral richness is an important predictor of foraging resource quality, in some circumstances, wildflower richness provides more explanatory capacity and comprises relatively better quality of floral resources. This result underscores the value of relatively large extents of restored prairie inside the urban matrix for bumble bees.
As cities grow, it is vital to maintain urban lands that provide bumble bee foraging resources, such that bumble bees can continue to maintain pollination services. This study demonstrated the importance of floral resource distribution on the prevalence of foraging bumble bees and potential disparities in plant and pollinator related ecosystem services across the city. The results of this study show that landscapes with high cover as well as floral diversity would maximize bumble bee foraging. In neighborhoods with ample floral resources or an abundance of ornamental cultivars, wildflower species presence is particularly important. Given that urban landscapes are not homogeneous and that floral resources are not consistently distributed across the cityscape, valuable pollinator habitat must be protected, and supplemented where gaps persist. Comprehensive consideration of cityscapes can help to prioritize conservation efforts to protect high value bumble bee foraging resources and ameliorate biodiversity “deserts.” Future work could contribute to the understanding of floral phenology and bumble bee foraging behavior throughout the season.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
VP planned and executed the study. JS and JZ reviewed the data analysis. VP drafted the manuscript and all authors reviewed it. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by the United States Department of Agriculture - Agricultural Research Service (Project #2090-32000-040-000-D). Mention of trade names or commercial products in this publication is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the USDA. The USDA is an equal opportunity provider and employer.
Conflict of interest
JS was employed by the Silvernail Studio for Geodesign, LLC.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/frsc.2023.1103721/full#supplementary-material
References
Bhattacharya, M., Primack, R. B., and Gerwein, J. (2003). Are roads and railroads barriers to bumblebee movement in a temperate suburban conservation area? Biol. Conserv. 109, 37–45. doi: 10.1016/S0006-3207(02)00130-1
Brosi, B. J., Armsworth, P. R., and Daily, G. C. (2008). Optimal design of agricultural landscapes for pollination services. Conserv. Lett. 1, 27–36. doi: 10.1111/j.1755-263X.2008.00004.x
Crone, E. E., and Williams, N. M. (2016). Bumble bee colony dynamics: quantifying the importance of land use and floral resources for colony growth and queen production. Ecol. Lett. 19, 460–468. doi: 10.1111/ele.12581
Darvill, B., O'Connor, S., Lye, G. C., Waters, J., Lepais, O., Goulson, D., et al., (2010). Cryptic differences in dispersal lead to differential sensitivity to habitat fragmentation in two bumblebee species. Mol. Ecol. 19, 53–63. doi: 10.1111/j.1365-294X.2009.04423.x
Dornhaus, A., and Chittka, L. (2005). Bumble bees (Bombus terrestris) store both food and information in honeypots. Behav. Ecol. 16, 661–666. doi: 10.1093/beheco/ari040
Galpern, P., Manseau, M., and Wilson, P. (2012). Grains of connectivity: analysis at multiple spatial scales in landscape genetics. Mol. Ecol 21, 3996–4009. doi: 10.1111/j.1365-294X.2012.05677.x
Garbuzov, M., and Ratnieks, F. L. W. (2014). Listmania: the strengths and weaknesses of lists of garden plants to help pollinators. BioScience 64, 1019–1026. doi: 10.1093/biosci/biu150
Garibaldi, L. A., Steffan-Dewenter, I., Winfree, R., Aizen, M. A., Bommarco, R., Cunningham, S. A., et al., (2013). Wild pollinators enhance fruit set of crops regardless of honey bee abundance. Science 339, 1608–1611. doi: 10.1126/science.1230200
Goddard, M. A., Dougill, A. J., and Benton, T. G. (2010). Scaling up from gardens: biodiversity conservation in urban environments. Trends Ecol. Evol. 25, 90–98. doi: 10.1016/j.tree.2009.07.016
Goulson, D., Lepais, O., O'Connor, S., Osborne, J. L., Sanderson, R. A., Cussans, J., et al., (2010). Effects of land use at a landscape scale on bumblebee nest density and survival. J. Appl. Ecol. 47, 1207–1215. doi: 10.1111/j.1365-2664.2010.01872.x
Goulson, D., Lye, G. C., and Darvill, B. (2008). Decline and conservation of bumble bees. Annu. Rev. Entomol 53, 191–208. doi: 10.1146/annurev.ento.53.103106.093454
Hemberger, J., and Gratton, C. (2018). Resource pulse decreases bumble bee foraging trip duration in central Wisconsin agroecosystem. Ecol. Entomol 43, 447–457. doi: 10.1111/een.12516
Jha, S., and Kremen, C. (2013). Bumble bee foraging in response to landscape heterogeneity. Proc. Natl. Acad. Sci. U.S.A. 8, 555–558. doi: 10.1073/pnas.1208682110
Kalogirou, S. (2018). “lctools” R package: Local correlation, spatial inequalities, geographically-weighted regression and other tools. Version 2, 2–6.
Klein, A. M., Vaissière, B. E., Cane, J. H., Steffan-Dewenter, I., Cunningham, S. A., Kremen, C., et al., (2007). Importance of pollinators in changing landscapes for world crops. P. Roy. Soc. B-Biol. Sci. 274, 303–313. doi: 10.1098/rspb.2006.3721
Locke, D. H., Ossola, A., Minor, E., and Lin, B. B. (2021). Spatial contagion structures urban vegetation from parcel to landscape. People Nat. 4, 88–102. doi: 10.1002/pan3.10254
Lowenstein, D. M., Matteson, K. C., and Minor, E. S. (2018). Evaluating the dependence of urban pollinators on ornamental, non-native, and “weedy” floral resources. Urban Ecosyst. 22, 293–302. doi: 10.1007/s11252-018-0817-z
Lowenstein, D. M., and Minor, E. S. (2016). Diversity in flowering plants and their characteristics: integrating humans as a driver of urban floral resources. Urban Ecosyst. 19, 1735–1748. doi: 10.1007/s11252-016-0563-z
Lynch, L., Kangas, M., Ballut, N., Doucet, A., Schoenecker, K., Johnson, P., et al., (2021). Changes in land use and land cover along an urban-rural gradient influence floral resource availability. Curr. Landscape Ecol. Rep. 6, 46–70. doi: 10.1007/s40823-021-00064-1
Mach, B. M., and Potter, D. A. (2018). Quantifying bee assemblages and attractiveness of flowering woody landscape plants for urban pollinator conservation. PLoS ONE 13, e0208428. doi: 10.1371/journal.pone.0208428
Matteson, K. C., Grace, J. B., and Minor, E. S. (2013). Direct and indirect effects of land use on floral resources an flower-visiting insects across an urban landscape. Oikos 122, 682–694. doi: 10.1111/j.1600-0706.2012.20229.x
McFrederick, Q. S., and LeBuhn, G. (2006). Are urban parks refuges for bumble bees Bombus spp. (Hymenoptera: Apidae)? Biol. Conserv. 129, 372–382. doi: 10.1016/j.biocon.2005.11.004
Minor, E., Lopez, B., Smith, A., and Johnson, P. (2023). Plant communities in Chicago residential neighborhoods show distinct spatial patterns. Landscape Urban Plan. 232, 104663. doi: 10.1016/j.landurbplan.2022.104663
O'Connell, M. C., Jordan, Z., McGilvray, E., Cohen, H., Liere, H., Lin, B. B., et al., (2021). Urban. Ecosystems 24, 391–404. doi: 10.1007/s11252-020-01043-w
Ollerton, J., Winfree, R., and Tarrant, S. (2011). How many flowering plants are pollinated by animals? Oikos 120, 321–326. doi: 10.1111/j.1600-0706.2010.18644.x
Osborne, J. L., Martin, A. P., Shortall, C. R., Todd, A. D., Goulson, D., Knight, M. E., et al.,(2008). Quantifying and comparing bumblebee nest densities in gardens and countryside habitats. J. Appl. Ecol. 45, 784–792. doi: 10.1111/j.1365-2664.2007.01359.x
Ossola, A., Locke, D., Lin, B., and Minor, E. (2019). Greening in style: Urban form, architecture and the structure of front and backyard vegetation. Landscape Urban Plan. 185, 141–157. doi: 10.1016/j.landurbplan.2019.02.014
Pfeiffer, V., Silbernagel, J., Guédot, C., and Zalapa, J. (2019). Woodland and floral richness boost bumble bee density in cranberry resource pulse landscapes. Landscape Ecol. 34, 979–996. doi: 10.1007/s10980-019-00810-1
Pope, N. S., and Jha, S. (2018). Seasonal food scarcity prompts long-distance foraging by a wild social bee. Am. Nat. 191, 45–57. doi: 10.5061/dryad.c6j8p
Potts, S. G., Roberts, S. P. M., Dean, R., Marris, G., Brown, M. A., Jones, R., et al., (2010). Declines of managed honey bees and beekeepers in Europe. J. Apic. Res. 49, 15–22. doi: 10.3896/IBRA.1.49.1.02
Pywell, R. F., Warman, E. A., Hulmes, L., Hulmes, S., Nuttall, P., Sparks, T., et al., (2006). Effectiveness of new agri-environment schemes in providing foraging resources for bumblebees in intensively farmed landscapes. Biol. Conserv. 129, 192–206. doi: 10.1016/j.biocon.2005.10.034
R Core Team. (2013). R: A Language and Environment for Statistical Computing. Vienna, Austria: R Foundation for Statistical Computing.
Redhead, J. W., Dreier, S., Bourke, A. F. G., Heard, M. S., Jordan, W. C., Sumner, S., et al., (2016). Effects of habitat composition and landscape structure on worker foraging distances of five bumble bee species. Ecol Appl. 26, 726–739. doi: 10.1890/15-0546
Requier, F., Jowanowitsch, K., Kallnik, K., and Steffan-Dewenter, I. (2020). Limitation of complementary resources affects colony growth, foraging behavior, and reproduction in bumble bees. Ecology 101, e02946. doi: 10.1002/ecy.2946
Rollings, R., and Goulson, D. (2019). Quantifying the attractiveness of garden flowers for pollinators. J. Insect Conserv. 23, 803–817. doi: 10.1007/s10841-019-00177-3
Spiesman, B. J., Bennett, A., Isaacs, R., and Gratton, C. (2016). Bumble bee colony growth and reproduction depend on local flower dominance and natural habitat area in the surrounding landscape. Biol. Conserv. 206, 217–223. doi: 10.1016/j.biocon.2016.12.008
Threlfall, C. G., Walker, K., Williams, S. G., Hahs, A. K., Mata, L., Stork, N., et al., (2015). The conservation value of urban green space habitats for Australian native bee communities. Biol. Conserv. 187, 240–248. doi: 10.1016/j.biocon.2015.05.003
Tuell, J. T., Fiedler, A. K., Landis, D., and Isaacs, R. (2008). Visitation by wild and managed bees (Hymenoptera: Apoidea) to eastern U.S. native plants for use in conservation programs. Environ. Entomol. 37, 707–718. doi: 10.1603/0046-225X(2008)37707:VBWAMB2.0.CO;2
Vasseur, C., Joannon, A., Aviron, S., Burel, F., Meynard, J., and Baudry, J. (2013). The cropping systems mosaic: How does the hidden heterogeneity of agricultural landscapes drive arthropod populations? Agric. Ecosyst. Environ. 166, 3–14. doi: 10.1016/j.agee.2012.08.013
Vaudo, A. D., Farrell, L. M., Patch, H. M., Grozinger, C. M., and Tooker, J. F. (2018). Consistent pollen nutritional intake drives bumble bee (Bombus impatiens) colony growth and reproduction across different habitats. Ecol Evol 8, 5765–5776. doi: 10.1002/ece3.4115
Vaudo, A. D., Patch, H. M., Mortensen, D. A., Tooker, J. H., and Grozinger, C. M. (2016). Macronutrient ratios in pollen shape bumble bee (Bombus impatiens) foraging strategies and floral preferences. Proc. Natl. Acad. Sci. U.S.A., 113, E4035–E4042. doi: 10.1073/pnas.1606101113
Williams, N. M., Regetz, J., and Kremen, C. (2012). Landscape-scale resources promote colony growth but not reproductive performance of bumble bees. Ecology 93, 1049–1058. doi: 10.1890/11-1006.1
Williams, N. M., Ward, K. L., Pope, N., Isaacs, R., Wilson, J., May, E. A., et al., (2015). Native wildflower plantings support wild bee abundance and diversity in agricultural landscapes across the United States. Ecol. Appl. 25, 2119–2131. doi: 10.1890/14-1748.1
Williams, P. H., and Osborne, J. L. (2009). Bumblebee vulnerability and conservation world-wide. Apidologie 40, 367–387. doi: 10.1051/apido/2009025
Williams, P. H., Thorp, R. W., Richardson, L. L., and Colla, S. R. (2014). An Identification Guide: Bumble Bees of North America. Princeton: Princeton University Press.
Winfree, R., Aguilar, R., Vázquez, D. P., LeBuhn, G., and Aizen, M. A. (2009). A meta-analysis of bees' responses to anthropogenic disturbance. Ecology 90, 2068–2076. doi: 10.1890/08-1245.1
Keywords: urban bees, floral resources, foraging ecology, geographically weighted regression (GWR), non-stationarity, bumble bees (Bombus), sustainable cities, pollinators
Citation: Pfeiffer VW, Zhu J, Poh K and Silbernagel J (2023) Local influence of floral resource attributes on urban bumble bee foraging activity. Front. Sustain. Cities 5:1103721. doi: 10.3389/frsc.2023.1103721
Received: 20 November 2022; Accepted: 23 June 2023;
Published: 13 July 2023.
Edited by:
Natacha P. Chacoff, Universidad Nacional de Tucumán, ArgentinaReviewed by:
Nicole Miller-Struttmann, Webster University, United StatesRodrigo M. Barahona-Segovia, University of Los Lagos, Chile
Copyright © 2023 Pfeiffer, Zhu, Poh and Silbernagel. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Vera W. Pfeiffer, dmVyYS53LnBmZWlmZmVyQGdtYWlsLmNvbQ==