 Nicole C. S. Bell
Nicole C. S. Bell John S. Ascher2
John S. Ascher2 Jen J.-M. Hayes
Jen J.-M. Hayes Mallory Mead
Mallory Mead Gail A. Langellotto
Gail A. Langellotto
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
SYSTEMATIC REVIEW article
Front. Sustain. Cities, 30 October 2023
Sec. Urban Greening
Volume 5 - 2023 | https://doi.org/10.3389/frsc.2023.1102360
This article is part of the Research TopicWorld Bee Day 2022: Pollinators in Urban EnvironmentsView all 10 articles
Urban garden spaces are potentially important habitats for bee conservation. Gardens can host diverse flora, which provide floral resources across foraging seasons for bee species. Recent reviews have focused on the impacts of cityscapes on urban bee assemblages in different green spaces. Urban gardens are distinct from other urban green spaces, and bee communities in urban spaces have been an increasing topic of study over the past few decades. We reviewed 28 urban garden bee studies spanning five decades and 14 countries to compile an original metadataset of bee species' functional traits to understand the conservation value of gardens, identify gaps in bee sampling efforts, and summarize the calls to action included by their authors. Studies of urban garden bees have documented between 674 (conservative count, excluding morphospecies) and 830 (liberal count, including morphospecies) bee species. Urban garden bee communities were taxonomically and functionally diverse, although bee species that were non-eusocial, ground-nesting, generalist foragers, and native were most common in garden habitats. The proportion of parasitic bee species and specialist foragers found in urban gardens was comparable to proportions for global bee taxa. This suggests that gardens contain the hosts and forage needed to support bees with specialized life history requirements, and thus represent high quality habitat for a subset of bee communities. Garden bee research was strongly biased toward the northern hemisphere, which signifies a large gap in our understanding of garden bee communities in other regions. The variety of, and non-standard sampling methods in garden bee research makes it difficult to directly compare results between studies. In addition, both intentional low taxonomic resolution and a lack of collaboration with taxonomists constrains our understanding of bee diversity. Our analyses highlight both successes of past urban garden bee studies, and areas of opportunity for future research as we move into a sixth decade of garden bee research.
Native bees are critically important organisms that support biodiversity and crop production via their pollination services (Klein et al., 2007; Winfree et al., 2008; Ollerton et al., 2011). Animals pollinate 87% of flowering plant species, with the majority of animal-mediated pollination conferred by bees (Ollerton et al., 2011; Christmann, 2019) making them the primary pollinators of most agricultural crops and wild plants (Potts et al., 2010). Substantial losses of bees have been widely reported (Biesmeijer et al., 2006; Potts et al., 2010; Goulson et al., 2015; Zattara and Aizen, 2021), although evidence is sparse for most species, outside of bumble bees (Bartomeous et al., 2013; Soroye et al., 2020) and mason bees (LeCroy et al., 2020). While large-scale seasonal losses of managed western honeybee colonies have been reported recently, the number of global colonies has increased by 45% (Potts et al., 2016).
Despite disagreement about the extent to which bee species (beyond bumble bees and mason bees) are in decline (Goulson and Nicholls, 2016) and about the causes of potential declines, media coverage about bee population losses has increased public attention and enthusiasm for bee conservation (Wilson et al., 2017). In particular, public attention on bees has highlighted opportunities to promote bee conservation in public and private urban spaces (Sirohi et al., 2015; Threlfall et al., 2015; Turo and Gardiner, 2019; Hall and Martins, 2020; Hane and Korfmacher, 2022). Cities can provide an array of habitat options for bees, including urban gardens and parks. Recent reviews have examined the influence of urban landscapes on bee communities across a broad range of habitats, including gardens, but also including cemeteries, vacant lots, wastelands, parks, and remnant native vegetation (Ayers and Rehan, 2021; Prendergast et al., 2022). Although these reviews have advanced our understanding of urban bee communities, we suggest that gardens are distinctly different from other urban habitats, and thus deserve separate consideration.
Urban gardens are characterized by a diverse and heterogeneous plant assemblage that is actively tended by gardeners (Threlfall et al., 2016). The extremely high diversity of plants in garden sites, relative to other urban greenspaces, reflects the large pool of plants available to gardeners via the nursery trade (Thompson et al., 2003). In addition, management and maintenance decisions made by gardeners ultimately contributes to high within-garden heterogeneity, compared to most other urban greenspaces (Thompson et al., 2003). For example, gardeners' decisions might result in areas dedicated to fruit trees, annual vegetables, lawn, shade trees and plants adapted to growing in shade, ornamental cultivars, and/or native plants, all within a single garden. Urban parks, golf courses, or cemeteries in contrast, are typically planted with species from a limited plant palette, and subject to management practices that tend to homogenize plant communities across urban green spaces (Threlfall et al., 2016). Other urban habitat types, including wastelands and vacant lots are often minimally cultivated or managed (Gardiner et al., 2013; Twerd and Banaszak-Cibicka, 2019), and offer limited opportunities for public engagement. Thus, garden plant assemblages vary considerably over small scales, whereas urban parks, golf courses, and cemeteries are typically more homogenous.
Gardens also offer opportunities to engage the general public in bee conservation efforts via science-based planting and management practices (Anderson et al., 2022), unlike vacant lots and other minimally managed urban greenspaces. The management of urban gardens, though, can also vary widely, both within and between individual gardens, including those gardens with vigorous maintenance and frequent mowing (Ayers and Rehan, 2021) and those that use more natural landscaping approaches (McCarthy, 2018). Smaller urban gardens, including home, allotment, or community gardens, can harbor nearly as much diversity as larger urban parks and adjacent natural areas (Fetridge et al., 2008; Normandin et al., 2017; Baldock et al., 2019). There is value, then, in zooming in to bee communities within urban gardens, to better understand their bee communities, and to share with gardeners which practices might enhance or impede bee conservation efforts.
The conservation value of gardens has repeatedly been referenced in the literature (Tommasi et al., 2004; Matteson et al., 2008; Pawelek et al., 2009; Lerman and Milam, 2016; Plascencia and Philpott, 2017; Lanner et al., 2020). In fact, an interest in urban gardens as a space for insect conservation dates back to 1941, when entomologist Frank Lutz documented 1,402 insect species in his suburban 15,000 m2 garden in Ramsey, NJ, a suburb of New York City, USA (Lutz, 1941). Gardens can provide nest sites (Cane, 2001; Tonietto et al., 2011) and diverse floral resources which can provide pollen and nectar across the entire foraging season for many bee species (Tommasi et al., 2004; Burdine and McCluney, 2019; Lanner et al., 2020). Gardens may also be sites of heavy pesticide use (Meftaul et al., 2020), which can have lethal or sublethal effects on native bees (Hladik et al., 2016). Highly bred ornamental plants, with reduced floral rewards of nectar and/or pollen, often dominate garden plant assemblages, which may limit the usefulness of urban gardens for bees (Comba et al., 1999; Corbet et al., 2001; Wenzel et al., 2020). However, gardeners' enthusiasm for bee conservation also has potential to drive plant purchases toward native species (Anderson et al., 2021), which tend to attract more diverse bee assemblages (Williams et al., 2011; Morandin and Kremen, 2013; Pardee and Philpott, 2014; Salisbury et al., 2015; Anderson et al., 2022). Although we have a general sense of the types of management practices that enhance or exclude bee taxa in gardens, we also have an opportunity to look at the types of bees that are relatively abundant or sparse in garden systems. Doing so may reveal more specific garden design and management practices that can enhance bee conservation efforts. A recently published review (Rahimi et al., 2022) focused on functional traits of bees in urban gardens, though only dominant garden bee species (the most abundant bee species from each study) were examined. Additionally, most bees were categorized to the family or genera level, with the exception of European honeybees (a non-native and managed species across most areas of the globe), which precludes consideration of species-specific patterns.
We thus compiled a metadataset (a dataset of datasets) of urban garden bee study characteristics and the functional traits of species identified, using studies which took place over the past 50 years. We used these data to address three objectives. First, we characterized the state of urban garden bee research, including geographic extent and methodologies used to study bees in home, community/allotment, or rooftop gardens. Second, we characterized the abundance and richness of bee species that have been found in urban gardens, as well as bees' functional traits, to better understand the types of bee taxa that are more or less common in garden bee communities. Finally, we evaluated the bee conservation recommendations that have emerged from these papers, which we refer to as a “call to action”.
To assemble the garden bee research literature, we conducted standardized searches of the databases WebofScience, AGRICOLA (EBSCOHost), and CAB Direct on January 19, 2022, using the Boolean search term: (pollinator* OR bee OR Apoidea) AND garden AND (urban OR community) AND (visitation OR richness OR diversity OR “functional diversity”) NOT (beekeeping OR apiculture). We used the Boolean operator of NOT (beekeeping OR apiculture) to exclude the multitude of studies on honeybee biology, which was not the focus of this review.
The initial search yielded 227 peer-reviewed publications after the removal of duplicates. To be included in the metadataset, studies had to have been conducted in urban garden spaces: home gardens (i.e., a home garden cultivated for the personal use of the tenants or homeowners); community/allotment gardens (i.e., shared, semi-public space, typically containing several different plots cultivated by different individuals); or rooftop gardens (garden on the rooftop of a multi-tenant building). Non-garden urban bee studies, including those conducted in parks, vacant lots, remnant habitats, or other non-garden green spaces, were not included in this review. Additional criteria for inclusion were that studies had to have identified at least 50% of bee taxa to species level and had to present bee data separate from (rather than pooled with) other taxa. Species level determinations were necessary in order to ensure that we were accurately categorizing bees' functional traits (e.g., nesting habit, sociality, native status, foraging breadth). Though we considered including two papers that were just under this 50% threshold (Lowenstein et al., 2015, 2019), these papers were ultimately excluded due to low taxonomic resolution in combination with only utilizing sight identification, thus making it difficult to evaluate identifications. Coarse groupings of bees as “small” and “large” bees (e.g., Fukase and Simons, 2016) precluded functional trait assignments to garden bees from some studies. It is possible that a lack of available taxonomists (Drew, 2011) might have limited taxonomic resolution in these and other urban garden bee studies. Papers that reported bee communities in both urban and non-urban sites, or both garden and non-garden sites, were included if it was possible to identify and specifically extract bee data associated with only the urban garden sites. Papers from all geographical regions and publication dates were included, provided the above criteria were met (Figure 1).
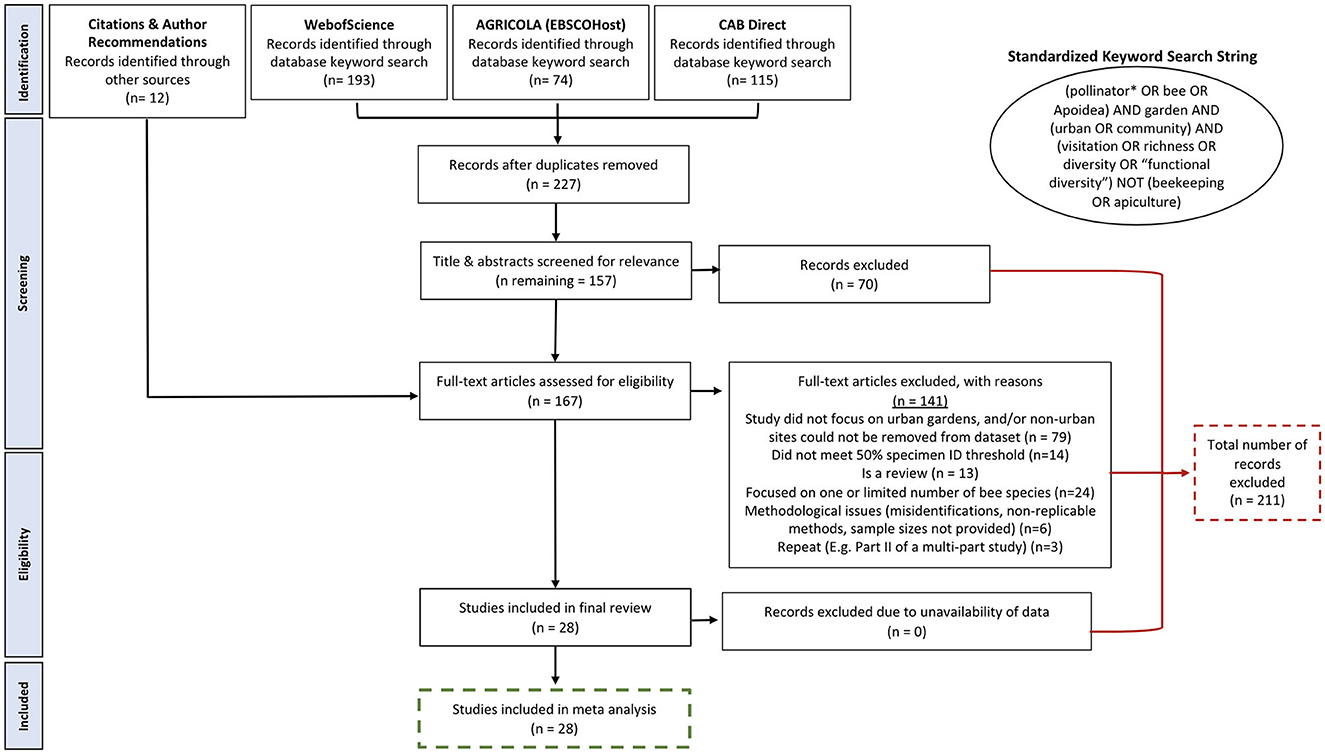
Figure 1. PRISMA flow diagram depicting the databases searched with a standardized keyword search string, and the process to finalize the 28 publications included in this review.
We initially screened abstracts for relevancy (i.e., urban garden studies that identified bees to species), and then screened the full text of papers that passed initial screening prior to coding them. After application of the inclusion and exclusion criteria, we retained 19 papers, including studies of bees on urban farms (Sivakoff et al., 2018), botanical gardens (Pardee and Philpott, 2014), and a university garden (Wojcik et al., 2008). These studies were retained in our metadataset because study sites were similar in size and flowering plant diversity to typical home or community gardens. Another six papers were included after searching the bibliographies of these 18 papers for additional references. Four papers were added during the review process, including two that were published after our initial literature search (Ollerton et al., 2022; Prendergast et al., 2022). The Ollerton et al. (2022) paper was a collaborative effort across the globe by different scientists, and some sites were gardens; we were able to include 40 garden sites from this paper. Two papers (Halder et al., 2013; Del Toro and Ribbons, 2020) were later removed from the metadataset due to concerns related to species identifications. Del Toro and Ribbons (2020) was retracted by the publisher in November 2022. Halder et al. (2013) was removed from our dataset due to multiple species misspellings and identifying species far outside of their known range (e.g., Xylocopa nigrita only occurs in Africa, and Xylocopa calens only occurs in Madagascar; Pauly, 2016). We also included one book in this review (Owen, 2010). Although not peer-reviewed, the extensive and decades-long sampling of a single garden space was meticulously documented and reported, such that the book met all inclusion criteria. The early documentation of insects from Lutz (1941) was excluded because only order-level identifications were listed. Our garden bee metadataset was thus drawn from 27 journal articles published from, 1990 to 2022, and one book published in 2010 (Table 1).
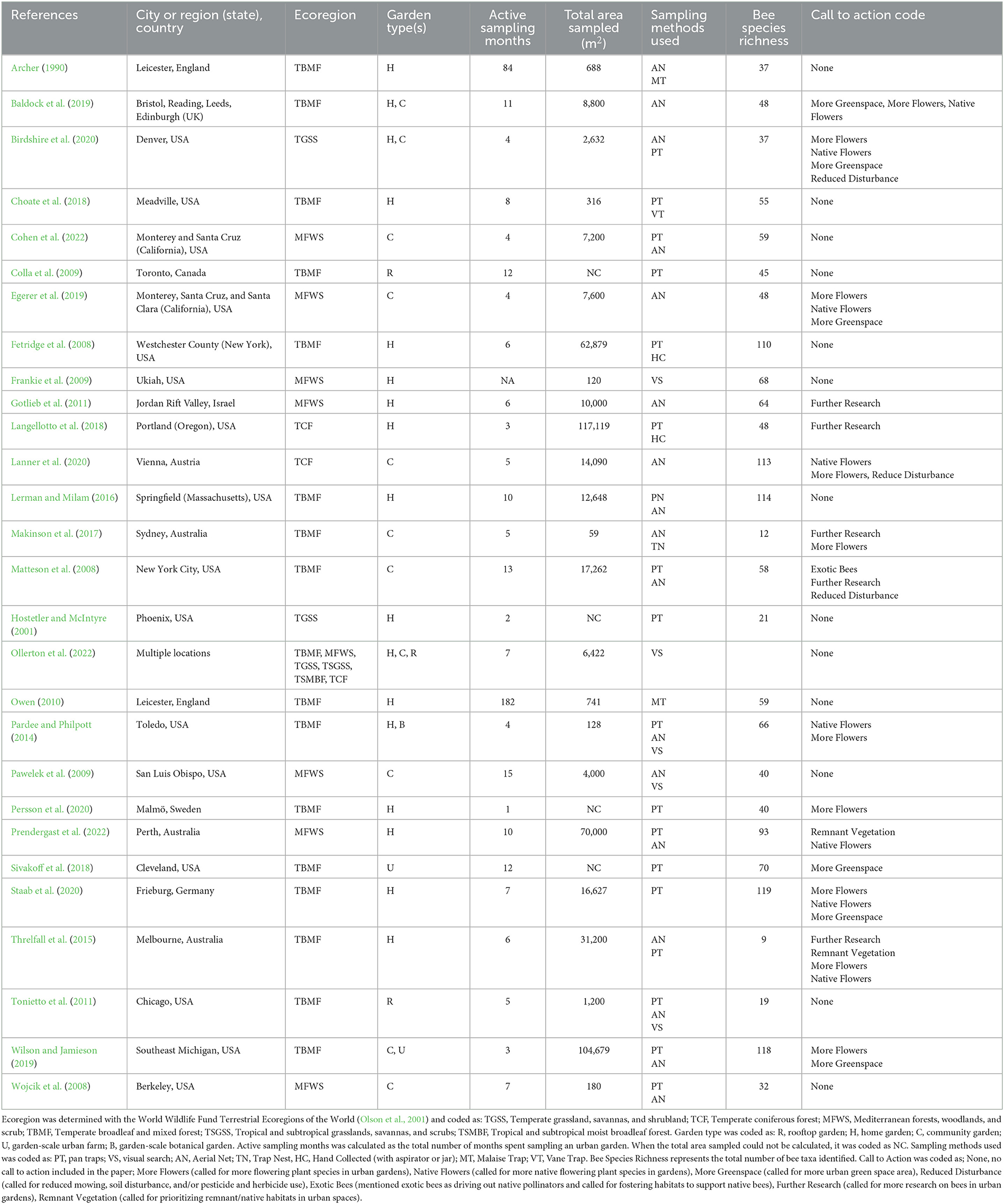
Table 1. List of garden bee studies included in this review, and study characteristics.
To better understand the current state of garden bee research, as well as any gaps, we extracted the following data from each paper: study location (city/cities, country) and biome; type of garden (community, home, garden-scale urban farm, garden-scale botanical garden, rooftop) and number of plots/gardens sampled; cumulative area (m2) of urban garden(s) studied and sampled and timeframe of the study (total number of active sampling months to account for those that spanned multiple years); type of sampling methods employed (pan traps, aerial netting, malaise trap, visual search, trap nest, vane trap, hand collection); number of bee species found (species richness), and bee species' functional traits, if identified (sociality, diet breadth, and nesting location). Biome was determined using the World Wildlife Fund Terrestrial Ecoregions of the World (Olson et al., 2001). When data were not available in the paper or supplementary material, we contacted authors for additional information. Information on site size was not available for Hostetler and McIntyre (2001), Colla et al. (2009), Sivakoff et al. (2018), and Persson et al. (2020). These studies were not included in estimates of cumulative urban garden area sampled. Active sampling months was not available for Frankie et al. (2009).
For all studies, we noted the prevalent research themes (including, but not limited to, baseline pollinator assessment, comparative landscape study, and effects of urbanization), to identify areas that have been investigated across urban garden bee studies (Table 2). We also recorded whether a study included a call to action. The Cambridge Dictionary defines a call to action as “something as a speech, piece of writing, or act that asks for or encourages people to take action about a problem” (Call to action, 2022). To identify a call to action from general recommendations, we searched for command words, including but not limited to “should,” “must,” “we ask,” “we recommend,”, followed by a set of actions (i.e., “gardeners should reduce the frequency of lawn mowing and plant more flowers where possible”). Calls to action were coded as being one or more of these categories: “more flowers” called for more flowering plant species in urban gardens; “native flowers” specifically called for more native flowering plant species in urban green spaces; “more greenspace” called for more urban green space area; “exotic bees” specifically mentioned exotic bees as driving out native pollinators and called for fostering habitats to support native bees; “further research” called for more research on bees in urban gardens; “remnant vegetation” called for prioritizing remnant/native habitats in urban spaces, and “reduced disturbance” called for reduced mowing, soil disturbance, and/or pesticide and herbicide use.
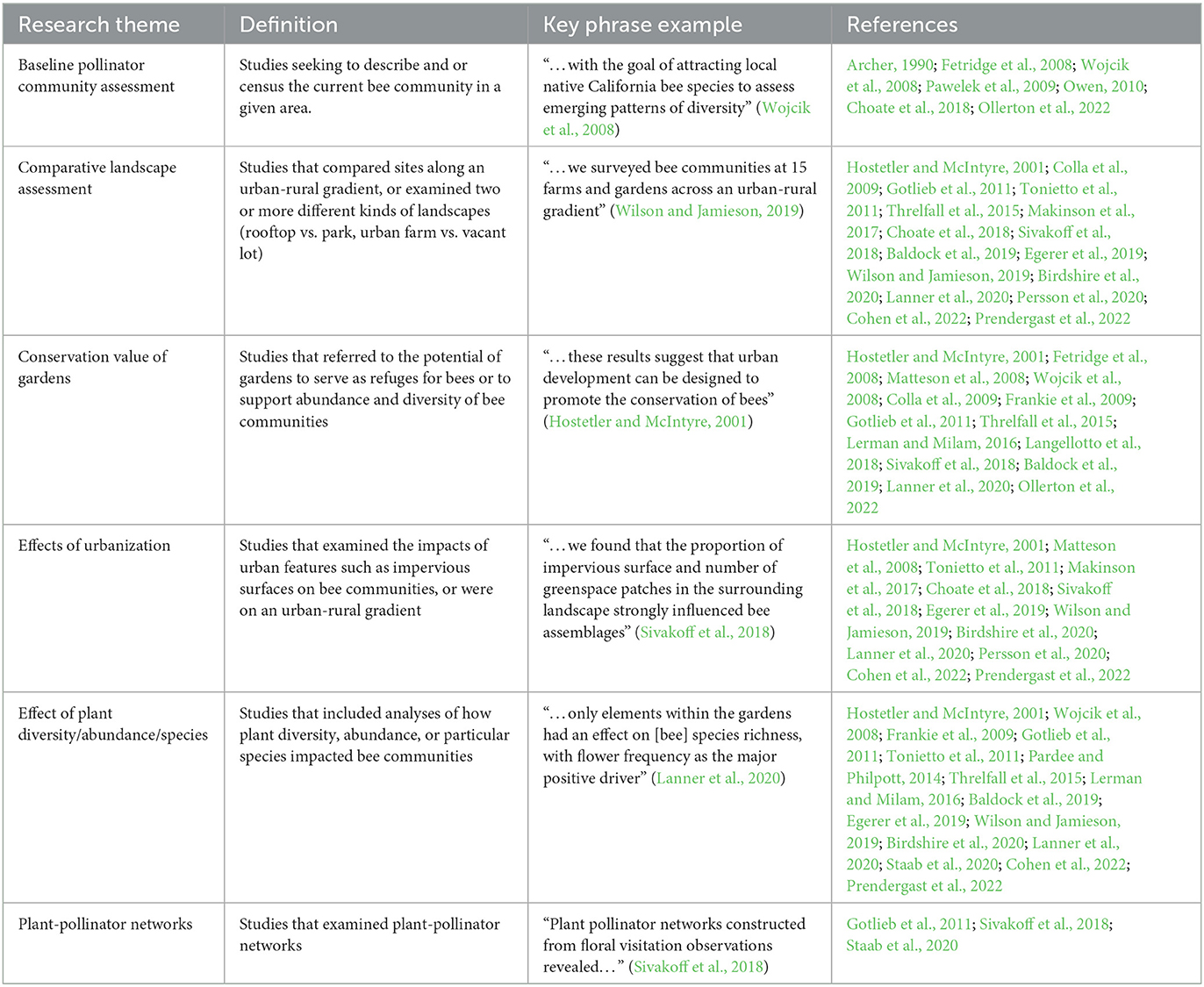
Table 2. Major research themes, definitions, and key-word examples emanating from the 28 studies included in this review.
To better understand the abundance, diversity, and types of bees that have been found in urban garden studies, we compiled, validated, and updated a master list of garden bee taxa reported across all papers (referred to in this paper as the garden bee metadataset). Validating and updating the master list involved correcting numerous misspellings, as well as updating genera and epithets to account for nomenclature changes and/or to correct synonyms. Examples include updating correct use of Lasioglossum heterognathum from Lasioglossum (Dialictus) heterognathus (Fetridge et al., 2008) and updating to the correct use of Pseudoanthidium nanum from Anthidium nanum (Lanner et al., 2020). Other examples include updating Afranthidium repetitum to Pseudoanthidium [Immanthidium] repetitum and updating Lasioglossum (Dialictus) mitchelli Gibbs to L. (Dialictus) hitchensi). In another case, Lasioglossum (Dialictus) tegulariforme (cited in Pawelek et al., 2009) was combined with L. helianthi due to likely misidentification after conferring with a taxonomic expert on the Lasioglossum genus (J. Gibbs, personal communication). One species, Lasioglossum (Sphecodogastra) leucopymatum, was removed from the dataset because its identification in Gotlieb et al. (2011) is unlikely in Israel; the native region of this species is not known to extend beyond Afghanistan and surrounding countries (Ebmer, 1995, as cited in Astafurova and Proshchalykin, 2020). These types of taxonomic updates and quality control checks help to ensure that future efforts to describe urban garden bee communities are as accurate and up to date as possible.
Bee taxon abundance and functional traits for each paper (if reported) were initially coded by N.B., J.H., M.M., and G.L. Entries were reviewed for accuracy by N.B. and J.H., and ultimately confirmed or corrected by J.A. To identify which bee species are relatively common in garden habitats (i.e., dominant garden bee species), we noted the number of studies where each bee species had been collected. In addition to noting frequency of species across studies, we also recorded relative abundance across studies, when available. We also recorded the geographic region and garden type where each study occurred. Species' functional traits were extracted from a publication (when listed) or were determined by cross-referencing bee taxa with species-specific ecological data embedded in Discover Life (Cane, 2003; Giles and Ascher, 2006; Lerman and Milam, 2016; Ascher and Pickering, 2020). We recorded the nesting substrate (e.g., soil, cavity, hive, wood excavator, aerial nest), sociality (e.g., eusocial, non-eusocial, subsocial, semi-social, parasitic), floral specificity (e.g., polylectic, oligolectic, or no pollen for parasitic species), and native status (e.g., native or exotic to the region where study was conducted) (Table 3). In instances where species' functional trait information was not available, we inferred traits from closely related congeneric species. Native status for bees found in North American studies was determined using Cane (2003) and Giles and Ascher (2006). Native status for bees found in studies outside of North America was sourced directly from studies or through Discover Life (Ascher and Pickering, 2020), and verified by J. A.
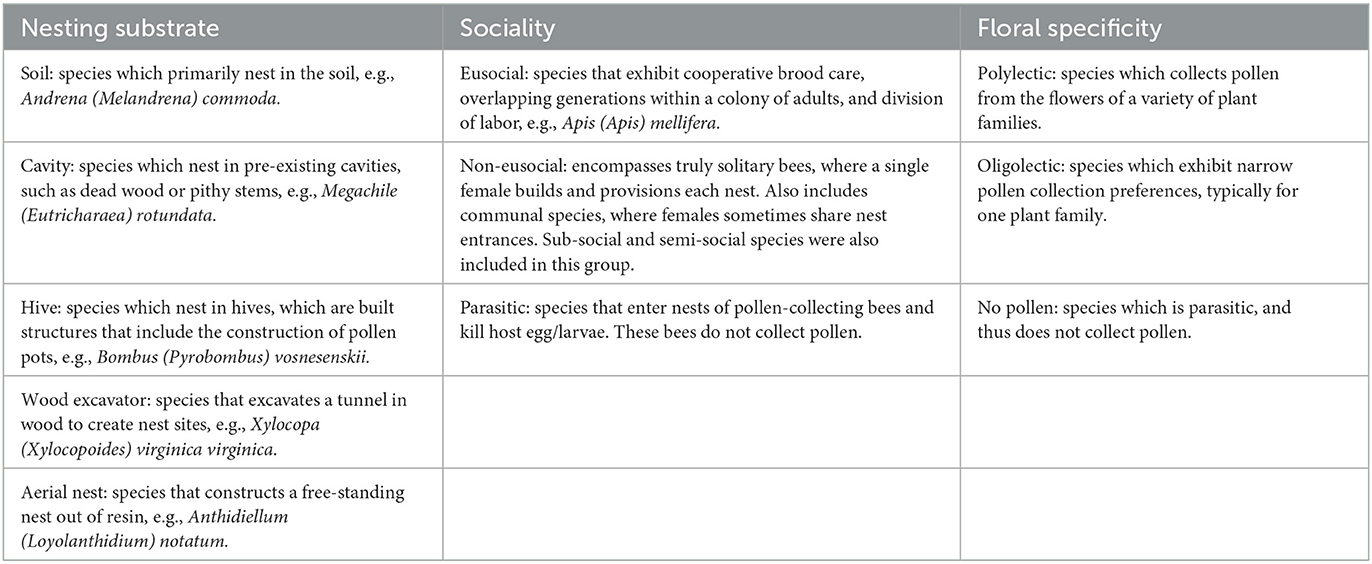
Table 3. Definitions used to code bees' ecological traits were coded for nesting substrate (soil, cavity, hive, wood excavator, or aerial nest), sociality (eusocial, non-eusocial, parasitic), and floral specificity (polylectic, oligolectic, no pollen).
We estimated the cumulative number of bee species collected from urban gardens, across all studies, in two ways. First, we generated a liberal estimate of the upper end of the range of species represented, by including specimens identified to the species level, the morphospecies level, and two species identified to the subspecies level. For example, Hylaeus (Hylaeus) mesillae, Hylaeus (Hylaeus) mesillae cressoni would count as two separate species, using this method. Likewise, Osmia (Osmia) lignaria lignaria and Osmia (Osmia) lignaria propinqua would account for two species in the liberal estimate. The conservative estimate was calculated by excluding specimens identified only to the morphospecies level (unless no species-level identifications for that genus were listed), and the compilation of the two species identified to the subspecies level. Species identified to the genus level were included in the analysis of bee functional traits and in the liberal estimate of species richness, but not in the conservative estimate of bee species richness.
Specimens identified as nr. (exact identification cannot be vertified; e.g., Megachile nr. relativa) or aff. (having affinities to a particular species, but may not be that species; e.g., Hylaeus aff. affinis) were not double counted. For example, Megachile nr. relativa and Megachile relativa were counted as one species. Those specimens that were identified as either/or species were not double counted, when that species was already represented in the dataset (e.g., Agapostemon angelicus/texanus, Agapostemon angelicus, and Agapostemon texanus were collectively counted as two species).
To better understand the types of bees that are common in urban gardens, we noted the five most abundant species in each paper. Bee abundance was not available for Frankie et al. (2009), Langellotto et al. (2018), Lanner et al. (2020), and Staab et al. (2020). Bee abundance was only sporadically reported in Ollerton et al. (2022), so it was not included in abundance counts. Pawelek et al. (2009) and Owen (2010) did not record the abundance of Apis mellifera, but honeybees were included in the top five most abundant species for these two papers, due to authors' noting that the species was highly abundant. Honeybees were excluded from analysis and abundance counts in Egerer et al. (2019) and Persson et al. (2020). The six most abundant species were included for Pardee and Philpott (2014), because two species had the same recorded abundance. Bees identified only to the genus level were not included in the bee abundance dataset. We performed the same functional trait analysis on the most abundant bees across papers as we did on the garden bee metadataset.
Summary statistics for the species functional traits were generated in R Studio (22.07.02) using the “dpylr” (Wickham et al., 2022) and “magrittr” (Bache et al., 2022) packages, and the “count” function to generate frequency counts, which were then used to manually calculate proportions within each of the functional trait categories.
Across the 27 articles and one book we used to compile our garden bee metadataset, 466 individual garden spaces were sampled, including 644 home gardens, 161 community gardens, 12 urban farms, and 9 rooftop gardens.
Most of the studies took place in the United States (n = 16). Other studies took place in Australia (n = 3), the United Kingdom (n = 3), and Canada, Germany, Austria, Sweden, and Israel (n = 1, each). All but four studies were conducted in the northern hemisphere (Threlfall et al., 2015; Makinson et al., 2017; Prendergast et al., 2022; sites in Ollerton et al., 2022; Figure 2). The most studied biome was temperate broadleaf and mixed forest (n = 16), followed by Mediterranean forests, woodlands, and scrubs (n = 7), temperate coniferous forest (n = 2), and temperate grassland, savannas, and shrubland (n = 2). Out of the 40 garden sites that met inclusion criteria in Ollerton et al. (2022) (see included sites in Supplementary Table 1), most were in Europe (United Kingdom, Ireland, Germany, Spain, Italy; n = 29 sites), though the United States (n = 4 sites), Australia (n = 3 sites), Brazil (n = 2 sites), Algeria (n = 1 site), and Mexico (n = 1 site) were also represented. The biomes represented in the included sites from Ollerton et al. (2022) were mostly temperate broadleaf and mixed forest (n = 32), though Mediterranean forest, woodlands and scrubs (n = 3), tropical and subtropical moist broadleaf forest (n = 2), temperate grasslands, savannas, and shrublands (n = 1), tropical and subtropical grasslands, savannas, and shrublands (n = 1), and temperate coniferous forest (n = 1) were also represented.
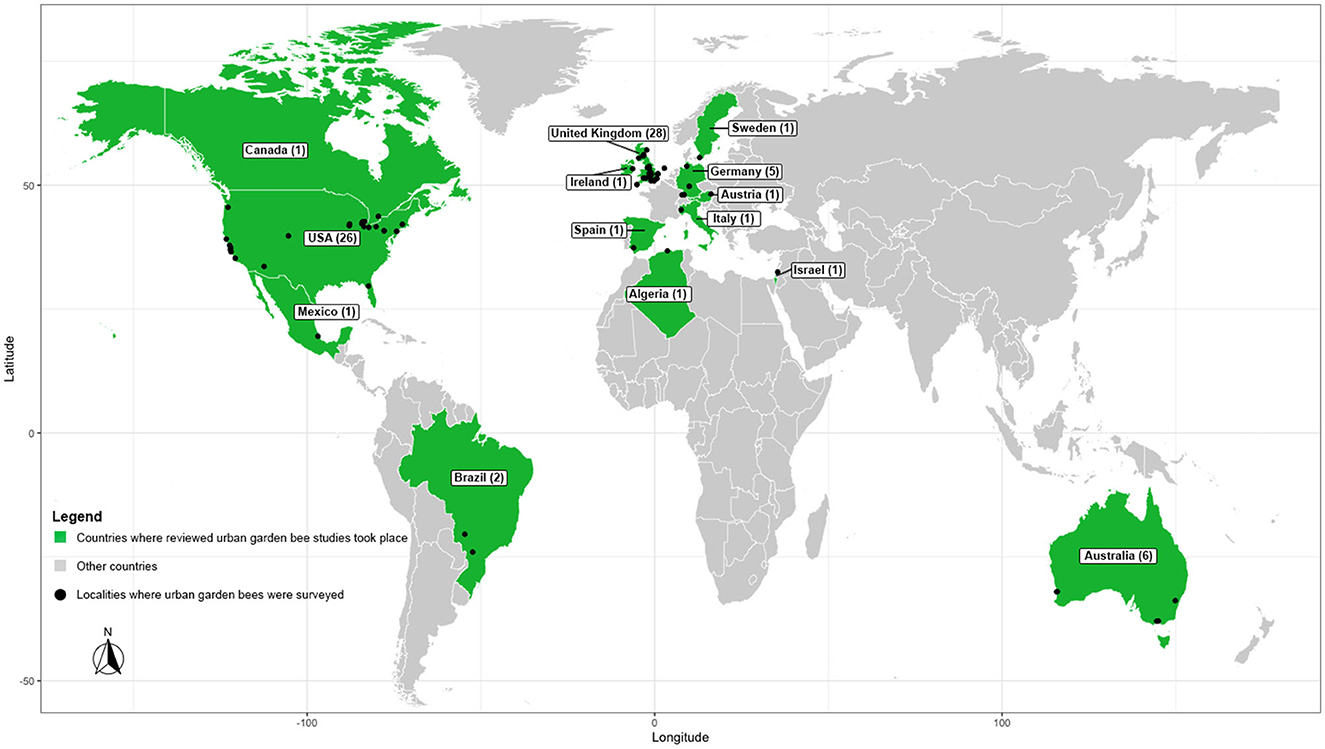
Figure 2. Map depicting where reviewed urban garden bee studies were conducted.
Netting was the most common method used to sample garden bees (n = 19), followed by pan traps (n = 17). Only six studies sampled bees by visual search. Six studies included other sampling methods, including hand collection with aspirators or collection jars. Most studies used either two sampling methods (n = 13), or one sampling method (n = 13), and three studies used three sampling methods.
Across the 27 studies that reported sampling period, the mean number of active sampling months was 16.1 months ± 35.8 (SD), whereas the median number of active sampling months was 6 months. The Owen (2010) study skewed the mean, since it took place over 30 years. Excluding the Owen study, the mean number of active sampling months was 9.8 months ± 15.3 (SD). Across the 24 studies that reported site size, the cumulative area of all study sites covered 2.9 km2. Mean cumulative area sampled was 20,991 m2 ± 32,527 m2 (median size 7,400 m2). Studies with multiple garden study sites, such as Langellotto et al. (2018) and Wilson and Jamieson (2019) skewed the mean, with cumulative areas sampled of over 100,000 m2 each.
The total number of bee species found across all urban garden studies was between 674 (excluding morphospecies) and 830 bee species (Supplementary Table 2). The mean number of species found per study was 63 ± 35.7 (median 57 species). Across all garden bee species, 18.6% were eusocial (n = 154), 64.9% were non-eusocial (n = 539), 13.3% were parasitic (n = 110), and 3.2% had unknown social behaviors (n = 27). Most nested in the soil (53.6%; n = 445), followed by cavity nesters (32.9%, n = 273), and species that nest in hives (5.8%, n = 48). The remaining species' nesting habits were unknown (6.1%, n = 51) or had other nesting habits (wood excavation or aerial nests; 1.6%, n = 13). Generalist foragers comprised 64.4% of bee species found in gardens (n = 535); specialist foragers comprised 13.3% of species found (n = 110). Other species were parasitic, and do not forage for pollen (13.3%, n = 110), or their diet breadth was unknown (9.0%, n = 75) (Figure 3). The most abundant bee family represented was Halictidae (31.3%, n = 260 species), followed by Megachilidae (22.5%, n = 187 species) and Apidae (21.6%, n = 179 species) (Figure 4). The families Colletidae (12.7%, n = 105 species), Andrenidae (11.1%, n = 92 species), and Melittidae (0.80%, n = 7 species) were also represented. Across all bees, with 16 species duplicated due to differing native/exotic status depending on region, only 2.9% of bee species were exotic to the region studied (n = 24 species). Most garden bee species (92.5%) were native (n = 768 species). The native status of the remaining 4.6% was unknown (n = 38 species). We identified the dominant garden bee species across studies by recording the number of papers in which a particular species was recorded. Across all studies, the five most frequently reported bee species were Apis (Apis) mellifera (n = 20 studies), Halictus (Odontalictus) ligatus (n = 17 studies), Anthidium (Anthidium) manicatum (n = 17 studies), Megachile (Eutricharaea) rotundata (n = 16 studies), and Halictus (Protohalictus) rubicundus (n = 15 studies). Although eusocial, non-native species were a minority in the full dataset, the most dominant garden bees were eusocial (60%, n = 3) and non-native to the region where they were studied (60%, n = 3). All five species were polylectic, but represented three different nesting strategies: soil (40%, n = 2), cavity (40%, n = 2), and hive (20%, n = 1).
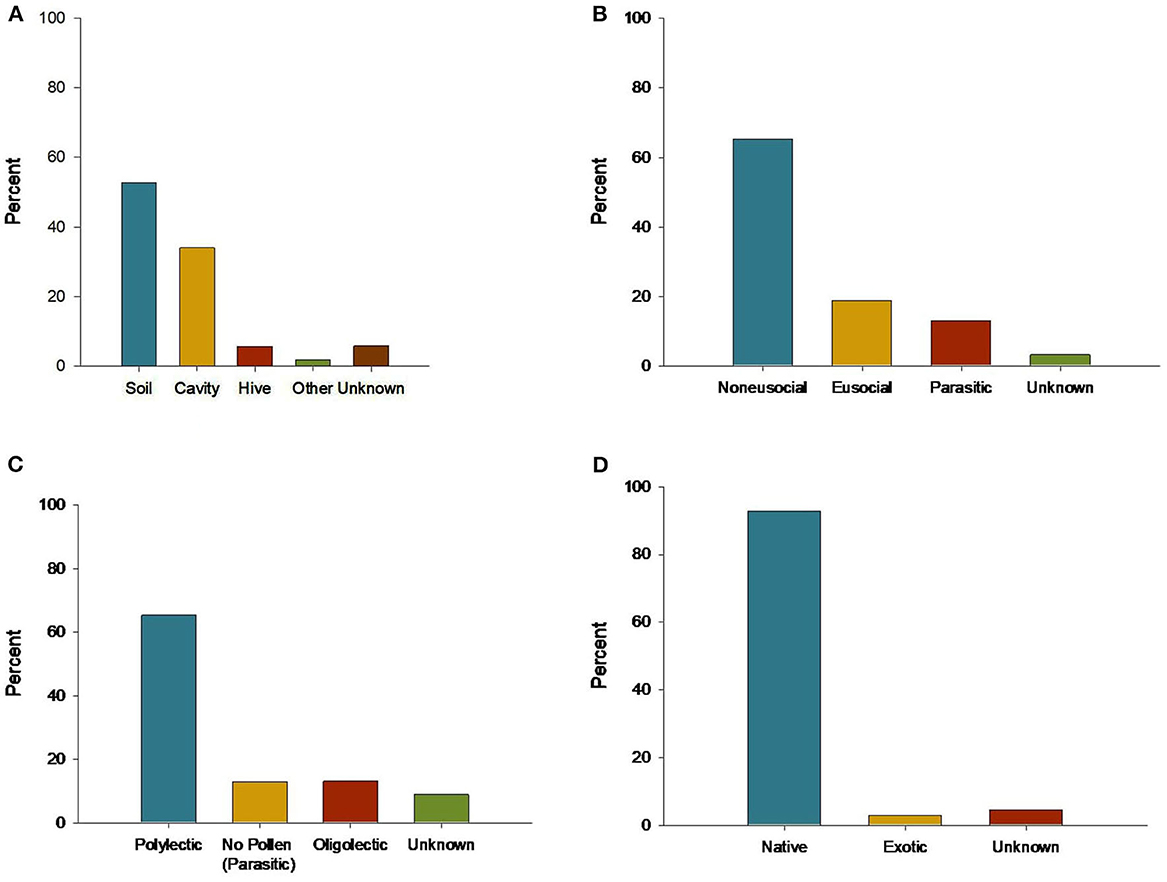
Figure 3. Distribution of functional traits across urban garden bee taxa: (A) nest type, (B) sociality, (C) floral specificity, and (D) native status.
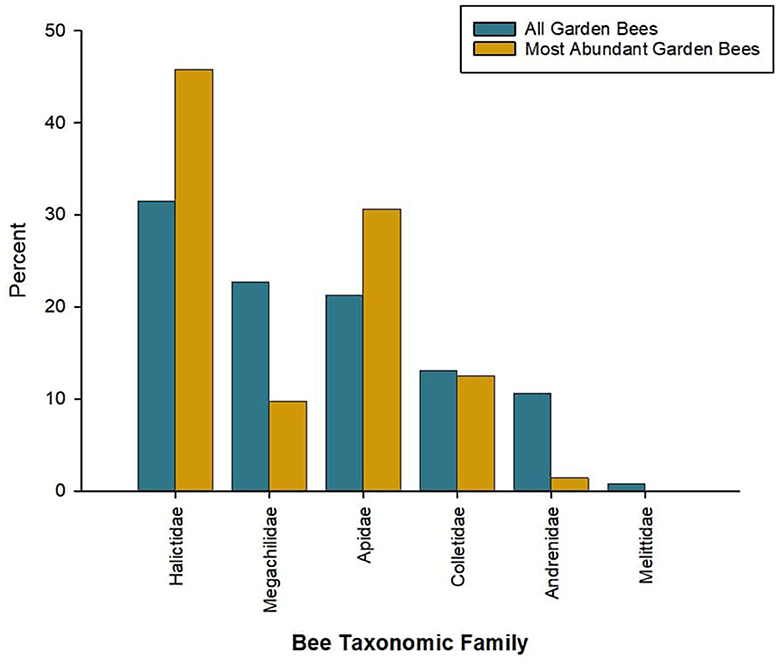
Figure 4. Percent of bee taxonomic family distribution for most abundant taxa collected from urban gardens, for all taxa collected from urban gardens, and globally. Abundant taxa were determined by selecting the five most abundant bee species from each study that reported abundance.
When looking at the 5 (or 6, for Pardee and Philpott, 2014) most abundant bee species within each study where abundance was reported (n = 23 papers), there were 73 species after the removal of duplicates (Supplementary Table 3). Across entries, 31.5% were eusocial (n = 23 species), 65.8% were non-eusocial (n = 48 species), and 2.7% had unknown social behaviors (n = 2 species). Most nested in the soil (53.4%, n = 39 species), followed by cavity nesters (31.5%, n = 23 species), and species that live in hives (12.3%, n = 9 species). The remaining species were wood excavating (1.4%, n = 1 species) or had unknown nesting habits (1.4%, n = 1 species). Most species were generalists (94.5%, n = 69 species), with only 4.1% of the most abundant species being specialist foragers (n = 3 species), and one species' diet was unknown (1.4%, n = 1 species). Most abundant garden bees were native (91.8%, n = 67 species), with 8.2% of abundant bee species being exotic to the region in which they were studied (n = 6 species). The most abundant bee family represented was Halictidae (46.6%, n = 34 species), followed by Apidae (30.1%, n = 22 species. The families Colletidae (12.3%, n = 9 species), Megachilidae (9.6%, n = 7 species), and Andrenidae (1.4%, n = 1 species) were also represented.
The most prevalent research themes from garden bee studies included investigations of the effects of plant diversity/abundance/species on bees (n = 16) and comparative landscape assessments (n = 16), followed by studies of the conservation value of gardens (n = 14), and the effects of urbanization on bees (n = 13). Other research themes included baseline pollinator community assessments (n = 8) and plant-pollinator networks (n = 3). While 13 of the 14 studies that included the conservation value of gardens as a major theme concluded that they are valuable conservation sites, Gotlieb et al. (2011) asserted that gardens do not promote species richness compared to more natural areas.
We identified 15 papers that included calls to action in the reviewed literature, with several papers containing more than one (Figure 5). The most common calls to action were for gardeners to plant more flowering plant species in gardens (n = 10), for gardeners to include more native flowering plant species in gardens (n = 8), advocating for more urban green space (n = 6), and for scientists to conduct further research (n = 5). Other calls for actions included suggesting gardeners reduce habitat disturbance (n = 3), leave remnant vegetation where possible (n = 2), and a call for habitat to specifically support native bees, rather than exotic bees (n = 1).
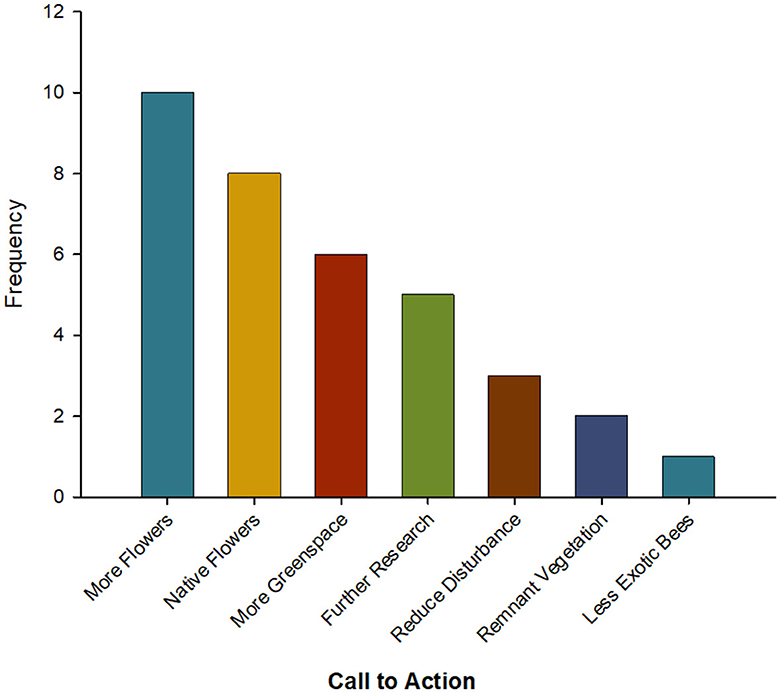
Figure 5. Proportion of various calls to action included in garden bee studies. Calls to action included “More Flowers” or “Native Flowers” (in urban gardens); “More Greenspace” (in urban landscapes), “Exotic Bees” (noted the negative impact of exotic bees on native pollinators and called for fostering habitats to support native bees); “Further Research” (on bees in urban gardens,) “Remnant Vegetation” (called for prioritizing remnant/native habitats in urban spaces); and “reduced disturbance” (including mowing, soil disturbance, and/or pesticide and herbicide use).
Between 674 (conservative estimate) and 830 (liberal estimate) bee species have been collected from urban garden habitats included in our review, representing six of the seven extant bee families. These bees have been identified from a relatively small number of studies, biased to the northern hemisphere. Bees from the family Stenotritidae were not represented. This is not surprising since this family is comprised of 21 species isolated to Australia (Danforth et al., 2019), although we did include four studies with sites located in Australia in our review (Threlfall et al., 2015; Makinson et al., 2017; Ollerton et al., 2022; Prendergast et al., 2022). While there are estimated to be over 20,000 bee species worldwide (Danforth et al., 2019; Orr et al., 2021), urban areas can present harsh conditions for many bees (Cardoso and Gonçalves, 2018), including heat stress (Hamblin et al., 2017), homogenization of forage plants (Groffman et al., 2014), increased landscape disturbance (Threlfall et al., 2015; Lerman and Milam, 2016), competition from exotic species (LeCroy et al., 2020), and a decrease in forage and nesting site availability (Bates et al., 2011; Choate et al., 2018; Birdshire et al., 2020; Lanner et al., 2020). This can result in significant declines in pollinator abundance and species richness, when compared with more rural sites (Bates et al., 2011; Birdshire et al., 2020; Millard et al., 2021), although this is not always the case (Kearns and Oliveras, 2009; Baldock et al., 2015; Sirohi et al., 2015).
Across all studies in the metadataset for which the total area sampled was available, ~2.9 km2 of cumulative garden area was sampled. Small garden spaces can, in fact, host bee communities that are representative of bee communities found in larger, more intact habitat (Fetridge et al., 2008; Kearns and Oliveras, 2009). This suggests the potential conservation value of urban gardens to bees, particularly if gardeners emphasize the availability of forage plants (Tommasi et al., 2004; Matteson and Langellotto, 2010; Burdine and McCluney, 2019; Lanner et al., 2020) and nesting resources (Cane, 2001; Tonietto et al., 2011).
Across all garden bee species documented in this review, 64.4% were polylectic (generalist foragers). This aligns with general estimates of bee foraging habits in the United States, where between 65 and 75% of bee species are estimated to be polylectic (Fowler, 2020a,b; Fowler and Droege, 2020). That oligolectic, specialist foragers made up 13.3% of the bees identified from garden study sites, suggests that gardens can support the specialized life history requirements of some bee species, which could be an area to build upon for continued urban bee conservation efforts. Fowler (2016) emphasizes that strategies to conserve pollinator populations should specifically target specialist species.
It is important to note that because the metadataset is biased to the northern hemisphere, the data compiled for the most abundant garden bee species is skewed toward North America and Europe. All the dominant garden bees documented in this review were generalist foragers. Three specialist bees were present, however, when considering the 5–6 most common garden bee species within each urban garden study. These abundant specialists included Colletes daviesanus, Megachile (Pseudomegachile) aff. flavipes, and Melissodes (Eumelissodes) robustior. C. daviesanus and M. robustior both specialize on plants in the Asteraceae (Müller and Kuhlmann, 2008; Fowler, 2020b). Many of the specialist bees found in urban gardens, such as bees from the family Andrenidae and bees from the genus Melissodes, also specialize on plants in the Asteraceae (Cameron et al., 1996; Michez et al., 2008). In a study of bee associations with native plants, Douglas' aster (Symphyotrichum subspicatum) was observed to support 19 different bee species, and estimated to support up to 74 bee species (Anderson et al., 2022). Though many bees specialize on Asteraceae, host plant specialization is not limited to this one botanical family (Larkin et al., 2008), suggesting that a broad representation of plant families in a garden may be best suited to supporting oligolectic species.
While some studies have found a high richness and abundance of exotic bee species in urban gardens (Matteson et al., 2008; Gruver and CaraDonna, 2021), we found a relatively low number of exotic species (n = 24, or 3%) across our metadataset. To date, the proportion of exotic species remains low in urban garden systems, though some exotic bee species are numerically abundant and dominant components in urban gardens. It is important to note, though, that the percent of exotic bee species increased as we examined the most abundant bees in urban garden studies (8.3%) and the dominant bees across all studies (60%), compared to just 3% of garden bee species in our metadataset, suggesting that exotic species are disproportionately benefitting from urbanization (Fitch et al., 2019). The most common exotic species (though specimen abundance was not reported for every paper) were Apis (Apis) mellifera (n = 3,206 specimens), Hylaeus (Spatulariella) hyalinatus (n = 207 specimens), and Hylaeus (Hylaeus) leptocephalus (n = 195 specimens).
Urban gardens also support a relatively high number of parasitic bee species (n = 110, 13.3% of urban garden bee species found in our review), which is reflective of estimated proportions of bee social parasites in North America (15%; Bohart, 1970). No parasitic bee species were represented when we examined the most abundant species in urban gardens. Parasitic bees (kleptoparasites) can act as indicator species for bee communities, because they respond to disturbances in a manner that is reflective of the entire bee community (Sheffield et al., 2013). As with specialist foragers, the relatively high proportion of parasitic bees collected from garden studies suggests that gardens can support the specialized life history requirements of at least some bee species.
Though floral resources are often emphasized as being predictive of pollinator abundance in urban spaces (Matteson and Langellotto, 2010; Plascencia and Philpott, 2017; Hyjazie and Sargent, 2022), less attention has been given to the importance of nest sites. Nest resources are particularly important for smaller-bodied bees, as body size can be predictive of foraging range (reviewed in Greenleaf et al., 2007). The existence of nest sites or nesting resources in gardens, then, may influence what bee species are able to persist in urban spaces. We found the percentage of cavity nesting bees in this metadataset relatively low (32.9% of species) in contrast with those of previous studies and reviews that have examined urban bee communities across a broad range of habitats, and have found cavity nesters to be dominant in urban environments (reviewed in Buchholz and Egerer, 2020; Ayers and Rehan, 2021; Fauviau et al., 2022). In contrast, soil nesting bees, the most common nesting strategy of all solitary bees (Danforth et al., 2019; Antoine and Forrest, 2021), were relatively abundant in urban gardens (53.6% of species), though ground nesting bees are estimated to represent between 65 and 70% of all bees (Danforth et al., 2019; Sgolastra et al., 2019). Opportunities to enhance nesting resources in gardens to support the abundance of wild bee species with varying nesting strategies including provisioning patches of bare soil (Cunningham-Minnick et al., 2019), and woody additions, such as small logs (Pawelek et al., 2015).
We found that bees from the family Halictidae were somewhat overrepresented (31.3% of bee species) in our metadataset compared to expected global representation of ~22% (Danforth et al., 2019). Others have found that urban bee assemblages are dominated by Halictidae (in particular Halictinae; Fortel et al., 2014; Geslin et al., 2016; Villalta et al., 2021). This may be in part explained by the bias of pan-traps toward smaller bees (Cane, 2001; Portman et al., 2020), given that 17 studies sampled with pan-traps. Another explanation could be the tendency for eusocial bees to dominate urban settings (Zanette et al., 2005) due to social traits enhancing the spread and competitiveness of certain species (Chapman and Bourke, 2001). Of all the halictids found in urban gardens, 41.1% were eusocial, 35.9% were non-eusocial, 13.0% were parasitic, with the social structure of 10.0% of the halictids unknown. Bees from the family Andrenidae are underrepresented in the dataset (11.1% of bee species) compared to global expected proportions of 15% (Danforth et al., 2019), particularly when we examine the most abundant and dominant bees in urban settings. Because most andrenids (in particular, the majority of Andrena species) are spring-flying bees, their underrepresentation could be related to sampling periods focusing more on summer months, or due to lack of spring-flowering forage in gardens (Matteson et al., 2008). Global totals for Andrenidae are also enhanced by a uniquely large radiation of perditine (genera Perdita and Macrotera) and protandrenine (Protandrena sensu lato) in deserts and of Andrena in Mediterranean areas (Wood, 2021; Bossert et al., 2022). All but four of the garden bee studies included in this review were from different regions and/or biomes less favorable to this family (Table 1). Bees within the Apidae were among the most abundant bees found in urban gardens. For example, the European honeybee (Apis mellifera), was documented as one of the most abundant species in seven papers and held exotic status in all of them. When honeybees are present, they may have negative impacts on native bee communities, including depletion of nectar and pollen resources (Carneiro and Martins, 2012), which particularly puts pressure on oligolectic species (Cane and Tepedino, 2017).
Urban gardens are often dominated by ornamentally modified and exotic plant species (Threlfall et al., 2016), and the impact of exotic plant species on native insect species is varied (Sunny et al., 2015). While generalist bees are more likely to forage on invasive or non-native plant species than specialists (Lopezaraiza-Mikel et al., 2007; Tepedino et al., 2008), there is abundant evidence to support generalist bees' preference for native plant species (Williams et al., 2011; Morandin and Kremen, 2013; Pardee and Philpott, 2014; Salisbury et al., 2015; Anderson et al., 2022), suggesting that even generalist bees may be facultative specialists (Synge, 1947; Percival, 1974; Wills et al., 1990). Preserving and planting native flowering plant species as a means to sustain wild bee communities was specifically recommended by five of the studies in our dataset (Table 1; Figure 5). In a recent study of bee associations with native and non-native garden plants, Anderson et al. (2022) documented significant associations between several native bees known to be polylectic (including Halictus ligatus, Halictus tripartitus, Bombus caliginosus) and specific native plants (Symphyootrichum subspicatum, Eschscholzia californica, and Phacelia heterophylla, respectively) even when bee-attractive, non-native garden plants were nearby. This suggests that generalist bees may prioritize foraging from certain native plants, perhaps to meet nutritional needs (Roulston et al., 2000; Wood et al., 2018) and/or to take advantage of efficient foraging opportunities (Williams et al., 2011). Despite the potential benefits of native plant species to garden bees, there is a general lack of concordance between the native plants that bees found most attractive, and the ones gardeners found most attractive (Anderson et al., 2022), with some of the top plants for bees described by gardeners as being “weedy” or “unattractive” (Anderson et al., 2021). Fortunately, gardeners' perception of native plants can significantly improve when short messages are shared regarding a plant's value to native bees (Anderson et al., 2021), highlighting the value of informal outreach and education efforts.
Beyond gardeners' perceptions, changing a garden's vegetative composition to include more native plants and other bee-friendly practices does not come without barriers. Many home gardens, particularly in the United States, are regulated entities, and municipal ordinances can limit the height of grasses, the presence of “weedy” looking species, and woody debris (Larson et al., 2020). Gardens come with their own sets of social norms that prioritize a tidy aesthetic that may require synthetic chemical inputs, and/or reduce bee nest site availability (Nassauer et al., 2009; Locke et al., 2018). Studies have also reviewed the potential benefits of adding “cues to care” (e.g., fences and tidy paths, bright flowers) in urban gardens, which imply the presence of a garden caretaker, thus creating a more ecologically-minded space that may appease societal, and sometimes municipal, expectations (Nassauer, 1995; Li and Nassauer, 2020).
Beyond any nutritional advantages that native plants may confer to native bees, increasing their planting in urban garden spaces might reduce exploitative and/or interference competition with exotic bees (Stout and Morales, 2009). For example, even though plants were cultivated at a common field site, non-native honeybees were much more abundant on non-native plants (e.g., “Grosso” lavender, Lavandula x intermedia “Grosso”; oregano, Origanum vulgare; and catnip, Nepeta cataria) than on native plants highly attractive to native bees (e.g., globe gilia, Gilia capitata; Douglas' aster, Symphyotrichum subspicatum; yarrow, Achillea millefolium; California poppy, Eschscholzia californica; and Oregon sunshine, Eriophyllum lanatum) (Anderson et al., 2022). This suggests that intermixing non-native with native plants in garden spaces might facilitate niche-partitioning and co-existence between non-native and efficient foragers, such as honeybees, and the native bee community (Comba et al., 1999; Salisbury et al., 2015; Pei et al., 2023).
The studies included in this review were biased to the northern hemisphere. Most study sites were located at mid-latitudes, which host the highest levels of bee biodiversity (Orr et al., 2021), and most studies were also located in either temperate or xeric regions, which are also hotspots of bee diversity (Cheng and Ashton, 2021; Orr et al., 2021). Studies are underway in regions not represented in this analysis, but they may not yet be published (Hui, 2021), did not meet inclusion criteria (Wen et al., 2013), or may have been filtered out of our search, since search terms were exclusively in English. No studies from the southern hemisphere were excluded solely for identifying fewer than 50% of specimens to the species level. Instead, studies were screened out because they did not occur in urban gardens (Sing et al., 2016; Stewart et al., 2018). Nonetheless, the lack of studies from the southern hemisphere, particularly Africa (De Palma et al., 2016), and less studied regions of the northern hemisphere, such as Asia (De Palma et al., 2016), represents a huge gap in our understanding of garden bee communities. Some taxonomic biases, such as the relative scarcity of Colletidae, may also reflect geographic biases, since this family is most species-rich in Australia and in temperate South America.
As our review and other studies have shown, urban bee data (including garden bee data), is centered around the northern hemisphere, especially the United States and Europe (De Palma et al., 2016; Brant et al., 2022). Although this is a recognized deficiency, it is important to note that this has been an identified area of concern in bee ecology for at least 20 years (Liow et al., 2001; Hernandez et al., 2009; Buchholz and Egerer, 2020; Shackleton et al., 2021; Prendergast et al., 2022). The rate of urbanization is increasing globally (United Nations, 2018), particularly in developing regions [United Nations Population Fund (UNFPA), 2007]. We know that urbanization leads to large-scale habitat loss and fragmentation (Morse et al., 2003; Miller et al., 2014; Baldock et al., 2019), and percent impervious surface cover is associated with declines in species richness (Choate et al., 2018; Burdine and McCluney, 2019; Birdshire et al., 2020), but bee species richness and abundance in urban areas is highly trait- and scale-dependent (Archer, 1990; Wenzel et al., 2020). City gardens have the potential to be a refuge for wild bees (Tommasi et al., 2004; Matteson et al., 2008; Lowenstein et al., 2014; Baldock et al., 2019; Hall and Martins, 2020), to provide important social benefits (Dunnett and Qasim, 2000), and to fulfill socio-cultural needs (Sturiale et al., 2020), creating a synergistic effect between social and ecological benefits (Dennis and James, 2017). Understanding urban garden bee communities in the southern hemisphere and other understudied regions, such as Asia, should be prioritized, to create more context- and region-specific recommendations for gardeners.
Urban garden bee research spans decades, and recommendations to create standardized sampling methods and conservation opportunities date back nearly as far (Cane et al., 2000; Cane, 2001; Frankie et al., 2009; Williams et al., 2011; Buchholz and Egerer, 2020; Wenzel et al., 2020). While standardized sampling methods have been developed for monitoring bee populations (LeBuhn et al., 2003; Droege et al., 2016), these recommendations are not amenable to urban garden habitats. Specifically, established protocols require long transects and/or large sampling spaces (e.g., 1 hectare), which are unrealistic, given the heterogeneity and relatively small size of most urban garden habitats. Studies included in this review had a broad range in active sampling months (1–182 mo), total area sampled (59–117, 119 m2), and the number of sampling methods employed (1–3 methods). There was a lack of standardized sampling across studies, with a total of seven different methods used. Because pan-trapping is known to be biased toward smaller bees and tends to miss specialist bees (Cane, 2001), supplementing pan-traps with aerial netting can provide better insight into an area's true species richness (Williams et al., 2011). Previous reviews of urban garden bee ecology also recommend that studies have temporal depth, or span over many seasons, to account for the variances in bee community structure over multiple years (Cane, 2001; Williams et al., 2011). Although the studies we examined display a wide range in sampling methods, garden bee sampling is complicated by needing access to dispersed, and often private or gated, parcels of land. Even if sampling access is granted for one season, it may be difficult to maintain access for multiple years, thus complicating the fulfillment a multi-year study. An example where access was not an issue, and thus there were 182 active sampling months, was Owen (2010). The study took place in her own backyard, leading to a 30-year dataset.
Based on the results of this review, we make the following recommendations that may benefit future urban garden bee studies:
1. Create standardized sampling methods for gardens. Although standardized sampling methods exist for bee communities (e.g., LeBuhn et al., 2003; Droege et al., 2016), they are not amenable to garden habitats. Consistency in sampling across studies would allow for comparison across studies, as well as comparisons across time.
2. Researchers should work with, and advocate for, collaboration with taxonomists. Collaboration is necessary to avoid identification mistakes (e.g., Halder et al., 2013; Del Toro and Ribbons, 2020), which can delay or misguide our understanding of bee communities. In addition, training taxonomists should be a priority to account for identification demands (Drew, 2011).
3. Prioritize and fund research of urban bee communities in the southern hemisphere and understudied regions of the northern hemisphere. This is of particular importance because the rate of urbanization is high in developing countries, amplifying pressures on bee populations.
The studies included in this review represent data collected over the past five decades. As we move into a sixth decade of extensive garden bee studies amidst massive global change events (e.g., urbanization, climate change), it becomes more important than ever to create and tend urban spaces that yield multiple benefits. Gardens provide important social benefits (Dunnett and Qasim, 2000) and fulfillment of socio-cultural needs (Sturiale et al., 2020), while also providing habitat for a diversity of wildlife (e.g., Owen, 2010; Marzluff, 2015; Hall and Martins, 2020) and urban plants (Doody et al., 2014). Thus, gardens are somewhat uniquely positioned for creating a synergistic effect between social and ecological benefits (Dennis and James, 2017). We hope that the metadataset we compiled, as well as our associated summary of key findings and current research gaps, might be useful to current and future urban ecologists who study urban garden spaces.
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
NB: Conceptualization, Data curation, Formal analysis, Methodology, Writing—original draft, Writing—review & editing. JA: Data curation, Validation, Writing—review & editing. JH: Data curation, Writing—review & editing. MM: Data curation, Writing—review & editing. GL: Conceptualization, Data curation, Funding acquisition, Methodology, Supervision, Writing—review & editing.
Funding for this study was provided by an Oregon State University (OSU) Department of Horticulture ARCO-Swallow Fellowship to NB, and contributions from Y. Sherry Sheng and Spike Wadsworth in support of the OSU Garden Ecology Lab.
We thank all of the authors and researchers whose work formed the basis of this review. We would also like to acknowledge all of the authors who reached out to share additional information about their research with us: Caragh Threlfall, Nancy McIntyre, Stacy Philpott, Gabriella Pardee, Rebecca Tonietto, Michael Staab, Kristen Birdshire, Katherine Baldock, Adrian Carper, and Gordon Frankie.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/frsc.2023.1102360/full#supplementary-material
Supplementary Table 1. Site information from Ollerton et al. (2022).
Supplementary Table 2. List of bee taxa found across studies in our metadataset, including bee functional trait data.
Supplementary Table 3. List of the most abundant bee taxa found across studies that reported abundance in our metadataset, including bee functional trait data.
Anderson, A. G., Costner, L., Best, L., and Langellotto, G. A. (2022). The bee fauna associated with Pacific Northwest (USA) native plants for gardens. Conserv. Sci. Pract. 4, e12801. doi: 10.1111/csp2.12801
Anderson, A. G., Messer, I., and Langellotto, G. A. (2021). Gardeners' perceptions of northwestern U.S. native plants are influenced by ecological information and garden grou15gp affiliation. HortTechology 31, 458–469. doi: 10.21273/HORTTECH04770-20
Antoine, C. M., and Forrest, J. R. K. (2021). Nesting habitat of ground-nesting bees: a review. Ecol. Entomol. 46, 143–159. doi: 10.1111/een.12986
Archer, M. E. (1990). The solitary aculeate wasps and bees (Hymenoptera: Aculeata) of an English suburban garden. Entomol. Gazette 41, 129–142.
Ascher, J. S., and Pickering, J. (2020). Discover Life Bee Species Guide and World Checklist (Hymenoptera: Apoidea: Anthophila). Available online at: http://www.discoverlife.org/mp/20q?guide=Apoidea_species (accessed November 15, 2022).
Astafurova, Y. V., and Proshchalykin, M. Y. (2020). The bees of the family Halictidae (Hymenoptera) described by Ferdinand Morawitz from the collection of Aleksey Fedtschenko. Zookeys 994, 35–104. doi: 10.3897/zookeys.994.58441.figure35
Ayers, A. C., and Rehan, S. M. (2021). Supporting bees in cities: how bees are influenced by local and landscape features. Insects 12, 128. doi: 10.3390/insects12020128
Bache, M. S., Wickham, H, and Henry, L. (2022). Magrittr: A Forward-Pipe Operator for R. Available online at: https://cran.r-project.org/web/packages/magrittr/magrittr.pdf (accessed June 1, 2023).
Baldock, K. C. R., Goddard, M. A., Hicks, D. M., Kunin, W. E., Mitschunas, N., Morse, H., et al. (2019). A systems approach reveals urban pollinator hotspots and conservation opportunities. Nat. Ecol. Evol. 3, 363–373. doi: 10.1038/s41559-018-0769-y
Baldock, K. C. R., Goddard, M. A., Hicks, D. M., Kunin, W. E., Mitschunas, N., Osgathorpe, L. M., et al. (2015). Where is the UK's pollinator biodiversity? The importance of urban areas for flower-visiting insects. Proc. R. Soc. Biol. Sci. 282, 20142849. doi: 10.1098/rspb.2014.2849
Bartomeous, I., Ascher, J., Gibbs, J., Danforth, B., Wagner, D., Hedtke, S., et al. (2013). Historical changes in northeastern US bee pollinators related to shared ecological traits. Proc. Nat. Acad. Sci. U. S. A. 110, 4656–4660. doi: 10.1073/pnas.1218503110
Bates, A. J., Sadler, J. P., Fairbrass, A. J., Falk, S. J., Hale, J. D., and Matthews, T. J. (2011). Changing bee and hoverfly pollinator assemblages along an urban-rural gradient. PLoS ONE 6, e23459. doi: 10.1371/journal.pone.0023459
Biesmeijer, J. S., Roberts, S. P. M., Reemer, M., Ohlemüller, R., Edwards, M., Peeters, T., et al. (2006). Parallel declines in pollinators and insect-pollinated plants in Britain and the Netherlands. Science 313, 351–354. doi: 10.1126/science.1127863
Birdshire, K. R., Carper, A. L., and Briles, C. E. (2020). Bee community response to local and landscape factors along an urban-rural gradient. Urban Ecosyst. 23, 689–702. doi: 10.1007/s11252-020-00956-w
Bohart, G. E. (1970). The Evolution of Parasitism Among Bees. 40. Faculty Honor Lectures. Paper 18. Available online at: https://digitalcommons.usu.edu/honor_lectures/18 (accessed November 15, 2022).
Bossert, S., Wood, T. J., Patiny, S., Michez, D., Almeida, E. A. B., Minckley, R. L., et al. (2022). Phylogeny, biogeography and diversification of the mining bee family Andrenidae'. Syst. Entomol. 47, 283–302. doi: 10.1111/syen.12530
Brant, R. A., Arduser, M., and Dunlap, A. S. (2022). There must bee a better way: a review of published urban bee literature and suggested topics for future study. Landsc. Urban Plan. 226, 104513. doi: 10.1016/j.landurbplan.2022.104513
Buchholz, S., and Egerer, M. H. (2020). Functional ecology of wild bees in cities: towards a better understanding of trait-urbanization relationships. Biodivers. Conserv. 29, 2779–2801. doi: 10.1007/s10531-020-02003-8
Burdine, J., and McCluney, K. (2019). Interactive effects of urbanization and local habitat characteristics influence bee communities and flower visitation rates. Oecologia 190, 715–723. doi: 10.1007/s00442-019-04416-x
Call to action (2022). In Cambridge English Dictionary: Definitions & Meanings [WWW Document], 2023. Available online at: https://dictionary.cambridge.org/us/dictionary/english/ (accessed July 6, 2022).
Cameron, S. A., Whitfield, J. B., Hulslander, C. L., Cresko, W. A., Isenberg, S. B., and King, R. W. (1996). Nesting biology and foraging patterns of the solitary bee Melissodes rustica (Hymenoptera: Apidae) in northwest Arkansas. J. Kans. Entomol. Soc. 69, 260–273.
Cane, J. (2003). Exotic Non-social Bees (Hymenoptera: Apoidea) in North America: Ecological Implications. For Non-native Crops, Whence Pollinators for the Future? Thomas Say Publications, Entomological Society of America. 113–126.
Cane, J. H. (2001). Habitat fragmentation and native bees: a premature verdict? Conserv. Ecol. 5, 3. doi: 10.5751/ES-00265-050103
Cane, J. H., Minckley, R. L., and Kervin, L. J. (2000). Sampling bees (Hymenoptera: Apiformes) for pollinator community studies: pitfalls of pan-trapping. J. Kansas Entomol. Soc. 73, 225–231.
Cane, J. H., and Tepedino, V. J. (2017). Gauging the effect of honey bee pollen collection on native bee communities. Conserv. Lett. 10, 205–210. doi: 10.1111/conl.12263
Cardoso, M. C., and Gonçalves, R. B. (2018). Reduction by half: the impact on bees of 34 years of urbanization. Urban Ecosyst. 21, 943–949. doi: 10.1007/s11252-018-0773-7
Carneiro, L. T., and Martins, C. F. (2012). Africanized honey bees pollinate and preempt the pollen of Spondias mombin (Anacardiaceae) flowers. Apidologie 43, 474–486. doi: 10.1007/s13592-011-0116-7
Chapman, R., and Bourke, A. (2001). The influence of sociality on the conservation biology of social insects. Ecol. Lett. 4, 650–662. doi: 10.1046/j.1461-0248.2001.00253.x
Cheng, W., and Ashton, L. (2021). Ecology: what affects the distribution of global bee diversity. Curr. Biol. 31, 127–128. doi: 10.1016/j.cub.2020.11.044
Choate, B. A., Hickman, P. L., and Moretti, E. A. (2018). Wild bee species abundance and richness across an urban–rural gradient. J. Insect Conserv. 22, 391–403. doi: 10.1007/s10841-018-0068-6
Christmann, S. (2019). Do we realize the full impact of pollinator loss on other ecosystem services and the challenges for any restoration in terrestrial areas? Restor. Ecol. 27, 720–725. doi: 10.1111/rec.12950
Cohen, H., Egerer, M., Thomas, S.-S., and Philpott, S. M. (2022). Local and landscape features constrain the trait and taxonomic diversity of urban bees. Landsc. Ecol. 37, 583–599. doi: 10.1007/s10980-021-01370-z
Colla, S., Willis, E., and Packer, L. (2009). Can green roofs provide habitat for urban bees (Hymenoptera: Apidae)? Cities Environ. 2. doi: 10.15365/cate.2142009
Comba, L., Corbet, S. A., Barron, A., Bird, A., Collinge, S., Miyazaki, N., et al. (1999). Garden flowers: insect visits and the floral reward of horticulturally-modified variants. Ann. Bot. 83, 73–86. doi: 10.1006/anbo.1998.0798
Corbet, S. A., Bee, J., Dasmahapatra, K., Gale, S., Gorringe, E., La Ferla, B., et al. (2001). Native or exotic? Double or single? Evaluating plants for pollinator-friendly gardens. Ann. Bot. 87, 219–232. doi: 10.1006/anbo.2000.1322
Cunningham-Minnick, M. J., Peters, V. E., and Crist, T. O. (2019). Nesting habitat enhancement for wild bees within soybean fields increases crop production. Apidologie 50, 833–844. doi: 10.1007/s13592-019-00691-y
Danforth, B. N., Minckley, R. L., Neff, J. L., and Fawcett, F. (2019). The Solitary Bees: Biology, Evolution, Conservation. Princeton, NJ: Princeton University Press.
De Palma, A., Abrahamczyk, S., Aizen, M., Albrecht, M., Basset, Y., Bates, A., et al. (2016). Predicting bee community responses to land-use changes: effects of geographic and taxonomic biases. Sci. Rep. 6, 31153. doi: 10.1038/srep31153
Del Toro, I., and Ribbons, R. (2020). No Mow May lawns have higher pollinator richness and abundances: an engaged community provides floral resources for pollinators. PeerJ 8, e10021. doi: 10.7717/peerj.10021
Dennis, M., and James, P. (2017). Ecosystem services of collectively managed urban gardens: exploring factors affecting synergies and trade-offs at the site level. Ecosyst. Serv. 26, 17–26. doi: 10.1016/j.ecoser.2017.05.009
Doody, B. J., Perkins, H. C., Sullivan, J. J., Meurk, C. D., and Stewart, G. H. (2014). Performing weeds: gardening, plant agencies and urban plant conservation. Geoforum 56, 124–136. doi: 10.1016/j.geoforum.2014.07.001
Drew, L. (2011). Are we losing the science of taxonomy? As need grows, numbers and training are failing to keep up. BioScience 61, 942–946. doi: 10.1525/bio.2011.61.12.4
Droege, S., Engler, J., Sellers, E., and O'Brien, L. (2016). National Protocol Framework for the Inventory and Monitoring of Bees. U.S. Fish and Wildlife Service, U.S. Department of the Interior. Available online at: https://static1.squarespace.com/static/61faf397da49d61281536985/t/628fd486a292a27b0991c0d9/1653593225179/National_Bee_Monitoring_Protocol_Framework_Final_06-17-2016.pdf (accessed November 15, 2022).
Dunnett, N., and Qasim, M. (2000). Perceived benefits to human well-being of urban gardens. Horttechnology 10, 40–45. doi: 10.21273/HORTTECH.10.1.40
Ebmer, A. (1995). Asiatische Halictidae, 3. Die Artengruppe der Lasioglossum carinate-Evylaeus (Insecta: Hymenoptera: Apoidea: Halictidae: Halictinae). Linzer Biologische Beiträge 27, 525–652.
Egerer, M., Cecala, J. M., and Cohen, H. (2019). Wild bee conservation within urban gardens and nurseries: effects of local and landscape management. Sustainability 12, 293. doi: 10.3390/su12010293
Fauviau, A., Baude, M., Bazin, N., Fiordaliso, W., Fisogni, A., Fortel, L., et al. (2022). A large-scale dataset reveals taxonomic and functional specificities of wild bee communities in urban habitats of Western Europe. Sci. Rep. 12, 18866. doi: 10.1038/s41598-022-21512-w
Fetridge, E. D., Ascher, J. S., and Langellotto, G. A. (2008). The bee fauna of residential gardens in a suburb of New York City (Hymenoptera: Apoidea). Ann. Entomol. Soc. Am. 101, 1067–1077. doi: 10.1603/0013-8746-101.6.1067
Fitch, G., Wilson, C. J., Glaum, P., Vaidya, C., Simao, M.-C., and Jamieson, M. A. (2019). Does urbanization favour exotic bee species? Implications for the conservation of native bees in cities. Biol. Lett. 15, 20190574. doi: 10.1098/rsbl.2019.0574
Fortel, L., Henry, M., Guilbaud, L., Guirao, A. L., Kuhlmann, M., Mouret, H., et al. (2014). Decreasing abundance, increasing diversity and changing structure of the wild bee community (Hymenoptera: Anthophila) along an urbanization gradient. PLoS ONE 9, e104769. doi: 10.1371/journal.pone.0104679
Fowler, J. (2016). Specialist bees of the Northeast: host plants and habitat conservation. Northeast. Nat. 23, 305–320. doi: 10.1656/045.023.0210
Fowler, J. (2020a). Pollen Specialist Bees of the Central United States. Available online at: https://jarrodfowler.com/bees_pollen.html (accessed October 10, 2022).
Fowler, J. (2020b). Pollen Specialist Bees of the Western United States. Available online at: https://jarrodfowler.com/pollen_specialist.html (accessed October 10, 2022).
Fowler, J., and Droege, S. (2020). Pollen Specialist Bees of the Eastern United States. Available online at: https://jarrodfowler.com/specialist_bees.html (accessed October 10, 2022).
Frankie, G. W., Thorp, R. W., Pawelek, J. C., Hernandez, J., and Coville, R. (2009). Urban bee diversity in a small residential garden in northern California. J. Hymenopt. Res. 18, 368–379.
Fukase, J., and Simons, M. M. (2016). Increased pollinator activity in urban gardens with more native flora. Appl. Ecol. Environ. Res. 14, 297–310. doi: 10.15666/aeer/1401_297310
Gardiner, M. M., Burkman, C. E., and Prajzner, S. P. (2013). The value of urban vacant land to support arthropod biodiversity and ecosystem services. Environ. Entomol. 42, 1123–1136. doi: 10.1603/EN12275
Geslin, B., Le Féon, V., Kuhlmann, M., Vaissière, B. E., and Dajoz, I. (2016). The bee fauna of large parks in downtown Paris, France. Ann. Soc. Entomol. France 51, 487–493. doi: 10.1080/00379271.2016.1146632
Giles, V., and Ascher, J. S. (2006). A survey of the bees of the Black Rock Forest Preserve, New York (Hymenoptera: Apoidea). J. Hymenopt. Res. 15, 208–231.
Gotlieb, A., Hollender, Y., and Mandelik, Y. (2011). Gardening in the desert changes bee communities and pollination network characteristics. Basic Appl. Ecol. 12, 310–320. doi: 10.1016/j.baae.2010.12.003
Goulson, D., and Nicholls, E. (2016). The canary in the coalmine; bee declines as an indicator of environmental health. Sci. Progr. 99, 312–326 doi: 10.3184/003685016X14685000479908
Goulson, D., Nicholls, E., Botías, C., and Rotherary, E. (2015). Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 347, 1255957. doi: 10.1126/science.1255957
Greenleaf, S. S., Williams, N. M., Winfree, R., and Kremen, C. (2007). Bee foraging ranges and their relationship to body size. Oecologia 153, 589–596. doi: 10.1007/s00442-007-0752-9
Groffman, P. M., Cavender-Bares, J., Bettez, N. D., Grove, J. M., Hall, S. J., Heffernan, J. B., et al. (2014). Ecological homogenization of urban USA. Front. Ecol. Environ. 12, 74–81. doi: 10.1890/120374
Gruver, A., and CaraDonna, P. (2021). Chicago bees: urban areas support diverse bee communities but with more non-native bee species compared to suburban areas. Environ. Entomol. 50, 982–994. doi: 10.1093/ee/nvab048
Halder, A., Dey, D., and Debnath, N. (2013). Pollinators of Karela in non-urbanized and urbanized fields. Environ. Ecol. 31, 1178–1184.
Hall, D. M., and Martins, D. J. (2020). Human dimensions of insect pollinator conservation. Curr. Opin. Insect Sci. 38, 107–114. doi: 10.1016/j.cois.2020.04.001
Hamblin, A. L., Youngsteadt, E., López-Uribe, M. M., and Frank, S. D. (2017). Physiological thermal limits predict differential responses of bees to urban heat-island effects. Biol. Lett. 13, 20170125. doi: 10.1098/rsbl.2017.0125
Hane, E. N., and Korfmacher, K. F. (2022). Insect “Bee&Bees” and pollinator penthouses: teaching students about pollinators and their services in an urban environment. Urban Ecosyst. 25, 1057–1064. doi: 10.1007/s11252-021-01186-4
Hernandez, J. L., Frankie, G. W., and Thorp, R. W. (2009). Ecology of urban bees: a review of current knowledge and directions for future study. Cate 2, 1–15. doi: 10.15365/cate.2132009
Hladik, M. L., Vandever, M., and Smalling, K. L. (2016). Exposure of native bees foraging in an agricultural landscape to current-use pesticides. Sci. Total Environ. 542, 469–477. doi: 10.1016/j.scitotenv.2015.10.077
Hostetler, N. E., and McIntyre, M. E. (2001). Effects of urban land use on pollinator (Hymenoptera: Apoidea) communities in a desert metropolis. Basic Appl. Ecol. 2, 209–218. doi: 10.1078/1439-1791-00051
Hui, V. L. S. (2021). Bee-Plant Networks in Allotment Gardens (Undergraduate thesis). Singapore, National University of Singapore.
Hyjazie, B. F., and Sargent, R. D. (2022). Floral resources predict the local bee community: implications for conservation. Biol. Conserv. 273, 109679. doi: 10.1016/j.biocon.2022.109679
Kearns, C. A., and Oliveras, D. M. (2009). Environmental factors affecting bee diversity in urban and remote grassland plots in Boulder, Colorado. J. Insect. Conserv. 13, 655–665. doi: 10.1007/s10841-009-9215-4
Klein, A.-M., Vassière, B., Cane, J., Steffan-Dewenter, I., Cunningham, S., Kremen, C., et al. (2007). Importance of pollinators in changing landscapes for world crops. Proc. R. Soc. B 274, 303–313. doi: 10.1098/rspb.2006.3721
Langellotto, G. A., Melathopoulos, A., Messer, I., Anderson, A., McClintock, N., and Costner, L. (2018). Garden pollinators and the potential for ecosystem service flow to urban and peri-urban agriculture. Sustainability 10, 2047. doi: 10.3390/su10062047
Lanner, J., Kratschmer, S., Petrovic, B., Gaulhofer, F., Meimberg, H., and Pachinger, B. (2020). City dwelling wild bees: how communal gardens promote species richness. Urban Ecosyst. 23, 271–288. doi: 10.1007/s11252-019-00902-5
Larkin, L. L., Neff, J. L., and Simpson, B. B. (2008). The evolution of a pollen diet: host choice and diet breadth of Andrena bees (Hymenoptera: Andrenidae). Apidologie 39, 133–145. doi: 10.1051/apido:2007064
Larson, K. L., Andrade, R., Nelson, K. C., Wheeler, M. M., Engebreston, J. M., Hall, S. J., et al. (2020). Municipal regulation of residential landscapes across US cities: patterns and implications for landscape sustainability. J. Environ. Manage. 275, 111132. doi: 10.1016/j.jenvman.2020.111132
LeBuhn, G., Griswold, T., Minckley, R., Droege, S., Roulston, T. A., Cane, J., et al. (2003). A Standardized Method for Monitoring Bee Populations–The Bee Inventory (BI) Plot, 5.
LeCroy, K. A., Savoy-Burke, G., Carr, D. E., Delaney, D. A., and Roulston, T. H. (2020). Decline of six native mason bee species following the arrival of an exotic congener. Sci. Rep. 10, 18745. doi: 10.1038/s41598-020-75566-9
Lerman, S., and Milam, J. (2016). Bee fauna and floral abundance within lawn-dominated suburban yards in Springfield, MA. Ann. Entomol. Soc. Am.109, 713–723. doi: 10.1093/aesa/saw043
Li, J., and Nassauer, J. I. (2020). Cues to care: a systematic analytical review. Landsc. Urban Plann. 201, 103821. doi: 10.1016/j.landurbplan.2020.103821
Liow, L. H., Sodhi, N., and Elmqvist, T. (2001). Bee diversity along a disturbance gradient in tropical lowland forest of South-East Asia. J. Appl. Ecol. 38, 180–192. doi: 10.1046/j.1365-2664.2001.00582.x
Locke, D. H., Roy Chowdhury, R., Grove, J. M., Martin, D. G., Goldman, E., Rogan, J., et al. (2018). Social norms, yard care, and the difference between front and back yard management: Examining the landscape mullets concept on urban residential lands. Soc. Nat. Resour. 31, 1169–1188. doi: 10.1080/08941920.2018.1481549
Lopezaraiza-Mikel, M. E., Hayes, R. B., Whalley, M. R., and Memmott, J. (2007). The impact of an alien plant on a native plant-pollinator network: an experimental approach. Ecol. Lett. 10, 539–550. doi: 10.1111/j.1461-0248.2007.01055.x
Lowenstein, D., Matteson, K., and Minor, E. (2015). Diversity of wild bees supports pollination services in an urbanized landscape. Oecologia179, 811–821. doi: 10.1007/s00442-015-3389-0
Lowenstein, D., Matteson, K., and Minor, E. (2019). Evaluating the dependence of urban pollinators on ornamental, non-native, and “weedy” floral resources. Urban Ecosyst. 22, 293–302. doi: 10.1007/s11252-018-0817-z
Lowenstein, D. M., Matteson, K. C., Xiao, I., Silva, A. M., and Minor, E. S. (2014). Humans, bees, and pollination services in the city: the case of Chicago, IL (USA). Biodivers. Conserv. 23, 2857–2874. doi: 10.1007/s10531-014-0752-0
Lutz, F. E. (1941). A lot of insects. Entomology in a suburban garden. J. N. Y. Entomol. Soc. 49, 366.
Makinson, J. C., Threlfall, C. G., and Latty, T. (2017). Bee-friendly community gardens: impact of environmental variables on the richness and abundance of exotic and native bees. Urban Ecosyst. 20, 463–476. doi: 10.1007/s11252-016-0607-4
Marzluff, J. (2015). Welcome to Subbirdia: Sharing Our Neighborhoods With Wrens, Robins, Woodpeckers, and Other Wildlife. New Haven, CT: Yale University Press, 320.
Matteson, K. C., Ascher, J. S., and Langellotto, G. A. (2008). Bee richness and abundance in New York city urban gardens. Ann. Entomol. Soc. Am. 101, 140–150. doi: 10.1603/0013-8746(2008)101[140:BRAAIN]2.0.CO;2
Matteson, K. C., and Langellotto, G. A. (2010). Determinates of inner city butterfly and bee species richness. Urban Ecosyst. 13, 333–347. doi: 10.1007/s11252-010-0122-y
Meftaul, I M. D., Venkateswarlu, K., Dharmarajan, R., Annamalai, P., and Megharaj, M. (2020). Pesticides in the urban environment: a potential threat that knocks at the door. Sci. Total Environ. 711, 134612. doi: 10.1016/j.scitotenv.2019.134612
Michez, D., Patiny, S., Rasmont, P., Timmermann, K., and Vereecken, N. J. (2008). Phylogeny and host-plant evolution in Melittidae s.l. (Hymenoptera: Apoidea). Apidologie 39, 146–162. doi: 10.1051/apido:2007048
Millard, J., Outhwaite, C. L., Kinnersley, R., Freeman, R., Gregory, R. D., Adedoja, O., et al. (2021). Global effects of land-use intensity on local pollinator biodiversity. Nat. Commun. 12, 2902. doi: 10.1038/s41467-021-23228-3
Miller, J. D., Kim, H., Kjeldsen, T. R., Packman, J., Grebby, S., and Dearden, R. (2014). Assessing the impact of urbanization on storm runoff in a peri-urban catchment using historical change in impervious cover. J. Hydrol. 515, 59–70. doi: 10.1016/j.jhydrol.2014.04.011
Morandin, L. A., and Kremen, C. (2013). Hedgerow restoration promotes pollinator populations and exports native bees to adjacent fields. Ecol. Appl. 23, 829–839. doi: 10.1890/12-1051.1
Morse, C. C., Huryn, A. D., and Cronan, C. (2003). Impervious surface area as a predictor of the effects of urbanization on stream insect communities in Maine, U.S.A. Environ. Monit. Assess. 89, 95–127. doi: 10.1023/A:1025821622411
Müller, A., and Kuhlmann, M. (2008). Pollen hosts of western palaearctic bees of the genus Colletes (Hymenoptera: Colletidae): the Asteraceae paradox. Biol. J. Linn. Soc. 95, 719–733. doi: 10.1111/j.1095-8312.2008.01113.x
Nassauer, J. I. (1995). Messy ecosystems, orderly frames. Landsc. J. 14, 161–170. doi: 10.3368/lj.14.2.161
Nassauer, J. I., Wang, Z., and Dayrell, E. (2009). What will the neighbors think? Cultural norms and ecological design. Landsc. Urban Plann. 92, 282. doi: 10.1016/j.landurbplan.2009.05.010
Normandin, E., Vereecken, N. J., Buddle, C. M., and Fournier, V. (2017). Taxonomic and functional trait diversity of wild bees in different urban settings. PeerJ 5, e3051. doi: 10.7717/peerj.3051
Ollerton, J., Trunschke, J., Havens, K., Landaverde-González, P., Keller, A., Gilpin, A.-M., et al. (2022). Pollinator-flower interactions in gardens during the COVID-19 pandemic lockdown of 2020. J. Pollinat. Ecol. 31, 87–96. doi: 10.26786/1920-7603(2022)695
Ollerton, J., Winfree, R., and Tarrant, S. (2011). How many flowering plants are pollinated by animals. Oikos 120, 321–326. doi: 10.1111/j.1600-0706.2010.18644.x
Olson, D. M., Dinerstein, E., Wikramanayake, E. D., Burgess, N. D., Powell, G. V. N., Underwood, E. C., et al. (2001). Terrestrial ecoregions of the world: a new map of life on Earth. Bioscience 51, 933–938. doi: 10.1641/0006-3568(2001)051[0933:TEOTWA]2.0.CO;2
Orr, M. C., Hughes, A. C., Chesters, D., Pickering, J., Zhu, C.-D., and Ascher, J. S. (2021). Global patterns and drivers of bee distribution. Curr. Biol. 31, 451–458.e4. doi: 10.1016/j.cub.2020.10.053
Pardee, G. L., and Philpott, S. M. (2014). Native plants are the bee's knees: local and landscape predictors of bee richness and abundance in backyard gardens. Urban Ecosyst. 17, 641–659. doi: 10.1007/s11252-014-0349-0
Pauly, A. (2016). Les Xylocopa d'Afrique. Atlas Hymenoptera. Available online at: http://www.atlashymenoptera.net/page.aspx??id=84 (accessed April 1, 2023).
Pawelek, J. C., Frankie, G. W., Frey, K., Guerrero, S. L., and Schindler, M. (2015). California Bee-Friendly Garden Recipes. Richmond, CA: University of California Agriculture and Natural Resources.
Pawelek, J. C., Frankie, G. W., Thorp, R. W., and Przybylski, M. (2009). Modification of a community garden to attract native bee pollinators in urban San Luis Obispo, California. Cities Environ. 2, 1–20. doi: 10.15365/cate.2172009
Pei, C. K., Hovick, T. J., Limb, R. F., Harmon, J. P., and Geaumont, B. A. (2023). Native and introduced pollinators vary in their seasonal floral resource visitation and selection between native and exotic plant species. J. Appl. Ecol. 60, 1424–1434. doi: 10.1111/1365-2664.14416
Percival, M. (1974). Floral ecology of coastal scrub in southeast Jamaica. Biotropica 6, 104–129. doi: 10.2307/2989824
Persson, A. S., Ekroos, J., Olsson, P., and Smith, H. G. (2020). Wild bees and hoverflies respond differently to urbanisation, human population density and urban form. Landsc. Urban Plan. 204, 103901. doi: 10.1016/j.landurbplan.2020.103901
Plascencia, M., and Philpott, S. (2017). Floral abundance, richness, and spatial distribution drive urban garden bee communities. Bul. Entomol. Res. 107, 658–667. doi: 10.1017/S0007485317000153
Portman, Z. M., Bruninga-Socolar, B., and Cariveau, D. P. (2020). The state of bee monitoring in the United States: a call to refocus away from bowl traps and towards more effective methods. Ann. Entomol. Soc. Am. 113, 337–342. doi: 10.1093/aesa/saaa010
Potts, S. G., Biesmeijer, J., Kremen, C., Neumann, P., Schweiger, O., and Kunin, E. (2010). Global pollinator declines: trends, impacts and drivers. Trends Ecol. Evol. 25, 345–353. doi: 10.1016/j.tree.2010.01.007
Potts, S. G., Imperatriz-Fonseca, V., Ngo, H. T., Aizen, M. A., Biesmeijer, J. C., Breeze, T. D., et al. (2016). Safeguarding pollinators and their values to human well-being. Nature 540, 220–229. doi: 10.1038/nature20588
Prendergast, K. S., Dixon, K. W., and Bateman, P. W. (2022). A global review of determinants of native bee assemblages in urbanized landscapes. Insect Conserv. Divers. 15, 385–405. doi: 10.1111/icad.12569
Rahimi, E., Barghjelveh, S., and Dong, P. (2022). A review of diversity of bees, the attractiveness of host plants and the effects of landscape variables on bees in urban gardens. Agric. Food Sec. 11, 6. doi: 10.1186/s40066-021-00353-2
Roulston, T. H., Cane, J. H., and Buchmann, S. L. (2000). What governs protein content of pollen: Pollinator preferences, pollen–pistil interactions, or phylogeny? Ecol. Monogr. 70, 617–643. doi: 10.1890/0012-9615(2000)070[0617:WGPCOP]2.0.CO;2
Salisbury, A., Armitage, J., Bostock, H., Perry, J., Tatchell, M., and Thompson, K. (2015). Editor's Choice: enhancing gardens as habitats for flower-visiting aerial insects (pollinators): should we plant native or exotic species? J. Appl. Ecol. 52, 1156–1164. doi: 10.1111/1365-2664.12499
Sgolastra, F., Hinarejos, S., Pitts-Singer, T. L., Boyle, N. K., Joseph, T., Luckmann, J., et al. (2019). Pesticide exposure assessment paradigm for solitary bees. Environ. Entomol. 48, 22–35. doi: 10.1093/ee/nvy105
Shackleton, C. M., Cilliers, S. S., du Toit, M. J., and Davoren, E. (2021). “The need for an urban ecology of the Global South,” in Urban Ecology in the Global South, Cities and Nature, eds C. M. Shackleton, S. S. Cilliers, E. Davoren, and M. J. du Toit. Cham: Springer International Publishing, 1–26.
Sheffield, C. S., Pindar, A., Packer, L., and Kevan, P. G. (2013). The potential of cleptoparasitic bees as indicator taxa for assessing bee communities. Apidologie 44, 501–510. doi: 10.1007/s13592-013-0200-2
Sing, K., Wang, W., Wan, T., Lee, P., Li, Z., Chen, X., et al. (2016). Diversity and human perceptions of bees (Hymenoptera: Apoidea) in Southeast Asian megacities. Genome 59, 827–839. doi: 10.1139/gen-2015-0159
Sirohi, M., Jackson, J., Edwards, M., and Ollerton, J. (2015). Diversity and abundance of solitary and primitively eusocial bees in an urban centre: a case study from Northampton (England). J. Insect Conserv. 19, 487–500. doi: 10.1007/s10841-015-9769-2
Sivakoff, F. S., Prajzner, S. P., and Gardiner, M. M. (2018). Unique bee communities within vacant lots and urban farms result from variation in surrounding urbanization intensity. Sustainability 10, 1926. doi: 10.3390/su10061926
Soroye, P., Newbold, T., and Kerr, J. (2020). Climate change contributes to widespread declines among bumble bees across continents. Science 367, 685–688. doi: 10.1126/science.aax8591
Staab, M., Pereira-Peixoto, M. H., and Klein, A.-M. (2020). Exotic garden plants partly substitute for native plants as resources for pollinators when native plants become seasonally scarce. Oecologia 194, 465–480. doi: 10.1007/s00442-020-04785-8
Stewart, A., Sritongchuay, T., Teartisup, P., Kaewsomboon, S., and Bumrungsri, S. (2018). Habitat and landscape factors influence pollinators in a tropical megacity, Bangkok, Thailand. PeerJ 6, e5335. doi: 10.7717/peerj.5335
Stout, J. C., and Morales, C. L. (2009). Ecological impacts of invasive alien species on bees. Apidologie 40, 388–409. doi: 10.1051/apido/2009023
Sturiale, L., Scuderi, A., Timpanaro, G., Foti, V. T., and Stella, G. (2020). “Social and inclusive “value” generation in metropolitan area with the “Urban Gardens” planning,” in Values and Functions for Future Cities, Green Energy and Technology, eds G. Mondini, A. Oppio, S. Stanghellini, M. Bottero, and F. Abastante (Cham: Springer International Publishing), 285–302.
Sunny, A., Diwakar, S., and Sharma, G. P. (2015). Native insects and invasive plants encounters. Arthropod. Plant Interact. 9, 323–331. doi: 10.1007/s11829-015-9384-x
Synge, A. D. (1947). Pollen collection by honeybees (Apis mellifera). J. Anim. Ecol. 16, 122–138. doi: 10.2307/1492
Tepedino, V. J., Bradley, B. A., and Griswold, T. L. (2008). Might flowers of invasive plants increase native bee carrying capacity? Intimations from Capitol Reef National Park, Utah. Nat. Areas J. 28, 44–50. doi: 10.3375/0885-8608(2008)28[44:MFOIPI]2.0.CO;2
Thompson, K., Austin, K. C., Smith, R. M., Warren, P. H., Angold, P. G., and Gaston, K. J. (2003). Urban domestic gardens (I): putting small-scale plant diversity in context. J. Veg. Sci. 14, 71–78. doi: 10.1111/j.1654-1103.2003.tb02129.x
Threlfall, C. G., Ossola, A., Hahs, A., Williams, N., Wilson, L., and Livesley, S. (2016). Variation in vegetation structure and composition across urban green space types. Front. Ecol. Evol. 4. doi: 10.3389/fevo.2016.00066
Threlfall, C. G., Walker, K., Williams, N., Hahs, A., Mata, L., and Livesley, S. (2015). The conservation value of urban green space habitats for Australian native bee communities. Biol. Conserv. 187, 240–248. doi: 10.1016/j.biocon.2015.05.003
Tommasi, D., Miro, A., Higo, H. A., and Winston, M. L. (2004). Bee diversity and abundance in an urban setting. Can. Entomol. 136, 851–869. doi: 10.4039/n04-010
Tonietto, R., Fant, J., Ascher, J., Ellis, K., and Larkin, D. (2011). A comparison of bee communities of Chicago green roofs, parks and prairies. Landsc. Urban Plan. 103, 102–108. doi: 10.1016/j.landurbplan.2011.07.004
Turo, K. J., and Gardiner, M. M. (2019). From potential to practical: conserving bees in urban public green spaces. Front. Ecol. Environ. 17, 167–175. doi: 10.1002/fee.2015
Twerd, L., and Banaszak-Cibicka, W. (2019). Wastelands: their attractiveness and importance for preserving the diversity of wild bees in urban areas. J. Insect Conserv. 23, 573–588. doi: 10.1007/s10841-019-00148-8
United Nations (2018). Revision of World Urbanization Prospects [WWW Document]. United Nations. Available online at: https://www.un.org/en/desa/2018-revision-world-urbanization-prospects (accessed October 15, 2022).
United Nations Population Fund (UNFPA) (2007). UNFPA Annual Report [WWW Document]. United Nations Population Fund. Available online at: https://www.unfpa.org/sites/default/files/pubpdf/ar07_eng.pdf (accessed October 15, 2022).
Villalta, I., Ledet, R., Baude, M., Genoud, D., Bouget, C., Cornillon, M., et al. (2021). A DNA barcode-based survey of wild urban bees in the Loire Valley, France. Sci. Rep. 11, 4770. doi: 10.1038/s41598-021-83631-0
Wen, Z., Soh, W., Wen, R., and Ngiam, R. (2013). Flower-visiting bees and wasps in Singapore parks (Insecta: Hymenoptera). Nat. Singap. 6, 153–172.
Wenzel, A., Grass, I., Belavadi, V., and Tscharntke, T. (2020). How urbanization is driving pollinator diversity and pollination – A systematic review. Biol. Conserv. 241, 108321. doi: 10.1016/j.biocon.2019.108321
Wickham, H., François, R., Henry, L., and Müller, K. (2022). dplyr: A Grammar of Data Manipulation. Available online at: https://dplyr.tidyverse.org, https://github.com/tidyverse/dplyr (accessed June 1, 2023).
Williams, N. M., Cariveau, D., Winfree, R., and Kremen, C. (2011). Bees in disturbed habitats use, but do not prefer, alien plants. Basic Appl. Ecol. 12, 332–341. doi: 10.1016/j.baae.2010.11.008
Wills, R. T., Lyons, M., and Bell, D. T. (1990). The European Honey Bee in Western Australian Kwongan: Foraging Preferences and Some Implications for Management.
Wilson, C. J., and Jamieson, M. A. (2019). The effects of urbanization on bee communities depends on floral resource availability and bee functional traits. PLoS ONE 14, e0225852. doi: 10.1371/journal.pone.0225852
Wilson, J. S., Forister, M. L., and Carril, O. M. (2017). Interest exceeds understanding in public support of bee conservation. Front. Ecol. Environ. 15, 460–466. doi: 10.1002/fee.1531
Winfree, R., Williams, N., Gaines, H., Ascher, J., and Kremen, C. (2008). Wild bee pollinators provide the majority of crop visitation across land-use gradients in New Jersey and Pennsylvania, USA. J. Appl. Ecol. 45, 793–802. doi: 10.1111/j.1365-2664.2007.01418.x
Wojcik, V. A., Frankie, G. W., Thorp, R. W., and Hernandez, J. L. (2008). Seasonality in bees and their floral resource plants at a constructed urban bee habitat in Berkeley, California. J. Kans. Entomol. Soc. 81, 15–28. doi: 10.2317/JKES-701.17.1
Wood, T. J. (2021). Fifteen new Andrena species from little-visited aris, Mediterranean, and mountainous parts of the Old World (Hymenoptera: Andrenidae). Zootaxa 4933, 451–492. doi: 10.11646/ZOOTAXA.4933.4.1
Wood, T. J., Kaplan, I., and Szendrei, Z. (2018). Wild bee pollen diets reveal patterns of seasonal foraging resources for honey bees. Front. Ecol. Evol. 6, 210. doi: 10.3389/fevo.2018.00210
Zanette, L. R. S., Martins, R. P., and Ribeiro, S. P. (2005). Effects of urbanization on Neotropical wasp and bee assemblages in a Brazilian metropolis. Landsc. Urban Plan. 71, 105–121. doi: 10.1016/j.landurbplan.2004.02.003
Keywords: Anthophila, pollinator, urban, ornamental landscapes, garden
Citation: Bell NCS, Ascher JS, Hayes JJ-M, Mead M and Langellotto GA (2023) Assessing five decades of garden bee studies. Front. Sustain. Cities 5:1102360. doi: 10.3389/frsc.2023.1102360
Received: 18 November 2022; Accepted: 02 October 2023;
Published: 30 October 2023.
Edited by:
Katherine Baldock, Northumbria University, United KingdomReviewed by:
Benoît Geslin, UMR7263 Institut méditerranéen de biodiversité et d'écologie marine et continentale (IMBE), FranceCopyright © 2023 Bell, Ascher, Hayes, Mead and Langellotto. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Nicole C. S. Bell, bmljb2xlY3NiZWxsQGdtYWlsLmNvbQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.