 Hannes Peter
Hannes Peter Grégoire Michoud
Grégoire Michoud Susheel Bhanu Busi
Susheel Bhanu Busi Tom J. Battin1
Tom J. Battin1
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiomes , 15 January 2024
Sec. Environmental Microbiomes
Volume 2 - 2023 | https://doi.org/10.3389/frmbi.2023.1279550
This article is part of the Research Topic Ecology, Evolution, and Biodiversity of Microbiomes and Viromes From Extreme Environments View all 8 articles
Viruses modulate the diversity and activity of microbial communities. However, little is known about their role for the structure of stream bacterial biofilm communities. Here, we present insights into the diversity and composition of viral communities in various streams draining three proglacial floodplains in Switzerland. Proglacial streams are characterized by extreme environmental conditions, including near-freezing temperatures and ultra-oligotrophy. These conditions select for few but well-adapted bacterial clades, which dominate biofilm communities and occupy niches via microdiversification. We used metagenomic sequencing to reveal a diverse biofilm viral assemblage in these streams. Across the different floodplains and streams, viral community composition was tightly coupled to that of the bacterial hosts, which was underscored by generally high host specificity. Combining predictions of phage-host interactions with auxiliary metabolic genes (AMGs), we identify specific AMGs shared by phages infecting microdiverse clade members. Our work provides a step towards a better understanding of the complex interactions among bacteria and phages in stream biofilm communities in general and streams influenced by glacier meltwaters and characterized by microdiversity in particular.
Viruses infecting bacteria, also called bacteriophages, play important roles in modulating the diversity and activity of bacterial assemblages in every biome on Earth (Dion et al., 2020). Particularly phage-induced mortality is known to control bacterial abundance and biomass, community composition and diversity (Suttle, 1994; Suttle, 2007; Roux et al., 2016). Metabolic reprogramming during lytic infections redirects host metabolism towards viral progeny production, with sizable impacts on ecosystem-scale biogeochemical cycles (reviewed by: Zimmerman et al., 2020). However, beyond altering host’ metabolic pathways and inducing mortality, phages influence the fitness of their bacterial hosts in numerous ways. Lysogenic infections, for example, can impede host functioning by insertion of phage genetic material into functional genes (reviewed by: Obeng et al., 2016). And phage-encoded sigma factors can influence bacterial spore formation, affecting important bacterial life-history traits such as dormancy (Schwartz et al., 2022).
While the consequences of viral infections for individual bacterial cells are often detrimental, beneficial effects of phage-host interactions typically arise at the level of populations, communities or even holobionts (Shkoporov et al., 2022). For instance, phage-driven diversification (Koskella and Brockhurst, 2014), lysogenic conversion (Howard-Varona et al., 2017) or auxiliary metabolic genes [AMGs (Kieft et al., 2021b)] can increase phenotypic plasticity or augment the metabolic repertoire of microbial assemblages (reviewed by: Shkoporov et al., 2022). In biofilms, where densely packed communities of microbes interact, phage genomic material has been shown to enter even phylogenetically distant hosts directly or during conjugative transfers, ultimately increasing the immunological memory (Hwang et al., 2023).
Besides the density, diversity, and activity of phages and their hosts, the eco-evolutionary consequences of phage infections depend on host specificity. While a notion of a predominantly narrow phage host range prevailed in early work, recent advances, including single-cell and metagenomics studies of hydrothermal biofilms (Jarett et al., 2020; Hwang et al., 2023), suggest indeed a continuum of host ranges (reviewed by: de Jonge et al., 2019; Holtappels et al., 2023).
Here, we present insights into the interactions between bacterial biofilm communities and their phages in high-altitude stream ecosystems. Alpine streams are extreme environments, characterized by ultra-oligotrophy, low water temperature, exposure to ultraviolet (UV) radiation, and short ice-free seasons, yet represent hotspots of microbial biodiversity (Freimann et al., 2013; reviewed by: Hotaling et al., 2017; Brandani et al., 2022; Busi et al., 2022; Fodelianakis et al., 2022). Glacier influence adds additional constraints for life in alpine streams. Close to the glacier terminus, glacier-fed streams (GFS) are permanently ice cold and the erosive activity of glaciers liberates large quantities of mineral particles, which render GFS turbid and limit light for phototrophic primary production. Compared to GFS, groundwater-fed streams (GWS) that drain elevated terrain towards the edges of the floodplains, are warmer, their streambeds more consolidated and lower turbidity allows photosynthetic primary producers to dwell (Freimann et al., 2013; Brandani et al., 2022).
Previous work on biofilm communities in GFS and GWS revealed that they are diverse with members of all domains of life present (reviewed by: Battin et al., 2016; Brandani et al., 2022), distinct from the microbial community suspended in the water column (Ezzat et al., 2022), and shaped predominantly by environmental selection (Fodelianakis et al., 2022; Brandani et al., 2023). More specifically, this means that ecologically successful taxa in both GFS and GWS are recruited from similar clades, including taxa classified as Polaromonas, Rhodoferax, Rhizobacter, Methylotenera, and Massilia. These genera are microdiverse (Fodelianakis et al., 2022), indicating that they efficiently fill available niches which cannot be occupied by less-well adapted taxa. Microdiversity is an intrinsic property of many microbial communities (reviewed by: Larkin and Martiny, 2017), including marine viral communities (Needham et al., 2017; Gregory et al., 2019).
By focusing on microdiverse biofilm communities in proglacial streams, we aim to shed new light on the role of phage-host interactions in microbial communities in extreme environments. We hypothesize that narrow phage host ranges predominate in communities dominated by microdiverse clades because selective constraints which lead to microdiversification may also shape bacteria-phage coevolution. Moreover, we speculate that interactions between phages and microdiverse clade members in extreme environments may be predominantly beneficial, thus contributing to the ecological success of microdiverse clades in extreme environments. The rationale behind this is that reciprocal feedbacks between beneficial effects of phage-host interactions and the density of hosts could stabilize the process of microdiversification over evolutionary timescales – in contrast to a situation where density-dependent phage predation would lead to turnover in host communities (and thus suppress microdiversification). We argue that studying microbial communities in alpine stream biofilms is ideally suited to address such questions, because of the pronounced microdiversity and limited dispersal due to geographic and topographic isolation, which can increase phage-host encounter probabilities and hence lessen the coevolutionary strength of phage-host associations.
We here present data from an extensive survey of benthic biofilm communities sampled from proglacial streams in the Swiss Alps in summer 2019. Detailed descriptions of the sampling design and environmental parameters as well as analyses of spatial diversity patterns (Brandani et al., 2022), assembly processes (Brandani et al., 2023) and the functional diversity (Michoud et al., 2023) are available elsewhere. In this field survey, a total of 259 benthic sediment samples were collected from the Otemma Glacier (OTE, 45.95E, 7.45N), the Valsorey Glacier (SOY, 45.91E, 7.27N) and Val Roseg Glacier (VAR, 46.39E, 9.84N) floodplains. The sampling sites were distributed along gradients from the glacier snout to the outflows of the respective proglacial floodplains, covering terrain which has been deglaciated since ca. 36 (OTE), 65 (VAR), and 70 (SOY) years. Samples of the coarse sandy sediment fraction were collected using flame-sterilized shovels (0-5 cm depth) and fractionated using flame-sterilized sieves (Retsch, 0.25 and 3.15 mm). Subsamples were immediately flash-frozen on dry ice and stored at -80°C until processing. Samples for bacterial cell counting were fixed using 1.8 mL of a mixture containing filter-sterilized paraformaldehyde (1% w/v) and 0.5% (w/w) glutaraldehyde, flash frozen and kept at -80°C (Brandani et al., 2022; Fodelianakis et al., 2022). After extraction from sediments using pyrophosphate, shaking and sonication, bacterial cells were stained using SYBR Green I (1 x final concentration; Thermo Fisher) for 15 minutes at 37°C and counted using flow cytometry (NovoCyte, ACEA) equipped with a 488 nm laser. Samples were analyzed with a reading time of 2 minutes at a flow rate of 14 µL min-1 and thresholds on the forward scatter channel set to 300 and the 530/30 nm fluorescence channel set to 1500. Cells were distinguished from background noise based on fluorescence signals on biplots of 530/30 and 725/40 nm using the NovoExpress (ACEA) software. Cell abundances were converted to cells per g dry sediment.
Here, we focus on the viral communities, which we assess using bulk metagenomic sequencing of a subset of samples. We obtained metagenomes for 47 samples from GFS (n=12) and GWS (n=35) from OTE (nGFS=3, nGWS=14), SOY (nGFS=6, nGWS=10) and VAR (nGFS=3, nGWS=11), respectively. Twenty-four of these samples were obtained in June–July, and 23 samples were collected in August–September 2019 (Supplementary Table S1).
DNA was extracted from 0.5 g of sediment using an optimized extraction protocol tailored to the low-biomass and mineral nature of these samples (Busi et al., 2020). Metabarcoding libraries were prepared using 2-3 ng µL-1 input DNA and primers targeting the V3-V4 hypervariable region of the 16S rRNA gene [341f (5’-CCTACGGGNGGCWGCAG-3’) and 785r (5’-GACTACHVGGGTATCTAATCC-3’) (Klindworth et al., 2013)]. Amplification was performed on a Biometra Trio (Biometra) using the KAPA HiFi DNA Polymerase (Hot Start and Ready Mix formulation) in a 25 μL-amplification reaction containing 1 x PCR buffer, 1 μM of each primer, 0.48 μg μL−1 bovine serum albumin (BSA), and 1.0 μL of template DNA. After an initial denaturation step at 95°C for 3min, 25 cycles of 94°C for 30 s, 55°C for 30 s, and 72°C for 30 s were followed by a final extension at 72°C for 5 min. Sequencing libraries were prepared using dual indices (Illumina). Prior to paired-end sequencing on a single MiSeq (Illumina) lane, library DNA concentrations were quantified, normalized, and pooled. Libraries were then sequenced at the Lausanne Genomic Technologies Facility (Switzerland), producing 300 bp paired end reads.
Metagenome libraries were prepared using the NEBNext Ultra II FS library kit, with 50 ng of input DNA. Briefly, DNA was enzymatically fragmented for 12.5 min and amplified using 6 PCR cycles. Qubit (Invitrogen) and Bioanalyzer (Agilent) were used to check the quality and insert size of the libraries (450 bp). The metagenomes were sequenced at the Functional Genomics Centre Zürich using an S4 flowcell on a NovaSeq (Illumina) platform, producing 150 bp paired end reads.
Amplicon sequences were quality checked using Trimmomatic v0.36 (Bolger et al., 2014) and assigned to exact Amplicon Sequence Variants (ASVs) using DADA2 v1.10.0 (Callahan et al., 2016) implemented in QIIME2 v2020.8 (Bolyen et al., 2019) with default parameters. Singleton ASVs were removed and taxonomy was assigned using the naïve Bayesian classifier implemented in QIIME2 and the SILVA v138.1 reference database. Non-bacterial ASVs (i.e. chloroplasts, mitochondria, and archaea) were discarded. We built de novo a phylogenetic tree using RAxML v8.2.12 (Stamatakis, 2014) and the GTRCAT substitution model and the rapid bootstrapping option. Phylogenetic distances among ASVs were calculated using function cophenetic implemented in R v4.1 (R Core Team, 2021).
Metagenomic sequence reads were preprocessed using trim_galore v0.6.6 (Krueger et al., 2021), which uses fastqc v0.11.9 (Andrews, 2010) for quality control and cutadapt v3.4 (Martin, 2011) for adapter removal. Trimmed reads were assembled separately for each sample with megahit v1.2.9 (Li et al., 2015) using default parameters and a minimum contig length of 1,000 bp. Prokaryotic Metagenome-Assembled Genomes (MAGs) were binned using MetaBAT2 v2.15 (Kang et al., 2019) and CONCOCT v1.1 (Alneberg et al., 2014). Reads were mapped to the assemblies with CoverM v0.6.1 (https://github.com/wwood/CoverM). DAS Tool v1.1.2 (Sieber et al., 2018) was used to obtain a non-redundant set of MAGs for each sample. All MAGs were then dereplicated with dRep v3.2.2 (Olm et al., 2017), considering a completeness level greater than 75% and a contamination level less than 10% as determined by checkM v1.1.3 (Parks et al., 2015). Taxonomy of MAGs was assigned using GTDB-Tk v1.7.0 (Chaumeil et al., 2019) based on release 202 obtained from https://data.ace.uq.edu.au/public/gtdb/data/releases/ (Quast et al., 2013). DefenseFinder v1.0.9 (Tesson et al., 2022) using default settings was used to identify antiviral systems on MAGs.
Putative viral contigs (n=57168) were identified from the assemblies using VIBRANT v1.2.1 (Kieft et al., 2020) with default parameters. VIBRANT also provides information about auxiliary metabolic genes on viral contigs based on KEGG annotations (i.e. including “metabolic pathways” and “sulfur relay system”). Complete viral contigs were then identified using CheckV v0.8.1 (Nayfach et al., 2021) and set aside (n=801). The remaining, non-complete viral contigs were binned into viral metagenome-assembled genomes (vMAGs) with PHAMB v1.0.1 (Johansen et al., 2022) which uses VAMB v3.0.2 (Nissen et al., 2021) and DeepVirFinder v1.0 (Ren et al., 2020) to obtain high-quality viral genomes. Subsequently, complete viral genomes and high-quality vMAGs were dereplicated using vRhyme v1.1.0 (Kieft et al., 2022) taking the longest bin and 97% identity to obtain a final set of vMAGs. CheckV was then used to assess vMAG completeness and quality. To obtain vMAG abundances, reads were mapped to the concatenated vMAG sequences using CoverM v0.6.1 in contig mode, using 100% minimum read identity, and the “trimmed_mean” method, which removes 5% of the highest and lowest coverages, respectively. The number of reads mapped to vMAGs are reported in Supplementary Table S1. An additional detection threshold of 5% of the contig length covered was applied and vMAG abundances below this threshold were set to zero. Coverage was then normalized (i.e. to range between 0 and 1) and treated as relative abundances in downstream analyses. BACPHLIP v0.9.6 (Hockenberry and Wilke, 2021) was used to assign temperate or virulent lifestyles to complete vMAGs. Viral taxonomy was assigned using kaiju v1.9.0 (Menzel et al., 2016) and the viruses database (only viruses from the NCBI RefSeq database, accessed 07/05/2022). PHIST v1.1.0 (Zielezinski et al., 2022), an alignment-free, k-mer frequency-based tool was used to predict phage-host interactions (based on padjusted <0.05). A snakemake workflow for our bioinformatic analyses can be found here: https://github.com/hpeter0803/viromes/tree/ensemble_paper.
The statistical software language R v4.1 was used to prepare figures and to perform all statistical analyses. Specifically, viral taxonomic composition was visualized using the heat_tree function of the metacoder v0.3.6 (Foster et al., 2017) R package. Differential abundance analysis for vMAGs was performed using DESeq2 v3.15 (Love et al., 2014). Multivariate analyses of bacterial (16S rRNA gene amplicon) and viral (vMAG) community composition included non-metric multidimensional scaling ordination (metaMDS), procrustes correlation (procrustes and protest), and permutational multivariate analysis of variance using distance matrices (PERMANOVA, adonis2) using Bray-Curtis dissimilarities and were performed using the R package vegan v 2.6-4 (Oksanen et al., 2022) with default settings. Prior to these analyses, Wisconsin double standardization was applied to relative abundances and AMG counts were Hellinger transformed prior to Principal Component Analysis. Phage-host interactions were visualized using Cytoscape v3.10.0 (Shannon et al., 2003).
After removal of singletons, we retained 35,170 bacterial ASVs across the three different floodplains and the different stream types. The bacterial communities were taxonomically dominated by members of Gamma- and Alphaproteobacteria (17.4 and 8.2% of ASVs, respectively), Bacteroidia (12.5%), Planctomycetes (7.8%), and Verrucomicrobiae (6.4%). This diversity was further classified into 925 different genera, of which members of Candidatus Nomurabacteria (1.8% of ASVs), Flavobacterium (1.7%), and Candidatus Saccharimonadales (1.4%) accounted for most ASV richness.
Previous work has identified environmental factors common to both GFS and GWS, such as low water temperature (average 6.5 ± 4.6°C) and ultra-oligotrophy to shape proglacial stream biofilm communities (Brandani et al., 2022). Yet, important environmental differences between GFS and GWS, such as turbidity and diurnal variation in stream flow select for different ASVs which account for the compositional (Freimann et al., 2013; Brandani et al., 2022; Brandani et al., 2023) and functional differences (Michoud et al., 2023) between stream types. Leveraging phylogenetic and ecological signatures, we previously related environmental selection to microdiversity, particularly among members of the Gammaproteobacterial family Comamonadacea in proglacial streams in New Zealand (Fodelianakis et al., 2022) and the European Alps (Brandani et al., 2023). Here, we re-iterate this and use phylogenetic distances between ASVs, prevalence and relative abundance to identify microdiverse bacterial clades. We found that the genera Polaromonas, Rhodoferax, Rhizobacter, Methylotenera, and Massilia featured consistently short phylogenetic distances among ASVs (Figure 1A) and are microdiverse. Short phylogenetic distances could be a consequence of the number of ASVs with the same taxonomic classification. Indeed, we found that genera with few ASVs had generally shorter phylogenetic distances than genera containing many ASVs. Polaromonas (nASVs=169), Rhodoferax (nASVs=386), Rhizobacter (nASVs=283), Methylotenera (nASVs=134), and Massilia (nASVs=121), however, contain numerous ASVs, comparable to other, non-microdiverse bacterial genera (Figure 1A). And while these five microdiverse genera account on average ( ± SD) for only 5.0 ± 2.3% of ASV richness detected in each sample, they contribute 14.5 ± 11.3% to relative abundance (Figures 1B, C).
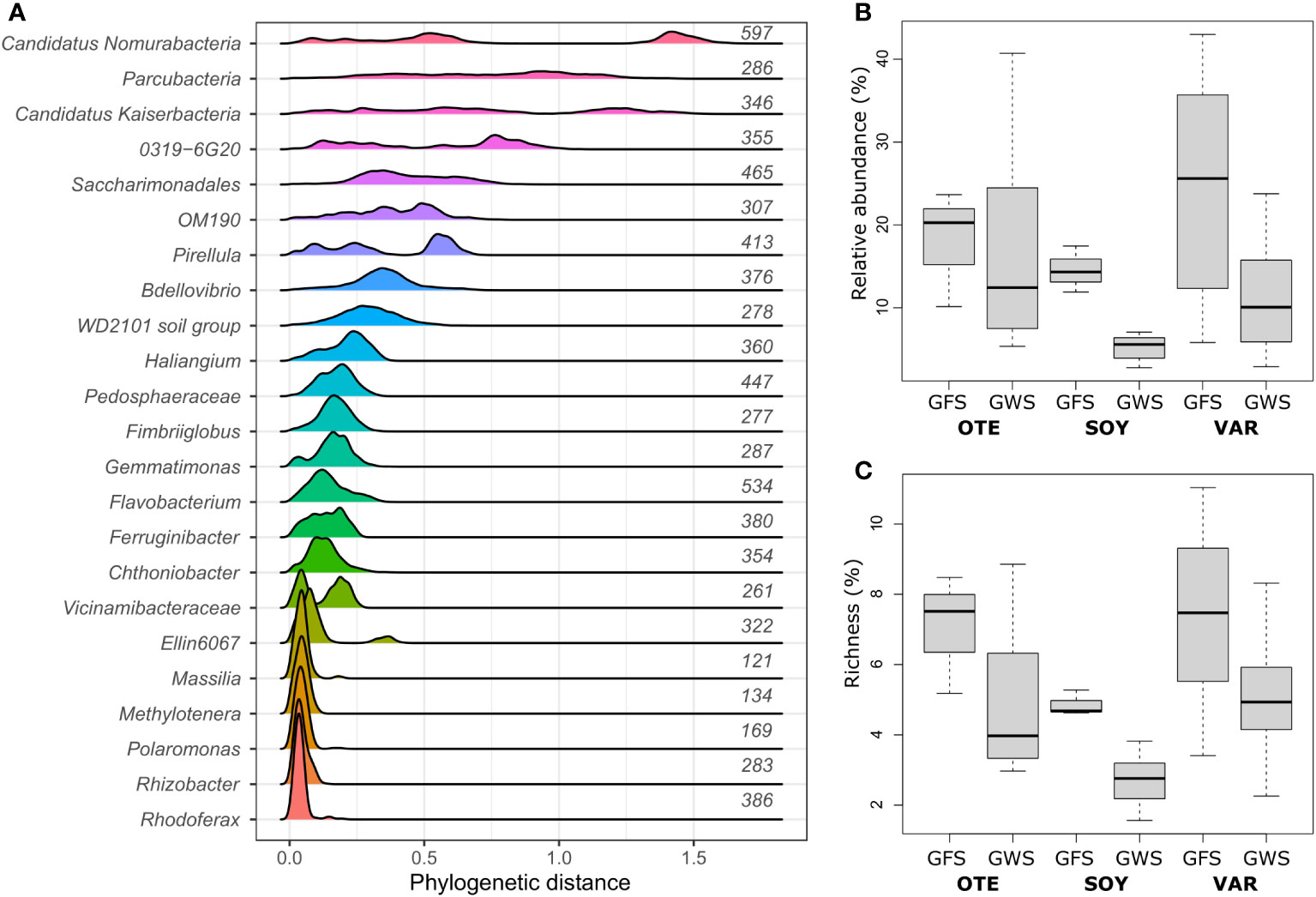
Figure 1 Microdiverse bacterial clades. Microdiverse bacterial genera in proglacial streams are abundant, prevalent and characterized by short pairwise phylogenetic distances among ASVs. Shown are the distributions of phylogenetic distances among ASVs with same genus-level taxonomy for the most abundant genera (A). Note that e.g. Parcubacteria, Saccharimonadales, Pedosphaeraceae, and Vicinamibacteraceae refer to genus-level taxonomic assignments of further unclassified ASVs. The number of ASVs per genus is given to the right of each distribution. Microdiverse genera, i.e. Rhodoferax, Rhizobacter, Polaromonas, Methylotenera, and Massilia, were particularly abundant in GFS (B) while they contributed disproportionately less to overall community richness (C).
We obtained 2732 high-quality bacterial MAGs and previous work unraveled the functional differences between MAGs found in GFS and GWS (Michoud et al., 2023). Here, we used MAGs to identify phage-host interactions and screened them for their anti-phage arsenal. Nevertheless, MAGs, while only resolving a subset of the ASVs-resolved microbial diversity, reflected the composition of bacterial communities (Supplementary Figure S1). Specifically, matching ASVs and MAGs with same genus-level classification, mean relative abundance of MAGs was well correlated with relative abundance of ASVs (Spearman’s rho = 0.65, p<0.01).
Generally, little is known about viral biodiversity in extreme environments, particularly in mountain streams and rivers (Payne et al., 2020; Bekliz et al., 2022; Busi et al., 2022). Here, we identified 1452 high-quality (n=870) or complete (n=582) vMAGs. vMAG genome size varied considerably, with complete vMAG genome size averaging 47.8 kbp (range 2.6 to 373.3 kbp) and high-quality vMAG size averaging 70.4 kbp (range: 5.0 to 622.1 kbp). Taxonomically, viral communities were dominated by viruses classified as Caudoviricetes (Figure 2). Caudoviricetes classified as Ralstonia phage, Tsukubavirus, and Gordonia phage were particularly abundant in GFS, whereas, unclassified viruses, Unahavirus and Koutsourovirus were relatively more abundant in GWS (Supplementary Figure S2A). Besides Caudoviricetes (n=930), Malgrandaviricetes (n=60), and Megaviricetes (n=6) constituted the viral communities.
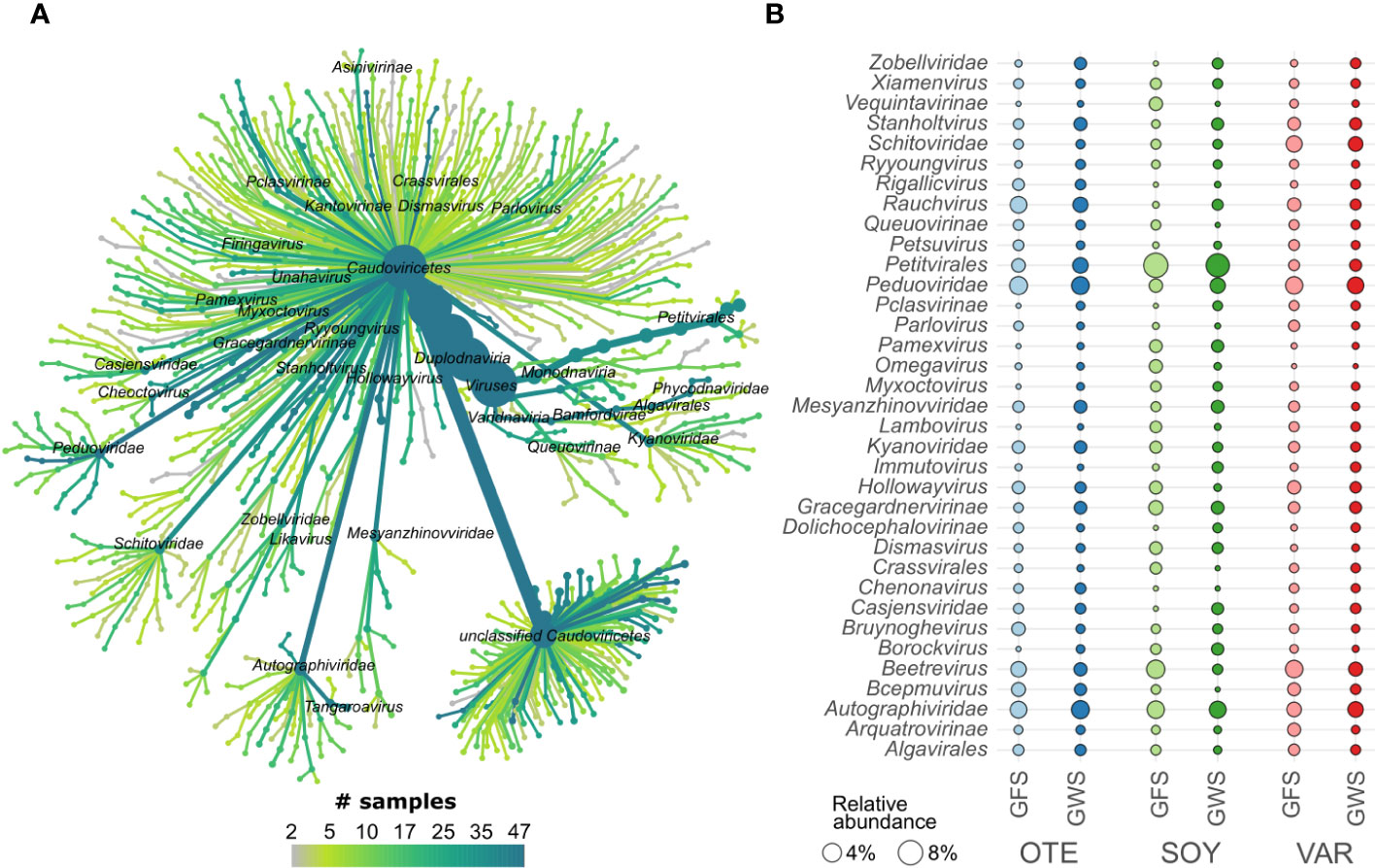
Figure 2 Viral taxonomic composition. vMAGs were classified mostly as members of Caudoviricetes (A). Displayed is a tree-like representation of viral taxonomy. We found high prevalence (the number of samples in which a specific taxonomic group was detected) across several taxonomic groups. The mean relative abundance (circle size, panel B) of the most abundant vMAGs highlights differences across floodplains (e.g. Autographiviridae, Peduoviridae, and the eukaryotic viruses Algalvirales) and differences between GFS and GWS (e.g. Petitvirales, Beetrevirus, and Kyanoviridae; see Supplementary Figure S2A for more detail).
On average, 225.4 ± 74.7 vMAGs were detected in each sample with no significant differences in viral richness across stream types (ANOVA, F=0.85, p=0.38) or floodplains (ANOVA, F=1.35, p= 0.26) (Supplementary Figure S3). Across the three floodplains, 831 vMAGs were present in GFS samples and 1350 vMAGs were present in GWS samples. While 15 vMAGs were present in all samples, 52 vMAGs were found in both stream types and across all three floodplains, forming the viral proglacial stream biofilm core community (Supplementary Figure S2B). These 52 core vMAGs accounted on average for 9.0% of vMAG relative abundance, with a notable overrepresentation in GFS of OTE, where they represented 33.5 ± 12.9% of vMAG relative abundance. A large number of vMAGs were exclusively found in GWS of SOY (n=264), VAR (n=150), and OTE (n=116) (Supplementary Figure S2B). In contrast, fewer vMAGs were exclusively present in GFS of VAR (n=45), SOY (n=31), and OTE (n=19).
Across all floodplains and stream types, most vMAGs were rare (i.e. low abundance) and only few vMAGs were abundant (i.e. reached more than 0.1% of relative abundance in any given sample), yet, these abundant vMAGs accounted on average for 96.8% of total viral relative abundance (Supplementary Figures S3A, C). Screening for lysogeny-associated protein domains (e.g. integrases and recombinases) on complete vMAG genomes (n=582), we found that 81.0% of them featured a virulent lifestyle, whereas 19.0% followed a temperate lifestyle (Supplementary Figure S4). Contrary to Killing-the-winner and Piggyback-the-winner model expectations (Thingstad and Lignell, 1997; Knowles et al., 2016; Silveira and Rohwer, 2016; reviewed by: Chen et al., 2021), we found no relationship between the ratio of temperate to virulent lifestyles (mean ratio: 0.41 ± 0.21) and bacterial abundance, despite a range of two orders of magnitude in bacterial abundance across our samples (Supplementary Figure S4). Additional work would be needed to better constrain these predictions (i.e. extending beyond complete vMAGs and accounting for novelty). Moreover, bulk metagenomics, as compared to viromics, may better capture actively replicating viruses but may offer only limited resolution to resolve non-infecting or rare viruses (i.e. viruses infecting rare hosts (Roux et al., 2021). Using metagenomics, low phage richness has previously been linked to Clustered Regularly Interspaced Short Palindromic Repeats (CRISPER)-Cas abundance (Meaden et al., 2022), suggesting a trade-off exists between this adaptive bacterial immune response and phage diversity. We screened the bacterial MAGs (n=2732) and identified a total of 18,840 antiviral defense genes on 1947 MAGs. Restriction-modification (ngenes=8,705), Cas (ngenes=4,483), abortive infection (ngenes=1,365), and the Septu defense system (ngenes=1,299) were the most commonly found defense types. MAGs contained up to 12 different antiviral defense system types (median number of defense types: 2) highlighting the general importance of antiviral defense for bacterial stream biofilm communities. Moreover, we found a significant relationship between viral diversity and the relative abundance of Cas-containing MAGs (Spearman’s rho = 0.49, p=0.0006, Supplementary Figure S5). Taken together, the diverse viral community and versatile antiviral defense arsenal in proglacial stream biofilms suggests a coevolutionary history of phages and biofilm-dwelling bacteria.
Bacterial community composition based on Bray-Curtis similarity obtained from ASV relative abundance significantly differed between floodplains (PERMANOVA, R2 = 0.08, p<0.001) and stream types (PERMANOVA, R2 = 0.03, p<0.001), with a significant interaction between floodplain and stream type (PERMANOVA, R2 = 0.05, p<0.001). Additional variation can be explained by the spatial positioning of samples (e.g. distance to the glacier snout (Brandani et al., 2022), but substantial variation in the bacterial host community remains inexplicable. This highlights the large spatial heterogeneity among nearby streams in proglacial floodplains and the importance of stochastic community assembly processes in nascent ecosystems (Brandani et al., 2023). Moreover, bacterial communities from the Valsorey Glacier catchment were compositionally more distinct from communities of Otemma and Val Roseg, whereas particularly GWS sites of Otemma and Val Roseg were more similar (Figure 3A). This compositional similarity was mirrored by the viral community (Figure 3B), with significant differences between floodplains (PERMANOVA, R2 = 0.10, p<0.001) and stream types (PERMANOVA, R2 = 0.04, p<0.001), and a significant interaction term (PERMANOVA, R2 = 0.06, p<0.001). Superimposition of non-metric multidimensional scaling ordinations further substantiated the congruence between viral and bacterial communities (Procrustes correlation: 0.74, p=0.001, Supplementary Figure S6). The strength of these associations is comparable to earlier reports on the coupling of stream biofilm bacterial and viral communities (Bekliz et al., 2022) and suggests that host availability dominates over environmental conditions in shaping viral communities.
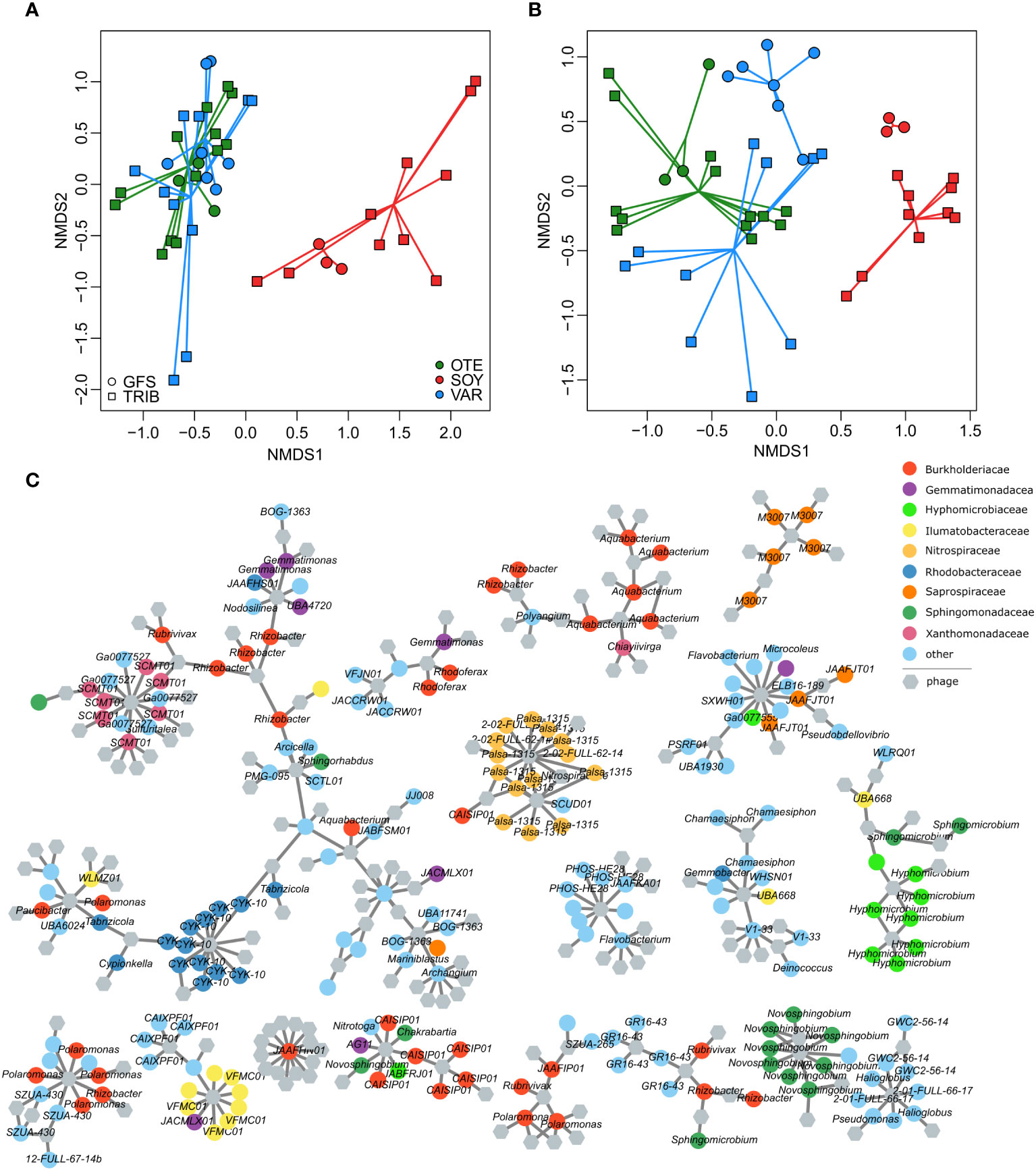
Figure 3 Phage-bacteria coupling. Non-metric multidimensional scaling ordinations based on vMAG relative abundance (A) and bacterial 16S rRNA gene ASV relative abundance (B) reveal the compositional dissimilarities among floodplains and stream types. This separation was more pronounced for the bacterial communities whereas viral communities were more similar (particularly samples of both GFS and GWS in OTE and VAR were similar). Resolving putative phage-host interactions (grey lines) among individual vMAGs (hexagons) and bacterial MAGs (colored circles) illustrate the generally high phage-host specificity (i.e. phages tend to interact with bacteria of similar taxonomy) (C). The 17 largest sub-networks of the entire interaction network are shown.
We used phage-host predictions (PHIST) to further resolve the coupling between bacterial and phage communities and predicted a total of 1435 phage-host interactions. Similar to other work (Luo et al., 2022), the majority of phage-host interactions (84.2%) involved a single phage-host pair and only six phages had more than 10 predicted interactions with different MAGs (maximum number of predicted interactions = 13). In line with generally high host specificity, vMAGs with multiple host interactions tended to interact with taxonomically similar MAGs (Figure 3C). For instance, 11 out of 13 interactions of a vMAG classified as Lacusarxvirus involved the alpha-proteobacterial CYK-10 genus. The other two interactions were predicted for MAGs classified as Sphingomonas (Alphaproteobacteria).
Phage-host interactions involving the microdiverse genera Polaromonas, Rhizobacter, Rhodoferax, Methylotenera, and Massilia were also dominated by single phage-host pairs (66 out of 110 interactions). Phages interacting with Rhizobacter, Polaromonas, and Rhodoferax also interacted with several other bacterial genera (Supplementary Figure S7). For instance, of the 38 vMAGs putatively interacting with Rhizobacter, 22 vMAGs exclusively interacted with Rhizobacter whereas 16 vMAGs also interacted with different bacterial genera. However, this was not consistent for all microdiverse bacterial clades: all of the 19 phages predicted to interact with the microdiverse genus Methylotenera, exclusively interacted with Methylotenera MAGs. Strikingly, only a single phage-host interaction was identified involving the microdiverse genus Massilia. Taken together, we discovered a continuum of potential phage host ranges but mostly found evidence for host specificity in proglacial streams.
While the limitations of in silico phage-host predictions need to be considered in this context (Brum and Sullivan, 2015; de Jonge et al., 2019), we report here a large number of putative phage-host interactions in stream biofilms, in line with reports from other extreme environments (Munson-McGee et al., 2018; Jarett et al., 2020). Moreover, the continuum of phage host range suggests that multiple eco-evolutionary processes shape bacteria-phage interactions in these communities (Weitz et al., 2013; de Jonge et al., 2019). On the one hand, trade-offs between virulence and host range (de Jonge et al., 2019) or resource limitation (Weitz et al., 2013) can explain narrow host ranges, whereas reduced dispersal and the dense packing of diverse bacteria have been associated with broader host ranges in biofilms (Hwang et al., 2023). In communities characterized by microdiversity, narrow host ranges may be beneficial for phages given the increased encounter probability with phylogenetically closely related members. Phages interacting with the microdiverse bacterial genus Methylotenera may, for instance, benefit from such narrow host range. On the other hand, broad host ranges may increase the likelihood of exchanging genetic material across distant bacterial populations. This may, for example, be the case for the microdiverse genus Rhizobacter in proglacial stream biofilms. We may thus hypothesize that beneficial effects of broad host ranges could contribute to the longer-term success of microdiverse bacterial clades. Microdiversity implies that phylogenetically closely related taxa occupy available niches. Typically, such closely related taxa are expected to compete for similar resources (Larkin and Martiny, 2017) and phage-mediated exchange of genetic material, including auxiliary metabolic genes, may alleviate competition among microdiverse clade members.
In total, we detected 938 AMGs on 448 vMAGs, broadly reflecting the taxonomic composition of the entire vMAG community. In contrast to a recent analysis of Pearl River Estuary viromes (Luo et al., 2022), we did not detect differences in the number of AMGs between virulent and temperate phages. Specifically, 318 out of 1041 virulent vMAGs and 130 out of 411 temperate vMAGs harbored at least one AMG. When accounting for viral lifestyle distribution (i.e. 81.0% of vMAGs were virulent), both viral lifestyles represented a similar number of AMGs. Moreover, AMGs abundant among virulent phages were also abundant among temperate phages (Spearman’s rho = 0.72, p<0.0087).
AMGs encoded for a large functional diversity, with in total 166 different KEGG orthologues (KOs) present. Particularly numerous were AMGs encoding dcm (159 AMGs) which in prokaryotes can be involved in DNA methylation in restriction-modification systems (Pingoud et al., 2014). However, in phages, so-called orphan methyltransferases (i.e., without endonucleases of the prokaryotic restriction-modification system) can contribute to the protection of phage DNA against digestion (Heyerhoff et al., 2022). Other common AMGs were queE, queD, queE, and folE genes (137 AMG), involved in the synthesis of the deazapurine nucleoside preQO, which also has been shown to protect viruses from restriction enzymes (Hutinet et al., 2019; Kieft et al., 2020). In contrast to pelagic systems (Kieft et al., 2021b; Heyerhoff et al., 2022) and despite the importance of cross-domain interactions in proglacial stream biofilms (Busi et al., 2022), however, AMGs encoding the core photosystem II proteins psbA and psbD were rare, with only 3 AMGs present across our entire dataset. The majority (34.4%) of AMGs encoded KOs involved in the metabolism of cofactors and vitamins including AMGs involved in folate biosynthesis, the metabolisms of nicotinate/nicotinamide, and porphyrine/chlorophyll. AMGs involved in amino acid metabolism (cysteine and methionine) accounted for another 20.7% of all AMGs. AMGs involved in carbohydrate metabolism (15.4%), glycan biosynthesis (8.4%), and sulfur metabolism (4.3%) were also commonly detected. Similar to marine sediments (Heyerhoff et al., 2022), cysH, which is involved in the synthesis of sulfite during assimilatory sulfate reduction, dominated AMGs involved in sulfur metabolism. cysH ranks among the globally conserved AMGs (Kieft et al., 2020) and has the potential to influence ecosystem-scale sulfur metabolism (Kieft et al., 2021a). This may be particularly relevant in GFS, where high turbidity and the absence of light fosters chemolithoautotrophic bacterial metabolisms. Chemolithoautotrophic bacteria can oxidize reduced sulfur to fix CO2 and to produce biomass. In the absence of photosynthetic organisms in GFS, chemolithoautotrophic carbon production may thus be an important ecosystem function, and phage AMGs may play a pivotal role.
We did not detect significant differences in AMGs between viral communities from different floodplains or stream types (Figure 4A). Abundant AMG families, including the above-mentioned AMGs, tended to be present and abundant in most samples, whereas other AMG families were only sporadically present and often at low relative abundance. However, when coupling AMG with phage-host predictions (17.3% of AMGs were on vMAGs with a host prediction), we found that common and abundant AMGs were found on phages associated with specific bacterial community members (Figure 4B). For instance, 19 AMGs encoding folate biosynthesis were found on phages associated with 6 bacterial genera, including members of Novosphingobium, Calothrix, and Rhodobacter. Similarly, AMGs encoding for cysteine and methionine metabolism were particularly prevalent among vMAGs infecting members of the bacterial genera Polaromonas, Microcoleus, Ferruginibacter, and Segetibacter. Finally, using Principal Component Analysis with all AMGs for which phage-host predictions were available, we found distinct groups of bacterial genera to be host to phages with specific AMGs (Figure 4C). For example, the two microdiverse bacterial genera Methylotenera and Rhizobacter, together with Lysobacter and Sandarakinorhabdus formed a distinct group dominated by AMGs involved in porphyrine and chlorophyll metabolism. Another microdiverse bacterial genus, Rhodoferax, together with several other, non-microdiverse bacterial genera formed a distinct cluster based on AMGs involved in the metabolism of cysteine and methionine. AMGs involved in the biosynthesis of folate and lipopolysaccharides, explained clusters of bacterial genera involving members such as Rhodobacter, Nitrospira, Novosphingobium, and Calothrix as well as a cluster composed of Methylibium, Pedosphaera, and Ideonella. Interestingly, one of the most common and microdiverse bacterial genus in proglacial stream biofilms, Polaromonas, did not associate with any of the clusters. These findings highlight that similar AMGs may be found on phages associated with microdiverse and non-microdiverse clades.
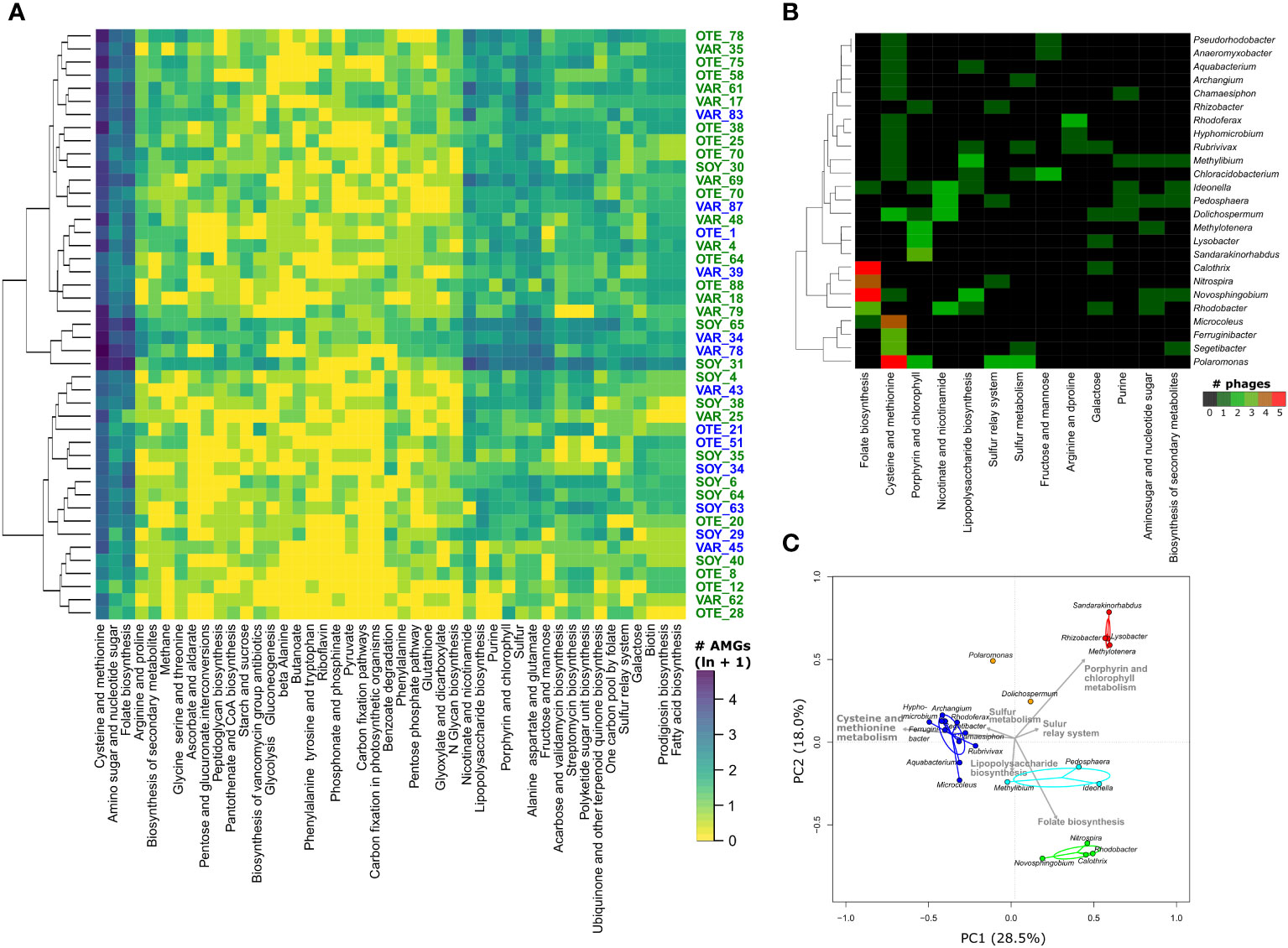
Figure 4 Auxiliary metabolic genes. A heatmap of the number of different AMGs found in the different samples highlights the prevalence of AMGs involved in the metabolism of cysteine and methionine, amino sugars and nucleotide sugar and folate biosynthesis (A). The colors represent ln(x+1) transformed number of AMGs. Despite the pronounced compositional difference between GFS and GWS, no differences in AMG composition between GFS (blue sample IDs) and GWS (green sample IDs) were apparent. The dendrogram to the left of the heatmap shows the results of single-linkage clustering, hierarchically connecting samples based on similar AMG distributions. Coupling AMGs with phage-host interactions revealed AMGs shared by phages putatively infecting bacterial MAGs (B). Shown are the 13 most common AMG-encoded pathways for 25 bacterial genera. Colors reflect the number of vMAGs. Considering all phage-host interactions and AMGs in a Principal Component Analysis, further highlighted the pronounced differences in phages carrying AMGs and infecting specific bacterial genera (C) Based on the number of AMGs on phages, different bacterial host genera (circles) cluster into distinct groups. Colored lines connect host genera to cluster centroids. Ellipses denote 95% confidence intervals for these clusters. Grey arrows denote KO pathways distinctive for the different clusters. For instance, the two microdiverse genera Rhizobacter and Methylotenera cluster with Sandarakinorhabdus and Lysobacter in terms of AMGs found on their respective phages. These AMGs are enriched in genes encoding porphyrin and chlorophyll metabolism. Note the distribution of microdiverse bacterial genera in different clusters and the absence of the microdiverse bacterial genus Massilia.
Despite their compact genomes, phages regularly encode AMGs which can provide them a fitness advantage by augmenting or redirecting specific metabolic processes (Dion et al., 2020; Kieft et al., 2021b). This highlights the reciprocal benefits of phage-host interactions, which manifest during coevolution (Koskella and Brockhurst, 2014). The selective environmental constraints in proglacial streams have persisted over geological time scales, shaping microbial diversity and putatively influencing phage-bacteria coevolution. Host specificity of AMGs has been previously reported (Luo et al., 2022) and here we report specific AMGs linked with microdiverse (and non-microdiverse) clades in proglacial stream biofilms. Arguably, microdiverse clades and their phages have shared a long evolutionary time in a selective environment, refining the AMGs library and putatively contributing to the eco-evolutionary success of both, the phages and their prokaryotic hosts. Microdiversity is an intrinsic property of many microbial communities and horizontal gene transfer can generate microdiversity (Larkin and Martiny, 2017). Phage-mediated HGT transfer may therefore contribute to microdiversification and narrow host range and AMG specificity suggests that in proglacial stream biofilms, virus-host interactions may foster ‘Maestro Microbes’ (Larkin and Martiny, 2017), which occupy niches by optimizing specific traits (in contrast to ‘Renaissance Microbes’ which acquire new traits (Larkin and Martiny, 2017).
Generally, little is known about viral biodiversity in extreme environments, particularly however in streams and rivers draining mountain regions (Payne et al., 2020; Bekliz et al., 2022; Busi et al., 2022). Here, we expand the current knowledge of phage diversity and its role in shaping microbial communities in streams and rivers. Biofilms are thought to represent a physical barrier for viruses (Abedon, 2011), and our work unravels viral communities, which are shaped by their bacterial host community composition. Extreme environmental conditions in proglacial streams lead to strong selective sorting of these host communities, ultimately resulting in communities dominated by few but well-adapted and microdiverse clades. Contrary to our expectation of generally narrow phage-host ranges in communities dominated by microdiverse bacterial clades, we found a continuum of host ranges, which we attribute to different eco-evolutionary dynamics, putatively associated with resource limitation in proglacial streams (explaining narrow host ranges) and the biofilm mode of life (putatively explaining broader host ranges). Phages infecting members of different microdiverse bacterial genera carry AMGs, which appear to be distinct and specific. Future work may further elucidate the role of non-microdiverse clades, which share similar AMGs and assess to which degree the coevolution between phages and their hosts shapes the AMG repertoire. This may be particularly relevant in extreme environments, where stark selective processes shape communities. On the other hand, microdiversity is a hallmark of many microbial communities, and beneficial phages-host interactions may play important roles in generating and maintaining microdiversity.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: https://www.ncbi.nlm.nih.gov/, PRJNA808857. Supplementary Table S1 lists individual BioSample accession numbers. Raw bioinformatic output is available at Figshare under doi 10.6084/m9.figshare.24511972.
HP: Conceptualization, Formal Analysis, Investigation, Methodology, Visualization, Writing – original draft. GM: Conceptualization, Data curation, Formal Analysis, Investigation, Methodology, Writing – original draft. SB: Conceptualization, Data curation, Formal Analysis, Investigation, Methodology, Writing – original draft. TB: Conceptualization, Funding acquisition, Project administration, Resources, Supervision, Writing – original draft.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. Funding supporting this work was provided by the Swiss National Science Foundation (grant CRSII5_180241) to TJB.
We thank Jade Brandani, Nicola Deluigi, Kevin Casellini, and Paraskevi Pramateftaki for assistance in the field and laboratory. We further thank Stylianos Fodelianakis, Tyler Kohler, Massimo Bourquin, Leila Ezzat, Martin Boutroux, and two reviewers for fruitful discussions and feedback on the manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) SB, GM declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/frmbi.2023.1279550/full#supplementary-material
Abedon S. T. (2011). Bacteriophages and biofilms: ecology, phage therapy, plaques. Nova Science Publishers.
Alneberg J., Bjarnason B. S., de Bruijn I., Schirmer M., Quick J., Ijaz U. Z., et al. (2014). Binning metagenomic contigs by coverage and composition. Nat. Methods 11, 1144–1146. doi: 10.1038/nmeth.3103
Andrews S. (2010) FastQC: A Quality Control Tool for High Throughput Sequence Data. Available at: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/.
Battin T. J., Besemer K., Bengtsson M. M., Romani A. M., Packmann A. I. (2016). The ecology and biogeochemistry of stream biofilms. Nat. Rev. Microbiol. 14, 251–263. doi: 10.1038/nrmicro.2016.15
Bekliz M., Pramateftaki P., Battin T. J., Peter H. (2022). Viral diversity is linked to bacterial community composition in alpine stream biofilms. ISME Commun 2, 27. doi: 10.1038/s43705-022-00112-9
Bolger A. M., Lohse M., Usadel B. (2014). Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics 30, 2114–2120. doi: 10.1093/bioinformatics/btu170
Bolyen E., Rideout J. R., Dillon M. R., Bokulich N. A., Abnet C. C., Al-Ghalith G. A., et al. (2019). Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 37, 852–857. doi: 10.1038/s41587-019-0209-9
Brandani J., Peter H., Busi S. B., Kohler T. J., Fodelianakis S., Ezzat L., et al. (2022). Spatial patterns of benthic biofilm diversity among streams draining proglacial floodplains. Front. Microbiol. 13. doi: 10.3389/fmicb.2022.948165
Brandani J., Peter H., Fodelianakis S., Kohler T. J., Bourquin M., Michoud G., et al. (2023). Homogeneous environmental selection structures the bacterial communities of benthic biofilms in proglacial floodplain streams. Appl. Environ. Microbiol. 89, e02010–e02022. doi: 10.1128/aem.02010-22
Brum J. R., Sullivan M. B. (2015). Rising to the challenge: accelerated pace of discovery transforms marine virology. Nat. Rev. Microbiol. 13, 147–159. doi: 10.1038/nrmicro3404
Busi S. B., Bourquin M., Fodelianakis S., Michoud G., Kohler T. J., Peter H., et al. (2022). Genomic and metabolic adaptations of biofilms to ecological windows of opportunity in glacier-fed streams. Nat. Commun. 13, 2168. doi: 10.1038/s41467-022-29914-0
Busi S. B., Pramateftaki P., Brandani J., Fodelianakis S., Peter H., Halder R., et al. (2020). Optimised biomolecular extraction for metagenomic analysis of microbial biofilms from high-mountain streams. PeerJ 8, e9973. doi: 10.7717/peerj.9973
Callahan B. J., McMurdie P. J., Rosen M. J., Han A. W., Johnson A. J. A., Holmes S. P. (2016). DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 13, 581–583. doi: 10.1038/nmeth.3869
Chaumeil P.-A., Mussig A. J., Hugenholtz P., Parks D. H. (2019). GTDB-Tk: a toolkit to classify genomes with the Genome Taxonomy Database. Bioinformatics 36, 1925–1927. doi: 10.1093/bioinformatics/btz848
Chen X., Weinbauer M. G., Jiao N., Zhang R. (2021). Revisiting marine lytic and lysogenic virus-host interactions: Kill-the-Winner and Piggyback-the-Winner. Sci. Bull. 66, 871–874. doi: 10.1016/j.scib.2020.12.014
de Jonge P. A., Nobrega F. L., Brouns S. J. J., Dutilh B. E. (2019). Molecular and evolutionary determinants of bacteriophage host range. Trends Microbiol. 27, 51–63. doi: 10.1016/j.tim.2018.08.006
Dion M. B., Oechslin F., Moineau S. (2020). Phage diversity, genomics and phylogeny. Nat. Rev. Microbiol. 18, 125–138. doi: 10.1038/s41579-019-0311-5
Ezzat L., Fodelianakis S., Kohler T. J., Bourquin M., Brandani J., Busi S. B., et al. (2022). Benthic biofilms in glacier-fed streams from Scandinavia to the Himalayas host distinct bacterial communities compared with the streamwater. Appl. Environ. Microbiol. 88, e00421–e00422. doi: 10.1128/aem.00421-22
Fodelianakis S., Washburne A. D., Bourquin M., Pramateftaki P., Kohler T. J., Styllas M., et al. (2022). Microdiversity characterizes prevalent phylogenetic clades in the glacier-fed stream microbiome. ISME J. 16, 666–675. doi: 10.1038/s41396-021-01106-6
Foster Z. S. L., Sharpton T. J., Grünwald N. J. (2017). Metacoder: An R package for visualization and manipulation of community taxonomic diversity data. PLoS Comput. Biol. 13, e1005404. doi: 10.1371/journal.pcbi.1005404
Freimann R., Bürgmann H., Findlay S. E., Robinson C. T. (2013). Bacterial structures and ecosystem functions in glaciated floodplains: contemporary states and potential future shifts. ISME J. 7, 2361–2373. doi: 10.1038/ismej.2013.114
Gregory A. C., Zayed A. A., Conceição-Neto N., Temperton B., Bolduc B., Alberti A., et al. (2019). Marine DNA viral macro- and microdiversity from pole to pole. Cell 177, 1109–1123.e14. doi: 10.1016/j.cell.2019.03.040
Heyerhoff B., Engelen B., Bunse C. (2022). Auxiliary metabolic gene functions in pelagic and benthic viruses of the Baltic Sea. Front. Microbiol. 13. doi: 10.3389/fmicb.2022.863620
Hockenberry A. J., Wilke C. O. (2021). BACPHLIP: predicting bacteriophage lifestyle from conserved protein domains. PeerJ 9, e11396. doi: 10.7717/peerj.11396
Holtappels D., Alfenas-Zerbini P., Koskella B. (2023). Drivers and consequences of bacteriophage host range. FEMS Microbiol. Rev. 47, fuad038. doi: 10.1093/femsre/fuad038
Hotaling S., Hood E., Hamilton T. L. (2017). Microbial ecology of mountain glacier ecosystems: biodiversity, ecological connections and implications of a warming climate. Environ. Microbiol. 19, 2935–2948. doi: 10.1111/1462-2920.13766
Howard-Varona C., Hargreaves K. R., Abedon S. T., Sullivan M. B. (2017). Lysogeny in nature: mechanisms, impact and ecology of temperate phages. ISME J. 11, 1511–1520. doi: 10.1038/ismej.2017.16
Hutinet G., Kot W., Cui L., Hillebrand R., Balamkundu S., Gnanakalai S., et al. (2019). 7-Deazaguanine modifications protect phage DNA from host restriction systems. Nat. Commun. 10, 5442. doi: 10.1038/s41467-019-13384-y
Hwang Y., Roux S., Coclet C., Krause S. J. E., Girguis P. R. (2023). Viruses interact with hosts that span distantly related microbial domains in dense hydrothermal mats. Nat. Microbiol. 8, 946–957. doi: 10.1038/s41564-023-01347-5
Jarett J. K., Džunková M., Schulz F., Roux S., Paez-Espino D., Eloe-Fadrosh E., et al. (2020). Insights into the dynamics between viruses and their hosts in a hot spring microbial mat. ISME J. 14, 2527–2541. doi: 10.1038/s41396-020-0705-4
Johansen J., Plichta D. R., Nissen J. N., Jespersen M. L., Shah S. A., Deng L., et al. (2022). Genome binning of viral entities from bulk metagenomics data. Nat. Commun. 13, 965. doi: 10.1038/s41467-022-28581-5
Kang D. D., Li F., Kirton E., Thomas A., Egan R., An H., et al. (2019). MetaBAT 2: an adaptive binning algorithm for robust and efficient genome reconstruction from metagenome assemblies. PeerJ 7, e7359. doi: 10.7717/peerj.7359
Kieft K., Adams A., Salamzade R., Kalan L., Anantharaman K. (2022). vRhyme enables binning of viral genomes from metagenomes. Nucleic Acids Res. 50, e83. doi: 10.1093/nar/gkac341
Kieft K., Breister A. M., Huss P., Linz A. M., Zanetakos E., Zhou Z., et al. (2021a). Virus-associated organosulfur metabolism in human and environmental systems. Cell Rep. 36, 109471. doi: 10.1016/j.celrep.2021.109471
Kieft K., Zhou Z., Anantharaman K. (2020). VIBRANT: automated recovery, annotation and curation of microbial viruses, and evaluation of viral community function from genomic sequences. Microbiome 8, 90. doi: 10.1186/s40168-020-00867-0
Kieft K., Zhou Z., Anderson R. E., Buchan A., Campbell B. J., Hallam S. J., et al. (2021b). Ecology of inorganic sulfur auxiliary metabolism in widespread bacteriophages. Nat. Commun. 12, 3503. doi: 10.1038/s41467-021-23698-5
Klindworth A., Pruesse E., Schweer T., Peplies J., Quast C., Horn M., et al. (2013). Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 41, e1. doi: 10.1093/nar/gks808
Knowles B., Silveira C. B., Bailey B. A., Barott K., Cantu V. A., Cobián-Güemes A. G., et al. (2016). Lytic to temperate switching of viral communities. Nature 531, 466–470. doi: 10.1038/nature17193
Koskella B., Brockhurst M. A. (2014). Bacteria-phage coevolution as a driver of ecological and evolutionary processes in microbial communities. FEMS Microbiol. Rev. 38, 916–931. doi: 10.1111/1574-6976.12072
Krueger F., James F., Ewels P., Afyounian E., Schuster-Boeckler B. (2021). TrimGalore: v0.6.7. doi: 10.5281/zenodo.5127899
Larkin A. A., Martiny A. C. (2017). Microdiversity shapes the traits, niche space, and biogeography of microbial taxa. Environ. Microbiol. Rep. 9, 55–70. doi: 10.1111/1758-2229.12523
Li D., Liu C.-M., Luo R., Sadakane K., Lam T.-W. (2015). MEGAHIT: an ultra-fast single-node solution for large and complex metagenomics assembly via succinct de Bruijn graph. Bioinformatics 31, 1674–1676. doi: 10.1093/bioinformatics/btv033
Love M. I., Huber W., Anders S. (2014). Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 550. doi: 10.1186/s13059-014-0550-8
Luo X.-Q., Wang P., Li J.-L., Ahmad M., Duan L., Yin L.-Z., et al. (2022). Viral community-wide auxiliary metabolic genes differ by lifestyles, habitats, and hosts. Microbiome 10, 190. doi: 10.1186/s40168-022-01384-y
Martin M. (2011). Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet.journal 17, 10–12. doi: 10.14806/ej.17.1.200
Meaden S., Biswas A., Arkhipova K., Morales S. E., Dutilh B. E., Westra E. R., et al. (2022). High viral abundance and low diversity are associated with increased CRISPR-Cas prevalence across microbial ecosystems. Curr. Biol. 32, 220–227.e5. doi: 10.1016/j.cub.2021.10.038
Menzel P., Ng K. L., Krogh A. (2016). Fast and sensitive taxonomic classification for metagenomics with Kaiju. Nat. Commun. 7, 11257. doi: 10.1038/ncomms11257
Michoud G., Kohler T. J., Peter H., Brandani J., Busi S. B., Battin T. J. (2023). Unexpected functional diversity of stream biofilms within and across proglacial floodplains despite close spatial proximity. Limnology Oceanography 68, 2183–2194. doi: 10.1002/lno.12415
Munson-McGee J. H., Peng S., Dewerff S., Stepanauskas R., Whitaker R. J., Weitz J. S., et al. (2018). A virus or more in (nearly) every cell: ubiquitous networks of virus-host interactions in extreme environments. ISME J. 12, 1706–1714. doi: 10.1038/s41396-018-0071-7
Nayfach S., Camargo A. P., Schulz F., Eloe-Fadrosh E., Roux S., Kyrpides N. C. (2021). CheckV assesses the quality and completeness of metagenome-assembled viral genomes. Nat. Biotechnol. 39, 578–585. doi: 10.1038/s41587-020-00774-7
Needham D. M., Sachdeva R., Fuhrman J. A. (2017). Ecological dynamics and co-occurrence among marine phytoplankton, bacteria and myoviruses shows microdiversity matters. ISME J. 11, 1614–1629. doi: 10.1038/ismej.2017.29
Nissen J. N., Johansen J., Allesøe R. L., Sønderby C. K., Armenteros J. J. A., Grønbech C. H., et al. (2021). Improved metagenome binning and assembly using deep variational autoencoders. Nat. Biotechnol. 39, 555–560. doi: 10.1038/s41587-020-00777-4
Obeng N., Pratama A. A., van Elsas J. D. (2016). The significance of mutualistic phages for bacterial ecology and evolution. Trends Microbiol. 24, 440–449. doi: 10.1016/j.tim.2015.12.009
Oksanen J., Simpson G. L., Blanchet G. F., Kindt R., Legendre P., et al. (2022) vegan: Community ecology package. R package version 2.6-4. Available at: https://CRAN.R-project.org/package=vegan.
Olm M. R., Brown C. T., Brooks B., Banfield J. F. (2017). dRep: a tool for fast and accurate genomic comparisons that enables improved genome recovery from metagenomes through de-replication. ISME J. 11, 2864–2868. doi: 10.1038/ismej.2017.126
Parks D. H., Imelfort M., Skennerton C. T., Hugenholtz P., Tyson G. W. (2015). CheckM: assessing the quality of microbial genomes recovered from isolates, single cells, and metagenomes. Genome Res. 25, 1043–1055. doi: 10.1101/gr.186072.114
Payne A. T., Davidson A. J., Kan J., Peipoch M., Bier R., Williamson K. (2020). Widespread cryptic viral infections in lotic biofilms. Biofilm 2, 100016. doi: 10.1016/j.bioflm.2019.100016
Pingoud A., Wilson G. G., Wende W. (2014). Type II restriction endonucleases–a historical perspective and more. Nucleic Acids Res. 42, 7489–7527. doi: 10.1093/nar/gku447
Quast C., Pruesse E., Yilmaz P., Gerken J., Schweer T., Yarza P., et al. (2013). The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Res. 41, D590–D596. doi: 10.1093/nar/gks1219
R Core Team (2021). R: A language and environment for statistical computing (Vienna, Austria: R Foundation for Statistical Computing). Available at: https://www.R-project.org/.
Ren J., Song K., Deng C., Ahlgren N. A., Fuhrman J. A., Li Y., et al. (2020). Identifying viruses from metagenomic data using deep learning. Quantitative Biol. 8, 64–77. doi: 10.1007/s40484-019-0187-4
Roux S., Brum J. R., Dutilh B. E., Sunagawa S., Duhaime M. B., Loy A., et al. (2016). Ecogenomics and potential biogeochemical impacts of globally abundant ocean viruses. Nature 537, 689–693. doi: 10.1038/nature19366
Roux S., Matthijnssens J., Dutilh B. E. (2021). Metagenomics in virology. Encyclopedia Virol. Dennis H. Bamford, Mark Zuckerman, Encyclopedia of Virology (Fourth Edition), Academic Press. 133–140. doi: 10.1016/B978-0-12-809633-8.20957-6
Schwartz D. A., Lehmkuhl B. K., Lennon J. T. (2022). Phage-encoded sigma factors alter bacterial dormancy. mSphere 7, e00297–e00222. doi: 10.1128/msphere.00297-22
Shannon P., Markiel A., Ozier O., Baliga N. S., Wang J. T., Ramage D., et al. (2003). Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res. 13, 2498–2504. doi: 10.1101/gr.1239303
Shkoporov A. N., Turkington C. J., Hill C. (2022). Mutualistic interplay between bacteriophages and bacteria in the human gut. Nat. Rev. Microbiol. 20, 737–749. doi: 10.1038/s41579-022-00755-4
Sieber C. M. K., Probst A. J., Sharrar A., Thomas B. C., Hess M., Tringe S. G., et al. (2018). Recovery of genomes from metagenomes via a dereplication, aggregation and scoring strategy. Nat. Microbiol. 3, 836–843. doi: 10.1038/s41564-018-0171-1
Silveira C. B., Rohwer F. L. (2016). Piggyback-the-Winner in host-associated microbial communities. NPJ Biofilms Microbiomes 2, 1–5. doi: 10.1038/npjbiofilms.2016.10
Stamatakis A. (2014). RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 30, 1312–1313. doi: 10.1093/bioinformatics/btu033
Suttle C. (1994). The significance of viruses to mortality in aquatic microbial communities. Microbial Ecol. 28, 237–243. doi: 10.1007/BF00166813
Suttle C. A. (2007). Marine viruses — major players in the global ecosystem. Nat. Rev. Microbiol. 5, 801–812. doi: 10.1038/nrmicro1750
Tesson F., Hervé A., Mordret E., Touchon M., d’Humières C., Cury J., et al. (2022). Systematic and quantitative view of the antiviral arsenal of prokaryotes. Nat. Commun. 13, 2561. doi: 10.1038/s41467-022-30269-9
Thingstad T., Lignell R. (1997). Theoretical models for the control of bacterial growth rate, abundance, diversity and carbon demand. Aquat. Microbial Ecol. 13, 19–27. doi: 10.3354/ame013019
Weitz J. S., Poisot T., Meyer J. R., Flores C. O., Valverde S., Sullivan M. B., et al. (2013). Phage-bacteria infection networks. Trends Microbiol. 21, 82–91. doi: 10.1016/j.tim.2012.11.003
Zielezinski A., Deorowicz S., Gudyś A. (2022). PHIST: fast and accurate prediction of prokaryotic hosts from metagenomic viral sequences. Bioinformatics 38, 1447–1449. doi: 10.1093/bioinformatics/btab837
Keywords: phage-host interactions, host range, auxiliary metabolic genes, environmental selection, coevolution
Citation: Peter H, Michoud G, Busi SB and Battin TJ (2024) The role of phages for microdiverse bacterial communities in proglacial stream biofilms. Front. Microbiomes 2:1279550. doi: 10.3389/frmbi.2023.1279550
Received: 18 August 2023; Accepted: 22 December 2023;
Published: 15 January 2024.
Edited by:
Janina Rahlff, Friedrich Schiller University Jena, GermanyReviewed by:
Ovidiu-Nicolae Popa, Heinrich Heine University Düsseldorf, GermanyCopyright © 2024 Peter, Michoud, Busi and Battin. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Hannes Peter, aGFubmVzLnBldGVyQGVwZmwuY2g=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.