 Ehab Kotb Elmahallawy1,2*
Ehab Kotb Elmahallawy1,2* Ahmed Gareh3
Ahmed Gareh3 Marwa M. I. Ghallab4Pamela C. Köster5,6,7
Marwa M. I. Ghallab4Pamela C. Köster5,6,7 Alejandro Dashti5Dina Aboelsoued8Nagwa Ibrahim Toaleb8Hind Alzaylaee9
Alejandro Dashti5Dina Aboelsoued8Nagwa Ibrahim Toaleb8Hind Alzaylaee9 Moisés Gonzálvez2,10
Moisés Gonzálvez2,10 Amira A. Saleh11
Amira A. Saleh11 Alaa S. Alhegaili12Ahmed Fathy Eldehn13
Alaa S. Alhegaili12Ahmed Fathy Eldehn13 Carolina Hernández-Castro5,14
Carolina Hernández-Castro5,14 Begoña Bailo5
Begoña Bailo5 David González-Barrio5
David González-Barrio5 David Carmena5,15*
David Carmena5,15*- 1Department of Zoonoses, Faculty of Veterinary Medicine, Sohag University, Sohag, Egypt
- 2Departamento de Sanidad Animal, Grupo de Investigación en Sanidad Animal y Zoonosis (GISAZ), Universidad de Córdoba, Córdoba, Spain
- 3Department of Parasitology, Faculty of Veterinary Medicine, Aswan University, Aswan, Egypt
- 4Department of Medical Parasitology, Faculty of Medicine, Kafrelsheikh University, Kafr El Sheikh, Egypt
- 5Parasitology Reference and Research Laboratory, National Centre for Microbiology, Majadahonda, Spain
- 6Faculty of Health Sciences, Alfonso X El Sabio University (UAX), Villanueva de la Cañada, Madrid, Spain
- 7Faculty of Medicine, Alfonso X El Sabio University (UAX), Villanueva de la Cañada, Madrid, Spain
- 8Department of Parasitology and Animal Diseases, Veterinary Research Institute, National Research Centre, Cairo, Egypt
- 9Department of Biology, College of Science, Princess Nourah bint Abdulrahman University, Riyadh, Saudi Arabia
- 10Departamento de Sanidad Animal, Facultad de Veterinaria, Campus de Excelencia Internacional Regional “Campus Mare Nostrum”, Universidad de Murcia, Murcia, Spain
- 11Department of Medical Parasitology, Faculty of Medicine, Zagazig University, Zagazig, Egypt
- 12Department of Medical Laboratory, College of Applied Medical Sciences, Prince Sattam bin Abdul Aziz University, Alkharj, Saudi Arabia
- 13Department of Otorhinolaryngology, Kasr Al-Ainy Medical School, Cairo University, Cairo, Egypt
- 14Parasitology Group, Faculty of Medicine, Academic Corporation for the Study of Tropical Pathologies, University of Antioquia, Medellín, Colombia
- 15Centre for Biomedical Research in Infectious Diseases (CIBER), Carlos III Health Institute (ISCIII), Madrid, Spain
Introduction: Giardiosis remains one of the most prevalent enteric parasitic infections globally. Earlier molecular-based studies conducted in Egypt have primarily focused on paediatric clinical populations and most were based on single genotyping markers. As a result, there is limited information on the frequency and genetic diversity of G. duodenalis infections in individuals of all age groups.
Methods: Individual stool samples (n = 460) from outpatients seeking medical care were collected during January–December 2021 in Kafr El-Sheikh governorate, northern Egypt. Initial screening for the presence of G. duodenalis was conducted by coprological examination. Microscopy-positive samples were further confirmed by real-time PCR. A multilocus sequence typing approach targeted amplification of the glutamate dehydrogenase (gdh), beta-giardin (bg), and triose phosphate isomerase (tpi) genes was used for genotyping purposes. A standardised epidemiological questionnaire was used to gather basic sociodemographic and clinical features of the recruited patients.
Results: Giardia duodenalis cysts were observed in 5.4% (25/460, 95% CI: 3.6–7.9) of the stool samples examined by conventional microscopy. The infection was more frequent in children under the age of 10 years and in individuals presenting with diarrhoea but without reaching statistical significance. Stool samples collected during the winter period were more likely to harbour G. duodenalis. All 25 microscopy-positive samples were confirmed by real-time PCR, but genotyping data was only available for 56.0% (14/25) of the isolates. Sequence analyses revealed the presence of assemblages A (78.6%, 11/14) and B (21.4%, 3/14). All assemblage A isolates were identified as sub-assemblage AII, whereas the three assemblage B sequences belonged to the sub-assemblage BIII. Patients with giardiosis presenting with diarrhoea were more frequently infected by the assemblage A of the parasite.
Conclusion: This is one of the largest epidemiological studies evaluating G. duodenalis infection in individuals of all age groups in Egypt. Our molecular data suggest that G. duodenalis infections in the surveyed population are primarily of anthropic origin. However, because assemblages A and B are zoonotic, some of the infections identified can have an animal origin. Additional investigations targeting animal (domestic and free-living) and environmental (water) samples are warranted to better understand the epidemiology of giardiosis in Egypt.
1 Introduction
Giardiosis by Giardia duodenalis (syn. G. lamblia, G. intestinalis) is the most frequently reported intestinal protozoan infection worldwide, with an estimated 280 million people being infected each year (1). The pathogen has a ubiquitous distribution, although most human cases occur predominantly in humid and temperate regions (2–4). As other members of the Giardia genus, G. duodenalis has a direct life cycle (monoxenus) without intermediate hosts. Infection occurs by ingestion of the transmissive cyst stage, either directly through contact with infected individuals or animals or indirectly from food or water contaminated with faecal material. In the duodenum, excystation releases vegetative trophozoites (two per cyst) into the small intestinal lumen, where they divide by binary fission and attach to the host epithelium via a ventral adhesive disc. Encystation of the trophozoite to the cyst stage occur in the colon and is promoted by exposure to bile. The life cycle is completed when cysts are excreted into the environment with the faeces of infected hosts (5). Giardia duodenalis cysts are environmental-resistant and can stay viable for prolonged periods of time under appropriate conditions of humidity and temperature. Giardia duodenalis cysts are also resistant to chemical disinfectants (e.g., chlorine and ozone) commonly used in the water industry (6, 7).
The clinical manifestations of human giardiosis vary from asymptomatic carriage to a wide diversity of symptoms including acute or chronic diarrhoea, dryness, abdominal discomfort, nausea, vomiting, flatulence, weight loss, and anaemia resulting from vitamin A, iron, and zinc deficiencies (8–10). In poor-resource areas giardiosis has been associated with impairment of growth and cognitive development during childhood (11). The pathogenic effect of giardiosis occurs through multiple mechanisms including functional disruption of the epithelial barrier, shortening of brush border microvilli, gut microbiota dysbiosis, apoptosis of enterocytes, electrolyte/nutrient/water malabsorption, and anion hypersecretion, among others (12, 13). The severity of the disease is possibly dogged by multiple factors such as the virulence of the parasite (14), the immunological and nutritional status of the host (15), the occurrence of coinfection with other pathogens (16), and the frequency of cases refractory to treatment (17). Young children, the older adults, and immunocompromised individuals are more susceptible to the infection (18, 19). In developing countries, infections by G. duodenalis are concomitant with poor sanitary conditions, poor water quality, and overcrowding (20, 21).
Diagnosis of giardiosis is primarily conducted by microscopy analysis of stool samples in most clinical microbiology laboratories, particularly in poor-resource settings (22). In recent years, PCR-based methods have been increasingly shown to be sensitive and cost-effective alternatives to microscopy examination of stools. Advantages of molecular assays over conventional microscopy include highly sensitive and specific identification of parasite DNA and rapid turnover with little hands-on work (23–26). When coupled to Sanger or next-generation amplicon sequencing, PCR methods allow species and genotype confirmation (27). Nucleotide sequence analyses of the small subunit ribosomal RNA (ssu rRNA), glutamate dehydrogenase (gdh), beta-giardin (bg), and triose phosphate isomerase (tpi) genes have evidenced that G. duodenalis is indeed a multi-species complex composed of eight (A-H) assemblages that differ widely in host range and specificity, with assemblages A and B responsible for ~95% of human infections (4, 28). Canine-adapted assemblages C-D, feline-adapted assemblage F, and ungulate-adapted assemblage E can occasionally cause human infections (4, 28). Giardiosis is endemic in Egypt. Human infections have been estimated at 9.2–80.2% by conventional microscopy, at 19.0–38.4% by ELISA, at 7.9% by rapid diagnostic tests, and at 11.5–42.3% by PCR (Table 1) (29–53). Most of the epidemiological studies were carried out on paediatric populations presenting with gastrointestinal manifestations. Molecular studies demonstrated that assemblage B (55.3%, 587/1,061) is more prevalent than assemblage A (36.9%, 392/1,061), with assemblage E (2.7%, 29/1,061), assemblage C (0.1%, 1/1,061) and mixed (4.9%, 52/1,061) infections being also detected at much lower rates (Table 1) (29–53). This study aimed to determine the prevalence and molecular diversity of G. duodenalis in outpatients seeking medical care in northern Egypt. To do so, conventional microscopy was used as screening method and molecular (PCR and Sanger sequencing) methods as confirmatory and genotyping methods.
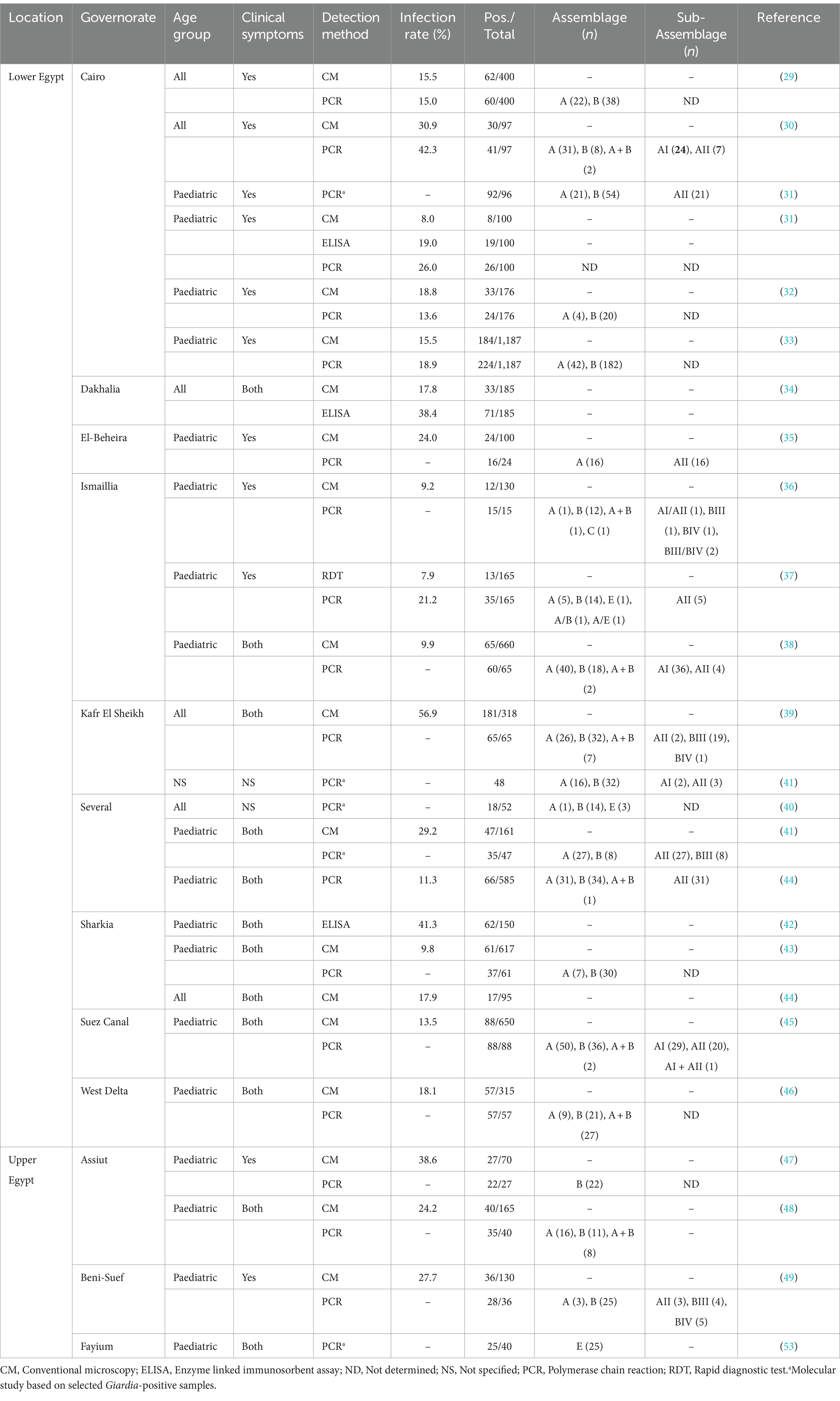
Table 1. Occurrence and genetic diversity of Giardia duodenalis in human populations in Egypt.
2 Materials and methods
2.1 Ethical statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by the Kafrelsheikh University Research Ethics Committee (protocol code MKSU 50–1-10). Signed informed consents were obtained from all patients that volunteer to participate in the study. Stool samples and associated epidemiological and clinical data were anonymized to protect the privacy of the participants.
2.2 Patient recruitment and sample collection
Single fresh stool specimens from outpatients seeking medical attention at the Kafrelsheikh University Hospital (Kafr El-Sheikh governorate, Egypt) were collected during the period January–December 2021 without specific exclusion criteria. Each stool sample was collected in a screw-capped container and labelled with a unique identifier code. Information regarding sex, age, location, and sampling date were also recorded. Faecal consistency was categorised as either diarrheic or formed. Samples were submitted for microscopy examination within 3 h of collection.
2.3 Microscopy examination
2.3.1 Direct wet smear
Direct saline (0.85% NaCl) and Lugol’s iodine wet mounts were prepared from freshly passed stool specimens for the detection of G. duodenalis cysts and trophozoites according to standard procedures (54). Cover slips were gently put over microscopy glass slides to spread out the emulsion and then examined under a light microscope using a low-power (10×) objective for screening and a high-power (40×) objective for confirmation of presumptive and positive findings.
2.3.2 Concentration using the formalin-ethyl acetate sedimentation method
The formyl-ether concentration method was used according to recommended procedures (54). Briefly, 4 g of fresh faecal material was homogenised in 10 mL of 10% formalin and sieved through surgical gauze as a mechanical filter to remove faecal debris. The sieved suspension was transferred into a clean 15 mL centrifuge tube and 3 mL of ethyl acetate were added. After vigorous shaking, the mixture was centrifuged for 10 min at 500 × g. The supernatant was carefully discarded, and faecal smears made from the sediment and examined as described in sub-section 2.3.1.
2.3.3 Concentration using the zinc sulphate flotation method
The zinc sulphate flotation method was used according to recommended procedures (54). Briefly, 4 g of fresh faecal material was homogenised in 10 mL of 10% formalin and sieved through surgical gauze as a mechanical filter to remove faecal debris. The sieved suspension was transferred into a clean 15 mL centrifuge tube and another 10 mL of 10% formalin were added. The mix was centrifuged for 10 min at 500 × g. After discarding the supernatant, a 33% zinc sulphate solution (specific gravity: 1.18) was added to the sediment, followed by homogenisation and centrifugation for 2 min at 500 × g. Faecal smears were made with 1–2 drops of the surface film and examined as described in sub-section 2.3.1.
Aliquots of all Giardia-positive stool samples at microscopy examination were stored in 70% ethanol and shipped to the National Centre for Microbiology, Health Institute Carlos III (Majadahonda, Spain) for downstream molecular testing.
2.4 DNA extraction and purification
Genomic DNA was isolated from about 200 mg of faecal samples yielding positive results for G. duodenalis at microscopy examination by using the QIAamp DNA Stool Mini Kit (Qiagen, Hilden, Germany) according to the manufacturer’s instructions. Extracted and purified DNA samples were eluted in 200 μL of PCR-grade water and kept at 4°C until subsequent molecular testing.
2.5 Molecular identification and characterization of Giardia duodenalis
Detection of G. duodenalis DNA was achieved using a real-time PCR (qPCR) method targeting a 62-bp region of the gene codifying the small subunit ribosomal RNA (ssu rRNA) gene of the parasite (55). For assessing its molecular diversity at the assemblage and sub-assemblage levels, we adopted a sequence-based multilocus genotyping (MLST) scheme targeting the genes encoding for the glutamate dehydrogenase (gdh), β-giardin (bg), and triose phosphate isomerase (tpi) proteins of G. duodenalis. To maximise the options of amplification success at these markers, only Giardia isolates that tested positive by qPCR and yielded cycle threshold (CT) values <32 were tested for genotyping purposes. A semi-nested PCR was used to amplify a 432-bp fragment of the gdh gene (56), and nested PCRs were used to amplify 511 and 530 bp fragments of the bg and tpi genes, respectively (57, 58).
2.6 PCR and gel electrophoresis standard procedures
Information regarding the oligonucleotides and PCR cycling conditions used for the detection and genotyping of G. duodenalis is summarised in Supplementary Tables 1, 2. The qPCR protocol described above was carried out on a Corbett Rotor Gene™ 6,000 real-time PCR system (Qiagen), with reaction mixes containing 2× TaqMan® Gene Expression Master Mix (Applied Biosystems, CA, United States). All the semi-nested and nested PCR procedures mentioned above were performed on a 2720 Thermal Cycler (Applied Biosystems). Reaction mixes consistently included 2.5 units of MyTAQ™ DNA polymerase (Bioline GmbH, Luckenwalde, Germany) and 5–10 μL of MyTAQ™ Reaction Buffer containing 5 mM dNTPs and 15 mM MgCl2. Laboratory-confirmed positive and negative DNA samples of human origin were routinely used as controls and included in each round of PCR. PCR amplicons were visualised on 1.5% D5 agarose gels (Condalab, Madrid, Spain) stained with Pronasafe (Condalab) nucleic acid staining solutions.
2.7 Sequence analyses
Amplicons of the anticipated size were directly sequenced in both directions using the corresponding internal primer sets (Supplementary Table 1) in 10 μL reactions. DNA sequencing was conducted by capillary electrophoresis using the BigDye® Terminator chemistry (Applied Biosystems) on an ABI 3730xl sequencer analyser (Applied Biosystems). Raw sequencing data was examined with Chromas Lite version 2.1 software1 to generate consensus sequences. These sequences were compared with reference sequences deposited at the National Centre for Biotechnology Information (NCBI) using the BLAST tool.2 Representative nucleotide sequences generated in this study have been deposited in GenBank under accession numbers PP035393–PP035398 (gdh locus), PP035399–PP035400 (bg locus), and PP035401 (tpi locus).
2.8 Phylogenetic analyses
To analyse the phylogenetic relationship among G. duodenalis assemblages and sub-assemblages at the gdh, bg, and tpi markers, maximum-likelihood trees were constructed using MEGA version 11 (59), based on substitution rates calculated with the general time reversible model and gamma distribution with invariant sites (G + I). Bootstrapping with 1,000 replicates was used to determine support for the clades.
2.9 Statistical analyses
Associations between G. duodenalis infections and potential risk factors (sex, age, sampling season, and stool consistency) were evaluated by bivariate analysis using the Pearson’s chi-square test or Fisher’s test, as required. The significance level was set at 0.05. All statistical analyses were performed using R free software (60).
3 Results
In the present survey, 460 individual stool samples were collected from outpatients seeking medical care. The male/female ratio was 0.84. Participating patients had a median age of 13 years (0.1–60; standard deviation: 18.8). Near half of the participating patients were children under the age of 10 (44.8%, 206/460) and presented with diarrhoea (54.1%, 249/460; Table 2).
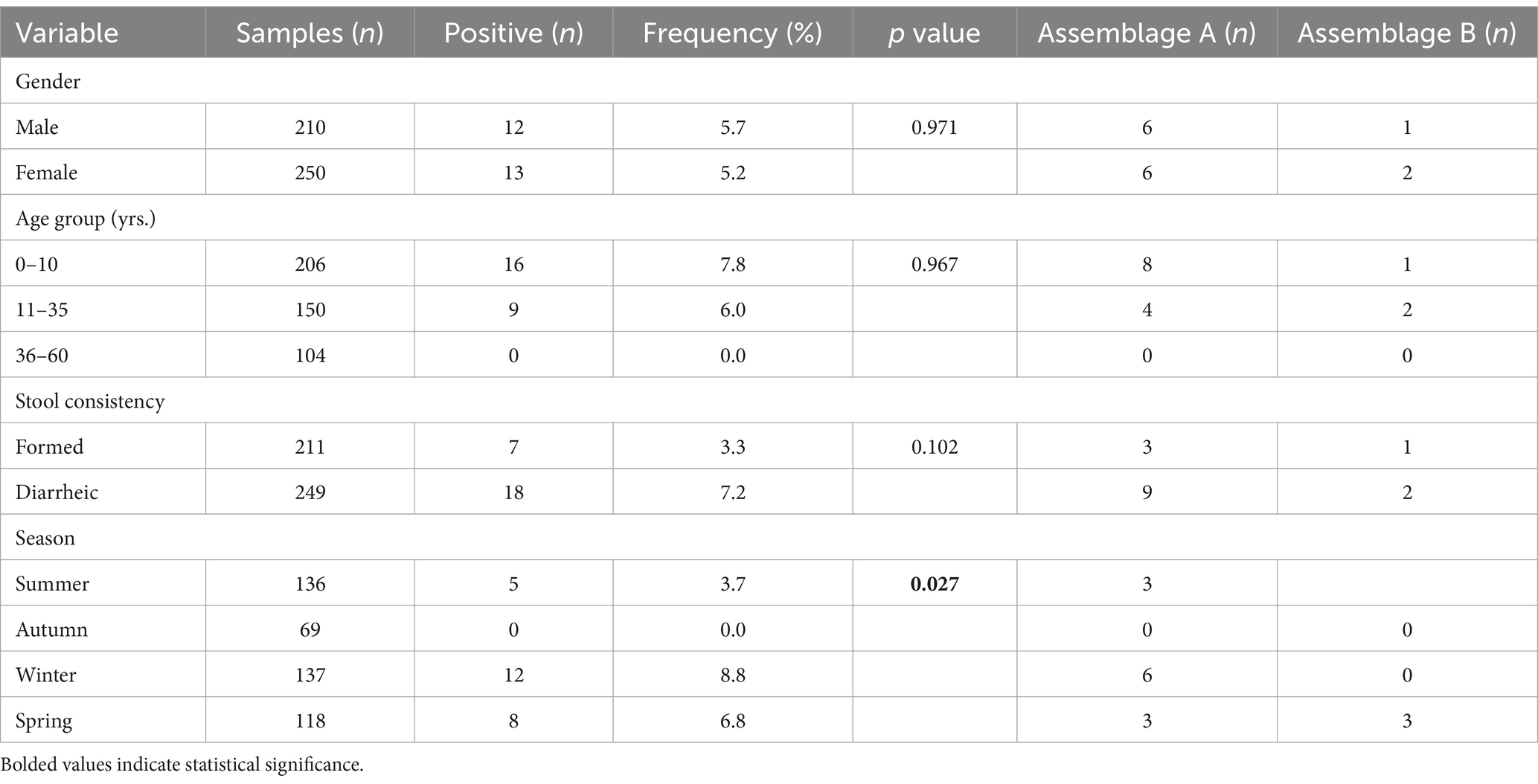
Table 2. Frequency of Giardia duodenalis infections and genotypes according to the gender, age group, stool consistency, and sampling season of the surveyed human population (n = 460).
3.1 Microscopy
Overall, 5.4% [25/460, 95% Confidence Interval (CI): 3.6–7.9] of faecal samples examined by microscopy tested positive for G. duodenalis. Neither gender, age group nor stools consistency were positively associated with a higher likelihood of having a G. duodenalis infection. Despite this lack of statistical significance, the pathogen was more frequently found in paediatric patients under 10 years of age (7.8%) and in patients presenting with diarrhoea (7.2%). Giardia infections were similarly present in males and in females (5.7% vs. 5.2%, respectively). In contrast, outpatients attended during the autumn period were less likely (p = 0.027) to harbour the pathogen (Table 2).
3.2 Confirmation of Giardia duodenalis by qPCR
All 25 faecal DNA samples with a Giardia-positive result by conventional microscopy examination tested also positive by qPCR. Yielded CT values ranged from 20.2 to 31.9 (median: 22.5; standard deviation: 4.0). Most (84.0%, 21/25) of the qPCR-positive samples yielded CT values <30 (Table 3).
3.3 Genotyping and sub-genotyping of Giardia duodenalis isolates
The molecular diversity of the 25 G. duodenalis-confirmed isolates was investigated at the assemblage and sub-assemblage levels at three (gdh, bg, and tpi) genetic markers. Successful PCR amplifications and sequencing data were generated for 48.0% (12/25, gdh), 20.0% (5/25, bg), and 12.0% (3/25, tpi) of the samples investigated at the three loci (Table 3). Overall, 56.0% (14/25) of the Giardia-positive samples were successfully genotyped at one locus at least. MLST data at the three assessed loci was available for a single sample (4.0%, 1/25). Subtyping data at a single locus and two loci were available for 36.0% (9/25) and 16.0% (4/25) of samples, respectively. No genotyping data could be obtained for 44.0% (11/25) of the Giardia-positive samples. Assemblage A (44.0%, 11/25) was more prevalent than assemblage B (12.0%, 3/25). No A + B mixed infections were detected. No host-adapted assemblages of canine (C, D), feline (F), or livestock (E) origin were identified circulating in the surveyed clinical population (Table 3).
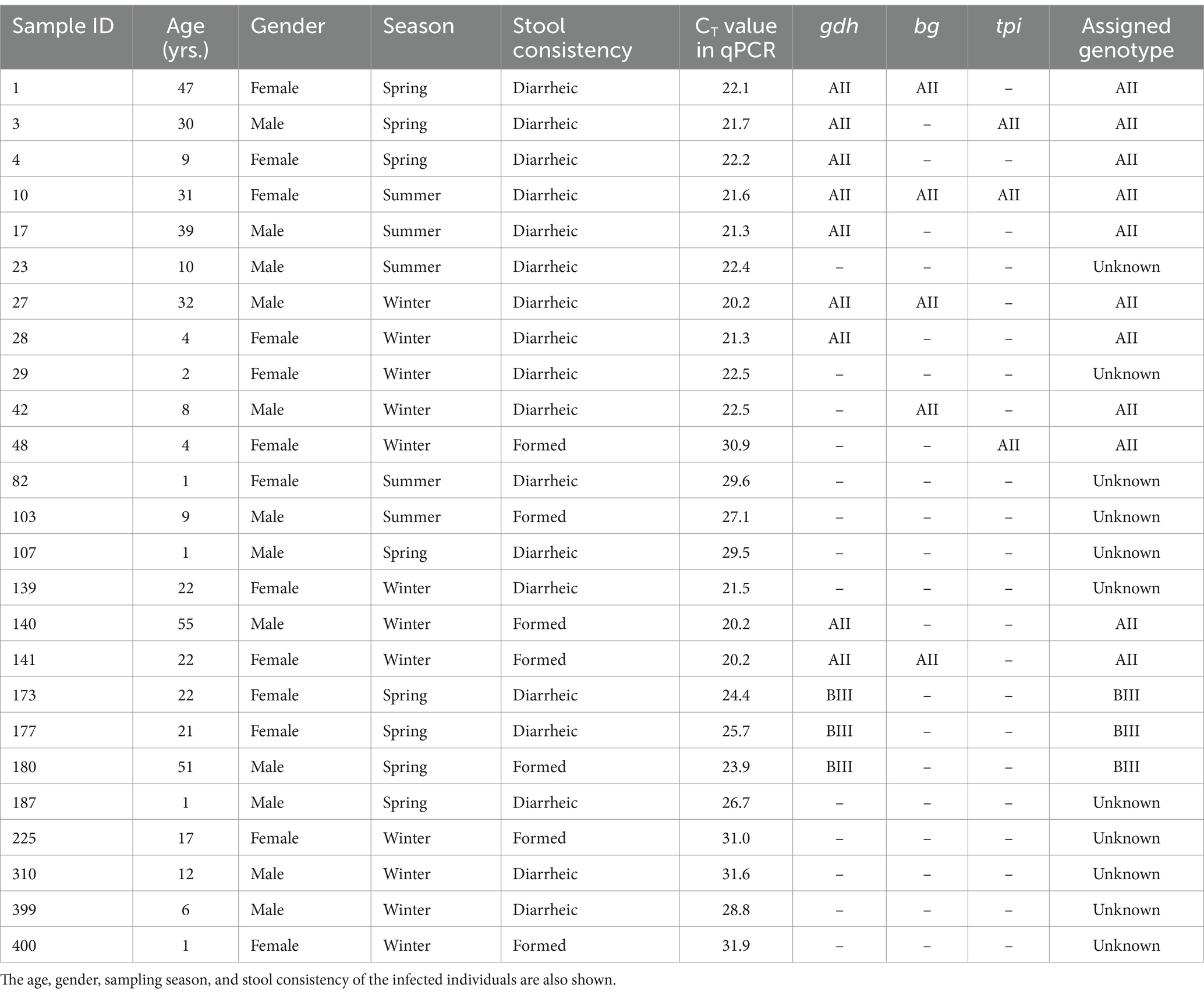
Table 3. Multilocus sequence typing results of the 25 G. duodenalis-positive human samples successfully genotyped at any of the three loci investigated in the present survey.
Table 4 shows the frequency and molecular diversity of G. duodenalis at the gdh, bg, and tpi loci. Out of the 12 gdh sequences, nine (75.0%) were assigned to the sub-assemblage AII. Of them, seven showed 100% identity with reference sequence L40510. The remaining two sequences differed from it by 1–2 single nucleotide polymorphisms (SNPs) in the form of ambiguous (double peak) positions. Sub-assemblage BIII was identified in three (25.0%) isolates, all of them differing among them and by 1–7 SNPs with reference sequence AF069059. Most of the SNPs detected correspond to clear mutations. No isolates belonging to sub-assemblage BIV were detected. All five bg sequences were identified as sub-assemblage AII. Four of them were identical to reference sequence AY072723, whereas the remaining one differed form it by a single SNP in the form of a double peak at chromatogram inspection. All three tpi sequences were identified as sub-assemblage AII and showed 100% identity with reference sequence U57897. No assemblage B isolates could be genotyped at the bg and tpi loci.
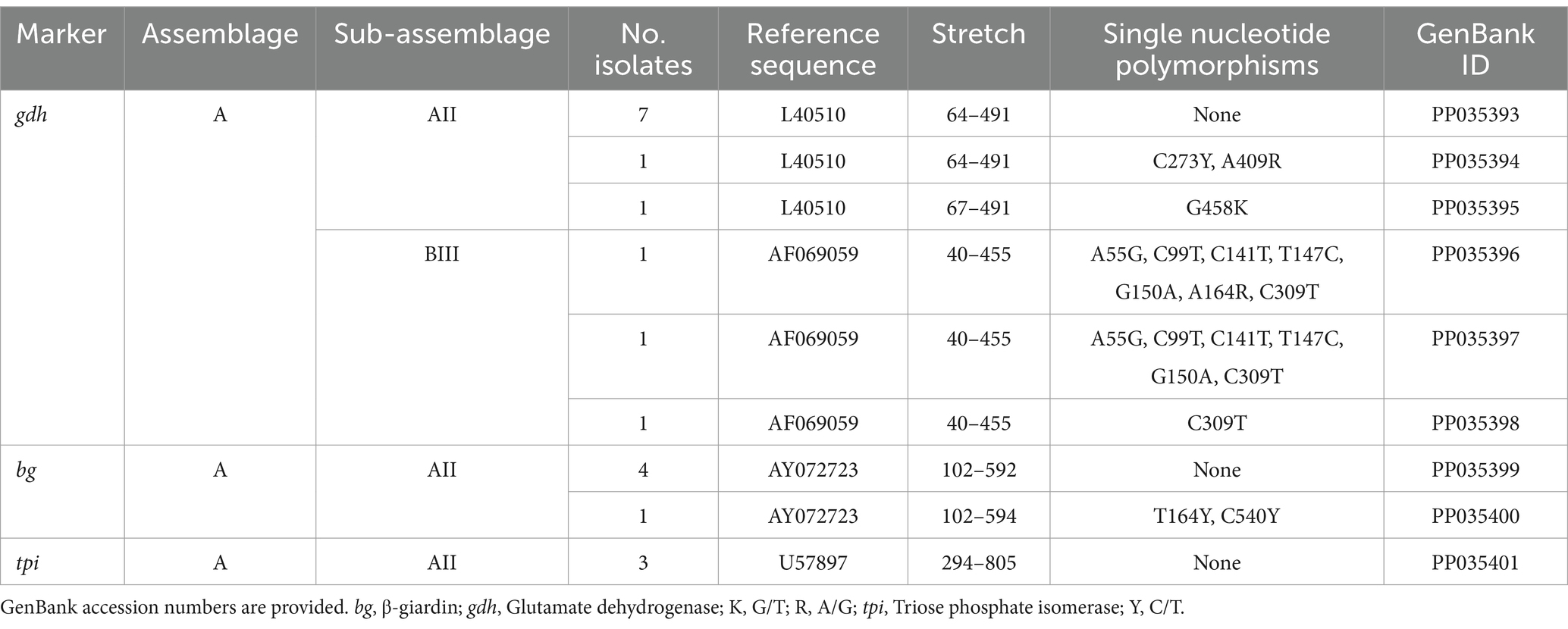
Table 4. Frequency and molecular diversity of G. duodenalis identified at the gdh, bg, and tpi loci in the human population under study.
Our statistical analyses also revealed that the distribution of assemblages A and B was independent of all the variables (gender, age group, stool consistency and sampling season) considered as potential risk factors in the present study (Table 2).
Phylogenetic analyses conducted at the gdh (Figure 1), bg (Figure 2) and tpi (Figure 3) loci clearly showed that our G. duodenalis sequences formed well-supported clades with appropriate reference sequences retrieved from GenBank.
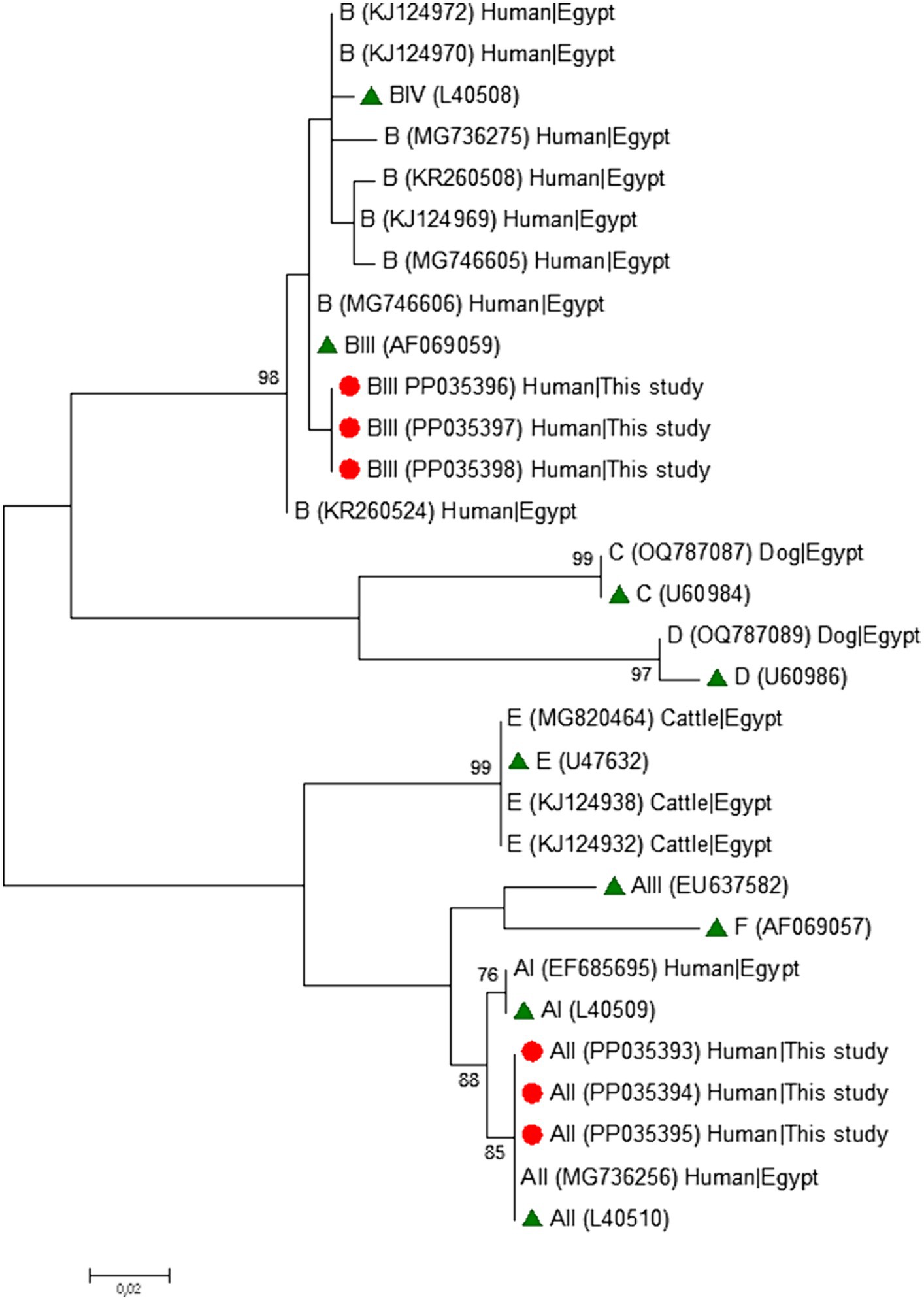
Figure 1. Phylogenetic relationship among Giardia duodenalis assemblages and sub-assemblages revealed by a maximum likelihood analysis of the partial gdh rDNA gene. Substitution rates were calculated by using the general time reversible model. Numbers on branches are percent bootstrapping values over 70% using 1,000 replicates. The filled red circle indicates the nucleotide sequence generated in the present study. The filled green triangles indicate reference sequences. Human and animal sequences from Egyptian origin retrieved from GenBank were included in the analysis for comparative purposes.
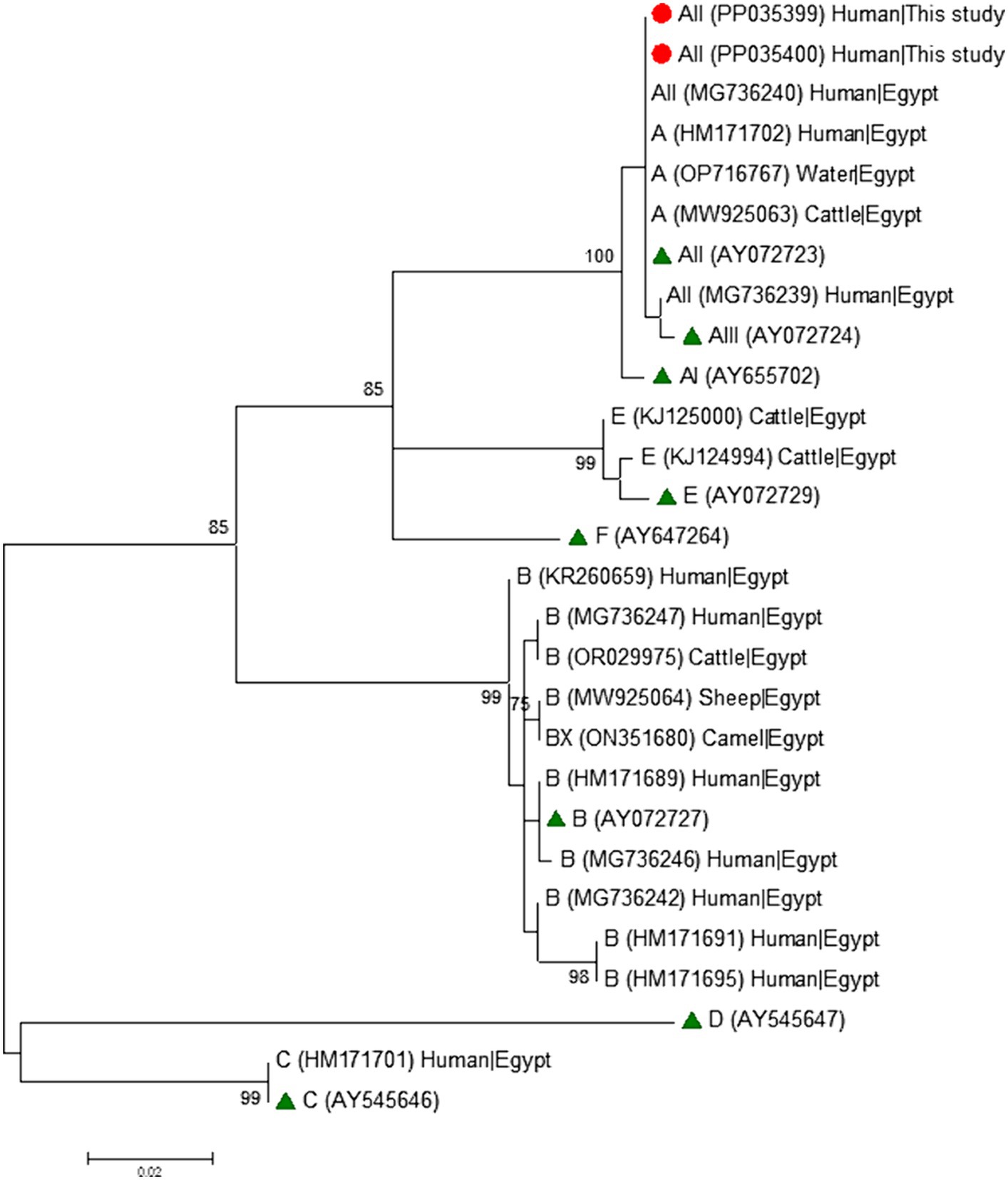
Figure 2. Phylogenetic relationship among Giardia duodenalis assemblages and sub-assemblages revealed by a maximum likelihood analysis of the partial bg rDNA gene. Substitution rates were calculated by using the general time reversible model. Numbers on branches are percent bootstrapping values over 70% using 1,000 replicates. The filled red circle indicates the nucleotide sequence generated in the present study. The filled green triangles indicate reference sequences. Human and animal sequences from Egyptian origin retrieved from GenBank were included in the analysis for comparative purposes.
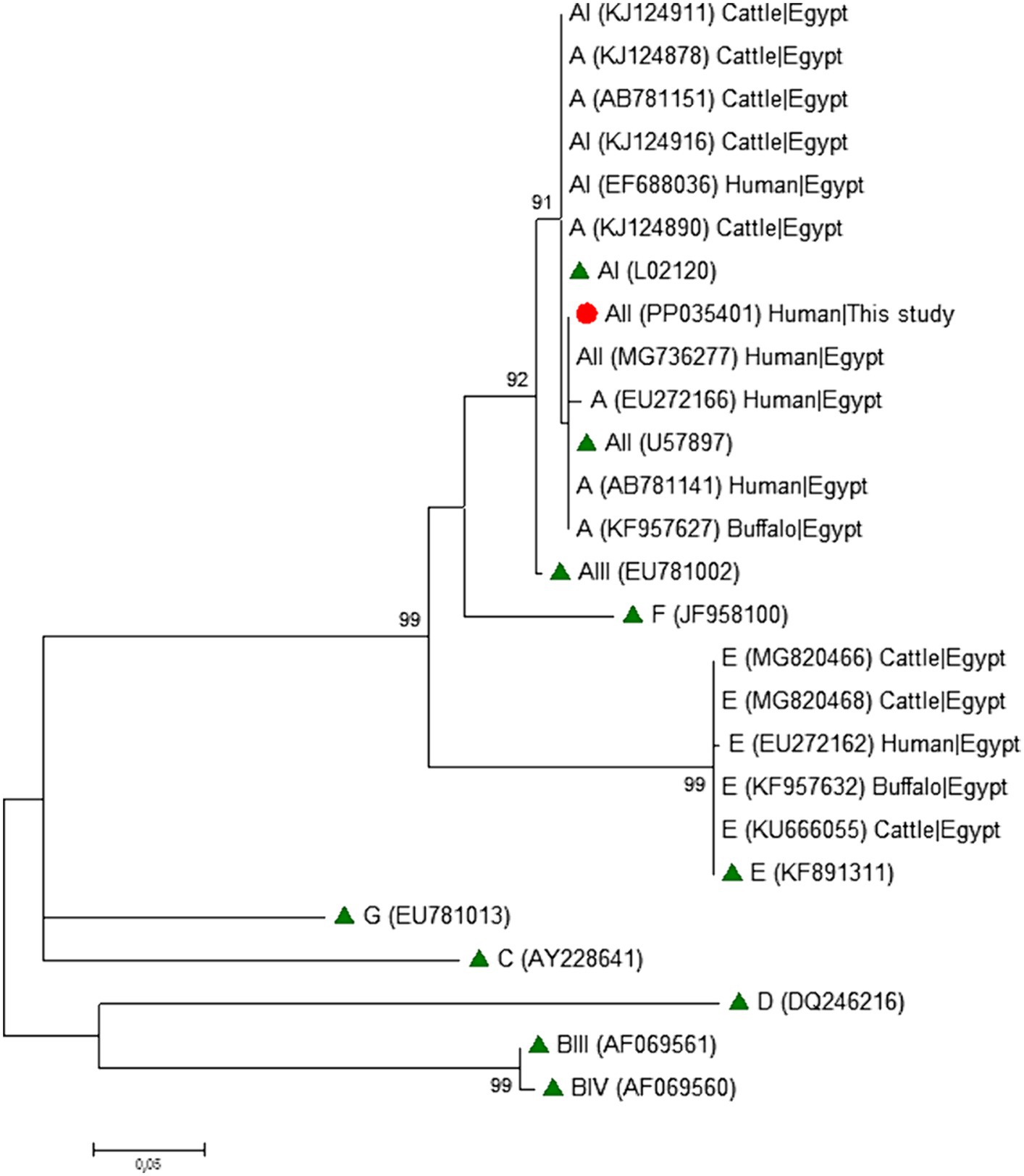
Figure 3. Phylogenetic relationship among Giardia duodenalis assemblages and sub-assemblages revealed by a maximum likelihood analysis of the partial tpi rDNA gene. Substitution rates were calculated by using the general time reversible model. Numbers on branches are percent bootstrapping values over 70% using 1,000 replicates. The filled red circle indicates the nucleotide sequence generated in the present study. The filled green triangles indicate reference sequences. Human and animal sequences from Egyptian origin retrieved from GenBank were included in the analysis for comparative purposes.
4 Discussion
Giardiosis remains a public health concern globally, with young children living in poor-resource settings carrying the bulk of the infection. Interestingly, large case/control and longitudinal studies conducted in endemic areas (most of them targeting paediatric populations) have shown that G. duodenalis infection was not associated with acute diarrhoea (61) or even has a protective effect against it (62, 63). These data highlighted to the need of clarifying the circumstances under which G. duodenalis infection may be involved in the development of diarrheal disease (either acute or persistent) and whether specific Giardia genotypes exhibit enhanced pathogenicity over other genotypes (61). This study assessed the frequency and genetic diversity of G. duodenalis infection in outpatients seeking medical care in northern Egypt. Strengths of the survey include (i) the recruiting of patients belonging to all age groups (allowing better insight of the infection through lifespan) with and without diarrhoea (allowing the study of the infection according to clinical status and the genetic variant of the parasite) and (ii) the adoption of a MLST approach to better understand the genetic diversity of the G. duodenalis isolates obtained.
In the present study, G. duodenalis infection rates were similarly found in males and females (5.7% vs. 5.2%). This finding aligns with those previously reported in studies conducted in the Egyptian provinces of El-Dakahlia, El-Gharbia, and Damietta (52) and the West Delta region (46). Conversely, males were more likely to harbour G. duodenalis infections in studies conducted in Egypt (30, 48) and other Mediterranean and Middle East countries including Algeria (64), Saudi Arabia (65) and Yemen (66). Taken together, these findings suggest that behavioural and occupational factors likely play a role in the risk of human transmission and infection. Our results also indicated that G. duodenalis infections were more frequent in children aged less than 10 years than in other age groups. This outcome is consistent with the results obtained in other microscopy-based studies conducted in Egypt (33, 46). Young children might be more vulnerable to G. duodenalis infections because of their poor personal hygiene practices, higher exposure to unsanitary environments, and immature immune system compared to adults (67, 68). However, it should be noted that other surveys have reported higher parasite frequency rates in the 10–20 years age group (30).
Despite the fact that G. duodenalis infections were more frequently identified in individuals presenting with diarrhoea than in those without diarrhoea (7.2% vs. 3.3%), the difference was not statistically significant. Remarkably, large cohort studies conducted in sub-Saharan countries including the Global Enteric Multicenter Study (GEMS) (69), the Malnutrition and Enteric Disease Study (MAL-ED) (70), and the Vaccine Impact on Diarrhea in Africa Case–Control Study (VIDA) (71) have evidenced that G. duodenalis detection was more common among non-diarrheal than diarrheal infected children. The reasons for this trend are unclear, but an indirect mechanism triggered by G. duodenalis leading to changes in colonisation/infection by other enteric pathogens associated with moderate-to-severe-diarrhoea has been suggested (71).
Regarding environmental factors, we found that G. duodenalis infections were significantly more reported during the winter (January–March) months. This is in contrast with other studies conducted in Egypt where the parasite was more frequently identified during mid-summer and late winter (33). In the absence of robust temporal series of human giardiosis in Egypt, assessing the seasonal variation of the infection is a difficult task, so available data (including those from this study) should be interpreted with caution.
In the present study, all 25 G. duodenalis-positive samples at microscopy examination were confirmed by qPCR with CT values <32, indicative of moderate-to-high parasite burdens. However, genotyping data were only available for 56.0% (14/25) of them. This moderate genotyping success rate can be explained by the fact that the gdh, bg, and tpi genotyping markers used in the present survey are all single-copy genes with limited diagnostic sensitivity. In contrast, the qPCR assay used here targeted the ssu rRNA marker, a multiple copy gene with high diagnostic sensitivity particularly suited for detection purposes (27, 28). Our genotyping analyses revealed that assemblage A was more prevalent than assemblage B (78.6% vs. 21.4%). Similar results have been reported in previous molecular surveys conducted in Cairo Governorate (30), El-Beheira Governorate (35), Ismaillia Governorate (38), and Sharkia Governorate (44) in Lower Egypt, and Assiut Governorate (48) in Upper Egypt. However, it should be noted that the opposite trend has been more frequently reported, with assemblage B accounting for 55.3% (587/1,061) of the human cases of giardiosis genotyped in the country (Table 1). No mixed A + B infections were detected in the present study, but seems a relatively common finding in other studies. Taken together, these data indicate that human G. duodenalis infections are primarily of anthropic nature in Egypt. However, and unknown fraction of these infections might be of animal origin, as both assemblages A and B are zoonotic. Although probably infrequent, zoonotic transmission events are also possible, as demonstrated by the occasional presence of canine-adapted assemblage C in a symptomatic children in Ismaillia Governorate (36) and hoofed-adapted assemblage E in human population from different Egyptian governorates (37, 40, 53). Unveiling the epidemiology of giardiosis in Egypt is a complex task hampered by our limited knowledge of which extent the human, animal, and environmental reservoirs contribute to the burden of human disease. Ideally, this task should be tackled under a One Health approach targeting all three reservoirs in the same spatiotemporal frame. Investigations should be directed towards the identification of sources of infection and transmission pathways including the identification of spillover events involving cross-species transmission in the wildlife-domestic interface. More research is also needed to investigate waterborne and foodborne transmission of G. duodenalis infections. Generating good quality molecular data is essential for these purposes.
This study unveiled variations in G. duodenalis assemblage frequencies among individuals of different ages and sexes. Assemblage A was more prevalently found in children less than 10 years of age. Of note, assemblage A was also the most common G. duodenalis genetic variant circulating in children with diarrhoea aged 2–8 years and was significantly associated with the duration and severity of the infection at Cairo Governorate (29). In contrast, children with diarrhoea below the age of 6 were more likely infected with assemblage B than with assemblage A at Beni-Suef Governorate (49). In other study conducted in Algeria, symptomatic children aged less than 7 years were predominantly infected with assemblage A, whereas assemblage B was more frequent in children older than 11 years (64). Overall, all the findings mentioned above pointed out to a mixed distribution pattern of G. duodenalis assemblages in Egypt regardless the geographical region of origin and the human population under study, very likely reflecting a complex epidemiological scenario characterised by multiple sources of infection and transmission pathways.
Currently there is no clear correlation between G. duodenalis assemblages and the outcome and severity of the infection, with limited studies on this topic (72). Large case–control studies of paediatric populations conducted in Ethiopia (73) and Mozambique (74) reported similar assemblage distributions among individuals with and without diarrhoea, suggesting that the genetic variant of the parasite is not an essential factor in the outcome of the infection. Although the present study lacks a case/control design, most children presenting with diarrhoea harboured the assemblage A of the parasite. This results is consistent with those reported in similar studies conducted in Australia (75), Bangladesh (76), Egypt (29), India (77), Iran (78), Spain (79), Syria (80), Turkey (81), and the UK (82).
This study has some design and methodological limitations that must be taken into account when evaluating the results obtained and the conclusions reached. First, the transversal nature of the study is not adequate to follow up the course of giardiosis and to capture seasonal variations of the infection. Second, initial screening of G. duodenalis infection was conducted by microscopy examination of a single stool sample per patient. Because of the limited sensitivity of this method, the infection rate reported here is likely an underestimation of the true one. Third, results obtained here might not be representative of other epidemiological scenarios or geographical areas in Egypt. And fourth, the relatively limited number of G. duodenalis isolates successfully genotyped (some of them at a single locus) might have biased the estimation of the true molecular diversity of G. duodenalis infections and the actual frequency of assemblages/sub-assemblages circulating in the investigated human population. This fact might have compromised the accuracy of some of the results obtained and the conclusions reached and warrants further investigations to corroborate the genotyping data presented here.
5 Conclusion
Endemic giardiosis continues to pose a significant public health threat in Egypt. Children under 10 years old are particularly vulnerable to the infection, so improved personal hygiene practices and promotion of healthy habits together with better access to safe drinking water and sanitary facilities are necessary to minimise the risk of transmission and infection. Our molecular findings revealed that most human cases of giardiosis were caused by assemblage A, which was the G. duodenalis genetic variant more prevalently found in individuals with diarrhoea. These results, together with the absence of animal-adapted assemblages C-F suggest that G. duodenalis infections in the surveyed population are primarily anthropic in nature. However, we cannot ruled out that an unknown proportion of the infections detected have an animal origin, as both assemblages A and B are zoonotic. Additional investigations are warranted to better understand the epidemiology of giardiosis in Egypt. Of particular interest would be conducting molecular-based epidemiological surveys in animal (domestic and free-living) and environmental (water) samples to better improve our knowledge on the sources of infection and transmission pathways of the parasite.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found at: [GenBank under accession numbers PP035393–PP035398 (gdh locus), PP035399–PP035400 (bg locus), and PP035401 (tpi locus)].
Ethics statement
The study was conducted in accordance with the Declaration of Helsinki and approved by the Kafrelsheikh University Research Ethics Committee (protocol code MKSU 50–1-10). Signed informed consents were obtained from all patients that volunteer to participate in the study. Stool samples and associated epidemiological and clinical data were anonymized to protect the privacy of the participants. The studies were conducted in accordance with the local legislation and institutional requirements. Written informed consent for participation in this study was provided by the participants’ legal guardians/next of kin.
Author contributions
EE: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Resources, Software, Validation, Visualization, Writing – original draft, Writing – review & editing. AG: Formal analysis, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. MIG: Data curation, Formal analysis, Methodology, Software, Validation, Visualization, Writing – review & editing. PK: Formal analysis, Investigation, Methodology, Software, Validation, Visualization, Writing – original draft, Writing – review & editing. AD: Data curation, Formal analysis, Methodology, Software, Visualization, Writing – review & editing. DA: Conceptualization, Data curation, Methodology, Software, Validation, Visualization, Writing – original draft. NT: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Software, Validation, Writing – original draft. HA: Data curation, Formal analysis, Software, Validation, Visualization, Writing – original draft, Writing – review & editing. MG: Data curation, Formal analysis, Software, Validation, Visualization, Writing – original draft, Writing – review & editing. AS: Conceptualization, Data curation, Formal analysis, Resources, Software, Validation, Writing – original draft, Writing – review & editing. AA: Data curation, Formal analysis, Resources, Software, Validation, Visualization, Writing – original draft. AE: Data curation, Formal analysis, Software, Validation, Writing – original draft. CH-C: Formal analysis, Methodology, Writing – review & editing. BB: Formal analysis, Methodology, Software, Validation, Writing – review & editing. DG-B: Data curation, Formal analysis, Software, Validation, Writing – review & editing. DC: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Software, Supervision, Validation, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This study was partially funded by the Health Institute Carlos III (ISCIII), Spanish Ministry of Economy and Competitiveness under project PI19CIII/00029. EE is the recipient of a postdoctoral fellowship funded by the Ministry of the Higher Education of the Arab Republic of Egypt. MG was supported by a postdoctoral contract Margarita Salas (University of Murcia) from the Programme of Requalification of the Spanish University System (Spanish Ministry of Universities) financed by the European Union-NextGenerationEU. DG-B is the recipient of a Sara Borrell Research Contract (CD19CIII/00011) funded by the Spanish Ministry of Science, Innovation, and Universities. AD is the recipient of a PFIS contract (FI20CIII/00002) funded by the Spanish Ministry of Science and Innovation and Universities. CH-C is the recipient of a fellowship funded by the Fundación Carolina (Spain) and the University of Antioquia, Medellín (Colombia).
Acknowledgments
We would like to acknowledge the Princess Nourah bint Abdulrahman University Researchers Supporting Project no. PNURSP2024R401, Princess Nourah Bint Abdulrahman University, Riyadh, Saudi Arabia.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpubh.2024.1377123/full#supplementary-material
Footnotes
References
1. WHO. The world health report. Fighting disease fostering development. Geneva: World Health Organization (1996).
2. Cai, W, Ryan, U, Xiao, L, and Feng, Y. Zoonotic giardiasis: an update. Parasitol Res. (2021) 120:4199–218. doi: 10.1007/s00436-021-07325-2
3. Sarria-Guzmán, Y, Chávez-Romero, Y, Bernal, JE, González-Jiménez, FE, Serrano-Silva, N, and Fusaro, C. Molecular identification of Giardia spp. in Latin America: an updated systematic review on reports from 2017 to 2021. J Infect Dev Ctries. (2022) 16:392–401. doi: 10.3855/jidc.15806
4. Tawana, M, Onyiche, TE, Ramatla, T, and Thekisoe, O. A 'One Health' perspective of Africa-wide distribution and prevalence of Giardia species in humans, animals and waterbodies: a systematic review and meta-analysis. Parasitology. (2023) 150:769–80. doi: 10.1017/S0031182023000513
5. Adam, RD. Giardia duodenalis: biology and pathogenesis. Clin Microbiol Rev. (2021) 34:e0002419. doi: 10.1128/CMR.00024-19
6. Betancourt, WQ, and Rose, JB. Drinking water treatment processes for removal of Cryptosporidium and Giardia. Vet Parasitol. (2004) 126:219–34. doi: 10.1016/j.vetpar.2004.09.002
7. Morrison, CM, Hogard, S, Pearce, R, Gerrity, D, von Gunten, U, and Wert, EC. Ozone disinfection of waterborne pathogens and their surrogates: a critical review. Water Res. (2022) 214:118206. doi: 10.1016/j.watres.2022.118206
8. Al-Mekhlafi, MS, Azlin, M, Nor Aini, U, Shaik, A, Sa'iah, A, Fatmah, MS, et al. Giardiasis as a predictor of childhood malnutrition in orang Asli children in Malaysia. Trans R Soc Trop Med Hyg. (2005) 99:686–91. doi: 10.1016/j.trstmh.2005.02.006
9. Bartelt, LA, and Sartor, RB. Advances in understanding Giardia: determinants and mechanisms of chronic sequelae. F1000Prime Rep. (2015) 7:62. doi: 10.12703/P7-62
10. Einarsson, E, Ma'ayeh, S, and Svärd, SG. An up-date on Giardia and giardiasis. Curr Opin Microbiol. (2016) 34:47–52. doi: 10.1016/j.mib.2016.07.019
11. Dougherty, M, and Bartelt, LA. Giardia and growth impairment in children in high-prevalence settings: consequence or co-incidence? Curr Opin Infect Dis. (2022) 35:417–23. doi: 10.1097/QCO.0000000000000877
12. Halliez, MC, and Buret, AG. Extra-intestinal and long term consequences of Giardia duodenalis infections. World J Gastroenterol. (2013) 19:8974–85. doi: 10.3748/wjg.v19.i47.8974
13. Allain, T, Fekete, E, Sosnowski, O, Desmonts de Lamache, D, Motta, JP, Leger, D, et al. High-fat diet increases the severity of Giardia infection in association with low-grade inflammation and gut microbiota dysbiosis. Sci Rep. (2021) 11:18842. doi: 10.1038/s41598-021-98262-8
14. Argüello-García, R, and Ortega-Pierres, MG. Giardia duodenalis virulence - "to be, or not to be". Curr Trop Med Rep. (2021) 8:246–56. doi: 10.1007/s40475-021-00248-z
15. Fink, MY, and Singer, SM. The intersection of immune responses, microbiota, and pathogenesis in giardiasis. Trends Parasitol. (2017) 33:901–13. doi: 10.1016/j.pt.2017.08.001
16. Wang, L, Xiao, L, Duan, L, Ye, J, Guo, Y, Guo, M, et al. Concurrent infections of Giardia duodenalis, Enterocytozoon bieneusi, and Clostridium difficile in children during a cryptosporidiosis outbreak in a pediatric hospital in China. PLoS Negl Trop Dis. (2013) 7:e2437. doi: 10.1371/journal.pntd.0002437
17. Mørch, K, and Hanevik, K. Giardiasis treatment: an update with a focus on refractory disease. Curr Opin Infect Dis. (2020) 33:355–64. doi: 10.1097/QCO.0000000000000668
18. Bartelt, LA, and Platts-Mills, JA. Giardia: a pathogen or commensal for children in high-prevalence settings? Curr Opin Infect Dis. (2016) 29:502–7. doi: 10.1097/QCO.0000000000000293
19. Krumrie, S, Capewell, P, Smith-Palmer, A, Mellor, D, Weir, W, and Alexander, CL. A scoping review of risk factors and transmission routes associated with human giardiasis outbreaks in high-income settings. Curr Res Parasitol Vector Borne Dis. (2022) 2:100084. doi: 10.1016/j.crpvbd.2022.100084
20. Younas, M, Shah, S, and Talaat, A. Frequency of Giardia lamblia infection in children with recurrent abdominal pain. J Pak Med Assoc. (2008) 58:171–4.
21. Shaima, SN, Das, SK, Ahmed, S, Jahan, Y, Khan, SH, Mamun, GMS, et al. Anthropometric indices of Giardia-infected under-five children presenting with moderate-to-severe diarrhea and their healthy community controls: data from the global enteric multicenter study. Child Aust. (2021) 8:1186. doi: 10.3390/children8121186
22. WHO. Basic laboratory methods in medical parasitology. Geneva: World Health Organization (1991).
23. de Boer, RF, Ott, A, Kesztyüs, B, and Kooistra-Smid, AM. Improved detection of five major gastrointestinal pathogens by use of a molecular screening approach. J Clin Microbiol. (2010) 48:4140–6. doi: 10.1128/JCM.01124-10
24. Verweij, JJ. Application of PCR-based methods for diagnosis of intestinal parasitic infections in the clinical laboratory. Parasitology. (2014) 141:1863–72. doi: 10.1017/S0031182014000419
25. van Lieshout, L, and Roestenberg, M. Clinical consequences of new diagnostic tools for intestinal parasites. Clin Microbiol Infect. (2015) 21:520–8. doi: 10.1016/j.cmi.2015.03.015
26. Formenti, F, Valerio, M, Guerriero, M, Perandin, F, Pajola, B, Mistretta, M, et al. Molecular biology can change the classic laboratory approach for intestinal protozoan infections. Front Microbiol. (2017) 8:2191. doi: 10.3389/fmicb.2017.02191
27. Ryan, U, Paparini, A, and Oskam, C. New technologies for detection of enteric parasites. Trends Parasitol. (2017) 33:532–46. doi: 10.1016/j.pt.2017.03.005
28. Ryan, UM, Feng, Y, Fayer, R, and Xiao, L. Taxonomy and molecular epidemiology of Cryptosporidium and Giardia - a 50 year perspective (1971-2021). Int J Parasitol. (2021) 51:1099–119. doi: 10.1016/j.ijpara.2021.08.007
29. El Basha, NR, Zaki, MM, Hassanin, OM, Rehan, MK, and Omran, D. Giardia assemblages a and B in diarrheic patients: a comparative study in Egyptian children and adults. J Parasitol. (2016) 102:69–74. doi: 10.1645/14-676
30. Helmy, MM, Abdel-Fattah, HS, and Rashed, L. Real-time PCR/RFLP assay to detect Giardia intestinalis genotypes in human isolates with diarrhea in Egypt. J Parasitol. (2009) 95:1000–4. doi: 10.1645/GE-1670.1
31. Madbouly, NA, Farid, A, El-Badry, AA, and El-Amir, AM. Immunomolecular identification of Giardia intestinalis in diarrhoeal children; comparison of three diagnostic methods. J Egypt Soc Parasitol. (2016) 46:253–60. doi: 10.21608/jesp.2016.88648
32. Taha, SA, Abd Al Aal, Z, Saleh, NS, and El-Badry, AA. Giardia intestinalis assemblages among Egyptian symptomatic children: prevalence and seasonal distribution in Cairo. Egypt J Egypt Soc Parasitol. (2018) 48:661–8. doi: 10.21608/jesp.2018.76585
33. Ismail, MA, El-Akkad, DM, Rizk, EM, El-Askary, HM, and El-Badry, AA. Molecular seasonality of Giardia lamblia in a cohort of Egyptian children: a circannual pattern. Parasitol Res. (2016) 115:4221–7. doi: 10.1007/s00436-016-5199-7
34. Elswaifi, SF, Palmieri, JR, El-Tantawy, N, El-Hussiny, M, Besheer, T, and Abohashem, E. Comparison of microscopic and immunoassay examination in the diagnosis of intestinal protozoa of humans in Mansoura. Egypt J Parasit Dis. (2016) 40:580–5. doi: 10.1007/s12639-014-0542-3
35. Abd El-Latif, NF, El-Taweel, HA, Gaballah, A, Salem, AI, and Abd El-Malek, AHM. Molecular characterization of Giardia intestinalis detected in humans and water samples in Egypt. Acta Parasitol. (2020) 65:482–9. doi: 10.2478/s11686-020-00176-4
36. Soliman, RH, Fuentes, I, and Rubio, JM. Identification of a novel assemblage B subgenotype and a zoonotic assemblage C in human isolates of Giardia intestinalis in Egypt. Parasitol Int. (2011) 60:507–11. doi: 10.1016/j.parint.2011.09.006
37. Helmy, YA, Klotz, C, Wilking, H, Krücken, J, Nöckler, K, Von Samson-Himmelstjerna, G, et al. Epidemiology of Giardia duodenalis infection in ruminant livestock and children in the Ismailia province of Egypt: insights by genetic characterization. Parasit Vectors. (2014) 7:321. doi: 10.1186/1756-3305-7-321
38. Hussein, EM, Ismail, OA, Mokhtar, AB, Mohamed, SE, and Saad, RM. Nested PCR targeting intergenic spacer (IGS) in genotyping of Giardia duodenalis isolated from symptomatic and asymptomatic infected Egyptian school children. Parasitol Res. (2017) 116:763–71. doi: 10.1007/s00436-016-5347-0
39. Yu, F, Amer, S, Qi, M, Wang, R, Wang, Y, Zhang, S, et al. Multilocus genotyping of Giardia duodenalis isolated from patients in Egypt. Acta Trop. (2019) 196:66–71. doi: 10.1016/j.actatropica.2019.05.012
40. Foronda, P, Bargues, MD, Abreu-Acosta, N, Periago, MV, Valero, MA, Valladares, B, et al. Identification of genotypes of Giardia intestinalis of human isolates in Egypt. Parasitol Res. (2008) 103:1177–81. doi: 10.1007/s00436-008-1113-2
41. Sadek, GS, El-Settawy, MA, and Nasr, SA. Genotypic characterization of Giardia duodenalis in children in Menoufiya and Sharkiya governorates. Egypt Life Sci J. (2013) 10:3006–15.
42. Mahmud, MA, Chappell, C, Hossain, MM, Habib, M, and Dupont, HL. Risk factors for development of first symptomatic Giardia infection among infants of a birth cohort in rural Egypt. Am J Trop Med Hyg. (1995) 53:84–8. doi: 10.4269/ajtmh.1995.53.84
43. Mohamed, AMA, Bayoumy, AM, Abo-Hashim, AH, Ibrahim, AA, and El-Badry, AA. Giardiasis in symptomatic children from Sharkia, Egypt: genetic assemblages and associated risk factors. J Parasit Dis. (2020) 44:719–24. doi: 10.1007/s12639-020-01254-0
44. Gharieb, RMA, and Abd El-Ghany, AM. Giardia lamblia in household persons and buffalo calves; prevalence, molecular identification and associated risk factors. Jpn J Vet Res. (2016) 64:S15–22.
45. Hussein, EM, Zaki, WM, Ahmed, SA, Almatary, AM, Nemr, NI, and Hussein, AM. Predominance of Giardia lamblia assemblage a among iron deficiency anaemic pre-school Egyptian children. Parasitol Res. (2016) 115:1537–45. doi: 10.1007/s00436-015-4888-y
46. Elhadad, H, Abdo, S, Tolba, M, Salem, AI, Mohamed, MA, El-Abd, EA, et al. Detection of Giardia intestinalis assemblages a and B among children from three villages in the West Delta region, Egypt using assemblage specific primers. J Parasit Dis. (2021) 45:655–63. doi: 10.1007/s12639-020-01338-x
47. Ahmed, SO, Hamed, MI, and Yones, DA. Molecular and conventional detection of zoonotic Giardia and Cryptosporidium in children and calves in upper Egypt. Am J Infect Dis. (2016) 4:91–4. doi: 10.12691/ajidm-4-4-4
48. Ahmad, AA, El-Kady, AM, and Hassan, TM. Genotyping of Giardia duodenalis in children in upper Egypt using assemblage-specific PCR technique. PLoS One. (2020) 15:e0240119. doi: 10.1371/journal.pone.0240119
49. Ghieth, MA, El-Badry, AA, Abu-Sarea, EY, Abdel Gawad, SS, and Elsharkawy, MM. Genotypic analysis of Giardia duodenalis in children at Egypt. Comp Clin Pathol. (2016) 25:1241–6. doi: 10.1007/s00580-016-2337-7
50. Fahmy, HM, El-Serougi, AO, El Deeb, HK, Hussein, HM, Abou-Seri, HM, Klotz, C, et al. Giardia duodenalis assemblages in Egyptian children with diarrhea. Eur J Clin Microbiol Infect Dis. (2015) 34:1573–81. doi: 10.1007/s10096-015-2389-7
51. Amer, SE. Genotypic and phylogenetic characterization of Giardia intestinalis from human and dairy cattle in Kafr El sheikh governorate, Egypt. J Egypt Soc Parasitol. (2013) 43:133–46. doi: 10.12816/0006373
52. Naguib, D, El-Gohary, AH, Roellig, D, Mohamed, AA, Arafat, N, Wang, Y, et al. Molecular characterization of Cryptosporidium spp. and Giardia duodenalis in children in Egypt. Parasit Vectors. (2018) 11:403. doi: 10.1186/s13071-018-2981-7
53. Abdel-Moein, KA, and Saeed, H. The zoonotic potential of Giardia intestinalis assemblage E in rural settings. Parasitol Res. (2016) 115:3197–202. doi: 10.1007/s00436-016-5081-7
54. Garcia, LS. Diagnostic Medical Parasitology. Manual of commercial methods in clinical microbiology. Washington, DC USA: ASM Press (2005).
55. Verweij, JJ, Schinkel, J, Laeijendecker, D, van Rooyen, MA, van Lieshout, L, and Polderman, AM. Real-time PCR for the detection of Giardia lamblia. Mol Cell Probes. (2003) 17:223–5. doi: 10.1016/s0890-8508(03)00057-4
56. Read, CM, Monis, PT, and Thompson, RC. Discrimination of all genotypes of Giardia duodenalis at the glutamate dehydrogenase locus using PCR-RFLP. Infect Genet Evol. (2004) 4:125–30. doi: 10.1016/j.meegid.2004.02.001
57. Lalle, M, Pozio, E, Capelli, G, Bruschi, F, Crotti, D, and Cacciò, SM. Genetic heterogeneity at the beta-giardin locus among human and animal isolates of Giardia duodenalis and identification of potentially zoonotic subgenotypes. Int J Parasitol. (2005) 35:207–13. doi: 10.1016/j.ijpara.2004.10.022
58. Sulaiman, IM, Fayer, R, Bern, C, Gilman, RH, Trout, JM, Schantz, PM, et al. Triose phosphate isomerase gene characterization and potential zoonotic transmission of Giardia duodenalis. Emerg Infect Dis. (2003) 9:1444–52. doi: 10.3201/eid0911.030084
59. Tamura, K, Stecher, G, and Kumar, S. MEGA11: molecular evolutionary genetics analysis version 11. Mol Biol Evol. (2021) 38:3022–7. doi: 10.1093/molbev/msab120
60. R Core Team. R: A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing (2023).
61. Muhsen, K, and Levine, MM. A systematic review and meta-analysis of the association between Giardia lamblia and endemic pediatric diarrhea in developing countries. Clin Infect Dis. (2012) 55:S271–93. doi: 10.1093/cid/cis762
62. Veenemans, J, Mank, T, Ottenhof, M, Baidjoe, A, Mbugi, EV, Demir, AY, et al. Protection against diarrhea associated with Giardia intestinalis is lost with multi-nutrient supplementation: a study in Tanzanian children. PLoS Negl Trop Dis. (2011) 5:e1158. doi: 10.1371/journal.pntd.0001158
63. Haque, R, Mondal, D, Karim, A, Molla, IH, Rahim, A, Faruque, AS, et al. Prospective case-control study of the association between common enteric protozoal parasites and diarrhea in Bangladesh. Clin Infect Dis. (2009) 48:1191–7. doi: 10.1086/597580
64. Rebih, N, Boutaiba, S, Aboualchamat, G, Souttou, K, Hakem, A, and Al, NS. Molecular and epidemiological characterization of Giardia intestinalis assemblages detected in Djelfa. Algeria J Parasit Dis. (2020) 44:281–8. doi: 10.1007/s12639-020-01206-8
65. Shalaby, I, Gherbawy, Y, and Banaja, A. Molecular characterization of Giardia parasite isolated from stool samples collected from different hospitals in Taif City (Saudi Arabia). Trop Biomed. (2011) 28:487–96.
66. Al-Mekhlafi, HM. Giardia duodenalis infection among rural communities in Yemen: a community-based assessment of the prevalence and associated risk factors. Asian Pac J Trop Med. (2017) 10:987–95. doi: 10.1016/j.apjtm.2017.09.011
67. Bauhofer, AFL, Cossa-Moiane, ILC, Marques, SDA, Guimarães, ELAM, Munlela, BA, Anapakala, EM, et al. Intestinal protozoa in hospitalized under-five children with diarrhoea in Nampula - a cross-sectional analysis in a low-income setting in northern Mozambique. BMC Infect Dis. (2021) 21:201. doi: 10.1186/s12879-021-05881-7
68. Gebru, H, Deyissia, N, Medhin, G, and Kloos, H. The association of sanitation and hygiene practices with intestinal parasitic infections among under-14 children in rural Dire Dawa, eastern Ethiopia: a community based cross-sectional study. Environ Health Insights. (2023) 17:11786302231180801. doi: 10.1177/11786302231180801
69. Kotloff, KL, Nataro, JP, Blackwelder, WC, Nasrin, D, Farag, TH, Panchalingam, S, et al. Burden and aetiology of diarrhoeal disease in infants and young children in developing countries (the global enteric multicenter study, GEMS): a prospective, case-control study. Lancet. (2013) 382:209–22. doi: 10.1016/S0140-6736(13)60844-2
70. Platts-Mills, JA, Babji, S, Bodhidatta, L, Gratz, J, Haque, R, Havt, A, et al. Pathogen-specific burdens of community diarrhoea in developing countries: a multisite birth cohort study (MAL-ED). Lancet Glob Health. (2015) 3:e564–75. doi: 10.1016/S2214-109X(15)00151-5
71. Marcenac, P, Traoré, A, Kim, S, Prentice-Mott, G, Berendes, DM, Powell, H, et al. Giardia detection and codetection with other enteric pathogens in young children in the vaccine impact on diarrhea in Africa (VIDA) case-control study: 2015-2018. Clin Infect Dis. (2023) 76:S106–13. doi: 10.1093/cid/ciac940
72. Zajaczkowski, P, Lee, R, Fletcher-Lartey, SM, Alexander, K, Mahimbo, A, Stark, D, et al. The controversies surrounding Giardia intestinalis assemblages a and B. Curr Res Parasitol Vector Borne Dis. (2021) 1:100055. doi: 10.1016/j.crpvbd.2021.100055
73. Alemu, Y, Abdissa, A, Mekonnen, Z, Sharew, B, Johansen, ØH, Bjørang, O, et al. Prevalence and assemblage of Giardia duodenalis in a case-control study of children under 5 years from Jimma, Southwest Ethiopia. Parasitol Res. (2023) 123:38. doi: 10.1007/s00436-023-08029-5
74. Messa, A Jr, Köster, PC, Garrine, M, Gilchrist, C, Bartelt, LA, Nhampossa, T, et al. Molecular diversity of Giardia duodenalis in children under 5 years from the Manhiça district, southern Mozambique enrolled in a matched case-control study on the aetiology of diarrhoea. PLoS Negl Trop Dis. (2021) 15:e0008987. doi: 10.1371/journal.pntd.0008987
75. Read, C, Walters, J, Robertson, ID, and Thompson, RC. Correlation between genotype of Giardia duodenalis and diarrhoea. Int J Parasitol. (2002) 32:229–31. doi: 10.1016/s0020-7519(01)00340-x
76. Haque, R, Roy, S, Kabir, M, Stroup, SE, Mondal, D, and Houpt, ER. Giardia assemblage a infection and diarrhea in Bangladesh. J Infect Dis. (2005) 192:2171–3. doi: 10.1086/498169
77. Ajjampur, SS, Sankaran, P, Kannan, A, Sathyakumar, K, Sarkar, R, Gladstone, BP, et al. Giardia duodenalis assemblages associated with diarrhea in children in South India identified by PCR-RFLP. Am J Trop Med Hyg. (2009) 80:16–9. doi: 10.4269/ajtmh.2009.80.16
78. Sarkari, B, Ashrafmansori, A, Hatam, GR, Motazedian, MH, Asgari, Q, and Mohammadpour, I. Genotyping of Giardia lamblia isolates from human in southern Iran. Trop Biomed. (2012) 29:366–71.
79. Sahagún, J, Clavel, A, Goñi, P, Seral, C, Llorente, MT, Castillo, FJ, et al. Correlation between the presence of symptoms and the Giardia duodenalis genotype. Eur J Clin Microbiol Infect Dis. (2008) 27:81–3. doi: 10.1007/s10096-007-0404-3
80. Aydin, AF, Besirbellioglu, BA, Avci, IY, Tanyuksel, M, Araz, E, and Pahsa, A. Classification of Giardia duodenalis parasites in Turkey into groups a and B using restriction fragment length polymorphism. Diagn Microbiol Infect Dis. (2004) 50:147–51. doi: 10.1016/j.diagmicrobio.2004.06.001
81. Skhal, D, Aboualchamat, G, Al Mariri, A, and Al, NS. Prevalence of Giardia duodenalis assemblages and sub-assemblages in symptomatic patients from Damascus city and its suburbs. Infect Genet Evol. (2017) 47:155–60. doi: 10.1016/j.meegid.2016.11.030
Keywords: Giardia , conventional microscopy, epidemiology, diarrhoea, human, assemblage, PCR
Citation: Elmahallawy EK, Gareh A, Ghallab MMI, Köster PC, Dashti A, Aboelsoued D, Toaleb NI, Alzaylaee H, Gonzálvez M, Saleh AA, Alhegaili AS, Eldehn AF, Hernández-Castro C, Bailo B, González-Barrio D and Carmena D (2024) Microscopy detection and molecular characterisation of Giardia duodenalis infection in outpatients seeking medical care in Egypt. Front. Public Health. 12:1377123. doi: 10.3389/fpubh.2024.1377123
Edited by:
Ponsiano Ocama, Makerere University, UgandaReviewed by:
Marius Stelian Ilie, Banat University of Agricultural Sciences and Veterinary Medicine, RomaniaAleksandra Uzelac, University of Belgrade, Serbia
Copyright © 2024 Elmahallawy, Gareh, Ghallab, Köster, Dashti, Aboelsoued, Toaleb, Alzaylaee, Gonzálvez, Saleh, Alhegaili, Eldehn, Hernández-Castro, Bailo, González-Barrio and Carmena. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ehab Kotb Elmahallawy, ZWVoYWFAdW5pbGVvbi5lcw==; David Carmena, ZGFjYXJtZW5hQGlzY2lpaS5lcw==