
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
CASE REPORT article
Front. Public Health , 21 August 2023
Sec. Infectious Diseases: Epidemiology and Prevention
Volume 11 - 2023 | https://doi.org/10.3389/fpubh.2023.1236384
 Davi E. R. Sousa1,2
Davi E. R. Sousa1,2 Tais M. Wilson1,2
Tais M. Wilson1,2 Isabel L. Macêdo1,2Alessandro P. M. Romano3Daniel G. Ramos3Pedro H. O. Passos3Gabriela R. T. Costa1,4
Isabel L. Macêdo1,2Alessandro P. M. Romano3Daniel G. Ramos3Pedro H. O. Passos3Gabriela R. T. Costa1,4 Vagner S. Fonseca5,6Maria Angélica M. M. Mares-Guia7
Vagner S. Fonseca5,6Maria Angélica M. M. Mares-Guia7 Marta Giovanetti6,8
Marta Giovanetti6,8 Luiz Carlos Junior Alcantara6,7
Luiz Carlos Junior Alcantara6,7 Ana Maria B. de Filippis7Giane R. Paludo1Cristiano B. Melo1
Ana Maria B. de Filippis7Giane R. Paludo1Cristiano B. Melo1 Márcio B. Castro1,2*
Márcio B. Castro1,2*Free-ranging non-human primates (NHP) can live in anthropized areas or urban environments in close contact with human populations. This condition can enable the emergence and transmission of high-impact zoonotic pathogens. For the first time, we detected a coinfection of the yellow fever (YF) virus with Toxoplasma gondii in a free-ranging NHP in a highly urbanized area of a metropolis in Brazil. Specifically, we observed this coinfection in a black-tufted marmoset found dead and taken for a necropsy by the local health surveillance service. After conducting an epidemiological investigation, characterizing the pathological features, and performing molecular assays, we confirmed that the marmoset developed an acute fatal infection caused by T. gondii in coinfection with a new YF virus South American-1 sub-lineage. As a result, we have raised concerns about the public health implications of these findings and discussed the importance of diagnosis and surveillance of zoonotic agents in urbanized NHPs. As competent hosts of zoonotic diseases such as YF and environmental sentinels for toxoplasmosis, NHPs play a crucial role in the One Health framework to predict and prevent the emergence of dangerous human pathogens.
Zoonotic infections have plagued populations since ancient civilizations and have again shown their drastic effects on public health recently in the COVID-19 pandemic. The surveillance of zoonotic diseases has become essential in a globalized world, especially considering wild animals that live or are close to people in anthropic environments. The early detection of epizootic yellow fever (YF) in non-human primates (NHPs) is a successful example of surveillance for triggering health actions to prevent YF cases in humans (1).
NHPs are important sentinels for YF that acutely die from the disease and play an essential role in disease transmission. In the sylvatic cycle of YF, New World NHPs are infected by mosquitoes in forests and may die. These animals and mosquitoes may also disseminate the YF virus (YFV) to anthropogenic areas such as cities through waves of epizootic outbreaks reaching urban NHP populations, where they can live as our daily neighbors and be potential hosts for the YFV and other zoonotic agents (1–6). In some countries, well-adapted species of NHPs are commonly found in cities and often have close contact with humans (1, 7, 8). Additionally, the high susceptibility of some free-ranging NHPs species to develop fatal Toxoplasma gondii infections can be a concern for public health services due to people’s outcry caused by the unexplained death of many animals during outbreaks (7, 9, 10).
The surveillance of deaths in free-ranging NHPs is an essential part of Brazil’s National Program to Control Yellow Fever (NPCYF) and also provides opportunities for the surveillance of other zoonotic agents in NHPs that live near human populations. In this study, for the first time, we detected an acute fatal case of toxoplasmosis in coinfection with the YFV in a free-ranging marmoset in an urbanized area of a metropolis in Brazil.
In December 2020, during a new re-emergence of YF in Midwestern Brazil, we investigated a black-tufted marmoset (Callithrix penicillata) found dead in an urbanized area of Brasilia (15°49′23.6”S 47°53′02.8”W), as part of the NPCYF at the Regional Reference Laboratory of the Brazilian Ministry of Health, University of Brasilia, Federal District, Midwestern Brazil. The marmoset carcass was fresh at necropsy and showed splenomegaly and hepatomegaly with an enhanced lobular pattern (Figure 1). Tissue samples were collected and stored at −20°C and also fixed in 10% neutral buffered formalin, paraffin-embedded, and stained with hematoxylin and eosin (H&E) stain for microscopic evaluation. Histologically, we observed multifocal random hepatocellular necrosis within scattered neutrophils and histiocytes and numerous free and intrahistiocytic intralesional protozoal tachyzoites (Figure 2). The lungs showed moderate interstitial pneumonia with bradyzoite cysts consistent with T. gondii (Figure 3), which were also similarly observed in the spleen.
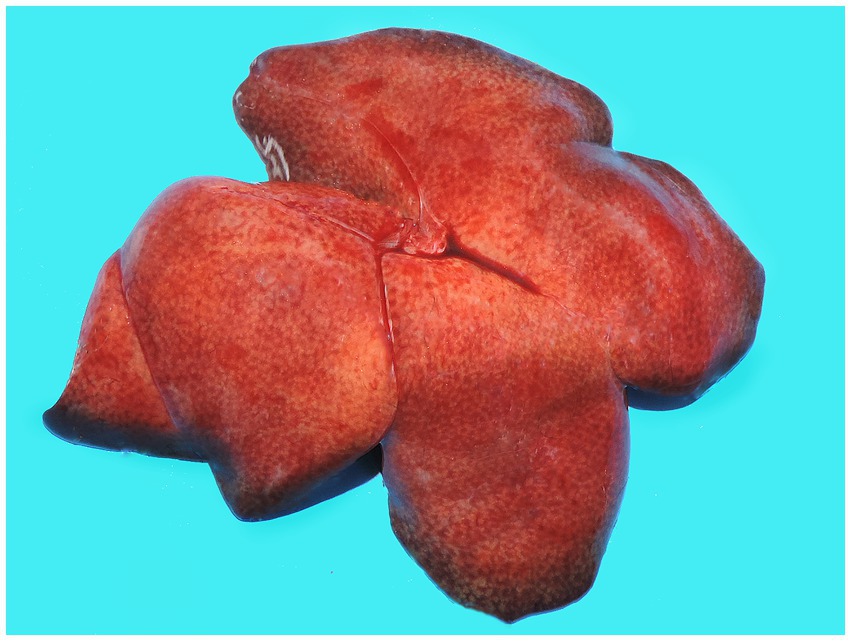
Figure 1. Gross hepatic changes in a case of acute fatal toxoplasmosis in coinfection with the YFV in an urbanized black-tufted marmoset (Callithrix penicillata). Hepatomegaly with an enhanced lobular pattern.
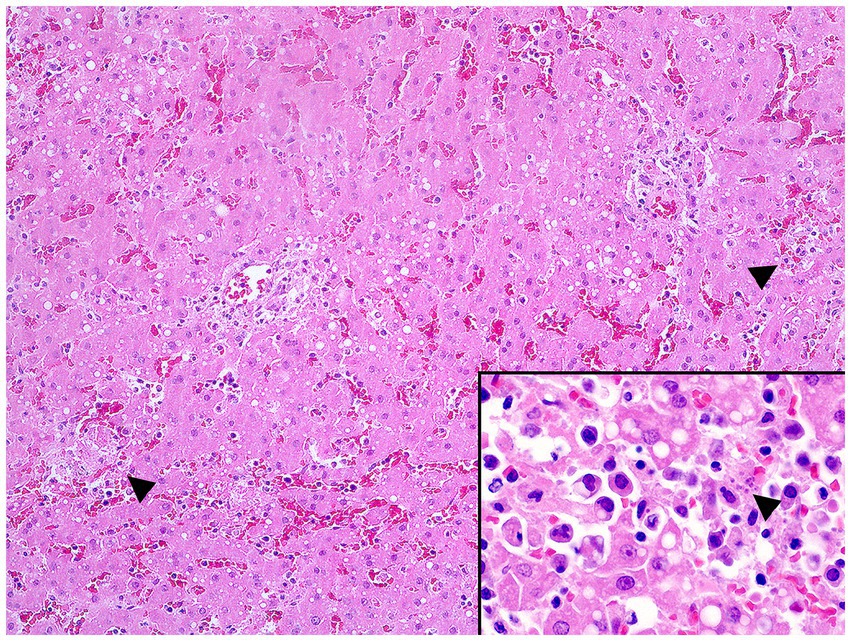
Figure 2. Multifocal random hepatocellular necrosis (arrowheads) (H&E, objective 10X) and free protozoal tachyzoites within a necrotic focus (arrowhead, inset) in the liver.
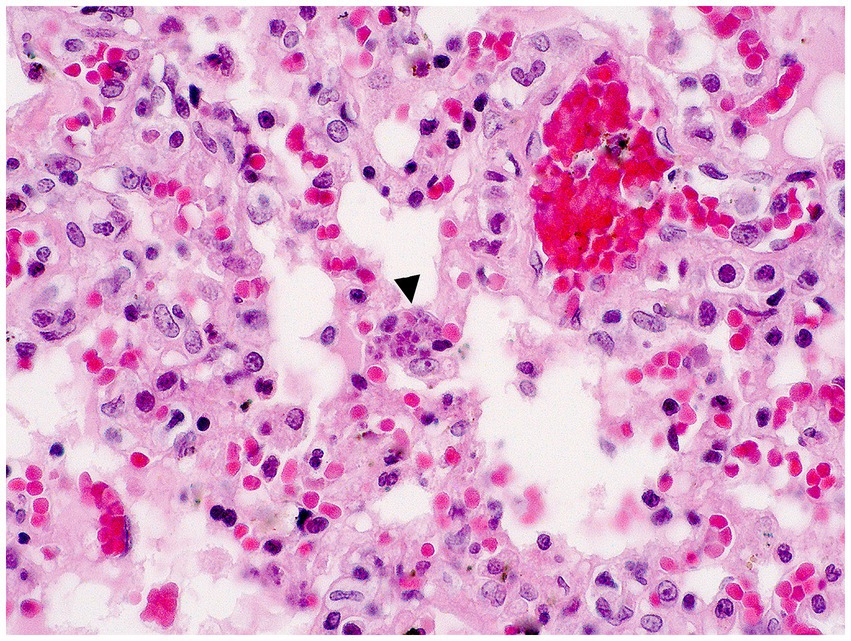
Figure 3. Interstitial pneumonia and intralesional protozoan tachyzoites (arrowhead) in the lung (H&E, objective 40X).
We conducted immunohistochemical (IHC) assays for T. gondii and the YFV in paraffin-embedded tissue samples using anti-T. gondii polyclonal antibody (VMRD USA, dilution 1:2,000, incubated overnight) and anti-YFV monoclonal antibody (ACM3A8-C12 in-house produced by Fiocruz/PR, Brazilian Ministry of Health, dilution 1:200, incubated for 2 hours), and the alkaline phosphatase method (MACH 4 Universal AP-Polymer Kit, Biocare Medical), reaction revealed with a red chromogen (Warp Red Chromogen Kit, Biocare Medical) counterstained with Mayer’s hematoxylin. Antigen retrieval was performed with citrate pH 6.0 solution, 125°C, for 3 min in a pressure cooker (11).
IHC revealed immunostaining of T. gondii tachyzoites and bradyzoite cysts in the liver (Figure 4), lungs, and spleen samples, while YFV antigens were not detected in the liver. We used conventional polymerase chain reaction (cPCR) in frozen liver samples to identify T. gondii DNA.

Figure 4. Free protozoan tachyzoites in the liver immunostained for Toxoplasma gondii within necrotic and inflammatory foci (IHC/AP, objective 20X) and in a close view (inset).
RNA was extracted from a tissue sample (liver) using the QIAamp Viral RNA Mini KitTM (Qiagen, Hilden, Germany) or the MagMAX Pathogen RNA/DNA Kit (Life Technologies, Carlsbad, United States) according to the manufacturer’s instructions. YFV RNA was detected using the RT-qPCR protocol previously described (12), with a cycle threshold value of 32.
The positive sample was submitted to a cDNA synthesis protocol (13) using a ProtoScript II First Strand cDNA Synthesis Kit. A multiplex tiling PCR was then performed using the previously published YFV primer scheme and 30 cycles of PCR using Q5 high-fidelity DNA polymerase (NEB) as previously described (14). Amplicons were purified using AMPure XP beads (Beckman Coulter), and cleaned-up PCR product concentrations were measured using a Qubit Double-stranded DNA (dsDNA) High Sensitivity (HS) Assay Kit on a Qubit 3.0 fluorometer (Thermo Fisher). DNA library preparation was performed using the Ligation Sequencing Kit (Oxford Nanopore Technologies) and the Native Barcoding Kit (NBD103; Oxford Nanopore Technologies, Oxford, United Kingdom). A sequencing library was generated from the barcoded products using the genomic DNA sequencing kit SQK-MAP007/SQK-LSK208 (Oxford Nanopore Technologies). The sequencing library was loaded onto a R9.4 flow cell (Oxford Nanopore Technologies).
Raw files were base called using Guppy v4.5.4, and barcode demultiplexing was performed using qcat. Consensus sequences were generated by de novo assembling using Genome Detective1 (15). The yellow fever typing tool (13) further revealed that the new genome sequence obtained in this study (GenBank accession number OP508570) represented a new sub-lineage belonging to the YFV South American-1 genotype (16) and was subsequently named YFV PA/MG sub-lineage (17).
Considering the complexity of interactions between NHPs and human populations in urbanized environments, the detection of a fatal case of toxoplasmosis in coinfection with the YFV in a free-ranging marmoset highlights the importance of robust and broad laboratory-based diagnosis for infectious agents of public health concern in a wildlife animal as a sentinel for zoonosis. This case also emphasizes the need for constant public health surveillance services improvement, especially since both YF and toxoplasmosis can cause fatal outbreaks in marmosets (9, 10, 18) and less frequently in humans (19–21), and the geographical and clinicopathological features overlap.
The genomic surveillance analysis showed a new sub-lineage of the YFV in Central Brazil that initially emerged in the Amazon (Pará State, Brazil) and, for the first time, two distinct YFV sub-lineages have caused simultaneous outbreaks in non-human primates (NHPs) in very distinct regions and biomes of Brazil (6, 22). This new YFV PA/MG sub-lineage identified in the Brazilian Cerrado (a savannah-like biome) has been spreading in a north-to-south axis from Amazonia (North Brazil) to other regions since 2017, which promoted fatal outbreaks in NHPs detected in 2020/2021 in Central Brazil (6, 17, 22). Concomitantly, a second sublineage named YFV MG/SP/RS has been detected since 2018, causing epizootics in NHPs in Southern Brazil (22).
Epizootics of YF and toxoplasmosis are primarily characterized by severe liver damage in non-human primates, with some similarities in the gross aspect, but they have distinct histopathological features (18, 23). Midzonal to panlobular necrosis, Councilman body formation, and microvesicular steatosis are the main features of YF virus-induced liver damage (18). While in toxoplasmosis, mild to moderate random liver necrosis with a variable lymphohistiocytic inflammatory infiltrate intensity and intralesional apicomplexan zoites are the most significant pathological hallmarks (10, 23). In the urbanized marmoset, all pathological findings supported the diagnosis of acute fatal toxoplasmosis and no morphological changes indicated the YFV infection.
Surprisingly, the YFV coinfection molecularly confirmed in the black-tufted marmoset and with unremarkable hepatic virus-induced injury was an unexpected finding with great public health concern. Previous studies have shown that natural YFV infections were detected without causing viral liver damage and with a low viral load in marmosets found dead from other causes in urban and natural areas (4, 24). YFV infections occur in susceptible human and non-human primate populations in areas where mosquito vectors are present (18). Haemagogus spp. and Sabethes spp. mosquitos are the primary vectors for the sylvatic cycle of YF but have also been detected in periurban and forested areas of highly urbanized cities in Brazil (25, 26).
Regarding the urban zoonotic transmission of YF and this case of a YFV-infected marmoset, several factors must be taken into account: the presence of favorable conditions for the exposure of susceptible people; unvaccinated susceptible individuals; most marmosets usually have a low viral load, and their role in the maintenance and transmission of YF is not fully understood; and others (1, 4, 25, 27). Therefore, it is crucial to consider the risk of infected vector mosquitoes in the YFV transmission and maintenance in urbanized marmoset populations and also in the zoonotic viral spread for unvaccinated humans in natural parks and forested areas in the cities.
As observed in the marmoset of this study, New World NHPs are considered very susceptible to acute fatal toxoplasmosis, developing severe liver damage, interstitial pneumonia, and other systemic lesions in different organs (7, 10, 23). Severe toxoplasmosis is often associated with immunosuppression in humans, causing systemic lesions such as encephalitis, pneumonia, and hepatitis (28–30), but no such association has been documented in NHPs (31).
T. gondii is considered one of the most successful parasites worldwide, infecting around a third of the global population due to its ability to affect humans and a variety of warm-blooded animals, which is considered a pillar in successful protozoan dissemination and survival (32). Toxoplasmosis is also among the main foodborne parasitic diseases in the US and Europe (31, 32) and is a global public health concern, especially in developing countries (33, 34). In similar conditions observed in the marmoset, anthropized areas may play a critical role in the T. gondii spread from domestic cats to wildlife where they may come into contact (35).
T. gondii infections have mainly been reported in captive NHPs (7, 23) and with a few outbreaks in free-ranging marmosets (9, 10). Even though NHPs are not considered to be directly involved in the chain of disease transmission to humans (7), outbreaks of fatal toxoplasmosis in urbanized marmosets may serve as an indicator of potential environmental risk for T. gondii infection to people in a determined area or region.
Coinfections can significantly impact the transmission, clinical progression, and control of human infectious diseases, such as in the case of immunocompromised patients (such as those with AIDS) experiencing acute toxoplasmosis or being exposed to multiple other opportunistic infections. To date, coinfection of the YFV with other pathogens has not been reported in humans worldwide, but it is important to include it in the differential diagnosis of other icterohemorrhagic diseases such as acute leptospirosis (27).
Assessing the risk of infection for human zoonotic pathogens, such as the YFV and T. gondii, in a metropolitan environment is challenging and still requires further research. NHPs can be both victims and reservoirs of zoonotic diseases such as YF and environmental sentinels for toxoplasmosis, playing a crucial role in the One Health framework. With robust complementary molecular methods, permanent epidemiological and pathological investigations of deaths in NHPs are essential for surveillance programs to prevent human cases of lethal zoonoses in vulnerable populations, particularly during YFV outbreaks. The role of NHPs as ecological sentinels for human toxoplasmosis is still overlooked, and further studies are needed to determine its relevance. Urbanized free-ranging NHPs living as our neighbors in backyards, gardens, and other forested areas in cities may serve as warning signs about the emergence of dangerous human pathogens.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found at: https://www.ncbi.nlm.nih.gov/genbank/, OP508570.
This study was conducted on an animal found dead in the City, collected by the Health Surveillance Service and taken for a necropsy at the Official Regional Laboratory for Diagnosis of YF, Brazillian Ministry of Health. This Lab is part of the National Program for the Control and Prevention of YF.
DS, TW, IM, and MC: data curation—original draft. AR, DR, PP, GC, VF, MM-G, MG, LA, AF, GP, and CM: writing—review. All authors contributed to the article and approved the submitted version.
This work was supported in part by the National Institutes of Health USA grant U01 AI151698 for the United World Arbovirus Research Network (UWARN).
The authors thank the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brazil (CAPES) for financing in part (Finance Code 001) the doctoral scholarships of Davi Emanuel Ribeiro de Sousa and Isabel Luana de Macêdo. The authors MC, LA, and MG thank the fellows of research productivity (PQ) granted by the National Council for Scientific and Technological Development (CNPq-Brazil). MG is funded by PON “Ricerca e Innovazione” 2014–2020. MG is supported in part by the CRP- ICGEB RESEARCH GRANT 2020 Project CRP/BRA20-03, Contract CRP/20/03.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Cunha, MS , Costa, AC , Fernandes, NCCA , Guerra, JM , Santos, FCP , Nogueira, JS, et al. Epizootics due to yellow fever virus in São Paulo state, Brazil: viral dissemination to new areas (2016–2017). Sci Rep. (2019) 7:1–13. doi: 10.1038/s41598-019-41950-3
2. Kotait, I , Oliveira, RN , Carrieri, ML , Castilho, JG , Macedo, CI , Pereira, PMC, et al. Non-human primates as a reservoir for rabies virus in Brazil. Zoonoses Public Health. (2019) 66:47–59. doi: 10.1111/zph.12527
3. Goes de Jesus, J , Gräf, T , Giovanetti, M , Mares-Guia, MA , Xavier, J , Lima Maia, M, et al. Yellow fever transmission in non-human primates, Bahia, Northeastern Brazil. PLoS Negl Trop Dis. (2020) 14:e0008405. doi: 10.1371/journal.pntd.0008405
4. Mares-Guia, M , Horta, MA , Romano, A , Rodrigues, C , Mendonça, M , Dos Santos, CC, et al. Yellow fever epizootics in non-human primates, Southeast and Northeast Brazil (2017 and 2018). Parasit Vectors. (2020) 13:90. doi: 10.1186/s13071-020-3966-x
5. Guerra, JM , Ferreira, CSDS , Díaz-Delgado, J , Takahashi, JPF , Kimura, LM , Araújo, LJT, et al. Concurrent yellow fever and pulmonary aspergillosis due to Aspergillus fumigatus in a free-ranging howler monkey (Alouatta sp). J Med Primatol. (2021) 50:201–4. doi: 10.1111/jmp.12522
6. Andrade, MS , Campos, FS , Oliveira, CH , Oliveira, RS , Campos, AAS , Almeida, MAB, et al. Fast surveillance response reveals the introduction of a new yellow fever virus sub-lineage in 2021, in Minas Gerais, Brazil. Mem Inst Oswaldo Cruz. (2022):117. doi: 10.1590/0074-02760220127
7. Dubey, JP , Murata, FH , Cerqueira-Cézar, CK , Kwok, OC , Yang, Y , and Su, C . Recent epidemiologic, clinical, and genetic diversity of toxoplasma gondii infections in non-human primates. Res Vet Sci. (2021) 136:631–41. doi: 10.1016/j.rvsc.2021.04.017
8. Pereira, AABG , Dias, B , Castro, SI , Landi, MFA , Melo, CB , Wilson, TM, et al. Electrocutions in free-living black-tufted marmosets (Callithrix penicillata) in anthropogenic environments in the Federal District and surrounding areas, Brazil. Primates. (2020) 61:321–9. doi: 10.1007/s10329-019-00760-x
9. Sousa, DER , Wilson, TM , Gonçalves, AAB , Ritter, JM , Passos, PHO , Romano, AP, et al. Investigating epizootics: acute fatal toxoplasmosis in urbanized free-ranging black-tufted marmosets (Callithrix penicillata). Chicago. IL: ACVP Virtual Meeting Abstract. American College of Veterinary Pathology (ACVP) Annual Meeting (2020).
10. Rodrigues, OA , Ritter, JM , Santos, O , Lucena, FP , Mattos, SA , Parente de Carvalho, TP, et al. Pathology and epidemiology of fatal toxoplasmosis in free-ranging marmosets (Callithrix spp.) from the Brazilian Atlantic forest. PLoS Negl Trop Dis. (2022) 16:e0010782. doi: 10.1371/journal.pntd.0010782
11. Bollweg, BC , Silva-Flannery, L , Spivey, P , and Hale, GL . Optimization of commercially available Zika virus antibodies for use in a laboratory-developed immunohistochemical assay. J Pathol Clin Res. (2017) 4:19–25. doi: 10.1002/cjp2.84
12. Domingo, C , Patel, P , Yillah, J , Weidmann, M , Méndez, JA , Nakouné, ER, et al. Advanced yellow fever virus genome detection in point-of-care facilities and reference laboratories. J Clin Microbiol. (2012) 50:4054–60. doi: 10.1128/JCM.01799-12
13. Faria, NR , Kraemer, MUG , Hill, SC , Goes de Jesus, J , Aguiar, RS , Iani, FCM, et al. Genomic and epidemiological monitoring of yellow fever virus transmission potential. Science. (2018) 361:894–9. doi: 10.1126/science.aat7115
14. Quick, J , Grubaugh, ND , Pullan, ST , Claro, IM , Smith, AD , and Gangavarapu, K . Multiplex PCR method for MinION and Illumina sequencing of Zika and other virus genomes directly from clinical samples. Nat Protoc. (2017) 12:1261–76. doi: 10.1038/nprot.2017.066
15. Vilsker, M , Moosa, Y , Nooij, S , Fonseca, V , Ghysens, Y , and Dumon, K . Genome detective: an automated system for virus identification from high-throughput sequencing data. Bioinformatics. (2019) 35:871–3. doi: 10.1093/bioinformatics/bty695
16. Giovanetti, M , Mendonça, MCL , Fonseca, V , Mares-Guia, MA , Fabri, A , Xavier, J, et al. Yellow fever virus re-emergence and spread in Southeast Brazil, 2016–2019. J Virol. (2019) 12:e01623–19. doi: 10.1128/JVI.01623-19
17. Oliveira, CH , Andrade, MS , Campos, FS , Cardoso, J , Gonçalves-Dos-Santos, ME , and Oliveira, RS . Yellow fever virus maintained by Sabethes mosquitoes during the dry season in Cerrado, a semiarid region of Brazil, in 2021. Viruses. (2023) 15:757. doi: 10.3390/v15030757
18. Passos, PHO , Ramos, DG , Romano, AP , Cavalcante, KRLJ , Miranda, LHM , Coelho, JMCO, et al. Hepato-pathological hallmarks for the surveillance of yellow fever in South American non-human primates. Acta Trop. (2022) 231:106468. doi: 10.1016/j.actatropica.2022.106468
19. Painter, JA , Hoekstra, RM , Ayers, T , Tauxe, RV , Braden, CR , Angulo, FJ, et al. Attribution of foodborne illnesses, hospitalizations, and deaths to food commodities by using outbreak data, United States, 1998–2008. Emerg Infect Dis. (2013) 19:407–15. doi: 10.3201/eid1903.111866
20. Scallan, E , Hoekstra, RM , Angulo, FJ , Tauxe, RV , Widdowson, MA , Roy, SL, et al. Foodborne illness acquired in the United States--major pathogens. Emerg Infect Dis. (2011) 17:7–15. doi: 10.3201/eid1701.p11101
21. Demar, M , Ajzenberg, D , Maubon, D , Djossou, F , Panchoe, D , Punwasi, W, et al. Fatal outbreak of human toxoplasmosis along the Maroni River: epidemiological, clinical, and parasitological aspects. Clin Infect Dis. (2007) 45:88–95. doi: 10.1086/521246
22. Andrade, MS , Campos, FS , Campos, AAS , Abreu, FVS , Melo, FL , Sevá, ADP, et al. Real-time genomic surveillance during the 2021 re-emergence of the yellow fever virus in Rio Grande do Sul state, Brazil. Viruses. (2021) 13:197. doi: 10.3390/v13101976
23. Santana, CH , Oliveira, AR , Dos Santos, DO , Pimentel, SP , Souza, LDR , and Moreira, LGA . Genotyping of toxoplasma gondii in a lethal toxoplasmosis outbreak affecting captive howler monkeys (Alouatta sp.). J Med Primatol. (2021) 50:99–107. doi: 10.1111/jmp.12506
24. Fernandes, N , Guerra, JM , Díaz-Delgado, J , Cunha, MS , Saad, LD , Iglezias, SD, et al. Differential yellow fever susceptibility in New World nonhuman primates, comparison with humans, and implications for surveillance. Emerg Infect Dis. (2021) 27:47–56. doi: 10.3201/eid2701.191220
25. Abreu, FVS , Ferreira-de-Brito, A , Azevedo, AS , Linhares, JHR , Oliveira Santos, V , Hime, ME, et al. Survey on non-human primates and mosquitoes does not provide evidences of spillover/spillback between the urban and sylvatic cycles of yellow fever and Zika viruses following severe outbreaks in Southeast Brazil. Viruses. (2020) 12:364. doi: 10.3390/v12040364
26. Carvalho, MESD , Naves, HAM , Carneiro, E , and Miranda, MF . Distribuição vertical de mosquitos dos géneros Haemagogus e Sabethes, em zona urbana de Goiânia-Goiás-Brasil. J Trop Pathol. (1997) 26:173–1. doi: 10.5216/rpt.v26i1.17371
27. Meregildo-Rodriguez, ED , and Villegas-Chiroque, M . Fiebre amarilla selvática con serología positiva para leptospira en un varon joven peruano. Rev Peru Med Exp Salud Publica. (2019) 36:700–4. doi: 10.17843/rpmesp.2019.364.4347
28. Hryzhak, IH . Invasion with toxoplasma gondii can promote pneumocystis pneumonia in individuals with HIV/AIDS. Folia Parasitol. (2021) 68:1–5. doi: 10.14411/fp.2021.018
29. Fard, SA , Khajeh, A , Khosravi, A , Mirshekar, A , Masoumi, S , Tabasi, F, et al. Fulminant and diffuse cerebral toxoplasmosis as the first manifestation of HIV infection: a case presentation and review of the literature. Case Rep Clin Pract Rev. (2020) 21:e919624:e919624–1. doi: 10.12659/AJCR.919624
30. Atılla, A , Aydin, S , Demırdöven, AN , and Kiliç, SS . Severe Toxoplasmic Hepatitis in an Immunocompetent Patient. Jpn J Infect Dis. (2015) 68:407–9. doi: 10.7883/yoken.JJID.2014.422
31. Dubey, JP . Outbreaks of clinical toxoplasmosis in humans: five decades of personal experience, perspectives and lessons learned. Parasit Vectors. (2021) 14:1–12. doi: 10.1186/s13071-021-04769-4
32. Djurković-Djaković, O , Dupouy-Camet, J , Van der Giessen, J , and Dubey, JP . Toxoplasmosis: overview from a one health perspective. Food Waterborne Parasitol. (2019) 15:e00054. doi: 10.1016/j.fawpar.2019.e00054
33. Mohamedahmed, KA , Eltayeb, EA , Omer, IT , Tamomh, AG , Talha, AA , and Abakar, AD . Toxoplasmosis in Sudan: an overview. Int J Med Parasitol Epidemiol. (2021) 2:31–2. doi: 10.34172/ijmpes.2021.09
34. Qamar, W , and Alsayeqh, AF . A review of foodborne toxoplasma gondii with a special focus on its prevalence in Pakistan from 2000 to 2022. Front Vet Sci. (2023) 9:1080139. doi: 10.3389/fvets.2022.1080139
Keywords: non-human primates, toxoplasma gondii , infectious diseases, surveillance, One Health, zoonosis
Citation: Sousa DER, Wilson TM, Macêdo IL, Romano APM, Ramos DG, Passos PHO, Costa GRT, Fonseca VS, Mares-Guia MAMM, Giovanetti M, Alcantara LCJ, de Filippis AMB, Paludo GR, Melo CB and Castro MB (2023) Case report: Urbanized non-human primates as sentinels for human zoonotic diseases: a case of acute fatal toxoplasmosis in a free-ranging marmoset in coinfection with yellow fever virus. Front. Public Health. 11:1236384. doi: 10.3389/fpubh.2023.1236384
Edited by:
Galileu Barbosa Costa, United States Food and Drug Administration, United StatesReviewed by:
Izabela Rezende, Stanford University, United StatesCopyright © 2023 Sousa, Wilson, Macêdo, Romano, Ramos, Passos, Costa, Fonseca, Mares-Guia, Giovanetti, Alcantara, de Filippis, Paludo, Melo and Castro. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Márcio B. Castro, bWJjYXN0cm9AdW5iLmJy
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.