 Katherine M. J. McMurray
Katherine M. J. McMurray Renu Sah
Renu Sah
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Psychiatry, 29 November 2022
Sec. Anxiety and Stress Disorders
Volume 13 - 2022 | https://doi.org/10.3389/fpsyt.2022.1015349
This article is part of the Research TopicWomen in Psychiatry 2022: Anxiety and Stress DisordersView all 3 articles
Panic disorder (PD) is unique among anxiety disorders in that the emotional symptoms (e.g., fear and anxiety) associated with panic are strongly linked to body sensations indicative of threats to physiological homeostasis. For example, panic attacks often present with feelings of suffocation that evoke hyperventilation, breathlessness, or air hunger. Due to the somatic underpinnings of PD, a major focus has been placed on interoceptive signaling and it is recognized that dysfunctional body-to-brain communication pathways promote the initiation and maintenance of PD symptomatology. While body-to-brain signaling can occur via several pathways, immune and humoral pathways play an important role in communicating bodily physiological state to the brain. Accumulating evidence suggests that neuroimmune mediators play a role in fear and panic-associated disorders, although this has not been systematically investigated. Currently, our understanding of the role of immune mechanisms in the etiology and maintenance of PD remains limited. In the current review, we attempt to summarize findings that support a role of immune dysregulation in PD symptomology. We compile evidence from human studies and panic-relevant rodent paradigms that indicate a role of systemic and brain immune signaling in the regulation of fear and panic-relevant behavior and physiology. Specifically, we discuss how immune signaling can contribute to maladaptive body-to-brain communication and conditioned fear that are relevant to spontaneous and conditioned symptoms of PD and identify putative avenues warranting future investigation.
A complex interplay and engagement of the central nervous system (CNS) and periphery is key to the genesis of emotional responses (1). Peripheral modulation of the-emotional responses was first postulated early under the peripheral feedback theory by William James (2) and later championed as interoceptive emotional modulation by several investigators (3, 4). Dysregulated interoceptive processing and maladaptive emotional responses are a hallmark of fear –associated disorders, particularly Panic Disorder (PD), a debilitating psychiatric illness which occurs in ∼4–8% of Americans (5–7). PD typically begins in the second decade of life (8) and is second only to major depressive disorder in terms of associated debility among psychiatric conditions in the United States (9). PD is characterized by recurrent panic attacks that consist of incapacitating periods of acute-onset respiratory, cardiovascular, gastrointestinal autonomic, and cognitive symptoms. According to the DSM-5 (10), recurrent panic attacks in PD are categorized as being either spontaneous (unexpected) or cued (expected). Recurrence of panic attacks leads to anticipatory anxiety, conditioned fear and avoidance of panic contexts, cues and reminders leading to compromised functioning and disability (11–15). Current treatments have limited therapeutic efficacy and a delayed onset of action (15–17). While studies in the past few decades have improved our understanding of panic neurobiology [reviewed in: (18–20)], the mechanistic basis of spontaneous panic and sustained fear is still poorly understood yet could lead to improvements in treatment outcomes.
The association of PD with dysregulated interoceptive processing suggests an important role for body-to-brain signaling in PD pathology. While body-to-brain signaling can occur via several pathways, immune and humoral pathways play an important role in communicating bodily physiological state to the brain (21). Immune activation in conjunction with humoral interoceptive mechanisms can mediate discrete changes in brain and behavior that may predispose to psychiatric disorders (21–24). Strong evidence supports a role for the immune system and associated cellular mediators in regulation of behaviors associated with depression and anxiety such as sickness behavior, anhedonia and learned helplessness [reviewed in (22–25)]. Accumulating evidence suggests that neuroimmune regulatory mechanisms also play a role in fear and panic-associated disorders, although this has not been well studied. Interestingly, mounting epidemiological evidence supports a high comorbidity of PD with inflammatory conditions such as Crohn’s disease, asthma, inflammatory bowel syndrome (IBS) and fibromyalgia (26–31), suggesting a potential role of dysregulated immune signaling in PD pathology. However, our understanding of the role of immune mediators in the etiology and maintenance of PD remains limited.
Thus, in the current review, we attempt to summarize findings that support a role of immune dysregulation in PD symptomology. We compile evidence from human studies and panic-relevant rodent paradigms that indicate a role of systemic and brain immune signaling, in the regulation of fear and panic-relevant behavior and physiology. Specifically, we discuss how immune signaling can contribute to maladaptive body-to-brain communication and conditioned fear that are associated with spontaneous and conditioned aspects in PD onset and maintenance.
Clinical observations and collective evidence from challenge studies in the laboratory, neuroimaging, symptomology, treatment responses and translational animal models have led to an increased understanding of PD (11, 12, 14, 18–20, 32–41). As illustrated in Figure 1, PD frequently originates with patients experiencing spontaneous panic attacks that seem to occur without an explicit trigger. Over time, PD develops as the result of associative conditioning processes that lead to fear and phobic avoidance as well as anticipatory anxiety of future attacks (15). Thus, to improve our understanding of PD and develop novel treatments, clinical and preclinical work has sought to understand the mechanisms underlying these various aspects of panic pathology, including both spontaneous panic attacks and attacks driven by conditioned responses, as well as the relationship between these processes.
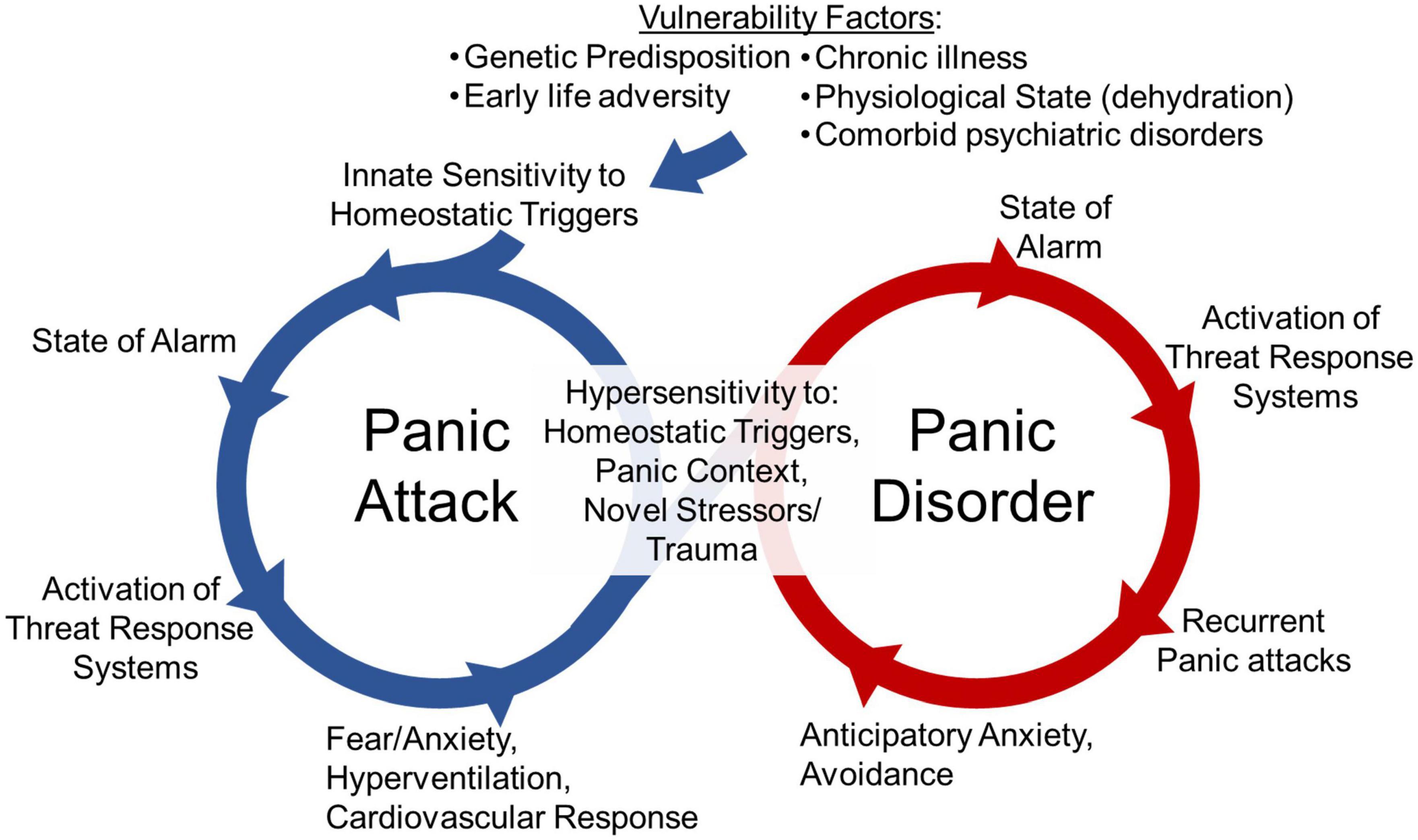
Figure 1. The cycle of panic disorder: Vulnerability factors such as genetic predisposition, early life adversity, chronic illness, physiological state (i.e., dehydration), or other psychiatric disorder diagnosis associate with innate sensitivity to homeostatic triggers. These homeostatic triggers are threats to internal homeostasis that lead to a heightened state of alarm, the activation of threat responses systems and a panic attack. The emotional, behavioral and physiological responses occurring during panic attacks such as fear, anxiety, hyperventilation and cardiovascular responses are evoked in an effort restore physiological homeostasis. The engagement of these systems and the conditioned responses to contexts where panic attacks occurred can lead to hypersensitivity to future homeostatic triggers, novel stressors or trauma, and exposure to previous panic contexts. This can lead to a cycle of recurrent panic attacks, and anticipatory anxiety and avoidance of panic-associated contexts, that ultimately facilitates the development of panic disorder.
Panic disorder is highly heterogenous with variable symptom profile and intensity in panic episodes experienced by the same individual and across patients (14, 15, 42). Interestingly, PD is unique among anxiety disorders in that fear and anxiety associated with panic are primarily directed toward somatic symptoms (32). In these individuals, body sensations or physiological signals linked to risk of suffocation (e.g., dyspnea, breathlessness, or air hunger) elicit dysfunctional defensive responding leading to anxious apprehension, fear, panic, contributing to the persistence of PD symptomatology. Due to the somatic underpinnings of PD, a major focus has been placed on interoceptive signaling and body-to-brain communication pathways, as interoceptive inputs serve an important regulatory function in generation of adaptive behaviors and physiology key to emotional regulation (3, 4, 37). Indeed, strong evidence now supports a primary role of homeostasis and interoception “an individual’s sensing and monitoring of the physiological condition of the body itself” in driving panic attacks (3, 4, 37, 43). In particular, it has been proposed that while the expected or cued panic attacks in PD are triggered by exteroceptive triggers (i.e., context of previous panic attack or other unrelated stressors, or traumatic experiences), the unexpected or spontaneous panic attacks may be provoked by interoceptive sensory triggers caused by fluctuations in the internal milieu that challenge homeostasis (37).
This theory of interoceptive signaling driving PD is primarily based on observations that patients with PD show heightened sensitivity to homeostatic disturbances, particularly those that induce acidosis such as CO2 inhalation and sodium lactate infusion (18, 38, 40, 44–62). In people with PD, these threats to internal homeostasis drive increased fear, respiratory, and cardiac responses that can result in panic attacks, suggesting this heightened sensitivity may confer vulnerability to panic attacks (38, 58). The ability to evoke panic attacks in clinical populations within the lab has greatly improved our understanding of the behavioral, emotional and physiological aspects of panic attacks. It has also allowed researchers to investigate the effects of pharmacological treatments or other therapies (i.e., cognitive behavioral therapy, etc.). Additionally, it has allowed for quantification of molecular biomarkers within blood or saliva immediately before and after panic attacks.
CO2 inhalation is the interoceptive stressor most commonly used to probe the mechanisms underlying panic attacks and PD. In this model, individuals are exposed to either low-dose CO2 inhalation (5–7.5% CO2) or a single beath of high-dose 35% CO2 by breathing in air composed of non-hypoxic levels of oxygen (∼21%) and varied nitrogen levels used to balance differing CO2 percentages (19, 34, 38, 45, 46, 49, 55, 56, 58, 59, 63, 64). CO2 inhalation evokes fear, anxiety and physiological responses in both patients with PD and in healthy individuals (38, 45, 46, 55). Responses to CO2 inhalation increase in intensity as the concentration of CO2 increases and tend to be stable across time (53). Interestingly, CO2 sensitivity seems to lie on a spectrum. Compared to individuals without PD, patients with PD show a left-shifted CO2 inhalation response curve, presenting with heightened emotional and physiological responses, and a greater likelihood of a resultant panic attack across all CO2 inhalation concentrations. First-degree family members of patients with PD present with an intermediate CO2 sensitivity phenotype between healthy individuals and those with PD suggesting there may be a genetic influence on this phenotype (38). Although not clearly defined, predisposition factors associated with PD such as genetics, early life adversity, underlying respiratory abnormalities and other factors are thought to promote sensitivity to interoceptive stressors like CO2 inhalation (18, 20, 52, 53, 61, 65, 66). Given the strength of the association between sensitivity to interoceptive stressors and PD, studies seeking to understand the mechanisms driving CO2 sensitivity may improve our understanding of PD and lead to identification of novel treatments.
Although spontaneous panic attacks are key to panic etiology, the development to PD also involves associative conditioning processes leading to fear and phobic avoidance (15, 32, 37, 67, 68). This can be particularly detrimental to quality of life in patients as they begin to avoid situations or contexts they believe may elicit panic attacks. Studies exploring conditioned fear responses, particularly contextual associations, have improved our understanding of PD and aided the development of behavioral therapies (32, 37, 67, 68). Studies investigating the transition from initial spontaneous panic attacks to development of conditioned fear are more limited yet understanding this transition may improve our ability to prevent the development of PD or improve treatments. Previous evidence suggests that initial panic attacks associated with dysfunctional interoceptive body-to-brain signaling could lead to sensitized fear-arousal-stress regulatory circuits to promote the chronicity and maintenance of PD (see Figure 1). However, the specific mechanisms by which dysfunctional interoceptive signaling could promote sensitized fear-arousal-stress responding are unclear.
Given the relevance of systemic and CNS interactions and crosstalk in panic genesis, the immune system provides an interesting regulatory pathway that can provide mechanistic insights on panic physiology. Historically, interoceptive signaling has largely focused on neural-mediated mechanisms (4, 33, 37, 41) however humoral signals are increasingly being recognized as important mediators in communicating bodily physiological state to the brain (21). Immune mediators and pathways can regulate neuronal activity and function leading to altered behavior and physiology (69–72). Reciprocally, sympathetic and neuroendocrine signals from the brain regulate immune response and function (24, 73–75). Relevance of immune mediators and inflammation in fear associated disorders such as PTSD is recognized (75–77). Whether dysfunctional immune signaling in the periphery or brain contributes to panic symptomology has not been systematically assessed. In the following sections we provide support for a potential PD-neuroimmune link based on evidence from genetic studies and cytokine measurements in human studies as well as panic-relevant animal paradigms simulating interoceptive and conditioned fear aspects of PD.
Mounting evidence from the clinic supports a potential role of dysregulated immune function in PD. Most work has focused on identifying specific genes, epigenetic associations or alterations in immune effectors comparing individuals with PD to those without a PD diagnosis that we will discuss below:
1) Immune-associated genes in Panic Disorder
Genetic studies indicate a heritability of PD at about 40% (78) suggesting a strong role for genetic variance in mediating risk to develop PD. Identifying the specific genes or gene clusters (groups of genes involved in similar functions or pathways) associated with PD could help improve our understanding of the mechanisms underlying PD and point to putative therapeutic targets. Multiple genetic studies have identified associations between immunomodulatory genes and PD. Single nucleotide polymorphisms (SNPs) within the INF-γ (+ 874 A/T), TNF-α (-308 G/A), and IL-10 (-1082 G/A) genes were investigated in patients with PD (79). The group reported that the G allele in IL-10-1082 G/A might have a protective role in reducing the manifestations of PD in female patients. IL-10 is an anti-inflammatory cytokine inhibiting the generation of several inflammatory cytokines (80, 81). Interestingly, IL-10 inhibits the nearly ubiquitous expression of indoleamine 2,3-dioxygenase (IDO), an enzyme responsible for directing tryptophan degradation, a pathway that has been implicated in anxiety disorders (82–84). Another study found an association of polymorphisms in the IKBKE (inhibitor of kappa light polypeptide gene enhancer in B cells, kinase epsilon) gene in patients with PD (85). IKBKE is involved in regulation of innate immunity, inhibiting NF-kappa B signaling in response to inflammatory cytokines, particularly IL-1 (85, 86), suggesting a potential role of innate immune signaling in PD. Association of a MASP-2 YA haplotype and Mannan-binding lectin (MBL) deficiency was reported in patients with PD, an observation explained to be associated with innate immune alterations that may increase susceptibility for infections and autoimmune states due to their roles in complement activation (87). In strong support of an immune dysfunction in PD, Shimada-Sugimoto et al. (88), performed pathway analyses in order to overcome the limitations of conventional single-marker analysis in identifying associated SNPs with modest effects. Using multiple pathway analyses the group reported that pathways related to immunity show the strongest association with PD. For further investigation, the group studied HLA polymorphisms in candidate susceptibility genes HLA-B and HLA-DRB1, associated with the major histocompatibility complex (MHC) and immune dysfunction in PD patients and control subjects (89). Patients with PD were significantly more likely to carry HLA-DRB1 further supporting links to immune regulation and genes involved in immune-related pathways are associated with PD.
In addition to gene polymorphism and pathway analysis studies, epigenetic contributions have been investigated in the context of immune modulation. A recent study investigated whether aberrant DNA methylation of inflammation-related genes was associated in the development of PD (90). Methylation levels of CCL3, CRP, CSF2, CXCL8, IFNG, IL12B, IL1A, IL-4, IL-6, TNF was investigated. Significantly higher methylation levels of the IL-4 gene were observed in PD patients than control subjects. Importantly, the methylation levels of IL-4 gene showed a significant positive correlation with the severity of panic and anxiety (90). In another study, (91), a significant association between panic severity score and methylation levels of Asb1, a member of the suppressors of cytokine signaling (SOCS) family was observed in PD subjects, suggesting a role of epigenetic factors. Furthermore, in a follow up mouse study, the authors found a correlation between peripheral Asb1 and IL-1β mRNA expression after acute social defeat stress suggesting a relationship between Asb1, IL-1β and stress responding (91). Assessment of T cell receptor excision circles (TRECs), Forkhead-Box-Protein P3 gene (FOXP3) methylation of regulatory T cells (Tregs) and relative telomere lengths (RTLs) was conducted in patients with PD and age- and sex-matched healthy controls in order to test for a potential dysfunction and premature aging of the immune system (92). Significantly reduced TRECs in PD patients and FOXP3 hypermethylation in female patients with PD was observed, reflecting immune system-related deficits in PD.
Other studies support differences in the expression of immune-associated genes in panic pathology. Our lab found significantly higher expression of immunomodulatory gene T-cell death associated gene 8 (TDAG8) in peripheral monocytes collected from PD subjects and healthy volunteers (93). TDAG8, also expressed in brain microglia, is an acid sensing GPCR regulating CO2-associated fear (94). Interestingly, a significant positive correlation was observed between monocytic TDAG8 expression and panic symptom severity score and TDAG8 expression was lower in individuals with symptom remission, suggesting potential utility of TDAG8 as a treatment response biomarker (93). Lastly, one group, Maron et al. (95) conducted gene expression profiling following cholecystokinin CCK-4, a commonly used panic provocation agent challenge in control subjects. Interestingly, several immune regulatory genes showed alterations between “panickers” and “non-panickers,” suggesting that acute panicogenesis may engage immune system targets.
Collectively, genetic evidence suggests dysregulation of immune function and possibly epigenetic mechanisms in panic etiology. Although some association with immunomodulatory genes have been reported in PD, more follow up investigation is warranted.
2) Immune effectors in PD
Measurement of alterations in immune mediators such as cytokines, chemokines and T cells have been conducted in individuals with PD. Several studies have reported altered cytokine concentrations in PD, however directionality and targets assessed differ between studies and are not always consistent. In general, a broad spectrum of cytokines appear to be upregulated in PD (96, 97) suggesting dysregulated immune signaling, however, specific associations and contribution to panic physiology is not well understood. Due to variability in study layouts and for correct interpretation, we have divided available evidence into measurements performed at baseline (no challenge) or following a panic-relevant challenge or stressor.
i) Immune alterations in panic disorder under non-challenge conditions
Proinflammatory mediators within the interleukin (IL) family have been the most studied in the context of PD with measurements reported on IL-6, IL-1β, IL-2, IL-3, IL-12, IL-10, and TNFα [see (97)]. Multiple studies have shown elevated IL-6 concentrations in the serum of PD subjects (96, 98–100). One study showed that while patients with both PD and generalized anxiety disorder (GAD) showed higher IL-6 compared to healthy individuals, PD subjects had even greater IL-6 than individuals with GAD (98). Interestingly, the utility of IL-6 as a potential treatment response biomarker is supported by significantly lower IL-6 concentrations in individual with remitting symptoms compared to those with current panic symptoms (99). Furthermore, another study observed as association of pretreatment IL-6 with poor treatment response (100). However, other studies have reported no significant changes in IL-6 in PD patients (101, 102), suggesting that factors other than a panic diagnosis may contribute to IL-6 alterations.
The IL-1 family of cytokines have also been investigated in PD. A study by Brambilla et al. (103) measured IL-1β plasma concentrations before and after treatment with alprazolam, and reported significantly higher IL1β both before and after treatment (103). However, other studies have reported no associations of IL-1α/IL-1β with PD (102, 104). These inconsistencies have been contributed to differences in assay methodologies (97).
C-reactive protein (CRP), an inflammatory marker shows significant elevation in patients with PD as compared to healthy controls (105). Following 8 weeks of selective serotonin reuptake inhibitor (SSRI) treatment, CRP concentrations decreased only in treatment-responsive individuals. Additionally, the authors found reduced γ globulin and higher cortisol levels in PD patients compared to controls prior to initiation of treatment. Collectively, these observations suggest engagement of the acute phase response in PD.
Measurement of broad spectrum panel of cytokines and inflammatory markers such IL−6, IL−1α, IL−1β, IL−8, MCP−1, MIP−1α, Eotaxin, GM−CSF, and IFN−α revealed that 87% of individuals with PD or PTSD had six or more detectable levels of these cytokines, compared with only 25% of control (96). In PD patients compared to controls, 17 of 20 cytokines and chemokines examined showed significant elevation, suggesting that PD may associate with a generalized inflammatory state and that specific “inflammatory signatures” may differ on an individual basis. This could associate with the general heterogeneity in symptomology shown within PD patients.
Among other members of the interleukin family, higher IL-2 concentrations were reported in PD subjects compared to healthy controls by one study (104), however no associations were reported by other studies (101, 102, 106). One study reported increased IL-18 concentrations in PD the magnitude of which was comparable to a concurrently tested group of depression subjects (107).
ii) Immune alterations post stressor in patients with PD
The ability to respond appropriately to stressors and adapt is critical to survival by increasing alertness and preparing the body to fight (108). Engagement of stress responses with the immune system may have originated from a need to prepare the body to fight infection from wounds (109). Mounting evidence suggests that immune system engagement may also regulate fear learning which may be beneficial to survival by helping an individual remember that a fearful environment should be avoided (109). Immune responses to stressors occur in healthy individuals, but evidence suggests these responses may be dysregulated in those with psychiatric diagnoses (22, 70, 73, 110). As both homeostatic and psychogenic stressors are relevant to panic physiology (Figure 1), investigating immune-associated alterations following panic-relevant stressors in individuals with PD can provide valuable information.
To date, only one study directly studied alterations in immune factors pre- and post -CO2 inhalation (101), a well-established interoceptive stressor in PD described above. No significant differences in either baseline or post CO2 concentrations of TNF-α, IL-6, IL-8, IL-10, IL-1RA, soluble sIL-2R, soluble sIL-6R or positive acute phase protein haptoglobin were observed. Lipopolysaccharide-stimulated cytokine production (TNF-α, IL-6, IL-1β, IL-10, IFN-γ) in whole blood was comparable for PD patients and their matched controls. Limitations of the study suggested by the authors included: the low number of subjects and a gender-biased sample with underrepresentation of women. Given the relevance of panic provocation challenges in PD, more larger “n” studies are warranted; not only for CO2 inhalation, but other interoceptive challenges such as sodium lactate.
Dysfunctional stress response systems such as the hypothalamic pituitary adrenal (HPA) axis and sympathetic nervous system (SNS) have been reported in PD [reviewed in (111)]. These stress response systems have bidirectional regulatory associations with the immune system, with either system engaging the other to regulate immune and stress responses (24, 112–114). Accumulating evidence indicates stress-neuroimmune interactions in anxiety disorders including PD (115). Given strong evidence of immune dysregulation within PD patients under non-stressful conditions, it is important to determine whether immune responses to stressful stimuli are dysregulated and whether immune dysregulation could contribute to CO2 sensitivity in patients with PD.
Exposure to acute psychological stressors have previously been reported to show engagement with the immune system, particularly cytokine release. For example, public speaking or performing complex tasks in public elevated plasma cytokines and CRP (116–118). One study compared immune mediators in healthy individuals and those with PD following the Trier Social Test (TSST) (119). They found PD patients have higher baseline and post stressor concentrations of IL-10, and also show blunted cortisol responses. Interestingly, peak IL-6 concentration associated with PD symptom severity. Heightened startle responses have also been reported in PD patients compared to healthy controls only under fear-associated threat conditions (120). Interestingly, there is an association between heightened startle reactivity and inflammation (75).
Collectively, evidence suggests altered immune mediators in PD, that may represent underlying immune dysfunction, however, additional studies especially following panic-relevant triggers and stress challenges are required for understanding their association with PD physiology. Human studies are limited and cannot provide mechanistic information on brain neuroimmune alterations and association with panic-relevant behaviors. Furthermore, it is difficult to probe neurocircuits orchestrating body to brain signaling. In the following section, we discuss selected paradigms that may be relevant to understanding the mechanistic association of immune dysregulation and panic associated behavior and physiology.
Several panic-relevant rodent models have been developed over the last few decades that provide valuable insights on panic pathophysiology. Consistent with the importance of uncued/spontaneous and cued triggers in generating behavioral and physiological responses in PD, these models have focused on homeostatic and stress challenges, respectively [reviewed in (18–20, 34, 48, 52, 121–124)]. Previously reviewed literature has primarily focused on acid-base homeostasis, PD-relevant neurotransmitter systems, region-targeted interventions, genetic and transgenic manipulations. Despite emerging evidence from human studies, contribution of immune mechanisms in panic and fear has not been systematically investigated using translational paradigms. In this section, we assess selected rodent paradigms that provide information on the potential role of immune cells, targets and signaling that may regulate panic-relevant responses and provide mechanistic insights on panic-fear-immune links (see Table 1).

Table 1. Animal studies of immune regulation of spontaneous/conditioned fear.
1) CO2 inhalation and microglial acid sensing by TDAG8
As described in preceding sections, homeostatic triggers that promote acid-base imbalance such as low-dose, non-hypoxic CO2 inhalation induce intense fear, anxiety, physiological responses and panic attacks in PD subjects (18, 19, 34, 38, 41, 47, 56). Several investigators including our laboratory, have used CO2 inhalation to simulate panic-relevant behavior and physiology in mice and rats (48, 65, 94, 125–133). The partial pressure of CO2 in the blood and CNS increases following CO2 inhalation challenge. In the extracellular fluid, CO2 is hydrolyzed to carbonic acid (H2CO3) by carbonic anhydrase which readily dissociates into bicarbonate (HCO3–) and H+ (134). The resulting acidosis constitutes a homeostatic threat that is sensed by acid-sensing chemosensors in the brain and periphery (18, 19, 34, 38, 41, 47, 56). This model has strong translational value due to its strong face, predictive and etiological validity [discussed in: (18, 19, 48, 127)].
Given its strong translational value, CO2 inhalation in rodents is commonly used to investigate mechanisms underlying PD to simulate panic-relevant behaviors and physiology. To date there have been somewhat limited investigations into the role of immune dysregulation in mediating PD-relevant behaviors and physiological outcomes in CO2 inhalation models. However, support for immune dysregulation in these models is growing. Our lab cloned acid-sensing G protein coupled receptor, T- cell death associated gene-8 (TDAG8) in rodent brain (135), and reported TDAG8 expression on microglia, innate immune cells of the CNS (94), that are recruited in physiological responses to homeostatic fluctuations (136–140). On sensing subtle imbalance in ionic homeostasis, microglia transform rapidly from a resting to an activated state in accordance with their role in maintenance of the CNS microenvironment. Interestingly, TDAG8-expressing microglia were enriched within the sensory circumventricular organs (CVOs) (94). Sensory CVOs such as the subfornical organ (SFO) are integrative sites that lack a blood-brain barrier and have access to systemic and CNS compartments for maintenance of homeostasis (141). Importantly, the SFO has been identified as a site where interoceptive stimuli can be sensed and relayed to panic-generating CNS areas, and has been implicated in regulating panic-relevant responses to other panicogens such as intravenous lactate (142–144). More directly, our data showed targeted infusion of acidified artificial cerebrospinal fluid (aCSF; a homeostatic stress) into the SFO triggers fear-relevant behaviors like freezing; an effect which is dependent on TDAG8 (145). Our data further revealed the necessity of microglial acid-sensor TDAG8 to orchestrate CO2-evoked behavioral (freezing) and cardiovascular responses relevant to panic via microglial activation, IL-1β signaling, and SFO neuronal firing (94). Thus, the SFO likely represents a primary locus for detecting interoceptive triggers in “body-to brain” transmission of panic.
Recruitment of SFO immunomodulatory mechanisms in panic-relevant outcomes is novel, although not surprising, as the SFO is also known to drive immune responses and sickness behaviors, particularly fever (141, 146–148). The SFO also regulates blood pressure (149) as well as respiration (150), and previous studies reported IL-1β-IL-1R signaling mediated cardiovascular activation via the SFO (151). More recent observations from our lab show significant attenuation of CO2-evoked freezing following SFO-targeted infusion of the IL-1 receptor antagonist, IL-1RA (152), suggesting a primary role of SFO IL-1R1 signaling in CO2-associated fear behaviors. Notably, the IL-1R1 receptor is localized on endothelial cells within the SFO (152, 153) and is required for CO2-evoked activation of SFO neurons (94). The SFO could be mediating these effects though its direct projections to brain areas mediating defensive behaviors and physiological responses relevant to panic such as the hypothalamus, prefrontal cortex, BNST, and periaqueductal gray (154, 155). Collectively, our data highlight a contributory role of neuroimmune signaling and specialized sites such as the SFO in regulation of panic-relevant behavior and physiology.
2) Early Life Stress- CO2 respiratory dysfunction paradigm
Strong evidence supports significant variance and individual differences in responding to CO2 inhalation (38, 47, 156). People with panic lie at one extreme of the sensitivity distribution, showing paroxysmal hyperventilation and panic when they undergo CO2 challenges (38, 52, 58, 61, 157). Modeling CO2 sensitivity in rodents as a proxy for human panic has unique strategic advantages such as an opportunity to dissect mechanistic contributions. Studies performed by Battaglia and coworkers have highlighted gene x environment interactions in mediating increased CO2 sensitivity [reviewed in (158–160)]. Chronic stress in the form of early life adversity for example childhood parental loss, contributes to these relationships (158, 161), and stressful life events occurring in childhood-adolescence heighten young adults’ CO2 sensitivity (162).
Translationally relevant paradigms such as neonatal maternal separation (NMS) and repeated cross fostering (RCF) model have effectively simulated the effects of early life stress on CO2 sensitivity and provided mechanistic insights [reviewed in (158, 160)]. Heightened ventilatory response to CO2 inhalation is observed in adult animals with a history of NMS or RCF as neonates (65, 126, 163–167), that may be regulated by gene x environment factors and epigenetic alterations in gene methylation patterns (163, 166). Interestingly, significant increase in microglial density and reduced arborization was observed within the nucleus of the solitary tract (cNTS) and the dorsal motor nucleus of the vagus (DMNV), two key areas regulating breathing (168) suggesting that NMS may compromise microglial ability to perform optimal synaptic pruning that could lead to aberrant respiratory control. In a more recent study (169), the group reported a significant effect of ovarian hormones on microglial activation within in CO2/H+ sensing brain stem areas, suggesting that hormonal fluctuations may influence anomalies of respiratory control via neuroimmune mechanisms. Given that effects of early life stress on immune signaling, neuroimmune physiology and function are well established [for review see (110, 170–172)], more investigation is warranted on the delineating the links between ELS-immune signaling and panic-relevant physiology.
3) Chronic Social Defeat paradigms - body to brain immune signaling in anxiety and fear
As described in preceding sections, exposure to exteroceptive stressors and aversive contexts can facilitate the recurrence of panic attacks. Furthermore, dysregulation of stress response systems has been implicated in PD, particularly the sympathetic nervous system that regulates the immune system via innervation of the spleen and lymphoid organs [Webster Marketon and Glaser (173); Dantzer (174)]. A primary role of chronic stress mediated immune dysfunction and CNS remodeling has been recognized in the genesis of psychiatric illnesses, including mood and anxiety disorders [reviewed in (24, 71, 72, 175)]. Recurrent or chronic stress engages intrinsic neuronal pathways that lead to physiological effects, such as neuroendocrine and sympathetic activation (176, 177). These systemic responses to stress further promote the release and trafficking of myeloid cells with enhanced inflammatory potential into various organs, including the brain (178, 179). Preclinical rodent models of stress-induced behavior and physiology can provide important information on brain-to-body as well as body-to-brain communication mechanisms. In this regard, psychosocial stress paradigms, particularly repeated social defeat (RSD) stress exposure [reviewed in (114, 179–181)] have yielded important mechanistic information on body-brain neuroimmune crosstalk in regulating fear extinction (182), anxiety and social avoidance behaviors [reviewed in (179)] that are relevant to persistent fear, anxiety and comorbid depression observed in PD. Coordinated events including activation of brain threat appraisal sites, microglial activation and peripheral immune signaling is reported to orchestrate RSD-mediated behavioral deficits. Exposure to RSD leads to increased sympathetic activation promoting elevated noradrenergic signaling in the periphery and increased production and release of glucocorticoid-insensitive monocytes into the circulation from the bone marrow and spleen [reviewed in (114, 183)]. Pharmacological and genetic intervention strategies using adrenergic receptor antagonists, microglial activation blockers, cell specific IL-1R transgenic mice highlight a key role of sympathetic activation, microglia-mediated trafficking of reactive IL-1β- releasing monocytes to the brain, and the recruitment of endothelial and neuronal IL-1R1 in promoting RDS induced expression of anxiety-associated behaviors and reduced social interaction (184–187). Although IL-1-mediated mechanisms appear to be important in the RDS model, it would be important to note that other cytokine mediators, like interleukin-6 have also been implicated in social defeat stress- induced blood brain barrier alterations and impaired social interaction behavior (188–191). In addition to adverse behavioral effects, social defeat exposure has long term effects on cardiovascular responses (192, 193) and aberrant respiratory patterns (194) that are of relevance to PD. Immune dysfunction and inflammatory mechanisms contribute to increased cardiovascular and autonomic activation in stressed mice (195, 196).
In summary, previous work has highlighted the relevance of social defeat stress paradigms to anxiety and depression physiology. However, the observed crosstalk of sympathetic activation, peripheral immune signaling and brain networks regulating behavior and physiological responses in this model may also relate to panic physiology.
4) Fear Conditioning paradigms
Pavlovian fear conditioning has been central to understanding the etiology of anxiety disorders such as PD [reviewed in (197)]. The clinical relevance of fear conditioning to PD is based on conditioned fear responses to panic-evoking stimuli and contexts, as well as generalized fear to resembling cues. Conditioning processes contribute by conferring fear and anxiogenic valence to these conditioning triggers that enable the maintenance of the disorder well after the termination of the unconditioned stimulus (i.e., the panic attack). The measurement of Pavlovian fear conditioning and extinction in rodents offers a relatively simple paradigm that is translatable as an approach to study biological underpinnings of fear-related disorders (198–201). While most rodent studies highlight the applicability of their findings to PTSD, they are also pertinent to PD symptomology. In this section we discuss evidence supporting a role of several immune factors and neuroimmune signaling in regulating conditioned fear associated behaviors. We discuss selected fear conditioning studies in this section excluding models of neurodegeneration, addiction or severe inflammatory insults.
A role of IL-6 in the maintenance of fear memory was reported using auditory cue fear conditioning, an effects dependent on fear extinction, suggesting that IL-6 and other IL-6 related pro-inflammatory cytokines may contribute to the persistence of fear memory (76). Another study reported impairment of acquisition and extinction of fear following intra-amygdala injection of IL-6 (202). Fear regulatory effects of other cytokines and T cells have also been investigated. Levels of TNF-α were increased in microglia from mice representing retention of fear memory, and returned to basal levels in mice representing extinction (203), suggesting that sustained fear is facilitated by microglial TNF-α signaling. A role of NLRP3 inflammasome activation and IL-1β has also been reported in the regulation of fear memory (204). Genetic knockout and pharmaceutical inhibition of the NLRP3 inflammasome enhanced the extinction of contextual fear memory. IL-1β administration inhibited extinction, while, IL-1RA (IL-1R antagonist) facilitated extinction. In addition to cytokines associated with innate immune signaling, the role of CD4(+) T cells in promoting fear responses by enhancing learning and memory processes has been reported (205). Lymphocyte deficient Rag2(-/-) mice showed attenuated fear responses in a cued fear conditioning paradigm compared to wild-type mice and reconstitution with CD4(+) T cells promoted fear learning and memory.
Stress-enhanced fear learning (SEFL), which encompasses both trauma and memory components in rodents, has emerged as a valuable preclinical model for PTSD [reviewed in (206)], and may be pertinent to the conditioned aspects of PD physiology. SEFL simulates the impact of traumatic stress in the form of several footshocks on subsequent fear conditioning to a single shock delivered in a novel context (207). Maladaptive behavioral outcomes following traumatic footshocks in the SEFL paradigm have been associated with immune dysregulation, specifically, a central role of hippocampal IL-1β has been reported (208–210). Exposure to footshocks induced a time-dependent increase in IL-1β expression within the hippocampal dentate gyrus, and IL-1RA treatment prevented the development of SEFL (208). Interestingly, hippocampal astrocytes were reported to be the source of IL-1β (209). To confirm a key role of astrocyte signaling in SEFL, the same group investigated the effects of glial-expressing DREADD construct [AAV8-GFAP-hM4Di(Gi)-mCherry] delivered in the dorsal hippocampus (210). Inhibition of astroglial Gi activation was sufficient to attenuate SEFL, suggesting that IL-1β signaling involves glial-neuronal interactions in stress potentiation of fear.
Collective evidence from these studies suggests that immune signaling has regulatory effects on fear learning and memory processes and that dysregulated immune status in PD could promote sustained and generalized fear.
Mounting evidence supports a role for neuroinflammation in PD, however, questions remain about the specific mechanisms and effectors contributing to pathology. Clinical observations suggest that PD associates with heightened peripheral inflammation at baseline. Patients with PD also present with hyperactive homeostatic stress responsivity, particularly in response to acid-base imbalance, which associates with dysregulated immune responses to stress. Our working model (see Figure 2) proposes a complex interplay between peripheral and central inflammatory mechanisms, and a hyperactive response system to homeostatic stress. While it is currently unclear if hyperactive homeostatic stress responsivity leads to dysregulated inflammatory signaling or if a heightened inflammatory state increases sensitivity to homeostatic stressors, it is likely the relationship is bidirectional with either facilitating the other. The exacerbation of sensitivity to homeostatic stressors like acidosis and peripheral inflammation then promotes the behavioral, emotional and physiological symptoms of panic attacks through neuroinflammatory mechanisms and dysregulated neuroimmune signaling within the brain and ultimately, leads to the development of PD.
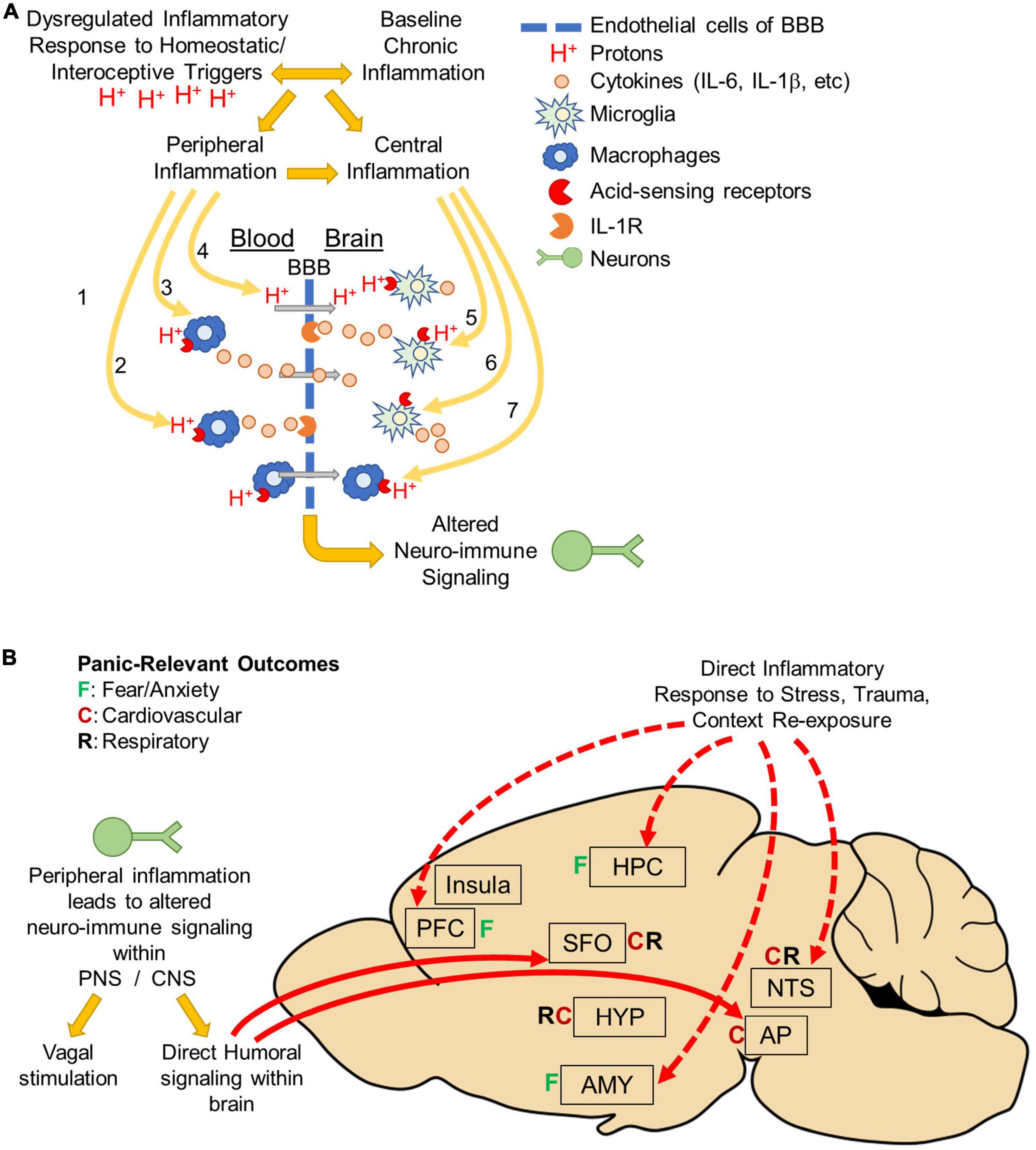
Figure 2. Body to brain signaling: Mechanisms of acidosis evoked peripheral and central inflammation: (A) panic disorder is associated with dysregulated inflammatory responses to homeostatic/interoceptive triggers like acidosis. Panic disorder is also associated with baseline chronic inflammation. Regardless of which inflammatory response occurs initially, either can facilitate the development of the other and result in peripheral and central inflammation. Acidosis could lead to altered neuroimmune signaling through a variety of mechanisms. In the periphery, acidosis may activate acid-sensing receptors on peripheral macrophages. This could lead to either (1) macrophage infiltration across the blood brain barrier (BBB), (2) cytokine release that activates cytokine receptors on endothelial cells, or (3) release of cytokines that cross the BBB. Peripheral acidosis could acidify the brain via protons (H+) that cross the BBB and activate acid-sensing receptors on microglia. Alternatively, acidosis could occur directly within the brain (5) activating acid-sensing receptors on microglia resulting in cytokine release and activation of cytokine receptors on endothelial cells or (6) directly on neurons or other cell types. It is also possible that acidosis could activate acid-sensing receptors on peripheral macrophages that have previously infiltrated the brain, possibly as a result of prior trauma or other risk factor (disease, etc.). Ultimately, Inflammation within the brain is thought to drive emotional, behavioral and physiological responses that occur during panic attacks and increase vulnerability to develop panic disorder. (B) Peripheral inflammation can alter neuronal activity either by neuro-immune signaling occurring either in the periphery or directly in brain. In the periphery, neuro-immune signaling could result in vagal stimulation that terminates in the nucleus tractus solitarius (NTS) and mediates cardiovascular and respiratory responses. There could also be direct humoral signaling within circumventricular organs the subfornical organ (SFO) or Area Postrema (AP). These areas are known to project directly to fear, respiratory and cardiovascular regulatory areas throughout the brain. Alternatively, inflammation could occur directly in the brain in response to stress (interoceptive/homeostatic or exteroceptive), trauma, or panic context re-exposure. Many studies have shown increased inflammation in fear-associated regions prefrontal cortex (PFC), hippocampus (HPC), amygdala (AMY) or the NTS. Inflammation in these areas could affect neuronal activity and drive the emotional, behavioral and physiological responses associated with panic disorder pathology.
An important question going forward involves the relationship between peripheral and central inflammation, and how inflammation reaches the brain to ultimately drive panic symptomology. Though it is unclear how inflammation could reach the brain in PD, homeostatic stressors like acidosis could lead to altered neuroimmune signaling within the brain through a variety of mechanisms (Figure 2A). In the periphery, acidosis may activate acid-sensing receptors on peripheral macrophages and lead to release of cytokines that either activate cytokine receptors on endothelial cells or cross the BBB to activate receptors directly within the brain. Alternatively, peripheral acidosis could acidify the brain via protons (H+) that cross the BBB and activate acid-sensing receptors on microglia. Another possibility is that acidosis occurs directly within the brain activating acid-sensing receptors on microglia resulting in cytokine release and activation of cytokine receptors on endothelial cells or directly on neurons or other cell types. The BBB generally protects against infiltration from peripheral immune cells, however mounting evidence suggests mood, anxiety and stress-associated psychiatric disorders like PTSD and depression associate with changes in BBB permeability (191, 211–213). In PD, it is possible that prior trauma, chronic stress or comorbidities such as PTSD or depression which associate with changes in BBB permeability could cause peripheral macrophages to penetrate the brain, which could then be activated by subsequent acidosis. It is also possible that repeated activation of peripheral macrophages could alter the BBB and lead to changes in how peripheral macrophages interact or engage with the peripheral-endothelial-BBB interphase. Future studies are needed to identify which, if any, of these mechanisms contribute to neuroinflammation in PD and determine whether targeting these mechanisms could improve treatment outcomes.
Future studies are also needed to identify how inflammatory signaling is translated into neuronal signaling within the brain to ultimately regulate panic symptomology. The conversion of immune signaling into neuronal signaling could originate either in the periphery or directly in brain (Figure 2B). In the periphery, neuro-immune signaling could result in vagal stimulation that terminates in the nucleus tractus solitarius (NTS) and mediates cardiovascular and respiratory responses (214). There could also be direct humoral signaling within circumventricular organs the subfornical organ (SFO) or area postrema (AP). These areas are known to project directly to fear, respiratory and cardiovascular regulatory areas throughout the brain (147). Alternatively, inflammation could occur directly in the brain in response to stress (interoceptive/homeostatic or exteroceptive), trauma, or panic context re-exposure. Many studies have shown increased inflammation in fear-associated regions prefrontal cortex (PFC), hippocampus (HPC), amygdala (AMY) or the NTS (75, 83, 204, 208, 209, 215–217). Inflammation in these areas could affect neuronal activity and drive the emotional, behavioral and physiological responses associated with PD pathology.
Another consideration is the level of inflammation associated with pathology. Historically, most studies on the role of innate immune cells within the brain have focused on the effects of high levels of neuroinflammation more typically associated sickness that can have more dramatic effects such as increased phagocytosis. Yet even mild perturbations in the immune milieu can have neuromodulatory effects. Innate immune cells contribute to homeostatic maintenance of the CNS and dysregulation of this functionality may lead to pathology (140). For example, microglia have an important role in the synaptic pruning (72). Dysregulated synaptic pruning could have lasting effects on neurotransmission and mounting evidence suggests an association of dysregulated synaptic pruning with psychiatric disorders like depression or autism. There is also an association between inflammation and increased oxidative stress in psychopathology which could exacerbate cell damage, alter cell signaling within the brain and result in behavioral changes (218–221). Additionally, at lower concentrations cytokines may act as neuromodulators within the brain, changing excitability and having more mild effects on cognition/memory/decision making rather than inducing robust sickness behaviors at higher concentrations (70). In the case of PD, a lack of evidence for drastic effects on immune function might be due to relatively mild changes on the innate immune system, resulting in changes to homeostatic maintenance or monocyte trafficking to endothelial cells at the BBB rather than increased cell death or infiltration of peripheral monocytes/T cells as a result of BBB degradation.
Lastly, the high level of co-morbidity between PD and inflammatory diseases such as multiple sclerosis (222), lupus (89, 223), asthma (30, 224), and irritable bowel syndrome (225), as well as other psychiatric disorders associated with inflammation like depression (6, 14, 226), PTSD (6, 227) and substance use disorders (6, 14) supports a role for dysregulated immune signaling in PD risk and pathology. However, the mechanisms driving high comorbidity are unclear and particularly complex as either PD or the inflammatory disorder may pre-date the other. For example, initial presentation with an inflammation-associated disorder could drive changes in homeostatic sensitivity and lead to increased risk for developing panic, innate sensitivity to homeostatic stressors could lead to development of a heightened inflammatory state and increase risk for developing inflammation-associated co-morbidities, or the same underlying inflammatory pathways may drive pathology in PD and commonly co-morbid inflammatory disorders and be dysregulated in affected individuals. Future studies are needed to determine the relationship between these disorders and the mechanisms through which one may lead to the other.
Overall, strong evidence supports the hypothesis that dysregulated body to brain signaling is central to the development of PD. Emerging clinical and preclinical evidence points to a strong role for dysregulated immune signaling in driving the dysfunctional body to brain signaling underlying spontaneous and cued panic attacks. However, further studies are needed to better understand neuroinflammatory mechanisms in PD.
KM and RS conceived of the project, wrote and edited the manuscript, and provided funding. Both authors contributed to the article and approved the submitted version.
This work was supported by R01MH093362 and R21MH083213 and VA Merit Grant I01-BX001075 to RS, as well as postdoctoral training award F32MH117913 and Pathway to Independence award K99AA029168 to KM.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Nord CL, Garfinkel SN. Interoceptive pathways to understand and treat mental health conditions. Trends Cogn Sci. (2022) 26:499–513. doi: 10.1016/j.tics.2022.03.004
3. Damasio A, Carvalho GB. The nature of feelings: evolutionary and neurobiological origins. Nat Rev Neurosci. (2013) 14:143–52. doi: 10.1038/nrn3403
4. Khalsa SS, Adolphs R, Cameron OG, Critchley HD, Davenport PW, Feinstein JS, et al. Interoception and mental health: a roadmap. Biol Psychiatry Cogn Neurosci Neuroimaging. (2018) 3:501–13. doi: 10.1016/j.bpsc.2018.04.007
5. Kessler RC, Chiu WT, Jin R, Ruscio AM, Shear K, Walters EE. The epidemiology of panic attacks, panic disorder, and agoraphobia in the national comorbidity survey replication. Arch Gen Psychiatry. (2006) 63:415–24. doi: 10.1001/archpsyc.63.4.415
6. de Jonge P, Roest AM, Lim CCW, Florescu SE, Bromet EJ, Stein DJ, et al. Cross-national epidemiology of panic disorder and panic attacks in the world mental health surveys. Depress Anxiety. (2016) 33:1155–77. doi: 10.1002/da.22572
7. Gros DF, Frueh BC, Magruder KM. Prevalence and features of panic disorder and comparison to posttraumatic stress disorder in VA primary care. Gen Hosp Psychiatry. (2011) 33:482–8. doi: 10.1016/j.genhosppsych.2011.06.005
8. Beesdo K, Pine DS, Lieb R, Wittchen HU. Incidence and risk patterns of anxiety and depressive disorders and categorization of generalized anxiety disorder. Arch Gen Psychiatry. (2010) 67:47–57. doi: 10.1001/archgenpsychiatry.2009.177
9. Gadermann AM, Alonso J, Vilagut G, Zaslavsky AM, Kessler RC. Comorbidity and disease burden in the national comorbidity survey replication (NCS-R). Depress Anxiety. (2012) 29:797–806. doi: 10.1002/da.21924
10. American Psychiatric Association. DSM 5. 2013. Washington, DC: American Psychiatric Association (2013).
11. Klein DF. False suffocation alarms, spontaneous panics, and related conditions. An integrative hypothesis. Arch Gen Psychiatry. (1993) 50:306–17. doi: 10.1001/archpsyc.1993.01820160076009
12. Smoller JW, Tsuang MT. Panic and phobic anxiety: defining phenotypes for genetic studies. Am J Psychiatry. (1998) 155:1152–62. doi: 10.1176/ajp.155.9.1152
13. Eaton WW, Kessler RC, Wittchen HU, Magee WJ. Panic and panic disorder in the United States. Am J Psychiatry. (1994) 151:413–20. doi: 10.1176/ajp.151.3.413
14. Craske MG, Kircanski K, Epstein A, Wittchen H-UU, Pine DS, Lewis-Fernández R, et al. Panic disorder: a review of DSM-IV panic disorder and proposals for DSM-V. Depress Anxiety. (2010) 27:93–112. doi: 10.1002/da.20654
15. Roy-Byrne PP, Craske MG, Stein MB. Panic disorder. Lancet. (2006) 368:1023–32. doi: 10.1016/S0140-6736(06)69418-X
16. Barlow DH, Gorman JM, Shear MK, Woods SW. Cognitive-behavioral therapy, imipramine, or their corn or their combination for panic disorder: a randomized controlled trial. J Am Med Assoc. (2000) 283:2529–36. doi: 10.1001/jama.283.19.2529
17. Yonkers KA, Bruce SE, Dyck IR, Keller MB. Chronicity, relapse, and illness – Course of panic disorder, social phobia, and generalized anxiety disorder: findings in men and women from 8 years of follow-up. Depress Anxiety. (2003) 17:173–9. doi: 10.1002/da.10106
18. Vollmer LL, Strawn JR, Sah R. Acid–base dysregulation and chemosensory mechanisms in panic disorder: a translational update. Transl Psychiatry. (2015) 5:e572–572. doi: 10.1038/tp.2015.67
19. Wemmie JA. Neurobiology of panic and pH chemosensation in the brain. Dialogues Clin Neurosci. (2011) 13:475–83. doi: 10.31887/DCNS.2011.13.4/jwemmie
20. Johnson PL, Federici LM, Shekhar A. Etiology, triggers and neurochemical circuits associated with unexpected, expected, and laboratory-induced panic attacks. Neurosci Biobehav Rev. (2014) 46:429–54. doi: 10.1016/j.neubiorev.2014.07.027
21. Savitz J, Harrison NA. Interoception and inflammation in psychiatric disorders. Biol Psychiatry Cogn Neurosci Neuroimaging. (2018) 3:514–24. doi: 10.1016/j.bpsc.2017.12.011
22. Capuron L, Miller AH. Immune system to brain signaling: neuropsychopharmacological implications. Pharmacol Ther. (2011) 130:226–38. doi: 10.1016/j.pharmthera.2011.01.014
23. Dantzer R. Cytokine, sickness behaviour, and depression. Immunol Allergy Clin North Am. (2009) 29:247–64. doi: 10.1016/j.iac.2009.02.002
24. Ménard C, Pfau ML, Hodes GE, Russo SJ. Immune and neuroendocrine mechanisms of stress vulnerability and resilience. Neuropsychopharmacology (2017) 42:62–80. doi: 10.1038/npp.2016.90
25. Miller AH, Raison CL. The role of inflammation in depression: from evolutionary imperative to modern treatment target. Nat Rev Immunol. (2015) 16:22–34. doi: 10.1038/nri.2015.5
26. Alciati A, Cirillo M, Masala IF, Sarzi-Puttini P, Atzeni F. Differences in depression, anxiety and stress disorders between fibromyalgia associated with rheumatoid arthritis and primary fibromyalgia. Stress Heal. (2021) 37:255–62. doi: 10.1002/smi.2992
27. Vork L, Mujagic Z, Drukker M, Keszthelyi D, Conchillo JM, Hesselink MAM, et al. The experience sampling method—evaluation of treatment effect of escitalopram in IBS with comorbid panic disorder. Neurogastroenterol Motil. (2019) 31:1–9. doi: 10.1111/nmo.13515
28. Tarter RE, Switala J, Carra J, Edwards KL. Inflammatory bowel disease: psychiatric status of patients before and after disease onset. Int J Psychiatry Med. (1987) 17:173–81. doi: 10.2190/CB02-2V8B-Y624-R781
30. Hasler G, Gergen PJ, Kleinbaum DG, Ajdacic V, Gamma A, Eich D, et al. Asthma and panic in young adults: a 20-year prospective community study. Am J Respir Crit Care Med. (2005) 171:1224–30. doi: 10.1164/rccm.200412-1669OC
31. Trkulja V, Barić H. Current research on complementary and alternative medicine (CAM) in the treatment of major depressive disorder: an evidence-based review. Adv Exp Med Biol. (2021) 1305:375–427. doi: 10.1007/978-981-33-6044-0_20
32. Bouton ME, Mineka S, Barlow DH. A modern learning theory perspective on the etiology of panic disorder. Psychol Rev. (2001) 108:4–32. doi: 10.1037/0033-295X.108.1.4
33. Gorman JM, Kent JM, Sullivan GM, Coplan JD. Neuroanatomical hypothesis of panic disorder, revised. Am J Psychiatry. (2000) 157:493–505. doi: 10.1176/appi.ajp.157.4.493
34. Leibold NK, van den Hove DLA, Esquivel G, De Cort K, Goossens L, Strackx E, et al. The brain acid-base homeostasis and serotonin: a perspective on the use of carbon dioxide as human and rodent experimental model of panic. Prog Neurobiol. (2015) 129:58–78. doi: 10.1016/j.pneurobio.2015.04.001
35. Coplan JD, Lydiard RB. Brain circuits in panic disorder. Biol Psychiatry. (1998) 44:1264–76. doi: 10.1016/S0006-3223(98)00300-X
36. Quagliato LA, Freire RC, Nardi AE. The role of acid-sensitive ion channels in panic disorder: a systematic review of animal studies and meta-analysis of human studies. Transl Psychiatry. (2018) 8:185. doi: 10.1038/s41398-018-0238-z
37. Van Diest I. Interoception, conditioning, and fear: the panic threesome. Psychophysiology. (2019) 56:1–27. doi: 10.1111/psyp.13421
38. Colasanti A, Esquivel G, Schruers KJ, Griez EJ. On the psychotropic effects of carbon dioxide. Curr Pharm Des. (2012) 18:5627–37. doi: 10.2174/138161212803530745
39. Boettcher H, Brake CA, Barlow DH. Origins and outlook of interoceptive exposure. J Behav Ther Exp Psychiatry. (2016) 53:41–51. doi: 10.1016/j.jbtep.2015.10.009
40. Maddock RJ. The lactic acid response to alkalosis in panic disorder?: an integrative review. J Neuropsychiatry Clin Neurosci. (2001) 13:22–34. doi: 10.1176/jnp.13.1.22
41. Dresler T, Guhn A, Tupak SV, Ehlis AC, Herrmann MJ, Fallgatter AJ, et al. Revise the revised? New dimensions of the neuroanatomical hypothesis of panic disorder. J Neural Transm. (2013) 120:3–29. doi: 10.1007/s00702-012-0811-1
42. Sheikh JI, Leskin GA, Klein DF. Gender differences in panic disorder: findings from the national comorbidity survey. Am J Psychiatry. (2002) 159:55–8. doi: 10.1176/appi.ajp.159.1.55
43. Pfeifer G, Garfinkel SN, Gould van Praag CD, Sahota K, Betka S, Critchley HD. Feedback from the heart: emotional learning and memory is controlled by cardiac cycle, interoceptive accuracy and personality. Biol Psychol. (2017) 126:19–29. doi: 10.1016/j.biopsycho.2017.04.001
44. do Amaral JMX, Spadaro PTM, Pereira VM, Silva AC, Nardi AE. The carbon dioxide challenge test in panic disorder: a systematic review of preclinical and clinical research. Rev Bras Psiquiatr. (2013) 35:318–31. doi: 10.1590/1516-4446-2012-1045
45. Harrison WM, Sandberg D, Gorman JM, Fyer M, Nee J, Uy J, et al. Provocation of panic with carbon dioxide inhalation in patients with premenstrual dysphoria. Psychiatry Res. (1989) 27:183–92. doi: 10.1016/0165-1781(89)90133-9
46. Bailey JE, Argyropoulos SV, Kendrick AH, Nutt DJ. Behavioral and cardiovascular effects of 7.5% CO2 in human volunteers. Depress Anxiety. (2005) 21:18–25. doi: 10.1002/da.20048
47. Fyer MR, Uy J, Martinez J, Goetz R, Klein DF, Fyer A, et al. CO2 challenge of patients with panic disorder. Am J Psychiatry. (1987) 144:1080–2. doi: 10.1176/ajp.144.8.1080
48. Leibold NK, van den Hove DLA, Viechtbauer W, Buchanan GF, Goossens L, Lange I, et al. CO2 exposure as translational cross-species experimental model for panic. Transl Psychiatry. (2016) 6:e885. doi: 10.1038/tp.2016.162
49. Coryell W, Rickels H. Effects of escitalopram on anxiety and respiratory responses to carbon dioxide inhalation in subjects at high risk for panic disorder: a placebo-controlled, crossover study. J Clin Psychopharmacol. (2009) 29:174–8. doi: 10.1097/JCP.0b013e31819a8d96
50. Rassovsky Y, Kushner MG. Carbon dioxide in the study of panic disorder: issues of definition, methodology, and outcome. J Anxiety Disord. (2003) 17:1–32. doi: 10.1016/S0887-6185(02)00181-0
51. Perna G, Brambilla F, Arancio C, Bellodi L. Menstrual cycle-related sensitivity to 35% CO2 in panic patients. Biol Psychiatry. (1995) 37:528–32. doi: 10.1016/0006-3223(94)00154-U
52. Battaglia M. Sensitivity to carbon dioxide and translational studies of anxiety disorders. Neuroscience. (2017) 346:434–6. doi: 10.1016/j.neuroscience.2017.01.053
53. Roberson-Nay R, Gorlin EI, Beadel JR, Cash T, Vrana S, Teachman BA. Temporal stability of multiple response systems to 7.5% carbon dioxide challenge. Biol Psychol. (2017) 124:111–8. doi: 10.1016/j.biopsycho.2017.01.014
54. Olsson M, Ho HP, Annerbrink K, Thylefors J, Eriksson E. Respiratory responses to intravenous infusion of sodium lactate in male and female wistar rats. Neuropsychopharmacology. (2002) 27:85–91. doi: 10.1016/S0893-133X(02)00296-8
55. Kellner M, Muhtz C, Nowack S, Leichsenring I, Wiedemann K, Yassouridis A. Effects of 35% carbon dioxide (CO2) inhalation in patients with post-traumatic stress disorder (PTSD): a double-blind, randomized, placebo-controlled, cross-over trial. J Psychiatr Res. (2018) 96:260–4. doi: 10.1016/j.jpsychires.2017.10.019
56. Papp LA, Martinez JM, Klein DF, Coplan JD, Norman RG, Cole R, et al. Respiratory psychophysiology of panic disorder: three respiratory challenges in 98 subjects. Am J Psychiatry. (1997) 154:1557–65.
57. Sanderson WC, Wetzler S. Five percent carbon dioxide challenge: valid analogue and marker of panic disorder? Biol Psychiatry. (1990) 27:689–701. doi: 10.1016/0006-3223(90)90584-O
58. Papp LA, Klein DF, Gorman JM. Carbon dioxide hypersensitivity, hyperventilation, and panic disorder. Am J Psychiatry. (1993) 150:1149–57. doi: 10.1176/ajp.150.8.1149
59. Gorman JM, Browne ST, Papp LA, Martinez J, Welkowitz L, Coplan JD, et al. Effect of antipanic treatment on response to carbon dioxide. Biol Psychiatry. (1997) 42:982–91. doi: 10.1016/S0006-3223(97)00160-1
60. Talesnik B, Berzak E, Ben-Zion I, Kaplan Z, Benjamin J. Sensitivity to carbon dioxide in drug-naïve subjects with post-traumatic stress disorder. J Psychiatr Res. (2007) 41:451–4. doi: 10.1016/j.jpsychires.2005.11.008
61. Rappaport LM, Sheerin C, Savage JE, Hettema JM, Roberson-Nay R. Clinical characteristics of latent classes of CO2hypersensitivity in adolescents and young adults. Behav Res Ther. (2017) 93:95–103. doi: 10.1016/j.brat.2017.03.015
62. Fishman SM, Carr DB, Beckett A, Rosenbaum JF. Hypercapneic ventilatory response in patients with panic disorder before and after alprazolam treatment and in pre- and postmenstrual women. J Psychiatr Res. (1994) 28:165–70. doi: 10.1016/0022-3956(94)90027-2
63. Leibold NK, Weidner MT, Ziegler C, Ortega G, Domschke K, Lesch KP, et al. DNA methylation in the 5-HTT regulatory region is associated with CO2-induced fear in panic disorder patients. Eur Neuropsychopharmacol. (2020) 36:154–9. doi: 10.1016/j.euroneuro.2020.04.011
64. Perna G, Bertani A, Caldirola D, Di Pasquale D, Migliarese G, Bellodi L. Modulation of hyperreactivity to 35% CO2 after one week of treatment with paroxetine and reboxetine: a double-blind, randomized study. J Clin Psychopharmacol. (2004) 24:277–82. doi: 10.1097/01.jcp.0000125682.97466.3c
65. D’Amato FR, Zanettini C, Lampis V, Coccurello R, Pascucci T, Ventura R, et al. Unstable maternal environment, separation anxiety, and heightened CO2 sensitivity induced by gene-by-environment interplay. PLoS One. (2011) 6:2–12. doi: 10.1371/journal.pone.0018637
66. Bandelow B, Späth C, Alvarez Tichauer G, Broocks A, Hajak G, Rüther E. Early traumatic life events, parental attitudes, family history, and birth risk factors in patients with panic disorder. Compr Psychiatry. (2002) 43:269–78. doi: 10.1053/comp.2002.33492
67. Lissek S, Rabin S, Heller RE, Lukenbaugh D, Geraci M, Pine DS, et al. Overgeneralization of conditioned fear as a pathogenic marker of panic disorder. Am J Psychiatry. (2010) 167:47–55. doi: 10.1176/appi.ajp.2009.09030410
68. Holtz K, Pané-Farré CA, Wendt J, Lotze M, Hamm AO. Brain activation during anticipation of interoceptive threat. Neuroimage. (2012) 61:857–65. doi: 10.1016/j.neuroimage.2012.03.019
69. Wu Y, Dissing-Olesen L, MacVicar BA, Stevens B. Microglia: dynamic mediators of synapse development and plasticity. Trends Immunol. (2015) 36:605–13. doi: 10.1016/j.it.2015.08.008
70. Kronfol Z, Remick DG. Cytokines and the brain: implications for clinical psychiatry. Am J Psychiatry. (2000) 157:683–94. doi: 10.1176/appi.ajp.157.5.683
71. Dwyer JB, Ross DA, Schafer DP, Lehrman EK, Stevens B, Wohleb ES, et al. Modern microglia: novel targets in psychiatric neuroscience. Biol Psychiatry. (2016) 80:e47–9. doi: 10.1016/j.biopsych.2016.08.006
72. Bollinger JL, Wohleb ES. The formative role of microglia in stress-induced synaptic deficits and associated behavioral consequences. Neurosci Lett. (2019) 711:134369. doi: 10.1016/j.neulet.2019.134369
73. Tsyglakova M, McDaniel D, Hodes GE. Immune mechanisms of stress susceptibility and resilience: lessons from animal models. Front Neuroendocrinol. (2019) 54:100771. doi: 10.1016/j.yfrne.2019.100771
74. McKim DB, Patterson JM, Wohleb ES, Jarrett BL, Reader BF, Godbout JP, et al. Sympathetic release of splenic monocytes promotes recurring anxiety following repeated social defeat. Biol Psychiatry. (2016) 79:803–13. doi: 10.1016/j.biopsych.2015.07.010
75. Michopoulos V, Powers A, Gillespie CF, Ressler KJ, Jovanovic T. Inflammation in fear-and anxiety-based disorders: PTSD, GAD, and beyond. Neuropsychopharmacology. (2017) 42:254–70. doi: 10.1038/npp.2016.146
76. Young MB, Howell LL, Hopkins L, Moshfegh C, Yu Z, Clubb L, et al. A peripheral immune response to remembering trauma contributes to the maintenance of fear memory in mice. Psychoneuroendocrinology. (2018) 94:143–51. doi: 10.1016/j.psyneuen.2018.05.012
77. Deslauriers J, Powell SB, Risbrough VB. Immune signaling mechanisms of PTSD risk and symptom development: insights from animal models. Curr Opin Behav Sci. (2017) 14:123–32. doi: 10.1016/j.cobeha.2017.01.005
78. Hettema JM, Neale MC, Kendler KS. A review and meta-analysis of the genetic epidemiology of anxiety disorders. Am J Psychiatry. (2001) 158:1568–78. doi: 10.1176/appi.ajp.158.10.1568
79. Kim HJ, Kim YK. The G allele in IL-10-1082 G/A may have a role in lowering the susceptibility to panic disorder in female patients. Acta Neuropsychiatr. (2016) 28:357–61. doi: 10.1017/neu.2016.25
80. Smith EM, Cadet P, Stefano GB, Opp MR, Hughes TK. IL-10 as a mediator in the HPA axis and brain. J Neuroimmunol. (1999) 100:140–8. doi: 10.1016/S0165-5728(99)00206-4
81. Fiorentino DF, Zlotnik A, Mosmann TR, Howard M, O’Garra A. IL-10 inhibits cytokine production by activated macrophages. J Immunol. (1991) 147:3815–22.
82. Walker AK, Wing EE, Banks WA, Dantzer R. Leucine competes with kynurenine for blood-to-brain transport and prevents lipopolysaccharide-induced depression-like behavior in mice. Mol Psychiatry. (2019) 24:1523–32. doi: 10.1038/s41380-018-0076-7
83. Dantzer R, O’Connor JC, Freund GG, Johnson RW, Kelley KW. From inflammation to sickness and depression: when the immune system subjugates the brain. Nat Rev Neurosci. (2008) 9:46–56. doi: 10.1038/nrn2297
84. Weiss G, Murr C, Zoller H, Haun M, Widner B, Ludescher C, et al. Modulation of neopterin formation and tryptophan degradation by Th1- and Th2-derived cytokines in human monocytic cells. Clin Exp Immunol. (1999) 116:435–40. doi: 10.1046/j.1365-2249.1999.00910.x
85. Traks T, Koido K, Balõtšev R, Eller T, Kõks S, Maron E, et al. Polymorphisms of IKBKE gene are associated with major depressive disorder and panic disorder. Brain Behav. (2015) 5:e00314. doi: 10.1002/brb3.314
86. Sankar S, Chan H, Romanow WJ, Li J, Bates RJ. IKK-i signals through IRF3 and NFκB to mediate the production of inflammatory cytokines. Cell Signal. (2006) 18:982–93. doi: 10.1016/j.cellsig.2005.08.006
87. Foldager L, Ko O, Steffensen R, Thiel S, Kristensen AS, Christian J, et al. Bipolar and panic disorders may be associated with hereditary defects in the innate immune system. J Affect Disord. (2014) 164:148–54. doi: 10.1016/j.jad.2014.04.017
88. Shimada-Sugimoto M, Otowa T, Miyagawa T, Umekage T, Kawamura Y, Bundo M, et al. Epigenome-wide association study of DNA methylation in panic disorder. Clin Epigenetics. (2017) 9:1–11. doi: 10.1186/s13148-016-0307-1
89. Shimada-Sugimoto M, Otowa T, Miyagawa T, Khor SS, Kashiwase K, Sugaya N, et al. Immune-related pathways including HLA-DRB1*13:02 are associated with panic disorder. Brain Behav Immun. (2015) 46:96–103. doi: 10.1016/j.bbi.2015.01.002
90. Zou Z, Huang Y, Wang J, Min W, Zhou B. DNA methylation of IL-4 gene and the association with childhood trauma in panic disorder. Psychiatry Res. (2020) 293:113385. doi: 10.1016/j.psychres.2020.113385
91. Emeny RT, Baumert J, Zannas AS, Kunze S, Wahl S, Iurato S, et al. Anxiety associated increased CpG methylation in the promoter of Asb1: a translational approach evidenced by epidemiological and clinical studies and a murine model. Neuropsychopharmacology. (2018) 43:342–53. doi: 10.1038/npp.2017.102
92. Prelog M, Hilligardt D, Schmidt CA, Przybylski GK, Leierer J, Almanzar G, et al. Hypermethylation of FOXP3 promoter and premature aging of the immune system in female patients with panic disorder? PLoS One. (2016) 11:1–14. doi: 10.1371/journal.pone.0157930
93. Strawn JR, Vollmer LL, McMurray KMJ, Mills JA, Mossman SA, Varney ST, et al. Acid-sensing T cell death associated gene-8 receptor expression in panic disorder. Brain Behav Immun. (2018) 67:36–41. doi: 10.1016/j.bbi.2017.07.014
94. Vollmer LL, Ghosal S, McGuire JL, Ahlbrand RL, Li K-Y, Santin JM, et al. Microglial Acid Sensing Regulates Carbon Dioxide-Evoked Fear. Biol Psychiatry. (2016) 80:541–51. doi: 10.1016/j.biopsych.2016.04.022
95. Maron E, Kallassalu K, Tammiste A, Kolde R, Vilo J, Tõru I, et al. Peripheral gene expression profiling of CCK-4-induced panic in healthy subjects. Am J Med Genet Part B Neuropsychiatr Genet. (2010) 153:269–74.
96. Hoge EA, Brandstetter K, Moshier S, Pollack MH, Wong KK, Simon NM. Broad spectrum of cytokine abnormalities in panic disorder and posttraumatic stress disorder. Depress Anxiety. (2009) 26:447–55. doi: 10.1002/da.20564
97. Quagliato LA, Nardi AE. Cytokine alterations in panic disorder: a systematic review. J Affect Disord. (2018) 228:91–6. doi: 10.1016/j.jad.2017.11.094
98. Zou Z, Zhou B, Huang Y, Wang J, Min W, Li T. Differences in cytokines between patients with generalised anxiety disorder and panic disorder. J Psychosom Res. (2020) 133:109975. doi: 10.1016/j.jpsychores.2020.109975
99. Belem Da Silva CT, Costa MDA, Bortoluzzi A, Pfaffenseller B, Vedana F, Kapczinski F, et al. Cytokine levels in panic disorder: evidence for a dose-response relationship. Psychosom Med. (2017) 79:126–32. doi: 10.1097/PSY.0000000000000384
100. Kim K, Jang EH, Kim AY, Fava M, Mischoulon D, Papakostas GI, et al. Pre-treatment peripheral biomarkers associated with treatment response in panic symptoms in patients with major depressive disorder and panic disorder: a 12-week follow-up study. Compr Psychiatry (2019) 95:152140. doi: 10.1016/j.comppsych.2019.152140
101. Van Duinen MA, Schruers KRJ, Kenis GRL, Wauters A, Delanghe J, Griez EJL, et al. Effects of experimental panic on neuroimmunological functioning. J Psychosom Res. (2008) 64:305–10. doi: 10.1016/j.jpsychores.2007.10.004
102. Tükel R, Arslan BA, Ertekin BA, Ertekin E, Oflaz S, Ergen A, et al. Decreased IFN-γ and IL-12 levels in panic disorder. J Psychosom Res. (2012) 73:63–7. doi: 10.1016/j.jpsychores.2012.04.012
103. Brambilla F, Bellodi L, Perna G, Bertani A, Panerai A, Sacerdote P. Plasma interleukin-1 beta concentrations in panic disorder. Psychiatry Res. (1994) 54:135–42. doi: 10.1016/0165-1781(94)90002-7
104. Rapaport MH, Stein MB. Serum cytokine and soluble interleukin-2 receptors in patients with panic disorder. Anxiety. (1994) 1:22–5. doi: 10.1002/anxi.3070010106
105. Herrán A, Sierra-Biddle D, García-Unzueta MT, Puente J, Vázquez-Barquero JL, Amado JA. The acute phase response in panic disorder. Int J Neuropsychopharmacol. (2005) 8:529–35. doi: 10.1017/S1461145705005432
106. Weizman R, Laor N, Wiener Z, Wolmer L, Bessler H. Cytokine production in panic disorder patients. Clin Neuropharmacol. (1999) 22:107–9. doi: 10.1097/00002826-199903000-00008
107. Kokai M, Kashiwamura S, Okamura H, Ohara K, Morita Y. Plasma interleukin-18 levels in patients with psychiatric disorders. J Immunother. (2002) 25(Suppl 1):68–71. doi: 10.1097/00002371-200203001-00011
108. McEwen BS, Akil H. Revisiting the stress concept: implications for affective disorders. J Neurosci. (2020) 40:12–21. doi: 10.1523/JNEUROSCI.0733-19.2019
109. Rachal Pugh C, Fleshner M, Watkins LR, Maier SF, Rudy JW. The immune system and memory consolidation: a role for the cytokine IL-1β. Neurosci Biobehav Rev. (2001) 25:29–41. doi: 10.1016/S0149-7634(00)00048-8
110. Nettis MA, Pariante CM, Mondelli V. Early-life adversity, systemic inflammation and comorbid physical and psychiatric illnesses of adult life. Curr Top Behav Neurosci. (2020) 44:207–25. doi: 10.1007/7854_2019_89
111. Abelson JL, Khan S, Liberzon I, Young EA. HPA axis activity in patients with panic disorder: review and synthesis of four studies. Depress Anxiety. (2007) 24:66–76. doi: 10.1002/da.20220
112. Moynihan J, Kruszewska B, Madden K, Callahan T. Sympathetic nervous system regulation of immunity. J Neuroimmunol. (2004) 147:87–90. doi: 10.1016/j.jneuroim.2003.10.018
113. Moynihan JA. Mechanisms of stress-induced modulation of immunity. Brain Behav Immun. (2003) 17:11–6. doi: 10.1016/S0889-1591(02)00060-0
114. Wohleb ES, McKim DB, Sheridan JF, Godbout JP. Monocyte trafficking to the brain with stress and inflammation: a novel axis of immune-to-brain communication that influences mood and behavior. Front Neurosci. (2015) 9:447. doi: 10.3389/fnins.2014.00447
115. Won E, Kim YK. Neuroinflammation-associated alterations of the brain as potential neural biomarkers in anxiety disorders. Int J Mol Sci. (2020) 21:1–19. doi: 10.3390/ijms21186546
116. Steptoe A, Hamer M, Chida Y. The effects of acute psychological stress on circulating inflammatory factors in humans: a review and meta-analysis. Brain Behav Immun. (2007) 21:901–12. doi: 10.1016/j.bbi.2007.03.011
117. Deinzer R, Granrath N, Stuhl H, Twork L, Idel H, Waschul B, et al. Acute stress effects on local Il-1β responses to pathogens in a human in vivo model. Brain Behav Immun. (2004) 18:458–67. doi: 10.1016/j.bbi.2003.11.008
118. Von Känel R, Kudielka BM, Preckel D, Hanebuth D, Fischer JE. Delayed response and lack of habituation in plasma interleukin-6 to acute mental stress in men. Brain Behav Immun. (2006) 20:40–8. doi: 10.1016/j.bbi.2005.03.013
119. Petrowski K, Wichmann S, Kirschbaum C. Stress-induced pro- and anti-inflammatory cytokine concentrations in panic disorder patients. Psychoneuroendocrinology. (2018) 94:31–7. doi: 10.1016/j.psyneuen.2018.05.005
120. Grillon C, Ameli R, Goddard A, Woods SW, Davis M. Baseline and fear-potentiated startle in panic disorder patients. Biol Psychiatry. (1994) 35:431–9. doi: 10.1016/0006-3223(94)90040-X
121. Santos M, D’Amico D, Dierssen M. From neural to genetic substrates of panic disorder: insights from human and mouse studies. Eur J Pharmacol. (2015) 759:127–41. doi: 10.1016/j.ejphar.2015.03.039
122. Schenberg LC, Schimitel FG, Armini Rde S, Bernabé CS, Rosa CA, Tufik S, et al. Translational approach to studying panic disorder in rats: hits and misses. Neurosci Biobehav Rev. (2014) 46:472–96. doi: 10.1016/j.neubiorev.2014.10.002
123. Kinkead R, Tenorio L, Drolet G, Bretzner F, Gargaglioni L. Respiratory manifestations of panic disorder in animals and humans: a unique opportunity to understand how supramedullary structures regulate breathing. Respir Physiol Neurobiol. (2014) 204:3–13. doi: 10.1016/j.resp.2014.06.013
124. Moreira FA, Gobira PH, Viana TG, Vicente MA, Zangrossi H, Graeff FG. Modeling panic disorder in rodents. Cell Tissue Res. (2013) 354:119–25. doi: 10.1007/s00441-013-1610-1
125. Tenorio-Lopes L, Fournier S, Henry MS, Bretzner F, Kinkead R. Disruption of estradiol regulation of orexin neurons: a novel mechanism in excessive ventilatory response to CO2 inhalation in a female rat model of panic disorder. Transl Psychiatry. (2020) 10:394. doi: 10.1038/s41398-020-01076-x
126. Battaglia M, Rossignol O, Bachand K, D’Amato FR, De Koninck Y. Amiloride modulation of carbon dioxide hypersensitivity and thermal nociceptive hypersensitivity induced by interference with early maternal environment. J Psychopharmacol. (2019) 33:101–8. doi: 10.1177/0269881118784872
127. McMurray KMJ, Strawn JR, Sah R. Fluoxetine modulates spontaneous and conditioned behaviors to carbon dioxide (CO2) inhalation and alters forebrain–midbrain neuronal activation. Neuroscience. (2019) 396:108–18. doi: 10.1016/j.neuroscience.2018.10.042
128. McMurray KMJ, Gray A, Horn P, Sah R. High behavioral sensitivity to carbon dioxide associates with enhanced fear memory and altered forebrain neuronal activation. Neuroscience. (2020) 429:92–105. doi: 10.1016/j.neuroscience.2019.12.009
129. Winter A, Ahlbrand R, Naik D, Sah R. Differential behavioral sensitivity to carbon dioxide (CO2) inhalation in rats. Neuroscience. (2017) 346:423–33. doi: 10.1016/j.neuroscience.2017.01.003
130. Ziemann AE, Allen JE, Dahdaleh NS, Drebot II, Coryell MW, Wunsch AM, et al. The amygdala is a chemosensor that detects carbon dioxide and acidosis to elicit fear behavior. Cell. (2009) 139:1012–21. doi: 10.1016/j.cell.2009.10.029
131. Taugher RJ, Lu Y, Wang Y, Kreple CJ, Ghobbeh A, Fan R, et al. The bed nucleus of the stria terminalis is critical for anxiety-related behavior evoked by CO2 and acidosis. J Neurosci. (2014) 34:10247–55. doi: 10.1523/JNEUROSCI.1680-14.2014
132. Olsson M, Annerbrink K, Bengtsson F, Hedner J, Eriksson E. Paroxetine influences respiration in rats: implications for the treatment of panic disorder. Eur Neuropsychopharmacol. (2004) 14:29–37. doi: 10.1016/S0924-977X(03)00044-0
133. Spiacci A, Vilela-Costa HH, Sant’Ana AB, Fernandes GG, Frias AT, da Silva GSF, et al. Panic-like escape response elicited in mice by exposure to CO2, but not hypoxia. Prog Neuro-Psychopharmacology Biol Psychiatry. (2018) 81:178–86. doi: 10.1016/j.pnpbp.2017.10.018
134. Huckstepp RTR, Dale N. Redefining the components of central CO 2 chemosensitivity - towards a better understanding of mechanism. J Physiol. (2011) 589:5561–79. doi: 10.1113/jphysiol.2011.214759
135. McGuire J, Jp H, Ghosal S, Ke E, Fr S, Sah R. Acid-sensing by the T cell death-associated gene 8 (TDAG8) receptor cloned from rat brain. Biochem Biophys Res Commun. (2009) 386:420–5. doi: 10.1016/j.bbrc.2009.05.133
136. Hanisch U-K, Kettenmann H. Microglia: active sensor and versatile effector cells in the normal and pathologic brain. Nat Neurosci. (2007) 10:1387–94. doi: 10.1038/nn1997
137. Kreutzberg GW. Microglia: a sensor for pathological events in the CNS. Trends Neurosci. (1996) 19:312–8. doi: 10.1016/0166-2236(96)10049-7
138. Nimmerjahn A, Kirchhoff F, Helmchen F. Resting microglial cells are highly dynamic surveillants of brain parenchyma in vivo. Science. (2005) 308:1314–8. doi: 10.1126/science.1110647
139. Rosin JM, Kurrasch DM. Emerging roles for hypothalamic microglia as regulators of physiological homeostasis. Front Neuroendocrinol. (2019) 54:100748. doi: 10.1016/j.yfrne.2019.100748
140. Li Q, Barres BA. Microglia and macrophages in brain homeostasis and disease. Nat Rev Immunol. (2018) 18:225–42. doi: 10.1038/nri.2017.125
141. Johnson AK, Gross PM. Sensory circumventricular organs and brain homeostatic pathways. FASEB J. (1993) 7:678–86. doi: 10.1096/fasebj.7.8.8500693
142. Shekhar A, Keim SR. The circumventricular organs form a potential neural pathway for lactate sensitivity: implications for panic disorder. J Neurosci. (1997) 17:9726–35. doi: 10.1523/JNEUROSCI.17-24-09726.1997
143. Johnson PL, Sajdyk TJ, Fitz SD, Hale MW, Lowry CA, Hay-Schmidt A, et al. Angiotensin II’s role in sodium lactate-induced panic-like responses in rats with repeated urocortin 1 injections into the basolateral amygdala: amygdalar angiotensin receptors and panic. Prog Neuropsychopharmacol Biol Psychiatry. (2013) 44:248–56. doi: 10.1016/j.pnpbp.2013.02.014
144. Shekhar A, Sajdyk TS, Keim SR, Yoder KK, Sanders SK. Role of the basolateral amygdala in panic disorder. Ann N Y Acad Sci. (1999) 877:747–50. doi: 10.1111/j.1749-6632.1999.tb09315.x
145. Winter A, Mcmurray KMJ, Ahlbrand R, Allgire E, Shukla S, Jones J, et al. The subfornical organ regulates acidosis- - evoked fear by engaging microglial acid- - sensor TDAG8 and forebrain neurocircuits in male mice. J Neurosci Res. (2022) 200:1732–46. doi: 10.1002/jnr.25059
146. Cartmell T, Luheshi GN, Rothwell NJ. Brain sites of action of endogenous interleukin-1 in the febrile response to localized inflammation in the rat. J Physiol. (1999) 518:585–94. doi: 10.1111/j.1469-7793.1999.0585p.x
147. McKinley MJ, McAllen RM, Davern P, Giles ME, Penschow J, Sunn N, et al. The sensory circumventricular organs of the mammalian brain. (Vol. 172). Berlin: Springer (2003). doi: 10.1007/978-3-642-55532-9
148. McKinley MJ, Allen AM, Burns P, Colvill LM, Oldfield BJ. Interaction of circulating hormones with the brain: the roles of the subfornical organ and the organum vasculosum of the lamina terminalis. Clin Exp Pharmacol Physiol. (1998) 25:S61–7. doi: 10.1111/j.1440-1681.1998.tb02303.x
149. Smith PM, Ferguson AV. Circulating signals as critical regulators of autonomic state–central roles for the subfornical organ. Am J Physiol Regul Integr Comp Physiol. (2010) 299:R405–15. doi: 10.1152/ajpregu.00103.2010
150. Ferguson AV, Beckmann LM, Fisher JT. Effects of subfornical organ stimulation on respiration in the anesthetized rat. Can J Physiol Pharmacol. (1989) 67:1097–101. doi: 10.1139/y89-173
151. Wei S-G, Zhang Z-H, Beltz TG, Yu Y, Johnson AK, Felder RB. Subfornical organ mediates sympathetic and hemodynamic responses to blood-borne proinflammatory cytokines. Hypertens. (2013) 62:118–25. doi: 10.1161/HYPERTENSIONAHA.113.01404
152. McMurray KMJ, Winter A, Ahlbrand R, Wilson A, Shukla S, Sah R. Subfornical organ interleukin 1 receptor: a novel regulator of spontaneous and conditioned fear associated behaviors in mice. Brain Behav Immun. (2022) 101:304–17. doi: 10.1016/j.bbi.2022.01.004
153. McMurray KM, Winter A, Ahlbrand R, Wilson A, Shukla S, Sah R. Interleukin 1 receptor within the subfornical organ regulates spontaneous and conditioned fear. Brain Behav Immunity. (2021) 98:34. doi: 10.1016/j.bbi.2021.08.131
154. Miselis RR. The efferent projections of the subfornical organ of the rat: a circumventricular organ within a neural network subserving water balance. Brain Res. (1981) 230:1–23. doi: 10.1016/0006-8993(81)90388-7
155. Swanson LW, Lind RW. Neural projections subserving the initiation of a specific motivated behavior in the rat: new projections from the subfornical organ. Brain Res. (1986) 379:399–403. doi: 10.1016/0006-8993(86)90799-7
156. Griez EJ, Colasanti A, van Diest R, Salamon E, Schruers K. Carbon dioxide inhalation induces dose-dependent and age-related negative affectivity. PLoS One. (2007) 2:e987. doi: 10.1371/journal.pone.0000987
157. Gorman JM, Kent J, Martinez J, Browne S, Coplan J, Papp LA. Physiological changes during carbon dioxide inhalation in patients with panic disorder, major depression, and premenstrual dysphoric disorder: evidence for a central fear mechanism. Arch Gen Psychiatry. (2001) 58:125–31. doi: 10.1001/archpsyc.58.2.125
158. Battaglia M, Ogliari A, D’Amato F, Kinkead R. Early-life risk factors for panic and separation anxiety disorder: insights and outstanding questions arising from human and animal studies of CO2 sensitivity. Neurosci Biobehav Rev. (2014) 46:455–64. doi: 10.1016/j.neubiorev.2014.04.005
159. Battaglia M, Marco B. Gene-environment interaction in panic disorder and posttraumatic stress disorder. Can J Psychiatry. (2013) 58:69–75. doi: 10.1177/070674371305800202
160. Battaglia M, Khan WU. Reappraising preclinical models of separation anxiety disorder, panic disorder, and CO2 sensitivity: implications for methodology and translation into new treatments. Curr Top Behav Neurosci. (2018) 40:195–217. doi: 10.1007/7854_2018_42
161. Battaglia M, Pesenti-Gritti P, Medland SE, Ogliari A, Tambs K, Spatola CAM. A genetically informed study of the association between childhood separation anxiety, sensitivity to CO2, panic disorder, and the effect of childhood parental loss. Arch Gen Psychiatry. (2009) 66:64–71. doi: 10.1001/archgenpsychiatry.2008.513
162. Ogliari A, Tambs K, Harris JR, Scaini S, Maffei C, Reichborn-Kjennerud T, et al. The relationships between adverse events, early antecedents, and carbon dioxide reactivity as an intermediate phenotype of panic disorder: a general population study. Psychother Psychosom. (2010) 79:48–55. doi: 10.1159/000259417
163. Cittaro D, Lampis V, Luchetti A, Coccurello R, Guffanti A, Felsani A, et al. Histone Modifications in a mouse model of early adversities and panic disorder: role for Asic1 and neurodevelopmental genes. Sci Rep. (2016) 6:25131. doi: 10.1038/srep25131
164. Luchetti A, Oddi D, Lampis V, Centofante E, Felsani A, Battaglia M, et al. Early handling and repeated cross-fostering have opposite effect on mouse emotionality. Front Behav Neurosci. (2015) 9:93. doi: 10.3389/fnbeh.2015.00093
165. Kinkead R, Gulemetova R. Neonatal maternal separation and neuroendocrine programming of the respiratory control system in rats. Biol Psychol. (2010) 84:26–38. doi: 10.1016/j.biopsycho.2009.09.001
166. Giannese F, Luchetti A, Barbiera G, Lampis V, Zanettini C, Knudsen GP, et al. Conserved DNA methylation signatures in early maternal separation and in twins discordant for CO2 sensitivity. Sci Rep. (2018) 8:1–12. doi: 10.1038/s41598-018-20457-3
167. Genest SE, Gulemetova R, Laforest S, Drolet G, Kinkead R. Neonatal maternal separation induces sex-specific augmentation of the hypercapnic ventilatory response in awake rat. J Appl Physiol. (2007) 102:1416–21. doi: 10.1152/japplphysiol.00454.2006
168. Baldy C, Fournier S, Boisjoly-Villeneuve S, Tremblay MÈ, Kinkead R. The influence of sex and neonatal stress on medullary microglia in rat pups. Exp Physiol. (2018) 103:1192–9. doi: 10.1113/EP087088
169. Marques DA, Gargaglioni LH, Joseph V, Bretzner F, Bícego KC, Fournier S, et al. Impact of ovariectomy and CO2 inhalation on microglia morphology in select brainstem and hypothalamic areas regulating breathing in female rats. Brain Res. (2021) 1756:147276. doi: 10.1016/j.brainres.2021.147276
170. Dutcher EG, Pama EAC, Lynall M-E, Khan S, Clatworthy MR, Robbins TW, et al. Early-life stress and inflammation: a systematic review of a key experimental approach in rodents. Brain Neurosci Adv. (2020) 4:239821282097804. doi: 10.1177/2398212820978049
171. Reid B, Danese A. Challenges in researching the immune pathways between early life adversity and psychopathology. Dev Psychopathol. (2020) 32:1597–624. doi: 10.1017/S0954579420001157
172. Elwenspoek MMC, Kuehn A, Muller CP, Turner JD. The effects of early life adversity on the immune system. Psychoneuroendocrinology. (2017) 82:140–54. doi: 10.1016/j.psyneuen.2017.05.012
173. Webster Marketon JI, Glaser R. Stress hormones and immune function. Cell Immunol. (2008) 252:16–26. doi: 10.1016/j.cellimm.2007.09.006
174. Dantzer R. Neuroimmune interactions: from the brain to the immune system and vice versa. Physiol Rev. (2018) 98:477–504. doi: 10.1152/physrev.00039.2016
175. Wohleb ES, Delpech JC. Dynamic cross-talk between microglia and peripheral monocytes underlies stress-induced neuroinflammation and behavioral consequences. Prog Neuro Psychopharmacol Biol Psychiatry. (2017) 79:40–8. doi: 10.1016/j.pnpbp.2016.04.013
176. Irwin MR, Cole SW. Reciprocal regulation of the neural and innate immune systems. Nat Rev Immunol. (2011) 11:625–32. doi: 10.1038/nri3042
177. Sternberg EM. Neural regulation of innate immunity: a coordinated nonspecific host response to pathogens. Nat Rev Immunol. (2006) 6:318–28. doi: 10.1038/nri1810
178. Dhabhar FS. Stress-induced augmentation of immune function–the role of stress hormones, leukocyte trafficking, and cytokines. Brain Behav Immun. (2002) 16:785–98. doi: 10.1016/S0889-1591(02)00036-3
179. Ramirez K, Fornaguera-Trías J, Sheridan JF. Stress-induced microglia activation and monocyte trafficking to the brain underlie the development of anxiety and depression. Curr Top Behav Neurosci. (2017) 31:155–72. doi: 10.1007/7854_2016_25
180. Weber MD, Godbout JP, Sheridan JF. Repeated social defeat, neuroinflammation, and behavior: monocytes carry the signal. Neuropsychopharmacology. (2017) 42:46–61. doi: 10.1038/npp.2016.102
181. Takahashi A, Flanigan ME, McEwen BS, Russo SJ. Aggression, social stress, and the immune system in humans and animal models. Front Behav Neurosci. (2018) 12:56. doi: 10.3389/fnbeh.2018.00056
182. Lisboa SF, Niraula A, Resstel LB, Guimaraes FS, Godbout JP, Sheridan JF. Repeated social defeat-induced neuroinflammation, anxiety-like behavior and resistance to fear extinction were attenuated by the cannabinoid receptor agonist WIN55,212-2. Neuropsychopharmacology. (2018) 43:1924–33. doi: 10.1038/s41386-018-0064-2
183. Reader BF, Jarrett BL, McKim DB, Wohleb ES, Godbout JP, Sheridan JF. Peripheral and central effects of repeated social defeat stress: monocyte trafficking, microglial activation, and anxiety. Neuroscience. (2015) 289:429–42. doi: 10.1016/j.neuroscience.2015.01.001
184. Wohleb ES, Patterson JM, Sharma V, Quan N, Godbout JP, Sheridan JF. Knockdown of interleukin-1 receptor type-1 on endothelial cells attenuated stress-induced neuroinflammation and prevented anxiety-like behavior. J Neurosci. (2014) 34:2583–91. doi: 10.1523/JNEUROSCI.3723-13.2014
185. DiSabato DJ, Nemeth DP, Liu X, Witcher KG, O’Neil SM, Oliver B, et al. Interleukin-1 receptor on hippocampal neurons drives social withdrawal and cognitive deficits after chronic social stress. Mol Psychiatry. (2021) 26:4770–82. doi: 10.1038/s41380-020-0788-3
186. McKim DB, Weber MD, Niraula A, Sawicki CM, Liu X, Jarrett BL, et al. Microglial recruitment of IL-1β-producing monocytes to brain endothelium causes stress-induced anxiety. Mol Psychiatry. (2018) 23:1421–31. doi: 10.1038/mp.2017.64
187. Wohleb ES, Hanke ML, Corona AW, Powell ND, Stiner LM, Bailey MT, et al. β-Adrenergic receptor antagonism prevents anxiety-like behavior and microglial reactivity induced by repeated social defeat. J Neurosci. (2011) 31:6277–88. doi: 10.1523/JNEUROSCI.0450-11.2011
188. Pfau ML, Menard C, Cathomas F, Desland F, Kana V, Chan KL, et al. Role of monocyte-derived microRNA106b∼25 in resilience to social stress. Biol Psychiatry. (2019) 86:474–82. doi: 10.1016/j.biopsych.2019.02.023
189. Hodes GE, Pfau ML, Leboeuf M, Golden SA, Christoffel DJ, Bregman D, et al. Individual differences in the peripheral immune system promote resilience versus susceptibility to social stress. Proc Natl Acad Sci U.S.A. (2014) 111:16136–41. doi: 10.1073/pnas.1415191111
190. Wang J, Hodes GE, Zhang H, Zhang S, Zhao W, Golden SA, et al. Epigenetic modulation of inflammation and synaptic plasticity promotes resilience against stress in mice. Nat Commun. (2018) 9:477. doi: 10.1038/s41467-017-02794-5
191. Menard C, Pfau ML, Hodes GE, Kana V, Wang VX, Bouchard S, et al. Social stress induces neurovascular pathology promoting depression. Nat Neurosci. (2017) 20:1752–60. doi: 10.1038/s41593-017-0010-3
192. Morais-Silva G, Costa-Ferreira W, Gomes-de-Souza L, Pavan JC, Crestani CC, Marin MT. Cardiovascular outcomes related to social defeat stress: new insights from resilient and susceptible rats. Neurobiol Stress. (2019) 11:100181. doi: 10.1016/j.ynstr.2019.100181
193. Wood S. individual differences in the neurobiology of social stress: implications for depression-cardiovascular disease comorbidity. Curr Neuropharmacol. (2014) 12:205–11. doi: 10.2174/1570159X11666131120224413
194. Brouillard C, Carrive P, Camus F, Bénoliel JJ, Similowski T, Sévoz-Couche C. Long-lasting bradypnea induced by repeated social defeat. Am J Physiol Regul Integr Comp Physiol. (2016) 311:R352–64. doi: 10.1152/ajpregu.00021.2016
195. Finnell JE, Lombard CM, Melson MN, Singh NP, Nagarkatti M, Nagarkatti P, et al. The protective effects of resveratrol on social stress-induced cytokine release and depressive-like behavior. Brain Behav Immun. (2017) 59:147–57. doi: 10.1016/j.bbi.2016.08.019
196. Moshfegh CM, Elkhatib SK, Collins CW, Kohl AJ, Case AJ. Autonomic and redox imbalance correlates with T-Lymphocyte inflammation in a model of chronic social defeat stress. Front Behav Neurosci. (2019) 13:103. doi: 10.3389/fnbeh.2019.00103
197. Lissek S. Toward an account of clinical anxiety predicated on basic, neurally mapped mechanisms of pavlovian fear-learning: the case for conditioned overgeneralization. Depress Anxiety. (2012) 29:257–63. doi: 10.1002/da.21922
198. Deslauriers J, Toth M, Der-Avakian A, Risbrough VB. Current status of animal models of posttraumatic stress disorder: behavioral and biological phenotypes, and future challenges in improving translation. Biol Psychiatry. (2018) 83:895–907. doi: 10.1016/j.biopsych.2017.11.019
199. Careaga MBL, Girardi CEN, Suchecki D. Understanding posttraumatic stress disorder through fear conditioning, extinction and reconsolidation. Neurosci Biobehav Rev. (2016) 71:48–57. doi: 10.1016/j.neubiorev.2016.08.023
200. Maren S, Holmes A. Stress and fear extinction. Neuropsychopharmacology. (2016) 41:58–79. doi: 10.1038/npp.2015.180
201. Singewald N, Hauschild M, Sartori SB, Muigg P, Landgraf R, Hefner K, et al. Rodent models of impaired fear extinction: therapeutic approaches. Eur Neuropsychopharmacol. (2018) 21:S224. doi: 10.1016/S0924-977X(11)70338-8
202. Hao Y, Jing H, Bi Q, Zhang J, Qin L, Yang P. Intra-amygdala microinfusion of IL-6 impairs the auditory fear conditioning of rats via JAK/STAT activation. Behav Brain Res. (2014) 275:88–95. doi: 10.1016/j.bbr.2014.08.052
203. Yu Z, Fukushima H, Ono C, Sakai M, Kasahara Y, Kikuchi Y, et al. Microglial production of TNF-alpha is a key element of sustained fear memory. Brain Behav Immun. (2017) 59:313–21. doi: 10.1016/j.bbi.2016.08.011
204. Dong Y, Li S, Lu Y, Li X, Liao Y, Peng Z, et al. Stress-induced NLRP3 inflammasome activation negatively regulates fear memory in mice. J Neuroinflammation. (2020) 17:1–16. doi: 10.1186/s12974-020-01842-0
205. Clark SM, Soroka JA, Song C, Li X, Tonelli LH. CD4 + T cells confer anxiolytic and antidepressant-like effects, but enhance fear memory processes in Rag2 -/- mice. Stress. (2016) 19:303–11. doi: 10.1080/10253890.2016.1191466
206. Conoscenti MA, Fanselow MS. Dissociation in effective treatment and behavioral phenotype between stress-enhanced fear learning and learned helplessness. Front Behav Neurosci. (2019) 13:104. doi: 10.3389/fnbeh.2019.00104
207. Rau V, Fanselow MS. Exposure to a stressor produces a long lasting enhancement of fear learning in rats. Stress. (2009) 12:125–33. doi: 10.1080/10253890802137320
208. Jones ME, Lebonville CL, Barrus D, Lysle DT. The role of brain interleukin-1 in stress-enhanced fear learning. Neuropsychopharmacology. (2015) 40:1289–96. doi: 10.1038/npp.2014.317
209. Jones ME, Lebonville CL, Paniccia JE, Balentine ME, Reissner KJ, Lysle DT. Hippocampal interleukin-1 mediates stress-enhanced fear learning: a potential role for astrocyte-derived interleukin-1β. Brain Behav Immun. (2018) 67:355–63. doi: 10.1016/j.bbi.2017.09.016
210. Jones ME, Paniccia JE, Lebonville CL, Reissner KJ, Lysle DT. chemogenetic manipulation of dorsal hippocampal astrocytes protects against the development of stress-enhanced fear learning. Neuroscience. (2018) 388:45–56. doi: 10.1016/j.neuroscience.2018.07.015
211. Dion-Albert L, Cadoret A, Doney E, Kaufmann FN, Dudek KA, Daigle B, et al. Vascular and blood-brain barrier-related changes underlie stress responses and resilience in female mice and depression in human tissue. Nat Commun. (2022) 13:1–18.
212. Wang X, Ma L, Luo Y, Yang Y, Upreti B, Cheng Y, et al. Increasing of blood brain barrier permeability and the association with depression and anxiety in systemic lupus erythematosus patients. Front Med. (2022) 9:852835. doi: 10.3389/fmed.2022.852835
213. Mah L, Szabuniewicz C, Fiocco AJ. Can anxiety damage the brain? Curr Opin Psychiatry. (2016) 29:56–63. doi: 10.1097/YCO.0000000000000223
214. Zoccal DB, Furuya WI, Bassi M, Colombari DSA, Colombari E. The nucleus of the solitary tract and the coordination of respiratory and sympathetic activities. Front Physiol. (2014) 5:238. doi: 10.3389/fphys.2014.00238
215. Bi Q, Shi L, Yang P, Wang J, Qin L. Minocycline attenuates interferon-α-induced impairments in rat fear extinction. J Neuroinflammation. (2016) 13:1–12. doi: 10.1186/s12974-016-0638-z
216. Amorim MR, De Deus JL, Cazuza RA, Mota CMD, Da Silva LEV, Borges GS, et al. Neuroinflammation in the NTS is associated with changes in cardiovascular reflexes during systemic inflammation. J Neuroinflammation. (2019) 16:1–15. doi: 10.1186/s12974-019-1512-6
217. Morrison FG, Miller MW, Wolf EJ, Logue MW, Maniates H, Kwasnik D, et al. Reduced interleukin 1A gene expression in the dorsolateral prefrontal cortex of individuals with PTSD and depression. Neurosci Lett. (2019) 692:204–9. doi: 10.1016/j.neulet.2018.10.027
218. Salim S. Oxidative stress: a potential link between emotional wellbeing and immune response. Curr Opin Pharmacol. (2016) 29:70–6. doi: 10.1016/j.coph.2016.06.006
219. Kim Y-K, Jeon SW. Neuroinflammation and the immune-kynurenine pathway in anxiety disorders. Curr Neuropharmacol. (2018) 16:574–82. doi: 10.2174/1570159X15666170913110426
220. Viveros M, Arranz L, Hernanz A, Miquel J, De la Fuente M. A model of premature aging in mice based on altered stress-related behavioral response and immunosenescenceitle. Neuroimmunomodulation. (2007) 14:157–62. doi: 10.1159/000110640
221. Frank MG, Weber MD, Watkins LR, Maier SF. Stress-induced neuroinflammatory priming: a liability factor in the etiology of psychiatric disorders. Neurobiol Stress. (2016) 4:62–70. doi: 10.1016/j.ynstr.2015.12.004
222. Algin DÝ, Kilicoglu A, Koplay M. Panic disorder as presenting symptoms of multiple sclerosis. Ann Gen Psychiatry. (2010) 9:2010. doi: 10.1186/1744-859X-9-S1-S138
223. Bachen EA, Chesney MA, Criswell LA. Prevalence of mood and anxiety disorders in women with systemic lupus erythematosus. Arthritis Care Res. (2009) 61:822–9. doi: 10.1002/art.24519
224. Alonso J, De Jonge P, Lim CCW, Aguilar-gaxiola S, Caldas-de-almeida JM, Liu Z, et al. Association between mental disorders and subsequent adult onset asthma. J Psychiatr Res. (2014) 56:179–88. doi: 10.1016/j.jpsychires.2014.09.007
225. Maunder RG. Panic disorder associated with gastrointestinal disease: review and hypotheses. J Psychosom Res. (1998) 44:91–105. doi: 10.1016/S0022-3999(97)00133-5
226. Roy-Byrne PP, Stang P, Wittchen HU, Ustun B, Walters EE, Kessler RC. Lifetime panic-depression comorbidity in the national comorbidity survey: association with symptoms, impairment, course and help-seeking. Br J Psychiatry. (2000) 176:229–35. doi: 10.1192/bjp.176.3.229
227. Cougle JR, Feldner MT, Keough ME, Hawkins KA, Fitch KE. Comorbid panic attacks among individuals with posttraumatic stress disorder: associations with traumatic event exposure history, symptoms, and impairment. J Anxiety Disord. (2010) 24:183–8. doi: 10.1016/j.janxdis.2009.10.006
228. Avitsur R, Hunzeker J, Sheridan JF. Role of early stress in the individual differences in host response to viral infection. Brain Behav Immun. (2006) 20:339–48.
229. Roque S, Mesquita AR, Palha JA, Sousa N, Correia-Neves M. The behavioral and immunological impact of maternal separation: a matter of timing. Front Behav Neurosci. (2014) 8:1–10.
230. Lubach GR, Coe CL, Ershler WB. Effects of early rearing environment on immune-responses of infant rhesus monkeys. Brain Behav Immun. (1995) 9:31–46.
231. Bakker J, Kavelaars A, Kamphuis P, Cobelens P, van Vugt H, van Bel F, et al. Neonatal dexamethasone treatment increases susceptibility to experimental autoimmune disease in adult rats. J Immunol. (2000) 165:5932–7.
232. Bakker JM, Kavelaars A, Kamphuis PJGH, Heijnen CJ, Van Bel F. Effects of early neonatal dexamethasone treatment on susceptibility to experimental autoimmune encephalomyelitis (EAE) and on the LPS-induced corticosterone response in adult rats. Oediatric Res. (1999) 45:48.
233. Daft JG, Ptacek T, Kumar R, Morrow C, Lorenz RG. Cross-fostering immediately after birth induces a permanent microbiota shift that is shaped by the nursing mother. Microbiome. (2015) 3:1–10.
234. Bartolomucci A, Gioiosa L, Chirieleison A, Ceresini G, Parmigiani S, Palanza P. Cross fostering in mice: behavioral and physiological carry-over effects in adulthood. Genes Brain Behav. (2004) 3:115–22.
235. Maslanik T, Mahaffey L, Tannura K, Beninson L, Greenwood B, Fleshner M. The inflammasome and danger associated molecular patterns (DAMPs) are implicated in cytokine and chemokine responses following stressor exposure. Brain Behav Immun. (2013) 28:54–62.
Keywords: panic, fear, neuroimmune, panic disorder, interoception, body-to-brain
Citation: McMurray KMJ and Sah R (2022) Neuroimmune mechanisms in fear and panic pathophysiology. Front. Psychiatry 13:1015349. doi: 10.3389/fpsyt.2022.1015349
Received: 09 August 2022; Accepted: 02 November 2022;
Published: 29 November 2022.
Edited by:
Joaquim Radua, Institut d’Investigacions Biomèdiques August Pi i Sunyer (IDIBAPS), SpainReviewed by:
Manfred Schedlowski, University Hospital Essen, GermanyCopyright © 2022 McMurray and Sah. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Katherine M. J. McMurray, bWNtdXJya21AdWNtYWlsLnVjLmVkdQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.