 Zhi-Peng Guo1
Zhi-Peng Guo1 Peter Sörös
Peter Sörös Chun-Hong Liu
Chun-Hong Liu
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Psychiatry , 15 December 2021
Sec. Neuroimaging
Volume 12 - 2021 | https://doi.org/10.3389/fpsyt.2021.765106
This article is part of the Research Topic Insights in Neuroimaging and Stimulation: 2021 View all 7 articles
The coronavirus disease 2019 (COVID-19) comprises more than just severe acute respiratory syndrome. It also interacts with the cardiovascular, nervous, renal, and immune systems at multiple levels, increasing morbidity in patients with underlying cardiometabolic conditions and inducing myocardial injury or dysfunction. Transcutaneous auricular vagus nerve stimulation (taVNS), which is derived from auricular acupuncture, has become a popular therapy that is increasingly accessible to the general public in modern China. Here, we begin by outlining the historical background of taVNS, and then describe important links between dysfunction in proinflammatory cytokine release and related multiorgan damage in COVID-19. Furthermore, we emphasize the important relationships between proinflammatory cytokines and depressive symptoms. Finally, we discuss how taVNS improves immune function via the cholinergic anti-inflammatory pathway and modulates brain circuits via the hypothalamic–pituitary–adrenal axis, making taVNS an important treatment for depressive symptoms on post-COVID-19 sequelae. Our review suggests that the link between anti-inflammatory processes and brain circuits could be a potential target for treating COVID-19-related multiorgan damage, as well as depressive symptoms using taVNS.
In December 2019, a novel coronavirus disease (COVID-19) outbreak emerged from Wuhan, Hubei Province, China, initiating a global health threat and posing a challenge to the psychological resilience of populations worldwide (1). Clinically, presentation of COVID-19 varies from being asymptomatic, to including mild symptoms such as fever, sore throat, headache, fatigue, to manifesting as severe acute respiratory distress syndrome (ARDS) (2). Moreover, it also interacts with the cardiovascular, nervous, renal, and immune systems at multiple levels (3). An extreme immune reaction resulting in elevated levels of inflammatory cytokines, often referred to as a cytokine storm, has been linked to an increased number of deaths from COVID-19 (4, 5). However, even worse than this, the COVID-19 pandemic has also led to an increased prevalence of mental health problems, such as difficulty sleeping, depression and anxiety, and hypomania (6). Although a number of vaccines have been proved to be effective (7, 8), evidence-based evaluations and interventions targeting mental health disorders are relatively scarce (9). Transcutaneous auricular vagus nerve stimulation (taVNS) is being explored as an adjuvant therapy to the depressive symptoms of COVID-19 during the pandemic to deal with these disorders.
The concept of taVNS as a therapy has emerged relatively recently. The technique makes use of the analgesic effects of the neuronal network that innervates the vagus nerve (10), which targets the cutaneous receptive field of the auricular branch of the vagus nerve at the outer ear (11). Promising results indicate that, following taVNS treatment, the symptoms of mood disorders can be alleviated painlessly and without the need for surgery (12). Ventureyra was the first to propose applying vagus nerve stimulation (VNS) using surgically implanted electrodes wrapped around the vagus nerve in the neck (10). In 2005, VNS was approved as a long-term adjunctive treatment for patients with refractory depression of more than 18 years of age (13, 14). From a neuroanatomical point of view, vagus nerve fibers project to the nucleus tractus solitarius (NTS) and the locus coeruleus (LC), where they form direct and indirect ascending projections to many brain regions, including the midbrain, hypothalamus, amygdala, hippocampus, and frontal lobe (15). The vagus nerve, which is the longest nerve in the body, connects the central nervous system to the body by innervating major visceral organs such as the liver, spleen, and gastrointestinal tract (16). Once an inflammatory response has been detected, taVNS may help to attenuate inflammatory responses via the cholinergic anti-inflammatory pathway and by modulating brain circuits via the hypothalamic–pituitary–adrenal (HPA) axis (3, 17). Acute respiratory distress syndrome (ARDS) or fulminant pneumonia can lead to widespread inflammation and very high concentrations of cytokines in the lungs, accompanied by activation of the anti-inflammatory pathways mentioned above (18). To date, clinical and laboratory research demonstrated that taVNS can improve lung function (19, 20). In addition, taVNS is commonly used to treat encephalopathy, encephalitis, ischemic infarcts, cerebral venous thrombosis, as well as peripheral nervous system pathologies [i.e., muscle injuries, and peripheral neuropathies; (21–26)].
In order to better understand the mechanisms underlying taVNS, we review the literature on proinflammatory cytokines and the brain imaging correlates of taVNS. To date, there have not been any reviews that considered in detail how taVNS might treat depressive symptoms, which develops from COVID-19, or its associated co-morbidities. We provide an integrated account of how the dysregulation of inflammatory and immunological responses affect brain circuits in COVID-19.
Auricular acupuncture originated in China during the Chou period (first millennium BCE) and has recently attracted scientific and public attention as it becomes increasingly accessible to the general public in modern China (Figure 1A) (28). The practice of auricular acupuncture is referenced in the Huangdi Neijing (The Yellow Emperor's Classics of Internal Medicine), which describes how the ear is not isolated but rather is directly or indirectly connected with 12 meridians (six yang and six yin) (29). In the 1950s, Dr. Paul Nogier, a French neurologist, proposed that the outer ear represents “an inverted fetus map” (Figure 1B) (30)]. In 1990, the World Health Organization (WHO) recognized auricular acupuncture as a self-contained microacupuncture system that maps all portions of the ear to specific parts of the body and to the internal organs (31). Having considered the anatomy of the neural pathways in the external auricle, Usichenko et al. proposed that the analgesic effects of auricular acupuncture could be explained by stimulation of the auricular branch of the vagus nerve (32). The vagus is known to be a mixed nerve, with about 80% of its fibers carrying sensory afferent information to the brain and about 20% carrying efferent motor information to the liver, spleen, and gastrointestinal tract (33). Thus, it is very likely that taVNS functions based on the Chinese system of energy circulation along the meridians, which connect “diseased” body organs with the external auricle. In addition to Asian countries, in which this technique is widely available and easy to apply, it may be possible to use taVNS to effectively respond to the COVID-19 pandemic-related depressive symptoms as well as multiorgan damage in environments where medical resources are limited.
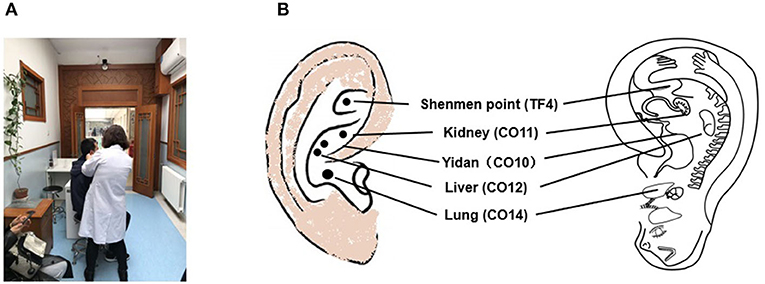
Figure 1. (A) Auricular acupuncture practice has recently been attracting the attention of the public in China and is commonly carried out within the Chinese medical hospital system. (B) TF4 and CO10-12 are used to stimulate the auricular branch of the vagus nerve, with the outer ear corresponding to an “inverted fetus map” [according to King and Hickey, 2013 (27)].
Several studies have suggested that the pathogenesis of COVID-19 involves an inability to resolve the inflammatory response along with the activation of immune cells and inflammatory cytokines (18, 34). In COVID-19 patients, an unregulated inflammatory response to the infection can result in the dysregulation of T cells with associated lymphopenia, high levels of the proinflammatory cytokines interleukin (IL)-6 and tumor necrosis factor (TNF)-α, and high levels of inflammatory chemokines, including C-C motif chemokine ligand (CCL-2) (35). In a study by Staats et al., 49-year-old man with excessive fatigue, mental cloudiness and body aches, and mental cloudiness had ceased after 5 days non-invasive VNS therapy (19). Furthermore, the authors also summarized five studies that used taVNS to treat COVID-19 and reported that the majority of patients obtain relief from respiratory distress after taVNS therapy. Three review papers have hypothesized that the cytokine storm and the worsening of patient health can be ameliorated or even prevented by taVNS (3, 18, 36). Therefore, targeting the inflammatory response and immune cells using taVNS might be a promising line of research in the fight against COVID-19-related inflammatory cytokine-induced multiorgan damage.
Current research indicates that COVID-19 might involve multiple organs including those in the central and peripheral nervous systems, rather than being restricted to the respiratory system (37). Recently, it has been noted that COVID-19 patients experience a number of different neurological symptoms, such as headache, dizziness, hyposmia, and hypogeusia during the course of the illness (38). Psychiatric symptoms, including post-traumatic stress disorder (PTSD), anxiety, and depressive symptoms, have also been reported in patients with COVID-19 (39, 40). Even worse than this, Kremer et al. found signal abnormalities in the medial temporal lobe and non-confluent multifocal white matter hyperintense lesions (41). Post-mortem brain imaging has demonstrated subcortical hemorrhagic and cortico-subcortical edematous changes, as well as olfactory impairment in patients who died of COVID-19 (42). Based on the results of published studies, COVID-19 encephalopathy appears to be more common in cases comorbid for encephalopathy, encephalitis, acute disseminated encephalomyelitis, myelitis, meningitis, ischemic infarcts, or cerebral venous thrombosis (43). In the peripheral nervous system, COVID-19 has been associated with dysfunction in the sense of smell and taste, and with muscle injury (41). Of note, the etiology of the encephalopathy in COVID-19 mentioned above is mostly linked to injury of the central and peripheral nervous systems by a cytokine storm, blood clots, or direct damage to specific receptors (41, 44). The pathogen that causes COVID-19, severe acute respiratory syndrome coronavirus 2, can invade the brain via vascular, peripheral nervous, lymphatic, cerebrospinal fluid pathways (45).
Several studies have suggested that inflammation or immune dysregulation are implicated in the pathophysiology of depression (46–51). It is now well-established that both the innate and adaptive immune systems become dysregulated in depressed patients and that controlling inflammation might be of therapeutic benefit (52). Two meta-analyses showed reliably higher levels of inflammatory markers in depression, namely IL-1β, IL-6, C-reactive protein (CRP), and TNF-α (53, 54). Plasma CRP in depression was not only positively associated with plasma levels of inflammatory cytokines (e.g., IL-6, TNF-α, sTNFR2, and IL-1ra), but also correlated with the level of CRP in cerebrospinal fluid (55). Both Alexopoulos et al. and Galecki et al. reported continual interactions between changes in the peripheral immune response and central immune activation [e.g., macrophage accumulation and microglial activation; (56, 57)]. These central and peripheral immune changes lead to increased production of proinflammatory cytokines (58, 59), which in turn lead to abnormalities in brain circuits. To some extent, this permits the relationship between abnormalities in brain circuits and inflammatory states in depression to be inferred. Hao et al. demonstrated that psychiatric patients were significantly higher in their levels of worry, anger, impulsivity, and intense suicidal ideation than healthy controls during the peak of the COVID-19 epidemic (60). Based on the psychological impact of the COVID-19 pandemic on psychiatric patients, targeting the cholinergic anti-inflammatory pathway and modulating brain circuits using taVNS is a rational approach to treating COVID-19 and its associated cytokine storm. Controlling inflammation might provide an overall therapeutic benefit, regardless of whether it is secondary to early life trauma, a more acute stress response, microbiome alterations, a genetic diathesis, or a combination of these and other factors.
Dysfunction of the cortico-limbic-striatal neural system, including cortical (anterior cingulate and prefrontal cortex) and limbic (amygdala, hippocampus, parahippocampal gyrus, cingulate gyrus, nucleus accumbens, and striatum) areas has been implicated in depression (61–63). Mayberg found dorsal and lateral cortical hypoactivity and ventral limbic hyperactivity in depression using positron emission tomography (64). Taylor and Liberzon also proposed a hypo-dorsal and -lateral cortical model of cognitive processes and a hyper-limbic model of emotional expression to account for the experience of depression (65). Using tasks requiring executive control and emotional information processing, Siegle et al. identified sustained increased amygdala activity in response to emotional information processing and decreased dorsal prefrontal cortex activity in response to executive cognitive tasks (66). Using a meta-analytic technique, Fitzgerald et al. identified two neural systems implicated in emotional regulation in depression, including reduced activity in dorsolateral prefrontal cortex and more dorsal regions of the anterior cingulate cortex (67). Furthermore, they found increased activity in medial prefrontal cortex and in subcortical regions related to emotional processing in the depressed state. All of these changes returned to normal after antidepressant treatment. Together, these studies imply that patients with depression may exhibit impairments in their cognitive control network, as evidenced by their inability to disengage from negative stimuli (68). In addition, they show impairments in their affective control network, as evidenced by the hyperactivity of their amygdala and hippocampus to negative stimuli and recall.
Discrepancies in stimulation locations exist among studies that stimulated the auricular branch of the vagus nerve (69). The location is often dictated by the geometry of an electrode, with clip electrodes typically attached to the tragus or cymba concha (70–73). The outer auditory canal is also reported as a stimulation site, without further clarification for the electrode location (74–76). Based on Peuker and Filler's anatomical studies, the auricular branch of the vagus nerve innervates the tragus, concha, and cymba concha (77). However, it is difficult to select an optimal stimulation site for any particular disorder. The taVNS devices are relatively inexpensive, small, and mobile, which will be performed at patient's home after training (78).
As taVNS is a novel treatment, there is currently no consensus on the appropriate stimulation parameters for its therapeutic use. According to the latest published International Consensus on taVNS (79), the points stimulated by taVNS are located in the auricular concha region, which contains a rich distribution of vagus nerve branches. Stimulation parameters used in taVNS studies have included: (1) a 20-Hz continuous sinusoidal wave (wave width, 0.2 ms) (80, 81); a 10-Hz continuous sinusoidal wave (73); a 20–30 Hz continuous sinusoidal wave (82, 83); a 4/20 Hz dense wave (between 0.8 and 1.5 mA) (84); a 20 Hz dense wave (between 4 and 6 mA) (72, 85); 1.5 Hz unipolar rectangular waves (0–600 mA) (69); a 120 Hz pulse wave (12 mA) (86); a 25 Hz monophasic rectangular waves (87); and (2) a gradually increasing stimulation intensity, starting from zero up to the highest point that the patients could tolerate (typically between 4 and 6 mA) (12). In terms of the safety of taVNS, a systematic review by Redgrave et al. reported the side effects of taVNS as local skin irritation, headache, nasopharyngitis, and a number of potentially serious adverse events [e.g., palpitations; (88)]. Indeed, the vagus nerve projects to the parabrachial nucleus, which can regulate heart rate, with one study showing that taVNS can cause side effects on heart rate when specific stimulation parameters (pulse width, 500 μs; frequency, 25 Hz) are used (89). However, in most cases, side effects were not apparent or disappeared after follow up (86, 90, 91).
VNS has greater effects in females in animal studies, probably because of the effect of estrogens on muscarinic acetylcholine receptors in the central nervous system (92). Similar effects would be expected in females human subjects due to both hormonal levels and the gender-dependent differences in the functions of the autonomic nervous system (93, 94). Age is associated with marked changes at the hormonal level, which in turn affect acetylcholine-mediated parasympathetic autonomic activity (95, 96). Fallgatter et al. reported that the vagus sensory-evoked potentials showed a trend toward reduction in the elderly, associated with age-related demyelination of neuronal structures or degenerative processes (97). In addition, sensitivity to electrical transcutaneous stimulation was found to be lower in the elderly (98).
There is now accumulating evidence that different forms of proinflammatory cytokine-mediated communication between the immune system and brain circuits modulate the inflammatory pathway in the brain (99–101). Rodent and human neuroimaging studies combined with experimental inflammatory challenges have been successful in clarifying the sensitivity of the insula and striatum to changes in peripheral inflammation in depression (102). Of note, neuroinflammation is associated with structural and functional anomalies in depression (103). A negative correlation was found between CRP levels and the cortical thickness of the right medial prefrontal cortex (mPFC) in depression (104). In a recent resting-state functional magnetic resonance imaging (fMRI) study, CRP level was negatively correlated with amygdala–ventromedial prefrontal cortex (vmPFC) connectivity in depressed patients with high levels of inflammation and symptoms of anxiety (105). Haroon et al. demonstrated that plasma CRP levels are significantly associated with glutamate levels in the left basal ganglia using magnetic resonance spectroscopy (MRS) (106), and increased glutamate in the left basal ganglia in turn correlated with anhedonia and psychomotor slowing. Haroon et al. further pointed out that patients with high levels of both inflammation and basal ganglia glutamate showed decreased local homogeneity in vmPFC, and in dorsal and ventral striatal regions (107). In their study of medically stable patients with depression, Felger et al. reported that levels of CRP, as well as those of IL-6, IL-1beta, and IL-1ra, were negatively associated with connectivity between ventral striatum and vmPFC, and that this decreased connectivity in turn correlated with increased anhedonia (108). Moreover, the level of CRP was negatively correlated with connectivity between dorsal striatum, vmPFC, and presupplementary motor area. Decreased connectivity between dorsal striatum, vmPFC, and presupplementary motor area were further correlated with motor speed and psychomotor slowing. More recently developed methods, such as large-scale network-based analyses, were used by Yin et al. to show that the increased level of CRP is associated with reduced connectivity in ventral striatum, amygdala, orbitofrontal and insular cortices, and posterior cingulate cortex (109). Using surface-based morphometry, Kakeda et al. demonstrated that cortical thicknesses, such as those of the superior frontal and medial orbitofrontal cortex, showed a significant inverse correlation with the level of IL-6 (110). Using automated cortical parcellation within the mPFC including Brodmann areas (BA) 9, 10, 11, 24, 25, and 32, Meier et al. found an inverse relationship between plasma CRP level and the thickness of BA32, with recurrent MDD patients having a thinner cortex in BA32 (104). Using voxel-based morphometry, Chen et al. found that orbitofrontal cortex, lingual gyrus, inferior frontal cortex, middle frontal cortex, and planum polare were negatively correlated with levels of IL-6 (111). Moreover, Frodl et al. reported an inverse relationship of IL-6 concentration and hippocampal volume in MDD (112). Doolin et al. provided additional evidence to support a negative association between CRP levels and hippocampal subfield volumes (113). Importantly, the striatum, vmPFC, and presupplementary motor area are part of the classical reward and motor circuitry that receives neurotransmitters such as glutamate, in addition to dopaminergic innervation (114–116). Furthermore, Nusslock et al. found that higher levels of inflammatory biomarkers (e.g., CRP, IL-6, IL-10, and TNF-α) were associated with lower connectivities within both the emotional network and the central executive network in urban African American youths, suggesting that inflammation or neuroimmunology may be involved in the pathogenesis of emotional and physical health problems (117). More importantly, Cosgrove et al. reported that higher levels of CRP were related to greater coupling of orbitofrontal cortical and anterior insular activity with increased appetite in depressed patients (118). Together, these studies imply that systemic low-grade inflammation is associated with the coupling of activity in striatum with that in reward- and interoceptive-related neural circuitry, and provide evidence for physiological subtypes within depression.
Macrophages, proinflammatory cytokines (such as interleukin (IL)-1β, IL-6, and tumor necrosis factor (TNF)-α) and chemokines released by respiratory epithelial and dendritic cells, are all known to play a role in the pathogenesis of critical patients with COVID-19 (119). Consequently, Bonaz et al. hypothesized that targeting the cholinergic anti-inflammatory pathway by vagus nerve stimulation could be a useful therapeutic option for patients with COVID-19. In support of this hypothesis, Staats et al. recently reported two patients with respiratory symptoms that were similar to those associated with COVID-19 who showed marked clinical benefit following the application of transcutaneous cervical vagus nerve stimulation (19). Research has also shown that the levels of proinflammatory cytokines, including IL-6, IL-10, IL-12, IL-13, and TNF-α, are elevated in MDD when compared to those of healthy controls (120). However, there is still a clear shortage of evidence supporting the neuroimaging findings of taVNS in the treatment of depressive symptoms in patients with COVID-19. Our previous review has validated taVNS may inhibit both peripheral and central inflammation and modulate multiple neural systems (121). Studies have demonstrated that taVNS increases connectivity of the nucleus accumbens (NAc) with bilateral mPFC/rostral anterior cingulate cortex (rACC); NAc with insula, occipital gyrus, and lingual/fusiform gyrus; amygdala with dorsolateral prefrontal cortex; and the default mode network (DMN) with precuneus and orbital prefrontal cortex. In addition, studies have reported decreased connectivity of medial hypothalamus (MH) with rACC, and DMN with anterior insula and parahippocampus (72, 85, 122, 123). Therefore, we argued that it was advantageous for treating the inflammatory processes associated with COVID-19 and modulate brain activity in the NAc, hypothalamus, DMN, amygdala, and rACC via the auricular branch of the vagus nerve (78). Further, it has been suggested that taVNS can attenuate inflammation by targeting the HPA axis (16).
Finally, since the beginning of the COVID-19 pandemic, various manifold neuroimaging features have been described for patients with COVID-19 and a range of interesting and helpful findings have been described across the globe (124). For example, Jain et al. found that acute stroke was the most common finding on neuroimaging; 92.5% of patients with positive neuroimaging studies also showed evidence of acute stroke on neuroimaging. Acute stroke is therefore a strong prognostic marker for a poor outcome (125). In another study, Mao et al. reported that 36.4% of patients had headache, dizziness, impaired consciousness, acute cerebrovascular disease, ataxia, and seizures, and that 8.9% of patients experienced specific manifestations in their senses, including taste, smell, vision impairment, and nerve pain (126). Furthermore, Brouwer et al. reported that acute cerebrovascular events were also detected in ~3% of patients and that 6% of patients with severe manifestations had cerebrovascular events (127). Similarly, Tsai et al. reported a wide range of neurological manifestations, including olfactory taste disorders, headache, acute cerebral vascular disease, dizziness, altered mental status, seizure, encephalitis, neuralgia, ataxia, Guillain-Barre syndrome, Miller Fisher syndrome, intracerebral hemorrhage, polyneuritis, and dystonic posture (128). In addition, Al-Olama et al. reported that COVID-19 infection can cause meningoencephalitis in right frontal intracerebral hematomas, subarachnoid hemorrhage, and in frontal and temporal lobe thin subdural hematomas (129). Therefore, obtaining detailed neurological examinations and neuroimaging for the early and accurate diagnosis of these often fatal neurological complications could significantly improve our understanding of COVID-19 and its neurological manifestations.
The cholinergic anti-inflammatory pathway via the vagus nerve has been proposed to be a key mediator of cross-communication between the peripheral immune system and the brain (130). Indeed, an increase of TNF-α in the liver and blood induced by an extreme immune reaction or cytokine storm was successfully dampened by stimulation of the vagus nerve, inducing an anti-inflammatory effect involving the release of acetylcholine (ACh) (131). Promisingly, Staats et al. reported clinically meaningful benefits of VNS in two COVID-19 patients with severe acute respiratory syndrome (19). The vagus nerve has a dual anti-inflammatory role, with 80% of the afferents targeting the cholinergic anti-inflammatory pathway and 20% of efferent fibers targeting the HPA axis (132). The efferent fibers of the vagus nerve activate the HPA axis, causing glucocorticoid release from the adrenal glands (133). Efferent fibers also run through the neck, connecting the brainstem to many organs, including the spleen, where they inhibit the release of TNF-α (16). Targeting the vagus nerve non-invasively may open up novel adjuvant approaches to treating COVID-19 patients. The various mechanisms by which taVNS may treat inflammation and related organ dysfunction in COVID-19 are illustrated in Figure 2.
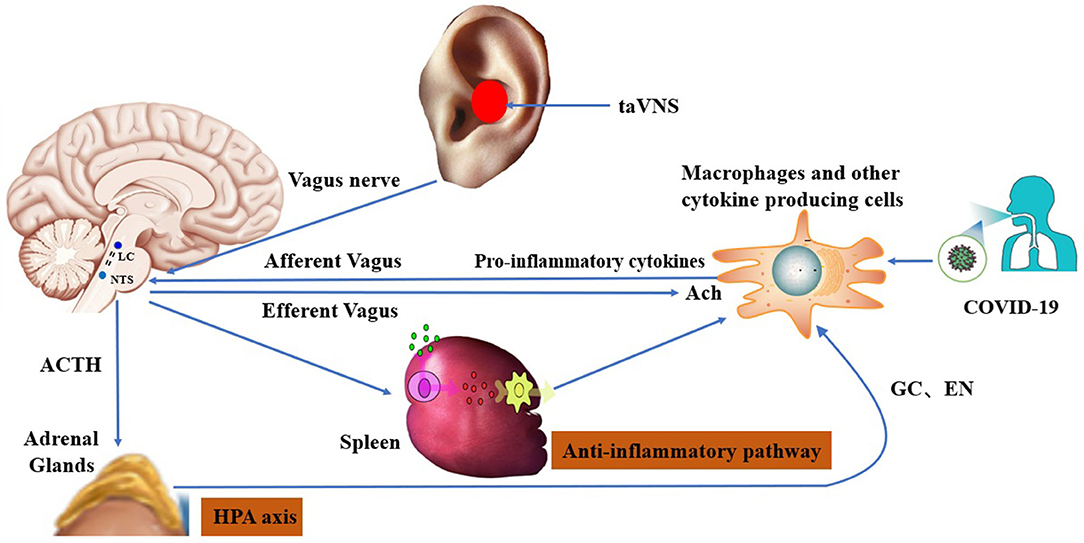
Figure 2. Hypothesized mechanisms of transcutaneous auricular vagus nerve stimulation in the treatment of post-COVID-19 sequelae: 1) improvement in immune function via the cholinergic anti-inflammatory pathway; and 2) modulation of brain circuits via the HPA axis [according to Bonaz and Sinniger, 2016 (134)].
TCM has a history of more than 2,000 years in the prevention and treatment of epidemics and plagues and the national health commission of China has recommended some patent Chinese medicine, such as Jinhua Qinggan granules, Lianhua Qingwen capsules, Xuebijing injections, a Qingfei Paidu decoction, a Huashi Baidu decoction, and a Xuanfei Baidu decoction (135). Patients with COVID-19 who took Jinhua Qinggan granules recovered faster than those who did not take the granules (136). Therapeutic efficacy was significantly higher in patients with COVID-19 taking Lianhua Qingwen capsules and Arbidol (umifenovir) than that in those taking Arbidol alone; moreover, the conversion rate to severe disease in patients taking these capsules was significantly lower than that in patients taking Arbidol alone (137). Furthermore, chest computed tomography images of patients with COVID-19 showed improvement after 6 days of treatment with Qingfei Paidu decoction (138). In addition, other therapies such as acupuncture might also play a beneficial role in treating breathlessness after COVID-19 (4). Thus, TCM could play an important role in fighting COVID-19 in China.
This review has provided a comprehensive evaluation of targets for taVNS that can be used to treat inflammation and related organ dysfunction in COVID-19. It is clear that COVID-19 involves interrelationships between proinflammatory cytokines and brain circuits. The research findings detailed here suggest that taVNS could be used as an adjuvant therapy for depressive symptoms during the COVID-19 pandemic. We present a rationale for targeting the anti-inflammatory process and modulating brain circuits to treat COVID-19 and its associated cytokine storm. The evidence we present suggests that in theory, in response to the respiratory symptoms and immune system damage caused by COVID-19, taVNS can be used to improve immune function and may be an important treatment for depressive symptoms on post-COVID-19 sequelae. We describe the multi-level mechanisms linking taVNS and regulation of systemic anti-inflammatory responses and prevention of neuroinflammation present so as to treat depressive symptoms during the COVID-19 pandemic. When pro-inflammatory cytokines are present due to an infection, taVNS can activate afferent vagal neurons through impacting the immune response (139, 140) and also efferent vagal neurons can release acetylcholine through the cholinergic anti-inflammatory pathway and HPA axis (132, 141). Then, we summarize how applying taVNS and targeting cognitive and mental distress through influencing the connectivity of neural networks (121). taVNS has been shown to be associated with improved the default mode network functioning, which has been implicated in cognitive as well as emotional functioning (72, 142). Further studies are needed to understand the relationship between the immune system and the brain, as well as the role of taVNS.
This paper was primarily written by C-HL, PS, and Z-PG. Figures were produced by C-HL, Z-PG, Z-QZ, DL, and M-HY. All authors read and approved the final manuscript.
This work was supported by grants from the Municipal Natural Science Foundation of Beijing of China (Grant No. 7212200) and National Natural Science Foundation of China (Grant Nos. 81871507 and 81471389).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
ACh, acetylcholine; AMY, amygdala; ARDS, acute respiratory distress syndrome; BA, Brodmann areas; CCL-2, C-C motif chemokine ligand; COVID-19, coronavirus disease 2019; CRP, C-reactive protein; EN, epinephrine; fMRI, functional magnetic resonance imaging; GC, glucocorticoid; HIP, hippocampus; HPA, hypothalamic pituitary adrenal; IL, interleukin; IL-1ra, interleukin-1 receptor antagonist; LC, locus coeruleus; MDD, major depressive disorder; MH, medial hypothalamus; mPFC, medial prefrontal cortex; MRS, magnetic resonance spectroscopy; NAc, nucleus accumbens; NE, noradrenaline; NTS, nucleus tractus solitarius; PFC, prefrontal cortex; PTSD, post-traumatic stress disorder; rACC, rostral anterior cingulate cortex; taVNS, transcutaneous auricular vagus nerve stimulation; TCM, Traditional Chinese Medicine; TNF, tumor necrosis factor; vmPFC, ventromedial prefrontal cortex; VNS, vagus nerve stimulation.
1. Wang C, Pan R, Wan X, Tan Y, Xu L, Ho CS, et al. Immediate psychological responses and associated factors during the initial stage of the 2019 coronavirus disease (COVID-19) epidemic among the general population in China. Int J Environ Res Public Health. (2020) 17:1729. doi: 10.3390/ijerph17051729
2. Huang C, Xu X, Cai Y, Ge Q, Zeng G, Li X, et al. Mining the characteristics of COVID-19 patients in China: analysis of social media posts. J Med Internet Res. (2020) 22:e19087. doi: 10.2196/19087
3. Fudim M, Qadri YJ, Ghadimi K, MacLeod DB, Molinger J, Piccini JP, et al. Implications for neuromodulation therapy to control inflammation and related organ dysfunction in COVID-19. J Cardiovasc Transl Res. (2020) 26:1–6. doi: 10.1007/s12265-020-10031-6
4. Zhang C, Wu Z, Li JW, Zhao H, Wang GQ. Cytokine release syndrome in severe COVID-19: interleukin-6 receptor antagonist tocilizumab may be the key to reduce mortality. Int J Antimicrob Agentsm. (2020) 55:105954. doi: 10.1016/j.ijantimicag.2020.105954
5. Zhou F, Yu T, Du R, Fan G, Liu Y, Liu Z, et al. Clinical course and risk factors for mortality of adult inpatients with COVID-19 in Wuhan, China: a retrospective cohort study. Lancet. (2020) 395:1054–62. doi: 10.1016/S0140-6736(20)30566-3
6. Guo J, Feng XL, Wang XH, van IJzendoorn MH. Coping with COVID-19: exposure to COVID-19 and negative impact on livelihood predict elevated mental health problems in Chinese adults. Int J Environ Res Public Health. (2020) 17:3857. doi: 10.3390/ijerph17113857
7. Polack FP, Thomas SJ, Kitchin N, Absalon J, Gurtman A, Lockhart S, et al. Safety and efficacy of the BNT162b2 mRNA covid-19 vaccine. N Engl J Med. (2020) 383:2603–15. doi: 10.1056/NEJMoa2034577
8. Doroftei B, Ciobica A, Ilie OD, Maftei R, Ilea C. Mini-review discussing the reliability and efficiency of COVID-19 vaccines. Diagnostics. (2021) 11:579. doi: 10.3390/diagnostics11040579
9. Lai J, Ma S, Wang Y, Cai Z, Hu J, Wei N, et al. Factors associated with mental health outcomes among health care workers exposed to coronavirus disease 2019. JAMA Netw Open. (2020) 3:e203976. doi: 10.1001/jamanetworkopen.2020.3976
10. Ventureyra EC. Transcutaneous vagus nerve stimulation for partial onset seizure therapy. A new concept. Childs Nerv Syst. (2000) 16:101–2. doi: 10.1007/s003810050021
11. Butt MF, Albusoda A, Farmer AD, Aziz Q. The anatomical basis for transcutaneous auricular vagus nerve stimulation. J Anat. (2020) 236:588–611. doi: 10.1111/joa.13122
12. Rong P, Liu J, Wang L, Liu R, Fang J, Zhao J, et al. Effect of transcutaneous auricular vagus nerve stimulation on major depressive disorder: a nonrandomized controlled pilot study. J Affect Disord. (2016) 195:172–9. doi: 10.1016/j.jad.2016.02.031
13. Helmers SL, Begnaud J, Cowley A, Corwin HM, Edwards JC, Holder DL, et al. Application of a computational model of vagus nerve stimulation. Acta Neurol Scand. (2012) 126:336–43. doi: 10.1111/j.1600-0404.2012.01656.x
14. Berry SM, Broglio K, Bunker M, Jayewardene A, Olin B, Rush AJ. A patient-level meta-analysis of studies evaluating vagus nerve stimulation therapy for treatment-resistant depression. Med Devices. (2013) 6:17–35. doi: 10.2147/MDER.S41017
15. Grimonprez A, Raedt R, Baeken C, Boon P, Vonck K. The antidepressant mechanism of action of vagus nerve stimulation: evidence from preclinical studies. Neurosci Biobehav Rev. (2015) 56:26–34. doi: 10.1016/j.neubiorev.2015.06.019
16. Bonaz B, Sinniger V, Pellissier S. The vagus nerve in the neuro-immune axis: implications in the pathology of the gastrointestinal tract. Front Immunol. (2017) 8:1452. doi: 10.3389/fimmu.2017.01452
17. Wang H, Yu M, Ochani M, Amella CA, Tanovic M, Susarla S, et al. Nicotinic acetylcholine receptor α7 subunit is an essential regulator of inflammation. Nature. (2013) 421:384–8. doi: 10.1038/nature01339
18. Kaniusas E, Szeles JC, Kampusch S, Alfageme-Lopez N, Yucuma-Conde D, Li X, et al. Non-invasive auricular vagus nerve stimulation as a potential treatment for Covid19-originated acute respiratory distress syndrome. Front Physiol. (2020) 11:890. doi: 10.3389/fphys.2020.00890
19. Staats P, Giannakopoulos G, Blake J, Liebler E, Levy RM. The use of non-invasive vagus nerve stimulation to treat respiratory symptoms associated with COVID-19: a theoretical hypothesis and early clinical experience. Neuromodulation. (2020) 23:784–8. doi: 10.1111/ner.13172
20. Krzyzaniak MJ, Peterson CY, Cheadle G, Loomis W, Wolf P, Kennedy V, et al. Efferent vagal nerve stimulation attenuates acute lung injury following burn: the importance of the gut-lung axis. Surgery. (2011) 150:379–89. doi: 10.1016/j.surg.2011.06.008
21. Ellrich J. Transcutaneous auricular vagus nerve stimulation. J Clin Neurophysiol. (2019) 36:437–42. doi: 10.1097/WNP.0000000000000576
22. Zhang Y, Huang Y, Li H, Yan Z, Zhang Y, Liu X, et al. Transcutaneous auricular vagus nerve stimulation (taVNS) for migraine: an fMRI study. Reg Anesth Pain Med. (2021) 46:145–50. doi: 10.1136/rapm-2020-102088
23. Li J, Zhang Q, Li S, Niu L, Ma J, Wen L, et al. α7nAchR mediates transcutaneous auricular vagus nerve stimulation-induced neuroprotection in a rat model of ischemic stroke by enhancing axonal plasticity. Neurosci Lett. (2020) 730:135031. doi: 10.1016/j.neulet.2020.135031
24. Wang Z, Yu L, Wang S, Huang B, Liao K, Saren G, et al. Chronic intermittent low-level transcutaneous electrical stimulation of auricular branch of vagus nerve improves left ventricular remodeling in conscious dogs with healed myocardial infarction. Circ Heart Fail. (2014) 7:1014–21. doi: 10.1161/CIRCHEARTFAILURE
25. Ma J, Zhang L, He G, Tan X, Jin X, Li C. Transcutaneous auricular vagus nerve stimulation regulates expression of growth differentiation factor 11 and activin-like kinase 5 in cerebral ischemia/reperfusion rats. J Neurol Sci. (2016) 369:27–35. doi: 10.1016/j.jns.2016.08.004
26. Wu D, Ma J, Zhang L, Wang S, Tan B, Jia G. Effect and safety of transcutaneous auricular vagus nerve stimulation on recovery of upper limb motor function in subacute ischemic stroke patients: a randomized pilot study. Neural Plast. (2020) 2020:8841752. doi: 10.1155/2020/8841752
27. King HC, Hickey AH, Connelly C. (2013). Auricular acupuncture: a brief introduction for military providers. Mil Med. 178(8):867–74. doi: 10.7205/MILMED-D-13-00075
28. Veith I. Acupuncture in traditional Chinese medicine. An historical review. Calif Med. (1973) 118:70–9.
29. Round R, Litscher G, Bahr F. Auricular acupuncture with laser. Evid Based Complement Alternat Med. (2013) 2013:984763. doi: 10.1155/2013/984763
30. Nogier P. Face to face with auriculotherapy. Acupunct Electrother Res. (1983) 8:99–100. doi: 10.3727/036012983816715055
31. World Health Organization. WHO Report of the Working Group on Auricular Nomenclature. Lyon: World Health Organization (1990).
32. Usichenko T, Hacker H, Lotze M. Transcutaneous auricular vagal nerve stimulation (taVNS) might be a mechanism behind the analgesic effects of auricular acupuncture. Brain Stimul. (2017) 10:1042–4. doi: 10.1016/j.brs.2017.07.013
33. Foley JO, Dubois FS. Quantitative studies of the vagus nerve in the cat. I. The ratio of sensory to motor fibers. J Comp Neurol. (1937) 67:49–67. doi: 10.1002/cne.900670104
34. Clerkin KJ, Fried JA, Raikhelkar J, Sayer G, Griffin JM, Masoumi A, et al. COVID-19 and cardiovascular disease. Circulation. (2020) 141:1648–55. doi: 10.1161/CIRCULATIONAHA
35. Ye T, Li Y, Xiong D, Gong S, Zhang L, Li B, et al. Combination of Danshen and ligustrazine has dual anti-inflammatory effect on macrophages and endothelial cells. J Ethnopharmacol. (2020) 266:113425. doi: 10.1016/j.jep.2020.113425
36. Bonaz B, Sinniger V, Pellissier S. Targeting the cholinergic anti-inflammatory pathway with vagus nerve stimulation in patients with Covid-19? Bioelectron Med. (2020) 6:15. doi: 10.1186/s42234-020-00051-7
37. Koralnik IJ, Tyler KL. COVID-19: a global threat to the nervous system. Ann Neurol. (2020) 88:1–11. doi: 10.1002/ana.25807
38. Zhou Z, Kang H, Li S, Zhao X. Understanding the neurotropic characteristics of SARS-CoV-2: from neurological manifestations of COVID-19 to potential neurotropic mechanisms. J Neurol. (2020) 267:2179–84. doi: 10.1007/s00415-020-09929-7
39. Bo HX, Li W, Yang Y, Wang Y, Zhang Q, Cheung T, et al. Posttraumatic stress symptoms and attitude toward crisis mental health services among clinically stable patients with COVID-19 in China. Psychol Med. (2020). 27:1–2. doi: 10.1017/S0033291720000999
40. Zhang J, Lu H, Zeng H, Zhang S, Du Q, Jiang T, et al. The differential psychological distress of populations affected by the COVID-19 pandemic. Brain Behav Immun. (2020) 87:49–50. doi: 10.1016/j.bbi.2020.04.031
41. Kremer S, Lersy F, de Sèze J, Ferré JC, Maamar A, Carsin-Nicol B, et al. Brain MRI findings in severe COVID-19: a retrospective observational study. Radiology. (2020) 297:E242–51. doi: 10.1148/radiol.2020202222
42. Coolen T, Lolli V, Sadeghi N, Rovai A, Trotta N, Taccone FS, et al. Early postmortem brain MRI findings in COVID-19 non-survivors. Neurology. (2020) 95:e2016–27. doi: 10.1212/WNL.0000000000010116
43. Paterson RW, Brown RL, Benjamin L, Nortley R, Wiethoff S, Bharucha T, et al. The emerging spectrum of COVID-19 neurology: clinical, radiological and laboratory findings. Brain. (2020) 8: awaa240. doi: 10.1093/brain/awaa240
44. Bridwell R, Long B, Gottlieb M. Neurologic complications of COVID-19. Am J Emerg Med. (2020) 38:1549.e3–7. doi: 10.1016/j.ajem.2020.05.024
45. Li Z, Liu T, Yang N, Han D, Mi X, Li Y, et al. Neurological manifestations of patients with COVID-19: potential routes of SARS-CoV-2 neuroinvasion from the periphery to the brain. Front Med. (2020) 14:533–41. doi: 10.1007/s11684-020-0786-5
46. Smith RS. The macrophage theory of depression. Med Hypotheses. (1991) 35:298–306. doi: 10.1016/0306-9877(91)90272-z
47. Maes M, Meltzer HY, Scharpé S, Bosmans E, Suy E, De Meester I, et al. Relationships between lower plasma L-tryptophan levels and immune-inflammatory variables in depression. Psychiatry Res. (1993) 49:151–65. doi: 10.1016/0165-1781(93)90102-m
48. Sluzewska A, Rybakowski J, Bosmans E, Sobieska M, Berghmans R, Maes M, et al. Indicators of immune activation in major depression. Psychiatry Res. (1996) 64:161–7. doi: 10.1016/s0165-1781(96)02783-7
49. Dantzer R, Kelley KW. Twenty years of research on cytokine-induced sickness behavior. Brain Behav Immun. (2007) 21:153–60. doi: 10.1016/j.bbi.2006.09.006
50. Kim YK, Na KS, Shin KH, Jung HY, Choi SH, Kim JB. Cytokine imbalance in the pathophysiology of major depressive disorder. Prog Neuropsychopharmacol Biol Psychiatry. (2007) 31:1044–53. doi: 10.1016/j.pnpbp.2007.03.004
51. Leonard B, Maes M. Mechanistic explanations how cell-mediated immune activation, inflammation and oxidative and nitrosative stress pathways and their sequels and concomitants play a role in the pathophysiology of unipolar depression. Neurosci Biobehav Rev. (2012) 36:764–85. doi: 10.1016/j.neubiorev.2011.12.005
52. Beurel E, Toups M, Nemeroff CB. The bidirectional relationship of depression and inflammation: double trouble. Neuron. (2020) 107:234–56. doi: 10.1016/j.neuron.2020.06.002
53. Dowlati Y, Herrmann N, Swardfager W, Liu H, Sham L, Reim EK, et al. A meta-analysis of cytokines in major depression. Biol Psychiatry. (2020) 67:446–57. doi: 10.1016/j.biopsych.2009.09.033
54. Howren MB, Lamkin DM, Suls J. Associations of depression with C-reactive protein, IL-1, and IL-6: a meta-analysis. Psychosom Med. (2009) 71:171–86. doi: 10.1097/PSY.0b013e3181907c1b
55. Felger JC. Imaging the role of inflammation in mood and anxiety-related disorders. Curr Neuropharmacol. (2018) 16:533–58. doi: 10.2174/1570159X15666171123201142
56. Alexopoulos GS, Morimoto SS. The inflammation hypothesis in geriatric depression. Int J Geriatr Psychiatry. (2011) 26:1109–18. doi: 10.1002/gps.2672
57. Galecki P, Talarowska M, Anderson G, Berk M, Maes M. Mechanisms underlying neurocognitive dysfunctions in recurrent major depression. Med Sci Monit. (2015) 21:1535–47. doi: 10.12659/MSM.893176
58. Wohleb ES, McKim DB, Sheridan JF, Godbout JP. Monocyte trafficking to the brain with stress and inflammation: a novel axis of immune-to-brain communication that influences mood and behavior. Front Neurosci. (2015) 8:447. doi: 10.3389/fnins.2014.00447
59. Wohleb ES, Delpech JC. Dynamic cross-talk between microglia and peripheral monocytes underlies stress-induced neuroinflammation and behavioral consequences. Prog Neuropsychopharmacol Biol Psychiatry. (2017) 79:40–8. doi: 10.1016/j.pnpbp.2016.04.013
60. Hao F, Tan W, Jiang L, Zhang L, Zhao X, Zou Y, et al. Do psychiatric patients experience more psychiatric symptoms during COVID-19 pandemic and lockdown? A case-control study with service and research implications for immunopsychiatry. Brain Behav Immun. (2020) 87:100–6. doi: 10.1016/j.bbi.2020.04.069
61. Jiang X, Dai X, Kale Edmiston E, Zhou Q, Xu K, Zhou Y, et al. Alteration of cortico-limbic-striatal neural system in major depressive disorder and bipolar disorder. J Affect Disord. (2017) 221:297–303. doi: 10.1016/j.jad.2017.05.025
62. Kong L, Chen K, Womer F, Jiang W, Luo X, Driesen N, et al. Sex differences of gray matter morphology in cortico-limbic-striatal neural system in major depressive disorder. J Psychiatr Res. (2013) 47:733–9. doi: 10.1016/j.jpsychires.2013.02.003
63. Pavuluri MN, Passarotti A. Neural bases of emotional processing in pediatric bipolar disorder. Expert Rev Neurother. (2020) 8:1381–7. doi: 10.1586/14737175.8.9.1381
64. Mayberg HS. Limbic-cortical dysregulation: a proposed model of depression. J Neuropsychiatry Clin Neurosci. (1997) 9:471–81. doi: 10.1176/jnp.9.3.471
65. Taylor SF, Liberzon I. Neural correlates of emotion regulation in psychopathology. Trends Cogn Sci. (2007) 11:413–8. doi: 10.1016/j.tics.2007.08.006
66. Siegle GJ, Thompson W, Carter CS, Steinhauer SR, Thase ME. Increased amygdala and decreased dorsolateral prefrontal BOLD responses in unipolar depression: related and independent features. Biol Psychiatry. (2007) 61:198–209. doi: 10.1016/j.biopsych.2006.05.048
67. Fitzgerald PB, Laird AR, Maller J, Daskalakis ZJ. A meta-analytic study of changes in brain activation in depression. Hum Brain Mapp. (2008) 29:683–95. doi: 10.1002/hbm.20426
68. Phillips C. Brain-derived neurotrophic factor, depression, and physical activity: making the neuroplastic connection. Neural Plast. (2017) 2017:7260130. doi: 10.1155/2017/7260130
69. Yap JYY, Keatch C, Lambert E, Woods W, Stoddart PR, Kameneva T. Critical review of transcutaneous vagus nerve stimulation: challenges for translation to clinical practice. Front Neurosci. (2020) 14:284. doi: 10.3389/fnins.2020.00284
70. Lehtimäki J, Hyvärinen P, Ylikoski M, Bergholm M, Mäkelä JP, Aarnisalo A, et al. Transcutaneous vagus nerve stimulation in tinnitus: a pilot study. Acta Otolaryngol. (2013) 133:378–82. doi: 10.3109/00016489.2012.750736
71. Straube A, Ellrich J, Eren O, Blum B, Ruscheweyh R. Treatment of chronic migraine with transcutaneous stimulation of the auricular branch of the vagal nerve (auricular t-VNS): a randomized, monocentric clinical trial. J Headache Pain. (2015) 16:543. doi: 10.1186/s10194-015-0543-3
72. Fang J, Rong P, Hong Y, Fan Y, Liu J, Wang H, et al. Transcutaneous vagus nerve stimulation modulates default mode network in major depressive disorder. Biol Psychiatry. (2016) 79:266–73. doi: 10.1016/j.biopsych.2015.03.025
73. Liu A, Rong P, Gong L, Song L, Wang X, Li L, et al. Efficacy and safety of treatment with transcutaneous vagus nerve stimulation in 17 patients with refractory epilepsy evaluated by electroencephalogram, seizure frequency, and quality of life. Med Sci Monit. (2018) 24:8439–48. doi: 10.12659/MSM.910689
74. Hasan A, Wolff-Menzler C, Pfeiffer S, Falkai P, Weidinger E, Jobst A, et al. Transcutaneous noninvasive vagus nerve stimulation (tVNS) in the treatment of schizophrenia: a bicentric randomized controlled pilot study. Eur Arch Psychiatry Clin Neurosci. (2015) 265:589–600. doi: 10.1007/s00406-015-0618-9
75. Sellaro R, van Leusden JW, Tona KD, Verkuil B, Nieuwenhuis S, Colzato LS. Transcutaneous vagus nerve stimulation enhances post-error slowing. J Cogn Neurosci. (2015) 27:2126–32. doi: 10.1162/jocn_a_00851
76. Steenbergen L, Sellaro R, Stock AK, Verkuil B, Beste C, Colzato LS. Transcutaneous vagus nerve stimulation (tVNS) enhances response selection during action cascading processes. Eur Neuropsychopharmacol. (2015) 25:773–8. doi: 10.1016/j.euroneuro.2015.03.015
77. Peuker ET, Filler TJ. The nerve supply of the human auricle. Clin Anat. (2002) 15:35–7. doi: 10.1002/ca.1089
78. Dedoncker J, Vanderhasselt MA, Ottaviani C, Slavich GM. Mental health during the COVID-19 pandemic and Beyond: the importance of the vagus nerve for biopsychosocial resilience. Neurosci Biobehav Rev. (2021) 125:1–10. doi: 10.1016/j.neubiorev.2021.02.010
79. Farmer AD, Strzelczyk A, Finisguerra A, Gourine AV, Gharabaghi A, Hasan A, et al. International consensus based review and recommendations for minimum reporting standards in research on transcutaneous vagus nerve stimulation (version 2020). Front Hum Neurosci. (2020) 14:568051. doi: 10.3389/fnhum.2020.568051
80. Aihua L, Lu S, Liping L, Xiuru W, Hua L, Yuping W. A controlled trial of transcutaneous vagus nerve stimulation for the treatment of pharmacoresistant epilepsy. Epilepsy Behav. (2014) 39:105–10. doi: 10.1016/j.yebeh.2014.08.005
81. La Marca R, Nedeljkovic M, Yuan L, Maercker A, Elhert U. Effects of auricular electrical stimulation on vagal activity in healthy men: evidence from a three-armed randomized trial. Clin Sci. (2010) 118:537–46. doi: 10.1042/CS20090264
82. Thompson SL, O'Leary GH, Austelle CW, Gruber E, Kahn AT, Manett AJ, et al. A review of parameter settings for invasive and non-invasive vagus nerve stimulation (VNS) applied in neurological and psychiatric disorders. Front Neurosci. (2021) 15:709436. doi: 10.3389/fnins.2021.709436
83. Kong J, Fang J, Park J, Li S, Rong P. Treating depression with transcutaneous auricular vagus nerve stimulation: state of the art and future perspectives. Front Psychiatry. (2018) 9:20. doi: 10.3389/fpsyt.2018.00020
84. Zhang S, He JK, Meng H, Zhao B, Zhao YN, Wang Y, et al. Effects of transcutaneous auricular vagus nerve stimulation on brain functional connectivity of medial prefrontal cortex in patients with primary insomnia. Anat Rec. (2021) 304:2426–35. doi: 10.1002/ar.24785
85. Tu Y, Fang J, Cao J, Wang Z, Park J, Jorgenson K, et al. A distinct biomarker of continuous transcutaneous vagus nerve stimulation treatment in major depressive disorder. Brain Stimul. (2018) 11:501–8. doi: 10.1016/j.brs.2018.01.006
86. Trevizol AP, Shiozawa P, Taiar I, Soares A, Gomes JS, Barros MD, et al. Transcutaneous vagus nerve stimulation (taVNS) for major depressive disorder: an open label proof-of-concept trial. Brain Stimul. (2016) 9:453–4. doi: 10.1016/j.brs.2016.02.001
87. Yakunina N, Kim SS, Nam EC. Optimization of transcutaneous vagus nerve stimulation using functional, MRI. Neuromodulation. (2017) 20:290–300. doi: 10.1111/ner.12541
88. Redgrave J, Day D, Leung H, Laud PJ, Ali A, Lindert R, et al. Safety and tolerability of transcutaneous vagus nerve stimulation in humans; a systematic review. Brain Stimul. (2018) 11:1225–38. doi: 10.1016/j.brs.2018.08.010
89. Badran BW, Mithoefer OJ, Summer CE, LaBate NT, Glusman CE, Badran AW, et al. Short trains of transcutaneous auricular vagus nerve stimulation (taVNS) have parameter-specific effects on heart rate. Brain Stimul. (2018) 11:699–708. doi: 10.1016/j.brs.2018.04.004
90. Wu C, Liu P, Fu H, Chen W, Cui S, Lu L, et al. Transcutaneous auricular vagus nerve stimulation in treating major depressive disorder: a systematic review and meta-analysis. Medicine. (2018) 97:e13845. doi: 10.1097/MD.0000000000013845
91. Huang F, Dong J, Kong J, Wang H, Meng H, Spaeth RB, et al. Effect of transcutaneous auricular vagus nerve stimulation on impaired glucose tolerance: a pilot randomized study. BMC Complement Altern Med. (2014) 4:203. doi: 10.1186/1472-6882-14-203
92. Du XJ, Dart AM, Riemersma RA. Sex differences in the parasympathetic nerve control of rat heart. Clin Exp Pharmacol Physiol. (1994) 21:485–93. doi: 10.1111/j.1440-1681.1994.tb02545.x
93. Koenig J, Thayer JF. Sex differences in healthy human heart rate variability: a meta-analysis. Neurosci Biobehav Rev. (2016) 64:288–310. doi: 10.1016/j.neubiorev.2016.03.007
94. Koenig J, Rash JA, Campbell TS, Thayer JF, Kaess M. A meta-analysis on sex differences in resting-state vagal activity in children and adolescents. Front Physiol. (2017) 8:582. doi: 10.3389/fphys.2017.00582
95. Moodithaya S, Avadhany ST. Gender differences in age-related changes in cardiac autonomic nervous function. J Aging Res. (2012) 2012:679345. doi: 10.1155/2012/679345
96. Krause B, Cohen Kadosh R. Not all brains are created equal: the relevance of individual differences in responsiveness to transcranial electrical stimulation. Front Syst Neurosci. (2014) 8:25. doi: 10.3389/fnsys.2014.00025
97. Fallgatter AJ, Ehlis AC, Ringel TM, Herrmann MJ. Age effect on far field potentials from the brain stem after transcutaneous vagus nerve stimulation. Int J Psychophysiol. (2005) 56:37–43. doi: 10.1016/j.ijpsycho.2004.09.007
98. Kemp J, Després O, Pebayle T, Dufour A. Age-related decrease in sensitivity to electrical stimulation is unrelated to skin conductance: an evoked potentials study. Clin Neurophysiol. (2014) 125:602–7. doi: 10.1016/j.clinph.2013.08.020
99. Carvalho LA, Torre JP, Papadopoulos AS, Poon L, Juruena MF, Markopoulou K, et al. Lack of clinical therapeutic benefit of antidepressants is associated overall activation of the inflammatory system. J Affect Disord. (2013) 148:136–40. doi: 10.1016/j.jad.2012.10.036
100. Kiraly DD, Horn SR, Van Dam NT, Costi S, Schwartz J, Kim-Schulze S, et al. Altered peripheral immune profiles in treatment-resistant depression: response to ketamine and prediction of treatment outcome. Transl Psychiatry. (2017) 7:e1065. doi: 10.1038/tp.2017.31
101. Ting EY, Yang AC, Tsai SJ. Role of interleukin-6 in depressive disorder. Int J Mol Sci. (2020) 21:2194. doi: 10.3390/ijms21062194
102. Harrison NA. Brain structures implicated in inflammation-associated depression. Curr Top Behav Neurosci. (2017) 31:221–48. doi: 10.1007/7854_2016_30
103. Yrondi A, Aouizerate B, El-Hage W, Moliere F, Thalamas C, Delcourt N, et al. Assessment of translocator protein density, as marker of neuroinflammation, in major depressive disorder: a pilot, multicenter, comparative, controlled, brain PET study (INFLADEP study). Front Psychiatry. (2018) 9:326. doi: 10.3389/fpsyt.2018.00326
104. Meier TB, Drevets WC, Wurfel BE, Ford BN, Morris HM, Victor TA, et al. Relationship between neurotoxic kynurenine metabolites and reductions in right medial prefrontal cortical thickness in major depressive disorder. Brain Behav Immun. (2016) 53:39–48. doi: 10.1016/j.bbi.2015.11.003
105. Mehta ND, Haroon E, Xu X, Woolwine BJ, Li Z, Felger JC. Inflammation negatively correlates with amygdala-ventromedial prefrontal functional connectivity in association with anxiety in patients with depression: preliminary results. Brain Behav Immun. (2018) 73:725–30. doi: 10.1016/j.bbi.2018.07.026
106. Haroon E, Fleischer CC, Felger JC, Chen X, Woolwine BJ, Patel T, et al. Conceptual convergence: increased inflammation is associated with increased basal ganglia glutamate in patients with major depression. Mol Psychiatry. (2016) 21:1351–7. doi: 10.1038/mp.2015.206
107. Haroon E, Chen X, Li Z, Patel T, Woolwine BJ, Hu XP, et al. Increased inflammation and brain glutamate define a subtype of depression with decreased regional homogeneity, impaired network integrity, and anhedonia. Transl Psychiatry. (2018) 8:189. doi: 10.1038/s41398-018-0241-4
108. Felger JC, Li Z, Haroon E, Woolwine BJ, Jung MY, Hu X, et al. Inflammation is associated with decreased functional connectivity within corticostriatal reward circuitry in depression. Mol Psychiatry. (2016) 21:1358–65. doi: 10.1038/mp.2015.168
109. Yin L, Xu X, Chen G, Mehta ND, Haroon E, Miller AH, et al. Inflammation and decreased functional connectivity in a widely-distributed network in depression: centralized effects in the ventral medial prefrontal cortex. Brain Behav Immun. (2019) 80:657–66. doi: 10.1016/j.bbi.2019.05.011
110. Kakeda S, Watanabe K, Katsuki A, Sugimoto K, Igata N, Ueda I, et al. Relationship between interleukin (IL)-6 and brain morphology in drug-naïve, first-episode major depressive disorder using surface-based morphometry. Sci Rep. (2018) 8:10054. doi: 10.1038/s41598-018-28300-5
111. Chen MH, Chang WC, Hsu JW, Huang KL, Tu PC, Su TP, et al. Correlation of proinflammatory cytokines levels and reduced gray matter volumes between patients with bipolar disorder and unipolar depression. J Affect Disord. (2019) 245:8–15. doi: 10.1016/j.jad.2018.10.106
112. Frodl T, Carballedo A, Hughes MM, Saleh K, Fagan A, Skokauskas N, et al. Reduced expression of glucocorticoid-inducible genes GILZ and SGK-1: high IL-6 levels are associated with reduced hippocampal volumes in major depressive disorder. Transl Psychiatry. (2012) 2:e88. doi: 10.1038/tp.2012.14
113. Doolin K, Allers KA, Pleiner S, Liesener A, Farrell C, Tozzi L, et al. Altered tryptophan catabolite concentrations in major depressive disorder and associated changes in hippocampal subfield volumes. Psychoneuroendocrinology. (2018) 95:8–17. doi: 10.1016/j.psyneuen.2018.05.019
114. Felger JC, Mun J, Kimmel HL, Nye JA, Drake DF, Hernandez CR, et al. Chronic interferon-α decreases dopamine 2 receptor binding and striatal dopamine release in association with anhedonia-like behavior in nonhuman primates. Neuropsychopharmacology. (2013) 38:2179–87. doi: 10.1038/npp.2013.115
115. Russo SJ, Nestler EJ. The brain reward circuitry in mood disorders. Nat Rev Neurosci. (2013) 14:609–25. doi: 10.1038/nrn3381
116. Diekhof EK, Kaps L, Falkai P, Gruber O. The role of the human ventral striatum and the medial orbitofrontal cortex in the representation of reward magnitude - an activation likelihood estimation meta-analysis of neuroimaging studies of passive reward expectancy and outcome processing. Neuropsychologia. (2012) 50:1252–66. doi: 10.1016/j.neuropsychologia.2012.02.007
117. Nusslock R, Brody GH, Armstrong CC, Carroll AL, Sweet LH, Yu T, et al. Higher peripheral inflammatory signaling associated with lower resting-state functional brain connectivity in emotion regulation and central executive networks. Biol Psychiatry. (2019) 86:153–62. doi: 10.1016/j.biopsych.2019.03.968
118. Cosgrove KT, Burrows K, Avery JA, Kerr KL, DeVille DC, Aupperle RL, et al. Appetite change profiles in depression exhibit differential relationships between systemic inflammation and activity in reward and interoceptive neurocircuitry. Brain Behav Immun. (2020) 83:163–71. doi: 10.1016/j.bbi.2019.10.006
119. Ye Q, Wang B, Mao J. The pathogenesis and treatment of the ‘cytokine storm’ in COVID-19. J Inf Secur. (2020) 80:607–13. doi: 10.1016/j.jinf.2020.03.037
120. Himmerich H, Patsalos O, Lichtblau N, Ibrahim MAA, Dalton B. Cytokine research in depression: principles, challenges, and open questions. Front Psychiatry. (2019) 10:30. doi: 10.3389/fpsyt.2019.00030
121. Liu CH, Yang MH, Zhang GZ, Wang XX, Li B, Li M, et al. Neural networks and the anti-inflammatory effect of transcutaneous auricular vagus nerve stimulation in depression. J Neuroinflammation. (2020) 17:54. doi: 10.1186/s12974-020-01732-5
122. Liu J, Fang J, Wang Z, Rong P, Hong Y, Fan Y, et al. Transcutaneous vagus nerve stimulation modulates amygdala functional connectivity in patients with depression. J Affect Disord. (2016) 205:319–26. doi: 10.1016/j.jad.2016.08.003
123. Wang X, Zhou X, Dai Q, Ji B, Feng Z. The role of motivation in cognitive reappraisal for depressed patients. Front Hum Neurosci. (2017) 11:516. doi: 10.3389/fnhum.2017.00516
124. Ladopoulos T, Zand R, Shahjouei S, Chang JJ, Motte J, Charles James J, et al. COVID-19: neuroimaging features of a pandemic. J Neuroimaging. (2021) 31:228–43. doi: 10.1111/jon.12819
125. Jain R, Young M, Dogra S, Kennedy H, Nguyen V, Jones S, et al. COVID-19 related neuroimaging findings: a signal of thromboembolic complications and a strong prognostic marker of poor patient outcome. J Neurol Sci. (2020) 414:116923. doi: 10.1016/j.jns.2020.116923
126. Mao L, Jin H, Wang M, Hu Y, Chen S, He Q, et al. Neurologic manifestations of hospitalized patients with coronavirus disease 2019 in Wuhan, China. JAMA Neurol. (2020) 77:683–90. doi: 10.1001/jamaneurol.2020
127. Brouwer MC, Ascione T, Pagliano P. Neurologic aspects of covid-19: a concise review. Infez Med. (2020) 28(Suppl. 1):42–5.
128. Tsai ST, Lu MK, San S, Tsai CH. The neurologic manifestations of coronavirus disease 2019 pandemic: a systemic review. Front Neurol. (2020) 11:498. doi: 10.3389/fneur.2020.00498
129. Al-Olama M, Rashid A, Garozzo D. COVID-19-associated meningoencephalitis complicated with intracranial hemorrhage: a case report. Acta Neurochir. (2020) 162:1495–9. doi: 10.1007/s00701-020-04402-w
130. Lei W, Duan Z. Advances in the treatment of cholinergic anti-inflammatory pathways in gastrointestinal diseases by electrical stimulation of vagus nerve. Digestion. (2021) 102:128–38. doi: 10.1159/000504474
131. Borovikova LV, Ivanova S, Zhang M, Yang H, Botchkina GI, Watkins LR, et al. Vagus nerve stimulation attenuates the systemic inflammatory response to endotoxin. Nature. (2000) 405:458–62. doi: 10.1038/35013070
132. Yu ZJ, Weller RA, Sandidge K, Weller EB. Vagus nerve stimulation: can it be used in adolescents or children with treatment-resistant depression? Curr Psychiatry Rep. (2008) 10:116–22. doi: 10.1007/s11920-008-0021-6
133. Bonaz B, Sinniger V, Pellissier S. Vagus nerve stimulation at the interface of brain-gut interactions. Cold Spring Harb Perspect Med. (2019) 9:a034199. doi: 10.1101/cshperspect.a034199
134. Bonaz B, Sinniger V, Pellissier S. (2016). Anti-inflammatory properties of the vagus nerve: potential therapeutic implications of vagus nerve stimulation. J Physiol. 594(20):5781–5790. doi: 10.1113/JP271539
135. Wang J, Qi F. Traditional Chinese medicine to treat COVID-19: the importance of evidence-based research. Drug Discov Ther. (2020) 14:149–50. doi: 10.5582/ddt.2020.03054
136. Ren JL, Zhang AH, Wang XJ. Traditional Chinese medicine for COVID-19 treatment. Pharmacol Res. (2020) 155:104743. doi: 10.1016/j.phrs.2020.104743
137. Yu L, Li YZ, Wang S, Wang Y. Observation of the Efficacy of Lian-hua Qingwen Granules Combined with Arbidol in the Treatment of Mild COVID-19. Chin Pharmaceut J. (2020) 55:1042–5. doi: 10.11669/cpj.2020.12.014
138. Zhang H, Zhu Y. (2020). One highly suspected case of novel coronavirus pneumonia treated by Integrated Traditional Chinese and Western medicine and nucleic acid analysis. Tianjin J. Tradit. Chin. Med. http://kns.cnki.net/kcms/detail/12.1349.R.20200227.0909.004.html
140. Wohleb ES, Godbout JP. Basic aspects of the immunology of neuroinflammation. Mod Trends Pharmacopsychiatry. (2013) 28:1–19. doi: 10.1159/000343964
141. Burger AM, D'Agostini M. Response to “the use of non-invasive vagus nerve stimulation to treat respiratory symptoms associated with COVID-19: a theoretical hypothesis and early clinical experience”. Neuromodulation. (2020) 23:1042–3. doi: 10.1111/ner.13253
Keywords: transcutaneous auricular vagus nerve stimulation, COVID-19, brain circuits, depression, epidemic
Citation: Guo Z-P, Sörös P, Zhang Z-Q, Yang M-H, Liao D and Liu C-H (2021) Use of Transcutaneous Auricular Vagus Nerve Stimulation as an Adjuvant Therapy for the Depressive Symptoms of COVID-19: A Literature Review. Front. Psychiatry 12:765106. doi: 10.3389/fpsyt.2021.765106
Received: 26 August 2021; Accepted: 12 November 2021;
Published: 15 December 2021.
Edited by:
Stefan Borgwardt, University of Lübeck, GermanyReviewed by:
Faisal Al-Otaibi, Alfaisal University, Saudi ArabiaCopyright © 2021 Guo, Sörös, Zhang, Yang, Liao and Liu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Chun-Hong Liu, bGl1Y2h1bmhvbmdAYmp6aG9uZ3lpLmNvbQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.