 Zaiquan Dong
Zaiquan Dong Xiaoling Shen
Xiaoling Shen Weihong Kuang
Weihong Kuang
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Psychiatry, 13 September 2021
Sec. Mood Disorders
Volume 12 - 2021 | https://doi.org/10.3389/fpsyt.2021.651536
This article is part of the Research TopicNovel Approaches to Improve Detection, Differentiation and Treatment in Mood Disorders View all 12 articles
Background: Major depressive disorder (MDD) and general anxiety disorder (GAD) share many common features, leading to numerous challenges in their differential diagnosis. Given the importance of the microbiota–gut–brain axis, we investigated the differences in gut microbiota between representative cases of these two diseases and sought to develop a microbiome-based approach for their differential diagnosis.
Methods: We enrolled 23 patients with MDD, 21 with GAD, and 10 healthy subjects (healthy crowd, HC) in the present study. We used 16S rRNA gene-sequencing analysis to determine the microbial compositions of the gut microbiome based on Illumina Miseq and according to the standard protocol.
Results: GAD showed a significant difference in microbiota richness and diversity as compared with HC. Additionally, Otu24167, Otu19140, and Otu19751 were significantly decreased in MDD relative to HC, and Otu2581 and Otu10585 were significantly increased in GAD relative to MDD. At the genus level, the abundances of Sutterella and Fusicatenibacter were significantly lower in MDD relative to HC, and the abundances of Fusicatenibacter and Christensenellaceae_R7_group were significantly lower in GAD than in HC. The abundance of Sutterella was significantly higher whereas that of Faecalibacterium was significantly lower in GAD relative to MDD. Moreover, we observed that Christensenellaceae_R7_group negatively correlated with the factor score (Limited to Hopelessness) and total score of HAMD-24 (p < 0.05), whereas Fusicatenibacter negatively correlated with FT4 (p < 0.05). Furthermore, the GAD group showed significant differences at the genus level for Faecalibacterium, which negatively correlated with PTC (p < 0.05).
Conclusions: This study elucidated a unique gut-microbiome signature associated with MDD and GAD that could facilitate differential diagnosis and targeted therapy.
Major depressive disorder (MDD) is characterized by deep sadness, reduced energy, vegetative nervous system dysregulation, cognitive dysfunction, and even a high suicidal tendency (1). Generalized anxiety disorder (GAD) is characterized by extreme anxiety about issues, such as security, money, and health, and accompanied by restlessness and autonomic dysfunction (2). Anxiety and depression are two common disorders that show high comorbidity (3–5). Although they share several causal and descriptive features, there are some associated differences in their clinical features and etiological factors (6). The separation of anxiety and depression disorders is extremely important for the elucidation of the underlying disease mechanisms and development of specific pharmacological and psychological treatments. Although many studies have distinguished anxiety and depression from the perspective of symptomatology and psychological, social, and physiological etiology (7–10), there remains no convincing evidence of their distinction. To further elucidate the substantial but incomplete overlap between these disorders, this study sought to determine whether patients with clinical diagnoses of MDD and GAD can be differentiated based on gut-microbiota features.
The studies conducted in recent decades indicate that gut microbiota play a crucial role in modulating brain function and human behavior (11). Furthermore, differences in gut microbiota have been identified in various psychiatric diseases, including depression, bipolar disorder, and schizophrenia (12, 13), as well as several animal models of psychiatric diseases (14–16). There is evidence for altered microbiota composition in depressed individuals (17–19), with levels of Faecalibacterium negatively correlating with symptom severity (20) and suggesting that the clinical phenotype of mental illness might be affected by gut microbiota. Additionally, studies show that probiotic administration of Bifidobacterium longum and Lactobacillus helveticus can decrease anxiety (21–23). Moreover, Chen et al. and Jiang et al. found several consistent taxonomic differences, including higher abundances of Enterobacterales, Bacteroidaceae, Escherichia/Shigella, Bacteroides, and Tyzerella and lower abundances of Firmicutes, Mollicutes, Prevotellaceae, Ruminococcaceae, Subdoligranulum, Coprococcus, and Dialister between participants with GAD and controls (24, 25).
In general, recent studies independently investigated the characteristics of gut flora in depression and anxiety patients but did not conduct a comparative analysis. In fact, few studies have explored the use of gut flora as a marker for disease diagnosis. For example, recent studies suggest that changes in intestinal microflora might be used as a biomarker for depression diagnosis and monitoring (26, 27). Zheng et al. recently identified distinct gut-microbial compositions in MDD as compared with bipolar disorder and provided a novel marker panel to distinguish MDD from bipolar disorder based on gut-microbiome signatures (28). To the best of our knowledge, there is currently no information concerning differences in intestinal flora as a biological marker identifying anxiety and depression.
Emerging evidence points to a bidirectional communication between the neuroendocrine system and gut microbiota (29). Gut microbiota can modulate central processes via endocrine pathways within the microbiota–gut–brain axis (30, 31). Sudo et al. found that elevations in plasma adrenocorticotropic hormone (ACTH) and corticosterone levels in response to stress were substantially higher in germ-free mice than in specific pathogen-free mice (32). Additionally, recent studies report that germ-free mice, which are devoid of any bacterial contamination, show reduced depression-like behaviors along with changes in the hypothalamic–pituitary–adrenal (HPA) axis as compared with specific pathogen-free mice (33, 34). On the other hand, numerous studies have verified the relationship between the hypothalamic–pituitary–thyroid (HPT) or HPA systems and the onset of depression and anxiety (32, 35). For example, Brownlie et al. proposed that a dynamic decrease in thyroid hormone levels, particularly FT3 and FT4 (36), could be related to depression. Moreover, increased activation of the HPA axis has been repeatedly observed in depressed patients, especially in the melancholic subtype (37). In most studies, these two relationships [between depression/anxiety and gut microbiota and between neuroendocrine (HPA/HPT) and gut microbiota] have been studied independently. However, whether the gut microbiota can affect the neuroendocrine system and lead to mental illnesses, including anxiety and depression, remains unclear.
In this study, we compared the differences in the intestinal flora of patients with anxiety diagnosis to those with depression diagnosis in order to determine whether intestinal flora can help distinguish between the two groups. To achieve this, we examined MDD and GAD patients without obvious anxiety and/or depressive symptoms, respectively, and used 16S rRNA gene-sequence analysis to help distinguish differences in their intestinal flora. Additionally, we analyzed the effects of different bacteria on clinical symptoms and the neuroendocrine system in order to further explore their function in these conditions.
Patients with MDD and GAD and normal control subjects participated in this study. Both MDD and GAD patients included a series of outpatients who received treatment at the West China Hospital from January to June 2019. All samples were from Chengdu, Sichuan, China, a relatively geographically closed area harboring residents with similar eating habits. The patients were diagnosed according to the Diagnostic and Statistical Manual of Mental Disorders (DSM-5) (38) at the first clinical examination, with diagnoses were confirmed by two psychiatrists (39). Patients <18 or >45 years of age and with organic etiology for their psychiatric symptoms, psychotic features, or intellectual disability were excluded. Patients included in the study were either newly diagnosed with depression or had not used psychotropic drugs for at least 6 months. The normal control subjects included 10 worker volunteers (aged 18–45 years) without current or past major psychiatric disorders. Subjects with the following conditions were also excluded: a lifetime history of bipolar disorder, schizophrenia, schizoaffective, or other psychiatric disorders; hypertension; cardiovascular disease; diabetes mellitus; obesity; liver cirrhosis; fatty liver disease; irritable bowel syndrome; inflammatory bowel disease; drug or alcohol abuse in the previous year; use of antibiotics, probiotics, prebiotics, or synbiotics in the 6 months before fecal sample collection; known active bacterial, fungal, or viral infections; and obvious dietary preferences (e.g., vegetarians). All patients completed the Hamilton Depression Rating Scale (HAMD-24) and the Hamilton Anxiety Scale (HAMA) to obtain a clinical rating of the severity of depression and anxiety (40, 41). Both scales were independently administered by two psychiatrists that were blinded to the clinical status of the participants and had attended a training session on how to administer the tests before the start of the study. To minimize the impact of accompanying symptoms, we also excluded GAD patients with HAMD-24 ≥20 and MDD patients with 14-item HAMA ≥14.
All procedures contributing to this work comply with the ethical standards of national and institutional committees on human experimentation and with the Helsinki Declaration of 1975, as revised in 2008. All procedures involving human subjects and patients were approved by the Ethics Committee of West China Hospital of Sichuan University (approval number: 2019-268). Written informed consent was obtained from all study subjects.
The HPT axis test indicators include thyroid-stimulating hormone (TSH; normal value: 0.27–4.2 mU/L), triiodothyronine (T3; normal value: 1.3–3.1 mmol/L), thyroxine (T4; normal values: 62.0–164.0 mmol/L), free triiodothyronine (FT3; normal value: 3.6–7.5 pmol/L), and free thyroxine (FT4; normal value: 12.0–22.0 pmol/L). The HPA axis test indicators include ACTH (normal value: 5.0–78.0 ng/L) and 8:00 A.M. cortisol (PTC; normal value: 147.3–609.3 mmol/L). Fasting venous blood was taken by drawing 4 mL of cubital venous blood at 8 A.M. after overnight fasting. All analyses were performed using a Roche Cobas e601 module (Roche, Basel, Switzerland) via electrochemiluminescence. All reagents and calibrations were performed according to manufacturer instructions.
Fecal samples were immediately frozen upon collection in a sterile plastic cup and stored at −80 °C before analysis. Microbial genomic DNA was extracted using the QIAamp DNA stool mini kit (Qiagen, Hilden, Germany) according to manufacturer instructions. The 16S rRNA V3-V4 amplicons were generated using the National Institutes of Health (NIH) Human Microbiome Project protocols (16S 454 Sequencing Protocol HMP Consortium; https://www.hmpdacc.org).
Libraries were prepared and paired-end sequenced with Illumina Miseq according to manufacturer instructions (42). These QIIME2 16S rRNA sequencing protocols were used to select and analyze operational taxonomic units (OTUs) (43). Sequences from this project were deposited in the NCBI Short Read Archive under BioProject ID PRJNA647236.
The sequence index file generated from the sequencing experiment was used to identify and extract the sample data saved in FASTQ format. Barcodes and the primers in the beginning and the end were used to identify and select sequence reads. The sequence number of each sample was normalized, and OTUs with 97% identity thresholds were used in the UPARSE (v.7.1; http://drive5.com/uparse/) software program. Chimeric sequences were identified and removed using UCHIME (v.4.1; http://drive5.com/uchime/). The taxonomy of each 16S rRNA gene sequence was analyzed with RDP Classifier (http://rdp.cme.msu.edu/) using the SILVA (SSU 138) 16S rRNA database at a confidence threshold of 70% (44).
Gut-microbiota-specific microbial characteristics were subjected to analysis of variance (ANOVA), emphasizing both statistical significance and biological relevance. ANOVA was used to compare the relative abundance of microbes identified with 16S rRNA sequencing.
Statistical analyses were performed using SPSS (v.21.0; IBM Corp., Armonk, NY, USA). One-way ANOVA was used to compare the continuous variables, including age, BMI, and clinical scales. Fisher's exact test was used to analyze contingency tables, and the chi-squared method was used to compare the variables of all three groups. A p < 0.05 was considered significant. The false recovery rate representing the threshold correction was generated using the Benjamini–Hochberg method (45).
The α-diversity was calculated by the ACE, Chao, Simpson, and Shannon indices. Mann–Whitney U tests were used to identify differences between the two groups. The β-diversity was calculated using the Bray–Curtis index as the distance method and reported according to principal component analysis. A hierarchical clustering tree was used to describe similarities among different data-point categories using the Bray–Curtis distance method and visualized using iTOL (https://itol.embl.de/).
PICRUSt, a bioinformatics software package that predicts metagenome functional content from marker gene (e.g., 16S rRNA) surveys and full genomes (46), was used to determine species function.
Circos software (http://circos.ca/) visualizes data in a circular layout (47) and was employed to visualize the relationship between samples and species.
Cytoscape is an open-source software platform for visualizing molecular-interaction networks and biological pathways and integrating these networks with annotations, gene-expression profiles, and other state data (48). We used this to determine the correlation between significantly different KEGG orthologs (KOs) and significantly different OTUs or species.
We collected 54 fecal samples from the study participants, including 10, 23 (18 newly diagnosed and 5 relapsed), and 21 (19 newly diagnosed and two relapsed) subjects in the healthy crowd (HC), MDD, and GAD groups, respectively. The mean age at assessment was 30.04 ± 5.90, 30.43 ± 7.95, and 30.22 ± 6.50 years for the MDD, GAD and HC groups, respectively, with no significant difference found between groups according to ANOVA. Moreover, the BMI, sex ratio, marital status, family history, and levels of neuroendocrine hormones did not differ significantly among the three groups (Table 1).
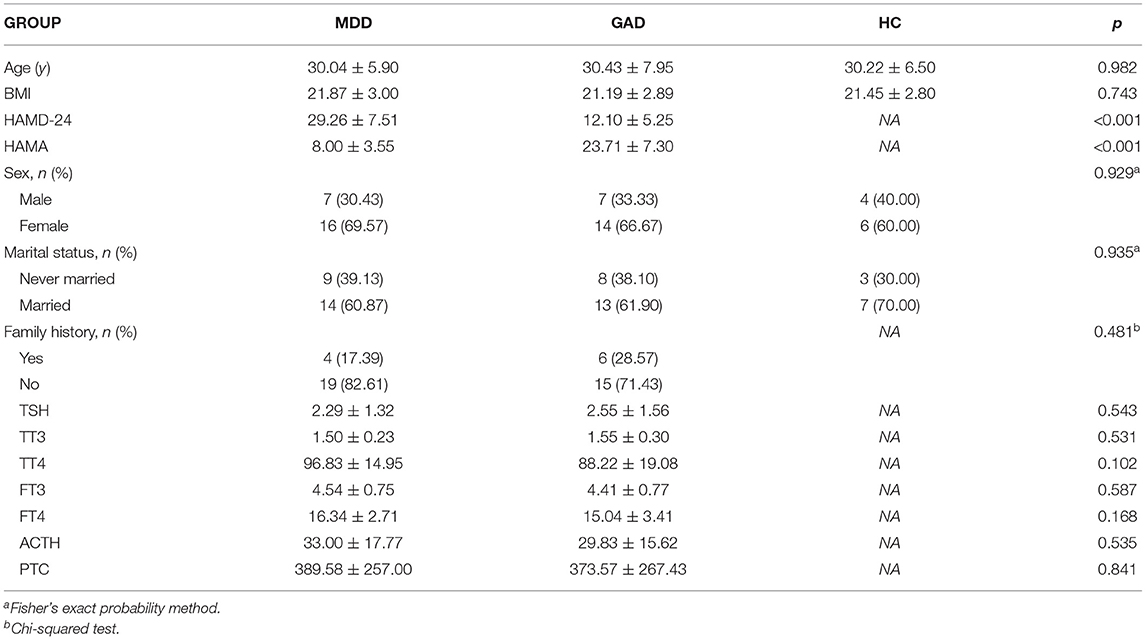
Table 1. Clinical and demographic characteristics of the MDD, GAD, and HC groups.
Accounting for 70% of the valid sequences, we obtained 1,620,000 high-quality sequences from the 54 fecal samples of all participants: the HC, MDD, and GAD groups contained 630,000, 690,000, and 300,000 sequences, respectively. The richness of gut bacterial communities in all three groups was estimated by ACE and Chao indices, and the diversity was estimated using the Shannon and Simpson diversity indices. ACE and Chao analysis showed that most of the gut microbial diversity in each sample had been captured with the current sequencing depth. After rarefying the sequencing depth among all samples using a bootstrap method (30,000 reads per sample), the Shannon and Simpson diversity index estimates were calculated, revealing no significant difference in richness and diversity between HC and MDD. However, GAD showed a significant difference in microbiota richness and diversity as compared with HC (Figures 1A–D). To explore the differences in the comprehensive microbial phenotypes of MDD, GAD, and HC, we performed β-diversity analysis. Among the three groups, statistical analysis of β-diversity at the genus level indicated that the distance was similar according to principal component analysis (Figure 1E). The hierarchical clustering tree used to describe the similarities among different data point categories using the Bray–Curtis distance method showed sample similarities between the three groups (Figure 1F).
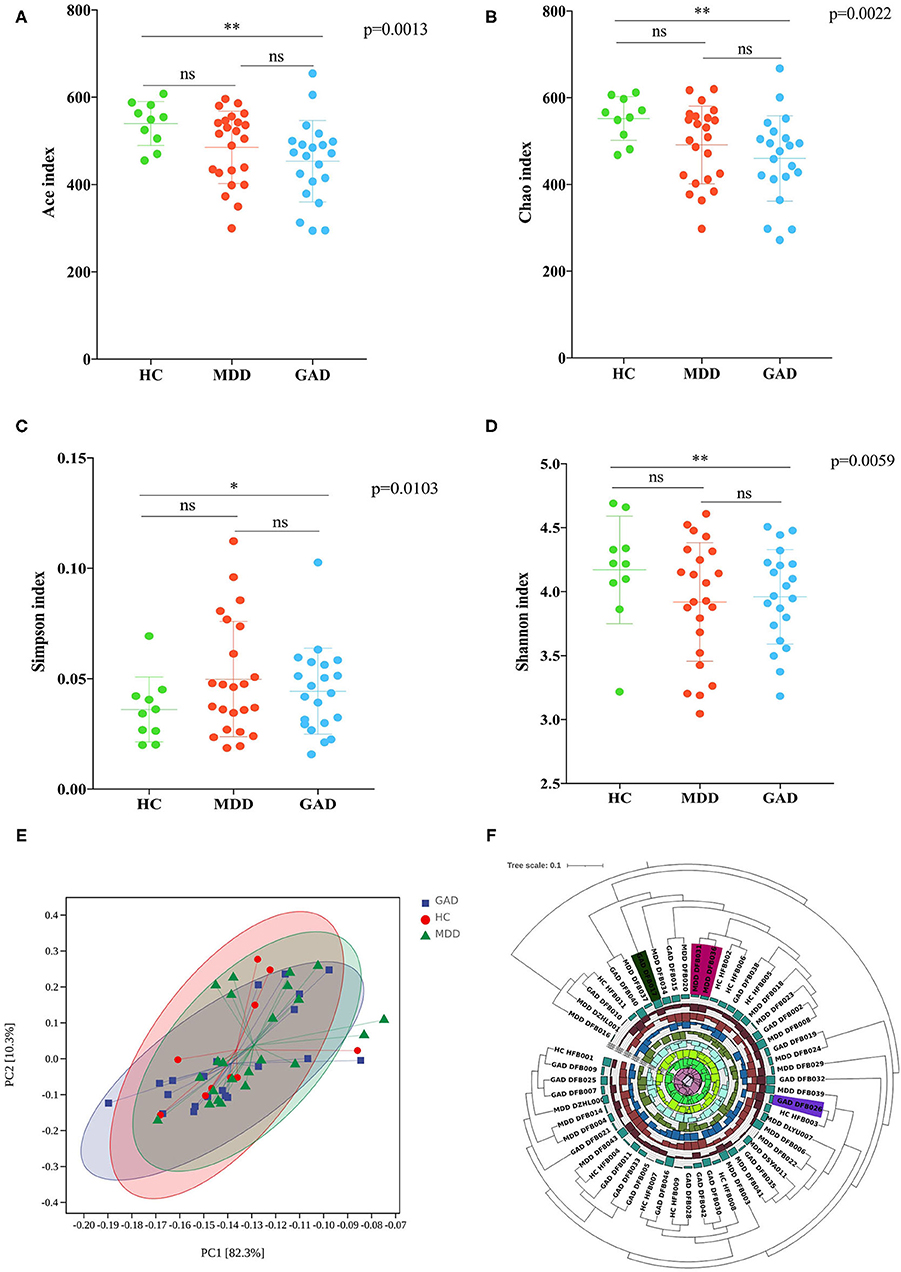
Figure 1. Diversity analysis (α and β). (A–D) Analysis of variations in richness (Chao and ACE indices) and diversity (Simpson and Shannon indices). MDD compared with HC revealed no significant difference in richness and diversity. GAD compared with HC revealed a significant difference in the richness and diversity. *p < 0.05 (Bonferroni < 0.017), **p < 0.01 (Bonferroni < 0.0033). MDD compared with GAD revealed no significant difference in richness and diversity. (E) Results of β-diversity visualized using principal component analysis (PCA; Bray–Curtis distance method). (F) Hierarchical clustering tree showing the similarities among different categories of data points by the Bray–Curtis distance method. Diversity analysis showed similar species diversity among the three groups.
We obtained 10,996, 14,406, and 15,010 species-level OTUs from the HC, MDD, and GAD groups, respectively, using the OTU cluster method (Table 2). Results of the Venn diagram showed that 5,069 OTUs were common to all three groups (Figure 2A). The relationship between samples and OTUs is presented as the Circos diagram (Figure 2B). The general overview of gut bacterial composition at the phylum and genus levels is shown in Figures 2C,E. At the phylum level, the groups were rich in Bacteroidetes, Proteobacteria, and Firmicutes, but there were differences in terms of abundance. Compared with that in the HC group, we found that the relative abundance of Proteobacteria and Firmicutes increased considerably, and that of Bacteroidetes decreased considerably in the MDD group. Additionally, the abundance of Firmicutes decreased, whereas that of Bacteroidetes and Proteobacteria increased in GAD relative to HC vs. MDD (Figure 2C). The relationship between samples and phylum-level species is presented as the Circos diagram (Figure 2D). At the genus level, we found that Bacteroides and Prevotella were abundant in the three groups. The abundance of Faecalibacterium decreased and Sutterella, Fusicatenibacter, and Christensenellaceae_R7_group increased in MDD relative to HC. The abundance of Fusicatenibacter and Christensenellaceae_R7_group decreased in GAD relative to HC. Compared with that in the MDD group, we found an increase in the relative abundance of Fusobacteria, Tenericutes, Verrucomicrobia, and Bacteroidetes but a decrease in that of Proteobacteria, Actinobacteria, and Firmicutes in the GAD group (Figure 2E). The relationship between samples and species is illustrated by the Circos diagram (Figure 2F). In summary, we found that patients with MDD or GAD showed considerable changes in gut microbiota, and that there were differences in the relative abundance of gut microbiota in patients with both disorders.
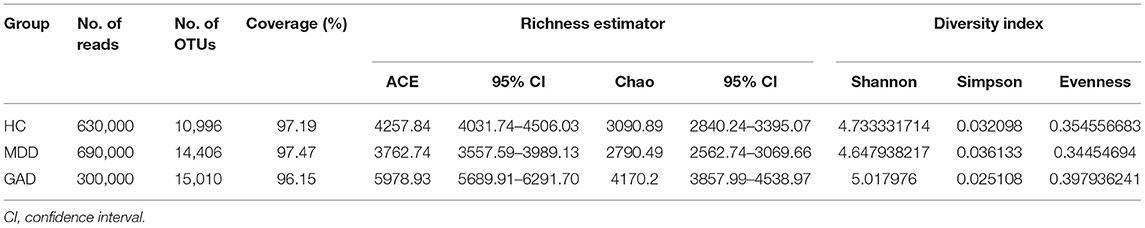
Table 2. Comparison of phylotype coverage and diversity estimation of the 16S rRNA gene libraries at 97% similarity from the sequencing analysis.
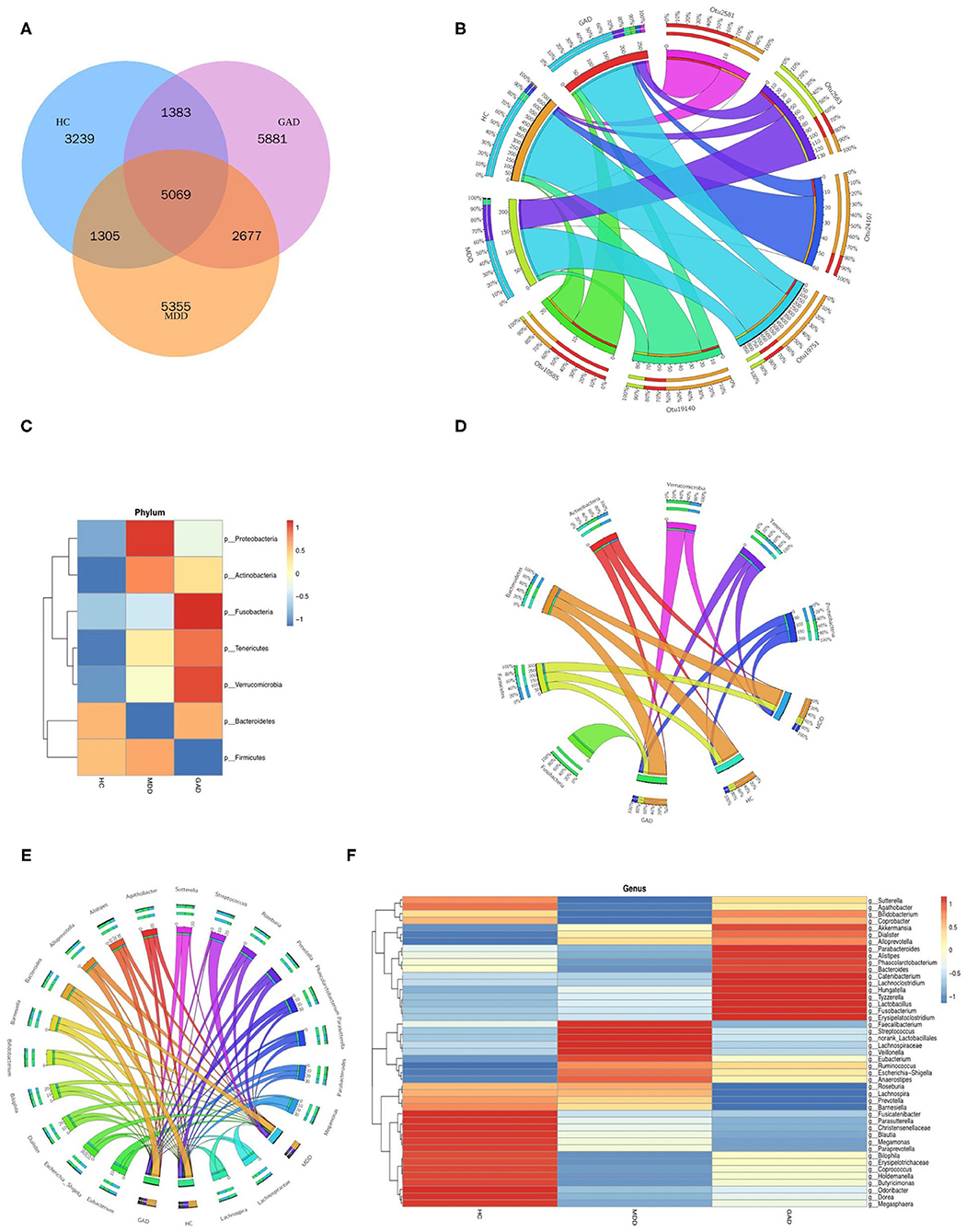
Figure 2. Species composition analysis. (A) Venn diagram of samples with common and unique OTUs. (B) Distribution of OTUs in the three groups. Data were visualized using Circos software (http://circos.ca/). The length of the bars for each sample on the outer ring represents the percentage of species in each sample. (C,E) Stacked bar plots at the phylum and genus levels show species composition according to the relative abundance of species. (D,F) Distribution of species in the three groups at the phylum and genus levels.
We the compared the relative abundance of microbial composition among the three groups in the discovery set at both the OTU and genus levels. We found taxonomic differences in fecal microbiota among the HC, MDD, and GAD groups and identified six significantly different OTUs that were altered among the three groups (Table 3 and Figure 3A). The levels of Otu24167, Otu19140, and Otu19751 were significantly decreased in MDD relative to HC, whereas Otu2563 levels were significantly increased in MDD relative to GAD or HC. Otu2581 and Otu10585 levels were significantly increased in GAD relative to MDD. Furthermore, we found no significant difference in the abundance of OTUs between GAD and HC. Similarly, at the genus level, the abundances of Sutterella and Fusicatenibacter were significantly lower in MDD relative to HC, and both Fusicatenibacter and Christensenellaceae_R7_group abundances were significantly lower in GAD relative to HC. Additionally, the abundance of Sutterella was significantly higher, whereas that of Faecalibacterium was significantly lower in GAD relative to MDD (Table 4 and Figure 3B).

Table 3. Statistical analysis of the OTUs by ANOVA.
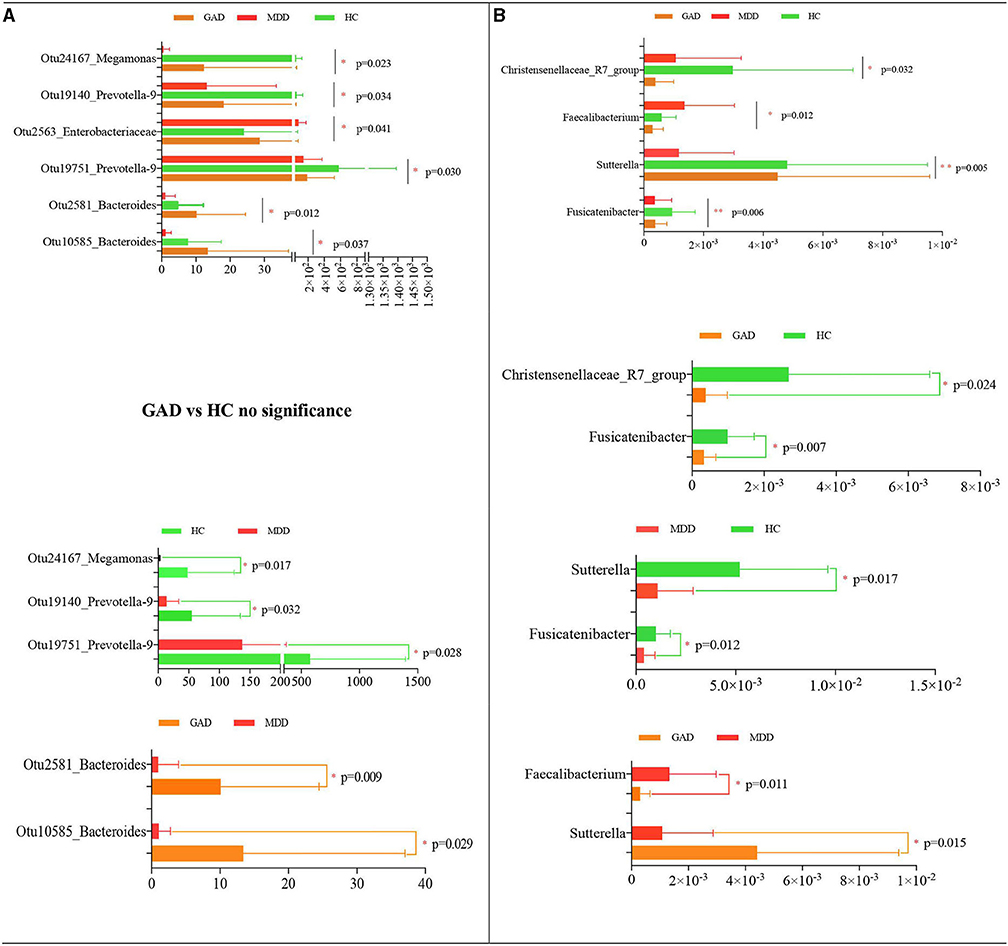
Figure 3. Statistical analysis of species differences. Taxonomic differences of fecal microbiota between HC, MDD, and GAD groups. Altered levels of specific bacterial OTUs and genera in MDD, GAD, and HC. (A) Six dominant bacterial OTUs were altered among the three groups. Otu24167, Otu19140, and Otu19751 were significantly decreased in MDD relative to HC. By contrast, Otu2563 was significantly upregulated in MDD relative to GAD or HC. Otu2581 and Otu0585 were significantly upregulated in GAD relative to MDD or HC. (B) Four significantly different species at the genus level across the three groups are shown. The abundances of Sutterella and Fusicatenibacter were significantly lower in MDD relative to HC. Fusicatenibacter and Christensenellaceae_R7_group abundances were significantly lower in GAD relative to HC. The abundance of Sutterella was higher, whereas that of Faecalibacterium was lower in GAD relative to MDD (all multiple comparisons; ANOVA tests).

Table 4. Statistical analysis of the species by ANOVA.
We obtained 6910 KOs and mapped them to the KEGG database using PICRUSt. A total of 69 significantly different KOs were obtained by ANOVA (Supplementary Material). The correlation between significantly different KOs and significantly different OTUs is depicted by a heatmap (Figure 4A). We found that the levels of two KOs (K01205 and K09011), which significantly correlated with Otu10585_Bacteroides, were significantly lower in MDD and higher in GAD relative to HC. The levels of one KO (K01201), which significantly correlated with Otu19751_Prevotella-9 and Otu19140_Prevotella-9, were lower in MDD and higher in GAD relative to HC. Another KO (K00163), which significantly correlated with Otu24167_Megamonas, showed higher levels in MDD relative to GAD and HC. Two KOs (K08281 and K03782), which significantly correlated with Otu2563_Enterobacteriaceae, showed higher levels in MDD relative to GAD and HC. Another KO (K07713), which significantly correlated with Otu2581_Bacteroides, showed lower levels in MDD relative to GAD.
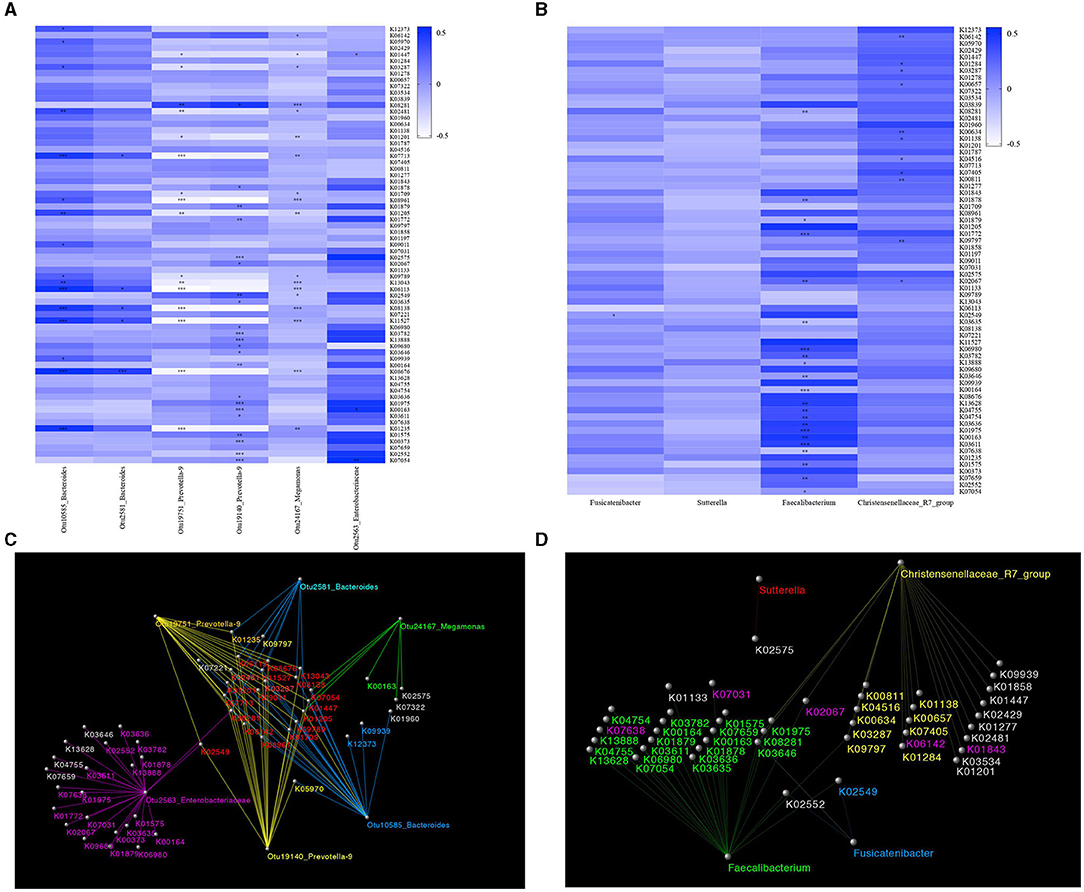
Figure 4. Functional prediction. Results of gut microbial functional pathway analyses. KOs were determined using PICRUSt. The significance of functional pathway prediction was determined by ANOVA (Supplementary Material). We obtained 69 significantly different KOs. The correlations between OTUs or species and KOs according to a heatmap are shown. Color variation indicates correlation, “+” indicates positive correlation, and “–” indicates negative correlation. *p < 0.05, **p < 0.01, ***p < 0.005. The network was used to show the correlation between significantly different KOs and OTUs or species using Cytoscape software. (A) The correlation between significantly different KOs and OTUs is shown in a heatmap. Sixteen KOs were significantly correlated with Otu10585_ Bacteroides, five KOs were significantly correlated with Otu2581_Bacteroides, 16 KOs were significantly correlated with Otu19751_Prevotella-9, 18 KOs were significantly correlated with Otu19140_Prevotella-9, 22 KOs were significantly correlated with Otu2563_Enterobacteriaceae, and three KOs were significantly correlated with Otu24167_Megamonas. (B) The correlation between significantly different KOs and species is shown in a heatmap. Eleven KOs were significantly correlated with Christensenellaceae_R7_group, 22 KOs were significantly correlated with Faecalibacterium, and one KO was significantly correlated with Fusicatenibacter. (C) Network diagram showing the relationship between significantly different KOs and OTUs. Different colors represent different OTUs. The same color between Kos and OTUs indicates a significant correlation. The red KOs represent significant correlations between KOs and multiple OTUs. (D) Network diagram showing the relationship between significantly different KOs and species. Different colors represent different species, and the same color indicates a significant correlation.
The correlation between the significantly different KOs and species is depicted by a heatmap (Figure 4B). We found that two KOs (K00657 and K04516) significantly correlated with Christensenellaceae_R7_group, and that their levels were higher in GAD and lower in MDD relative to HC. Two KOs (K08281 and K02067) significantly correlated with Faecalibacterium, and their levels were higher in MDD and lower in GAD relative to HC. One KO (K02549) significantly correlated with Fusicatenibacter, and its levels were higher in MDD and lower in GAD relative to HC. We used a network diagram to explain the relationship between the significantly different KOs and OTUs/species (Figures 4C,D).
As noted, we found four bacterial genera (Christensenellaceae_R7_group, Faecalibacterium, Fusicatenibacter, and Sutterella) with differences in MDD and GAD relative to HC or between them. These four bacterial genera can be considered as important genera that affect the disease phenotype. To further explore the functions of these different bacterial genera, we evaluated correlations among the relative abundance of bacteria, hormones (including PTC, ACTH, FT3, FT4, TT3, TT4, and TSH), and the total and factor scores of HAMD (Hopelessness, Sleep disturbance, Block, Diurnal/variation, Cognitive impairment, Weight, and Anxiety/somatic) in the MDD group and those of HAMA (Psychic anxiety and Somatic anxiety) in the GAD group. We found significant differences in correlations at the genus level for Fusicatenibacter and Christensenellaceae_R7_group in MDD patients (Figure 5A). We observed that Christensenellaceae_R7_group negatively correlated with the HAMD factor score (Limited to Hopelessness) and total score (p < 0.05), Fusicatenibacter negatively correlated with FT4 (p < 0.05), and other key phylotypes showed no strong correlation. Further, the GAD group showed significant differences at the genus level for Faecalibacterium, which negatively correlated with PTC (p < 0.05), whereas other key phylotypes showed no strong correlation (Figure 5B). The columnar stack diagram shows the correlation between clinical parameters and the significantly different species (Figures 5C,D).
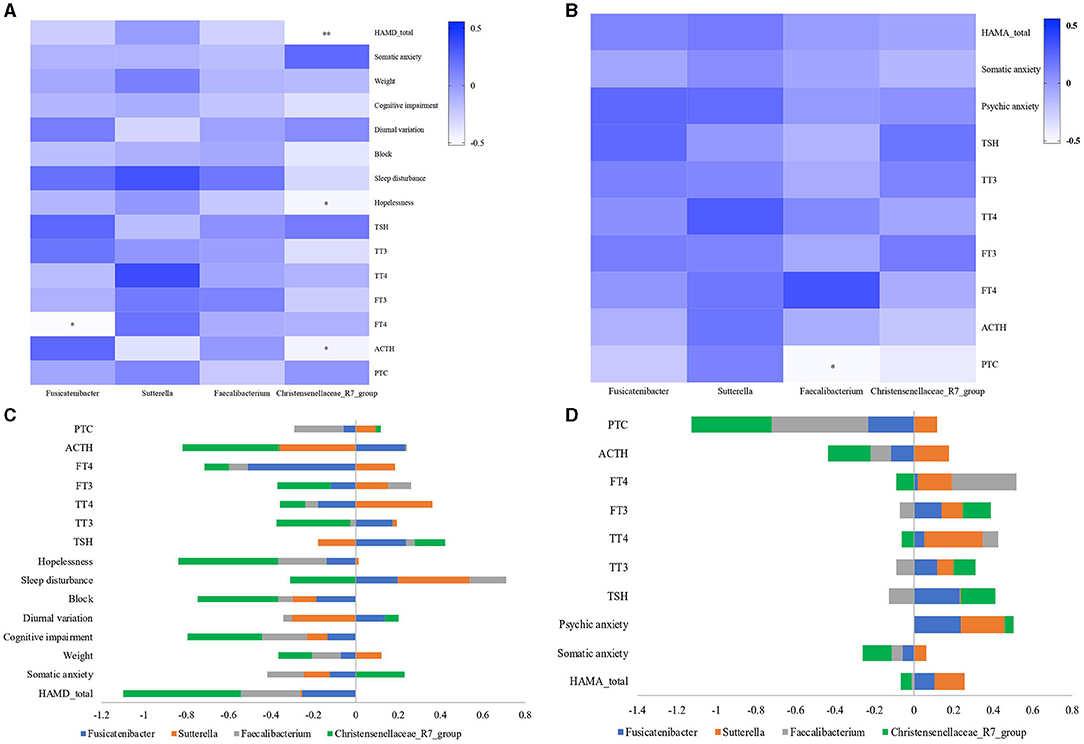
Figure 5. Relationship between gut microbiota and clinical parameters. Heatmaps were used to show relationships between significant differences in microbiota and clinical parameters, including hormones (PTC, ACTH, FT3, FT4, TT3, TT4, and TSH), the total and factor scores of HAMD (Hopelessness, Sleep disturbance, Block, Diurnal/variation, Cognitive impairment, Weight and Anxiety/somatic) in the MDD group, and the total and factor scores of HAMA (Psychic anxiety and Somatic anxiety) in the GAD group. (A) In the MDD group, Fusicatenibacter was significantly negatively correlated with FT4 (p < 0.05), and Christensenellaceae_R7_group was significantly negatively correlated with HAMD (p < 0.01), Hopelessness, and ACTH (p < 0.05). (B) In the GAD group, Faecalibacterium was significantly negatively correlated with PTC (p < 0.05). (C,D) The stacked bar plot was used to show the relationship between gut microbiota and clinical parameters (“–” shows a negative correlation, and “+” shows a positive correlation).
This study is the first to characterize and compare the gut-microbial compositions of patients with MDD and GAD. Our findings provide a better understanding of the differences between these two diseases in terms of their underlying mechanisms and will help in identifying novel therapeutic targets for better treatments.
We identified unique microbial signatures of patients with MDD and GAD relative to the HC. There was no significant difference in richness and diversity between the HC and patients with MDD. Consistent with this finding, numerous studies have reported no differences between MDD and control groups across all examined indices (49–51). However, some studies found decreased α-diversity in depressive disorders using the Shannon index (52, 53), as well as significant differences in β-diversity between participants with a depressive disorder and those in the control group (19, 52, 54–58). Based on the taxonomic findings in this study, Otu24167, Otu19140, and Otu19751 were significantly decreased in MDD as compared with HC, and the abundance of Sutterella and Fusicatenibacter at the genus level was significantly lower in patients with MDD than in the HC. Previous studies reported a lower Sutterella abundance in patients with depressive disorders than in the HC (19, 53–55); however, the association of Fusicatenibacter with depression has not previously been reported.
We found that the GAD group showed significantly higher microbiota richness and diversity than the HC, but there was no significant difference in OTU abundance between the two groups. This result differs significantly from previous reports. Chen et al. and Jiang et al. reported no difference in α-diversity between participants with an anxiety disorder and those in the control group, and that participants with GAD showed lower microbiota richness than control group subjects (24, 25). This heterogeneity of results might be attributed to numerous factors, including sample size, dietary intake, demographic characteristics of the participants, clinical status, sequencing methods, statistical methods, and/or the statistical significance threshold chosen to determine the disease-associated gut microbiota (59, 60). Based on the taxonomic findings (Figure 3B), Fusicatenibacter and Christensenellaceae_R7_group abundances were lower in the GAD group than in the HC. Mancabelli et al. reported Christensenellaceae as one of five taxa considered as a signature of a healthy gut (61). It is possible that its family might be related to affective disorders and neurological diseases (62). For example, patients with Parkinson's disease, multiple sclerosis, and autism have a remarkably lower relative abundance of Christensenellaceae (63–65).
The most important focus of this study was on distinguishing between MDD and GAD, and several significant differences were observed. Compared with patients with GAD, Otu2581 and Otu10585 levels were significantly reduced, whereas the abundance of Sutterella was decreased and that of Faecalibacterium was increased at the genus level in patients with MDD. However, no significant differences were observed in the α-diversity and richness of the intestinal floras between patients with GAD and patients with MDD, indicating that their intestinal floras were similar. Additionally, we found that Otu10585_Bacteroides significantly correlated with K09011 (map00290, valine, leucine, and isoleucine biosynthesis). Various studies also reported that MDD is associated with aberrant branched-chain amino acid and energy metabolism (66), suggesting that these amino acids (valine, leucine, and isoleucine) might serve as appropriate biomarkers for depression (67). We speculate that depression might be caused by the influence of Otu10585 on branched-chain amino acids metabolism. Moreover, the present results showed that Faecalibacterium was significantly correlated with K08281 (map00760, nicotinate, and nicotinamide metabolism) and K02067 (map02010, ABC transporters). Niacin deficiency is reportedly a contributing factor in mental-illness development and symptom alleviation (66). We speculate that the decreased abundance of Faecalibacterium might affect nicotinate and nicotinamide metabolism, leading to variations in correlative metabolism that result in MDD or GAD. ABC transporters exert notable effects on pathogen–host interactions and bacterial physiology (68), which might indicate another pathway of Faecalibacterium that results in GAD or MDD; however, the specific mechanism requires further study. Furthermore, the roles of Sutterella and Otu2581 remain unclear, although previous studies report that Sutterella is an intestinal flora associated with inflammatory responses and is found in abundance in autistic patients (69–71).
To determine why intestinal flora affect the clinical phenotype, we analyzed the correlation of some representative floras in patients with MDD or GAD with respect to the HPA or HPT axis. Our findings (Figure 5) implied that changes in intestinal flora might first induce changes in the HPA and/or HPT axis, which ultimately lead to the different clinical phenotypes of MDD and GAD. Previous reports indicated that gut-microbiota deficiency exacerbates the neuroendocrine and behavioral responses to acute stress (72–74). Other studies have also found a close relationship between HPT/HPA-axis dysfunction and depression/anxiety (32, 35, 36, 75, 76). For example, a dynamic decrease in thyroid hormone levels (particularly FT3 and FT4) is reportedly closely related to depression (36). These observations are consistent with the present findings and implications.
Additionally, we observed that Christensenellaceae_R7_group negatively correlated with factor (Limited to Hopelessness) and total scores of HAMD, suggesting that although Christensenellaceae_R7_group has not been observed as enriched in patients with MDD, it might affect the clinical manifestations and severity of MDD. Similar conclusions have been confirmed in other studies (20, 54).
This study has some limitations. First, the sample size was relatively small with no power calculation, which might have resulted in sampling bias. Second, the 16S rRNA gene sequencing used in this study resulted in limited functional information; therefore, whole-genome and whole-macrotranscriptome sequencing need to be performed in future studies. Third, we did not use standardized diagnostic tools to diagnose patients, assess the mental state of HCs and exclude comorbidities, which may weaken the reliability of the results. Last, other influential factors, such as food intake and physical activity, were not considered, which might also cause bias.
In summary, this study characterized and identified different gut-microbial compositions in subjects with MDD, subjects with GAD, and the HC. We identified a correlation between the bacteria and clinical symptoms, including a significant negative correlation between Christensenellaceae_R7_group and HAMD score. Moreover, we conducted a preliminary analysis of possible mechanisms underlying intestinal flora-affecting diseases. Our findings suggest that intestinal microflora might serve as molecular markers for distinguishing MDD from GAD.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
The studies involving human participants were reviewed and approved by the Ethics Committee of West China Hospital of Sichuan University. The patients/participants provided their written informed consent to participate in this study.
ZD and WK: conception of the work, final approval of the version to be published, and agreement to be accountable for all aspects of the work. ZD, XS, YH, JL, HL, HX, and LY: acquisition of data. ZD and LY: analysis and interpretation of data for the work. ZD: writing. WK: revising the work. All authors contributed to the article and approved the submitted version.
This work was supported by the National Natural Science Foundation of China [Grant Nos. 81621003, 81801350, and 81801357] and the Key R&D Projects of Science and Technology Department of Sichuan Province (Grant No. 2019YFS0217).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We thank Asbios (Tianjin) Biotechnology Co., LTD. for their help with microbiological analysis. We would also like to thank Editage for English language editing.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpsyt.2021.651536/full#supplementary-material
1. American Psychological Association. Diagnostic and statistical manual of mental disorders-text revision (DSM-IV-TR). Washington: APA. (2000)
2. Decker ML, Turk CL, Hess B, Murray CE. Emotion regulation among individuals classified with and without generalized anxiety disorder. J Anx Disord. (2008) 22:485–94. doi: 10.1016/j.janxdis.2007.04.002
3. Costello EJ, Mustillo S, Erkanli A, Keeler G, Angold A. Prevalence and development of psychiatric disorders in childhood and adolescence. Arch Gen Psychiatry. (2003) 60:837–44. doi: 10.1001/archpsyc.60.8.837
4. Godfrey K, Yung A, Killackey E, Cosgrave E, Yuen HP, Stanford C, et al. Patterns of current comorbidity in young help-seekers: implications for service planning and delivery. Australas Psychiatry. (2005) 13:379–83. doi: 10.1080/j.1440-1665.2005.02217.x
5. Reiter S, Emodi-Perlman A, Goldsmith C, Friedman-Rubin P, Winocur E. Comorbidity between depression and anxiety in patients with temporomandibular disorders according to the research diagnostic criteria for temporomandibular disorders. J Oral Facial Pain Headache. (2015) 29:135–43. doi: 10.11607/ofph.1297
6. Goldberg D. Towards DSM-5: the relationship between generalized anxiety disorder and major depressive episode. Psychol Med. (2008) 38:1671–5. doi: 10.1017/s003329170800295x
7. Clark LA, Watson D. Tripartite model of anxiety and depression: psychometric evidence and taxonomic implications. J Abnorm Psychol. (1991) 100:316–36. doi: 10.1037//0021-843x.100.3.316
8. Cohen JR, Young JF, Gibb BE, Hankin BL, Abela JR. Why are anxiety and depressive symptoms comorbid in youth? A multi-wave, longitudinal examination of competing etiological models. J Affect Disord. (2014) 161:21–9. doi: 10.1016/j.jad.2014.02.042
9. Krueger RF. The structure of common mental disorders. Arch Gen Psychiatry. (1999) 56:921–6. doi: 10.1001/archpsyc.56.10.921
10. Krueger RF, Caspi A, Moffitt TE, Silva PA. The structure and stability of common mental disorders (DSM-III-R): a longitudinal-epidemiological study. J Abnorm Psychol. (1998) 107:216–27. doi: 10.1037//0021-843x.107.2.216
11. Mayer EA, Knight R, Mazmanian SK, Cryan JF, Tillisch K. Gut microbes and the brain: paradigm shift in neuroscience. J Neurosci. (2014) 34:15490–6. doi: 10.1523/JNEUROSCI.3299-14.2014
12. Dickerson F, Severance E, Yolken R. The microbiome, immunity, and schizophrenia and bipolar disorder. Brain Behav Immun. (2017) 62:46–52. doi: 10.1016/j.bbi.2016.12.010
13. Rogers GB, Keating DJ, Young RL, Wong ML, Licinio J, Wesselingh S. From gut dysbiosis to altered brain function and mental illness: mechanisms and pathways. Mol Psychiatry. (2016) 21:738–48. doi: 10.1038/mp.2016.50
14. Sharon G, Cruz NJ, Kang DW, Gandal MJ, Wang B, Kim YM, et al. Human gut microbiota from autism spectrum disorder promote behavioral symptoms in mice. Cell. (2019) 177:1600–18. doi: 10.1016/j.cell.2019.05.004
15. Wong ML, Inserra A, Lewis MD, Mastronardi CA, Leong L, Choo J, et al. Inflammasome signaling affects anxiety- and depressive-like behavior and gut microbiome composition. Mol Psychiatry. (2016) 21:797–805. doi: 10.1038/mp.2016.46
16. Zheng P, Zeng B, Liu M, Chen J, Pan J, Han Y, et al. The gut microbiome from patients with schizophrenia modulates the glutamate-glutamine-GABA cycle and schizophrenia-relevant behaviors in mice. Sci Adv. (2019) 5:eaau8317. doi: 10.1126/sciadv.aau8317
17. Chen Z, Li J, Gui S, Zhou C, Chen J, Yang C, et al. Comparative metaproteomics analysis shows altered fecal microbiota signatures in patients with major depressive disorder. Neuroreport. (2018) 29:417–25. doi: 10.1097/WNR.0000000000000985
18. Valles-Colomer M, Falony G, Darzi Y, Tigchelaar EF, Wang J, Tito RY, et al. The neuroactive potential of the human gut microbiota in quality of life and depression. Nat Microbiol. (2019) 4:623–32. doi: 10.1038/s41564-018-0337-x
19. Zheng P, Zeng B, Zhou C, Liu M, Fang Z, Xu X, et al. Gut microbiome remodeling induces depressive-like behaviors through a pathway mediated by the host's metabolism. Mol Psychiatry. (2016) 21:786–96. doi: 10.1038/mp.2016.44
20. Jiang H, Ling Z, Zhang Y, Mao H, Ma Z, Yin Y, et al. Altered fecal microbiota composition in patients with major depressive disorder. Brain Behavior & Immunity. (2015) 48:186–94. doi: 10.1016/j.bbi.2015.03.016
21. Jang HM, Lee HJ, Jang SE, Han MJ, Kim DH. Evidence for interplay among antibacterial-induced gut microbiota disturbance, neuro-inflammation, and anxiety in mice. Mucosal Immunol. (2018) 11:1386–97. doi: 10.1038/s41385-018-0042-3
22. Messaoudi M, Lalonde R, Violle N, Javelot H, Desor D, Nejdi A, et al. Assessment of psychotropic-like properties of a probiotic formulation (Lactobacillus helveticus R0052 and Bifidobacterium longum R0175) in rats and human subjects. Br J Nutr. (2011) 105:755–64. doi: 10.1017/S0007114510004319
23. Mohammadi AA, Jazayeri S, Khosravi-Darani K, Solati Z, Mohammadpour N, Asemi Z, et al. The effects of probiotics on mental health and hypothalamic-pituitary-adrenal axis: a randomized, double-blind, placebo-controlled trial in petrochemical workers. Nutr Neurosci. (2016) 19:387–95. doi: 10.1179/1476830515Y.0000000023
24. Chen Y, Bai J, Wu D, Yu S, Qiang X, Bai H, et al. Association between fecal microbiota and generalised anxiety disorder: Severity and early treatment response. J Affect Disord. (2019) 259:56–66. doi: 10.1016/j.jad.2019.08.014
25. Jiang H, Zhang X, Yu Z, Zhang Z, Deng M, Zhao J, et al. Altered gut microbiota profile in patients with generalised anxiety disorder. J Psychiatr Res. (2018) 104:130–6. doi: 10.1016/j.jpsychires.2018.07.007
26. Chen JJ, Zheng P, Liu YY, Zhong XG, Wang HY, Guo YJ, et al. Sex differences in gut microbiota in patients with major depressive disorder. Neuropsychiatr Dis Treat. (2018) 14:647–55. doi: 10.2147/NDT.S159322
27. Lin P, Ding B, Feng C, Yin S, Zhang T, Qi X, et al. Prevotella and Klebsiella proportions in fecal microbial communities are potential characteristic parameters for patients with major depressive disorder. J Affect Disord. (2017) 207:300–4. doi: 10.1016/j.jad.2016.09.051
28. Zheng Q, Wang Y, Lu J, Lin W, Chen F, Jiao N. Metagenomic and metaproteomic insights into photoautotrophic and heterotrophic interactions in a Synechococcus Culture. MBio. (2020) 11:e03261–19. doi: 10.1128/mBio.03261-19
29. Farzi A, Fröhlich EE, Holzer P. Gut microbiota and the neuroendocrine system. Neurotherapeutics. (2018) 15:5–22. doi: 10.1007/s13311-017-0600-5
30. Cryan JF, Dinan TG. Mind-altering microorganisms: the impact of the gut microbiota on brain and behaviour. Nat Rev Neurosci. (2012) 13:701–12. doi: 10.1038/nrn3346
31. Kelly JR, Kennedy PJ, Cryan JF, Dinan TG, Clarke G, Hyland NP. Breaking down the barriers: the gut microbiome, intestinal permeability and stress-related psychiatric disorders. Front Cell Neurosci. (2015) 14:9:392. doi: 10.3389/fncel.2015.00392
32. Sudo N, Chida Y, Aiba Y, Sonoda J, Oyama N, Yu XN, et al. Postnatal microbial colonization programs the hypothalamic-pituitary-adrenal system for stress response in mice. J Physiol. (2004) 558:263–75. doi: 10.1113/jphysiol.2004.063388
33. Huo R, Zeng B, Li Z, Cheng K, Li B, Luo Y, et al. Microbiota modulate anxiety-like behavior and endocrine abnormalities in hypothalamic-pituitary-adrenal axis. Front Cell Infect Microbiol. (2017) 7:489. doi: 10.3389/fcimb.2017.00489
34. Luo YY, Zeng BH, Zeng L, Du XY, Li B, Hua R, et al. Gut microbiota regulates mouse behaviors through glucocorticoid receptor pathway genes in the hippocampus. Transl Psychiatry. (2018) 8:187. doi: 10.1038/s41398-018-0240-5
35. Holsboer F. The corticosteroid receptor hypothesis of depression. Neuropsychopharmacology. (2000) 23:477–501. doi: 10.1016/S0893-133X(00)00159-7
36. Brownlie BE, Rae AM, Walshe JW. Wells JE. Psychoses associated with thyrotoxicosis- ‘thyrotoxic psychosis’ A report of 18 cases, with statistical analysis of incidence. Eur J Endocrinol. (2000) 142:438–44. doi: 10.1530/eje.0.1420438
37. Juruena MF, Bocharova M, Agustini B, Young AH. Atypical depression and non-atypical depression: Is HPA axis function a biomarker? A systematic review J Affect Disord. (2018) 233:45–67. doi: 10.1016/j.jad.2017.09.052
38. American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders (DSM-5). Philadelphia: American Psychiatric Pub (2013).
39. Aizawa E, Tsuji H, Asahara T, Takahashi T, Teraishi T, Yoshida S, et al. Possible association of bifidobacterium and lactobacillus in the gut microbiota of patients with major depressive disorder. J Affect Disord. (2016) 202:254–7. doi: 10.1016/j.jad.2016.05.038
40. Hamilton M. Rating depressive patients. J Clin Psychiatry. (1980) 41:21–4. doi: 10.1111/j.1469-7610.1980.tb00039.x
41. Hamilton M. The assessment of anxiety states by rating. Br J Med Psychol. (1959) 32:50–5. doi: 10.1111/j.2044-8341.1959.tb00467.x
42. Bartram AK, Lynch MD, Stearns JC, Moreno-Hagelsieb G, Neufeld JD. Generation of multimillion-sequence 16S rRNA gene libraries from complex microbial communities by assembling paired-end illumina reads. Appl Environ Microbiol. (2011) 77:3846–52. doi: 10.1128/AEM.02772-10
43. Kuczynski J, Stombaugh J, Walters WA, Gonzalez A, Caporaso JG, Knight R. Using QIIME to analyze 16S rRNA gene sequences from microbial communities. Curr Protoc Microbiol. (2012) 1:1E 5. doi: 10.1002/9780471729259.mc01e05s27
44. Yilmaz P, Parfrey LW, Yarza P, Gerken J, Pruesse E, Quast C, et al. The SILVA and “All-species Living Tree Project (LTP)” taxonomic frameworks. Nucleic Acids Res. (2014) 42:D643–8. doi: 10.1093/nar/gkt1209
45. Ghosh D. Wavelet-based benjamini-hochberg procedures for multiple testing under dependence. MBE. (2019) 17:56–72. doi: 10.3934/mbe.2020003
46. Langille MGI, Zaneveld J, Caporaso JG, Mcdonald D, Knights D, Reyes JA, et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat Biotechnol. (2013) 31:814–21. doi: 10.1038/nbt.2676
47. Bunting SF, Nussenzweig A. End-joining, translocations and cancer. Nat Rev Cancer. (2013) 13:443–54. doi: 10.1038/nrc3537
48. Shannon P, Markiel A, Ozier O, Baliga NS, Wang JT, Ramage D, et al. Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res. (2003) 13:2498–504. doi: 10.1101/gr.1239303
49. Chahwan B, Kwan S, Isik A, van Hemert S, Burke C, Roberts L. Gut feelings: A randomised, triple-blind, placebo-controlled trial of probiotics for depressive symptoms. J Affect Disord. (2019) 253:317–26. doi: 10.1016/j.jad.2019.04.097
50. Mason BL, Li Q, Minhajuddin A, Czysz AH, Coughlin LA, Hussain SK, et al. Reduced anti-inflammatory gut microbiota are associated with depression and anhedonia. J Affect Disord. (2020) 266:394–401. doi: 10.1016/j.jad.2020.01.137
51. Vinberg M, Ottesen NM, Meluken I, Sørensen N, Pedersen O, Kessing LV, et al. Remitted affective disorders and high familial risk of affective disorders associate with aberrant intestinal microbiota. Acta Psychiat Scand. (2019) 139:174–84. doi: 10.1111/acps.12976
52. Huang Y, Shi X, Li Z, Shen Y, Shi X, Wang L, et al. Possible association of firmicutes in the gut microbiota of patients with major depressive disorder. Neuropsychiatr Dis Treat. (2018) 14:3329–37. doi: 10.2147/NDT.S188340
53. Liu Y, Zhang L, Wang X, Wang Z, Zhang J, Jiang R, et al. Similar fecal microbiota signatures in patients with Diarrhea-predominant irritable bowel syndrome and patients with depression. Clin Gastroentorol Hepatol. (2016) 14:1602–11.e5. doi: 10.1016/j.cgh.2016.05.033
54. Ye X, Wang D, Zhu H, Wang D, Li Jing, Tang Yanqing, et al. Gut microbiota changes in patients with major depressive disorder treated with vortioxetine. Front Psychiatry. (2021) 12:641491. doi: 10.3389/fpsyt.2021.641491
55. Chung YE, Chen H, Chou HL, Chen I, Lee M, Chuang L, et al. Exploration of microbiota targets for major depressive disorder and mood related traits. J Pstchiatr Res. (2019) 111:74–82. doi: 10.1016/j.jpsychires.2019.01.016
56. Kelly JR, Borre Y, O'Brien C, Patterson E, El Aidy S, Deane J, et al. Transferring the blues: Depression-associated gut microbiota induces neurobehavioural changes in the rat. J Psychiatr Res. (2016) 82:109–18. doi: 10.1016/j.jpsychires.2016.07.019
57. Lai W, Deng W, Xu S, Zhao J, Xu D, Liu Y, et al. Shotgun metagenomics reveals both taxonomic and tryptophan pathway differences of gut microbiota in major depressive disorder patients. Psychol Med. (2019) 51:1–12. doi: 10.1017/s0033291719003027
58. Sanada K, Nakajima S, Kurokawa S, Barcelo-Soler A, Ikuse D, Hirata A, et al. Gut microbiota and major depressive disorder: a systematic review and meta-analysis. J Affect Disord. (2020) 266:1–13. doi: 10.1016/j.jad.2020.01.102
59. Cheung SG, Goldenthal AR, Uhlemann AC, Mann JJ, Miller JM, Sublette ME. Systematic review of gut microbiota and major depression. Front Psychiatry. (2019) 10:34. doi: 10.3389/fpsyt.2019.00034
60. Busche S, Shao X, Caron M, Kwan T, Allum F, Cheung WA, et al. Population whole-genome bisulfite sequencing across two tissues highlights the environment as the principal source of human methylome variation. Genome Biol. (2015) 16:290. doi: 10.1186/s13059-015-0856-1
61. Mancabelli L, Milani C, Lugli GA, Turroni F, Cocconi D, van Sinderen D, et al. Identification of universal gut microbial biomarkers of common human intestinal diseases by meta-analysis. FEMS Microbiol Ecol. (2017) 93:12. doi: 10.1093/femsec/fix153
62. Waters JL, Ley RE. The human gut bacteria Christensenellaceae are widespread, heritable, and associated with health. BMC Biol. (2019) 17:83. doi: 10.1186/s12915-019-0699-4
63. Pedrosa Carrasco AJ, Timmermann L, Pedrosa DJ. Management of constipation in patients with Parkinson's disease. NPJ Parkinsons Dis. (2018) 4:6. doi: 10.3109/9780203008454-42
64. Wiesel PH, Norton C, Glickman S, Kamm MA. Pathophysiology and management of bowel dysfunction in multiple sclerosis. Eur J Gastroenterol Hepatol. (2001) 13:441–8. doi: 10.1097/00042737-200104000-00025
65. Dan Z, Mao X, Liu Q, Guo M, Zhuang Y, Liu Z, et al. Altered gut microbial profile is associated with abnormal metabolism activity of Autism Spectrum Disorder. Gut Microbes. (2020) 11:1246–67. doi: 10.1080/19490976.2020.1747329
66. Liu LY, Zhang HJ, Luo LY, Pu JB, Liang WQ, Zhu CQ, et al. Blood and urinary metabolomic evidence validating traditional Chinese medicine diagnostic classification of major depressive disorder. Chin Med. (2018) 13:53. doi: 10.1186/s13020-018-0211-z
67. Baranyi A, Amouzadeh-Ghadikolai O, von Lewinski D, Rothenhausler HB, Theokas S, Robier C, et al. Branched-chain amino acids as new biomarkers of major depression–a novel neurobiology of mood disorder. PLoS ONE. (2016) 11:e0160542. doi: 10.1371/journal.pone.0160542
68. Durmort C, Brown JS. Streptococcus pneumoniae lipoproteins and ABC transporters. Streptococcus Pneumoniae. (2015) 181–206. doi: 10.1016/B978-0-12-410530-0.00010-7
69. Mangin I, Bonnet R, Seksik P, Rigottier-Gois L, Sutren M, Bouhnik Y, et al. Molecular inventory of faecal microflora in patients with Crohn's disease. FEMS Microbiol Ecol. (2004) 50:25–36. doi: 10.1016/j.femsec.2004.05.005
70. Gophna U, Sommerfeld K, Gophna S, Doolittle WF. Veldhuyzen van Zanten SJ. Differences between tissue-associated intestinal microfloras of patients with Crohn's disease and ulcerative colitis. J Clin Microbiol. (2006) 44:4136. doi: 10.1128/JCM.01004-06
71. Williams BL, Hornig M, Parekh T, Lipkin WI. Application of novel PCR-based methods for detection, quantitation, and phylogenetic characterization of Sutterella species in intestinal biopsy samples from children with autism and gastrointestinal disturbances. MBio. (2012) 3:e00261–11. doi: 10.1128/mBio.00261-11
72. Crumeyrolle-Arias M, Jaglin M, Bruneau A, Vancassel S, Cardona A, Dauge V, et al. Absence of the gut microbiota enhances anxiety-like behavior and neuroendocrine response to acute stress in rats. Psychoneuroendocrinology. (2014) 42:207–17. doi: 10.1016/j.psyneuen.2014.01.014
73. Hryhorczuk C, Sharma S, Fulton SE. Metabolic disturbances connecting obesity and depression. Front Neurosci. (2013) 7:177. doi: 10.3389/fnins.2013.00177
74. Pellegrini C, Antonioli L, Calderone V, Colucci R, Fornai M, Blandizzi C. Microbiota-gut-brain axis in health and disease: Is NLRP3 inflammasome at the crossroads of microbiota-gut-brain communications? Prog Neurobiol. (2020) 191:101806. doi: 10.1016/j.pneurobio.2020.101806
75. Burke HM, Davis MC, Otte C, Mohr DC. Depression and cortisol responses to psychological stress: A meta-analysis. Psychoneuroendocrinology. (2005) 30:846–56. doi: 10.1016/j.psyneuen.2005.02.010
Keywords: gut microbiome, anxiety, depression, 16S ribosomal RNA, differential diagnosis
Citation: Dong Z, Shen X, Hao Y, Li J, Li H, Xu H, Yin L and Kuang W (2021) Gut Microbiome: A Potential Indicator for Differential Diagnosis of Major Depressive Disorder and General Anxiety Disorder. Front. Psychiatry 12:651536. doi: 10.3389/fpsyt.2021.651536
Received: 10 January 2021; Accepted: 19 August 2021;
Published: 13 September 2021.
Edited by:
Andrés Herane-Vives, University College London, United KingdomReviewed by:
Kiran Veer Sandhu, University College Cork, IrelandCopyright © 2021 Dong, Shen, Hao, Li, Li, Xu, Yin and Kuang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Weihong Kuang, a3doaGxqQDE2My5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.