 Meng Chen†
Meng Chen† Linfeng Sun
Linfeng Sun Shutang Tan
Shutang Tan
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Plant Sci., 12 March 2025
Sec. Plant Physiology
Volume 16 - 2025 | https://doi.org/10.3389/fpls.2025.1563571
This article is part of the Research TopicChemistry Bolsters Plant Physiology Research and Agricultural TechniquesView all articles
The phytohormone auxin plays a central role in plant growth and development. D6PK, a member of the AGC kinase family, phosphorylates PIN-FORMED (PIN) auxin transporters, thereby regulating PIN activity and polar auxin transport. In this study, we demonstrated that gravacin, a synthetic compound, functions as an inhibitor that targets D6PK in plants. Physiological and cell biology experiments revealed that the phenotypes of gravacin-treated plants were similar to those of d6pk d6pkl1 d6pkl2 (d0 d1 d2) triple mutants. Furthermore, in vitro kinase assays confirmed that gravacin directly inhibited the kinase activity of D6PK. Thus, by combining phenotypic analysis with cell biological and biochemical experiments, this research revealed that gravacin is an inhibitor of D6PK and elucidated the underlying mechanism. Our work provides a chemical tool that can be used to further dissect the role of D6PK and related physiological processes.
The phytohormone auxin plays an important role in governing nearly all facets of plant growth and patterning processes, from the embryonic stage to fruit development (Blázquez et al., 2020; Friml, 2022; Yu et al., 2022; Carrillo-carrasco et al., 2023). The directional transport of auxin is crucial for ensuring explicit plant development and tropic growth. The formation and maintenance of local auxin minima, maxima, and gradients depend on directional intercellular auxin transport, a process that is mediated by a group of auxin influx carriers, namely AUXIN1 (AUX1)/LIKE AUX1s (LAXs), and PIN-FORMED (PIN) auxin efflux carriers (Bennett et al., 1996; Vanneste and Friml, 2009; Tan et al., 2021; Ung et al., 2023). The PIN proteins consists of 10 transmembrane helices and a lengthy cytoplasmic loop, which harbors numerous phosphosites crucial for the regulation of PIN activity, polar distribution, and trafficking (Tan et al., 2021; Ung et al., 2023). In recent years, studies have confirmed that AGC (named after the cAMP-dependent [PKA] and cGMP dependent protein kinases [PKG], and protein kinase C [PKC]) kinases, mitogen-activated protein kinases (MAP kinases), calcium/calmodulin-dependent protein kinase-related kinases (Ca2+/CALMODULIN-DEPENDENT PROTEIN KINASE-RELATED KINASEs), and receptor kinases are capable of phosphorylating PIN proteins (Tan et al., 2021; Cui et al., 2024; Zhang et al., 2023; Luschnig and Friml, 2024; Wang et al., 2024).
D6PK, a member of the AGC kinase family, is capable of phosphorylating PIN family proteins, thereby increasing their activity in auxin transport and playing a crucial role in the directional transport of auxin (Zourelidou et al., 2009; Barbosa et al., 2014; Zourelidou et al., 2014; Weller et al., 2017). The D6PK/D6PKL subclade of AGC kinases comprise four homologous proteins, namely, D6PK, D6PKL1, D6PKL2, and D6PKL3. Loss-of-function D6PK/D6PKL mutants, such as d6pk d6pkl1 d6pkl2 (d0 d1 d2), d6pk d6pkl1 d6pkl3 (d0 d1 d3), and d6pk d6pkl1 d6pkl2 d6pkl3 (d0 d1 d2 d3), present a range of auxin-deficient phenotypes, including shoot dwarfism, reduced lateral branching, defective lateral root formation, and weakened tropism (Zourelidou et al., 2009; Barbosa et al., 2014; Zourelidou et al., 2014; Weller et al., 2017). D6PK is polarly localized at the basal side of the root cell and can directly phosphorylate the intracellular hydrophilic loops of the PIN1, PIN3, PIN4, and PIN7 proteins and activate their activities, thereby promoting polar auxin transport (Zourelidou et al., 2009). Unlike the phosphorylation mediated by PINOID (PID)/WAVY ROOT GROWTHs (WAGs), the phosphorylation cascade triggered by D6PK does not impact the polar orientation of PIN proteins but rather their ability to transport auxin across the cell membrane (Zourelidou et al., 2014; Weller et al., 2017).
Specific inhibitors play crucial roles in the study of the transport mechanism of auxin. As a classic auxin transport inhibitor, N-1-naphthylphthalamic acid (NPA) has been demonstrated to bind directly and inhibit the transport activity of PIN proteins (Abas et al., 2021; Teale et al., 2021; Lam Ung et al., 2022; Su et al., 2022; Yang et al., 2022; Xia et al., 2023). Consequently, it suppresses the efflux of auxin and interferes with plant responses mediated by auxin transport. Another classic inhibitor, 2,3,5-triiodobenzoic acid (TIBA), not only directly suppresses auxin transport mediated by PIN proteins but also disrupts the trafficking of PIN vesicles and the localization of PIN proteins at the plasma membrane (Dhonukshe et al., 2008; Zou et al., 2019; Tan et al., 2020b; Xia et al., 2023). Furthermore, naproxen suppresses auxin transport, notably PIN-mediated auxin efflux, by directly binding to PIN proteins (Tan et al., 2020b; Xia et al., 2023). A range of compounds, such as endosidin4 (ES4) (Kania et al., 2018), brefeldin A (BFA) (Geldner et al., 2001; Naramoto et al., 2014), and wortmannin (Jaillais et al., 2006; Peng et al., 2024), affect the localization of PIN proteins within the endomembrane system through indirect mechanisms (Semeradova et al., 2020; Tan et al., 2021; Peng et al., 2024). These compounds have been widely used in auxin-related studies as well as in agriculture.
Gravacin was originally isolated to inhibit the negative gravitropic growth of hypocotyls in dark-grown Arabidopsis seedlings (Rojas-Pierce et al., 2007). Previous studies have demonstrated that ABCB19 (also called P-glycoprotein 19 (PGP19)) is a potential molecular target for gravacin, 3-(5-[3,4-dichlorophenyl]-2-furyl)-acrylic acid (PubChem compound ID 776105, Chem-Bridge ID 5850247) (Rojas-Pierce et al., 2007; Kim et al., 2010). It was proposed that gravacin plays a role in the inhibition of gravitropism by suppressing the potential auxin transport activity of the PGP19 and PGP19-PIN complexes (Rojas-Pierce et al., 2007). However, the etiolated seedlings of the pgp19 mutants presented an overbent phenotype under dark conditions, which contradicts the proposed model in which gravacin inhibits PGP19 function (Rojas-Pierce et al., 2007). Moreover, pgp19-4 mutants exhibit a wild-type sensitivity to the impact of gravacin on the trafficking of the tonoplast protein marker GFP-dTIP (Rojas-Pierce et al., 2007). Recent studies revealed that the ABC transporters ABCB19 and ABCB1 preferentially transport brassinosteroids rather than auxins, as supported by both biochemical assays and structural analysis (Wei et al., 2024; Ying et al., 2024). These observations suggest that a target distinct from PGP19 might be involved in this phenotype of gravacin-treated seedlings. Here, we revealed that gravacin is an auxin transport inhibitor that targets the AGC kinase D6PK, which provides novel insights into the molecular mechanisms of this compound as well as the functions of D6PK.
Auxin plays a major role in regulating plant gravitropism in both roots and shoots, which relies on its asymmetric distribution in response to gravi-stimulation. To delineate the molecular mechanisms underlying the functions of gravacin in plants, we investigated its physiological effects in detail. When wild-type (Col-0) Arabidopsis seedlings were cultivated on Murashige and Skoog (MS) plates supplemented with varying concentrations of gravacin (1, 2, 5, and 10 µM), we found that Arabidopsis seedlings presented shorter primary roots and a lower number of lateral roots (LRs) than did those treated with the DMSO control (Figures 1A–C). To study the role of gravacin in the initiation of lateral root development in detail, lateral root primordia-probed fluorescent reporters, including pPIN1::PIN1-GFP (Benková et al., 2003) and pARF19::NLS-GFP (Rademacher et al., 2012), were used for gravacin treatment. Consistently, the results revealed that the formation of lateral root primordia was significantly lower in the gravacin treatment group than in the DMSO control group (Supplementary Figures S1A-D).
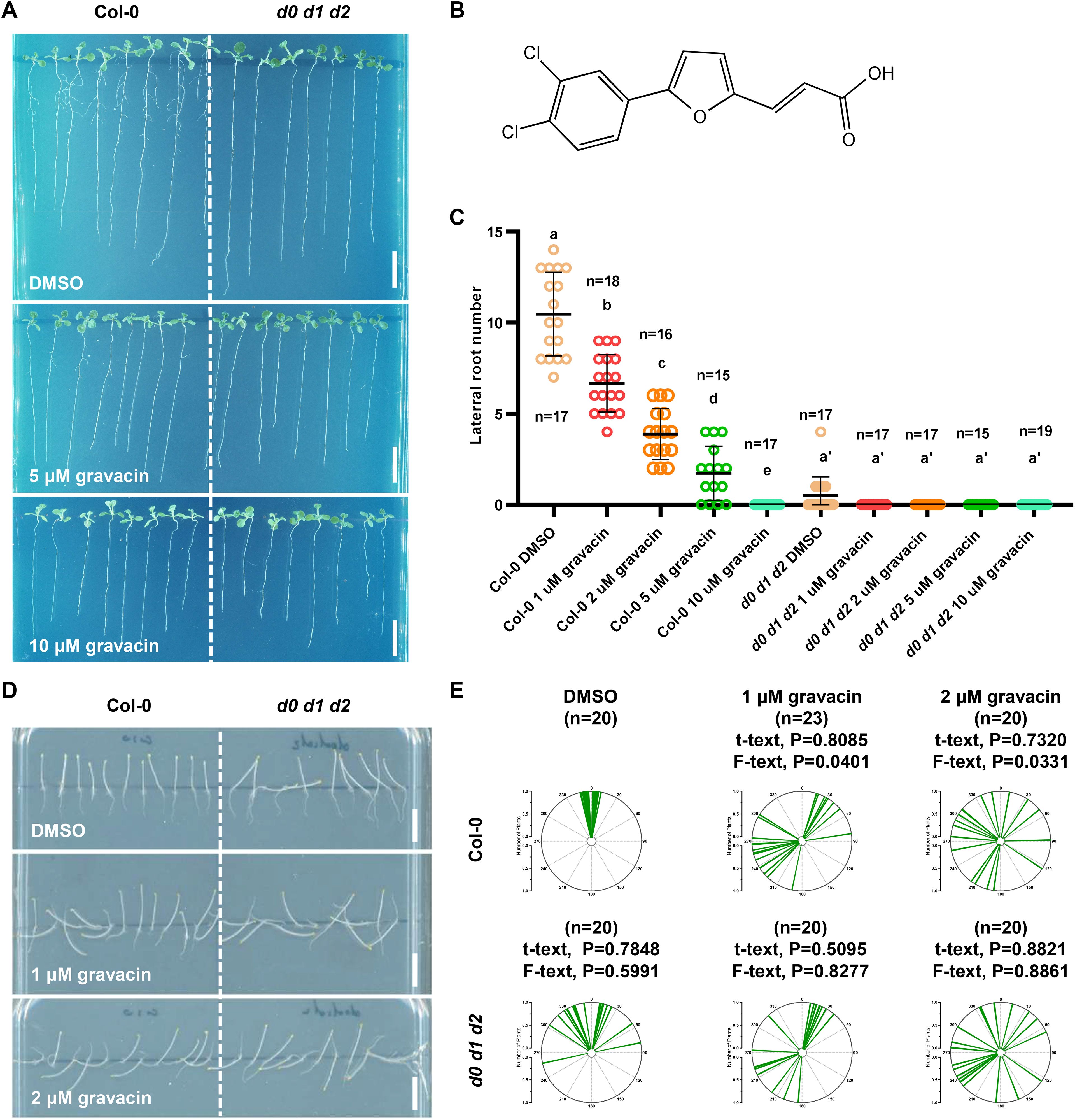
Figure 1. Gravacin inhibits lateral root formation and disrupts the gravitropism of Arabidopsis plants in the dark. (A), Representative images showing the morphological changes in 11-day-old Col-0 and d0 d1 d2 seedlings grown on MS media supplemented with gravacin. Scale bars, 1 cm. (B), The structure of gravacin. (C), Gravacin suppressed lateral root formation. The emerged lateral roots of 11-day-old Col-0 seedlings treated with gravacin were counted. The dots represent individual values, and the lines indicate the means ± SDs. Different letters represent significant differences; p < 0.05; one-way ANOVA with Tukey’s multiple comparison test. (D), Gravacin interferes with hypocotyl gravitropism under dark conditions. Four-day-old etiolated Col-0 and d0 d1 d2 seedlings were grown constantly on MS media supplemented with gravacin at the indicated concentrations. Scale bars, 1 cm. (E), The angles of Arabidopsis hypocotyls were measured via Fiji and are shown as polar charts. Each line represents the root tip angle of an individual seedling in polar charts. Unpaired t tests indicate the difference in the mean value, and F tests indicate the difference in variance. For Col-0, gravacin treatments were compared with the DMSO control, and the d0 d1 d2 groups were compared with Col-0 under the same gravacin treatment.
The lateral root phenotype under gravacin treatment closely resembled that of d0 d1 d2 triple mutants. Interestingly, d0 d1 d2 triple mutants also exhibit defects in the negative gravitropism of hypocotyls (Zourelidou et al., 2009; Barbosa et al., 2014; Tan et al., 2020a). Consistent with these findings, when wild-type Col-0 Arabidopsis seedlings were grown on MS media supplemented with the DMSO solvent control in darkness, their hypocotyls grew upright. In contrast, those seedlings that grew on MS plates supplemented with gravacin presented less gravitropic hypocotyls (Figures 1D, E). Notably, gravacin did not affect hypocotyl length, which is consistent with the phenotype of d0 d1 d2 (Supplementary Figure S3B). These findings suggest that gravacin disrupts the gravitropic response of Arabidopsis seedlings in darkness. To confirm these findings, 4-day-old etiolated Col-0 and d0 d1 d2 seedlings were transferred to plates containing DMSO or gravacin, after which they were turned 90 degrees. After 24 h of gravistimulation, the etiolated Col-0 seedlings bent upward, whereas the d0 d1 d2 seedlings bent less toward the DMSO control, and both genotypes lost gravitropism under gravacin treatment (Supplementary Figures S2A, B). Additionally, gravacin treatment impaired apical hook formation, similar to the defects observed in d0 d1 d2 mutants (Supplementary Figures S3A, C).
These findings indicate that gravacin may interfere with the same molecular pathway as that affected in the d0 d1 d2 mutants, which prompted us to explore whether D6PK is a potential target of gravacin.
To investigate the potential mechanisms by which gravacin exerts its physiological effects, we conducted RNA sequencing (RNA-seq) profiling of 6-day-old light-grown Arabidopsis Col-0 seedlings and 4-day-old dark-grown etiolated Col-0 seedlings, with each group treated with gravacin or the solvent control DMSO. Analysis of the RNA-seq data via Gene Ontology (GO) enrichment bubble plots revealed significant changes in the expression patterns of genes associated with auxin (Figures 2A, B), including genes related to auxin influx, auxin efflux, and the response to auxin. To evaluate the impact of gravacin on auxin signaling, we conducted RT-qPCR assays on several representative auxin-related genes. The results demonstrated that in the gravacin-treated Col-0 seedlings, there was a notable reduction in the expression of AUX1, IAA6, and IAA19. Conversely, the expression levels of IAR3 (IAA-amino acid hydrolase ILR1-like 4) and ABCB4 (ABC transporter B family member 4) were observed to be upregulated. In addition, we did not detect any change in D6PK/D6PKL expression under gravacin treatment in our RNA-seq data. These findings were consistent with our RNA-seq data, suggesting that gravacin modulated the auxin signaling pathway in plants (Supplementary Figures S4A, B).
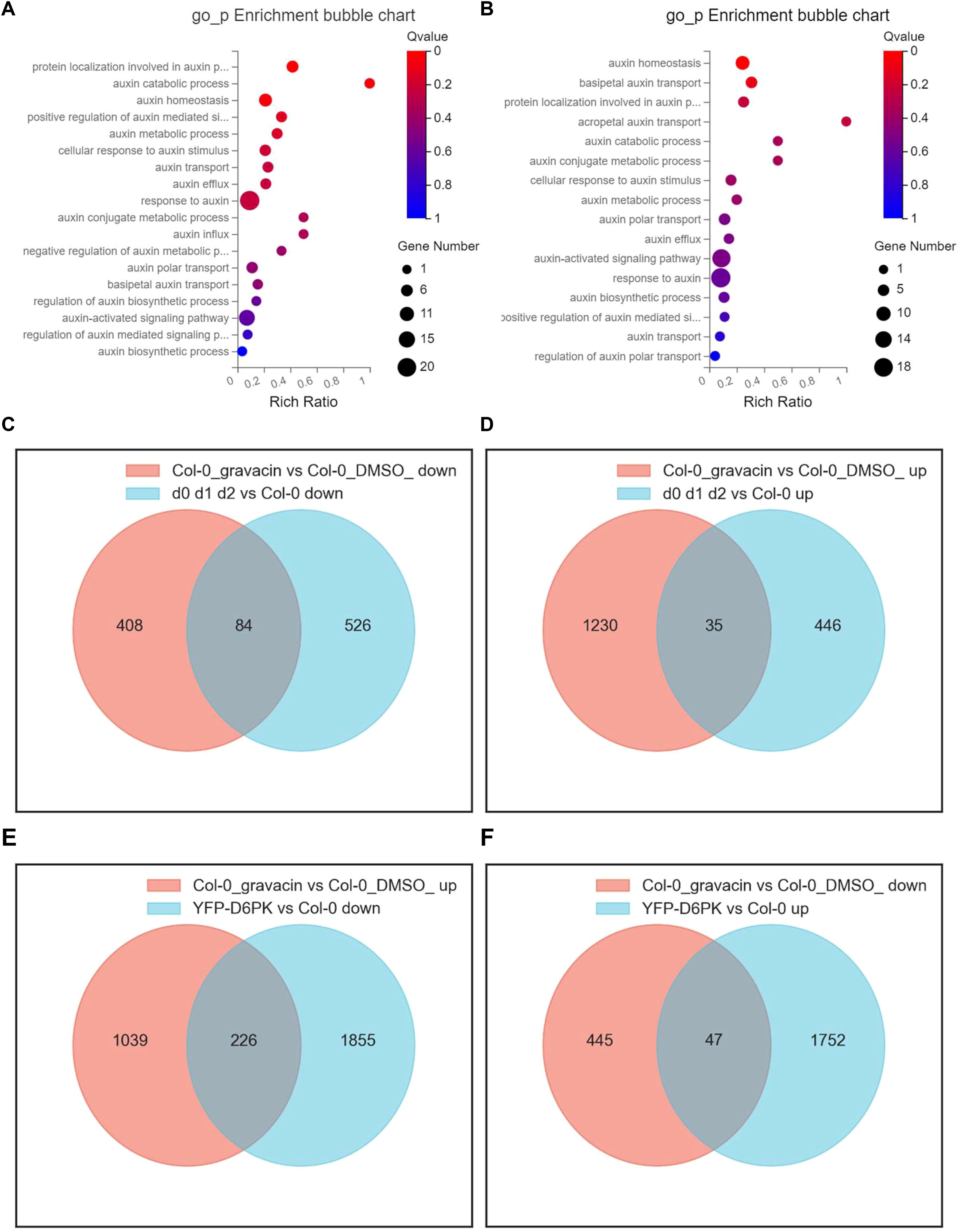
Figure 2. Gravacin treatment induces widespread changes in the transcriptional landscape, including the expression of auxin-related genes, in Arabidopsis seedlings and hypocotyls. (A), GO enrichment analysis of a schematic representation of the RNA-seq data of 6-day-old Col-0 seedlings grown vertically on MS media and treated with 10 µM gravacin or DMSO (as the mock control) for 3 hours. Differentially expressed mRNAs in Col-0 seedlings treated with gravacin versus those treated with DMSO. The X-axis label represents the enrichment ratio, and the Y-axis label represents the pathway. The size and color of the bubble represent the number of DEGs enriched in the pathway and enrichment significance, respectively. (B), GO enrichment analysis of a schematic representation of the RNA-seq data of 4-day-old Col-0 seedlings grown vertically in MS media in the dark and treated with 2 µM gravacin or DMSO (as the mock control). (C), Venn diagram showing the overlap of downregulated DEGs in the transcriptional data of gravacin-treated etiolated Col-0 seedlings versus etiolated d0 d1 d2 seedlings. Each colored circle represents a different dataset, and areas of overlap indicate shared DEGs. (P value < 0.05 and absolute log2 (fold change) > 0) in the translational data. (D), Venn diagram showing the overlap of upregulated DEGs in the transcriptional data of gravacin-treated etiolated Col-0 seedlings versus etiolated d0 d1 d2 seedlings. (E), Venn diagram showing the overlap of upregulated DEGs in the transcriptional data of gravacin-treated etiolated Col-0 seedlings versus downregulated DEGs in etiolated p35S::YFP-D6PK seedlings. (F), Venn diagram showing the overlap of downregulated DEGs in the transcriptional data of gravacin-treated etiolated Col-0 seedlings versus upregulated DEGs in etiolated p35S::YFP-D6PK seedlings.
Furthermore, Venn diagrams provided insights into the unique DEGs modulated by gravacin, as well as a distinct subset of genes that overlap with those affected in etiolated d0 d1 d2 seedlings and in etiolated p35S::YFP-D6PK (Zourelidou et al., 2009) seedlings. The data revealed that 226 genes, which were upregulated in gravacin-treated etiolated Col-0 seedlings, were subsequently downregulated in etiolated p35S::YFP-D6PK seedlings (Figure 2E). Notably, the genes whose expression was downregulated by gravacin presented pronounced changes in etiolated d0 d1 d2 seedlings (Figures 2C, D), suggesting that gravacin might be involved in D6PK-regulated pathways in plants.
Previous studies have shown that D6PK controls auxin transport and distribution by phosphorylating basally localized PIN auxin transporters. Decreased activity of PIN transporters in mutants of activating kinases, such as d6pk d6pkl1 d6pkl2 and pdk1.1 pdk1.2, resulted in an expansion of root tip regions with an enlarged DR5::GUS or DR5rev::GFP (an auxin responsive fluorescent reporter) region owing to the overproliferation of columellar cells (Zourelidou et al., 2009; Gao et al., 2020; Tan et al., 2020a; Xia et al., 2023). Since gravacin treatment affects genes related to auxin, we investigated whether exogenous auxin treatment could reverse the decrease in lateral root number observed in d0 d1 d2 mutants and gravacin-treated seedlings. As expected, auxin treatment was able to rescue the lateral root phenotype in these seedlings (Supplementary Figures S5A, B), implying that the disruption of auxin homeostasis caused by gravacin may inhibit lateral root formation. To test this hypothesis further, we used the DR5rev::GFP auxin-responsive reporter (Friml et al., 2003) as an indicator of auxin distribution and signaling. Consistent with these findings, gravacin-treated roots presented broader DR5 signal, suggesting that, compared with d0 d1 d2, gravacin had a similar effect on auxin distribution. (Figures 3A, B).
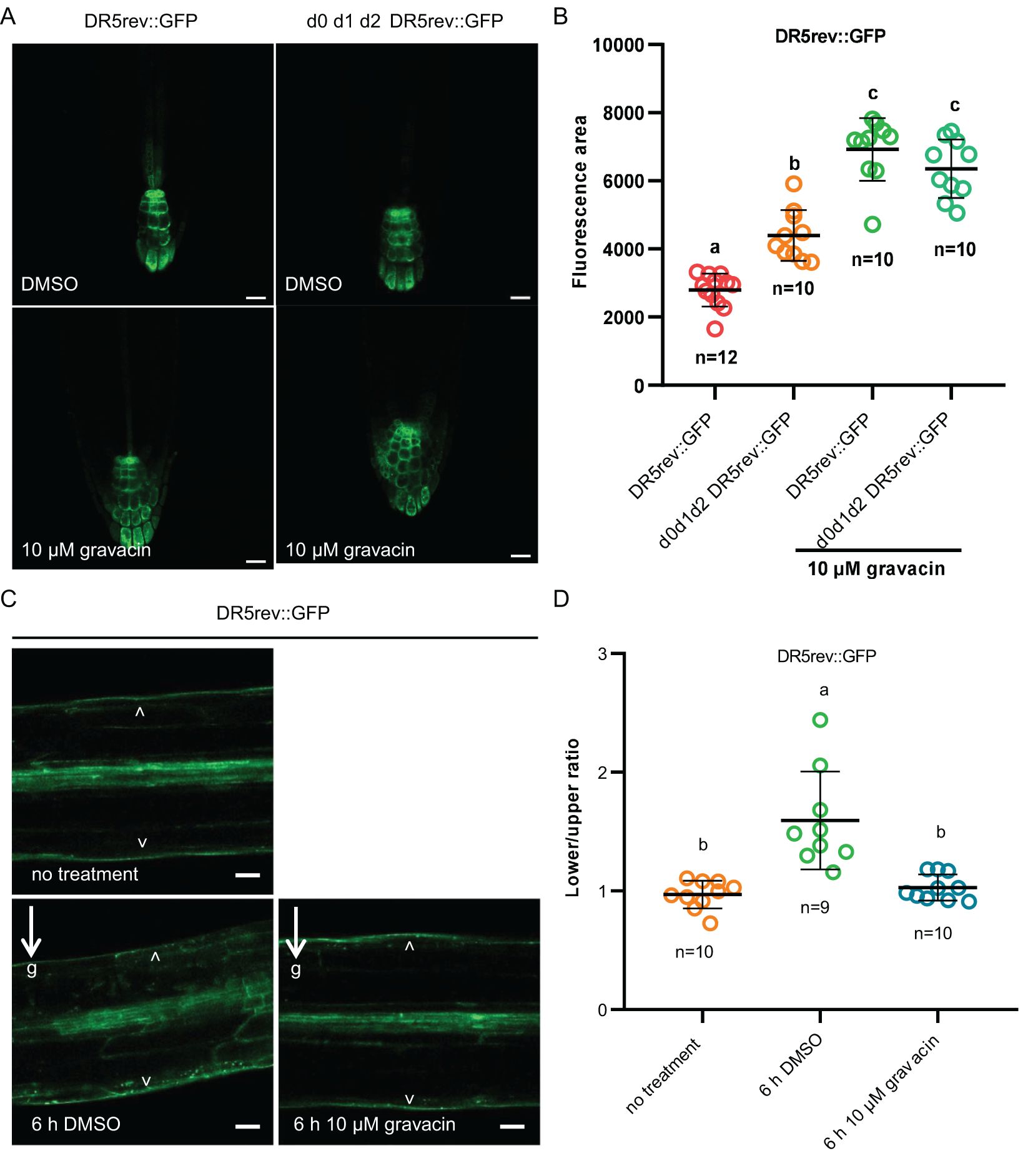
Figure 3. Gravacin interferes with the auxin response in plants. (A), 5-day-old DR5rev::GFP seedlings or d0 d1 d2 DR5rev::GFP seedlings grown on MS plates containing DMSO (control) or 10 µM gravacin were imaged via confocal laser scanning microscopy, n =10-12; scale bars, 20 µm. (B), The relative fluorescence intensities of DR5rev::GFP were measured in (A). P values were calculated via one-way ANOVA with Tukey’s multiple comparison test. The dots represent individual values, and the lines indicate the means ± SDs. Different letters represent significant differences. (C), 4-day-old etiolated DR5rev::GFP seedlings treated with DMSO (control) or 10 µM gravacin after 0 h and 6 h of gravity stimulation. The DR5 activity gradient along the gravity vector was clearly visible in DR5rev::GFP seedlings treated with DMSO after 6 h of gravity stimulation but not in the seedlings treated with gravacin. The arrowhead points to DR5rev::GFP accumulation. The arrows and “g” indicate the direction of gravity. Scale bars, 50 µm. (D), Quantification of DR5rev::GFP fluorescence intensity in 4-day-old etiolated DR5rev::GFP seedlings treated with DMSO (control) or 10 µM gravacin after 0 h and 6 h of gravistimulation. The DR5rev::GFP fluorescence intensity was compared between the outer side of the endodermal cells at the lower and upper sides of the hypocotyls. n = 9-10, respectively. P values were calculated via one-way ANOVA with Tukey’s multiple comparison test. The dots represent individual values, and the lines indicate the means ± SDs. Different letters represent significant differences.
To understand the mechanism underlying the gravitropic response of gravacin-treated hypocotyls, we examined the change in auxin distribution. Analysis of the DR5rev::GFP reporter revealed that prior to gravity stimulation, the DR5 signal was evenly distributed on both sides of the hypocotyl. After 6 hours of gravi-stimulation, a pronounced gradient of DR5 activity along the gravity vector at the lower side was observed. However, in hypocotyls treated with gravacin, this asymmetric distribution was less pronounced (Figures 3C, D). These results imply that gravacin may interfere with auxin transport by regulating the activity of the D6PK protein kinase.
It was reported previously that light-grown p35S::YFP-D6PK seedlings have more twisted roots than wild-type seedlings do, and dark-grown seedlings have severely shortened and thickened hypocotyls. To further investigate whether gravacin potentially disrupts auxin transport by regulating the activity of D6PK, we grew Col-0 seedlings and p35S::YFP-D6PK seedlings on MS media supplemented with gravacin or the solvent control DMSO. Phenotypic analysis revealed that p35S::YFP-D6PK seedlings, when treated with gravacin, presented a reduced root wavy growth phenotype under light conditions (Figures 4A–C), and their shortened hypocotyls, similarly, were partially restored by gravacin in darkness (Figures 4D, E). These results suggest that gravacin functions through D6PK proteins.
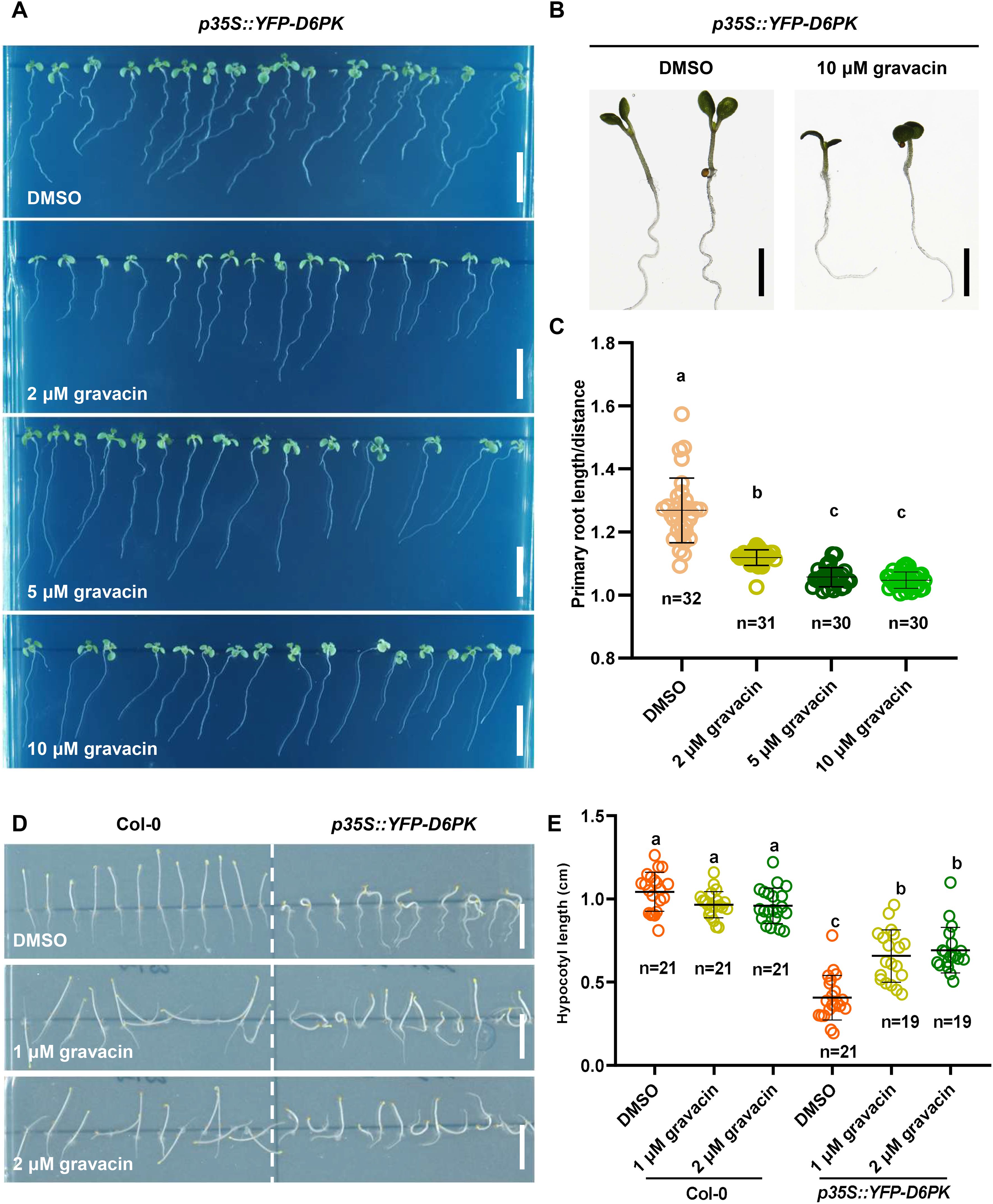
Figure 4. Gravacin can restore the phenotype of p35S::YFP-D6PK. (A), 4-day-old etiolated Col-0 and p35S::YFP-D6PK seedlings grown on MS media supplemented with gravacin. Scale bars, 1 cm. (B), The hypocotyl lengths of etiolated Col-0 seedlings and p35S::YFP-D6PK seedlings grown on MS media supplemented with gravacin were measured; n =19-21; P values were calculated via one-way ANOVA with Tukey’s multiple comparison test. The dots represent individual values, and the lines indicate the means ± SDs. Different letters represent significant differences. (C), Seven-day-old Col-0 and p35S::YFP-D6PK seedlings grown on MS media supplemented with gravacin. Scale bars, 1 cm. (D), Representative images of 7-day-old p35S::YFP-D6PK seedlings grown on plates supplemented with the indicated concentrations of gravacin were acquired with a stereomicroscope. Scale bars, 500 µm. (E), Root curvature of p35S::YFP-D6PK could be restored by gravacin. The distance: primary root length ratio was measured to indicate the degree of bending; n=30-32; P values were calculated via one-way ANOVA with Tukey’s multiple comparison test. The dots represent individual values, and the lines indicate the means ± SDs. Different letters represent significant differences.
PINs are phosphorylated at multiple serine/threonine residues by the protein kinase D6PK. To investigate whether gravacin influences D6PK activity in phosphorylating PINs, we conducted in vitro kinase assays. Our results revealed that His-tagged D6PK (His-D6PK) was capable of phosphorylating His-tagged PIN1 hydrophilic loops (His-PIN1-HL) (Tan et al., 2020a). However, the addition of gravacin to the reaction system with His-D6PK led to a decrease in the phosphorylation level of PIN1-HL compared with that in the reaction with D6PK alone (Figure 5A). This result implies that gravacin may target D6PK protein directly. To further validate this hypothesis, we utilized surface plasmon resonance (SPR) technology to measure the binding between D6PK and gravcin. The results revealed that gravacin indeed bound to the D6PK protein, with a kinetic dissociation constant (Kd) value of approximately 334 µM (Figures 5B, C). To gain insight into the inhibition mechanism of gravacin, we computed gravacin docking to the predicted Arabidopsis D6PK structure. The results indicated that the furan ring core of the gravacin was embedded within a hydrophobic cavity formed by Val124, Ala136, and Leu241, while its ortho-dichlorophenyl group located in another hydrophobic region formed by Phe188, Leu115, and Leu241, with both chlorine atoms of the ortho-dichlorophenyl group facing the solvent area. Additionally, the terminal carboxyl group of the compound might strengthen the binding stability to the target through a salt bridge interaction with the side chain amino group of Lys138, providing extra support for the overall stability of the binding conformation (Supplementary Figures 6A, B). In summary, the phenotypic analysis, in vitro kinase assays and SPR indicate that gravacin can directly binds to the D6PK, thereby inhibiting its activity and interfering with polar auxin transport.
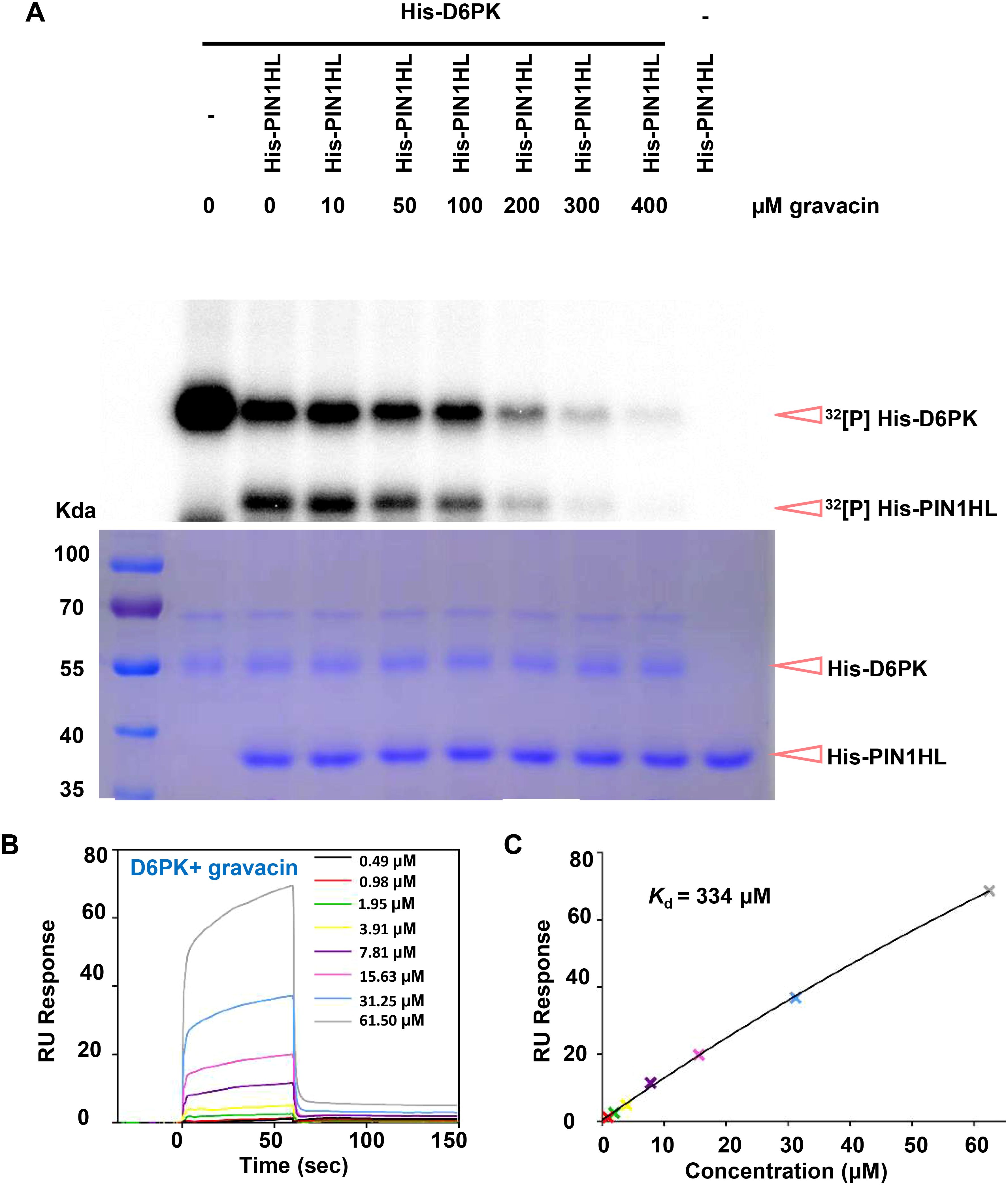
Figure 5. Gravacin can directly interact with D6PK and inhibit the phospholipid-D6PK-PIN cascade. (A), An in vitro kinase assay using [32P] ATP revealed that His-D6PK-induced phosphorylation of His-PIN1 was partially suppressed by gravacin in a dose-dependent manner. Reactions lacking the specified components (-) were used as controls. Top, recombinant proteins were separated by 10%SDS-PAGE after incubation in protein kinase buffer containing [32P] ATP. Phosphorylated His-PIN1 and His-D6PK was detected by autoradiography after gel electrophoresis. Bottom, recombinant His-PIN1, His-D6PK were detected by Coomassie brilliant blue (CBB) staining. (B), SPR signals for the association and dissociation of gravacin to D6PK. RU, resonance units. (C), Data from (B) were analyzed with a steady state affinity binding model. The resulting dissociation constant Kd was determined to be 334 μM.
This study provides compelling evidence that D6PK is a key target of gravacin. Notably, the phenotypes of gravacin-treated plants resembled those of d0 d1 d2 mutants, which presented defects in lateral root development and compromised gravitropism. Furthermore, transcriptome analysis and cytological experiments revealed that gravacin impacts the auxin pathway, aligning with D6PK’s capacity to modulate auxin transport. Additionally, the phenotype of p35S::YFP-D6PK can be suppressed by the application of exogenous gravacin, suggesting that gravacin can inhibit the D6PK function in vivo. Finally, an in vitro kinase assay directly revealed that gravacin can inhibit the D6PK-PIN cascade.
Our study demonstrated that the application of a high concentration of gravacin (100 µM) effectively inhibited the D6PK-PIN1 complex, and that the binding affinity (with a Kd value of approximately 334 µM) is relatively low. In contrast, a lower concentration of gravacin (2 µM) resulted in a complete loss of the gravitropic response in etiolated wild-type seedlings. These findings suggest: 1) D6PK is likely just one of several cellular targets of gravacin, suggesting that its interference with other cellular components is essential for the disruption of gravity perception and related signaling in seedlings. 2) Gravacin may modulate protein−protein interactions of unknown proteins associated with D6PK, thereby extending its influence on cellular processes beyond auxin transport. Moreover, we cannot exclude that additional target proteins besides D6PK/D6PKLs might exist for gravacin. For example, the shortened primary root is likely mediated by additional targets other than D6PK/D6PKL1/D6PKL2, based on the remained sensitivity of the d0 d1 d2 triple mutants to gravacin treatments.
Recent biochemical and structural studies have demonstrated that NPA is directly associated with PIN auxin transporters, thereby inhibiting their activity (Abas et al., 2021; Teale et al., 2021; Lam Ung et al., 2022; Su et al., 2022; Yang et al., 2022; Xia et al., 2023). Nevertheless, in addition to the direct interaction between NPA and PIN proteins, a variety of other protein targets have been suggested to mediate NPA function in planta. These include B-type ATP-binding cassette (ABC) transporters, TWISTED DWARF1 (TWD1, also known as FKBP42) and aminopeptidase 1 (APM1) (Teale and Palme, 2018). Recent studies have revealed that the ABC transporters ABCB19 and ABCB1 preferentially transport brassinosteroids rather than auxins. However, abcb19 mutants were reported to exhibit an overbent phenotype in the dark, suggesting a potential complex role of ABCB19 in the shoot gravitropic response as well as in auxin signaling. The potential role of these non-PIN proteins as targets for NPA has been previously discussed (Teale and Palme, 2018; Kong et al., 2022). Our research identified gravacin as a direct inhibitor of D6PK. This finding offers a precise tool for dissecting the molecular mechanisms of auxin distribution, differing from the mode of action of NPA, as gravacin does not appear to interact with PIN proteins.
The Arabidopsis thaliana (L.) mutants and transgenic plants were all of the Columbia-0 (Col-0) ecotype background. The transgenic marker lines pPIN1::PIN1-GFP (Benková et al., 2003), p35S::YFP-D6PK (Zourelidou et al., 2009), pARF19::NLS-GFP (Rademacher et al., 2012), and DR5rev::GFP (Friml et al., 2003) were previously reported. The mutant d0 d1 d2 in the Col-0 background was also reported previously. A list of all of the plant lines, including mutants, reporters and crosses, is provided in Supplementary Table S1. A list of the primers used to genotype the T-DNA mutants is provided in Supplementary Table S3. For phenotyping of seedlings or pharmacological experiments, surface-sterilized seeds (using 75% ethanol) were sown on Murashige and Skoog (MS) media supplemented with 1% (w/v) sucrose and 0.8% (w/v) phytoagar (pH 5.9), stratified at 4°C for 2 d and then grown in a growth chamber at 21°C with a long-day photoperiod (16–8 h light−8 h dark).
For long-term growth experiments, Arabidopsis seeds were sown on MS plates supplemented with the indicated chemicals. After stratification for 2 d at 4°C, the plates were moved to a growth chamber under the conditions described in the ‘Plant materials and growth conditions’ section. The phenotype of the roots was observed after an additional 7 or 11 days of growth. For lateral root induction assays, 6-day-old plants were grown on MS plates and subsequently transferred to MS plates supplemented with various concentrations of compounds. After 5 days, the number of lateral roots was counted. For PIN1::PIN1-GFP and ARF19::NLS-GFP observations, 6-day-old plants were transferred to MS plates supplemented with gravacin for 48 h. For DR5rev::GFP observation, DR5rev::GFP seedlings and d0 d1 d2 DR5rev::GFP seedlings were grown on MS plates with gravacin for 5 days. To observe the polarization of DR5rev::GFP, 5-day-old DR5rev::GFP seedlings grown on normal MS plates were transferred to MS plates containing gravacin and reoriented to 90°for 6 h. For the gravitropic response assay, 4-day-old etiolated seedlings were rotated 90° and subjected to gravity stimulation for a period of time before hypocotyl bending was observed. Afterwards, the samples were imaged via CLSM. A list of all of the reagents used in this study is provided in Supplementary Table S2.
Images were taken via a Zeiss LSM980 CLSM instrument equipped with a GaAsP detector (Zeiss). The manufacturer’s default settings (smart mode) were used for imaging proteins tagged with GFP (excitation, 488 nm; emission, 495–545 nm) or mCherry (excitation, 561 nm; emission, 580–650 nm). All of the images were obtained at an 8-bit depth with 2× line averaging. The images were analyzed and visualized with the Fiji program (Schindelin et al., 2012; Schneider et al., 2012).
For RNA-seq analysis, 6-day-old Arabidopsis seedlings were treated with 10 µM gravacin or DMSO for 3 h. Four-day-old etiolated Col-0 seedlings were grown constantly on MS media supplemented with 2 µM gravacin or DMSO. Each RNA sample was derived from a pool of Col-0 plants (weighing 100 mg) treated with the compounds. For subsequent analyses, three separate RNA samples for each condition were used Lists of DEGs were provided in Supplementary Tables S4–S7.
We utilized transcription-quantitative PCR (RT-qPCR) to investigate the transcript levels of IAA6, IAA19, AUX1, IAR3 and ABCB4 in Col-0 plants treated with 10 µM gravacin or DMSO, respectively. ACTIN7 (AT5G09810) was used as an internal reference. In detail, total RNA was extracted from the indicated tissues using Universal Plant Total RNA Isolation Kit (Vazyme). Then, we converted 1 µg of the RNA into cDNA using the Takara kit (RR036A). The resulting cDNA of the corresponding genes and ACTIN7 was analyzed using SYBR qPCR Master Mix (Tolobio, 22204) with a Bio-Rad CFX Connect Real-Time System. Expression values were normalized to the expression of ACTIN7 and the data are presented as the mean ± SD. from three biological replicates.
The recombinant proteins His-D6PK and His-PIN1-HL (Tan et al., 2020a) were expressed in the E. coli strain BL21 (DE3). Protein expression was induced by 0.5 mM isopropylβ-D-1-thiogalactopyranoside (IPTG) at 16°C for 20 h. Cell lysates were derived from approximately 200 mL of E. coli culture and subjected to affinity purification with HisSep Ni-NTA Agarose Resin (Yeasen) according to the manufacturer’s instructions. The concentration of the purified proteins was determined via a DeNovix DS-C device.
The procedure for in vitro phosphorylation assays followed a previously established protocol. Recombinant His-D6PK and His-PIN1-HL proteins were added together (5 µg of kinase and 10 µg of substrate in a 40 µL reaction system) in kinase reaction buffer (50 mM Tris-HCl, pH 7.5; 10 mM MgCl2; 10 mM ATP; and 1 mM DTT) in the presence of 5 µCi [32P]-ATP (NEG502A001MC; China Isotope & Radiation Corporation) and incubated in the presence of gravacin at 25°C for 60 minutes. The reactions were then terminated by the addition of SDS loading buffer. To terminate the reaction, SDS loading buffer was added. The samples were subsequently separated via 10% SDS−PAGE, developed overnight with a phosphor plate, and finally visualized via a Fujifilm FLA 3000 plus DAGE system.
A Biacore 8K system (Cytiva) was used to measure the binding affinity at 25°C with a flow rate of 30 µl min−1. Purified D6PK proteins were immobilized onto the series S CM5 sensor chips (Cytiva) by amine-coupling chemistry. Immobilized D6PK proteins were used to capture the gravacin at different concentrations. The running buffer contained HBS-EP (10 mM HEPES, pH 7.4, 150 mM NaCl, 3 mM EDTA, 0.05% surfactant P20) and 5% DMSO. Data were analyzed with the Biacore Insight Evaluation Software Version 3.0.12 using steady state affinity binding model.
The molecular docking experiment was conducted using AutoDock Vina (Version 1.2.6). Firstly, the D6PK protein structure was predicted by AlphaFold2. The compound structures were obtained by downloading from the public database PubChem and were constructed into three-dimensional structures with energy minimization using OpenBabel. The structures of the protein and compounds were further processed using AutoDockTools (version 1.5.7) to generate PDBQT format files for subsequent docking analysis by AutoDock Vina. During the docking simulation, the grid center was located at the ATP binding site of the D6PK protein to ensure effective contact between the small molecule and the target.
For seedling phenotyping, photographs were captured via a Sony A6000 camera equipped with a macro lens. The primary root length was then measured with ImageJ software, and the number of lateral roots was counted manually.
Most experiments were repeated at least three times independently, with similar results obtained. To measure primary root length, photographs were analyzed via ImageJ (https://imagej.nih.gov/ij/download.html) (Schneider et al., 2012). The fluorescence intensity of the CLSM images was quantified via Fiji (https://fiji.sc/) (Schindelin et al., 2012). Data visualization and statistical analysis were mostly performed with GraphPad Prism 9. Polar bar graphs were generated via Origin 8.0.
The sequence data from this study can be found in the Arabidopsis Genome Initiative or enBank/EMBL databases. The accession numbers are as follows: D6PK (AT5G55910), D6PKL1 (AT4G26610), D6PKL2 (AT5G47750), D6PKL3 (AT3G27580), PIN1 (AT1G73590), PIN2 (AT5G57090), PIN3 (AT1G70940), and PIN7 (AT1G23080), AUX1 (AT2G38120), IAA6 (AT1G52830), IAA19 (AT3G15540), IAR3 (AT1G51760), ABCB4 (AT2G47000). The RNA-seq data for this study are available online at the NCBI Sequence Read Archive (accession number: PRJNA1221602).
MC: Investigation, Methodology, Writing – original draft. ZY: Data curation, Investigation, Methodology, Writing – review & editing. YP: Data curation, Investigation, Methodology, Writing – review & editing. LHS: Investigation, Methodology, Writing – review & editing. XL: Funding acquisition, Supervision, Writing – review & editing. LFS: Funding acquisition, Methodology, Supervision, Writing – review & editing. ST: Conceptualization, Funding acquisition, Investigation, Supervision, Writing – review & editing, Writing – original draft.
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported by the National Natural Science Foundation of China (grants 32321001 to ST and LS, 32322041 to LS, grant 32471279 to XL), the Forestry Bureau of Anhui Province (AHLYJBGS-2024-01 to ST), the USTC Research Funds of the Double First-Class Initiative (YD9100002016), the Research Funds from the Center for Advanced Interdisciplinary Science and Biomedicine of IHM, the Division of Life Sciences and Medicine, the University of Science and Technology of China (QYPY20220012 to ST and QYPY20230034 to LS), the Strategic Priority Research Program of the Chinese Academy of Sciences (XDB37020103 to LS), the Natural Science Foundation of Anhui Province (2008085MC90 to XL, 2008085J15 to LS), the Double First-Class Initiative (YD9100002016 to ST, YD9100002004 to LS), the start-up funding from the University of Science and Technology of China (GG9100007007, KY9100000026, KY9100000051, and KJ2070000079 to ST), and the Fundamental Research Funds for the Central Universities (WK9100000021 to ST, WK9100000031 to LS).
We acknowledge Prof. Claus Schwechheimer (Technical University of Munich), Prof. Dolf Weijers (Wageningen University), and Prof. Mark Estelle (UCSD) for providing published Arabidopsis lines.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
The author(s) declare that no Generative AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2025.1563571/full#supplementary-material
Abas, L., Kolb, M., Stadlmann, J., Janacek, D. P., Lukic, K., Schwechheimer, C., et al. (2021). Naphthylphthalamic acid associates with and inhibits PIN auxin transporters. Proc. Natl. Acad. Sci. U. S. A. 118, e2020857118. doi: 10.1073/pnas.2020857118
Barbosa, I. C. R., Zourelidou, M., Willige, B. C., Weller, B., Schwechheimer, C. (2014). D6 PROTEIN KINASE activates auxin transport-dependent growth and PIN-FORMED phosphorylation at the plasma membrane. Dev. Cell 29, 674–685. doi: 10.1016/j.devcel.2014.05.006
Benková, E., Michniewicz, M., Sauer, M., Teichmann, T., Seifertová, D., Jürgens, G., et al. (2003). Local, efflux-dependent auxin gradients as a common module for plant organ formation. Cell 115, 591–602. doi: 10.1016/S0092-8674(03)00924-3
Bennett, M., Marchant, A., Green, H. G., May, S. T., Ward, S. P., Millner, P. A., et al. (1996). Arabidopsis AUX1 gene: a permease-like regulator of root gravitropism. Science 273, 14–17. doi: 10.1126/science.273.5277.948
Blázquez, M. A., Nelson, D. C., Weijers, D. (2020). Evolution of plant hormone response pathways. Annu. Rev. Plant Biol. 71, 327–353. doi: 10.1146/annurev-arplant-050718-100309
Carrillo-carrasco, V. P., Hernandez-garcia, J., Mutte, S. K., Weijers, D. (2023). The birth of a giant : evolutionary insights into the origin of auxin responses in plants. EMBO J. 42, e113018. doi: 10.15252/embj.2022113018
Cui, X., Wang, J., Li, K., Lv, B., Hou, B., Ding, Z. (2024). Protein post-translational modifications in auxin signaling. J. Genet. Genomics 51, 279–291. doi: 10.1016/j.jgg.2023.07.002
Dhonukshe, P., Grigoriev, I., Fischer, R., Tominaga, M., Robinson, D. G., Hašek, J., et al. (2008). Auxin transport inhibitors impair vesicle motility and actin cytoskeleton dynamics in diverse eukaryotes. Proc. Natl. Acad. Sci. U. S. A. 105, 4489–4494. doi: 10.1073/pnas.0711414105
Friml, J. (2022). Fourteen stations of auxin. Cold Spring Harb. Perspect. Biol. 14, a039859. doi: 10.1101/cshperspect.a039859
Friml, J., Vieten, A., Sauer, M., Weijers, D., Schwarz, H., Hamann, T., et al. (2003). Efflux-dependent auxin gradients establish the apical-basal axis of Arabidopsis. Nature 426, 147–153. doi: 10.1038/nature02085
Gao, Q., Yan, T., Zhang, Z. J., Liu, S. Y., Fang, X. D., Gao, D. M., et al. (2020). Casein kinase 1 regulates cytorhabdovirus replication and transcription by phosphorylating a phosphoprotein serine-rich motif. Plant Cell 32, 2878–2897. doi: 10.1105/tpc.20.00369
Geldner, N., Friml, J., Stierhof, Y. D., Jürgens, G., Palme, K. (2001). Auxin transport inhibitors block PIN1 cycling and vesicle trafficking. Nature 413, 425–428. doi: 10.1038/35096571
Jaillais, Y., Fobis-Loisy, I., Miège, C., Rollin, C., Gaude, T. (2006). AtSNX1 defines an endosome for auxin-carrier trafficking in Arabidopsis. Nature 443, 106–109. doi: 10.1038/nature05046
Kania, U., Lu, Q., Hicks, G. R., Nerinckx, W., Mishev, K., Cherfils, J., et al. (2018). The inhibitor endosidin 4 targets SEC7 domain-type ARF GTPase exchange factors and interferes with subcellular trafficking in eukaryotes. Plant Cell 30, 2553–2572. doi: 10.1105/tpc.18.00127
Kim, J. Y., Henrichs, S., Bailly, A., Vincenzetti, V., Sovero, V., Mancuso, S., et al. (2010). Identification of an ABCB/P-glycoprotein-specific inhibitor of auxin transport by chemical genomics. J. Biol. Chem. 285, 23309–23317. doi: 10.1074/jbc.M110.105981
Kong, M., Liu, X., Sun, L., Tan, S. (2022). Molecular mechanisms of N−1 −naphthylphthalamic acid, a chemical tool in plant biology and agriculture. Mol. Hortic. 2, 22. doi: 10.1186/s43897-022-00043-y
Lam Ung, K., Winkler, M., Schulz, L., Kolb, M., Janacek, D. P., Dedic, E., et al. (2022). Structures and mechanism of the plant PIN-FORMED auxin transporter. Nature 609, 605–610. doi: 10.1038/s41586-022-04883-y
Luschnig, C., Friml, J. (2024). Over 25 years of decrypting PIN-mediated plant development. Nat. Commun. 15, 9904. doi: 10.1038/s41467-024-54240-y
Naramoto, S., Otegui, M. S., Kutsuna, N., de Rycke, R., Dainobu, T., Karampelias, M., et al. (2014). Insights into the localization and function of the membrane trafficking regulator GNOM ARF-GEF at the Golgi apparatus in Arabidopsis. Plant Cell 26, 3062–3076. doi: 10.1105/tpc.114.125880
Peng, Y., Ji, K., Mao, Y., Wang, Y., Korbei, B., Luschnig, C., et al. (2024). Polarly localized Bro1 domain proteins regulate PIN-FORMED abundance and root gravitropic growth in Arabidopsis. Commun. Biol. 7, 1085. doi: 10.1038/s42003-024-06747-9
Rademacher, E. H., Lokerse, A. S., Schlereth, A., Llavata-Peris, C. I., Bayer, M., Kientz, M., et al. (2012). Different auxin response machineries control distinct cell fates in the early plant embryo. Dev. Cell 22, 211–222. doi: 10.1016/j.devcel.2011.10.026
Rojas-Pierce, M., Titapiwatanakun, B., Sohn, E. J., Fang, F., Larive, C. K., Blakeslee, J., et al. (2007). Arabidopsis P-glycoprotein19 participates in the inhibition of gravitropism by gravacin. Chem. Biol. 14, 1366–1376. doi: 10.1016/j.chembiol.2007.10.014
Schindelin, J., Arganda-Carreras, I., Frise, E., Kaynig, V., Longair, M., Pietzsch, T., et al. (2012). Fiji: An open-source platform for biological-image analysis. Nat. Methods 9, 676–682. doi: 10.1038/nmeth.2019
Schneider, C. A., Rasband, W. S., Eliceiri, K. W. (2012). NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 9, 671–675. doi: 10.1038/nmeth.2089
Semeradova, H., Montesinos, J. C., Benkova, E. (2020). All roads lead to auxin: Post-translational regulation of auxin transport by multiple hormonal pathways. Plant Commun. 1, 100048. doi: 10.1016/j.xplc.2020.100048
Su, N., Zhu, A., Tao, X., Ding, Z. J., Chang, S., Ye, F., et al. (2022). Structures and mechanisms of the Arabidopsis auxin transporter PIN3. Nature 609, 616–621. doi: 10.1038/s41586-022-05142-w
Tan, S., Di Donato, M., Glanc, M., Zhang, X., Klíma, P., Liu, J., et al. (2020b). Non-steroidal anti-inflammatory drugs target TWISTED DWARF1-regulated actin dynamics and auxin transport-mediated plant development. Cell Rep. 33, 108463. doi: 10.1016/j.celrep.2020.108463
Tan, S., Luschnig, C., Friml, J. (2021). Pho-view of auxin: Reversible protein phosphorylation in auxin biosynthesis, transport and signalling. Mol. Plant 14, 151–165. doi: 10.1016/j.molp.2020.11.004
Tan, S., Zhang, X., Kong, W., Yang, X.-L., Molnár, G., Vondráková, Z., et al. (2020a). The lipid code-dependent phosphoswitch PDK1–D6PK activates PIN-mediated auxin efflux in Arabidopsis. Nat. Plants 6, 556–569. doi: 10.1038/s41477-020-0648-9
Teale, W., Palme, K. (2018). Naphthylphthalamic acid and the mechanism of polar auxin transport. J. Exp. Bot. 69, 303–312. doi: 10.1093/jxb/erx323
Teale, W. D., Pasternak, T., Dal Bosco, C., Dovzhenko, A., Kratzat, K., Bildl, W., et al. (2021). Flavonol-mediated stabilization of PIN efflux complexes regulates polar auxin transport. EMBO J. 40, e104416. doi: 10.15252/embj.2020104416
Ung, K. L., Schulz, L., Pedersen, B. P., Stokes, D. L., Hammes, U. Z. (2023). Substrate recognition and transport mechanism of the PIN-FORMED auxin exporters. Trends Biochem. Sci. 48, 937–948. doi: 10.1016/j.tibs.2023.07.006
Vanneste, S., Friml, J. (2009). Auxin: A trigger for change in plant development. Cell 136, 1005–1016. doi: 10.1016/j.cell.2009.03.001
Wang, X., Chen, M., Li, J., Kong, M., Tan, S. (2024). The SCOOP-MIK2 immune pathway modulates Arabidopsis root growth and development by regulating PIN-FORMED abundance and auxin transport. Plant J. 120, 318–334. doi: 10.1111/tpj.v120.1
Wei, H., Zhu, H., Ying, W., Janssens, H., Kvasnica, M., Ma, Q., et al. (2024). Structural insights into brassinosteroid export mediated by the Arabidopsis ABC transporter. Plant Commun. Adv. Access. 6, 101181. doi: 10.1016/j.xplc.2024.101181
Weller, B., Zourelidou, M., Frank, L., Barbosa, I. C. R., Fastner, A., Richter, S., et al. (2017). Dynamic PIN-FORMED auxin efflux carrier phosphorylation at the plasma membrane controls auxin efflux-dependent growth. Proc. Natl. Acad. Sci. 114, E887–E896. doi: 10.1073/pnas.1614380114
Xia, J., Kong, M., Yang, Z., Sun, L., Peng, Y., Mao, Y., et al. (2023). Chemical inhibition of Arabidopsis PIN-FORMED auxin transporters by the anti-inflammatory drug naproxen. Plant Commun. 6, 100632. doi: 10.1016/j.xplc.2023.100632
Yang, Z., Xia, J., Hong, J., Zhang, C., Wei, H., Ying, W., et al. (2022). Structural insights into auxin recognition and efflux by Arabidopsis PIN1. Nature 609, 611–615. doi: 10.1038/s41586-022-05143-9
Ying, W., Wang, Y., Wei, H., Luo, Y., Ma, Q., Zhu, H., et al. (2024). Structure and function of the Arabidopsis ABC transporter ABCB19 in brassinosteroid export. Science 383, eadj4591. doi: 10.1126/science.adj4591
Yu, Z., Zhang, F., Friml, J., Ding, Z. (2022). Auxin signaling: Research advances over the past 30 years. J. Integr. Plant Biol. 64, 371–392. doi: 10.1111/jipb.13225
Zhang, W. J., Zhou, Y., Zhang, Y., Su, Y. H., Xu, T. (2023). Protein phosphorylation: A molecular switch in plant signaling. Cell Rep. 42, 112729. doi: 10.1016/j.celrep.2023.112729
Zou, M., Ren, H., Li, J. (2019). An auxin transport inhibitor targets Villin-mediated actin dynamics to regulate polar auxin transport. Plant Physiol. 181, 161–178. doi: 10.1104/pp.19.00064
Zourelidou, M., Absmanner, B., Weller, B., Barbosa, I. C. R., Willige, B. C., Fastner, A., et al. (2014). Auxin efflux by PIN-FORMED proteins is activated by two different protein kinases, D6 PROTEIN KINASE and PINOID. Elife 3, e02860. doi: 10.7554/eLife.02860.035
Keywords: auxin transport, gravacin, D6PK, PIN, arabidopsis
Citation: Chen M, Yang Z, Peng Y, Sun L, Liu X, Sun L and Tan S (2025) Gravacin as an inhibitor of the auxin transport-activating protein kinase D6PK in Arabidopsis. Front. Plant Sci. 16:1563571. doi: 10.3389/fpls.2025.1563571
Received: 20 January 2025; Accepted: 17 February 2025;
Published: 12 March 2025.
Edited by:
Kai Jiang, Yunnan University, ChinaReviewed by:
Da Cao, Ghent University, BelgiumCopyright © 2025 Chen, Yang, Peng, Sun, Liu, Sun and Tan. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xin Liu, bHgwMjNAdXN0Yy5lZHUuY24=; Linfeng Sun, c3VubGYxN0B1c3RjLmVkdS5jbg==; Shutang Tan, c3R0YW5AdXN0Yy5lZHUuY24=
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.