 Avinash Jha
Avinash Jha D. K. Jayswal
D. K. Jayswal Deep Shikha1
Deep Shikha1 Adyant Kumar
Adyant Kumar
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Plant Sci. , 24 February 2025
Sec. Plant Nutrition
Volume 16 - 2025 | https://doi.org/10.3389/fpls.2025.1519673
Increasing the nutritional value of any crop plant through various Conventional or non-Conventional methods is known as Biofortification. Deficiency of proteins, essential amino acids, vitamins and minerals leads to ailing health and increased vulnerability to various diseases, which in turn lead to uncountable and unpredicted loss in Gross Domestic Product leading to poor economic growth of the country. It is forthcoming and cost-effective approach that will provide a balance of micronutrient deficiency among the people of developing & underdeveloped nations not having the availability to diverse nutritional access. The Biofortified varieties not only provide required calories but also essential nutrients needed for proper growth and development of an individual. It is advantageous in combating malnutrition and hidden hunger by enhancing the micronutrient content of commonly consumed fruits. By increasing essential vitamins, minerals, and beneficial compounds through methods like traditional breeding, genetic engineering, and agronomic practices, biofortified fruits provide a sustainable solution to address deficiencies in regions with limited access to diverse foods. For instance, mango, guava, papaya, and citrus have been improved to offer higher levels of nutrients such as iron, zinc, vitamin C, and beta-carotene. This makes biofortified fruits a cost-effective way to enhance nutrition, particularly for vulnerable populations, helping to reduce the risks associated with hidden hunger and malnutrition. One of the important targets of United Nation is to provide fortified food enriched with important minerals to the targeted undernourished population in different parts of the world. The lack of essential nutrients, notably minerals such as iron (Fe), zinc (Zn), and vitamin A, is one of the main causes of “hidden hunger”, especially in underdeveloped nations. The review covers most of the important aspects of Biofortification in important fruit crops.
In the present scenario food security is major concern for most of the countries due to non-predictive Climate Change and increasing Population. Fortification of various crop plants in present and future scenario will be a major player in overcoming global hunger. According to United Nation, malnutrition & hidden hunger is severely affecting the people of under developed and developing nations like Africa and Southern Asia. Out of the total Infants deaths, maximum is attributed by these two factors only. Some of the Developed countries like Canada, USA & Europe also face deficiencies of Iron, Zinc, Iodine, and Vitamin A & D. Micronutrient malnutrition affects over 2 billion people around the world and causes serious health problems due to a lack of important vitamins and minerals like iodine, iron, zinc, and vitamin A. These nutrients are needed in small amounts for the body to function properly, but poor diet and problems with absorption, along with factors like food insecurity and poor healthcare, lead to deficiencies, especially in pregnant women and children. Iron deficiency, which causes anemia, affects more than 2 billion people (Benoist et al., 2008), while zinc deficiency affects about 1.1 billion people, leading to weakened immunity and growth problems (Frassinetti et al., 2006). Solving micronutrient deficiencies is important for improving health, especially in developing countries.
One of the major goals of World Health Organization (WHO) and Consultative Group on International Agricultural Research (CGIAR) is focusing on development of high yielding Biofortified crops. Considering the global scenario more than two billion people suffer micronutrient deficiency, 820 million people are undernourished, out of which 149 million children’s below 5 years of age are affected, and the condition is more serious in South Asian Countries. Coming on to the Indian scenario where 21.9% of population lives in extreme poverty hidden hunger and malnutrition is becoming serious day by day. In India 15.2% of people are undernourished, 38.4% of the children less than five years of age are stunted, 21.0% are wasted and 35.7% of the children are under-weight, 58.4% of the children below 5 years, 53% of the adult women and 22.7% of adult men are affected due to anemia. In countries like India a loss of 12 billion dollars is estimated every year due to these deficiencies in direct or indirect forms (Yadava et al., 2020). The high global consumption of bananas and mangoes highlights their significant potential for biofortification, which can play a crucial role in addressing micronutrient deficiencies. In Uganda, where banana consumption is exceptionally high, biofortifying bananas with essential nutrients like vitamin A, iron, and zinc could greatly enhance public health by improving nutrition in populations heavily reliant on bananas. Similarly, with mangoes being the second most consumed tropical fruit globally, biofortification can increase their nutritional value, helping to combat common deficiencies in regions where they are a dietary staple. As countries with high fruit consumption, such as India, the U.S., and Dominica, continue to rely on bananas and mangoes, biofortifying these widely consumed fruits offers an efficient, sustainable approach to combating malnutrition and improving overall health (FAO, 2020; World Population Review, 2023; Food and Agriculture Organization, 2023).
Malnutrition, particularly in developing countries, is not only driven by a lack of macronutrients—carbohydrates, proteins, and fats—but also by micronutrient deficiencies. While macronutrients are essential for providing energy and supporting body functions, micronutrient deficiencies, despite an adequate food supply, can lead to serious health problems. People in these regions often rely on staple foods like cereals and beans, which provide sufficient macronutrients but are low in essential vitamins and minerals. This imbalance exacerbates malnutrition, especially for vulnerable groups like children and pregnant women. Biofortification, which enhances the nutrient content of staple crops, can help improve the micronutrient intake without altering the availability of macronutrients. This strategy can address hidden hunger by improving the overall nutritional quality of the diet and reducing reliance on expensive or diverse foods (Kiran et al., 2022).
Nutrient movement and distribution within fruit trees are key to their growth, fruit production, and quality. Macronutrients like nitrogen (N), phosphorus (P), potassium (K), and calcium (Ca) are essential for these processes. While most macronutrients can move throughout the plant, calcium is less mobile, and its movement depends on water flow and areas of the plant with higher transpiration. Research has shown that calcium moves mainly through the xylem, and its distribution is influenced by the plant’s need and water flow. Nutrients from older leaves, pruning, and decomposing plant material also help maintain nutrient levels. Effective management of these nutrients, especially calcium, is important for fruit quality, as deficiencies can affect development. Ongoing studies focus on improving how nutrients are absorbed and distributed to boost tree health and fruit yield. New technologies to track nutrient flow and better nutrient management strategies aim to make fruit production more efficient and environmentally friendly (Kalcsits et al., 2020).
The work has been practiced in most of crops but this article mainly deals with Biofortification of the horticultural crops like Pomegranate, Banana, Mango, Grape, Papaya, Apple, Pear etc.
The various Biofortification strategies as presented in the Figure 1 include Agronomic, Conventional Breeding approaches and non-conventional breeding approaches like Genetic Engineering. Biofortification of fruit crops can be done through the procedure as explained in the Figure 2. All the strategies have been explained
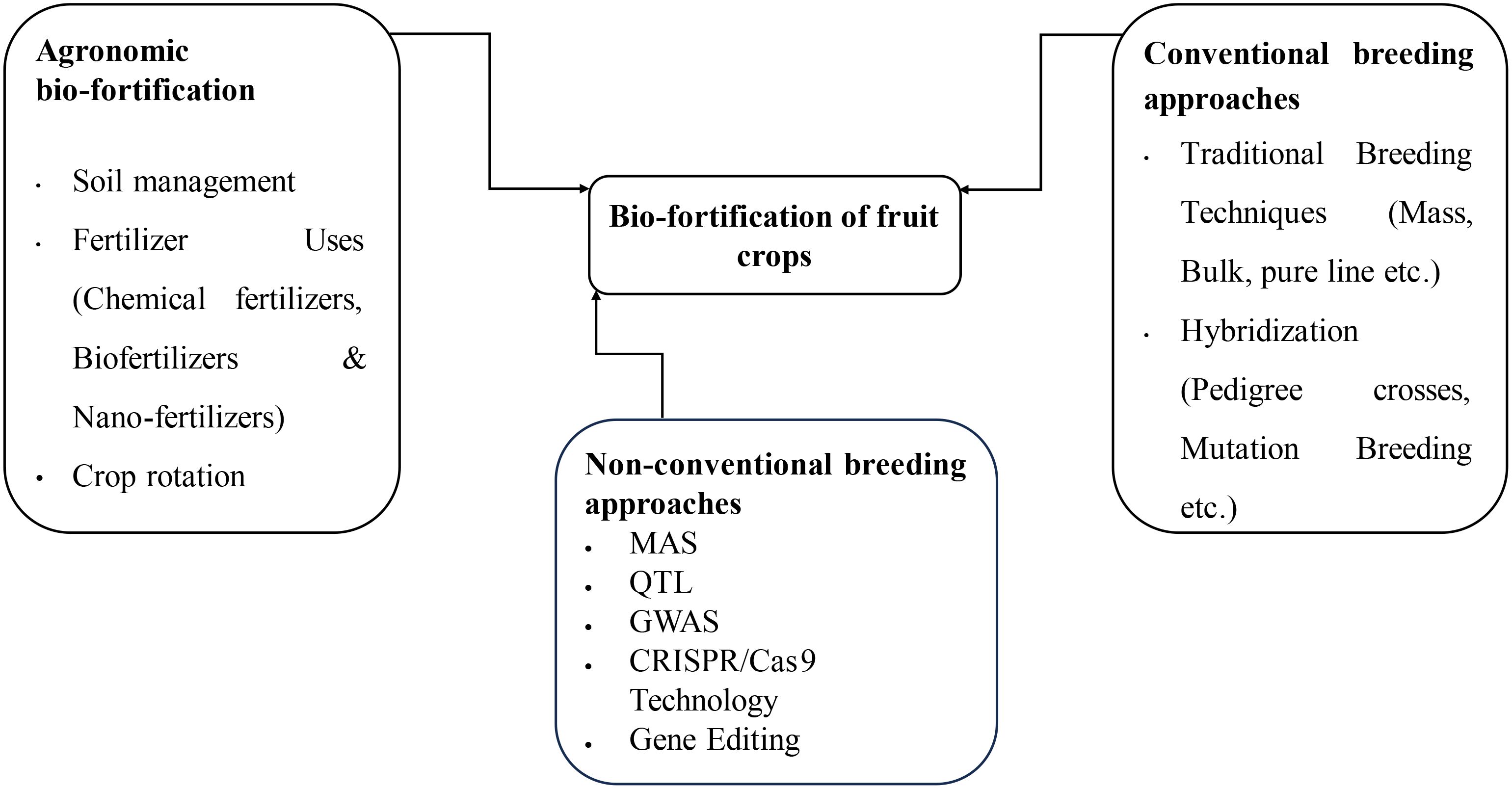
Figure 1. Methods of bio-fortification of fruit crops.
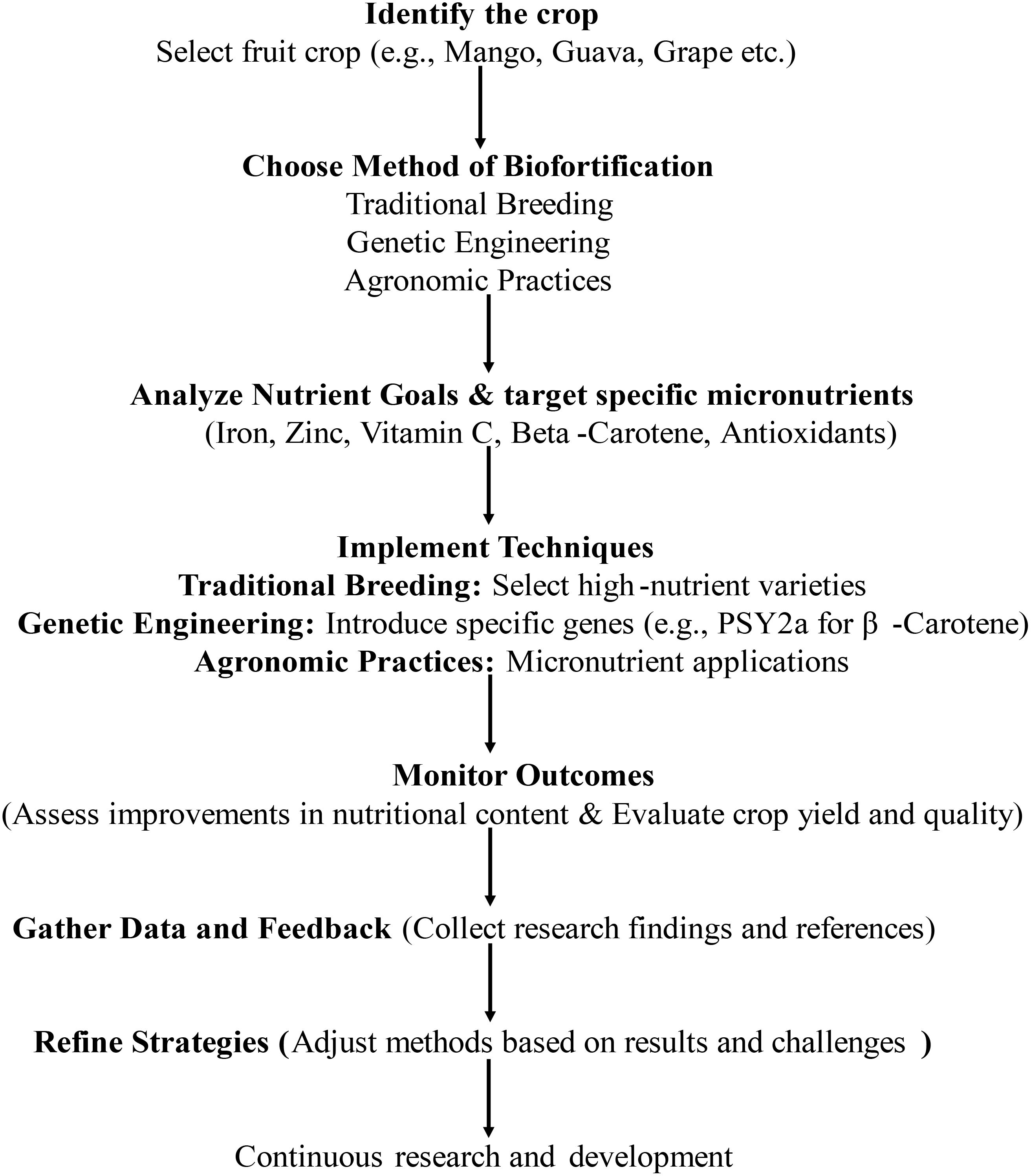
Figure 2. Procedure for bio-fortification of fruit crops.
Increasing the nutrient content in commercially adapted varieties through various agronomic means like fertigation, foliar application or through normal fertilizer application without affecting the threshold limit of other nutrients is known as Agronomic Biofortification. It is the fast safe and economical effective way of Biofortification with minimum efforts to enrich crops along with public acceptance. Most of the present-day experiments are based on this practice however, commercial exploitation has certain limitations like Low nutrient use efficiency (1-5%) for micronutrients like iron, zinc, copper, etc. which limit the uptake of applied micronutrients by plants. However, using heavy dosages of these micronutrients over a long period of time can lead to toxicity as most of the micronutrients remain in soil for longer duration of time. Some of the uncontrolled factors like changing weather conditions and Geographic variation in soil micronutrient deficiencies limit its usage up to some extent.
Agronomic biofortification focuses on improving the nutritional content of crops through farming practices. For example, pomegranate has been enhanced through agronomic techniques to increase iron, zinc, and vitamin C levels (Yadava et al., 2020). Similarly, grapes have been improved through agronomy to boost polyphenols, anthocyanins, resveratrol, and vitamins C and K (Di Lorenzo et al., 2019). Watermelon has also been biofortified using agronomic practices to increase lycopene, protein, and amino acids (Perkins et al., 2001). Additionally, crops like citrus, pear, and strawberry have been improved through agronomic methods as explained in Table 1 to increase vitamins, minerals, and antioxidants (Salonia et al., 2020; Zhang et al., 2021; Singh et al., 2022). These techniques are essential in enhancing the nutritional quality of fruit crops.
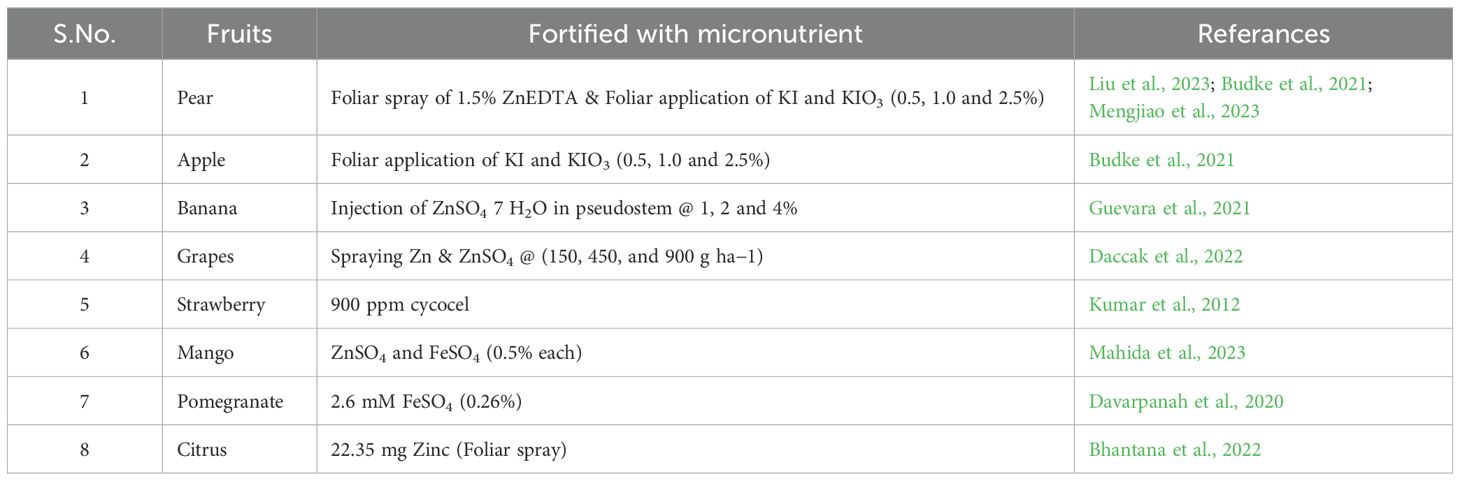
Table 1. Nutritional fortification of important fruit crops.
Plants primarily absorb nutrients through their roots, which transport them via the xylem. Understanding the availability of micronutrients in soil and their uptake by plants is crucial for enhancing plant nutrition, as confirmed by Bhardwaj et al. (2022). Different plant types employ distinct strategies for nutrient absorption. For instance, non-grassy plants (dicots) enhance iron (Fe) solubility in low-iron conditions, while grassy plants (monocots) utilize compounds called phytosiderophores to capture iron from the soil (Hell and Stephan, 2003).
Various transport proteins, such as those in the ZIP family, facilitate the uptake of key nutrients like zinc (Zn) and copper (Cu) (Eckhardt et al., 2001). When nutrients flow into the shoots, they promote root absorption and help regulate nutrient levels within the plant. After entering the xylem, nutrients are pulled up to the leaves, where they first act as sinks (nutrient importers) and later as sources (nutrient exporters) once photosynthesis exceeds the leaves’ energy needs (White and Broadley, 2009).
Nutrient movement differs within the plant: roots send nutrients to leaves via the xylem, while leaves transport them to fruits and seeds through the phloem. Some nutrients, like selenium (Se) and magnesium (Mg), move quickly through the phloem, whereas others, like iron and zinc, exhibit limited mobility. This limitation makes fruits and grains less nutrient-rich compared to leafy vegetables (White and Broadley, 2003). Foliar applications-spraying nutrients on leaves-can efficiently deliver micronutrients directly to the phloem, making this method valuable for biofortification.
Grains receive nutrients through a single vascular bundle, primarily storing iron and zinc in the embryo and aleurone layer. Specific proteins aid in transporting and storing these nutrients (Zhang et al., 2007). Enhancing beneficial substances, such as vitamins, while reducing harmful compounds called antinutrients (like phytates) can significantly improve nutrient absorption in the human diet. These strategies are supported by the findings of Bhardwaj et al. (2022) and can greatly assist in biofortification efforts, making crops more nutritious (Sancenon et al., 2003).
Biofertilizers are microbial products made from beneficial microorganisms that boost the growth and productivity of plants (Sahoo et al., 2013). These organisms are known as plant growth-promoting microorganisms and are valuable because they are inexpensive, easy to produce, and environmentally friendly. They enhance the supply and availability of nutrients in the soil, leading to better nutrient content in plants (Sahoo et al., 2013).
For instance, zinc (Zn) biofortification in crops like corn is aided by cyanobacteria such as Azotobacter and Anabaena, as well as the bacterium Bacillus aryabhattai (Prasanna et al., 2015). Similar benefits have been observed in wheat and soybeans (Ramesh et al., 2014). Using microbial interventions is recommended to address zinc deficiencies in soils (Dotaniya et al., 2016).
Arbuscular mycorrhizal fungi, like Rhizophagus irregularis, improve root development and help plants absorb essential nutrients like phosphorus (P), nitrogen (N), zinc (Zn), copper (Cu), manganese (Mn), and iron (Fe). This fungus has been shown to increase minerals in medicinal plants such as Mentha pulegium and Petroselinum hortense (Gashgari et al., 2020). Additionally, species of Pseudomonas, including Pseudomonas chlororaphis, enhance iron uptake and overall plant health (Sharma and Johri, 2003).
However, there are challenges in using biofertilizers. Identifying the right microorganisms for specific crops can be difficult, and biofertilizers may have a short shelf life due to variations in different environments. Improper storage and application can also hinder the benefits of biofortification.
Given these advantages, biofertilizers can be effectively applied to fruit crops such as strawberries and citrus fruits. For instance, using Rhizophagus irregularis can enhance root development and nutrient uptake, leading to better yields and nutrient-rich fruit in strawberries (Flores-Felix et al., 2015). Additionally, applying Pseudomonas chlororaphis could improve iron absorption in citrus trees, boosting fruit quality. Similarly, beneficial cyanobacteria can help address zinc deficiencies in strawberries, ensuring healthier plants and more flavorful berries. By leveraging these microbial interventions, fruit growers can improve growth and productivity while facing challenges posed by climate change and soil nutrient deficiencies.
Nano-fertilizers are fertilizers where the active ingredients are reduced to tiny particles, typically ranging from 1 to 100 nanometers. This size allows them to be effectively dispersed or encapsulated in host materials (Bhardwaj et al., 2022). Recently, nano biofortification has gained traction as a successful method to improve nutrition in crops like wheat (Khan et al., 2021). These tailored fertilizers have the potential to revolutionize agriculture by offering safe, targeted, and easy-to-apply nutrient delivery systems (Dimkpa and Bindraban, 2016; Manjunatha et al., 2016; Rai et al., 2018).
The high surface area of nano-fertilizers makes them more efficient and allows for slow-release nutrient supply to crops. They can include various nanoparticles, such as zinc oxide, silica, iron, and titanium dioxide, as well as core-shell quantum dots containing essential nutrients like Zn, Fe, Mn, and Cu (Prasad et al., 2015; Bhardwaj et al., 2022). The effectiveness of these nano-fertilizers depends on factors such as the plant species, size, concentration, and composition of the nanomaterials (Thakur et al., 2018). They are specifically engineered to address nutrient deficiencies, ensuring that plants not only grow but also accumulate essential nutrients in their edible parts (Li et al., 2016).
For example, a study by Dapkekar et al. (2018) showed that durum wheat treated with zinc complexed chitosan nanoparticles (Zn-CNP) increased grain zinc content significantly, even at lower concentrations compared to traditional zinc sulfate. Similarly, Hussain et al. (2021) found that foliar application of zinc oxide nanofertilizer boosted zinc levels in wheat plants. The use of chitosan-complexed zinc nano-fertilizers also led to higher grain zinc content and specific nutrient accumulation in seeds (Doolette et al., 2020).
Nanotechnology can be effectively applied to fruit crops to enhance their nutritional quality. By utilizing tiny particles known as nanoparticles, we can improve the ability of fruit plants to absorb essential nutrients. These nanoparticles can be applied to the seeds or sprayed on the leaves, facilitating better nutrient uptake.
For example, nanoparticles can induce a movement called “Brownian Motion,” which helps distribute nutrients more efficiently within the fruit plants. The smaller the particles, the more effectively the plants can absorb nutrients through their roots. This approach has shown promising results in increasing the nutrient levels in fruit crops, making them healthier for consumption (Jiang et al., 2018; Torre-Roche et al., 2020).
Further studies have demonstrated that using nano-iron and nano-manganese fertilizers can improve nutrient uptake and plant growth. For instance, applying nano-Fe2O3 stimulated root growth better than conventional iron forms, while nano-Mn showed increased manganese accumulation in seeds and enhanced overall plant growth (Pradhan et al., 2013; Dimkpa et al., 2018). These advancements in nano fertilizers can also be effectively applied to fruit crops. For example, using zinc and iron nano fertilizers can enhance nutrient uptake in fruit trees like apple and citrus, leading to improved fruit quality and yield. The slow-release properties of these fertilizers can ensure that fruit crops receive consistent nutrition throughout their growing season, ultimately increasing their resistance to stress and enhancing flavor and nutritional content. By adopting nano-fertilizers, fruit growers can address specific nutrient deficiencies while promoting healthier, more productive crops.
Breeding interference had played a significant role in Biofortification of important crops, where several traits are identified based on multiple environment stability assessment Models affecting the fortification pathway. Various breeding approaches like exploring variability present in natural population via Mass, Bulk and pure line selection or creation of variability through pedigree crosses or through induced, spontaneous or natural mutations in a population by targeted exploitation of mechanism favoring Biofortification, as it has been used to create grape varieties with more nutrients. By selecting specific plants, breeders aim to increase the levels of important compounds like polyphenols and anthocyanins in grape berries. These compounds help to improve the antioxidant properties and health benefits of grapes (Di Lorenzo et al., 2019). The transfer of required gene favoring Biofortification pathways through Backcross breeding will serve as an important tool in present and future times to enhance the existing varieties and lines of commercially important hybrids with increased nutrient content without hampering its original yield. The use of wild relatives of crop plant for transferring required gene favoring Biofortification is yet to be used at larger scale as it is a time-consuming process and due to linkage drag of unwanted genes associated with wild species. Sometimes desirable genes of interest rarely express due to gene interactions and other uncontrolled factors.
Various fruit varieties have been developed through conventional breeding to improve their nutritional value. In Uganda, HarvestPlus developed banana varieties like Apantu, Bira, Pelipita, Lai, and To’o (Table 2), which are rich in Vitamin A. In India, IARI developed several mango varieties such as Amarpali, Pusa Arunima, Pusa Surya, Pusa Pratibha, Pusa Peetamber, Pusa Lalima, and Pusa Shreshth, all high in beta-carotene and Vitamin C (Table 2). The Ataulfo mango, known for its beta-carotene and Vitamin C content, was developed by the USDA Agricultural Research Service in Mexico. Additionally, IARI in India developed the Pusa Navrang grape, which is packed with antioxidants.
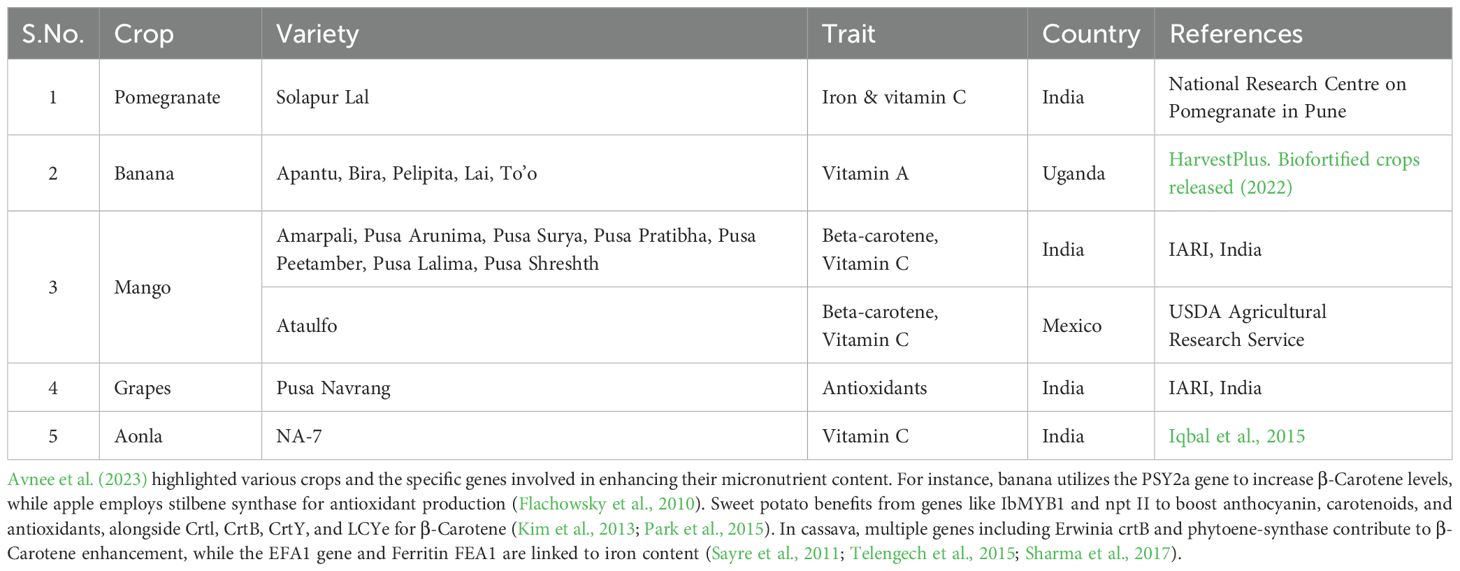
Table 2. Biofortified Varieties developed in important fruit crops.
To combat micronutrient malnutrition sustainably, dietary diversification is key. This approach encourages consuming a wide variety of healthy foods, including fruits, vegetables, and different grains. Success depends on factors like affordability, dietary habits, accessibility, and bioavailability of nutrients. Raising awareness about nutrition through media can help promote diverse diets.
Enhancing dietary diversity can be achieved by improving agricultural production, offering a greater variety of foods, and promoting better food processing and preparation methods. Additionally, combining dietary diversification with supplementation (like tablets) and fortification can effectively address specific nutrient deficiencies in the population (Lee et al., 2011).
Microbial biofortification is a useful way to improve the nutritional quality of crops by using helpful microorganisms like Plant Growth-Promoting Rhizobacteria (PGPR) and Arbuscular Mycorrhizal Fungi (AMF). PGPR can boost the yield and nutrient content of crops, especially legumes, by helping them fix nitrogen, which is important for their growth. AMF work with plant roots to help them absorb essential nutrients like phosphorus and micronutrients such as selenium. For example, garlic treated with Glomus intraradices has shown a tenfold increase in selenium content, showing how effective these microbes can be in improving nutrition (Roriz et al., 2020; Larsen et al., 2006).
Additionally, specific types of microbes have been successfully used on different crops to increase their micronutrient levels. For instance, using zinc-solubilizing bacteria has raised the zinc content in tomatoes from 2.06 mg to 2.87 mg per 100 g. This method not only makes crops healthier but also reduces the need for chemical fertilizers, which is better for the environment. However, there are challenges, such as not knowing enough about the micronutrient levels in seeds and how climate change affects nutrient levels, which make it hard to widely use microbial biofortification (Thakur et al., 2022).
This approach can also be applied to many fruit crops, helping to improve their nutritional quality and address vitamin and mineral deficiencies in people’s diets around the world.
Marker-assisted breeding (MAB) is an effective method used to improve the nutritional quality of fruit crops. This technique uses molecular markers to identify and select specific traits that enhance nutrient content in plants. By using MAB, breeders can quickly add desirable traits to fruit varieties, ensuring they are not only high-yielding but also rich in essential vitamins and minerals.
Biotechnology tools, such as genetic engineering and genome editing, work well with marker-assisted breeding. These tools allow for precise changes in the plant’s genetic makeup, enabling the introduction of specific genes that boost the availability of important nutrients. For example, transgenic methods can be used to add genes that increase the levels of vital nutrients like iron and zinc in the edible parts of fruit crops (Bouis et al., 2017; Athar et al., 2020).
The CRISPR/Cas9 genome editing technology is another innovative method for improving the nutritional quality of fruit crops. This advanced technique allows for targeted changes in the plant’s DNA, making it easier to enhance specific traits related to nutrient content. By combining marker-assisted breeding with these biotechnological tools, researchers can develop fruit varieties that are not only more nutritious but also better able to withstand environmental challenges as presented in Table 3.
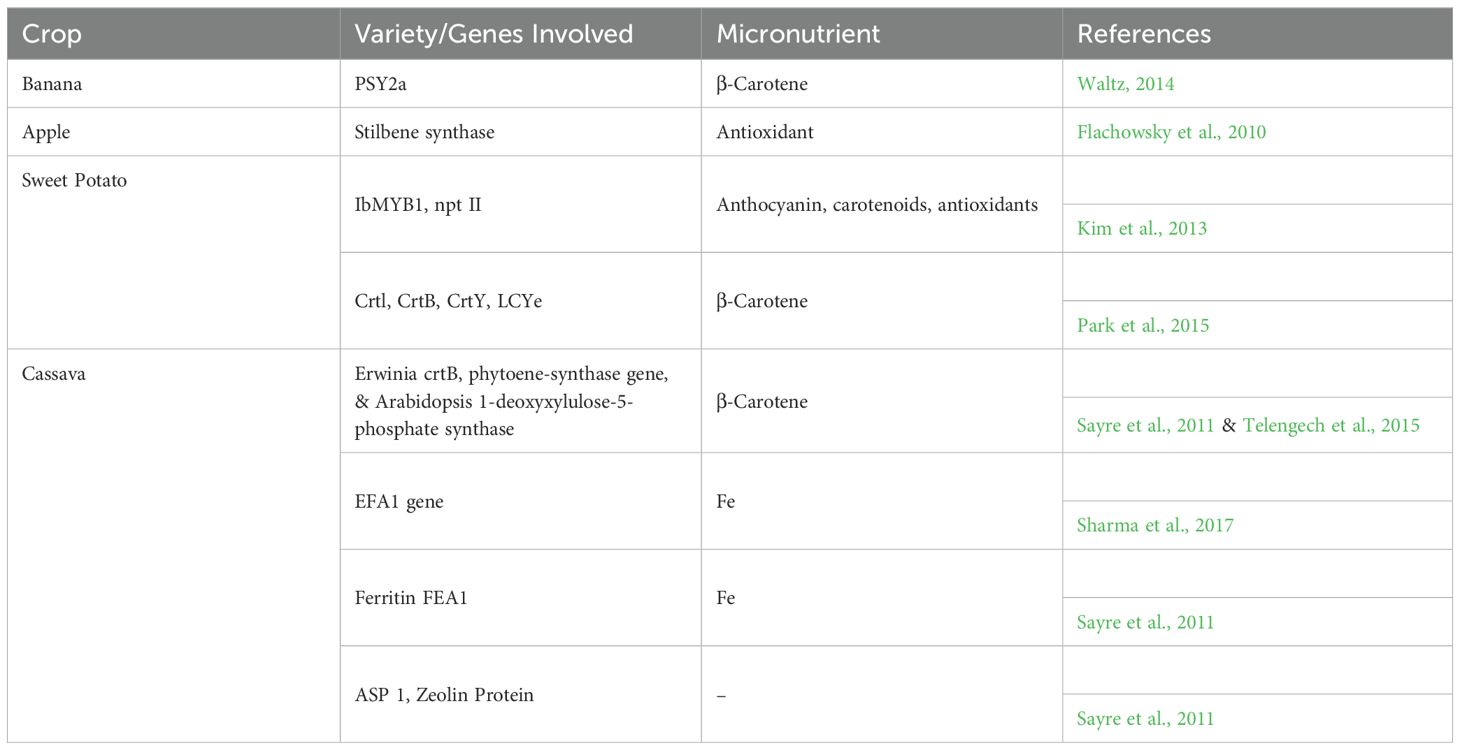
Table 3. Details of the fortified crops with the help of Genetic Engineering are as follows.
RNA interference (RNAi) technology has been used to create cassava plants that are resistant to cassava mosaic disease (CMD) and cassava brown streak disease (CBSD). This technology works by introducing small RNA molecules that target and break down the viral RNA, stopping the virus from spreading in the plant. Field trials have shown that these genetically modified cassava plants are resistant to both diseases and grow normally, even in conditions where the diseases would usually cause major damage. This innovation shows the potential of RNAi in controlling viral diseases in important crops, helping to ensure food security in areas where cassava is a key crop (Jain, 2024).
According to Bharti and Chimata (2019), New Plant-Breeding Techniques (NBTs) such as CRISPR and ZFN offer promising solutions for biofortifying fruit crops by improving their nutritional content more efficiently. These techniques allow for the development of fruit varieties that are richer in essential vitamins and minerals like vitamin C, iron, and zinc. Besides enhancing nutrient levels, NBTs can help address challenges such as pest resistance and environmental stresses, making fruit production more sustainable. By reducing the need for pesticides and optimizing resource use, these techniques contribute to the creation of nutrient-dense fruits that benefit farmers, consumers, and the environment. According to the study, Site-Directed Nucleases (SDN) and Oligonucleotide-Directed Mutagenesis (ODM) are two methods under NBTs that can be used to precisely modify genes in fruit crops, improving nutrient uptake and enhancing health benefits without introducing foreign DNA. These techniques offer non-GMO solutions for biofortifying fruits, making them an acceptable option for public concerns.
Bharti and Chimata (2019), also found that Cisgenesis and Intragenesis are other innovative techniques that can be used for transferring beneficial genes within the same or closely related species. These approaches allow for the introduction of traits like disease resistance and higher nutrient content without the need for foreign DNA, making biofortified fruits similar to conventionally bred ones. Additionally, RNA-Dependent DNA Methylation (RdDM) is a technique that can regulate gene expression to enhance nutrient production, improving vitamin and antioxidant content in fruits. Since RdDM modifies gene expression rather than the genetic sequence, it provides a way to biofortify fruits without foreign DNA. Techniques like grafting, combining genetically improved rootstocks with non-GM scions, can further enhance fruit yield and nutritional quality. According to the study, agro-infiltration, a method to test biofortification traits in fruit crops, offers a way to evaluate the effects of genetic modifications quickly, allowing for more effective breeding programs. Overall, these techniques can be used in the future to transform fruit crops, making them more nutritious, sustainable, and beneficial in addressing global malnutrition challenges.
Exploring the Role of Organic Fertilizers, Biofertilizers, and Nutri-Priming in Micronutrient Enhancement of Crops”: Biofortification through non-transgenic methods (Marques et al., 2021). offers several benefits for increasing micronutrient content in crops, utilizing approaches such as organic fertilizers, biofertilizers, and nutri-priming. Organic fertilizers, like manure and compost, are eco-friendly alternatives to inorganic fertilizers and have been shown to boost micronutrient levels in crops such as barley, wheat, and rice. However, they come with limitations, including the potential presence of harmful pathogens, antibiotics, and heavy metals, and the highly variable nutrient content, which makes precise nutrient delivery challenging. Similarly, biofertilizers, which use plant growth-promoting microorganisms, can enhance nutrient availability and crop growth, but their effectiveness is limited by factors like soil conditions, environmental factors, and the challenge of selecting the right microorganisms for specific crops. Despite the promise of biofertilizers, they often fail to fully replace traditional fertilizers in agroecosystems. Nutri-priming, another approach where seeds are soaked in nutrient solutions before planting, can increase micronutrient content, such as zinc in wheat and chickpeas. This method is low-cost and farmer-friendly but is also influenced by crop type, genotype, and environmental conditions, which can limit its effectiveness. Overall, while these non-transgenic biofortification methods have potential, their limitations, such as nutrient availability, environmental dependency, and lack of consistency, must be addressed to optimize their use in improving micronutrient intake, particularly in resource-limited settings.
The use of biotechnological tools like marker-assisted selection (MAS) and quantitative trait locus (QTL) maps by breeders or biotechnologists to identify gene of interest in large populations for use in breeding program is also one of the important aspects. Marker aided selection (MAS) leads to indirect selection of desired nutrient pathway favoring selection for Biofortification based on banding pattern of linked molecular markers. Various natural or created populations like Recombinant inbred lines (RILs), near isogenic lines (NILs) of interest are used for it. By the use of Genetic engineering technique or transgenic Breeding we can transfer gene of interest after cloning it across Genera and Species. Various Biological methods like Agro bacterium mediated gene transfer and non-biological methods like gene gun method, liposome mediated gene transfer, Micro injection method are yet to be used at larger scales for proper gene deployment favoring nutrient enhancement in required crop plants. For map-based cloning of genes of interest special mapping strategies can be used for rapidly targeting specific regions of the genome (Yu et al., 2021).
Various Molecular methodologies will serve as an important tool in future for development of Biofortified varieties by evaluation of genetic resources at molecular level and providing the clear-cut ideas of beneficial genetic resources for usage in further breeding programme (Shah et al., 2023). The use of CRISPR-Cas and Genome wide association mapping (GWAS) are the most advanced tool for success under present scenario. The CRISPR-Cas technology enables crop genome editing for qualitative enhancement in aroma, shelf life, oleic acid, anthocyanin, phytic acid, gluten etc. contents in several crop plants (Liu et al., 2021). Various genome edited golden crops like rice cultivar Kitaake has been developed by Knock-in, a 5.2-kb carotenogenesis cassette consisting of CrtI and maize PSY genes. The variety contains 7.9 μg/g dry weight (DW) β-carotene in the endosperm (Dong et al., 2020).
The use of new tools like Genome wide association mapping (GWAS) has opened the new door for fortification in important crops adapted to specific regions. GWAS can identify the loci associated with observed variations in phenotype for various agronomic traits among the Germplasm having nutrient deficit, which plays important role in regulation of the traits for using it as a genetic tool for pre breeding activities. Genome-wide association studies (GWAS) use high-throughput genomic technologies to scan entire genomes of large numbers of targets quickly, in order to find genetic variants correlated with a trait of interest. Accelerating breeding efforts for developing Biofortified bread wheat varieties necessitates understanding the genetic control of grain zinc concentration (GZnC) and grain iron concentration (GFeC) (Juliana et al., 2022). Scientists are trying to explore similar technique in most of the commercially important fruit crop.
The Pomegranate Solapur Lal is a high-nutrient hybrid (Bhagawa x [(Ganesh x Nana) x Daru]) variety developed by the ICAR-National Research Centre on Pomegranate in Pune (Table 2). Released in 2017, it is designed for semi-arid regions and yields 23 to 27 tons of fruit per hectare. It contains significantly higher levels of iron (5.6–6.1 mg/100g), zinc (0.64–0.69 mg/100g), and vitamin C (19.4–19.8 mg/100g) compared to the Ganesh and Bhagwa varieties, which have lower nutrient levels (Yadava et al., 2020). This variety matures 15-20 days earlier than Bhagwa, with a growing period of 160-165 days and an average yield of 35-39 kg per plant. It also has higher amounts of vitamin C, anthocyanins, and zinc, though it has a harder seed coat than Bhagwa.
Davarpanah et al. (2020) found that early season foliar fertilization with 2.6 mM FeSO4 significantly increased pomegranate fruit yield and quality. The treatment led to a 20–31% increase in yield, improved the number of fruits per tree, and enhanced juice content, total soluble solids, and sugar levels. FeSO4 also lowered juice acidity and raised the maturity index, though it slightly reduced antioxidant activity and phenolic compounds in the juice. This study highlights FeSO4 as an effective method for boosting pomegranate yield and fruit quality in high pH soils.
Efforts to develop biofortified bananas focus on improving their nutritional content through traditional breeding and genetic engineering. Traditional breeding aims to increase levels of provitamin A carotenoids, like beta-carotene, to help combat vitamin A deficiency, especially in regions where bananas are a staple food (Davey et al., 2009). Genetic engineering has also been used to enhance bananas’ nutritional value. For example, the Golden Banana has been developed to have higher levels of provitamin A carotenoids to address vitamin A deficiency (Arango et al., 2010). Additionally, researchers are working on increasing other nutrients like iron and zinc in bananas (Naqvi et al., 2009). Efforts are also being made to reduce harmful compounds, such as acrylamide, which can form during cooking and may be a carcinogen (Joshi et al., 2017a). Red bananas, such as “Red Dacca” and “Red Cavendish,” offer additional health benefits compared to yellow bananas. They have higher vitamin C content, potassium, dietary fiber, and antioxidants like anthocyanins, which can provide antioxidant and anti-inflammatory benefits (Lobo et al., 2010). Researchers are also using RNA interference to combat viral infections in bananas. For instance, transgenic plants with silenced viral components have shown promise in eliminating the bunchy top virus (Shekhawat et al., 2012).
Breeding of bananas is challenging because most commercial varieties are sterile and cannot crossbreed easily. However, genetic engineering is effective for bananas since they are typically grown from plant parts, which reduces the risk of spreading modified genes. Genetically modified bananas can be grown alongside regular ones without affecting their diversity. Unfortified bananas have 0.4 mg of iron per 100 grams, while fortified bananas can have up to 2.6 mg. Efforts are being made to increase bananas’ vitamin A, vitamin E, and iron levels. Research at Queensland University of Technology (QUT) has developed bananas with higher levels of these nutrients and improved resistance to diseases like Banana Bunchy Top Virus (QUT 2024). Similar research is underway in India with varieties such as Grand Nain and Rasthali. The Donald Danforth Plant Science Center is also developing bananas with high provitamin A for Nigeria and Kenya, with a new variety expected to be available in future.
The Coordinated efforts of TNAU, Coimbatore and IIHR Bangalore along with Queensland University of Technology, Australia under banana Biofortification programme Grand Nain and Rasthali Varieties of Banana will be Fortified with (Provitamin A) in first phase and later these varieties will be incorporated with tolerance for Banana Bunchy Top virus and Fusarium Wilt disease, similar work has been done to increase provitamin A content to 20 ppm by Donald Danforth Plant Science Center for locally adapted varieties for release in Nigeria & Kenya (Prasad et al., 2015). The identification and diffusion of well adapted varieties with high Provitamin A content especially in endemic regions (Bouis and Saltzman, 2017) has been adopted for the dissemination of high Provitamin A content of cooking and dessert bananas in Burundi, Rwanda and DR Congo (Andersson et al., 2017 and Ekesa et al., 2017). Some researchers have tried to reduce acrylamide which is a carcinogenic toxic substance produced in banana while cooking without hampering the original nutritional quality (Joshi et al., 2017b).
Biofortification of mangoes (Mangifera indica) aims to boost their nutritional value by increasing key micronutrients and phytochemicals. Although research on mango biofortification is limited, both traditional breeding and modern biotechnological methods have been explored. A study by Padmesh et al. (2013) examined how conventional breeding could improve the nutrient content of mangoes. The researchers found significant differences in mineral levels like iron, zinc, and calcium among various mango types, suggesting that breeding could produce mango varieties with higher mineral content. Genetic engineering is another approach being used to enhance mango nutrition. Saranya et al. (2017) worked on genetically modifying mango plants to increase their beta-carotene, a precursor to vitamin A. They introduced a gene from another plant that helps produce carotenoids, resulting in mangoes with higher beta-carotene levels. Agronomic practices also affect mango nutrition. Dhotra et al. (2021) tested the effect of applying micronutrients such as calcium nitrate, boric acid, and zinc sulfate to mango trees before flowering. Their study found that these nutrients enhanced the growth, yield, and quality of Dashehari mango fruits.
Mahida et al. (2023) found that agronomic biofortification, particularly using foliar sprays of zinc and iron, is an effective method to boost nutrient levels in crops. Their 2016-2017 field study on mango cv. Kesar showed that applying 0.5% FeSO4 and 0.5% ZnSO4 increased the nutrient content in both pulp and peel. The highest concentrations of zinc and iron were observed with the combination of both micronutrients, offering a promising approach to enhance mango nutrition.
Biofortification of guava aims to increase its nutritional value by boosting levels of essential nutrients and beneficial compounds. Traditional breeding methods have focused on developing guava varieties with higher concentrations of vitamin C, beta-carotene, iron, and zinc. These efforts help address micronutrient deficiencies and enhance the fruit’s overall nutritional profile (Navarro-Tarazaga et al., 2016). Additionally, genetic engineering techniques have been applied to further enhance guava’s nutritional content. By introducing specific genes into guava plants, researchers can increase the production of important bioactive compounds. For instance, the Paluma guava cultivar, known for its rich crimson pulp and high soluble solids content, has been developed to offer improved nutritional benefits and antioxidant properties. Alongside these genetic approaches, optimizing agricultural practices such as proper fertilization and irrigation can significantly boost nutrient uptake and accumulation in guava plants, contributing to effective biofortification (Sathya et al., 2019).
Grapes have been improved using both traditional breeding and modern genetic techniques to boost their health benefits. Traditional breeding focuses on increasing the levels of beneficial compounds like polyphenols and anthocyanins, which have antioxidant properties and are good for health (Di Lorenzo et al., 2019). This method involves selecting grape varieties that naturally have higher amounts of these health-promoting compounds. Modern genetic engineering also plays a role in enhancing grapes. This approach involves adding or modifying specific genes to increase the production of desirable compounds, such as resveratrol, which is known for its health benefits (Vezzulli et al., 2007). By making precise changes to the grape’s genetic makeup, scientists can improve the grapes’ nutritional value. Additionally, how grapes are grown—through different fertilization, irrigation, and canopy management techniques—can also impact their nutritional content (Cortell and Kennedy, 2006). Grapes are rich source of antioxidant with high levels of vitamin C and K. Pusa Navrang a new Cultivar of Grape introduced by Indian Agricultural Research Institute is rich in soluble solids like carbohydrates, organic acids, proteins, lipids, minerals and antioxidants (Table 2).
Zhao et al. (2020) conducted a study on biofortifying grapes with selenium (Se) and lithium (Li) using foliar application of organic fertilizers rich in these micronutrients. The study focused on five grape cultivars and evaluated the impact on vine vigour, fruit quality, and the levels of micronutrients and phenolic compounds. The results showed that the biofortification significantly increased the Se and Li content in the grapes, particularly in the skin, without adversely affecting vine vigour or fruit quality. However, the biofortification did influence the ions (mineral nutrients and trace elements) and phenolic compounds, with variations across cultivars. This method proved to be an effective strategy for enhancing the nutritional value of grapes, potentially improving their health benefits and increasing consumer awareness and consumption of these enriched fruits, especially in the Midwestern United States.
Daccak et al (2022) studied the biofortification of grapes with zinc (Zn) to help address micronutrient deficiencies in food. Zinc is important for human health, and the research focused on the Fernão Pires grape variety. The vines were sprayed with two types of zinc fertilizers, ZnO and ZnSO4, at three different concentrations (150, 450, and 900 g ha−1) during the growing season. The study looked at how zinc accumulated in the grapes, especially in the skin and seeds, and how it affected grape quality and winemaking. The results showed that the zinc levels in treated grapes were much higher than in the control grapes, with ZnO increasing zinc in the skin and ZnSO4 in the seeds. After winemaking, zinc content in the wine also increased by about 1.59 times with the 450 g ha−1 ZnSO4 treatment. While there were no significant changes in sugar, fatty acids, or color, the study concluded that biofortifying grapes with zinc is a good way to increase their nutritional value without harming their quality, making it a useful strategy for improving both grapes and wine.
Biofortification of papaya focuses on improving its nutritional value through genetic engineering. One key approach is increasing provitamin A carotenoids, like beta-carotene, by introducing genes that enhance carotenoid production (Shankar et al., 2010). This helps address vitamin A deficiency in populations that rely heavily on papaya. Another focus is boosting essential minerals, such as iron and zinc, in papaya fruits using genetic engineering techniques (Maxwell et al., 2013).
Non-conventional tools like Genetic Engineering led the introduction of specific genes involved in carotenoid biosynthesis in locally adapted papaya Variety that has enhanced the beta-carotene content (Shankar et al., 2010). After collecting material from Centre for Tropical Agriculture, Southedge, QLD Australia (Devit et al., 2010), observed that the expression level of both lcy-β1 and lcy-β2 genes is similar and low in leaves, but lcy-β2 expression increases markedly in ripe fruit. Isolation of the lcy-β2 gene from papaya, that is preferentially expressed in fruit and is correlated with fruit colour, will facilitate marker-assisted breeding for fruit colour in papaya and should create possibilities for metabolic engineering of carotenoid production in papaya fruit to alter both colour and nutritional properties.
Researchers have identified apple varieties with increased levels of antioxidants such as phenolic compounds, flavonoids, and anthocyanins, which are associated with health benefits due to their antioxidant and anti-inflammatory properties (Strand et al., 2018). Proper nutrient management, like balanced fertilization, can improve the nutritional quality of apples by influencing their nutrient content (Tagliavini et al., 2019). Genetic engineering is being explored to enhance apples by introducing specific genes. For example, incorporating a stilbene synthase gene from grapevines can increase resveratrol, a potent antioxidant, in apples (Tian et al., 2017). This genetic modification aims to improve the antioxidant capacity of apples. Scientists have also modified apple plants to produce higher levels of anthocyanins and flavan-3-ols, beneficial compounds that contribute to the apple’s nutritional value (Flachowsky et al., 2010). Additionally, Arctic® apples have been engineered to prevent browning by inhibiting polyphenol oxidases (PPOs), the enzymes responsible for browning. Arctic varieties include Fuji, Granny Smith, and Golden Delicious, with different types being released over the years (Lobato-Gomez et al., 2021).
Biofortification in pears aims to boost their nutritional content and help with nutrient deficiencies. Although research on pears is less developed compared to other crops, there are some key strategies being explored. One method is conventional breeding, where pear varieties are selected and crossed to increase their levels of vitamins, minerals, and antioxidants. Another method is genetic modification, which involves adding or enhancing specific genes to improve nutrient levels. For example, scientists have studied boosting genes related to antioxidants like anthocyanins to enhance the antioxidant properties of pears (Han et al., 2016). Agronomic Biofortification in Pyrus communis L. variety Rocha, has been successfully deployed with Foliar application of 0.6 kg Ca (NO3)2 ha−1 or 1.6 kg CaCl2 ha−1, that promotes Ca Biofortification in leaves without any phytotoxity symptoms (Cardoso et al., 2018). Successful attempts for Zinc fortification were done by (Liu et al., 2023) with chelated and non-chelated Zinc, however it was observed that chelated ZnEDTA can be safely used at a higher spraying rate of 1.5% as compared to 0.1%–0.4% for non-chelated Zn sources. Pear is a major fruit crop spreaded worldwide, but its breeding is time taking process. To facilitate molecular breeding and gene identification (Zhang et al., 2021), have performed genome-wide association studies (GWAS) on eleven fruit traits. They identified 37 loci associated with eight fruit quality traits and five loci associated with three fruit phenological traits. Over the period of time new advantageous mutation eliminates variation in lined neutral sites indicating that traits like fruit stone cell content, organic acid and sugar contents might have been under continuous selection during breeding improvement. One candidate gene, PbrSTONE, identified in GWAS, has been functionally verified to be involved in the regulation of stone cell formation, which is one of the most important fruit quality traits in pear.
It is well-regarded for its high vitamin C content and other healthful compounds. Traditional breeding methods focus on improving its nutritional profile through selection and cross-breeding techniques, aiming to enhance its beneficial properties further (Singh et al., 2022). In addition, genetic engineering offers the potential to introduce specific genes that could boost nutrient levels in aonla. However, this approach faces challenges, including regulatory hurdles and public concerns about genetically modified organisms (Chaurasia et al., 2009). Moreover, the use of black polythene mulch has been found to improve the quality of aonla fruits, particularly for the NA-7 variety (Table 2), by positively affecting various quality parameters (Iqbal et al., 2015).
Efforts to improve strawberries focus on making them more nutritious and beneficial for health (Singh et al., 2022). Genetic engineering has helped boost the vitamin C content and increase the plants’ resistance to diseases and environmental challenges (Borowski et al., 2016). For instance, scientists altered strawberries to have up to 47% less starch and 37% more soluble sugar by blocking a specific enzyme that converts sugar into starch (Park et al., 2006). Additionally, applying a 900 ppm cycocel spray to strawberries has been shown to enhance their quality by increasing their sweetness, vitamin C content, juice yield, and lowering their acidity (Kumar et al., 2012).
To improve the nutritional content of citrus fruits, traditional breeding methods involve selecting and crossbreeding varieties that naturally have higher nutrient levels. For example, breeders focus on increasing the vitamin C content in citrus fruits (Salonia et al., 2020). Genetic engineering also plays a role in enhancing citrus nutrition. Researchers have introduced genes to boost iron levels in citrus by increasing iron transporters, which can help combat iron deficiency (Zhang et al., 2019). They have also worked on increasing pro-vitamin A carotenoids, like beta-carotene, and folate in citrus fruits (Pons et al., 2018). Additional techniques include using specific fertilizers and treatments to improve the fruit quality, such as applying nitrogen, potassium sulfate, and calcium chloride (Bakshi et al., 2017; Devi et al., 2018). These efforts aim to provide citrus fruits with better nutritional value, especially in areas with high vitamin A deficiency.
Bhantana et al. (2022) emphasized the importance of optimizing micronutrient concentrations, particularly zinc (Zn), for improving the mineral nutrition, yield, and quality of citrus fruits. The study found that the soil had a Zn concentration of 22.35 mg/Kg, which is considered marginally available for plant uptake. Before treatment, the top leaves of the citrus varieties Wenzhou and Nanfeng showed relatively low concentrations of essential nutrients like nitrogen (N), phosphorus (P), potassium (K), and Zn. The study concluded that biofortification with Zn (either through foliar or soil application) is effective when soil Zn levels are below the critical threshold of 20 mg/Kg. Micronutrient supplementation under these conditions can improve the yield and nutritional quality of citrus fruits.
Lycopene is a carotenoid compound (Perkins et al., 2001) responsible for the red color in watermelon and is associated with various health benefits, including reducing the risk of certain cancers and cardiovascular diseases. Researchers have focused on increasing lycopene content through conventional breeding methods and genetic engineering techniques. Studies have shown success in developing watermelon varieties with higher levels of lycopene, contributing to enhanced nutritional value (Deng et al., 2023). highlights the potential of using nanoscale CuO particles to enhance disease management and nutritional quality of crops. Specifically, negatively charged nanospikes demonstrated significant benefits in both disease control and nutrient enhancement. Investigation from (Joshi et al., 2021) highlighted considerable natural variation in the concentrations of free amino acids and total protein across different watermelon accessions and regions, through a genome-wide association study (GWAS), they identified 188 SNPs linked to 167 candidate genes involved in seed-bound amino acids and total protein content. Analysis revealed that SNP clustering was independent of watermelon speciation and cultivar types, suggesting these genetic variations were not influenced by the domestication process. Significant candidate genes, such as Argininosuccinate synthase, were associated with 7% of the variation in arginine content, confirming their functional relevance. This study provides critical insights and a robust platform for future research aimed at developing watermelon varieties with superior nutritional attributes.
In the study by (Mengist et al., 2020), a high-throughput in vitro gastrointestinal digestion model was used to assess the bioaccessibility of phenolic compounds in various cultivated highbush blueberry accessions. The results showed significant differences in the bioaccessibility of flavonoids and phenolic acids among different accessions and years, with some accessions exhibiting high relative and absolute bioaccessibility values. Acylated anthocyanins were found to have higher relative bioaccessibility compared to non-acylated anthocyanins. The study also suggested that both genetic and environmental factors influence phenolic bioaccessibility, with broad sense heritability estimates indicating a moderate genetic contribution. These findings emphasize the importance of integrating genetic and genomic approaches to enhance the bioactive delivery of blueberries and identify key genotypes and genetic factors. Biofortification of fruit crops can be done by using the different techniques for enhancing the targeted nutrients and vitamins are as given in the Table 4.
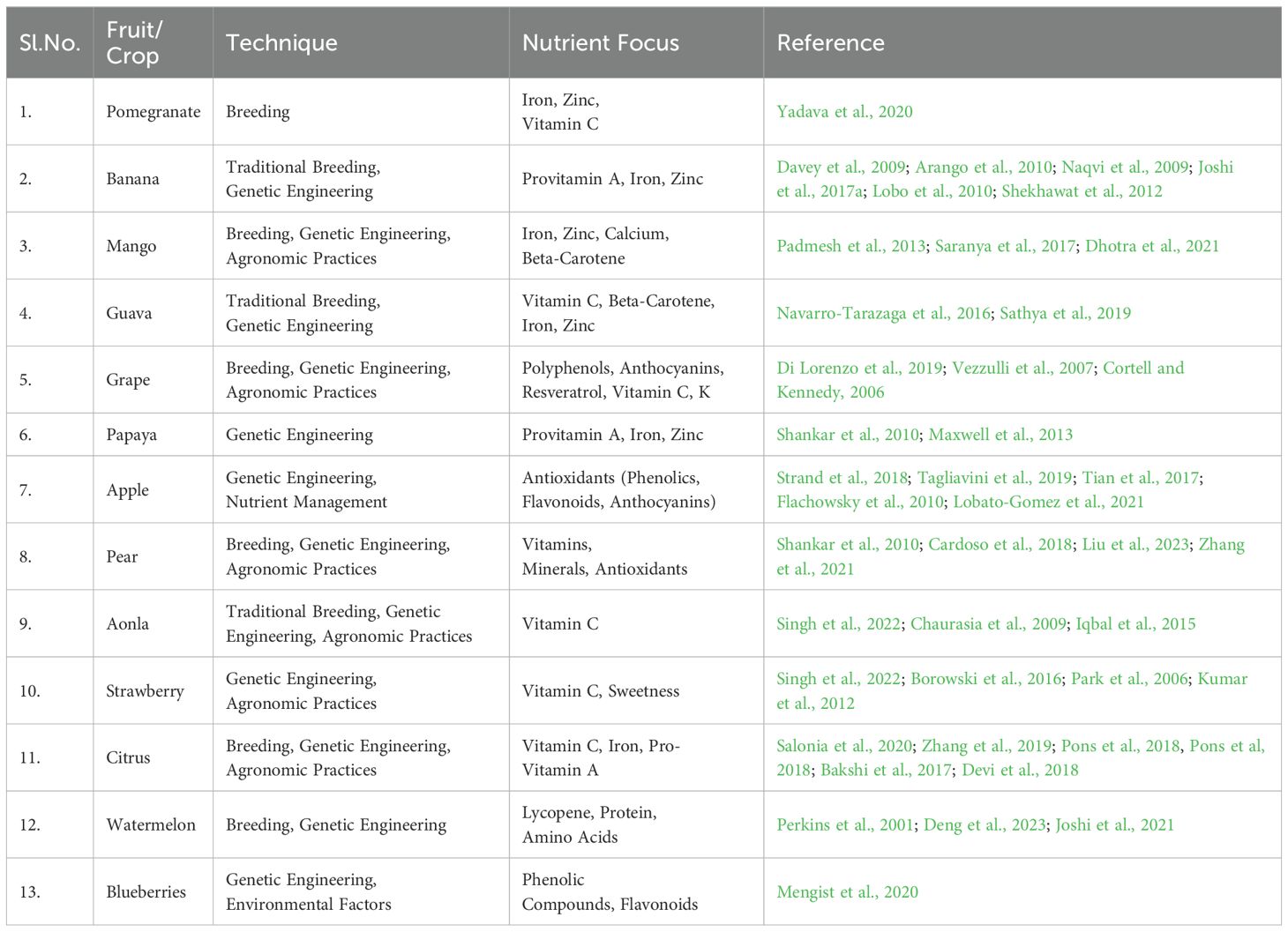
Table 4. Overview of Empowering Vital Fruit Crops with Enhanced Nutritional Contents.
Biofortification is a vital strategy for enhancing the nutritional value of fruits by increasing essential vitamins, minerals, and beneficial compounds. For instance, Mangifera indica (mango) has benefited from traditional breeding techniques that identify varieties rich in minerals like iron and zinc (Padmesh et al., 2013). Additionally, genetic engineering has successfully increased beta-carotene levels (Saranya et al., 2017), while agronomic practices such as micronutrient applications further enhance fruit quality (Dhotra et al., 2021). Similarly, Psidium guajava (guava) utilizes both traditional breeding for higher vitamin C and zinc content and genetic engineering to improve antioxidant levels, notably in the Paluma cultivar (Navarro-Tarazaga et al., 2016; Sathya et al., 2019).
Traditional breeding and genetic engineering also play significant roles in Vitis vinifera (grapes), where breeding increases antioxidants like polyphenols, and genetic modifications boost resveratrol levels (Di Lorenzo et al., 2019; Vezzulli et al., 2007). Agronomic practices contribute further to the nutritional content of these fruits (Cortell and Kennedy, 2006). In Carica papaya (papaya), genetic engineering aims to elevate provitamin A carotenoids and enhance disease resistance through targeted gene modifications (Shankar et al., 2010; Devit et al., 2010). Likewise, Malus domestica (apple) benefits from both traditional breeding and genetic engineering, with Arctic® varieties offering resistance to browning and enhanced antioxidant levels (Strand et al., 2018; Lobato-Gomez et al., 2021).
Pyrus communis (pear) demonstrates a similar integration of traditional breeding and genetic modifications aimed at boosting vitamins and antioxidants, complemented by agronomic practices such as foliar calcium and zinc applications (Han et al., 2016; Cardoso et al., 2018; Liu et al., 2023). Meanwhile, Phyllanthus emblica (aonla) relies primarily on traditional breeding and agronomic practices for quality improvements, facing regulatory challenges in genetic engineering (Singh et al., 2022; Iqbal et al., 2015).
Advancements in Fragaria × ananassa (strawberry) include improvements in vitamin C content and sweetness achieved through genetic modifications and agronomic techniques like cycocel spraying (Borowski et al., 2016; Kumar et al., 2012). Citrus species (Citrus spp.) have similarly benefited from breeding and genetic engineering to enhance vitamin C and iron content, further improved by agronomic practices (Salonia et al., 2020; Zhang et al., 2019). For Citrullus lanatus (watermelon), breeding and genetic modifications have increased lycopene content, while emerging techniques such as nanoscale particles for nutrient enhancement show promise (Perkins et al., 2001; Deng et al., 2023). Lastly, Vaccinium corymbosum (blueberries) highlights the variability in phenolic compound bioaccessibility, influenced by genetic and environmental factors, underscoring the necessity for integrated approaches (Mengist et al., 2020).
These examples collectively illustrate the potential of combining traditional, genetic, and agronomic methods to comprehensively improve fruit nutrition. Continued research is crucial for optimizing these approaches to meet global nutritional needs effectively.
Micronutrient malnutrition affects over 2 billion people globally, with deficiencies in vitamins and minerals like iodine, iron, zinc, and vitamin A posing significant health risks. Unlike macronutrients, which are needed in large amounts, micronutrients are essential in smaller quantities for proper bodily functions and development. Poor intake and absorption, exacerbated by factors such as inadequate food security and health services, lead to widespread deficiencies, particularly among pregnant women and young children. With the world population projected to reach 8 billion by 2030, addressing micronutrient malnutrition is crucial for improving public health, especially in developing countries.
A. Iron: Iron is essential for various cell functions, notably oxygen transport in hemoglobin. It is the leading cause of anemia, particularly affecting young women and children, with over 2 billion people impacted worldwide (Benoist et al., 2008). Symptoms of iron deficiency include fatigue and metabolic disruptions, while severe cases can lead to significant health complications.
B. Zinc: Zinc is a vital mineral required for cell division, growth, and immune function, yet it cannot be stored in the body (Frassinetti et al., 2006). Approximately 1.1 billion people are affected by zinc deficiency due to inadequate dietary intake. This deficiency is linked to various health issues, including impaired immunity, delayed wound healing, and emotional disturbances (Evans, 1986).
C. Iodine: Iodine is crucial for the production of thyroid hormones, which regulate metabolism. More than 2 billion people globally suffer from iodine deficiency, leading to conditions like goiter and developmental impairments in children (Delange, 1994). Insufficient iodine during pregnancy can result in neuro developmental issues for the offspring.
D. Vitamin A: Vitamin A is important for immune health, vision, and cell growth. Its deficiency affects 100–400 million preschool-aged children globally, resulting in significant health consequences, including blindness. Among pregnant women, vitamin A deficiency can led to night blindness and increased maternal mortality. Many developing countries rely on plant foods to meet their vitamin A needs.
E. Vitamin B: Vitamin B comprises eight forms, including B1, B2, B3, B5, B6, B8, B9, and B12, each acting as a co-factor in essential metabolic processes like carbohydrate metabolism and protein synthesis. Vitamin B6, in particular, is vital for protein metabolism, immune function, and neurotransmitter synthesis, but humans cannot produce it and must obtain it from dietary sources (Bryan et al., 2002). The rising prevalence of B6 deficiencies can lead to symptoms such as skin inflammation, fatigue, and depression.
F. Vitamin C: Vitamin C is a water-soluble vitamin predominantly sourced from plants, well-known for its immune-boosting properties and antioxidant effects. It acts as a co-factor in synthesizing collagen, cholesterol, and amino acids, and is also involved in energy metabolism (Perez-Massot et al., 2013; Maggini et al., 2017). Deficiency in vitamin C can lead to joint pain, connective tissue disorders, poor wound healing, and weakened immune response (Maggini et al., 2017).
G. Vitamin E: Vitamin E is a fat-soluble vitamin found in oil-rich foods like peanuts and sunflower seeds, and it can be stored in the body’s fat reserves. It serves as an antioxidant, regulates membrane lipid packaging, and is important for preventing diseases like cancer and cardiovascular conditions (Fitzpatrick et al., 2012). Deficiency typically occurs in individuals with fat metabolism disorders and can result in muscle weakness, hemolytic anemia, and neurological issues.
A. Soil and Plant Factors: Micronutrients travel from the soil to plants and then to humans, so the quality of the soil is very important. Key factors include soil texture, pH level, organic matter, and moisture. How well plants absorb nutrients depends on their root structure and connections with fungi (mycorrhizae). Some plants adapt to low nutrient levels by growing more roots or releasing acids to make nutrients more available (Zhang et al., 2010; Marschner, 2012).
B. Diet-Related Factors: When we eat plant foods, various factors affect how well our bodies absorb nutrients. These include the form of the nutrient, the type of food, and how different nutrients interact with each other. For example, vitamin C can help with iron absorption, while compounds like phytic acid can block it (Gibson, 2007). Individual factors like age, health, and genetics also play a role. If someone is low in iron or zinc, their body may absorb these nutrients better during that time (Hallberg and Hulthen, 2000). Malnutrition and infections can further hinder nutrient absorption (Johnson et al., 2008).
C. Food Fortification: To fight micronutrient malnutrition, adding essential nutrients to everyday foods—called food fortification—is a useful strategy (Allen et al., 2006). This method has been widely used in richer countries and is becoming more common in developing nations. For instance, Pakistan has started programs to add vitamins and minerals to wheat flour and cooking oil (Awan et al., 2013). These initiatives aim to improve nutrition without requiring people to change their diets significantly.
D. Biofortification of fruit crops faces several limitations, including issues related to bioavailability and the presence of antinutrients, which can hinder the absorption of essential micronutrients. For instance, compounds like phytic acid can form complexes with minerals such as iron and zinc, reducing their solubility and absorption in the human body. Additionally, consumer acceptance of biofortified crops, particularly those developed through transgenic methods, is often low due to concerns regarding genetically modified organisms. Regulatory challenges further complicate the production and distribution of these crops, as numerous regulations govern transgenic crops, creating barriers to market entry. Moreover, agronomic biofortification relies on the regular application of mineral fertilizers, which can be costly and may not be feasible for all farmers, potentially leading to environmental concerns and toxicity from repeated applications. Thus, while biofortification holds promise for improving nutritional quality, these limitations must be addressed to enhance its effectiveness and accessibility (Ofori et al., 2022).
To take the edge off Malnutrition, Biofortification is the most economical way to reinforce the nutritional security to undernourished people across India and world. The technology is not only confined to Agronomic fortification but also it has explored various conventional and biotechnological interventions leading to Biofortification in commercially important crops. Collaboration of interdisciplinary subject specialists along with their dedicated effort is needed. As compared to conventional breeding, biotechnological approaches to fortify crops are less time consuming but on the other hand lacks socio-economic and public acceptance as some of its advance approaches like transgenic breeding involves transfer of gene across genera or species and approval from genetic engineering approval committee (GEAC) in countries like India is also difficult. A Biofortification effort based on multiple coordinated efforts from interdisciplinary group is required for reducing the malnutrition in different populations thereby reducing the global hunger index and nutrition.
Van et al., 2017 found that biofortification through genetic engineering is generally well accepted by the majority of beneficiaries, with consumers willing to pay a premium for biofortified crops, which is crucial for the successful implementation of biofortification as a health policy intervention. They emphasized that this approach offers good value for money, particularly when multiple micronutrients are targeted in a single crop, thereby enhancing its economic viability and appeal to both producers and consumers. Furthermore, the authors highlighted the necessity of upgrading local supply chains to fully exploit the potential of biofortification and ensure effective delivery to consumers, especially in domestic markets where trade issues may be less relevant. Lastly, they underscored the importance of addressing ethical considerations related to biofortification, which is essential for informing policymakers and stakeholders about the implications of genetic engineering and ensuring the equitable distribution of biofortified crops.
In order to make use of Biofortified crops in seed chain, strengthening of formal and informal farming system is needed which most of the countries are using for reducing malnutrition. Intensive efforts have to be made by the policy makers like fixation of minimum support price (MSP) in public sector to support and promote Biofortified varieties. Considering these challenges the coordinated efforts by public sector, private sector and farmers will serve as a powerful tool for eradication of malnutrition all over the world.
Biofortification of fruits through traditional breeding, genetic engineering, and agronomic practices has substantially improved their nutritional content. Traditional breeding enhances fruit quality by selecting varieties with naturally higher nutrient levels, while genetic engineering enables precise modifications to boost specific vitamins and compounds. Agronomic practices further support these improvements by optimizing cultivation conditions. Combining these methods provides a comprehensive approach to addressing nutrient deficiencies and enhancing fruit quality. Continued research and development are essential to refine these strategies and ensure their widespread application, ultimately improving global fruit nutrition and health outcomes.
Many organizations are working to improve the nutritional quality of crops through biofortification. These include both national and international groups focused on fighting malnutrition and enhancing food security. Some key organizations involved in these efforts are CGIAR, which conducts agricultural research to reduce poverty and improve food security; CIAT, which aims to boost agricultural productivity in tropical areas; and CIMMYT, which specializes in maize and wheat research. The Food and Agriculture Organization (FAO) works globally to eliminate hunger and improve nutrition, while Harvest Plus is a leader in biofortification, focusing on increasing nutrients in staple crops. Other important organizations include GAIN, which aims to improve nutrition for vulnerable populations, UNICEF, which focuses on children’s nutrition, and the World Health Organization (WHO), which addresses global health issues, including malnutrition.
AJ: Conceptualization, Writing – original draft, Writing – review & editing. DJ: Conceptualization, Writing – original draft, Writing – review & editing. DS: Writing – review & editing. AK: Writing – review & editing. FA: Writing – review & editing.
The author(s) declare that no financial support was received for the research, authorship, and/or publication of this article.
The authors acknowledge to the Vice-chancellor, Bihar Agricultural University, Sabour for their gracious help and support.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declare that no Generative AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Allen, L. H., De Benoist, B., Dary, O., Hurrell, R., Organization, W. H (2006). Guidelines on food fortification with micronutrients (Geneva: World Health Organization).
Andersson, M. S., Saltzman, A., Virk, P. S., Pfeiffer, W. H. (2017). Progress update: crop development of biofortified staple food crops under Harvestplus. Afr. J. Food Agriculture Nutr. Dev. 17, 11905–11935. doi: 10.18697/ajfand.78.HarvestPlus05
Arango, J., Salazar, B., Garcia-Bastidas, F., Ortiz, J. F. (2010). Engineering banana and plantain to resist Fusarium wilt disease caused by Fusarium oxysporumf. sp. cubense. Trop. Plant Biol. 3, 88–102.
Athar, H. R., Khan, M. I., Pandey, A., Yilmaz, A., Gezgin, S. (2020). Genetic engineering for biofortification: A review. J. Plant Nutr. 43, 1–15. doi: 10.1080/01904167.2019.1671234
Avnee, S., Sood, S., Chaudhary, D. R., Jhorar, P., Rana, R. S. (2023). Biofortification: An approach to eradicate micronutrient deficiency. Front. Nutr. 10. doi: 10.3389/fnut.2023.1233070
Awan, J. A., Anjum, F. M., Huma, N. (2013). “Food fortification programmes in Pakistan,” in Handbook of food fortification and health. Eds. Preedy, V. R., Srirajaskanthan, R., Patel, V. B. (Springer, New York, NY), 347–360.
Bakshi, M., Wali, V. K., Bakshi, P., Sharma, A., Sharma, D., Shah, R. A. (2017). Integrated nutrient management induced changes in nutrient uptake, fruit yield and quality of Kinnow mandarin. Indian J. Agric. Sci. 87, 414–418. doi: 10.56093/ijas.v87i3.68790
Benoist, B., McLean, E., Egll, I., Cogswell, M. (2008). “Worldwide prevalence of anaemia 1993-2005,” in WHO global database on anaemia (World Health Organization, Geneva).
Bhantana, P., Moussa, M. G., Malla, R., Khadka, D., Shrestha, R. K., Vista, S. P., et al. (2022). Foliar versus soil biofortification of zn in citrus (Citrus reticulata blanco): effect on mineral nutrition and fruit yield and quality. BioMed. J. Sci. Tech Res. 41, 6612. doi: 10.26717/BJSTR.2022.41.006612
Bhardwaj, A. K., Geeta, A., Raj, K., Lamy, H., Hadi, P. A., Poonam, J. (2022). Switching to nanonutrients for sustaining agroecosystems and environment: The challenges and benefits in moving up from ionic to particle feeding. J. Nanobiotechnology 20, 1–28. doi: 10.1186/s12951-021-01177-9
Bharti, G., Chimata, M. K. (2019). Review on new plant breeding techniques. Int. J. Sci. Res. 8, 1–10.
Borowski, J. M., Bostan, H., Santelices, A., Gomez, G., Osorio, S. (2016). Genetic engineering of strawberry: Current status and future perspectives. Plant Cell Rep. 35, 1129–1151.
Bouis, H. E., Saltzman, A. (2017). Improving nutrition through biofortification: a review of evidence from HarvestPlus through 2016. Glob. Food Secur. 12, 49–58. doi: 10.1016/j.gfs.2017.01.009
Bouis, H. E., Saltzman, A., Low, J. W., Ball, A., Covic, N. (2017). Biofortification: A new tool to reduce micronutrient malnutrition. Food Nutr. Bull. 38, 1–12. doi: 10.1177/0379572116680010
Bryan, J., Calvaresi, E., Hughes, D. (2002). Short-term folate, vitamin B-12 or vitamin B-6 supplementation slightly affects memory performance but not mood in women of various ages. J. Nutr. 132, 1345–1356. doi: 10.1093/jn/132.6.1345
Budke, C., Dierend, W., Schön, H. G., Hora, K., Hermann, M. K., Diemo, D. (2021). Iodine biofortification of apples and pears in an orchard using foliar sprays of different composition. Front. Plant Sci. 12. doi: 10.3389/fpls.2021.638671
Cardoso, P., Mateus, T. C., Velu, G., RP, S., JP, S., ML, C., et al. (2018). Localization and distribution of Zn and Fe in grains of biofortified bread wheat lines through micro- and triaxial- X-ray fluorescence spectrometry Spectrochim. Acta B 141, 70–79. doi: 10.1016/j.sab.2018.01.006
Chaurasia, A. K., Subramaniam, V. R., Krishna, B., Sane, P. V. (2009). RAPD based genetic variability among cultivated varieties of Aonla (Indian Gooseberry, Phyllanthus emblica L.). Physiol. Mol. Biol. Plants 15, 169–173. doi: 10.1007/s12298-009-0019-5
Cortell, J. M., Kennedy, J. A. (2006). Effect of shading on accumulation of flavonoid compounds in (Vitis vinifera L.) Pinot Noir fruit and extraction in a model system. J. Agric. Food Chem. 54, 8510–8520. doi: 10.1021/jf0616560
Daccak, D., Lidon, F. C., Luís, I. C., Marques, A. C., Coelho, A. R. F., Pessoa, C. C., et al. (2022). Zinc biofortification in Vitis vinifera: Implications for quality and wine production. Plants 11, 2442. doi: 10.3390/plants11182442
Dapkekar, A., Deshpande, P., Oak, M. D., Paknikar, K. M., Rajwade, J. M. (2018). Zinc use efficiency is enhanced in wheat through nano fertilization. Sci. Rep. 8, 1–7. doi: 10.1038/s41598-018-25247-5
Davarpanah, S., Tehranifar, A., Zarei, M., Aran, M., Davarynejad, G., Abadía, J. (2020). Early season foliar iron fertilization increases fruit yield and quality in pomegranate. Agronomy 10, 852. doi: 10.3390/agronomy10060852
Davey, M. W., Van den Bergh, I., Markham, R., Swennen, R., Keulemans, J., Henshaw, G. G. (2009). The molecular biology of banana improvement: current status and future perspectives. In Vitro Cell. Dev. Biology-Plant 45, 297–321.
Delange, F. (1994). The disorders induced by iodine deficiency. Thyroid 4, 107–128. doi: 10.1089/thy.1994.4.107
Deng, C., Protter, C. R., Wang, Y., Borgatta, J., Zhou, J., Wang, P., et al. (2023). Nanoscale CuO charge and morphology control Fusarium suppression and nutrient biofortification in field-grown tomato and watermelon. Sci. Total Environment 905 167799. doi: 10.1016/j.scitotenv.2023.167799
Devi, K., Kumar, R., Wali, V. K., Bakshi, P., Sharma, N., Arya, V. M. (2018). Effect of foliar nutrition and growth regulators on nutrient status and fruit quality of Eureka lemon (Citrus limon). Indian J. Agric. Sci. 88, 704–708. doi: 10.56093/ijas.v88i5.80056
Devit, L. C., Fanning, K., Dietzgen, R. G., Timothy, A., Holton (2010). Isolation and functional characterization of a lycopene β-cyclase gene that controls fruit colour of papaya (Carica papaya L.). J. Exp. Bot. 61, 33–39.
Dhotra, B., Bakshi, P., Jeelani, M. I., Kumar, V., Rai, P. K., Khajuria, S., et al. (2021). Effect of foliar application of micronutrients on fruit growth, yield and quality of mango (Mangifera indica L.) cv Dashehari. Environ. Ecol. 39, 873–881.
Di Lorenzo, C., Colombo, F., Biella, S., Belgiovine, G., Ballabio, D., Tessarin, P., et al. (2019). Bioactive constituents and health effects of grapes. Molecules 24 4530. doi: 10.3390/molecules24234530
Dimkpa, C. O., Bindraban, P. S. (2016). Fortification of micronutrients for efficient agronomic production: A review. Agron. Sustain. Dev. 36, 1–27. doi: 10.1007/s13593-015-0346-6
Dimkpa, C. O., Singh, U., Adisa, I. O., Bindraban, P. S., Elmer, W. H., Gardea, -.-T., et al. (2018). Effects of manganese nanoparticle exposure on nutrient acquisition in wheat (Triticum aestivum L.). Agronomy 8, 158. doi: 10.3390/agronomy8090158
Dong, O. X., Yu, S., Jain, R., Zhang, N., Duong, P. Q., Butler, C. (2020). Marker-free carotenoid-enriched rice generated through targeted gene insertion using CRISPR-cas9. Nat. Commun. 11, 1178. doi: 10.1038/s41467-020-14981-y
Doolette, C. L., Read, T. L., Howell, N. R., Cresswell, T., Lombi, E. (2020). Zinc from foliar-applied nanoparticle fertilizer is translocated to wheat grain: A Zn radio-labelled translocation study comparing conventional and novel foliar fertilizers. Sci. Total Environ. 749, 142369. doi: 10.1016/j.scitotenv.2020.142369
Dotaniya, M. L., Datta, S. C., Biswas, D. R., Dotaniya, C. K., Meena, B. L., Rajendiran, S., et al. (2016). Use of sugarcane industrial by-products for improving sugarcane productivity and soil health. Int. J. Recycling Organic Waste Agric. 5, 185–194. doi: 10.1007/s40093-016-0132-8
Eckhardt, U., Marques, A. M., Buckhout, T. J. (2001). Two iron-regulated cation transporters from tomato complement metal uptake-deficient yeast mutants. Plant Mol. Biol. 45, 437–448. doi: 10.1023/A:1010620012803
Ekesa, B., Nabuuma, D., Gand, K., van den Bergh, I. (2017). Sensory evaluation of provitamin A carotenoid-rich banana cultivars on trial for potential adoption in Burundi and Eastern democratic republic of Congo. Fruits 72, 261–272. doi: 10.17660/th2017/72.5.1
Fitzpatrick, T. B., Basset, G. J., Borel, P., Carrari, F., Della, P. D., Fraser, et al. (2012). Vitamin deficiencies in humans: can plant science help? Plant Cell 24, 395–414. doi: 10.1105/tpc.111.093120
Flachowsky, H., Szankowski, I., Fischer, T. C., Richter, K., Peil, A., Hofer, M., et al. (2010). Transgenic apple plants overexpressing the Lc gene of maize show an altered growth habit and increased resistance to apple scab and fire blight. Planta 231, 623–635. doi: 10.1007/s00425-009-1074-4
Flores-Félix, J. D., Silva, L. R., Rivera, L. P., Marcos-García, M. (2015). Plants probiotics as a tool to produce highly functional fruits: The Case of Phyllobacterium and vitamin C in strawberries. PloS One 10, e0122281. doi: 10.1371/journal.pone.0122281
Food and Agriculture Organization (FAO) (2020). Food and agriculture organization statistical database. Available online at: http://www.fao.org/faostat/en/data/QC.
Food and Agriculture Organization of the United Nations (FAO) (2023). Per capita consumption of fruits. Available online at: https://ourworldindata.org/grapher/fruit-consumption-by-fruit-type?tab=table.
Frassinetti, S., Bronzetti, G. L., Caltavuturo, L., Cini, M., Della Croce, C. (2006). The role of zinc in life: a review. J. Environ. Pathol. Toxicol. Oncol. 25, 597–610. doi: 10.1615/JEnvironPatholToxicolOncol.v25.i3.40
Gashgari, R., Selim, S., Abdel-Mawgoud, M., Warrad, M., Habeeb, T. H., Saleh, A. M., et al. (2020). Arbuscular mycorrhizae induce a global metabolic change and improve the nutritional and health benefits of pennyroyal and parsley. Acta Physiologiae Plantarum 42, 1–11. doi: 10.1007/s11738-020-03091-3
Gibson, R. S. (2007). The role of diet-and host-related factors in nutrient bioavailability and thus in nutrient-based dietary requirement estimates. Food Nutr. Bull. 28, 77–100. doi: 10.1177/15648265070281S108
Guevara, J. E. M., Sanches, A. G., Pedrosa, V. M. D., Cruz, M. C. P., Silva, J. A., Teixeira, G. H. (2021). The injection of zinc sulfate into banana tree pseudostem can triple the zinc content and it is an effective method for fruit biofortification. J. Food Compos Anal. 102, 104020. doi: 10.1016/j.jfca.2021.104020
Hallberg, L., Hulthen, L. (2000). Prediction of dietary iron absorption: an algorithm for calculating absorption and bioavailability of dietary iron. Am. J. Clin. Nutr. 71, 1147–1160. doi: 10.1093/ajcn/71.5.1147
Han, Y., Vimolmangkang, S., Soria-Guerra, R. E., Rosales-Mendoza, S., Zheng, D., Lygin, A. V., et al. (2016). Overexpression of the apple anthocyanidin synthase gene in pear (Pyruscommuniscv. ‘White Doyenne’) induces visible changes in fruit color. Plant Cell Tissue Organ Culture 127, 641–650.
HarvestPlus (2022). Biofortified crops released. Available online at: https://bcr.harvestplus.org/varieties_released_list?crop.
Hell, R., Stephan, U. W. (2003). Iron uptake, trafficking and homeostasis in plants. Planta 216, 541–551. doi: 10.1007/s00425-002-0920-4
Hussain, A., Rizwan, M., Ali, S., Rehman, M. Z. U., Qayyum, M. E., Nawaz, R., et al. (2021). Combined use of different nanoparticles effectively decreased cadmium (Cd) concentration in grains of wheat grown in a field contaminated with Cd. Ecotoxicology Environ. Saf. 215, 112139. doi: 10.1016/j.ecoenv.2021.112139
Iqbal, M., Bakshi, P., Kumar, R., Wali, V. K., Bhushan, B. (2015). Influence of mulching on fruit quality of aonla (EmblicaofficinalisGaertn.) cv. NA-7. Ecology Environ. Conserv. 21, 263–268.
Jain, S. (2024). RNA interference technology for developing virus-resistant cassava. Pomology Encyclopedia, 223–231.
Jiang, J. Z., Zhang, S., Liu, L., Sun, B. M. (2018). A microscopic experimental study for nanoparticle motion for the enhancement of oxygen absorption in nanofluids. Nanotechnol. Rev. 7 (6), 529–539. doi: 10.1515/ntrev-2018-007
Johnson, A. A., Kyriacou, B., Callahan, D. L., Carruthers, L., Stangoulis, J., Katona, P. (2008). The interaction between nutrition and infection. Clin. Infect. Dis. 46, 1582–1588. doi: 10.1086/587658
Joshi, V., Nimmakayala, P., Song, Q., Abburi, V., Natarajan, P., Levi, A., et al. (2021). Genome-wide association study and population structure analysis of seed-bound amino acids and total protein in watermelon. PeerJ 9, e12343. doi: 10.7717/peerj.12343
Joshi, S., Yadav, P. K., Wusirika, R., Xu, L., Tang L and Avula, B. (2017a). Biofortification of banana to enhance its micronutrient content and tolerance to abiotic stresses. Front. Plant Sci. 8, 1–17.
Joshi, S., Yadav, P. K., Wusirika, R., Xu, L., Tang L and Avula, B. (2017b). Biofortification of banana to enhance its micronutrient content and tolerance to abiotic stresses. Front. Plant Sci. 8 p, 1–17.
Juliana, P., Govindan, V., Crespo-Herrera, L., Mondal, S., Huerta-Espino, J., Shrestha, S., et al. (2022). Genome-wide association mapping identifies key genomic regions for grain zinc and iron biofortification in bread wheat. Front. Plant Sci. Vol 13. doi: 10.3389/fpls.2022.903819
Kalcsits, L., Lotze, E., Tagliavini, M., Hannam, K. D., Mimmo, T., Neilsen, D., et al. (2020). Recent achievements and new research opportunities for optimizing macronutrient availability, acquisition, and distribution for perennial fruit crops. Agronomy 10, 1–28. doi: 10.3390/agronomy10111738
Khan, M. K., Pandey, A., Hamurcu, M., Gezgin, S., Athar, T., Rajput, V. D., et al. (2021). Insight into the prospects for nanotechnology in wheat biofortification. Biology 10, 1123. doi: 10.3390/biology10111123
Kim, S. H., Kim, Y. H., Ahn, Y. O., Ahn, M. J., Jeong, J. C., Lee, H. S., et al. (2013). Downregulation of the lycopene ϵ-cyclase gene increases carotenoid synthesis via the β-branch-specific pathway and enhances salt-stress tolerance in sweetpotato transgenic calli. Physiol. Plant 147, 432–432. doi: 10.1111/j.1399-3054.2012.01688.x
Kiran, A., Wakeel, A., Mahmood, K., Mubaraka, R., Hafsa, Haefele, S. M. (2022). Biofortification of staple crops to alleviate human malnutrition: Contributions and potential in developing countries. Front. Nutr. 12, 452. doi: 10.3390/agronomy12020452
Kumar, R., Bakshi, P., Srivastava, J. N., Sarvanan, S. (2012). Influence of plant growth regulators on growth, yield and quality of strawberry (Fragaria × ananassa Duch) cv. Sweet Charlie. Asian J. Horticulture 7, 40–43.
Larsen, E. H., Lobinski, R., Burger-Meÿer, K., Hansen, M. (2006). Uptake and speciation of selenium in garlic cultivated in soil amended with symbiotic fungi (mycorrhiza) and selenate. Anal. Bioanal. Chem. 385 (6), 1098–1108. doi: 10.1007/s00216-006-0535-x
Lee, S., Persson, D. P., Hansen, T. H., Husted, S., Schjoerring, J. K., Kim, Y. S., et al. (2011). Bio-available zinc in rice seeds is increased by activation tagging of nicotianamine synthase. Plant Biotechnol. J. 9, 865–873. doi: 10.1111/j.1467-7652.2011.00606.x
Li, J., Hu, J., Ma, C., Wang, Y., Wu, C., Huang, J., et al. (2016). Uptake, translocation and physiological effects of magnetic iron oxide (γ-Fe2O3) nanoparticles in corn (Zea mays L.). Chemosphere 159, 326–334. doi: 10.1016/j.chemosphere.2016.05.083
Liu, M., Xu, M., Yu, H., Fu Tang, H. S., Ma, Q. (2023). Spraying ZnEDTA at high concentrations: An ignored potential for producing zinc-fortified pear (Pyrus spp.) fruits without causing leaf and fruitlet burns. Sci. Hortic. 322, 112380. doi: 10.1016/j.scienta.2023.112380
Liu, Q., Yang, F., Zhang, J., Liu, H., Rahman, S., Islam, S., et al. (2021). Application of CRISPR/cas9 in crop quality improvement. Int. J. Mol. Sci. 22, 4206. doi: 10.3390/ijms22084206
Lobato-Gomez, M., Hewitt, S., Capell, T., Christou, P., Dhingra, A., Giron-Calva, P. S. (2021). Transgenic and genome-edited fruits: background, constraints, benefits, and commercial opportunities. Horticulture Research8 1), 1–3. doi: 10.1038/s41438-021-00601-3
Lobo, V., Patil, A., Phatak, A., Chandra, N. (2010). Free radicals, antioxidants and functional foods: Impact on human health. Pharmacognosy Rev. 4, 118–126. doi: 10.4103/0973-7847.70902
Maggini, S., Maldonado, P., Cardim, P., Fernandez Newball, C., Sota Latino, E. (2017). Vitamins C, D and Zinc: synergistic roles in immune function and infections. Vitamin Miner. 6, 167. doi: 10.4172/2376-1318.1000167
Mahida, A., Tandel, Y. N., Patel, D. P., Kumar, M. (2023). Agronomic biofortification with zinc and iron to enhance nutrient concentrations in mango. Environ. Conserv. J. 24, 192–196. doi: 10.36953/ECJ.17432548
Manjunatha, S. B., Biradar, D. P., Aladakatti, Y. R. (2016). Nanotechnology and its applications in agriculture: A review. J. Farm Sci. 29, 1–3. doi: 10.4236/oalib.1102455"10.4236/oalib.1102455
Marques, E., Darby, H. M., Kraft, J. (2021). Benefits and limitations of non-transgenic micronutrient biofortification approaches. Agronomy 11, 464. doi: 10.3390/agronomy11030464
Marschner, P. (2012). “Rhizosphere biology,” in Marschner’s mineral nutrition of higher plants, 3rd Edn (Elsevier, London).
Maxwell, D. P., Vincelli, P., McNellis, T. W., Randles, J. W., Bauw, G. (2013). Advances in genetic transformation of papaya: Past, present and future. GM Crops Food 4, 1–7.
Mengist, M. F., Burtch, H., Debelo, H., Pottorff, M., Bostan, H., Nunn, C., et al. (2020). Bioaccessibility of phenolics in highbush blueberries: Variation among accessions and years. Sci. Rep. 10, 7311. doi: 10.1038/s41598-020-74385-3
Mengjiao, L., Xu, M., Yu, H., Fu, H., Tang, S., Ma, Q., et al. (2023). Spraying ZnEDTA at high concentrations: an ignored potential for producing zinc-fortified pear (Pyrus spp.) fruits without causing leaf and fruitlet burns. Scientia Horticulturae. 322. doi: 10.1016/j.scienta.2023.112380
Naqvi, S., Zhu, C., Farre, G., Ramessar, K., Bassie, L., Breitenbach, J., et al. (2009). Transgenic multivitamin corn through biofortification of endosperm with three vitamins representing three different classes. Proc. Natl. Acad. Sci. 106, 7762–7767. doi: 10.1073/pnas.0901412106
Navarro-Tarazaga, M. L., Gomez-Maqueo, A., Montilla-Bascon, G., Castillo, S., Miras-Moreno, B., Nurez-Gomez, D. (2016). Nutritional value and antioxidant capacity of ten Spanish persimmon cultivars (Diospyros kaki Thunb.). Int. J. Food Sci. Nutr. 67, 848–857.
Ofori, K. F., Antoniello, S., English, M. M., Aryee, A. N. A. (2022). Improving nutrition through biofortification–A systematic review. Front. Nutr. 9. doi: 10.3389/fnut.2022.1043655
Padmesh, P., Mohanan, K. V., Thomas, G. (2013). Evaluation of nutrient composition of mango (Mangifera indica L.) genotypes. Int. J. Fruit Sci. 13, 453–466.
Park, S. C., Kim, Y. H., Kim, S. H., Jeong, Y. J., Kim, C. Y., Lee, J. S., et al. (2015). Overexpression of the IbMYB1 gene in an orange-fleshed sweet potato cultivar produces a dual-pigmented transgenic sweet potato with improved antioxidant activity. Physiol. Plant 153, 525–527. doi: 10.1111/ppl.12281
Park, J. I., Lee, Y. K., Chung, W. I., Lee, I. H., Choi, J. H., Lee, W. M., et al. (2006). Modification of sugar composition in strawberry fruit by antisense suppression of an ADP-glucose pyrophosphorylase. Mol. Breed. 17, 269–279. doi: 10.1007/s11032-005-5682-9
Perez-Massot, E., Banakar, R., Gómez-Galera, S., Zorrilla-López, U., Sanahuja, G., Arjó, G. (2013). The contribution of transgenic plants to better health through improved nutrition: opportunities and constraints. Genes Nutr. 8, 29–41. doi: 10.1007/s12263-012-0315-5
Perkins, P., Collins, J. K., Davis, A. R. (2001). Lycopene content differs among red-fleshed watermelon cultivars. J. Sci. Food. Agric. 81 (10), 983–987. doi: 10.1002/jsfa.880
Pons, E., Alquezar, B., Rodriguez, A., Martorell, P., Genoves, S., Ramon, D., et al. (2018). Metabolic engineering of β-carotene in orange fruit increases its in vivo antioxidant properties. Plant Biotechnol. J. 12 (1), 17–27. doi: 10.1111/pbi.12112
Pradhan, S., Patra, P., Das, S., Chandra, S., Mitra, S., Dey, K. K., et al. (2013). Photochemical modulation of biosafe manganese nanoparticles on Vigna radiata: A detailed molecular, biochemical, and biophysical study. Environ. Sci. Technol. 47, 13122–13131. doi: 10.1021/es402659t
Prasad, B. V. G., Mohanta, S., Rahaman, S., Prerna (2015). Bio-fortification in horticultural crops. Journal of agricultural engineering and food technology. Bareily 2, 95–99.
Prasanna, R., Bidyarani, N., Babu, S., Hossain, F., Shivay, Y. S., Nain, L. (2015). Cyanobacterial inoculation elicits plant defence response and enhanced Zn mobilization in maize hybrids. Cogent Food Agric. 1, 998507. doi: 10.1080/23311932.2014.998507.00
Rai, P. K., Kumar, V., Lee, S., Raza, N., Kim, K. H., Ok, Y. S., et al. (2018). Nanoparticle-plant interaction: Implications in energy, environment, and agriculture. Environ. Int. 119, 1–19. doi: 10.1016/j.envint.2018.06.012
Ramesh, A., Sharma, S. K., Sharma, M. P., Yadav, N., Joshi, O. P. (2014). Inoculation of zinc solubilizing Bacillus aryabhattai strains for improved growth, mobilization, and biofortification of zinc in soybean and wheat cultivated in vertisols of central India. Appl. Soil Ecol. 73, 87–96. doi: 10.1016/j.apsoil.2013.08.009
Roriz, C. L., Carvalho, A. M., Castro, I., Vasconcelos, M. W. (2020). Legume biofortification and the role of plant growth-promoting bacteria in a sustainable agricultural era. Agronomy 10 (3), 435. doi: 10.3390/agronomy10030435
Sahoo, R. K., Bhardwaj, D., Tuteja, N. (2013). “Biofertilizers: A sustainable eco-friendly agricultural approach to crop improvement,” in Plant acclimation to environmental stress (Springer, New York, NY, USA), 403–432.
Salonia, F., Ciacciulli, A., Poles, L., Pappalardo, H. D., Malfa, S. L., Licciardello, C. (2020). New plant breeding techniques in citrus for the improvement of important agronomic traits: A review. Frontiers 11. doi: 10.3389/fpls.2020.01234
Sancenon, V., Puig, S., Mira, H., Thiele, D. J., Penarrubia, L. (2003). Identification of a copper transporter family in Arabidopsis thaliana. Plant Mol. Biol. 51, 577–587. doi: 10.1023/A:1022345507112
Saranya, P., Vijayan, K., Rajeswari, P., Rajam, M. V. (2017). Biofortification of mango (Mangifera indica L.) with betacarotene by genetic transformation. Scientia Hortic. 225, 217–224.
Sathya, S., Selvi, A., Manivannan, V., Balamohan, T. (2019). Influence of nutrient management practices on growth and yield of guava cv. Lucknow-49 under sub-humid condition of southern India. Int. J. Curr. Microbiol. Appl. Sci. 8, 2102–2110.
Sayre, R., Beeching, J. R., Cahoon, E. B., Egasi, C., Fauquet, C., Fregene, M., et al. (2011). The BioCassava plus program: biofortification of cassava for sub-Saharan Africa. Annu. Rev. Plant Biol. 62, 251–252. doi: 10.1146/annurev-arplant-042110-103751
Shah, R. A., Bakshi, P., Jasrotia, A., Itoo, H., Padder, B. A., Anjum, I., et al. (2023). A review. Morphological to molecular markers, plant genetic diversity studies in walnut (Juglans regia L.). Erwerbs-Obstbau 65, 1499–1511. Springer. doi: 10.1007/s10341-023-00892-x
Shankar, A. V., Antony, U., Bhaskarachary, K., Shetty, P. H. (2010). Carotenoid composition and vitamin A value in commercially available papaya cultivars grown in Hawaii. J. Food Composition Anal. 23, 222–227.
Sharma, P., Aggrwal, P., Kaur, A. (2017). Biofrotifcation: a new approach to eradicate hidden hunger. Food Rev. Intl. 33, 1–21. doi: 10.1080/87559129.2015.1137309
Sharma, A., Johri, B. N. (2003). Growth-promoting influence of siderophore-producing Pseudomonas strains GRP3A and PRS9 in maize (Zea mays L.) under iron-limiting conditions. Microbiological Res. 158, 243–248. doi: 10.1078/0944-5013-00197
Shekhawat, U. K. S., Ganapathi, T. R., Hadapad, A. B. (2012). Transgenic banana plants expressing small interfering RNAs targeted against viral replication initiation gene display high-level resistance to banana bunchy top virus infection. J. Gen. Virol. 93, 1804–1813. doi: 10.1099/vir.0.041871-0
Singh, G., Bakshi, P., Kour, K., Sharma, N., Sinha, B. K., Kumar, R., et al. (2022). Effect of pollination methods on fruiting of different strawberry cultivars. Agric. Mechanization Asia Afr. Latin America 53, 9365–9372.
Strand, M., Hörnell, A., Byrnelius, H., Wold, A. B., Bjorck, I. (2018). Identification of apple cultivars with high levels of antioxidants and their implications for human health. PloS One 13, e0197989.
Tagliavini, M., Abadía, J., Marangoni, B. (2019). Fertilizer management effects on fruit composition in apple: A review. Horticulturae 5, 62.
Telengech, P. K., Malinga, J. N., Nyende, A. B., Gichuki, S. T., Wanjala, B. W. (2015). Gene expression of beta carotene genes in transgenic biofortified cassava. Biotech. 5, 465–462. doi: 10.1007/s13205-014-0243-8
Thakur, V., Sharma, A., Sharma, P., Kumar, P., Shilpa (2022). Improving vegetable crops for vitamins, minerals, and other quality traits. J. Hortic. Sci. Biotechnol. 97, 417–428. doi: 10.1080/14620316.2022.2036254
Thakur, S., Thakur, T., Kumar, R. (2018). Bio-nanotechnology and its role in agriculture and food industry. J. Mol. Genet. Med. 12, 1–5.
Tian, L., Duan, X., Wang, P. (2017). Bioengineering of the apple stilbene pathway for production of resveratrol in apple fruit. Plant Biotechnol. J. 15, 367–378.
Torre-Roche, R. D. L., Cantu, J., Tamez, C., Zuverza-Mena, N., Hamdi, H., Elmer, W., et al. (2020). Seed biofortification by engineered nanomaterials: A pathway to alleviate malnutrition? J. Agric. Food Chem. 4, 12189–12202. doi: 10.1021/acs.jafc.0c04881
Van, D., Fitzpatrick, T. B., De Steur, H. (2017). Editorial overview: Biofortification of crops: achievements, future challenges, socio-economic, health and ethical aspects. Curr. Opin. Biotechnol. 44, 1–4. doi: 10.1016/j.copbio.2017.03.007
Vezzulli, S., Bavaresco, L., Lovisolo, C. (2007). Skin pectin metabolism during grapevine berry ripening. Funct. Plant Biol. 34, 889–898.
Waltz, E. (2014). Vitamin A super banana in human trials. Nat. Biotechnol. 322023), 857. doi: 10.1038/nbt0914-857World Population Review
White, P. J., Broadley, M. R. (2003). Calcium in plants. Ann. Bot. 92, 487–511. doi: 10.1093/aob/mcg164
World Population Review. (2023). Available online at: https://www.nab.com.na/wp-content/uploads/2024/04/Market-Intelligence-Report-Bananas-revised-NAB-29032024.pdf.
White, P. J., Broadley, M. R. (2009). Biofortification of crops with seven mineral elements often lacking in human diets—iron, zinc, copper, calcium, magnesium, selenium, and iodine. New Phytol. 182, 49–84. doi: 10.1111/j.1469-8137.2008.02738.x
Yadava, D., Choudhury, P., Hossain, F., Kumar D and Mohapatra, T. (2020). Biofortified varieties. Sustainable way to alleviate malnutrition. 3rd ed. (New Delhi: Indian Council of Agricultural Research), 86.
Yu, L., Nie, Y., Jiao, J., Jian, L., Zhao, J. (2021). The sequencing based mapping method for effectively cloning plant mutated genes. Int. J. Mol. Sci. 22, 6224. doi: 10.3390/ijms22126224
Zhang, Y., Fu, X., Li, J., Zhu B and Tian, S. (2019). Iron accumulation in transgenic citrus fruits expressing Arabidopsis ferritin gene AtFer1. Front. Plant Sci. 10, 1514. doi: 10.3389/fpls.2019.01090
Zhang, F., Shen, J., Zhang, J., Zuo, Y., Li, L., Chen, X. (2010). Rhizosphere processes and management for improving nutrient use efficiency and crop productivity: implications for China. Adv. Agron. 107, 1–32. doi: 10.1016/S0065-2113(10)07001-X
Zhang, M., Xue, C., Hu, H., Li, J., Xue, Y., Wang, R., et al. (2021). Genome-wide association studies provide insights into the genetic determination of fruit traits of pear. Nat. Commun. 12, 1144. doi: 10.1038/s41467-021-21378-y
Zhang, W. H., Zhou, Y. C., Dibley, K. E., Tyerman, S. D., Furbank, R. T., Patrick, J. W. (2007). Nutrient loading of developing seeds. Funct. Plant Biol. 34, 314–331. doi: 10.1071/FP06271
Keywords: biofortification, underdeveloped nations, gross domestic product, climate change, malnutrition, hidden hunger, conventional etc
Citation: Jha A, Jayswal DK, Shikha D, Kumar A and Ahmad F (2025) Empowering vital fruit crops with enhanced nutritional contents. Front. Plant Sci. 16:1519673. doi: 10.3389/fpls.2025.1519673
Received: 30 October 2024; Accepted: 07 January 2025;
Published: 24 February 2025.
Edited by:
Anoop Kumar Srivastava, Central Citrus Research Institute (ICAR), IndiaReviewed by:
Heiplanmi Rymbai, ICAR Research Complex for NEH Region, IndiaCopyright © 2025 Jha, Jayswal, Shikha, Kumar and Ahmad. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: D. K. Jayswal, ZGtqYXlzd2FsLnBvbW9sb2dpc3RAZ21haWwuY29t
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.