 Mônica N. Alves1,2
Mônica N. Alves1,2 Juan Cifuentes-Arenas2
Juan Cifuentes-Arenas2 Regina Niñoles3
Regina Niñoles3 Laudecir Lemos Raiol-Junior1
Laudecir Lemos Raiol-Junior1 Everton Carvalho1,4Isabel Quirós-Rodriguez3
Everton Carvalho1,4Isabel Quirós-Rodriguez3 Jesus A. Ferro1
Jesus A. Ferro1 Concetta Licciardello5
Concetta Licciardello5 Berta Alquezar2,3
Berta Alquezar2,3 Lourdes Carmona3Javier Forment3
Lourdes Carmona3Javier Forment3 Aureliano Bombarely3
Aureliano Bombarely3 Nelson A. Wulff2
Nelson A. Wulff2 Leandro Peña2,3*
Leandro Peña2,3* José Gadea3*
José Gadea3*- 1Universidade Estadual Paulista (Unesp), Faculdade de Ciências Agrárias e Veterinárias (FCAV), Jaboticabal, SP, Brazil
- 2Fundo de Defesa da Citricultura (Fundecitrus), Araraquara, SP, Brazil
- 3Instituto de Biologia Molecular y Celular de Plantas (IBMCP), Universidad Politécnica de Valencia (UPV-CSIC), Valencia, Spain
- 4Helix Sementes e Biotecnologia, Patos de Minas, MG, Brazil
- 5Research Center for Olive Fruit and Citrus Crops, Council for Agricultural Research and Economics, Acireale, Italy
Huanglongbing (HLB) is a devastating disease of citrus plants caused by the non-culturable phloem-inhabiting bacterium Candidatus Liberibacter ssp., being Ca. Liberibacter asiaticus (CLas) the most aggressive species. CLas is vectored by the psyllid Diaphorina citri and introduced into sieve cells, establishing a successful infection in all Citrus species. Partial or complete resistance has been documented in the distant relatives Murraya paniculata and Bergera koenigii, respectively, providing excellent systems to investigate the molecular basis of HLB-resistance. It has been shown previously that the first weeks after bacterial release into the phloem are critical for the establishment of the bacterium. In this study, a thorough transcriptomic analysis of young flushes exposed to CLas-positive and negative psyllids has been performed in Citrus × sinensis, as well as in the aforementioned resistant species, along the first eight weeks after exposure. Our results indicate that the resistant species do not deploy a classical immunity response upon CLas recognition. Instead, transcriptome changes are scarce and only a few genes are differentially expressed when flushes exposed to CLas-positive and negative psyllid are compared. Functional analysis suggests that primary metabolism and other basic cellular functions could be rewired in the resistant species to limit infection. Transcriptomes of young flushes of the three species are very different, supporting the existence of distinct biochemical niches for the bacterium. These findings suggest that both intrinsic metabolic inadequacies to CLas survival, as well as inducible reprogramming of physiological functions upon CLas recognition, could orchestrate together restriction of bacterial multiplication in these resistant hosts.
Introduction
During the last decade, compelling models of plant–pathogen interactions have been established, mostly based on molecular studies of mesophyll cell-infecting pathogens (Dodds and Rathjen, 2010; Asai and Shirasu, 2015; Ceulemans et al., 2021). Local immune activation involves the activity of two classes of receptors: plasma membrane-localized receptors that typically recognize conserved microbial patterns in the apoplast, named as pathogen-associated-molecular-patterns (PAMPs). PAMPs detection constitutes the first tier of active plant immunity, which involves an influx of extracellular Ca2+ into the cytosol, an oxidative burst, and a massive reprogramming of the transcriptome in the first hours after PAMP recognition. Some pathogens overcome this PAMP-triggered immunity (PTI) by secreting effectors that manipulate cellular processes and suppress PTI. In resistant plants, targeted recognition of these effectors by cytoplasmic receptors forms the second tier of active plant immunity, called effector-triggered immunity (ETI). Both responses are triggered very quickly, ensuring, for many interactions, the complete deployment of defenses withing the first days after pathogen detection. The ability of a pathogen to overcome these two defensive lines within this time frame determines its success in establishing the infection (Thomma et al., 2011; Bigeard et al., 2015; Remick et al., 2023).
Some of the most devastating plant pathogens, however, do not infect mesophyll cells. They are delivered into the sieve cells of the phloem by feeding insects using specialized mouth parts called stylets, which pierce the plant tissue and directly access the nutrient-rich phloem vessels. The stylets travel through the apoplast to reach the phloem, secreting saliva to form a hard sheath that seals off plant cell leaks caused by the penetration process, causing limited wounding to the plant. Thus, those pathogens are directly delivered to the sieve cell cytoplasm, which lacks a nucleus, and its potential to mount an effective immune response is uncertain (Jiang et al., 2019; Huang et al., 2020). In addition, the high genome reduction rate of these pathogens, which makes them very dependent on host metabolism for survival, implies a lack of typical transmembrane systems for secretion of effectors (Duan et al., 2009; Oshima et al., 2013; Huang et al., 2020; Rattner et al., 2021). Under this scenario, which aspects of the aforementioned plant immunity are applicable to these interactions remain unknown.
One of the most ravaging diseases caused by phloem-inhabiting prokaryotic pathogens is Huanglongbing (HLB), also known as citrus greening. Since 2004 and 2005, when it was respectively detected in São Paulo (Brazil) and in Florida (USA), two of the most significant citrus-growing regions worldwide, it has caused billions of dollars in losses. In Brazil, despite successful management strategies adopted in the last years, the HLB incidence in commercial groves continues to increase, now affecting more than 44% of sweet orange trees in the citrus belt of São Paulo and Minas Gerais states (Fundecitrus, 2024). In Florida, the citrus industry relies on the limited production of more than 99% of infected trees (Li et al., 2020b). The disease has spread to almost all citrus-producing regions worldwide, except Australia and Mediterranean Basin.
The predominant HLB pathogen is the non-culturable, gram-negative α-proteobacterium ‘Candidatus Liberibacter asiaticus’ (CLas), which is mainly vectored by the Asian citrus psyllid (ACP) Diaphorina citri (Kuwayama) (Sternorrhyncha: Psyllidae) (Burckardt et al., 2021). Foliar symptoms include asymmetrical chlorosis known as blotchy mottle, enlarged veins, and intense canopy defoliation (Bové, 2006), caused by disturbance of the transport system between source and sinks, with starch accumulation in plastids, chloroplast disruption, and plugging of sieve cells via callose deposition (Bové, 2006; Andrade et al., 2019; Pandey and Wang, 2019). In field trees, these symptoms appear within some months to years after infection, depending on the tree’s age and size (Gottwald, 2010; Lopes et al., 2009). During the asymptomatic phase, infected plants remain indistinguishable from CLas-free plants and are an important source for the disease to spread within the grove, which complicates disease management (Lopes et al., 2009; Lee et al., 2015). How CLas can evade immunity and remain undetected by its host for such a long period is still unknown. CLas does not possess the conserved secretion mechanism to deliver effectors into plant cells, although most components needed to generate a functional Sec-dependent secretion system are present in its genome (Duan et al., 2009). The Sec-dependent secretion system has been hypothesized to be essential for pathogens like CLas or phytoplasmas to infect plants (Sugio et al., 2011; Yan et al., 2013) and it has been reported that CLas genes belonging putatively to this Sec-dependent system are upregulated in planta, but not in CLas-infected psyllids (Yan et al., 2013).
Although sensitivity to CLas varies among Citrus species and varieties, no complete resistance has been documented in this genus (Folimonova et al., 2009). However, Oceanian Citrus species formerly included by Swingle and Reece (1967) within the Microcitrus and Eremocitrus genera have been reported to be resistant to CLas (Ramadugu et al., 2016; Alves et al., 2021b, Alves et al., 2022). Moreover, partial or complete resistance has been reported in certain related genera within the family Rutaceae subfamily Aurantioideae (Ramadugu et al., 2016; Beloti et al., 2018; Cifuentes-Arenas et al., 2019; Alves et al., 2021a, Alves et al., 2021b, Alves et al., 2022). The monoembryonic orange jasmine (Murraya paniculata (L.) Jack) and curry leaf (Bergera koenigii L.), are good hosts for ACP (Damsteegt et al., 2010; Teck et al., 2011; Westbrook et al., 2011), being even more attractive to ACP females than sweet orange (Beloti et al., 2017; Tomaseto et al., 2019; Eduardo et al., 2022). M. paniculata is considered a transient host for CLas while B. koenigii is CLas-immune (Damsteegt et al., 2010; Beloti et al., 2018; Cifuentes-Arenas et al., 2019), providing a unique opportunity to explore distinct molecular mechanisms potentially involved in HLB-resistance. Recently, in a simulation of the natural entry of CLas in the plant host, Alves et al. (2021a) studied CLas dynamics in sweet orange Citrus × sinensis (L.) Osbeck ‘Valencia’, in M. paniculata, and in B. koenigii in the first stages of these plant-bacteria interactions. Using a well-controlled challenge-inoculation system with infectious psyllids, critical time points for these plant species with different responses to CLas infection were identified. In all three experimental systems, CLas was successfully released into the phloem by ACP, and bacterial titers decreased during the first days after inoculation. Subsequent increases in bacterial titers were observed for each host: while an exponential increase occurred in C. × sinensis up to the 40th day post-inoculation (DAI), from which a plateau was reached, CLas failed to successfully replicate in B. koenigii. In M. paniculata, CLas titers were generally lower than in C. × sinensis, and titer dynamics resembled that of C. × sinensis up to the 30th DAI, from which bacterial titers progressively decreased to undetectable levels (Alves et al., 2021a). These observations suggest that the first weeks after bacterial release into the phloem by ACP are critical for the establishment of the bacterium in each host, providing an excellent system to investigate molecular mechanisms leading to either resistance or susceptibility.
Here, a thorough transcriptomic analysis of new flushes of C. × sinensis, B. koenigii and M. paniculata was performed to study host gene expression dynamics after the ACP-vectored transmission of CLas. Our results suggest that the different responses to the bacterium in the three species are not due to the deployment of classical immunity in the resistant species pointing to a mechanism in which primary metabolism and other basic cellular functions are rewired to limit bacterial infection.
Materials and methods
Plants and growth conditions
Rearing of insects and plant maintenance were carried out in a controlled environmental room with 55% and 78% relative humidity, and daily temperature of 24°C-27°C. Lights were 20-30 cm above the plants with a photoperiod of 12 h of light (300 µmol m-2 s-1) and 12 h of darkness. Plants were grown in 300 mL conical tubes (6.5 x 5.9 cm, upper x lower diameter; 16 cm, height), filled with coconut fiber. In total, 51 two-year-old plants of Citrus × sinensis (L.) Osbeck ‘Valencia’ grafted on C. × limonia Osbeck (‘Rangpur’ lime), 39 two-year-old seedlings of M. paniculata and 39 of B. koenigii were used.
Rearing of CLas-infected and CLas-negative psyllids
Insect rearing and inoculation followed the methodology described by Lopes and Cifuentes-Arenas (2021). CLas-positive (real-time polymerase chain reactions (qPCR) Ct average of 21.82 ± 0.79) ‘Valencia’ sweet oranges grafted on ‘Swingle’ citrumelo (C. × paradisi Macfad. × Poncirus trifoliata L. Raf.) were used for obtaining CLas-positive insects. Those CLas-infected plants were pruned and when new flushes reached the V2 stage (Cifuentes-Arenas et al., 2018), four 10 to 20-days-old CLas-negative psyllids were confined per shoot. After 7 days of oviposition, adults were removed, and the eggs developed into nymphs and hatched into adults (named F1). When these F1 adults reached ~5 days, CLas presence was assessed by qPCR in a random sampling of three insects per plant. The remaining insects were kept confined in the CLas-positive plants until they were 15 to 20 days-old, and used for challenge inoculation experiments. CLas-negative insects were obtained using the same methodology on qPCR-negative plants. Adult insects used for oviposition were obtained from a colony kept at Fundecitrus, in which insects are continuously reared on healthy M. paniculata seedlings (Skelley and Hoy, 2004).
Psyllid challenge inoculation
C. × sinensis ‘Valencia’ plants, M. paniculata and B. koenigii seedlings were pruned at 15-25 cm to promote new flushes to sprout. A V2 flush was selected, and the remaining ones were eliminated. Five 15-20day-old CLas-positive or negative adult psyllids were confined to a single flush per plant for 48 h (inoculation access period, IAP). Twenty-four ‘Valencia’ sweet orange plants and 18 of each M. paniculata or B. koenigii seedlings were exposed to CLas-positive psyllids and the same number to CLas-negative ones. After the IAP, adults were removed and stored at -20°C. Fifteen days after psyllid removal, all plants were sprayed with insecticide (Abamectin EC, 7.2 g of active ingredient per 1 L of water) to eliminate any eggs or nymphs present.
Plant sampling procedures
Three plants per species were randomly selected as controls prior to the confinement with the insect (called here as “prior”). Samples were randomly collected at 0 (t0, day of psyllid removal), 10 (t10), 20 (t20), 30 (t30), 60 (t60),120 (t120) and 180 (t180) days after psyllid removal. The tissue that was in contact with the psyllid was frozen in liquid nitrogen and stored at -80°C. Due to shoot’s growth over time, the total collected tissue varied between samples. Each plant was used at a single time point, with 5-8 plants being used per time point for each species.
Evaluation of CLas multiplication
Total DNA was extracted from 0.1 g of frozen tissue. Psyllid DNA was extracted as described by Murray and Thompson (1980), with modifications by Alves et al. (2021a). After DNA precipitation with 0.6 V of isopropanol, DNA was washed twice with 70% ethanol and resuspended in 50 uL of Milli-Q® water. CLas presence was assayed by qPCR using CLas 16S rRNA gene sequence, using 100 ng of total DNA, TaqMan® PCR Master Mix (1x) (Invitrogen, Carlsbad, CA, United States), and HLBas primer-probe (0.5 µM/0.2 µM) in a StepOnePlus thermocycler (Applied Biosystems, California, USA) (Li et al, 2006). The mitochondrial gene cytochrome oxidase (COX) and a psyllid wingless (wg) gene region were used as internal controls to assess plants and psyllid DNA quality, respectively (Li et al., 2006; Manjunath et al., 2008). For CLas quantification, the linear relationship between the cycle threshold (Ct) and the 16S rRNA log was used (Lopes et al., 2013). Samples were considered CLas-positive when the Ct was lower than 34.0.
RNA extraction and sequencing
Samples for transcriptomic analysis were taken before the exposure to psyllids (prior), just after removal of the psyllids (t0) and after 10 (t10), 20 (t20), 30 (t30) and 60 (t60) days. Three individual flushes per species and time-point, which were exposed to either CLas-positive or negative psyllids, were selected. Each one was considered a biological replicate. Additionally, for time-point t10, three more replicates per species were used. Total RNA was extracted using the RNeasy Mini Kit (Qiagen, Valencia, CA, USA).
Total RNA concentration was calculated by Quant-IT RiboGreen (Invitrogen, #R11490). To assess the integrity of the total RNA, samples are run on the TapeStation RNA screentape (Agilent, #5067-5576). Only high-quality RNA preparations, with RIN greater than 7.0, were used for RNA library construction. A library was independently prepared with 0.5ug of total RNA for each sample by Illumina TruSeq Stranded Total RNA Library Prep Plant Kit (Illumina, Inc., San Diego, CA, USA, # 20020611). The first step involves removing the rRNA in the total RNA. Following this step, the remaining mRNA is fragmented into small pieces using divalent cations under elevated temperature. The cleaved RNA fragments are copied into first strand cDNA using SuperScript II reverse transcriptase (Invitrogen, #18064014) and random primers. Second strand cDNA synthesis was performed using DNA Polymerase I, RNase H and dUTP. These cDNA fragments were repaired by addition of a single ‘A’ base, and ligation of the adapters. The products are purified and enriched with PCR to create the final cDNA library. The libraries were quantified using KAPA Library Quantification kits for Illumina Sequencing platforms according to the qPCR Quantification Protocol Guide (KAPA BIOSYSTEMS, #KK4854) and qualified using the TapeStation D1000 ScreenTape (Agilent Technologies, # 5067-5582). Indexed libraries were then submitted to an Illumina NovaSeq (Illumina, Inc., San Diego, CA, USA), and the paired-end (2×150) sequencing was performed by the Macrogen Incorporated.
Bergera koenigii transcriptome assembly
B. koenigii transcriptome was assembled with Trinity v2.15.2 (Grabherr et al., 2011) using default parameters. Transcripts were collapsed into SuperTranscripts using the corresponding script from the Trinity package. Analysis of gene space completeness was performed with BUSCO “transcripts” mode (Manni et al., 2021) and the Viridiplantae Odb10 dataset. Presence of contaminants was assessed using Blobtools, version 1.1.1 (Challis et al., 2020). Reads were remapped against the created supertranscripts using the BWA (Burrows-Wheeler Aligner) program v.0.7.17-r1188 (Li and Durbin, 2009). Then, bam files were merged and indexed using Samtools (Li et al., 2009). Finally, Diamond Blastx against Uniprot Trembl database was run (Buchfink et al., 2015) and a Blobtools database was created. Results were filtered by removing sequences not associated with the Streptophyta Tag.
Transcriptome analysis
Adapter removal and quality trimming of raw reads was done with Cutadapt v3.7 (Martin, 2011). Clean reads were analyzed using FastQC v0.11.9 (Andrews, 2010). Clean read pairs longer than 20 nt were mapped to the combined genome of Candidatus Liberibacter asiaticus (https://www.ncbi.nlm.nih.gov/nuccore/NZ_CP019958) with either the C. × sinensis SWO.v3.0 (http://citrus.hzau.edu.cn/download.php), or the M. paniculata genome assembly (provided by Dr. Concetta Licciardello, Council for Agricultural Research and Economics, CREA), or the Bergera koenigii de novo transcriptome assembly (this study, see above) using STAR (Dobin et al., 2013). Number of reads mapped to one and only one of the annotated genes of the genome (uniquely mapped) was also obtained with STAR. In total, more than 855 million uniquely-mapped gene-assigned reads were obtained for C. × sinensis, 742 million for M. paniculata and 1376 million for B. koenigii (Supplementary Table 1). Differential expression analysis was performed with DESeq2 (Love et al., 2014). Principal component analysis was done on the NetworkAnalyst 3.0 suite (Zhou et al., 2019). Gene Set Enrichment Analysis (GSEA) (Subramanian et al., 2005) was used for pairwise comparisons within a species and Over-Representation Analysis (ORA) (Boyle et al., 2004) for pairwise comparisons among orthologous groups. GO categories were summarized by removing redundant terms according to ReviGO (Supek et al., 2011). Categories with frequency < 5 were selected and ranked by uniqueness. Bubble and UpSet plots were done using SRplot (Tang et al., 2023).
Results
CLas establishment and shoot ontogeny differs between C. × sinensis, M. paniculata and B. koenigii
After the 48h IAP (t0), psyllid survival rate (avg. 98.5%) and CLas titer in psyllids (avg. 5.1 ± 0.15 log cells/g of tissue) were similar regardless of the plant species (Supplementary Table 2). Similarly, the amount of initial bacterial load released in the plant vasculature after IAP did not differ between species, with averages of 2.42 ± 0.12 log cells/g of tissue, which is close to the limit of detection of the qPCR technique employed (Figure 1A; Supplementary Table 3). This was followed by a steep decrease up to t20. Subsequently, in C. × sinensis, CLas titer increased exponentially, until t30, increasing ~1 log until t60. In M. paniculata, the initial decrease was gradual, as well as the subsequent exponential increase until t30. Maximum titer was observed at t60, slowly decreasing thereafter. In B. koenigii, a drastic reduction in CLas titer following psyllid removal was observed at t10, and the subsequent increase noticed for the other two species from t20 onwards was not observed (Figure 1A).
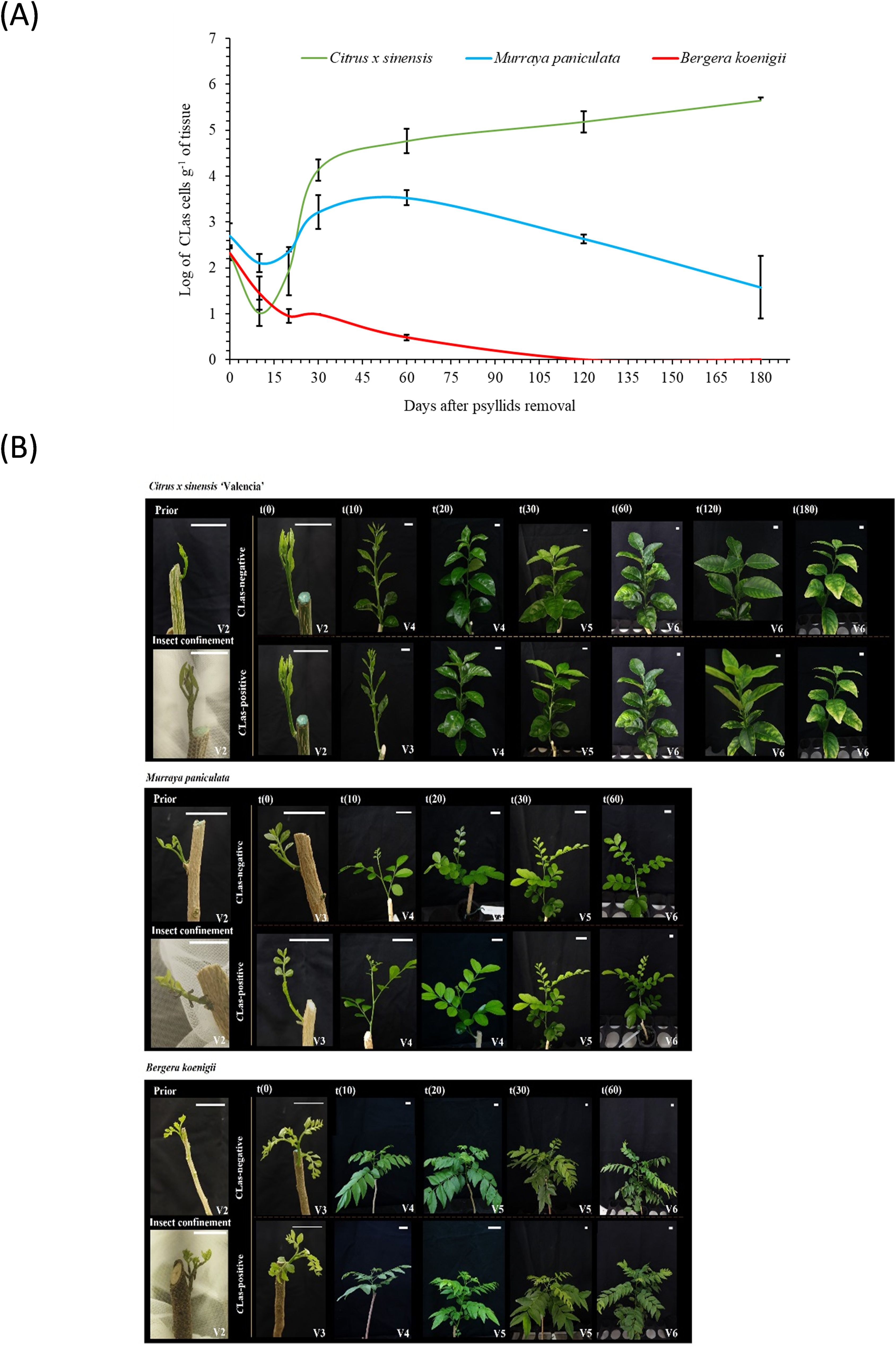
Figure 1. Shoot ontogeny and ‘Candidatus Liberibacter asiaticus’ population dynamics. (A) Evolution of the ‘Candidatus Liberibacter asiaticus’ population dynamics over time in new shoots of Citrus × sinensis, Murraya paniculata, and Bergera koenigii, following a 48 h period of CLas-exposed Diaphorina citri confinement. (B) Representative aspect of shoots from Citrus × sinensis, (top panels) Murraya paniculata, (middle panels) and Bergera koenigii (bottom panels) prior to insect confinement, at the confinement day, 48 h after exposure to CLas-positive or CLas-negative Diaphorina citri (t0) and 10 (t10), 20 (t20), 30 (t30), 60 (t60), 120 (t120) and 180 (t180) days after insect removing. White bar length in each photo corresponds to 2 cm.
Flush ontogeny along the experiment was determined according to Cifuentes-Arenas et al. (2018). The three species followed different ontogeny patterns, and the presence of the bacterium affected their development differently. C. × sinensis V2 flushes exposed to CLas-negative psyllids entered the maturation phase (V4) at t10 with leaves fully expanded at t30 (V5), and dormancy stage (V6) from t60 onwards. Flushes exposed to CLas-infected psyllids remained in developmental phase (V3) at t10, reached maturation phase (V4) at t20 and then followed the same dynamics as CLas-negative flushes. For M. paniculata and B. koenigii, developmental patterns were different to those of C. × sinensis. In both species and for either CLas-negative or positive flushes, initial V2 flushes entered the developmental phase (V4) at t10 with fully-expanded leaves at t20 (V5), earlier than in C. × sinensis (Figure 1B).
Transcriptomes of C. × sinensis, M. paniculata and B. koenigii are dramatically reprogrammed during flush growth
A thorough transcriptomic experiment was designed to evaluate the response to CLas in the three species. C. × sinensis and M. paniculata clean reads were mapped against the C. × sinensis SWO.v3.0 genome or the M. paniculata genome. Cross-mapping of B. koenigii clean reads against these two genomes revealed mapping percentages of 55% and 43%, respectively. Therefore, a de novo B. koenigii transcriptome was assembled using the whole set of reads obtained in this study. This assembly yielded a total of 25200 supercontigs, with a genome completeness of 89%. This de novo transcriptome was used to map B. koenigii reads.
First, transcriptome changes occurring in flushes along the first 60 days after exposure to CLas-negative psyllids were analyzed. Overall, gene expression was dramatically altered as compared to that of flushes prior to the inoculation period in the three species (Figures 2A–C; Supplementary Figure 1). Massive transcriptome changes were first observed at t10, with thousands of differentially expressed genes (DEGs), and a substantial number of changes also found at later time points, when flushes were at V5 or V6 stages. Similarly, flushes exposed to CLas-positive psyllids also exhibited massive gene-expression changes at t10, except for flushes of B. koenigii exposed to CLas-positive psyllids, whose transcriptional reprogramming was mainly triggered at t20 (Supplementary Table 4). These results suggest that, in general, gene expression is highly mobilized upon flush growth in shoots exposed to ACP, regardless of whether they were exposed to CLas-negative or positive psyllids.
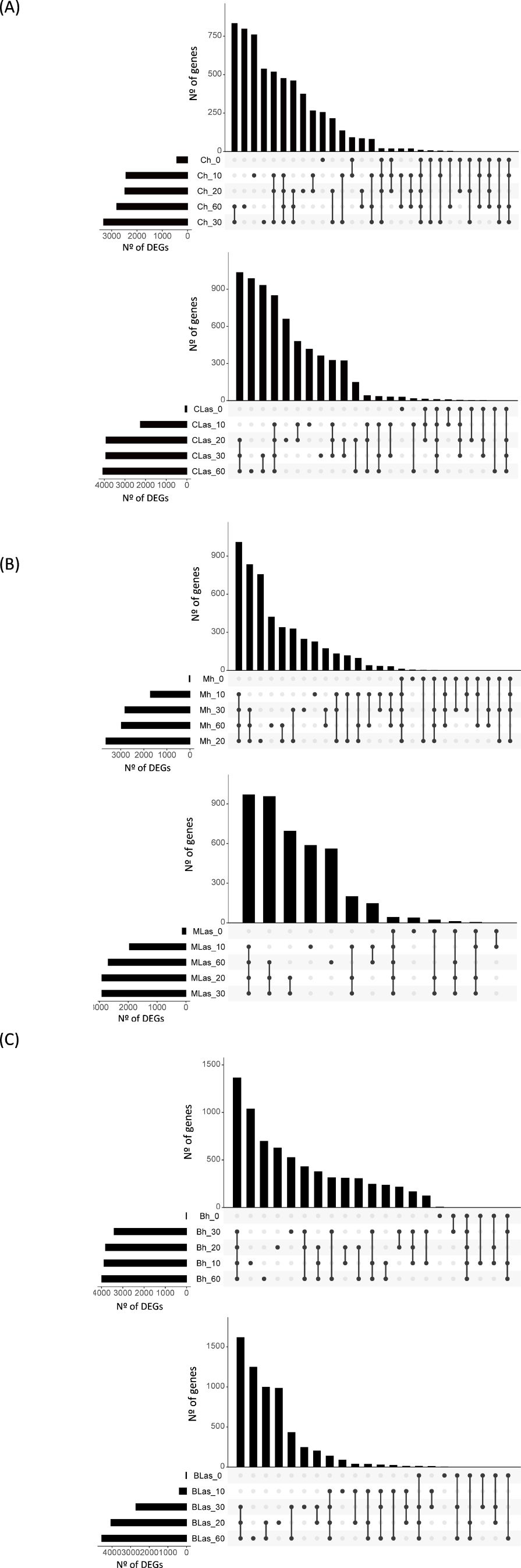
Figure 2. Transcriptome changes in Citrus ×sinensis, Murraya paniculata and Bergera koenigii flushes along the first 60 days after exposure to psyllids. (A) Number of up-regulated genes in Citrus × sinensis samples taken immediately (CLas_0, Ch_0), and after 10 (CLas_10, Ch_10), 20 (CLas_20, Ch_20), 30 (CLas_30, Ch_30) or 60 (CLas_60, Ch_60) days after exposure to CLas-negative (Ch, top) or CLas-positive (CLas, bottom) psyllids when compared against flushes before the exposure to psyllids (prior). (B) Number of up-regulated genes in Murraya paniculata samples taken immediately (MLas_0, Mh_0), and after 10 (MLas_10, Mh_10), 20 (MLas_20, Mh_20), 30 (MLas_30, Mh_30) or 60 (MLas_60, Mh_60) days after exposure to CLas-negative (Mh, top) or CLas-positive (MLas, bottom) psyllids when compared against flushes before the exposure to psyllids (prior). (C) Number of up-regulated genes in Bergera koenigii samples taken immediately (BLas_0, Bh_0), and after 10 (BLas_10, Bh_10), 20 (BLas_20, Bh_20), 30 (BLas_30, Bh_30) or 60 (BLas_60, Bh_60) days after exposure to CLas-negative (Bh, top) or CLas-positive (BLas, bottom) psyllids when compared against flushes before the exposure to psyllids (prior). CLas: Candidatus Liberibacter asiaticus. DEGs: Differentially expressed genes. In each graph, total number of DEGs (X axis) at each time point (Y axis) is shown in bottom-left panels. Intersection of sets of genes at multiple time points is shown in top right panels. Each column corresponds to a time point or set of time points (dots connected by lines below the X axis) containing the same DEGs. The time points shared are indicated in the graphic below the column, with the time points on the left.
C. × sinensis flush transcriptome is not dramatically altered upon CLas infection up to 60 days after inoculation
To identify differentially expressed genes (DEGs) in C. × sinensis upon CLas inoculation, paired-wise comparisons were performed between flushes exposed to CLas-negative or CLas-positive psyllids at every time-point. At t0, 167 genes were found differentially expressed after exposure to the bacterium, 123 genes at t10, 8 genes at t30 and 48 genes at t60 (Supplementary Table 5). This low number of differences was also reflected in PCA biplots shown in Figure 3, in which samples were roughly clustered according to flush developmental stage but not to exposure to CLas. Despite the low number of DEGs, some gene ontology categories were found enriched after gene-set-enrichment-analysis (GSEA) for every paired-wise comparison (Supplementary Table 6). A reduction of metabolic activity and growth-related processes in t0 flushes exposed to CLas was observed. Other enriched categories include an increase in DNA damage response in t10, in cell wall remodeling pathways in t30, and a decrease in translation and energy processes at t60. Our data suggest that the presence of the bacterium in C. × sinensis flushes did not cause massive transcriptional changes up to 60 days after inoculation.
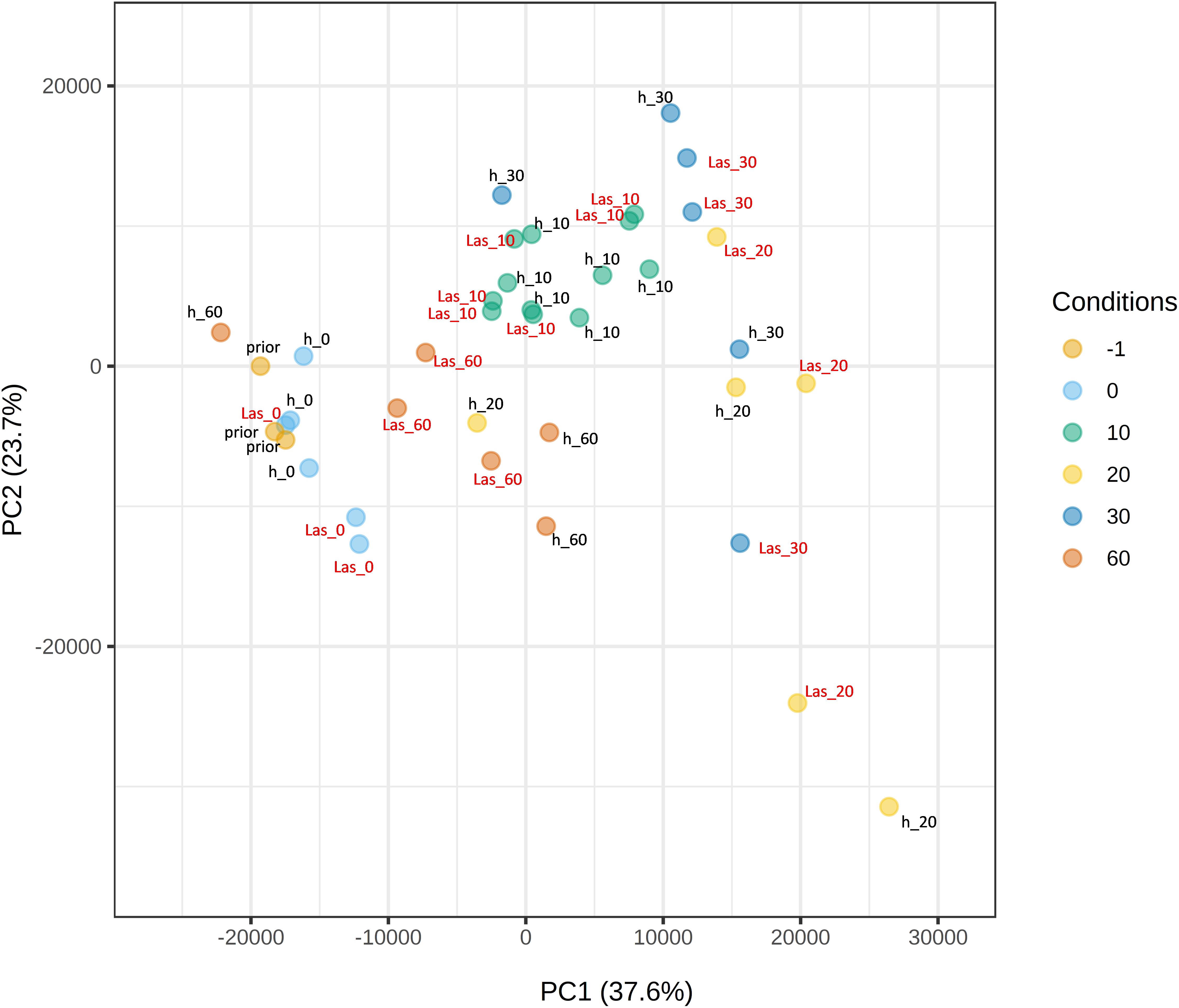
Figure 3. Principal Component Analysis (PCA) plot of Citrus × sinensis samples. Samples were taken before the exposure to psyllids (prior), immediately after exposure (0) or 10, 20, 30 or 60 days after exposure. h: samples exposed to CLas-negative psyllids; Las: samples exposed to CLas-positive psyllids. PC, principal component.
Flush transcriptome is not dramatically altered upon CLas infection in M. paniculata or B. koenigii along 60 days after inoculation
A parallel analysis was performed for the transient host M. paniculata and the fully resistant B. koenigii. In both species, paired-wise comparison again indicated that only a few significant changes were occurring between flushes that were exposed to CLas-negative psyllids and those that were exposed to CLas-positive ones, with less than 100 DEGs per time-point from t10 to t30 (Supplementary Tables 7, S8). In M. paniculata, 17 genes were up-regulated and only 5 down-regulated at t0 after inoculation with CLas-positive psyllids, 4 and 23 genes at t10, 14 and 79 at t20, 24 and 70 at t30 and 41 and 90 at t60 (up- and down-regulated, respectively). In B. koenigii, 28 genes were up-regulated and 85 genes down-regulated after inoculation with CLas-positive psyllids, 19 and 4 genes at t10, 14 and 34 at t20, 16 and 12 at t30, and 560 and 117 at t60 (up- and down-regulated, respectively). PCA plots revealed that samples were clustered according to flush developmental stage and not to the exposure or not to CLas (Figures 4A, B). In B. koenigii, a closer view to the PCA plot revealed that, for CLas-positive challenge, three out of the six replicates in t10 had a transcriptional signature that was closer to samples at t0, whereas in two of them it was closer to t10 samples exposed to CLas-negative psyllids. The remaining replicate displayed a signature unrelated to the rest of the samples and is not plotted (Figure 4B). This observation suggests the existence in this time point of samples of different nature among those exposed to CLas. GSEA analysis identified enriched categories which give insight into the transcriptional alterations taking place in CLas-infected flushes. Among others, it brought our attention the repression of photosynthesis-related processes when CLas was present, observed at t0, t10, t30 and t60 for M. paniculata, and t0 and t10 for B. koenigii, or the downregulation of pathways related to sulfur amino acids in t0 and t10 in B. koenigii (Figures 5, 6). The complete list of categories is shown in Supplementary Tables 9, 10. No categories related to plant immunity or defense responses to bacteria were detected in any time point in any of the two species., In summary, CLas presence on M. paniculata or B. koenigii flushes seems not to cause massive changes in gene expression up to 60 days post-inoculation, with some biochemical reprogramming events taking place.
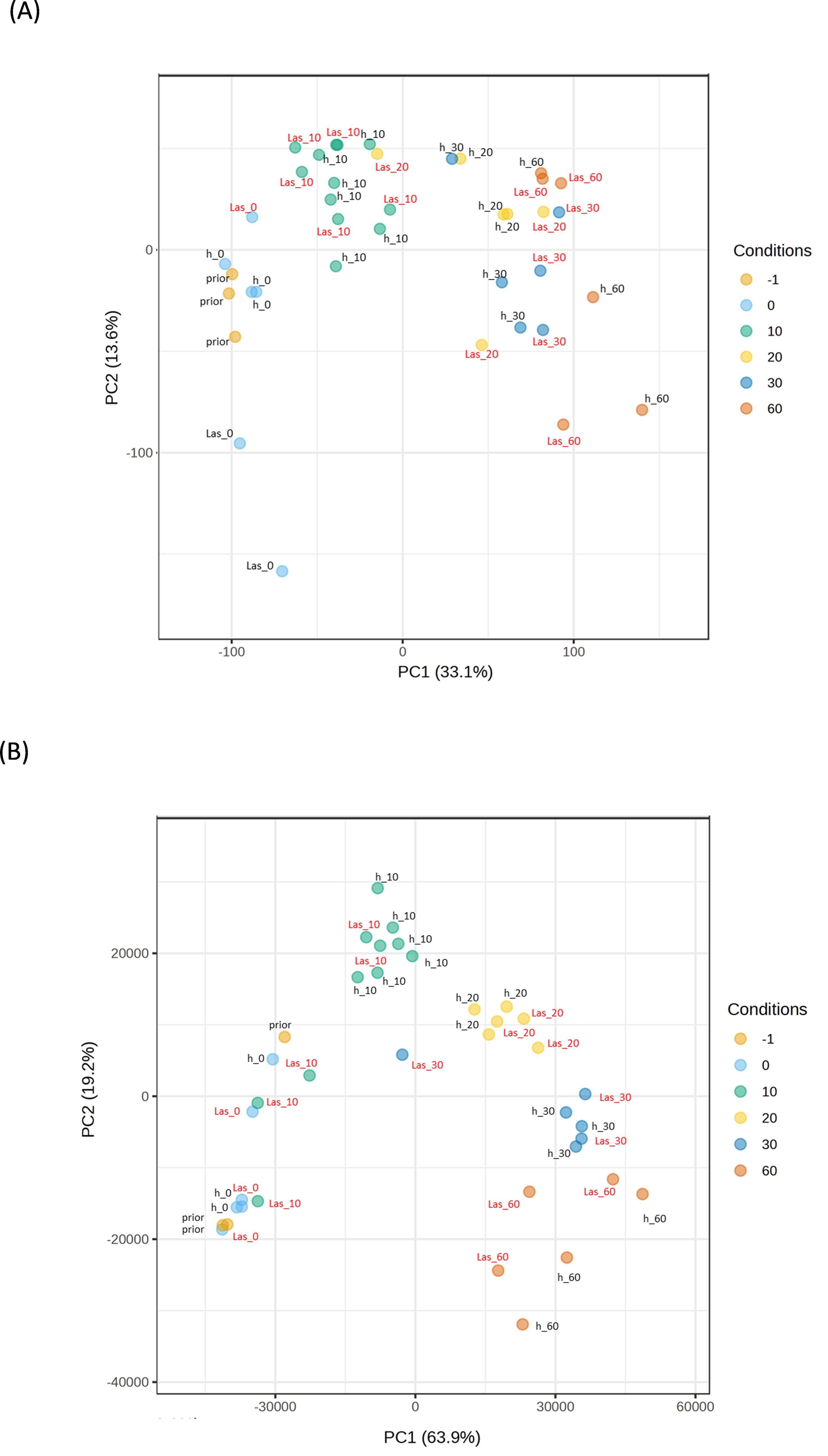
Figure 4. Principal Component Analysis (PCA) plot of Murraya paniculata and Bergera koenigii samples. (A) PCA plot for Murraya paniculata samples. (B) PCA plot for Bergera koenigii samples. Samples were taken before the exposure to psyllids (prior), immediately after exposure (0) or 10, 20, 30 or 60 days after exposure. h: samples exposed to CLas-negative psyllids; Las: samples exposed to CLas-positive psyllids. PC, principal component.
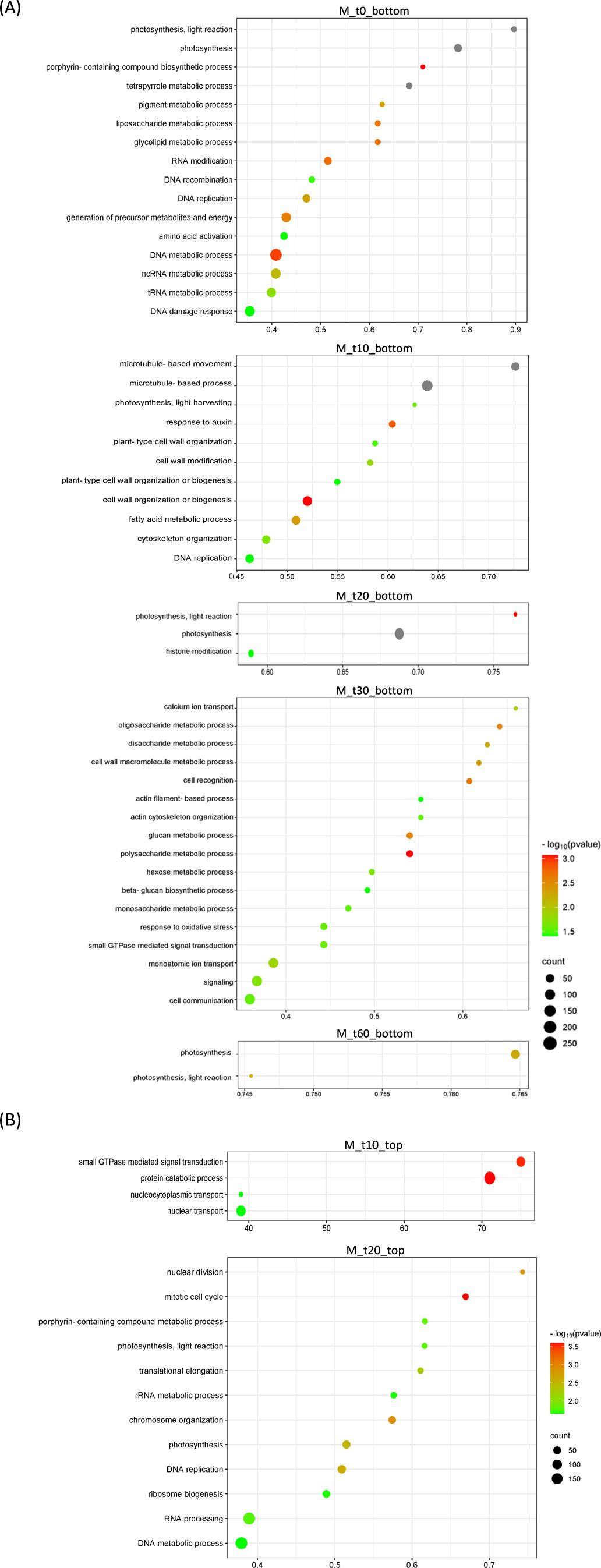
Figure 5. Bubble plots showing Gene Ontology (GO) terms enriched in Murraya paniculata differentially expressed genes. (A) Top-20 nonredundant enriched GO terms after Gene Set Enrichment Analysis in the bottom-ranked genes (bottom) after Deseq2 statistical test. These represent genes less expressed in CLas-infected flushes at different time points after psyllid exposure (M_t0 to M_t60). (B) Top-20 nonredundant enriched GO terms after Gene Set Enrichment Analysis in the top-ranked genes (top) after Deseq2 statistical test. These represent genes more expressed in CLas-infected flushes at different time points after psyllid exposure (M_t10 and M_t20). GO terms for the remaining time points are detailed in Supplementary Table 7. GO terms were obtained using Panther and filtered using ReviGO to remove semantically redundant terms. GO terms with frequency<10% were selected and ranked by dispensability. Gene ratio: Ratio of upregulated genes in a given category divided by total number of genes in this category. Counts, number of upregulated genes in a given category. −log P (P-value in log scale after false discovery rate correction).
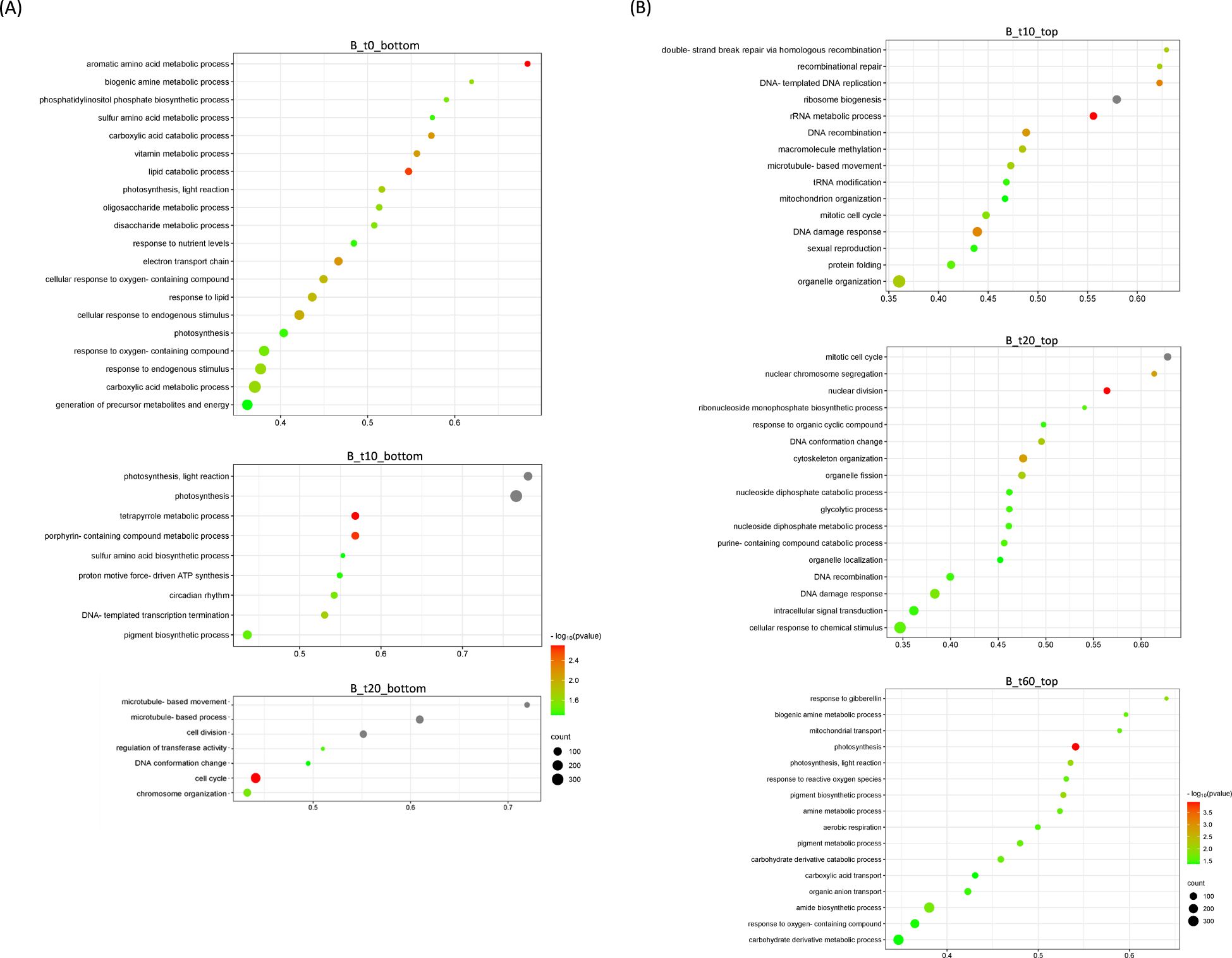
Figure 6. Bubble plots showing Gene Ontology (GO) terms enriched in Bergera koenigii differentially expressed genes. (A) Top-20 nonredundant enriched GO terms after Gene Set Enrichment Analysis in the bottom-ranked genes (bottom) after Deseq2 statistical test. These represent genes less expressed in CLas-infected flushes at different time points after psyllid exposure (B_t0, B_t10; B_t20). (B) Top-20 nonredundant enriched GO terms after Gene Set Enrichment Analysis in the top-ranked genes (top) after Deseq2 statistical test. These represent genes more expressed in CLas-infected flushes at different time points after psyllid exposure (B_t10, B_t20; B_t60). Number of GO terms for the remaining time points are detailed in Supplementary Table 8. GO terms were obtained using Panther and filtered using ReviGO to remove semantically redundant terms. GO terms with frequency<10% were selected and ranked by dispensability. Gene ratio: Ratio of genes in a given category divided by total number of genes in this category. Counts, number of genes in a given category. −log P (P-value in log scale after false discovery rate correction).
Transcriptomes of young flushes reveal distinct biochemical potentials for the three species that could explain differences in their response to CLas
The previous results suggest that M. paniculata and B. koenigii are not deploying a canonical immune response after CLas inoculation. It also suggests that gene expression changes upon CLas detection in flushes in both species are associated to biochemical rearrangements. To assess whether survival and multiplication of CLas in the different hosts is also influenced by the biochemical potential of the tissues in which CLas was initially inoculated by ACP, flush transcriptomes before bacterial inoculation (prior) were compared.
Gene expression was analyzed for groups of orthologous genes. Nearly 14000 and 9000 orthologous groups were identified between C. × sinensis, M. paniculata and B. koenigii, respectively. Many of those genes were found differentially expressed in non-inoculated flushes of the three species. More than 2500 genes were found more expressed in B. koenigii than in C. × sinensis flushes, and more than 2000 were less expressed. Similarly, more than 2600 genes were found more expressed in M. paniculata than in C. × sinensis, whereas more than 2900 genes were less expressed (Supplementary Table 11). Functional analysis upon differentially-expressed genes suggest the existence of many biochemical pathways that could be activated at different levels in the different species (Figure 7). For example, phosphate transport is more active in C. × sinensis than in M. paniculata, and genes related to cellular response to phosphate starvation are more expressed in B. koenigii than in C. × sinensis, whereas genes related to the serine metabolic pathway or the ‘tricarboxylic acid cycle’ are more expressed in C. × sinensis than in B. koenigii. The complete list of categories is shown in Supplementary Table 12. These results suggest that the biological composition of the phloem sap from the flushes of the three species is very different and could also be contributing to their distinct response to CLas inoculation.
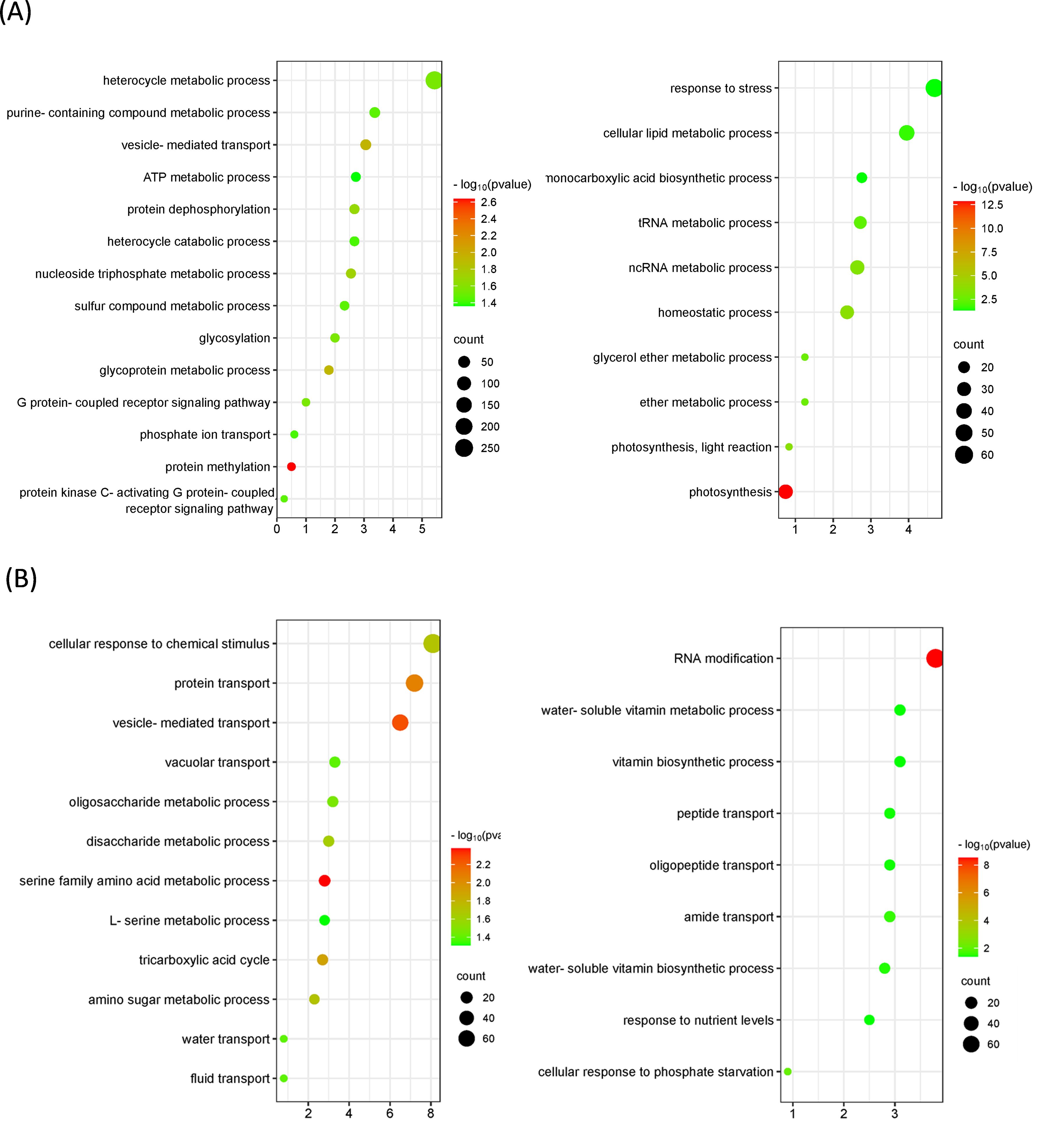
Figure 7. Bubble plots showing Gene Ontology (GO) terms enriched in orthologous groups between Citrus × sinensis and Murraya paniculata or Bergera koenigii flushes before psyllid exposure. (A) Top-20 nonredundant enriched GO terms among orthologous genes more (left) or less (right) expressed in Citrus × sinensis as compared to Murraya paniculata. (B) Top-20 nonredundant enriched GO terms among orthologous genes more (left) or less (right) expressed in Citrus × sinensis as compared to Bergera koenigii. GO terms were obtained using Panther and filtered using ReviGO to remove semantically redundant terms. GO terms with frequency<5% were selected and ranked by dispensability. Gene ratio, Ratio of genes in a given category divided by total number of genes in this category. Counts, number of genes in a given category. −log P (P-value in log scale after false discovery rate correction).
Discussion
The current model of plant resistance settles mostly on studies of mesophyll-infecting pathogens, and perhaps its most relevant feature is the activation of inducible responses by the plant upon pathogen recognition (Zhou and Zhang, 2020). Therefore, it is not surprising that many transcriptomic studies designed to evaluate the responses to CLas in susceptible or tolerant Citrus species centered the discussions on the deployment or not of complete defensive responses (Arce-Leal et al., 2020; Weber et al., 2022; Li et al., 2024). To our understanding, two essential aspects have been overlooked in many of these studies. On one hand, if CLas recognition by tolerant plants leads to PTI or ETI, a transcriptomic response should be activated very early after bacteria recognition. The foundation of the deployment of a robust PTI lies in the activation of stress-responsive transcription factors and the gene networks under their control, which begins soon after pathogen recognition (Li et al., 2020a). PTI is rapidly evaded by successful pathogens through the action of secreted effectors acting into the plant cells. When these effectors are recognized by resistant plans, ETI is activated, characterized by a marked transcriptional response within hours after pathogen recognition (Mine et al., 2018). Omics studies on HLB symptomatic samples, in which CLas infection took place months earlier, are not targeting the first molecular events after CLas recognition. In addition, CLas-infected material inoculated by grafting requires the earliest time point to be sampled weeks after inoculation to allow vascular recovery and CLas translocation to become detectable by qPCR (Raiol-Junior et al., 2021). Consequently, it is also an inadequate experimental system to evaluate early events of CLas infection. Simulation of natural entry by ACP-mediated inoculation using young shoots (Alves et al., 2021a; Lopes and Cifuentes-Arenas, 2021), as the one used by Wei et al. (2021) or in this work, seems to be more realistic to elucidate the molecular events taking place in this plant-pathogen interaction.
On the other hand, a complete immune response involves a massive transcriptional reprogramming of the cell, in the range of thousands of differentially-expressed genes (Lewis et al., 2015; Li et al., 2016; Mine et al., 2018). For example, in Arabidopsis, up to 25% of the genes presents altered gene expression after inoculation with the virulent Pseudomonas syringae (Tao et al, 2003), and in Citrus, more than 6000 genes were differentially expressed in the citrus canker resistant Kumquat 72 hours after recognition of Xanthomonas citri subs. citri, its causal agent (Ferrara et al., 2020). Therefore, up-regulation of some defense-related genes does not necessarily mean that a complete defense response has been deployed.
The relatively low number of DEGs found in the early stages after pathogen inoculation into the susceptible C. × sinensis observed in this study aligns with other reports analyzing the response to CLas in asymptomatic plants. For example, Fan et al. (2012) reported no significant changes in gene expression at 5 weeks after grafting inoculation to either C. × limon or C. × sinensis. Similarly, Ramsey et al. (2020) identified only 313 DEGs at 2 weeks after grafting inoculation to C. × limon, and Lombardi et al. (2024) found only 223 DEGs 4 weeks after inoculation to C. × sinensis, this last report using also ACP-vectored inoculation. Interestingly, a limited transcriptional reprogramming is also observed in response to other phloem-infecting pathogens of citrus, such as Spiroplasma citri (McNeil et al., 2023), suggesting that this might be a common feature of vascular pathogens that share similar infection pathways. By contrast, Wei et al. (2021) detected a high number of DEGs as early as 1 day after CLas inoculation in ‘Valencia’ sweet orange.
Our results suggest the idea that exposure of susceptible plants to CLas is not dramatically altering gene expression and indicate that a canonical and complete transcriptional immune response is not being activated in this susceptible host, although the possibility of waves of transcriptional changes occurring between the time intervals used in this experimental design cannot be discarded. Two non-exclusive hypotheses could explain these observations: a) the secretion of bacterial effectors suppress PTI, as it is proposed for several proteins putatively secreted by CLas to suppress PTI (Clark et al., 2018; Shi et al., 2019; Clark et al., 2020; Basu et al., 2022; Shi et al., 2023a, Shi et al., 2023b). So far, the mechanism for secretion is unknown, as CLas does not have the main machinery for effector secretion present in most plant pathogens (Cui et al., 2015; Jain et al., 2015). These effectors could be part of an orchestrated strategy in which CLas could also secrete other proteins that may interfere with specific plant metabolites produced by nucleated cells, altering plant cell homeostasis for its own benefit, and finally inducing symptoms, as observed for other phloem-limited pathogens (MacLean et al., 2011). b) CLas remains unnoticed for mesophyll cells, due to the apoplastic entry and exclusive vascular localization mediated by ACP, and the nuclei-free sieve element are unable to mount an effective immune response (Jiang et al., 2019; Huang et al., 2020; Lewis et al., 2022).
The use of resistant species such as M. paniculata and B. koenigii, in which CLas establishment is clearly limited (Alves et al., 2021a) opens the possibility to explore the molecular basis for resistance. Monoembryonic plants may exhibit genetic differences attributable to seed propagation, which might influence gene expression profiles. However, both Aurantioideae species were introduced into Brazil many decades ago, are seed-propagated and widely used as ornamentals, being highly homozygous (Nguyen et al., 2019; our unpublished results). In any case, as observed in the PCA, samples for each time point were closely grouped, indicating consistency among samples. Surprisingly, the limited number of DEGs observed in these two interactions in the first weeks after ACP-mediated bacterial inoculation, as well as the lack of immune defense categories enriched after functional analysis, suggest that canonical PTI or ETI responses are not the main mechanisms responsible for resistance in these two species. However, functional analysis revealed that the bacterium was not completely evading transcriptional responses in the plants. Both M. paniculata and B. koenigii tissues react to the presence of the bacterium by reprogramming the transcriptome of basic cellular functions. In both species, CLas presence provoked a subtle but consistent decrease in the expression of photosynthesis-related genes, photosynthesis being an enriched category upon less-expressed genes as soon as 48 hours after exposure to the bacterium. Enrichment of photosynthesis-related categories after transcriptomic analysis has been reported upon CLas infection in susceptible plants (Albrecht and Bowman, 2008; Zhong et al., 2015; Hu et al., 2017; Liu et al., 2019), but these experiments were performed in symptomatic plants, in which starch accumulation had already impaired photosynthetic processes and induced chloroplast degradation (Etxeberria et al., 2009). As mentioned before, this transcriptomic signature can be considered an effect of visual symptoms and therefore unrelated to the events investigated here. A decrease in photosynthesis in M. paniculata and B. koenigii at these very early stages after CLas inoculation, however, could be perceived as a mechanism triggered by the plants to hamper pathogen colonization. Curiously, this was also observed by Wei et al. (2021) in sweet orange at 5 days post inoculation with Clas-infected psyllids. Photosynthesis reduction has been reported in the incompatible interaction between tobacco and Phytophthora nicotianae (Scharte et al., 2005), as well as in resistant barley plants after inoculation with powdery mildew (Swarbrick et al., 2006). In M. paniculata or B. koenigii, a decrease in photosynthesis in the early stages of CLas-infection could be a mechanism to regulate source/sink relationships to limit pathogen spread (McIntyre et al., 2021). CLas moves from source to sink tissues through the phloem vessels. After inoculation, CLas remains in the young flushes until they are mature enough to become source tissues (Raiol-Junior et al., 2017; Alves et al., 2021a). In citrus, young flushes are nutritionally richer than mature flushes, with higher concentrations of total amino acids in the phloem sap, positively associated with CLas and ACP fitness (Setamou et al., 2016, Setamou et al., 2017). In this study, C. × sinensis flush maturation took longer than in M. paniculata and B. koenigii, and C. × sinensis flushes exposed to CLas-positive psyllids reached maturity slower than those exposed to CLas-negative psyllids. The combination of these two factors would increase the time window in which CLas have the available nutrients crucial for its establishment. However, in young, infected flushes of M. paniculata or B. koenigii, photosynthesis decay could translate into a retardation in the sink to source transition, compromising the establishment of CLas in hosts presenting nutrient inadequacy (Killiny, 2016). In this regard, the observation at t10 that two out of five B. koenigii samples exposed to CLas-positive psyllids cluster to samples exposed to t10 CLas-negative psyllids might be events in which the bacterium had failed to multiply in these plants.
Other biochemical processes altered after CLas exposure in B. koenigii could hamper bacterial survival and multiplication, such as the decrease in the biosynthesis of sulfur-containing amino acids. CLas is not able to synthesize cysteine de novo and must overcome its lack by acquiring it from the surrounding environment (Lai et al., 2016; Merfa et al., 2019). Interestingly, the expression of cysteine biosynthesis genes was also higher in non-inoculated C. × sinensis young flushes than in Bergera’s ones, suggesting that this metabolic constraint could be determinant for CLas multiplication in the resistant species. Although these cellular rearrangements are taking place upon CLas perception, the lack of nuclei in sieve elements suggests that these responses should be activated in adjacent nucleated cells. Similarly, in the case of C. × sinensis and other susceptible species, effector proteins, trafficking through the plasmodesmata (Cheval and Faulkner, 2018), could suppress immune signaling in nucleated cells and impact the production of systemic signals (Huang et al., 2024).
The lack of a bona fide defense response in M. paniculata and B. koenigii prompted us to investigate whether these species could also have physiological or metabolic constraints that could limit CLas establishment. Comparison of expression of orthologous genes provides an estimation of biological processes that are active in young flushes of any of the three species. Our analysis revealed that the three transcriptomes are very different, and consequently, their biochemical composition should also differ, as reported by Killiny (2016). The small genome content of CLas and the lack of core metabolic pathways indicate that this bacterium heavily depends on the host nutrition (Wang and Trivedi, 2013; Lai et al., 2016), which could impact on the suitability of different species as host. Our functional analysis suggest that the higher expression of genes related to phosphate transport in C. × sinensis or related to the response to phosphate starvation in B. koenigii, which is essential for Liberibacter optimal growth (Cruz-Munoz et al., 2019), differ between the three species. Moreover, C. × sinensis young flushes have a higher expression of genes of the tricarboxylic acid cycle (TCA) than B. koenigii. The ATP metabolism was also enriched upon genes more expressed in C. × sinensis than in M. paniculata. These findings likely reflect a higher activity of this metabolic pathway in the susceptible C. × sinensis. Citrate is the preferred carbon source for Liberibacter (Cruz-Munoz et al., 2018) and evidence suggests that CLas obtains energy by exploiting TCA cycle intermediates from the plant (Wang and Trivedi, 2013; Jain et al., 2017) and psyllid hosts (Killiny and Nehela, 2017). An ATP translocase has been identified in CLas, suggesting the potential to import ATP directly from their environment (Vahling et al., 2010). It could be hypothesized that a lower activity of the TCA cycle in B. koenigii and M. paniculata flushes could be determining their less suitability for CLas establishment. In the most extreme case of unsuitability for CLas survival, B. koenigii may be considered as a non-host of the bacterium.
Overall, our data support the hypothesis that most C. × sinensis cells do not recognize the presence of CLas after inoculation, likely due to the ACP strategy to introduce the bacterium directly into the phloem in less than two hours (Wu et al., 2016), which results in a very limited - transcriptional response. The fortuitous protection obtained by CLas in the vascular tissue would allow it to remain unnoticed for mesophyll cells, thus limiting the possibility to orchestrate a robust and effective defense response. This, together with the putative effector-mediated suppression of PTI would explain the long incubation period and the difficulty in managing this devastating disease. This model could also explain why foliar spray of inducers of pathogen resistance, such as salicylic acid, have proven to be ineffective in reducing bacterial titer in HLB-infected plants (Atwood and Brlansky, 2009; Li et al., 2021) as well as in other phloem-limited pathogens (Rodriguez-Saona et al., 2021). The same scheme would apply for B. koenigii and M. paniculata, although in these cases the presence of the bacterium in the phloem would induce biochemical rearrangements in the surrounding cells, suggesting the existence of signaling events initiated in the sieve elements upon CLas recognition.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material. The datasets generated for this study can be found in the GEO database under the accession number GSE277583.
Author contributions
MA: Conceptualization, Formal analysis, Validation, Investigation, Writing – original draft, Writing – review & editing. JC-A: Investigation, Writing – review & editing. RN: Investigation, Writing – review & editing. LR-J: Investigation, Writing – review & editing. EC: Investigation, Writing – review & editing. IQ-R: Investigation, Writing – review & editing. JAF: Conceptualization, Writing – review & editing. CL: Formal analysis, Resources, Writing – review & editing. BA: Formal analysis, Investigation, Validation, Writing – review & editing. LC: Formal analysis, Investigation, Writing – review & editing. JF: Formal analysis, Investigation, Writing – review & editing. AB: Formal analysis, Investigation, Writing – review & editing. NW: Formal analysis, Writing – review & editing, Funding acquisition. LP: Conceptualization, Formal analysis, Validation, Writing – original draft, Writing – review & editing, Funding acquisition. JG: Conceptualization, Formal analysis, Validation, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This study was partly funded by the Horizon 2020 of the European Union (H2020-SFS-2018-2020), under the project PRE-HLB: Preventing HLB epidemics for ensuring citrus survival in Europe and by Fundecitrus (04 3108). MA was recipient of a Ph.D. fellowship from CAPES.
Acknowledgments
We are grateful to André Luís Sanches team for their excellent assistance with grafting for propagation and plant maintenance, and to Fernanda Benedito for her support with DNA extraction and qPCR analysis.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2025.1502953/full#supplementary-material
Supplementary Figure 1 | Transcriptome changes in Citrus × sinensis, Murraya paniculata and Bergera koenigii flushes along the first 60 days after exposure to psyllids. (A) Number of down-regulated genes in Citrus × sinensis samples taken immediately (CLas_0, Ch_0), and after 10 (CLas_10, Ch_10), 20 (CLas_20, Ch_20), 30 (CLas_30, Ch_30) or 60 (CLas_60, Ch_60) days after exposure to CLas-negative (Ch, top) or CLas-positive (CLas, bottom) psyllids when compared against flushes before the exposure to psyllids (prior). (B) Number of down-regulated genes in Murraya paniculata samples taken immediately (MLas_0, Mh_0), and after 10 (MLas_10, Mh_10), 20 (MLas_20, Mh_20), 30 (MLas_30, Mh_30) or 60 (MLas_60, Mh_60) days after exposure to CLas-negative (Mh, top) or CLas-positive (MLas, bottom) psyllids when compared against flushes before the exposure to psyllids (prior). (C) Number of down-regulated genes in Bergera koenigii samples taken immediately (BLas_0, Bh_0), and after 10 (BLas_10, Bh_10), 20 (BLas_20, Bh_20), 30 (BLas_30, Bh_30) or 60 (BLas_60, Bh_60) days after exposure to CLas-negative (Bh, top) or CLas-positive (BLas, bottom) psyllids when compared against flushes before the exposure to psyllids (prior). CLas: Candidatus Liberibacter asiaticus. DEGs: Differentially expressed genes. In each graph, total number of DEGs (X axis) at each time point (Y axis) is shown in bottom-left panels. Intersection of sets of genes at multiple time points is shown in top right panels. Each column corresponds to a time point or set of time points (dots connected by lines below the X axis) containing the same DEGs. The time points shared are indicated in the graphic below the column, with the time points on the left.
References
Albrecht, U., Bowman, K. D. (2008). Gene expression in Citrus sinensis (L.) Osbeck following infection with the bacterial pathogen Candidatus Liberibacter asiaticus causing Huanglongbing in Florida. Plant Science. 175, 291–306. doi: 10.1016/j.plantsci.2008.05.001
Alves, M. N., Cifuentes-Arenas, J. C., Raiol-Junior, L. L., Ferro, J. A., Peña, L. (2021a). Early population dynamics of “Candidatus Liberibacter asiaticus” in susceptible and resistant genotypes after inoculation with infected Diaphorina citri feeding on young shoots. Front. Microbiol. 12. doi: 10.3389/fmicb.2021.683923
Alves, M. N., Lopes, S. A., Raiol-Junior, L. L., Wulff, N. A., Girardi, E. A., Ollitrault, P., et al. (2021b). Resistance to ‘Candidatus Liberibacter asiaticus’, the Huanglongbing associated bacterium, in sexually and/or graft-compatible Citrus relatives. Front. Plant Sci. 11. doi: 10.3389/fpls.2020.617664
Alves, M. N., Raiol-Junior, L. L., Girardi, E. A., Miranda, M., Wulff, N. A., Carvalho, E. V., et al. (2022). Insight into resistance to ‘Candidatus Liberibacter asiaticus,’ associated with Huanglongbing, in Oceanian citrus genotypes. Front. Plant Sci. 13. doi: 10.3389/fpls.2022.1009350
Andrade, M. O., Pang, Z., Achor, D. S., Wang, H., Yao, T., Singer, B. H., et al. (2019). The flagella of ‘Candidatus Liberibacter asiaticus’ and its movement in planta. Mol. Plant Pathol. 21, 109–123. doi: 10.1111/mpp.12884
Andrews, S. (2010). FastQC: a quality control tool for high throughput sequence data. Available online at: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (Accessed November 01, 2023]).
Arce-Leal, Á.P., Bautista, R., Rodríguez-Negrete, E. A., Manzanilla-Ramírez, M.Á., Velázquez-Monreal, J. J., Santos Cervantes, M. E., et al. (2020). Gene Expression Profile of Mexican Lime (Citrus aurantifolia) Trees in Response to Huanglongbing Disease caused by Candidatus Liberibacter asiaticus. Microorganisms 8:4, 528. doi: 10.3390/microorganisms8040528
Asai, S., Shirasu, K. (2015). Plant Cells under siege: Plant Immune system versus pathogen effectors. Curr. Opin. Plant Biol. 28, 1–8. doi: 10.1016/j.pbi.2015.08.008
Atwood, R., Brlansky, R. H. (2009). A trial to determine if salicylic acid and7or K-phite on young citrus trees grafter with HLB-infected budwood could prevent transmission of the bacteria. Proc. Florida State Hortic. Society. 122, 39. doi: 10.5555/20183248055
Basu, S., Huynh, L., Zhang, S., Rabara, R., Nguyen, H., Velásquez Guzmán, J., et al. (2022). Two liberibacter proteins combine to suppress critical innate immune defenses in citrus. Front. Plant Sci. 2:13. doi: 10.3389/fpls.2022.869178
Beloti, V. H., Alves, G. R., Coletta-Filho, H. D., Yamamoto, P. T. (2018). The Asian Citrus psyllid host Murraya koenigii is immune to Citrus huanglongbing pathogen ‘Candidatus Liberibacter asiaticus’. Phytopathology 108, 1089–1094. doi: 10.1094/PHYTO-01-18-0012-R
Beloti, V. H., Santo, F., Alves, G. R., Bento, J. M. S., Yamamoto, P. T. (2017). Curry leaf smells better than citrus to females of Diaphorina citri (Hemiptera: Liviidae). Arthropod-Plant Interact. 11, 709–716. doi: 10.1007/s11829-017-9524-6
Bigeard, J., Colcombet, J., Hirt, H. (2015). Signaling mechanisms in pattern-triggered immunity (PTI). Mol. Plant 8, 521–539. doi: 10.1016/j.molp.2014.12.022
Bové, J. M. (2006). Huanglongbing: a destructive, newly-emerging, century-old disease of citrus. J. Plant Pathol. 88, 7–37. doi: 10.4454/jpp.v88i1.828
Boyle, E. I., Weng, S., Gollub, S., Jin, H., Botstein, D., Cherry, J. M., et al. (2004). GO::TermFinder–open source software for accessing gene ontology information and finding significantly enriched gene ontology terms associated with a list of genes. Bioinf. (Oxford England) 208, 3710–3715. doi: 10.1093/bioinformatics/bth456
Buchfink, B., Xie, C., Huson, D. H. (2015). Fast and sensitive protein alignment using DIAMOND. Nat. Methods 12:1, 59–60. doi: 10.1038/nmeth.3176
Burckardt, D., Ouvrard, D., Percy, D. M. (2021). An updated classification of the jumping plant-lice (Hemiptera: Psylloidea) integrating molecular and morphological evidence. Eur. J. Taxonomy 736, 137–182. doi: 10.5852/ejt.2021.736.1257
Ceulemans, E., Ibrahim, H. M. M., De Coninck, B., Goossens, A. (2021). Pathogen effectors: exploiting the promiscuity of plant signaling hubs. Trends Plant Sci. 26, 780–795. doi: 10.1016/j.tplants.2021.01.005
Challis, R., Richards, E., Rajan, J., Cochrane, G., Blaxter, M. (2020). BlobToolKit–interactive quality assessment of genome assemblies. G3: Genes Genomes Genet. 10:4, 1361–1374. doi: 10.1534/g3.119.400908
Cheval, C., Faulkner, C. (2018). Plasmodesmal regulation during plant-pathogen interactions. New Phytol. 217, 62–67. doi: 10.1111/nph.14857
Cifuentes-Arenas, J. C., Beattie, G. A. C., Peña, L., Lopes, S. A. (2019). Murraya paniculata and Swinglea glutinosa as short-term transient hosts of ‘Candidatus Liberibacter asiaticus’ and implications for the spread of huanglongbing. Phytopatology 109, 2064–2073. doi: 10.1094/PHYTO-06-19-0216-R
Cifuentes-Arenas, J. C., de Goes, A., de Miranda, M. P., Beattie, G. A. C., Lopes, S. A. (2018). Citrus flush shoot ontogeny modulates biotic potential of Diaphorina citri. PloS One 13, e0190563. doi: 10.1371/journal.pone.0190563
Clark, K., Franco, J. Y., Schwizer, S., Pang, Z., Hawara, E., Liebrand, T. W. H., et al. (2018). An effector from the Huanglongbing-associated pathogen targets citrus proteases. Nat. Commun. 9, 1718. doi: 10.1038/s41467-018-04140-9
Clark, K. J., Pang, Z., Trinh, J., Wang, N., Ma, W. (2020). Sec-delivered effector 1 (SDE1) of ‘Candidatus liberibacter asiaticus’ Promotes citrus huanglongbing. Mol. Plant Microbe Interact. 33, 1394–1404. doi: 10.1094/MPMI-05-20-0123-R
Cruz-Munoz, M., Munoz-Beristain, A., Petrone, J. R., Robinson, M. A., Triplett, E. W. (2019). Growth parameters of Liberibacter crescens suggest ammonium and phosphate as essential molecules in the Liberibacter-plant host interface. BMC Microbiol. 2, 222. doi: 10.1186/s12866-019-1599-z
Cruz-Munoz, M., Petrone, J. R., Cohn, A. R., Munoz-Beristain, A., Killiny, N., Drew, J. C., et al. (2018). Development of chemically defined media reveals citrate as preferred carbon source for liberibacter growth. Front. Microbiol. 5 9. doi: 10.3389/fmicb.2018.00668
Cui, H., Tsuda, K., Parker, J. E. (2015). Effector-triggered immunity: from pathogen perception to robust defense. Annu. Rev. Plant Biol. 66, 487–451. doi: 10.1146/annurev-arplant-050213-040012
Damsteegt, V. D., Postnikova, E. N., Stone, A. L. (2010). Murraya paniculata and related species as potential hosts and inoculum reservoirs of ‘Candidatus Liberibacter asiaticus’, causal agent of Huanglongbing. Plant Dis. 94, 528–533. doi: 10.1094/PDIS-94-5-0528
Dobin, A., Davis, C. A., Schlesinger, F., Drenkow, J., Zaleski, C., Jha, S., et al. (2013). STAR: ultrafast universal RNA-seq aligner. Bioinformatics. 29, 15–21. doi: 10.1093/bioinformatics/bts635
Dodds, P. N., Rathjen, J. P. (2010). Plant immunity: towards an integrated view of plant-pathogen interactions. Nat. Rev. Genet. 11, 539–548. doi: 10.1038/nrg2812
Duan, Y., Zhou, L., Hall, D. G., Li, W., Doddapaneni, H., Lin, H., et al. (2009). Complete genome sequence of Citrus Huanglongbing bacterium, ‘Candidatus Liberibacter asiaticus’ obtained through metagenomics. MPMI 22, 1011–1020. doi: 10.1094/MPMI-22-8-1011
Eduardo, W. I., Miranda, M. P., Volpe, H. X. L., Garcia, R. B., Girardi, E. A., Alquezar, B., et al. (2022). Resistance of True Citrus species to Diaphorina citri. Pest Manag. Sci. 78, 4783–4792. doi: 10.1002/ps.7098
Etxeberria, E., Gonzalez, P., Achor, D., Albrigo, G. (2009). Anatomical distribution of abnormally high levels of starch in HLB-affected Valencia orange trees. Physiol. Mol. Plant Pathol. 74, 76–83. doi: 10.1016/j.pmpp.2009.09.004
Fan, J., Chen, C., Yu, Q., Khalaf, A., Achor, D. S., Brlansky, R. H., et al. (2012). Comparative transcriptional and anatomical analyses of tolerant rough lemon and susceptible sweet orange in response to ‘Candidatus Liberibacter asiaticus’ infection. Mol. Plant Microbe Interact. 25:11, 1396–1407. doi: 10.1094/MPMI-06-12-0150-R
Ferrara, A., Murata, M. M., Cofre, T. D. C. G., Cavallini, J. S., Peron, G., Julião, M. H. M., et al. (2020). CitrusKB: a comprehensive knowledge base for transcriptome and interactome of Citrus spp. infected by Xanthomonas citri subsp. citri at different infection stages. Database (Oxford). 1, baaa08. doi: 10.1093/database/baaa081
Folimonova, S. Y., Robertson, C. J., Gamsey, S. M., Dawson, W. O. (2009). Examination of the responses of different genotypes of citrus to huanglongbing (citrus greening) under different conditions. Phytopathology 23, 1346–1354. doi: 10.1094/PHYTO-99-12-1346
Fundecitrus (2024). Doenças: greening/HLB. Available online at: https://www.fundecitrus.com.br/levantamentos/greening (Accessed December 3, 2024).
Gottwald, T. R. (2010). Current epidemiological understanding of citrus Huanglongbing. Annu. Rev. Phytopathol. 48, 119–139. doi: 10.1146/annurevphyto-073009-114418
Grabherr, M. G., Haas, B. J., Yassour, M., Levin, J. Z., Thompson, D. A., Amit, I., et al. (2011). Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 15:29, 644–652. doi: 10.1038/nbt.1883
Hu, Y., Zhong, X., Liu, X., Lou, B., Zhou, C., Wang, X. (2017). Comparative transcriptome analysis unveils the tolerance mechanisms of Citrus hystrix in response to ‘Candidatus Liberibacter asiaticus’ infection. PloS One 12:12, e0189229. doi: 10.1371/journal.pone.0189229
Huang, G., Chang, X., Hu, Y., Li, F., Wang, N., Li, R. (2024). SDE19, a SEC-dependent effector from ‘Candidatus Liberibacter asiaticus’ suppresses plant immunity and targets Citrus sinensis Sec12 to interfere with vesicle trafficking. PloS Pathog. 10:20, e1012542. doi: 10.1371/journal.ppat.1012542
Huang, W., Reyes-Caldas, P., Mann, M., Seifbarghi, S., Kahn, A., Almeida, R. P. P., et al. (2020). Bacterial vector-borne plant diseases: unanswered questions and future directions. Mol. Plant 13, 1379–1393. doi: 10.1016/j.molp.2020.08.010
Jain, M., Fleites, L., Gabriel, D. (2015). Prophage-encoded peroxidade in ‘Candidatus liberibacter asiaticus’ Is a screted effector that suppresses plant defenses. MPMI 28, 1330–1337. doi: 10.1094/MPMI-07-05-0145-R
Jain, M., Munoz-Bodnar, A., Gabriel, D. W. (2017). Concomitant loss of the glyoxalase system and glycolysis makes the uncultured pathogen “Candidatus liberibacter asiaticus” an energy scavenger. Appl. Environ. Microbiol. 16:83, e01670–e01617. doi: 10.1128/AEM.01670-17
Jiang, Y., Zhang, C.-X., Chen, R., He, S. Y. (2019). Challenging battles of plants with phloem-feeding insects and prokaryotic pathogens. PNAS. 116, 23390–23397. doi: 10.1073/pnas.1915396116
Killiny, N. (2016). Metabolomic comparative analysis of the phloem sap of curry leaf tree (Bergera koenegii), orange jasmine (Murraya paniculata), and Valencia sweet orange (Citrus sinensis) supports their differential responses to Huanglongbing. Plant Signal. Behav. 11, e1249080. doi: 10.1080/15592324.2016.1249080
Killiny, N., Nehela, Y. (2017). Metabolomic Response to Huanglongbinh: Role of Carboxylic Compound in Citrus sinensis Response to ‘Candidatus Liberibacter asiaticus’ and its vector. Diaphorina citri. 30:8, 666–678. doi: 10.1094/MPMI-05-17-0106-R
Lai, K. K., Davis-Richardson, A. G., Dias, R., Triplett, E. W. (2016). Identification of the genes required for the culture of liberibacter crescens, the closest cultured relative of the liberibacter plant pathogens. Front. Microbiol. 207. doi: 10.3389/fmicb.2016.00547
Lee, J. A., Halbert, S. E., Dawson, W. O., Robertson, C. J., Keesling, J. E., Singer, B. H. (2015). Asymptomatic spread of huanglongbing and implications for disease control. Proc. Natl. Acad. Sci. 12:24, 7605–7610. doi: 10.1073/pnas.1508253112
Lewis, J. D., Knoblauch, M., Turgeon, R. (2022). The phloem as an arena for plant pathogens. Annu. Rev. Phytopathol. 60, 77–96. doi: 10.1146/annurev-phyto-020620-100946
Lewis, L. A., Polanski, K., de-Torres-Zabala, M., Jayaraman, S., Bowden, L., Moore, J., et al. (2015). Transcriptional dynamics driving MAMP-triggered immunity and pathogen effector-mediated immunosuppression in Arabidopsis leaves following infection with Pseudomonas syringae pv tomato DC3000. Plant Cell 27, 3038–3064. doi: 10.1105/tpc.15.00471
Li, H., Durbin, R. (2009). Fast and accurate short read alignment with Burrows–Wheeler transform. Bioinformatics 25:14, 1754–1760. doi: 10.1093/bioinformatics/btp324
Li, H., Handsaker, B., Wysoker, A., Fennell, T., Ruan, J., Homer, N., et al. (2009). 1000 Genome Project Data Processing Subgroup. The sequence alignment/map format and SAMtools. Bioinformatics 25:16, 2078–2079. doi: 10.1093/bioinformatics/btp352
Li, W., Hartung, J. S., Levy, L. (2006). Quantitative real-time PCR for detection and identification of Candidatus Liberibacter species associated with citrus Huanglongbing. J. Microbiol. Methods 66, 104–115. doi: 10.1016/j.mimet.2005.10.018
Li, J., Kolbasov, V. G., Pang, Z., Duan, S., Lee, D., Huang, Y., et al. (2021). Evaluation of the control effect of SAR inducers against citrus Huanglongbing applied by foliar spray, soil drench or trunk injection. Phytopathol. Res. 3, 2. doi: 10.1186/s42483-020-00079-2
Li, P., Lu, Y. J., Chen, H., Day, B. (2020b). The lifecycle of the plant immune system. CRC Crit. Rev. Plant Sci. 39:1, 72–100. doi: 10.1080/07352689.2020.1757829
Li, B., Meng, X., Shan, L., He, P. (2016). Transcriptional regulation of pattern-triggered immunity in plants. Cell Host Microbe 19, 641–650. doi: 10.1016/j.chom.2016.04.011
Li, R., Wang, X., Hu, Y., Huang, G. (2024). Analysis of huanglongbing-associated RNA-seq data reveals disturbances in biological processes within Citrus spp. triggered by Candidatus Liberibacter asiaticus infection. Front. Plant Sci. 10, 15. doi: 10.3389/fpls.2024.1388163
Li, S., Wu, F., Duan, Y., Singerman, A., Guan, Z. (2020a). Citrus greening: management strategies and their economic impact. HortScience horts 55:5, 604–612. doi: 10.21273/HORTSCI14696-19
Liu, X., Zheng, Y., Wang-Pruski, G., Gan, Y., Zhang, B., Hu, Q., et al. (2019). Transcriptome profiling of periwinkle infected with Huanglongbing (‘Candidatus Liberibacter asiaticus’). Eur. J. Plant Pathol. 153, 891–906. doi: 10.1007/s10658-018-01607-9
Lombardi, R. L., Ramsey, J. S., Mahoney, J. E., MacCoss, M. J., Heck, M. L., Slupsky, C. M. (2024). Longitudinal Transcriptomic, Proteomic, and Metabolomic Response of Citrus sinensis to Diaphorina citri Inoculation of Candidatus Liberibacter asiaticus. J. Proteome Res. 23, 2857–2869. doi: 10.1021/acs.jproteome.3c00485
Lopes, S. A., Cifuentes-Arenas, J. C. (2021). A protocol for successful transmission of ‘Candidatus Liberibacter asiaticus’ from citrus to citrus using Diaphorina citri. Phytopathology 111, 12. doi: 10.1094/PHYTO-02-21-0076-R
Lopes, S. A., Frare, G. F., Bertolni, E., Cambra, M., Fernandes, N. G., Ayres, A. J., et al. (2009). Liberibacters associated with citrus huanglongbing in Brazil: ‘Candidatus Liberibacter asiaticus’ is heat tolerant, ‘Ca. L. americanus’ is heat sensitive. Plant Dis. 3, 257–262. doi: 10.1094/PDIS-93-3-0257
Lopes, S. A., Luiz, F. Q. B. Q., Martins, E. C., Fassini, C. G., Barbosa, J. C., Beattie, G. A. C. (2013). Candidatus Liberibacter asiaticus titers in citrus and acquisition rates by Diaphorina citri are decreased by higher temperature. Plant Dis. 97, 1563–1570. doi: 10.1094/PDIS-11-12-1031-RE
Love, M. I., Huber, W., Anders, S. (2014). Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 550. doi: 10.1186/s13059-014-0550-8
MacLean, A. M., Sugio, A., Makarova, O. V., Findlay, K. C., Grieve, V. M., Tóth, R., et al. (2011). Phytoplasma effector SAP54 induces indeterminate leaf-like flower development in Arabidopsis plants. Plant Physiol. 157:2, 831–834. doi: 10.1104/pp.111.181586
Manjunath, K. L., Halbert, S. E., Ramadugu, C., Weeb, S., Lee, R. F. (2008). Detection of ‘Candidatus Liberibacter asiaticus’ in Diaphorina citri and its importance in the management of Citrus Huanglongbing in Florida. Phytopathology 98, 387–396. doi: 10.1094/PHYTO-98-4-0387
Manni, M., Berkeley, M. R., Seppey, M., Zdobnov, E. M. (2021). ). BUSCO: assessing genomic data quality and beyond. Curr. Protoc. 1:12, e323. doi: 10.1002/cpz1.323
Martin, M. (2011). Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet.journal 17:1, 10–12. doi: 10.14806/ej.17.1.200
McIntyre, K. E., Bush, D. R., Argueso, C. T. (2021). Cytokinin regulation of source-sink relationships in plant-pathogen interactions. Front. Plant Sci. 24:12. doi: 10.3389/fpls.2021.677585
McNeil, C. J., Araujo, K., Godfrey, K., Slupsky, C. M. (2023). Metabolite Signature and Differential Expression of Genes in Washington Navel Oranges (Citrus sinensis) Infected by Spiroplasma citri. Phytopathology 113:2, 299–308. doi: 10.1094/PHYTO-05-22-0177-R
Merfa, M. V., Pérez-López, E., Naranjo, E., Jain, M., Gabriel, D. W., de la Fuente, L. (2019). Progress and obstacles in culturing ‘Candidatus liberibacter asiaticus’, the bacterium associated with huanglongbing. Phytopathology. 109:7, 1092–1110. doi: 10.1094/PHYTO-02-19-0051-RVW
Mine, A., Seyfferth, C., Kracher, B., Berens, M. L., Becker, D., Tsuda, K. (2018). The defense phytohormone signaling network enables rapid, high-amplitude transcriptional reprogramming during effector-triggered immunity. Plant Cell. 30, 1199–1219. doi: 10.1105/tpc.17.00970
Nguyen, C. H., Beattie, G. A. C., Haigh, A. M., Astuti, I. P., Mabberley, D. J., Weston, P. H., et al. (2019). Molecular differentiation of the Murraya paniculata Complex (Rutaceae: Aurantioideae: Aurantieae). BMC Evol. Biol. 30;19, 236. doi: 10.1186/s12862-019-1555-4
Oshima, K., Maejima, K., Namba, S. (2013). Genomic and evolutionary aspects of phytoplasmas. Front. Microbiol. 4, 230. doi: 10.3389/fmicb.2013.00230
Pandey, S. S., Wang, N. (2019). Targeted early detection of Citrus Huanglongbing causal agent ‘Candidatus Liberibacter asiaticus’ before symptom expression. Phytopathology 109, 952–959. doi: 10.1094/PHYTO-11-18-0432-R
Raiol-Junior, L. L., Baia, A. D. B., Luiz, F. Q. B. F., Fassini, C. G., Marques, V. V., Lopes, S. A. (2017). Improvement in the excised citrus leaf assay to investigate inoculation of ‘Candidatus Liberibacter asiaticus’ by the Asian Citrus Psyllid Diaphorina citri. Plant Dis. 101, 409–413. doi: 10.1094/PDIS-08-16-1093-RE
Raiol-Junior, L. L., Cifuentes-Arenas, J. C., Carvalho, E. V., Girardi, E. A., Lopes, S. A. (2021). Evidence that ‘Candidatus Liberibacter asiaticus’ moves predominantly towards new tissue growth in citrus plants. Plant Dis. 105, 34–42. doi: 10.1094/PDIS-01-20-0158-RE
Ramadugu, C., Keremane, M. L., Halbert, S. E., Duan, Y. P., Roose, M. L., Stover, E., et al. (2016). Long-term field evaluation reveals huanglongbing resistance in Citrus relatives. Plant Dis. 9, 1858–1869. doi: 10.1094/PDIS-03-16-0271-RE
Ramsey, J. S., Chin, E. L., Chavez, J. D., Saha, S., Mischuk, D., Mahoney, J., et al. (2020). Longitudinal Transcriptomic, Proteomic, and Metabolomic Analysis of Citrus limon Response to Graft Inoculation by Candidatus Liberibacter asiaticus. J. Proteome Res. 19:6, 2247–2263. doi: 10.1021/acs.jproteome.9b00802
Rattner, R., Thapa, S. P., Dang, T., Osman, F., Selvaraj, V., Maheshwari, Y., et al. (2021). Genome analysis of Spiroplasma citri strains from different host plants and its leafhopper vectors. BMC Genomics 22:1, 373. doi: 10.1186/s12864-021-07637-8
Remick, B. C., Gaidt, M. M., Vance, R. E. (2023). Effector-triggered immunity. Annu. Rev. Immunol. 26:41, 453–481. doi: 10.1146/annurev-immunol-101721-031732
Rodriguez-Saona, C., Polashock, J. J., Kyryczenko-Roth, V., Holdcraft, R., Jimenez-Gonzalez, G., De Moraes, C. M., et al. (2021). Application of plant defense elicitors fails to enhance herbivore resistance or mitigate phytoplasma infection in cranberries. Front. Plant Sci. . 12:12. doi: 10.3389/fpls.2021.700242
Scharte, J., Schö, H., Weis, E. (2005). Photosynthesis and carbohydrate metabolism in tobacco leaves during an incompatible interaction with Phytophthora nicotianae. Plant Cell Environ. 28, 1421–1435. doi: 10.1111/j.1365-3040.2005.01380.x
Setamou, M., Alabi, O. J., Simpson, C. R., Jifon, J. L. (2017). Contrasting amino acid profiles among permissive and non-permissive hosts of Candidatus Liberibacter asiaticus, putative causal agent of Huanglongbing. PloS One 13:12, e0187921. doi: 10.1371/journal.pone.0187921
Setamou, M., Simpson, C. R., Alabi, O. J., Nelson, S. D., Telagamsetty, S., Jifon, J. L. (2016). Quality matters: influences of citrus flush physicochemical characteristics on population dynamics of the asian citrus psyllid (Hemiptera: liviidae). PloS One 28:12, e0168997. doi: 10.1371/journal.pone.0168997
Shi, J., Gong, Y., Shi, H., Ma, X., Zhu, Y., Yang, F., et al. (2023b). Candidatus Liberibacter asiaticus’ secretory protein SDE3 inhibits host autophagy to promote Huanglongbing disease in citrus. Autophagy. 19:9, 2558–2574. doi: 10.1080/15548627.2023.2213040
Shi, Q., Pitino, M., Zhang, S., Krystel, J., Cano, L. M., Shatters, R. G., Jr, et al. (2019). Temporal and spatial detection of Candidatus Liberibacter asiaticus putative effector transcripts during interaction with Huanglongbing-susceptible, -tolerant, and -resistant citrus hosts. BMC Plant Biol. 2:19, 122. doi: 10.1186/s12870-019-1703-4
Shi, H., Yang, Z., Huang, J., Wu, H., Fu, S., Li, W., et al. (2023a). An effector of ‘Candidatus Liberibacter asiaticus’ manipulates autophagy to promote bacterial infection. J. Exp. Bot. 17:74, 4670–4684. doi: 10.1093/jxb/erad176
Skelley, L. H., Hoy, M. A. (2004). A synchronous rearing method for the Asian citrus psyllid and its parasitoids in quarantine. Biol. Control. 29, 14–23. doi: 10.1016/S1049-9644(03)00129-4
Subramanian, A., Tamayo, P., Mootha, V. K., Mukherjee, S., Ebert, B. L., Gillette, M. A., et al. (2005). Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. U S A. 25:102, 15545–15550. doi: 10.1073/pnas.0506580102
Sugio, A., MacLean, A. M., Kingdom, H. N., Grieve, V. M., Manimekalai, R., Hogenhout, S. A. (2011). Diverse targets of phytoplasma effectors: from plant development to defense against insects. Annu. Rev. Phytopathol. 49, 175–195. doi: 10.1146/annurev-phyto-072910-095323
Supek, F., Bošnjak, M., Škunca, N., Šmuc, T. (2011). REVIGO summarizes and visualizes long lists of gene ontology terms. PloS One 6:7, e21800. doi: 10.1371/journal.pone.0021800
Swarbrick, P. J., Schulze-Lefert, P., Scholes, J. D. (2006). Metabolic consequences of susceptibility and resistance (race-specific and broad-spectrum) in barley leaves challenged with powdery mildew. Plant Cell Environ. 29, 1061–1076. doi: 10.1111/j.1365-3040.2005.01472.x
Swingle, W. T., Reece, C. (1967). “The botany of Citrus and its wild relatives,” in The citrus industry. Eds. Reuther, W., Webber, H. J., Batchelor, L. D. (Univ. Calif. Press, Berkeley and Los Angeles), 191–430.
Tang, D., Chen, M., Huang, X., Zhang, G., Zeng, L., Zhang, G., et al. (2023). SRplot: A free online platform for data visualization and graphing. PloS One 9:18, e0294236. doi: 10.1371/journal.pone.0294236
Tao, Y., Xie, Z., Chen, W., Glazebrook, J., Chang, H. S., Han, B., et al. (2003). Quantitative nature of Arabidopsis responses during compatible and incompatible interactions with the bacterial pathogen Pseudomonas syringae. Plant Cell 15:2, 317–330. doi: 10.1105/tpc.007591
Teck, S. L. C., Fatimah, A., Beattie, A., Heng, R. K. J., King, W. S. (2011). Influence of host plant species and flush growth stage on the asian citrus psyllid, diaphorina citri kuwayama. Am. J. Agric. Biol. Sci. 6:4, 536–543. doi: 10.3844/ajabssp.2011.536.543
Thomma, B. P., Nürnberger, T., Joosten, M. H. (2011). Of PAMPs and effectors: the blurred PTI-ETI dichotomy. Plant Cell. 23:1, 4–15. doi: 10.1105/tpc.110.082602
Tomaseto, A. F., Marques, R. N., Fereres, A., Zanardi, O. A., Volpe, H. X. L., Alquézar, B., et al. (2019). Orange jasmine as a trap crop to control Diaphorina citri. Sci. Rep. 9, 2070. doi: 10.1038/s41598-019-38597-5
Vahling, C. M., Duan, Y., Lin, H. (2010). Characterization of an ATP translocase identified in the destructive plant pathogen “Candidatus Liberibacter asiaticus. J. Bacteriol 192:3, 834–840. doi: 10.1128/JB.01279-09
Wang, N., Trivedi, P. (2013). Citrus huanglongbing: a newly relevant disease presents unprecedented challenges. Phytopathology 103:7, 652–665. doi: 10.1094/PHYTO-12-12-0331-RVW
Weber, K. C., Mahmoud, L. M., Stanton, D., Welker, S., Qiu, W., Grosser, J. W., et al. (2022). Insights into the mechanism of Huanglongbing tolerance in the Australian finger lime (Citrus australasica). Front. Plant Sci. 21:13. doi: 10.3389/fpls.2022.1019295
Wei, X., Mira, A., Yu, Q., Gmitter, F. G., Jr. (2021). The Mechanism of Citrus Host Defense Response Repression at Early Stages of Infection by Feeding of Diaphorina citri Transmitting Candidatus Liberibacter asiaticus. Front. Plant Sci. 8;12. doi: 10.3389/fpls.2021.635153
Westbrook, C. J., Hall, D. G., Stover, E., Duan, Y. P., Lee, R. F. (2011). Colonization of Citrus and Citrus-related germplasm by Diaphorina citri (Hemiptera: Psyllidae). Hortscience 46, 997–1005. doi: 10.21273/HORTSCI.46.7.997
Wu, T., Lou, X., Xu, C., Wu, F., Qureshil, W. A., Cen, Y. (2016). ). Feeding behavior of Diaphorina citri and its transmission of ‘Candidatus Liberibacter asiaticus’ to citrus. E. ntomol. Exp. Appl. 161, 104–111. doi: 10.1111/eea.12496
Yan, Q., Sreedharan, A., Wei, S., Wang, J., Pelz-Stelinski, K., Folimonova, S., et al. (2013). Global gene expression changes in Candidatus Liberibacter asiaticus during the transmission in distinct hosts between plant and insect. Mol. Plant Pathol. 14, 391–404. doi: 10.1111/mpp.12015
Zhong, Y., Cheng, C. Z., Jiang, N. H., Jiang, B., Zhang, Y. Y., Wu, B., et al. (2015). Comparative Transcriptome and iTRAQ Proteome Analyses of Citrus Root Responses to Candidatus Liberibacter asiaticus Infection. PloS One 5:10, e0126973. doi: 10.1371/journal.pone.0126973
Zhou, G., Soufan, O., Ewald, J., Hancock, R. E. W., Basu, N., Xia, J. (2019). NetworkAnalyst 3.0: a visual analytics platform for comprehensive gene expression profiling and meta-analysis. Nucleic Acids Res. 47:W1, W234–W241. doi: 10.1093/nar/gkz240
Keywords: Citrus, Murraya, Bergera, HLB, transcriptomics, Liberibacter
Citation: Alves MN, Cifuentes-Arenas J, Niñoles R, Raiol-Junior LL, Carvalho E, Quirós-Rodriguez I, Ferro JA, Licciardello C, Alquezar B, Carmona L, Forment J, Bombarely A, Wulff NA, Peña L and Gadea J (2025) Transcriptomic analysis of early stages of ‘Candidatus Liberibacter asiaticus’ infection in susceptible and resistant species after inoculation by Diaphorina citri feeding on young shoots. Front. Plant Sci. 16:1502953. doi: 10.3389/fpls.2025.1502953
Received: 27 September 2024; Accepted: 27 January 2025;
Published: 20 February 2025.
Edited by:
Ute Albrecht, University of Florida, United StatesReviewed by:
Veronica Ancona, Texas A&M University Kingsville, United StatesFred Gmitter, University of Florida, United States
Copyright © 2025 Alves, Cifuentes-Arenas, Niñoles, Raiol-Junior, Carvalho, Quirós-Rodriguez, Ferro, Licciardello, Alquezar, Carmona, Forment, Bombarely, Wulff, Peña and Gadea. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: José Gadea, amdhZGVhdkBpYm1jcC51cHYuZXM=; Leandro Peña, bHBlbmFAaWJtY3AudXB2LmVz